
Contact with fibronectin enhances preservation of normal but not chronic myelogenous leukemia...
9
Experimental Hematology 30 (2002) 324–332 0301-472X/02 $–see front matter. Copyright © 2002 International Society for Experimental Hematology. Published by Elsevier Science Inc. PII S0301-472X(01)00799-8 Contact with fibronectin enhances preservation of normal but not chronic myelogenous leukemia primitive hematopoietic progenitors Ravi Bhatia, Andrea D. Williams, and Heidi A. Munthe Division of Hematology and Bone Marrow Transplantation, City of Hope National Medical Center, Duarte, Calif., USA (Received 26 October 2001; revised 14 December 2001; accepted 18 December 2001) Objective. Coculture with stromal cells enhances preservation and self-renewal of primitive progenitor potential in hematopoietic cells during ex vivo culture with growth factors (GF). However, the respective roles of growth factors, stromal contact, and extracellular matrix (ECM) ligands in this effect are not clear. Here we investigated the role of direct contact with stroma and the ECM protein fibronectin (FN) in these effects, and investigated whether ab- normal integrin receptor function in chronic myelogenous leukemia (CML) progenitors was associated with perturbation in these responses. Methods. Normal bone marrow CD34 cells were cultured in GF-containing medium with or without contact with stromal layers, glutaraldehyde-fixed stromal layers (stroma-contact), or integrin-binding FN fragments for 7 days. Progeny cells were assayed for primitive progeni- tors in week-6 long-term culture-initiating cell (LTC-IC) and week-10 extended LTC-IC (ELTC-IC) assays. Results. Increased LTC-IC and ELTC-IC preservation was seen following coculture with stroma, and was also observed after culture in contact with fixed stromal layers and FN. Both 4 1 and 5 1-integrin binding FN fragments enhanced LTC-IC preservation. Analysis of single CD34 CD38 cells showed that coculture with FN resulted in significantly reduced cell division, but enhanced retention of LTC-IC capacity in divided cells. FN also increased LTC-IC frequency in undivided cells. CML progenitors demonstrate deficient integrin-medi- ated adhesion, migration, and signaling. Coculture of CML CD34 cells with stroma and FN failed to enhance LTC-IC preservation. Conclusion. We conclude that 1 integrin-FN interactions enhance normal primitive progenitor preservation with or without cell division, and that these mechanisms are impaired in CML primitive progenitors. © 2002 International Society for Experimental Hematology. Published by Elsevier Science Inc. Although short-term culture with hematopoietic growth fac- tors (GF) may result in modest increases in primitive he- matopoietic progenitor cells, as measured in assays for long-term culture-initiating cells (LTC-IC) and nonobese diabetic/severe combined immunodeficient (NOD/SCID) mouse repopulating cells (SRC), extended exposure to GF is associated with depletion of primitive progenitor capacity [1–4]. Coculture with stromal cells can enhance maintenance of primitive progenitor capacity during ex vivo culture with GF, suggesting that additional microenvironmental factors might be required for optimal primitive progenitor preserva- tion and renewal during GF-induced cell-cycle transit [5–8. Recent studies suggest that stromal cells can retard the differ- entiation of proliferating primitive hematopoietic cells and promote self-renewing divisions [9]. The ability of stroma to enhance maintenance of primitive hematopoietic cells appears to be related in part to the pro- duction of diffusible factors by stromal cells, which may in- clude additional growth factors, chemokines, and proteogly- cans [10–12]. However, in addition to diffusible stromal factors, direct contact with stromal elements may be required for optimal preservation of primitive progenitor capacity dur- ing ex vivo culture [13]. Adhesive interactions between he- matopoietic stem and progenitor cells and stromal ligands, in addition to determining progenitor localization within the bone marrow stromal microenvironment, may regulate he- Offprint requests to: Ravi Bhatia, M.D., Division of Hematology and Bone Marrow Transplantation, City of Hope National Medical Center, 1500 E Duarte Road, Duarte, CA 91010; E-mail: [email protected]
-
Upload
ravi-bhatia -
Category
Documents
-
view
215 -
download
3
Transcript of Contact with fibronectin enhances preservation of normal but not chronic myelogenous leukemia...

Experimental Hematology 30 (2002) 324–332
0301-472X/02 $–see front matter. Copyright © 2002 International Society for Experimental Hematology. Published by Elsevier Science Inc.PII S0301-472X(01)00799-8
Contact with fibronectin enhances preservation of normalbut not chronic myelogenous leukemia primitive hematopoietic progenitors
Ravi Bhatia, Andrea D. Williams, and Heidi A. Munthe
Division of Hematology and Bone Marrow Transplantation, City of Hope National Medical Center, Duarte, Calif., USA
(Received 26 October 2001; revised 14 December 2001; accepted 18 December 2001)
Objective.
Coculture with stromal cells enhances preservation and self-renewal of primitiveprogenitor potential in hematopoietic cells during ex vivo culture with growth factors (GF).However, the respective roles of growth factors, stromal contact, and extracellular matrix(ECM) ligands in this effect are not clear. Here we investigated the role of direct contact withstroma and the ECM protein fibronectin (FN) in these effects, and investigated whether ab-normal integrin receptor function in chronic myelogenous leukemia (CML) progenitors wasassociated with perturbation in these responses.
Methods.
Normal bone marrow CD34
�
cells were cultured in GF-containing medium with orwithout contact with stromal layers, glutaraldehyde-fixed stromal layers (stroma-contact), orintegrin-binding FN fragments for 7 days. Progeny cells were assayed for primitive progeni-tors in week-6 long-term culture-initiating cell (LTC-IC) and week-10 extended LTC-IC(ELTC-IC) assays.
Results.
Increased LTC-IC and ELTC-IC preservation was seen following coculture withstroma, and was also observed after culture in contact with fixed stromal layers and FN. Both
�
4
�
1 and
�
5
�
1-integrin binding FN fragments enhanced LTC-IC preservation. Analysis ofsingle CD34
�
CD38
�
cells showed that coculture with FN resulted in significantly reducedcell division, but enhanced retention of LTC-IC capacity in divided cells. FN also increasedLTC-IC frequency in undivided cells. CML progenitors demonstrate deficient integrin-medi-ated adhesion, migration, and signaling. Coculture of CML CD34
�
cells with stroma and FNfailed to enhance LTC-IC preservation.
Conclusion.
We conclude that
�
1 integrin-FN interactions enhance normal primitive progenitorpreservation with or without cell division, and that these mechanisms are impaired in CMLprimitive progenitors. © 2002 International Society for Experimental Hematology. Published
by Elsevier Science Inc.
Although short-term culture with hematopoietic growth fac-tors (GF) may result in modest increases in primitive he-matopoietic progenitor cells, as measured in assays forlong-term culture-initiating cells (LTC-IC) and nonobesediabetic/severe combined immunodeficient (NOD/SCID)mouse repopulating cells (SRC), extended exposure to GFis associated with depletion of primitive progenitor capacity[1–4]. Coculture with stromal cells can enhance maintenanceof primitive progenitor capacity during ex vivo culture withGF, suggesting that additional microenvironmental factorsmight be required for optimal primitive progenitor preserva-
tion and renewal during GF-induced cell-cycle transit [5–8.Recent studies suggest that stromal cells can retard the differ-entiation of proliferating primitive hematopoietic cells andpromote self-renewing divisions [9].
The ability of stroma to enhance maintenance of primitivehematopoietic cells appears to be related in part to the pro-duction of diffusible factors by stromal cells, which may in-clude additional growth factors, chemokines, and proteogly-cans [10–12]. However, in addition to diffusible stromalfactors, direct contact with stromal elements may be requiredfor optimal preservation of primitive progenitor capacity dur-ing ex vivo culture [13]. Adhesive interactions between he-matopoietic stem and progenitor cells and stromal ligands, inaddition to determining progenitor localization within thebone marrow stromal microenvironment, may regulate he-
Offprint requests to: Ravi Bhatia, M.D., Division of Hematology andBone Marrow Transplantation, City of Hope National Medical Center,1500 E Duarte Road, Duarte, CA 91010; E-mail: [email protected]

R. Bhatia et al./Experimental Hematology 30 (2002) 324–332
325
matopoiesis by directly transducing growth regulatory signals[14,15]. Adhesion to stroma involves multiple receptors onnormal hematopoietic progenitors, including
�
4
�
1 and
�
5
�
1integrin receptors, CD44 and selectins, and a variety of stro-mal adhesive ligands, including cell-surface receptors such asVCAM-1 and ICAM-1, and extracellular matrix (ECM) pro-teins such as fibronectin (FN) and thrombospondin [16].
�
1integrin receptor binding to FN has been shown to play a crit-ical role in normal primitive and committed progenitors’ ad-herence to stroma [17–19]. Engagement of integrin receptorsby FN can result in transduction of signals that alter prolifera-tion and apoptosis in committed hematopoietic progenitors[15,20–23]. Dao et al. have shown that contact with FN en-hances transduction with retroviral vectors and supports pres-ervation of cells capable of long-term engraftment of immun-odeficient mice [24]. Enhanced transduction could be relatedto immobilization and colocalization of progenitors and retro-viral particles on FN [25]. Further studies are required to clar-ify whether direct interactions with stroma and FN can en-hance preservation of primitive hematopoietic progenitorsduring GF culture and promote self-renewing divisions ofproliferating primitive hematopoietic cells, as has been de-scribed for stromal coculture [9].
Chronic myelogenous leukemia is characterized by ab-normally increased proliferation and vast expansion of ma-lignant hematopoietic progenitors, their enhanced releaseinto the circulation, and extramedullary hematopoiesis. Ab-normal trafficking of malignant progenitors in CML may berelated to impaired
�
4
�
1 and
�
5
�
1 integrin–mediated adhe-sion to stroma and FN and abnormal mobility on FN sub-strates [26–29]. Integrin receptor abnormalities may alsocontribute to the vast expansion of malignant progenitorsand their progeny in CML, since malignant committed pro-genitors fail to respond to integrin-mediated inhibition ofprogenitor proliferation [30,31]. However, the occurrenceand nature of abnormalities in integrin-mediated growth reg-ulation have not been studied in CML primitive progenitors.
In this study we explored the hypothesis that integrin-mediated interactions with stroma and FN enhance preser-vation of normal primitive progenitors by increasing theirself-renewal after cell division and that these mechanismswere impaired in CML primitive progenitors. We comparedthe effect on preservation of primitive progenitor capacityof coculture with stroma, fixed metabolically inactivatedstromal layers, and FN during GF culture. We used single-cell analysis to further determine the mechanisms underly-ing the effects of FN on primitive progenitor preservation.Finally, we investigated the effect of contact with stromaand FN on preservation of CML primitive progenitors.
Methods
Selection of CD34
�
progenitors
Bone marrow samples were obtained from 17 normal individualsand 9 CML patients, who were enrolled in this study after in-
formed consent was obtained using guidelines approved by the In-stitutional Review Board of the City of Hope National MedicalCenter. Seven CML patients were in chronic phase and two in ac-celerated phase. All patients had received hydroxyurea and two pa-tients had also received interferon-
�
, which had been discontinuedfor 4 or more weeks prior to obtaining marrow samples.
Bone marrow mononuclear cells (BMMNC) were isolated byFicoll-Hypaque (Sigma Diagnostics, St. Louis, MO, USA) densitygradient separation (specific gravity 1.077) for 30 minutes at 400
g
.CD34
�
cell–enriched populations were selected from BMMNCusing immunomagnetic column separation (Miltenyi Biotech Inc.,Auburn, CA, USA).
Preparation of stromal layers
Bone marrow stromal layers were established in T-75 or T-150flasks by culturing normal BMMNC in long-term bone marrowculture (LTBMC) medium as previously described. Confluentstromal layers, formed after 4 to 5 weeks of culture, were irradi-ated at 1250 cGy using a Cesium irradiator to eliminate endoge-nous hematopoiesis and subcultured in 24-well plates (Costar,Cambridge, MA, USA) [32]. Fixation of stromal layers with glu-taraldehyde was carried out as described by Hurley et al. [15]. Glu-taraldehyde fixation leads to the inability of stromal layers to pro-duce diffusible factors such as GF, but preserves adhesive ligandssuch as FN, its CS1 binding site, thrombospondin, collagen, andVCAM and can support integrin receptor–mediated progenitor ad-hesion and inhibit normal colony-forming cell (CFC) proliferationto the same extent as intact, unfixed stromal layers [15].
Preparation of fibronectin substrates
Tryptic fragments of human plasma FN linking a COOH-terminalFN fragment containing the heparin binding and CS1 domains(FN-40) and a fragment containing the cell binding sequenceRGDS (FN-120) were obtained from Gibco. FN-CH296, a recom-binant FN fragment containing
�
4
�
1 and
�
5
�
1 integrin-bindingdomains, was obtained from PanVera Corporation, Madison, WI,USA. The FN fragments were adsorbed to wells of 24-well or 96-well plates at a concentration of 12
�
g/cm
2
(FN-40 and FN-120) or8
�
g/cm
2
(FN-CH296) as previously described [30].
Exposure to stroma and fibronectin
CD34
�
cells were cultured in serum-free medium (SFM; StemPro)supplemented with interleukin-3 (IL-3, 2 ng/mL), stem cell factor(SCF, 10 ng/mL), and Flt-3 ligand (FL, 20 ng/mL) (PeproTechInc., Rocky Hill, NJ, USA) in 24-well tissue culture plates. Cellswere cultured with GF alone, with preestablished viable stromallayers or glutaraldehyde-fixed stromal layers, or with fibronectin(FN-40, FN-120, and FN-CH296) substrates. Cells were harvestedafter 7 days of culture and assayed for progenitors as described be-low. Changes in progenitor number were evaluated by comparingthe number of CFC and LTC-IC present in the progeny of 10,000initially plated cells after culture with the number of CFC andLTC-IC present in 10,000 cells at initiation of culture.
Progenitor assaysColony-forming cells.
Cells were plated in semisolid methylcellu-lose progenitor culture for 14–18 days and assessed for the presenceof colony-forming unit granulocyte-macrophage and burst-formingunit erythroid (BFU-E) colonies as previously described [33].
Long-term culture-initiating cells.
We have used the term “long-term culture-initiating cells” (LTC-IC) to refer to the absolute

326
R. Bhatia et al./Experimental Hematology 30 (2002) 324–332
LTC-IC frequency calculated from limiting dilution assays and“LTC-CFC” (LTC-IC–derived CFC) to refer to the number ofCFC generated from LTC-IC during bulk cultures.
Bulk cultures.
Cells were plated in LTBMC medium on M2-10B4 murine fibroblast feeders previously subcultured in 24-wellplates. Cultures were maintained at 37
�
C in a humidified atmo-sphere with 5% CO
2
and fed at weekly intervals by removal of halfthe medium from the wells and replacement with fresh medium.After 6 weeks (for LTC-CFC assays) or 10 weeks [for extendedLTC-CFC (ELTC-CFC) assays], all nonadherent and adherentcells were harvested, pooled, and plated in methylcellulose pro-genitor culture and the number of CFC evaluated after 14 days.
Limiting dilution assay.
Limiting dilution analyses wereperformed as previously described by plating cells on M2-10B4feeders subcultured in 96-well plates. Cells were suspended in LT-BMC medium and plated at four different concentrations with 22replicates for each concentration. Cultures were maintained at37
�
C in a humidified atmosphere with 5% CO
2
with weekly half-medium exchanges with fresh medium for 6 weeks. Subsequently,80% of the medium was carefully removed without agitation andwells were overlaid with CFC growth-supporting medium. Wellswere scored as positive or negative after 14 days. The frequency ofLTC-IC was calculated, using L-CALC software (Stem Cell Tech-nologies, Vancouver, BC, Canada) from the reciprocal of the cellconcentration that gave 37% negative wells.
Single-cell assays.
Single CD34
�
CD38
�
cells were sorted intowells of 96-well plates with or without FN-40 precoating, using aMoFlow flow cytometer (Cytomation, Fort Collins, CO, USA).Cells were cultured in SFM with the following GF combination:FL (20 ng/mL), SCF (10 ng/mL) and interleukin (IL)-3 (2 ng/mL).After 7 days of culture, each well was examined for the number ofcells present, and wells in which cell division had taken place wereidentified. Next, the contents of each well were divided equallyand transferred to 4 wells containing preestablished M210-B4 stro-mal layers. Cells were cultured in standard LTBMC conditions for6 weeks. Subsequently, supernatant LTBMC medium was care-fully removed and wells were overlaid with medium supportingCFC growth and evaluated for CFC after 14 days. The presence ofLTC-IC in 2 or more of the 4 wells derived from a single well al-lowed us to detect whether LTC-IC replication had occurred. Sin-gle CD34
�
CD38
�
cells were also deposited directly onto M210-B4feeder layers to determine the LTC-IC frequency in the initiatingpopulation.
Statistical analysis.
Results of experimental points obtained frommultiple experiments were reported as mean
�
SEM. Significancelevels for differences between samples were determined using thetwo-tailed Student’s
t
-test. For single-cell analysis, the Fisher’s ex-act test was used to determine significance of differences in resultsobtained with or without FN.
Results
To investigate whether direct contact with stroma enhancedpreservation of primitive hematopoietic progenitors, normalmarrow CD34
�
progenitors were cultured in SFM containingIL-3 (2 ng/mL), SCF (10 ng/mL), and FL (20 ng/mL) for 7days, with or without viable stromal layers or glutaraldehyde-fixed, metabolically inactivated stromal layers. Progeny cells
were assayed for primitive and committed progenitors inLTC-IC and CFC assays, respectively. Exposure to both via-ble and fixed stromal layers during GF culture resulted in en-hanced expansion of week-6 LTC-CFC compared with GFalone as assessed in bulk culture (Fig. 1). Exposure to stromaduring GF culture also enhanced absolute numbers of week-6 LTC-IC as assessed by limiting dilution analysis [1.16
�
0.16-fold change in week-6 LTC-IC numbers with GF alone,2.0
�
0.23-fold expansion with fixed stroma, and 1.84
�
0.24-fold expansion with viable stroma (n
5,
p
0.01compared with no stroma)]. Stroma also enhanced preserva-tion of more primitive week-10 ELTC-CFC (Fig. 1). Theseresults indicate that direct contact–mediated interactionscontribute to enhanced preservation of primitive progenitorcapacity during stromal coculture. Preservation of primitiveprogenitors was not significantly different after exposure tofixed stroma vs viable stroma. In contrast, there was a trendtowards enhanced CFC expansion after coculture with via-ble stroma compared with fixed stroma, suggesting an addi-tional role for diffusible stromal-derived factors (
p
0.08).We also investigated the role of FN-integrin interactions
in primitive progenitor preservation, by culturing CD34
�
cells for 7 days in plates coated with an
�
4
�
1 integrin bind-ing FN fragment (FN-40). Exposure to FN-40 resulted intwofold enhancement of week-6 LTC-CFC numbers and a2.4-fold enhancement of week-10 LTC-CFC numbers butonly a 1.1-fold change in CFC numbers (Fig. 1). These re-sults were confirmed by limiting dilution analysis of abso-lute week-6 LTC-IC frequency [1.16
�
0.16-fold change inweek-6 LTC-IC numbers with GF alone vs 2.05
�
0.23with FN-40 (
p
0.01) and 0.9
�
0.13-fold change in week-10 LTC-IC numbers with GF alone vs 1.7
�
0.33 with FN-40 (n
5,
p
0.01)]. Exposure to FN-120 (
�
5
�
1 binding)and FN-CH296 (with both
�
4
�
1 and
�
5
�
1 integrin bindingdomains) also enhanced LTC-IC preservation (Fig. 2).These results indicate that engagement of both
�
4
�
1 and
�
5
�
1 integrin receptors by FN could enhance primitive pro-genitor preservation during GF culture. Subsequent experi-ments evaluating the effects of FN on primitive progenitorswere performed using FN-40. The ability of FN-40 to en-hance week-6 LTC-IC numbers was not seen following co-culture at higher GF concentrations [IL-3 (10–50 ng/mL),SCF (20–100 ng/mL), FL (50–200 ng/mL)] (Fig. 3). How-ever, week-10 LTC-IC numbers were enhanced at all GFconcentrations studied, indicating increased importance ofFN-mediated signals for preservation of these more primi-tive progenitors (Fig. 3).[FIG 3] Culture with FN-40 in theabsence of GF did not significantly affect LTC-IC numbers(not shown), suggesting that optimal LTC-IC preservationrequired combinatorial signaling through GF and adhesionreceptors.
We directly evaluated the effect of FN on the relationshipbetween proliferation and preservation of LTC-IC capacityby studying single CD34
�
CD38
�
cells cultured with GFwith or without FN. Cells were sorted into wells of 96-well

R. Bhatia et al./Experimental Hematology 30 (2002) 324–332
327
plates containing SFM and GF, with or without FN pread-sorbtion. After 7 days, wells in which cell division had takenplace were identified. The number of CD34
�
CD38
�
cellsthat underwent division was significantly reduced followingcoculture with FN compared with culture with GF alone[13% (68/528) vs 22% (117/528),
p
0.001] (Fig. 4). Thenumber of divisions undergone by proliferating cells wasalso reduced with FN coculture (Table 1). The number ofwells in which no cells were seen was similar with and with-out FN, suggesting that cell survival was not affected. Thecontents of each well were replated on preestablished stromallayers to initiate LTC-IC culture. The proportion of prolifer-ating clones that contained LTC-IC was significantly in-creased in cells cultured on FN [12.8% (9/68) with FN vs 6%(6/117) with GF alone,
p
0.05], indicating that FN enhancedpreservation of LTC-IC capacity following GF-induced celldivision (
increased LTC-IC renewal
) (Fig. 4). This reflectedboth LTC-IC-replicative divisions that generated two ormore LTC-IC from a single cell and non–LTC-IC-replica-tive divisions in which only one of the progeny cells re-tained LTC-IC capacity. The LTC-IC capacity of cells thatdid not divide following FN and GF coculture was not sig-nificantly higher than with GF alone [4.4% (18/411)] butwas significantly increased compared with CD34
�
CD38
�
cells plated directly on stroma [3% (13/440) in the initiatingpopulation vs 6.1% (28/460) after GF and FN culture,
p
0.05]. Overall, a significantly increased number of LTC-ICwere seen in CD34
�
CD38
�
cells cultured on FN [42 LTC-IC generated from 528 initially plated cells (8%)] comparedwith cells cultured with GF alone [26 LTC-IC from 528 ini-
Figure 1. Contact with stroma and FN enhances normal primitive progen-itor numbers during ex vivo culture with GF. Normal marrow CD34� pro-genitors were cultured in medium containing IL-3 (2 ng/mL), SCF (10 ng/mL), and FL (20 ng/mL) for 7 days, with or without coculture with viablestromal layers, glutaraldehyde-fixed stromal layers, or FN-40 preadsorbedplates. Progeny cells were assayed for primitive progenitors in week-6LTC-CFC (n 4–10) and week-10 ELTC-CFC (n 5–8) assays, andcommitted progenitors in CFC assays (n 6–7), respectively. Data repre-sent fold change in progenitor numbers after 7 days of culture comparedwith the number of progenitors present in input CD34� cells at day 0, priorto initiation of culture. Results shown are mean � SEM of separate experi-ments. * p 0.02, ** p 0.07; † p 0.006, †† p 0.01, ††† p 0.003; ^ p 0.001; ^^ p 0.06, ^^^ p 0.01 compared with GF alone. The progenitorfrequency per 10,000 input CD34� cells were as follows: CFC at initiation1097 � 289, after 7 days culture with GF alone 4392 � 1217, � viablestroma 10,086 � 2686, � fixed stroma 6450 � 1265, � FN 5246 � 2195;week-6 LTC-CFC at initiation 205 � 32, after 7 days culture with GFalone 164 � 32, � viable stroma 464 � 242, � fixed stroma 250 � 56, �FN 342 � 82; week-10 ELTC-IC at initiation 82 � 33, after 7 days culturewith GF alone 24 � 11, � viable stroma 129 � 99, � fixed stroma 71 �37, � FN 54 � 24.
Figure 2. Engagement of both �4�1 and �5�1 integrin receptors by FNcould enhance primitive progenitor preservation during GF culture. Nor-mal marrow CD34� progenitors were cultured on plates preadsorbed withFN-40, FN-120, and FN-CH296 in medium containing IL-3 (2 ng/mL),SCF (10 ng/mL), and FL (20 ng/mL) for 7 days. Progeny cells wereassayed for CFC, week-6 LTC-IC, and week-10 ELTC-IC as described inMethods. Data represent fold change in progenitor numbers after 7 days ofculture compared with cells cultured in GF alone. Results shown are mean �SEM of 5 experiments. * p 0.05, compared with GF alone. The week-6LTC-IC frequency in 10,000 CD34� cells was 162 � 60 at initiation ofculture, and 129 � 48 after 7 days of culture with GF alone.

328 R. Bhatia et al./Experimental Hematology 30 (2002) 324–332
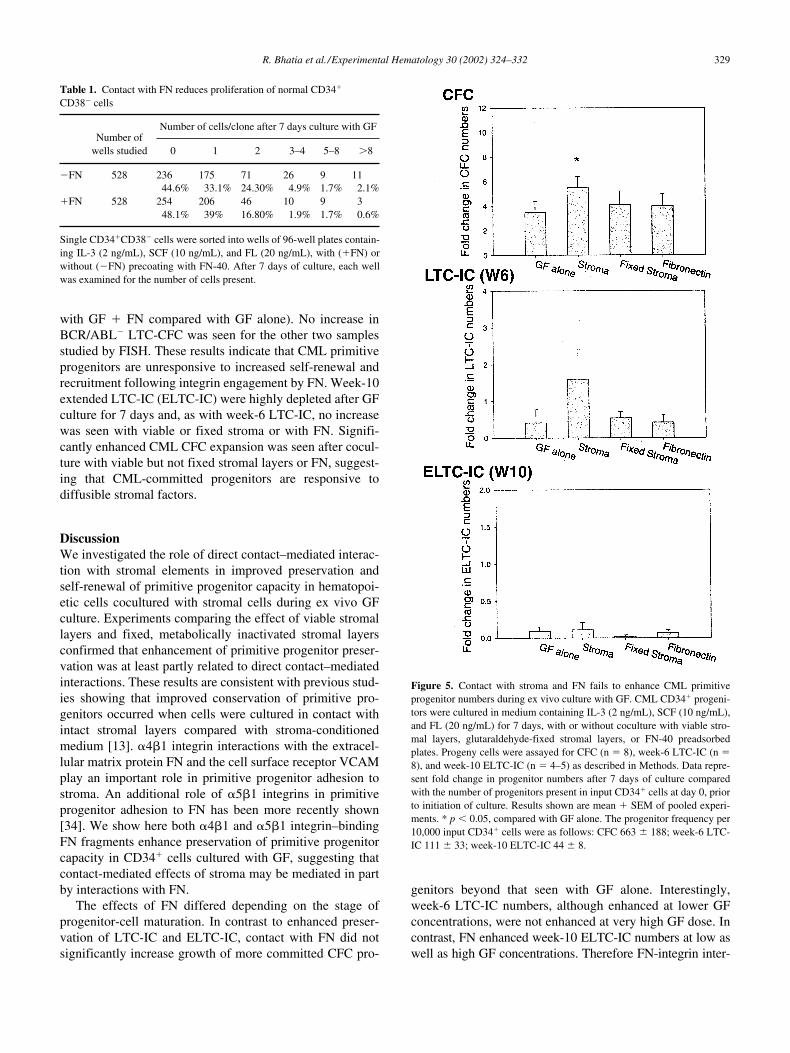
peared to increase CML LTC-IC preservation, this differ-ence was not statistically significant (Fig. 5). Fluorescencein situ hybridization (FISH) analysis of colonies generatedin LTC-IC assays for BCR/ABL status showed that in-creased LTC-IC preservation did not reflect outgrowth ofBCR/ABL� cells (data not shown). In contrast to normalLTC-IC, CML LTC-IC numbers were not significantly in-creased following coculture with fixed stromal layers (Fig.5), indicating unresponsiveness to contact-mediated en-hancement of primitive progenitor capacity. Similarly,CML LTC-IC numbers were not enhanced following cul-ture with FN-40. Enhanced outgrowth of BCR/ABL� cellswas seen on FISH analysis of LTC-CFC colonies from 3 of5 patients (BCR/ABL� cells in LTC-CFC increased from 8to 81%, 15 to 55% and 83 to 94% when cells were cultured
Figure 3. Effect of GF concentration on the ability of contact with FN toenhance normal primitive progenitor numbers. Normal marrow CD34�
progenitors were cultured on plates preadsorbed with FN-40 in mediumcontaining standard, “low” GF [IL-3 (2 ng/mL), SCF (10 ng/mL), and FL(20 ng/mL)], or “high” GF [IL-3 (50 ng/mL), SCF (100 ng/mL), and FL(200 ng/mL)] for 7 days. Progeny cells were assayed for CFC (n 5–6),week-6 LTC-IC (n 6–7), and week-10 ELTC-IC (n 6–7), as describedin Methods. Data represent fold change in progenitor numbers after 7 daysof culture compared with cells cultured in GF alone. Results shown aremean � SEM of separate experiments. * p 0.05, compared with GFalone. The CFC frequency per 10,000 CD34� cells at initiation of culturewas 1097 � 289, after 7 days of culture with low GF was 4421 � 1206,and with high GF was 4671 � 1167. The week-6 LTC-IC frequency per10,000 CD34� cells at initiation of culture was 194 � 34, after 7 days ofculture with low GF was 174 � 41, and with high GF was 512 � 130. Theweek-10 LTC-IC frequency per 10,000 CD34� cells at initiation of culturewas 99 � 42, after 7 days of culture with low GF was 27 � 14, and withhigh GF was 34 � 18.
tially plated cells (4.9%) (p 0.05)], confirming results ofbulk and limiting-dilution LTC-IC assays.
Finally, we evaluated the effect of coculture with fixedstromal layers and FN-40 on preservation of CML primitiveprogenitors. CML committed and primitive progenitors areknown to demonstrate abnormal �1 integrin function[18,28]. Although coculture with viable stromal layers ap-
Figure 4. FN enhances preservation of LTC-IC self-renewal followingGF-induced cell division. Single CD34�CD38� cells were sorted intowells of 96-well plates containing medium with IL-3 (2 ng/mL), SCF (10ng/mL), and FL (20 ng/mL), with or without precoating with FN-40. After7 days of culture, each well was examined for the number of cells present,and wells in which cell division had taken place were identified. The con-tents of each well were then divided equally and transferred to 4 wells con-taining preestablished stromal layers and cultured for 6 weeks in long-termculture, followed by assessment for CFC. The presence of LTC-IC in 2 ormore of the 4 wells derived from a single cell allowed detection of LTC-ICreplication. Significance value for differences in results obtained aftercoculture with GF and FN compared with GF alone are shown: * p
0.0001, **p 0.05. The LTC-IC frequency in CD34�CD38� cells plateddirectly on stroma was 3% (13 LTC-IC in 440 cells). Results representcombined data from 3 experiments.
R. Bhatia et al./Experimental Hematology 30 (2002) 324–332 329
with GF � FN compared with GF alone). No increase inBCR/ABL� LTC-CFC was seen for the other two samplesstudied by FISH. These results indicate that CML primitiveprogenitors are unresponsive to increased self-renewal andrecruitment following integrin engagement by FN. Week-10extended LTC-IC (ELTC-IC) were highly depleted after GFculture for 7 days and, as with week-6 LTC-IC, no increasewas seen with viable or fixed stroma or with FN. Signifi-cantly enhanced CML CFC expansion was seen after cocul-ture with viable but not fixed stromal layers or FN, suggest-ing that CML-committed progenitors are responsive todiffusible stromal factors.
DiscussionWe investigated the role of direct contact–mediated interac-tion with stromal elements in improved preservation andself-renewal of primitive progenitor capacity in hematopoi-etic cells cocultured with stromal cells during ex vivo GFculture. Experiments comparing the effect of viable stromallayers and fixed, metabolically inactivated stromal layersconfirmed that enhancement of primitive progenitor preser-vation was at least partly related to direct contact–mediatedinteractions. These results are consistent with previous stud-ies showing that improved conservation of primitive pro-genitors occurred when cells were cultured in contact withintact stromal layers compared with stroma-conditionedmedium [13]. �4�1 integrin interactions with the extracel-lular matrix protein FN and the cell surface receptor VCAMplay an important role in primitive progenitor adhesion tostroma. An additional role of �5�1 integrins in primitiveprogenitor adhesion to FN has been more recently shown[34]. We show here both �4�1 and �5�1 integrin–bindingFN fragments enhance preservation of primitive progenitorcapacity in CD34� cells cultured with GF, suggesting thatcontact-mediated effects of stroma may be mediated in partby interactions with FN.
The effects of FN differed depending on the stage ofprogenitor-cell maturation. In contrast to enhanced preser-vation of LTC-IC and ELTC-IC, contact with FN did notsignificantly increase growth of more committed CFC pro-
genitors beyond that seen with GF alone. Interestingly,week-6 LTC-IC numbers, although enhanced at lower GFconcentrations, were not enhanced at very high GF dose. Incontrast, FN enhanced week-10 ELTC-IC numbers at low aswell as high GF concentrations. Therefore FN-integrin inter-
Table 1. Contact with FN reduces proliferation of normal CD34�
CD38� cells
Number ofwells studied
Number of cells/clone after 7 days culture with GF
0 1 2 3–4 5–8 �8
�FN 528 236 175 71 26 9 1144.6% 33.1% 24.30% 4.9% 1.7% 2.1%
�FN 528 254 206 46 10 9 348.1% 39% 16.80% 1.9% 1.7% 0.6%
Single CD34�CD38� cells were sorted into wells of 96-well plates contain-ing IL-3 (2 ng/mL), SCF (10 ng/mL), and FL (20 ng/mL), with (�FN) orwithout (�FN) precoating with FN-40. After 7 days of culture, each wellwas examined for the number of cells present.
Figure 5. Contact with stroma and FN fails to enhance CML primitiveprogenitor numbers during ex vivo culture with GF. CML CD34� progeni-tors were cultured in medium containing IL-3 (2 ng/mL), SCF (10 ng/mL),and FL (20 ng/mL) for 7 days, with or without coculture with viable stro-mal layers, glutaraldehyde-fixed stromal layers, or FN-40 preadsorbedplates. Progeny cells were assayed for CFC (n 8), week-6 LTC-IC (n 8), and week-10 ELTC-IC (n 4–5) as described in Methods. Data repre-sent fold change in progenitor numbers after 7 days of culture comparedwith the number of progenitors present in input CD34� cells at day 0, priorto initiation of culture. Results shown are mean � SEM of pooled experi-ments. * p 0.05, compared with GF alone. The progenitor frequency per10,000 input CD34� cells were as follows: CFC 663 � 188; week-6 LTC-IC 111 � 33; week-10 ELTC-IC 44 � 8.

330 R. Bhatia et al./Experimental Hematology 30 (2002) 324–332
actions may play a greater role in promoting preservation andexpansion of the most primitive myeloid progenitors and maybecome less critical with increasing maturation. On the otherhand, the role of GF stimulation may assume greater impor-tance with progenitor maturation. These results are consistentwith studies showing that stromal support or coculture withFN was essential to maintain long-term but not short-term en-graftment capacity in progenitor cells during ex vivo cultureand transduction [24,35].
Integrin engagement by FN results in inhibition of com-mitted progenitor proliferation [15,20. We demonstrate herethat primitive progenitor proliferation is also decreased inthe presence of FN. However, FN also enhanced preserva-tion of primitive progenitors in dividing cells, suggesting thatit prevents loss of primitive progenitor capacity associatedwith GF-induced proliferation, presumably by preventingdifferentiation. Increased primitive progenitor preservationfollowing cell division resulted from both LTC-IC–replica-tive divisions (that generated two or more LTC-IC from onecell), and LTC-IC–nonreplicative divisions (that generatedonly a single LTC-IC). These effects are similar to thosepreviously reported for stromal cells, which counteract theloss of LTC-IC induced by GF by retarding differentiation,independent of proliferation [8,9]. LTC-IC capacity in undi-vided CD34�CD38� cells was also enhanced after coculturewith FN and GF compared with LTC-IC frequency in inputcells prior to GF culture. Transit through cell cycle has beenassociated with reduction in regenerating capacity of primi-tive hematopoietic cells [4,36,37]. Increased primitive pro-genitor recruitment by GF (i.e., activation of quiescent cellsto grow in LTC-IC assays) [38], in combination with inhibi-tion of cell-cycle progression by FN, may explain enhancedLTC-IC frequency in cells cocultured with GF and FN.However, other mechanisms such as increased survival can-not be ruled out [39]. These results do not exclude the possi-bility that interactions with ligands for other ECM proteinsor other cell surface receptors may also play a role.
The molecular mechanisms by which integrin engage-ment by FN may retard differentiation and enhance self-re-newal of primitive hematopoietic progenitors undergoingcell division are not clear. Integrin engagement can regulatedifferentiation in other cell types through alteration in keytranscription factors. For example, �2�1 integrins regulateOsf2 expression and activity in osteoblastic cells [40], andadhesion to vitronectin regulates sonic hedgehog activityduring cerebellar neuron development [41]. Signals through�1 integrins and MAP kinase cooperate to maintain the epi-dermal stem cell compartment in vitro [42]. The effect of in-tegrin engagement on key transcription factors that regulatemyeloid differentiation of primitive hematopoietic progeni-tors needs further investigation. Although we have mea-sured direct effects of integrin engagement by FN in thisstudy, integrins could also regulate primitive progenitorgrowth by playing an essential role in proper microenviron-mental localization.
Primitive and committed hematopoietic progenitors fromCML patients demonstrate abnormal �1 integrin–mediatedadhesion [26,27]. Malignant cells in CML are characterizedby the presence of the BCR/ABL fusion oncogene [43,44],which plays a critical role in the pathogenesis of CML [45–47]. Defects in integrin function in CML are directly relatedto BCR/ABL expression, since they can be reversed by in-hibiting BCR/ABL expression or kinase activity and in-duced in normal cells by introduction of the BCR/ABL gene[48–50]. In addition to defects in adhesion and migration,integrin signaling leading to inhibition of proliferation isalso impaired in committed CML progenitors [30,31]. Theresults of the present study demonstrate that signalingmechanisms induced following contact with stroma or FNthat enhance primitive progenitor preservation and self-renewal are also impaired in CML. The mechanisms under-lying these abnormalities are not clear. It is possible thatconstitutive activation by BCR/ABL of signaling through theRas-MAPK and PI-3 kinase pathways, that play importantroles in combinatorial signaling through GF and integrin re-ceptors, could antagonize normal signaling through thesepathways and contribute to these growth regulatory abnor-malities [51,52].
CML primitive progenitors demonstrate increased prolif-eration and maturation in response to GF stimulation com-pared with their normal counterparts [38,53,54. Impairedpreservation of primitive progenitor potential in response tosignals from stroma and FN could enhance the increasedgeneration of malignant committed progenitors and theirprogeny from primitive progenitors in CML. The combina-tion of increased primitive progenitor proliferation and mat-uration in response to GF stimulation, and reduced inhibi-tion of differentiation and proliferation in response tointegrin engagement, could contribute to the vast expansionof differentiated hematopoietic cells characteristic of CML.Differences in the effects of integrin-FN interactions onnormal and CML primitive progenitor growth could be usedin strategies for selective expansion/preservation of nonma-lignant progenitors from CML progenitor populations.
We conclude that integrin-FN interactions enhancedpreservation of primitive progenitor potential during exvivo culture with GF as a result of increased self-renewingdivisions as well as through enhancement of primitive pro-genitor capacity in undivided cells. Impaired integrin func-tion in CML primitive progenitors are associated with unre-sponsiveness to the growth regulatory effects of integrinengagement by FN.
AcknowledgmentsThis work was supported in part by the Public Health Servicesgrants CA74455 from the National Cancer Institute, AmericanCancer Society grant RPG-99-202-01-LBC, and The Leukemiaand Lymphoma Society Translational Research Grant 6468 toRavi Bhatia.

R. Bhatia et al./Experimental Hematology 30 (2002) 324–332 331
We are grateful to Dr. Marilyn Slovak and Feiyu Zhang in theCytogenetics Laboratory for FISH analysis for BCR/ABL. We ac-knowledge the excellent technical support of Lucy Brown in theAnalytical Cytometry Core. We would also like to express ourgratitude to Alison Ahlers, R.N. and Jeanine Stevenson, R.N., andthe physicians and staff in the Division of Hematology/BMT forassistance with patient samples.
References1. Peters SO, Kittler EL, Ramshaw HS, Quesenberry PJ (1996) Ex vivo
expansion of murine marrow cells with interleukin-3 (IL-3), IL-6, IL-11, and stem cell factor leads to impaired engraftment in irradiatedhosts. Blood 87:30
2. Habibian HK, Peters SO, Hsieh CC, et al. (1998) The fluctuating phe-notype of the lymphohematopoietic stem cell with cell cycle transit. JExp Med 188:393
3. Glimm H, Eaves CJ (1999) Direct evidence for multiple self-renewaldivisions of human in vivo repopulating hematopoietic cells in short-term culture. Blood 94:2161
4. Glimm H, Oh IH, Eaves CJ (2000) Human hematopoietic stem cellsstimulated to proliferate in vitro lose engraftment potential during theirS/G(2)/M transit and do not reenter G(0). Blood 96:4185
5. Brandt JE, Galy AH, Luens KM, et al. (1998) Bone marrow repopula-tion by human marrow stem cells after long-term expansion culture ona porcine endothelial cell line. Exp Hematol 26:950
6. Emerson SG (1996) Ex vivo expansion of hematopoietic precursors,progenitors, and stem cells: the next generation of cellular therapeu-tics. Blood 87:3082
7. Moore KA, Ema H, Lemischka IR (1997) In vitro maintenance of highlypurified, transplantable hematopoietic stem cells. Blood 89:4337
8. Bennaceur-Griscelli A, Tourino C, Izac B, Vainchenker W, Coulom-bel L (1999) Murine stromal cells counteract the loss of long-term cul-ture-initiating cell potential induced by cytokines in CD34�CD38low/�)human bone marrow cells. Blood 94:529
9. Bennaceur-Griscelli A, Pondarre C, Schiavon V, Vainchenker W,Coulombel L (2001) Stromal cells retard the differentiation ofCD34�CD38low/� human primitive progenitors exposed to cytokinesindependent of their mitotic history. Blood 97:435
10. Verfaillie CM, Catanzarro PM, Li WN (1994) Macrophage inflamma-tory protein 1 �, interleukin-3 and diffusible marrow stromal factorsmaintain human hematopoietic stem cells for at least eight weeks invitro. J Exp Med 179:643
11. Gupta P, McCarthy JB, Verfaillie CM (1996) Stromal fibroblast hepa-ran sulfate is required for cytokine-mediated ex vivo maintenance ofhuman long-term culture-initiating cells. Blood 87:3229
12. Lewis ID, Almeida-Porada G, Du J, et al. (2001) Umbilical cord bloodcells capable of engrafting in primary, secondary, and tertiary xenoge-neic hosts are preserved after ex vivo culture in a noncontact system.Blood 97:3441
13. Breems DA, Blokland EAW, Siebel KE, Mayen AEM, Engels LJA,Ploemacher RE (1998) Stroma-contact prevents loss of hematopoieticstem cell quality during ex vivo expansion of CD34� mobilized pe-ripheral blood stem cells. Blood 91:111
14. Verfaillie C, Hurley R, Bhatia R, McCarthy JB (1994) Role of bonemarrow matrix in normal and abnormal hematopoiesis. Crit Rev OncolHematol 16:201
15. Hurley RW, McCarthy JB, Verfaillie CM (1995) Direct adhesion tobone marrow stroma via fibronectin receptors inhibits hematopoieticprogenitor proliferation. J Clin Invest 96:511
16. Verfaillie CM (1998) Adhesion receptors as regulators of the hemato-poietic process. Blood 92:2609
17. Williams DA, Rios M, Stephens C, Patel VP (1991) Fibronectin andVLA-4 in hematopoietic stem cell–microenvironment interactions.Nature 352:438
18. Verfaillie CM, McCarthy JB, McGlave PB (1991) Differentiation ofprimitive human multipotent hematopoietic progenitors into single lin-eage clonogenic progenitors is accompanied by alterations in their ad-hesion to fibronectin. J Exp Med 174:693
19. Teixido J, Hemler ME, Greenberger JS, Anklesaria P (1992) Role of�1 and �2 integrins in the adhesion of human CD34hi stem cells tobone marrow stroma. J Clin Invest 90:358
20. Hurley RW, McCarthy JB, Wayner EA, Verfaillie CM (1997) Mono-clonal antibody crosslinking of the �4 or �1 integrin inhibits commit-ted clonogenic hematopoietic progenitor proliferation. Exp Hematol25:321
21. Schofield KP, Humphries MJ, de Wynter E, Testa N, Gallagher JT(1998) The effect of �4�1-integrin binding sequences of fibronectin ongrowth of cells from human hematopoietic progenitors. Blood 91:3230
22. Yokota T, Oritani K, Mitsui H, et al. (1998) Growth-supporting activi-ties of fibronectin on hematopoietic stem/progenitor cells in vitro andin vivo: structural requirement for fibronectin activities of CS1 andcell-binding domains. Blood 91:3263
23. Kapur R, Cooper R, Zhang L, Williams DA (2001) Cross-talk between�4�1/�5�1 and c-kit results in opposing effect on growth and survivalof hematopoietic cells via the activation of focal adhesion kinase, mi-togen-activated protein kinase, and Akt signaling pathways. Blood97:1975
24. Dao MA, Hashino K, Kato I, Nolta JA (1998) Adhesion to fibronectinmaintains regenerative capacity during ex vivo culture and transduc-tion of human hematopoietic stem and progenitor cells. Blood 92:4612
25. Hanenberg H, Xiao XL, Dilloo D, Hashino K, Kato I, Williams DA(1996) Colocalization of retrovirus and target cells on specific fi-bronectin fragments increases genetic transduction of mammaliancells. Nat Med 2:876
26. Gordon MY, Dowding CR, Riley GP, Goldman JM, Greaves MF(1984) Altered adhesive interactions with marrow stroma of hemato-poietic progenitor cells in chronic myelogenous leukaemia. Nature328:342
27. Verfaillie CM, McCarthy JB, McGlave PB (1992) Mechanisms under-lying abnormal trafficking of malignant progenitors in chronic myel-ogenous leukemia: Decreased adhesion to stroma and fibronectin butincreased adhesion to the basement membrane components lamininand collagen type IV. J Clin Invest 90:1232
28. Salgia R, Li J-L, Ewaniuk DS, et al. (1997) BCR/ABL induces multi-ple abnormalities of cytoskeletal function. J Clin Invest 100:46
29. Salgia R, Quackenbush E, Lin J, et al. (1999) The BCR/ABL oncogenealters the chemotactic response to stromal-derived factor-1�. Blood94:4233
30. Bhatia R, McCarthy JB, Verfaillie CM (1996) Interferon-� restoresnormal �1 integrin–mediated inhibition of hematopoietic progenitorproliferation by the marrow microenvironment in chronic myeloge-nous leukemia. Blood 87:3883
31. Jiang Y, Zhao RC, Verfaillie CM (2000) Abnormal integrin-mediatedregulation of chronic myelogenous leukemia CD34� cell proliferation:BCR/ABL upregulates the cyclin-dependent kinase inhibitor, p27Kip,which is relocated to the cell cytoplasm and incapable of regulatingcdk2 activity. Proc Natl Acad Sci U S A 97:10538
32. Bhatia R, McGlave PB, Dewald GW, Blazar BR, Verfaillie CM (1995)Abnormal function of the bone marrow microenvironment in chronicmyelogenous leukemia: role of malignant stromal macrophages. Blood85:3636
33. Bhatia R, Wayner EA, McGlave PB, Verfaillie CM (1994) Interferon-� restores normal adhesion of chronic myelogenous leukemia hemato-poietic progenitors to bone marrow stroma by correcting impaired �1integrin receptor function (see comments). J Clin Invest 94:384
34. van der Loo JC, Xiao X, McMillin D, Hashino K, Kato I, Williams DA(1998) VLA-5 is expressed by mouse and human long-term repopulat-ing hematopoietic cells and mediates adhesion to extracellular matrixprotein fibronectin. J Clin Invest 102:1051
35. Nolta JA, Smogorzewska EM, Kohn DB (1995) Analysis of optimal

332 R. Bhatia et al./Experimental Hematology 30 (2002) 324–332
conditions for retroviral-mediated transduction of primitive human he-matopoietic cells. Blood 86:101
36. Habibian HK, Peters SO, Hsieh CC, et al. (1998) The fluctuating phe-notype of the lymphohematopoietic stem cell with cell cycle transit. JExp Med 188:393
37. Gothot A, van der Loo JC, Clapp DW, Srour EF (1998) Cell cycle–related changes in repopulating capacity of human mobilized periph-eral blood CD34� cells in nonobese diabetic/severe combined im-mune-deficient mice. Blood 92:2641
38. Bhatia R, Munthe HA, Williams AD, Zhang F, Forman SJ, Slovak ML(2000) Increased sensitivity of chronic myelogenous leukemia primi-tive hematopoietic progenitors to growth factor–induced cell divisionand maturation. Exp Hematol 12:1401
39. Wang MW, Consoli U, Lane CM, et al. (1998) Rescue from apoptosisin early (CD34-selected) vs late (non–CD34-selected) human hemato-poietic cells by very late antigen 4- and vascular cell adhesion mole-cule (VCAM) 1–dependent adhesion to bone marrow stromal cells.Cell Growth Differ 9:105
40. Xiao G, Wang D, Benson MD, Karsenty G, Francheski RT (1998)Role of the �2-integrin in osteoblast-specific gene expression and acti-vation of Osf2 transcription factor. J Biol Chem 273:32988
41. Pons S, Trejo JL, Martinez-Morales JR, Marti E (2001) Vitronectinregulates Sonic hedgehog activity during cerebellum developmentthrough CREB phosphorylation. Development 128:1481
42. Zhu AJ, Haase I, Watt FM (1999) Signaling via �1 integrins and mito-gen-activated protein kinase determines human stem cell fate in vitro.Proc Natl Acad Sci U S A 96:6728
43. Rowley JD (1973) A new consistent chromosome abnormality inchronic myelogenous leukemia identified by quinacrine fluorescenceand Giemsa staining. Nature 243:209
44. DeKlein A, Van Kessel AG, Grosveld G, et al. (1982) A cellular onco-gene is translocated to the Philadelphia chromosome in chronic myelo-cytic leukemia. Nature 300:765
45. Daley GQ, Baltimore D (1988) Transformation of an interleukin-3–depen-
dent hematopoietic cell line by the chronic leukemia-specific P210 bcr/ablprotein. Proc Natl Acad Sci U S A 85:9312
46. Daley GQ, Van Etten RA, Baltimore D (1990) Induction of chronicmyelogenous leukemia in mice by the p210 bcr/abl gene of the Phila-delphia chromosome. Science 247:824
47. Pear WS, Miller JP, Xu L, et al. (1998) Efficient and rapid inductionof a chronic myelogenous leukemia–like myeloproliferative diseasein mice receiving P210 bcr/abl-transduced bone marrow. Blood 92:3780
48. Bhatia R, Verfaillie CM (1998) Inhibition of BCR-ABL expressionwith antisense oligonucleotides restores �1 integrin–mediated adhe-sion and proliferation inhibition in chronic myelogenous leukemia he-matopoietic progenitors. Blood 91:3414
49. Bhatia R, Munthe HA, Verfaillie CM (1998) Tyrphosin AG 957, a ty-rosine kinase inhibitor with anti-BCR/ABL tyrosine kinase activity, re-stores �1 integrin–mediated adhesion and inhibitory signaling in chronicmyelogenous leukemia hematopoietic progenitors. Leukemia 12:1708
50. Zhao RC, Fan E, Verfaillie CM (1999) Introduction of BCR/ABL innormal CD34� cells recreates the phenotype characteristics of CML.Blood 94:388A (abstr.)
51. Goga A, McLaughlin J, Afar DEH, Saffran DC, Witte ON (1995) Al-ternative signals to RAS for hematopoietic transformation by theBCR-ABL oncogene. Cell 82:981
52. Skorski T, Bellacosa A, Nieborowska-Skorska M, et al. (1997) Trans-formation of hematopoietic cells by BCR/ABL requires activation of aPI-3k/Akt-dependent pathway. EMBO J 16:6151
53. Petzer AL, Eaves CJ, Barnett MJ, Eaves AC (1997) Selective expan-sion of primitive normal hematopoietic cells in cytokine-supplementedcultures of purified cells from patients with chronic myeloid leukemia.Blood 90:64
54. Traycoff CM, Halstead B, Rice S, McMahel J, Srour EF, Cornetta K(1998) Chronic myelogenous leukaemia CD34� cells exit G0/G1phases of cell cycle more rapidly than normal marrow CD34� cells. BrJ Haematol 102:759


















