
Conservation and Developmental Control of Alternative Splicing in maebl Among Malaria Parasites
11
Conservation and Developmental Control of Alternative Splicing in maebl Among Malaria Parasites Naresh Singh 1 , Peter Preiser 2 †, Laurent Re ´nia 3 †, Bharath Balu 1 John Barnwell 4 , Peter Blair 1 , William Jarra 2 , Tatiana Voza 5 , Ire ` ne Landau 5 and John H. Adams 1 * 1 Department of Biological Sciences, University of Notre Dame, 220 Galvin, PO Box 369 Notre Dame, IN 46556, USA 2 Division of Parasitology National Institute for Medical Research, The Ridgeway, Mill Hill NW7 1AA, UK 3 De ´partement d’Immunologie Institut Cochin, INSERM U567 CNRS 8104, Universite ´ Rene ´ Descartes, Ho ˆpital Cochin Ba ˆtiment Gustave Roussy 75014 Paris, France 4 Division of Parasitic Diseases National Center for Infectious Diseases, Centers for Disease Control & Prevention, Mailstop F-13, Bldg. 22B, 4770 Buford Highway, NE, Chamblee, GA 30341, USA 5 Museum National d’Histoire Naturelle, F-75231 Paris France Genes of malaria parasites and other unicellular organisms have larger exons with fewer and smaller introns than metaozoans. Such differences in gene structure are perceived to extend to simpler mechanisms for transcriptional control and mRNA processing. Instead, we discovered a surprisingly complex level of post-transcriptional mRNA processing in analysis of maebl transcripts in several Plasmodium species. Mechanisms for internal alternative cis-splicing and exon skipping were active in multiple life cycle stages to change exon structure in the deduced coding sequence (CDS). The major alternatively spliced transcript utilized a less favorable acceptor splice site, which shifted codon triplet usage to a different CDS with a hydrophilic C terminus, changing the canonical type I membrane MAEBL product to a predicted soluble isoform. We found that develop- mental control of the alternative splicing pattern was distinct from the canonical splicing pattern. Western blot analysis indicated that MAEBL expression was better correlated with the appearance of the canonical ORF1 transcript. Together these data reveal that RNA metabolism in unicellular eukaryotes like Plasmodium is more sophisticated than believed and may have a significant role regulating gene expression in Plasmodium. q 2004 Elsevier Ltd. All rights reserved. Keywords: malaria; Plasmodium; alternative splicing; MAEBL; ligand *Corresponding author Introduction Malaria is one of the most serious human diseases causing several million deaths and clinical illness in hundreds of millions of people every year. The biological complexity of these pathogenic proto- zoans is extraordinary in the intricate life cycles, as the malaria parasite must develop in two very different host organisms, infecting different cell types, and its motile stages must often traverse different tissues before infecting a new cell. 1 Never- theless, the Plasmodium genome is estimated to have less than 6000 genes and the number of unique genes is still less since a considerable part of the Plasmodium genome is occupied by multi-gene families (e.g. var , rif, stevor). 2,3 Compared to the 0022-2836/$ - see front matter q 2004 Elsevier Ltd. All rights reserved. † P.P. and L.R. contributed equally to this work. Present addresses:P. Preiser, Nanyang Technological University, School of Biological Sciences, 60 Nanyang Drive, Singapore 637551; P. Blair, Department of Biology, Earlham College, 801 National Road West, Richmond, IN 47374, USA. Abbreviations used: AMA-1, apical membrane antigen- 1; DBP, Duffy antigen binding protein; DBL, duffy binding-like; EBA-175, erythrocyte binding antigen-175; ebl, erythrocyte binding-like; EBP, erythrocyte binding protein; IFA, indirect immunofluorescence assay; ORF, open reading frame. E-mail address of the corresponding author: [email protected] doi:10.1016/j.jmb.2004.08.047 J. Mol. Biol. (2004) 343, 589–599
-
Upload
naresh-singh -
Category
Documents
-
view
214 -
download
2
Transcript of Conservation and Developmental Control of Alternative Splicing in maebl Among Malaria Parasites

doi:10.1016/j.jmb.2004.08.047 J. Mol. Biol. (2004) 343, 589–599
Conservation and Developmental Control of AlternativeSplicing in maebl Among Malaria Parasites
Naresh Singh1, Peter Preiser2†, Laurent Renia3†, Bharath Balu1
John Barnwell4, Peter Blair1, William Jarra2, Tatiana Voza5, Irene Landau5
and John H. Adams1*
1Department of BiologicalSciences, University of NotreDame, 220 Galvin, PO Box 369Notre Dame, IN 46556, USA
2Division of ParasitologyNational Institute for MedicalResearch, The Ridgeway, MillHill NW7 1AA, UK
3Departement d’ImmunologieInstitut Cochin, INSERM U567CNRS 8104, Universite ReneDescartes, Hopital CochinBatiment Gustave Roussy75014 Paris, France
4Division of Parasitic DiseasesNational Center for InfectiousDiseases, Centers for DiseaseControl & Prevention, MailstopF-13, Bldg. 22B, 4770 BufordHighway, NE, Chamblee, GA30341, USA
5Museum National d’HistoireNaturelle, F-75231 ParisFrance
0022-2836/$ - see front matter q 2004 E
† P.P. and L.R. contributed equallPresent addresses: P. Preiser, Nan
University, School of Biological ScieDrive, Singapore 637551; P. Blair, DEarlham College, 801 National Road47374, USA.Abbreviations used: AMA-1, apic
1; DBP, Duffy antigen binding protebinding-like; EBA-175, erythrocyteebl, erythrocyte binding-like; EBP, eprotein; IFA, indirect immunofluoreopen reading frame.E-mail address of the correspond
Genes of malaria parasites and other unicellular organisms have largerexons with fewer and smaller introns than metaozoans. Such differences ingene structure are perceived to extend to simpler mechanisms fortranscriptional control and mRNA processing. Instead, we discovered asurprisingly complex level of post-transcriptional mRNA processing inanalysis of maebl transcripts in several Plasmodium species. Mechanisms forinternal alternative cis-splicing and exon skipping were active in multiplelife cycle stages to change exon structure in the deduced coding sequence(CDS). The major alternatively spliced transcript utilized a less favorableacceptor splice site, which shifted codon triplet usage to a different CDSwith a hydrophilic C terminus, changing the canonical type I membraneMAEBL product to a predicted soluble isoform. We found that develop-mental control of the alternative splicing pattern was distinct from thecanonical splicing pattern. Western blot analysis indicated that MAEBLexpression was better correlated with the appearance of the canonicalORF1 transcript. Together these data reveal that RNA metabolism inunicellular eukaryotes like Plasmodium is more sophisticated than believedand may have a significant role regulating gene expression in Plasmodium.
q 2004 Elsevier Ltd. All rights reserved.
Keywords: malaria; Plasmodium; alternative splicing; MAEBL; ligand
*Corresponding authorlsevier Ltd. All rights reserve
y to this work.yang Technologicalnces, 60 Nanyangepartment of Biology,West, Richmond, IN
al membrane antigen-in; DBL, duffybinding antigen-175;rythrocyte bindingscence assay; ORF,
ing author:
Introduction
Malaria is one of themost serious human diseasescausing several million deaths and clinical illness inhundreds of millions of people every year. Thebiological complexity of these pathogenic proto-zoans is extraordinary in the intricate life cycles, asthe malaria parasite must develop in two verydifferent host organisms, infecting different celltypes, and its motile stages must often traversedifferent tissues before infecting a new cell.1 Never-theless, the Plasmodium genome is estimated to haveless than 6000 genes and the number of uniquegenes is still less since a considerable part of thePlasmodium genome is occupied by multi-genefamilies (e.g. var, rif, stevor).2,3 Compared to the
d.

590 RNA Processing in Malaria Parasites
simple unicellular yeast, which is predicted to haveO6000 genes, the estimated size of the Plasmodiumgenome does not seem to adequately reflect theremarkable biological complexity in a malariaparasite’s life cycle.
Developmental processes of malaria parasites aretightly coordinated with gene expression in a “justin time” strategy so that transcript abundancepatterns closely mirror the time in the life cyclewhere products are needed.4–6 Gene expression ineukaryotes occurs at several levels, includinginitiation of transcription, elongation, mRNA pro-cessing, RNA stability, translation, and post-translation processing. Profiling of global geneexpression patterns of Plasmodium falciparumrevealed that the temporal control is fairlyrigid.5–7. Recent evidence indicates that chromatinassembly or remodeling operates as an importantmechanism to control transcription in malariaparasites.8 Cis-acting regulatory motifs are presentin the flanking non-coding regions adjacent to genecoding sequences, although these structures arepoorly defined in Plasmodium.9–11 However, theability to identify such characteristic cis elements asthe TATA box is hindered by the strong AT bias inthe P. falciparum genome. Still core trans elements ofthe transcription machinery, such as TATA-bindingprotein, are clearly present in Plasmodium genome.
In the post-transcriptional phase of gene expres-sion, splicing of precursor mRNA is controlled inthe splicesome by a series of small nuclear ribo-nucleoproteins (snRNP) to remove introns andbring together exon sequences in a very precisemanner. Similar to most other eukaryotes, exonboundaries of Plasmodium are demarcated byconsensus donor and acceptor splicing junctions,GU and AG, respectively.12 The acceptor is oftenpreceded by a poly-pyrimidine tract, although thiscan be veiled somewhat because introns generallyhave elevated AT nucleotide composition relative tothe coding sequence. To date, alternative splicinghas been discovered for only a few Plasmodiumgenes and the complexity of alternative splicing isrelatively limited.13–17 Alternative splicing isimportant in metazoans as a mechanism to createdifferent isoform products from the same gene,which is often developmentally coordinated andoccurs in a tissue-specific manner.18–21 Morerecently appreciated is the role of alternativesplicing as a gene regulatory mechanism targetingmRNA for degradation.22,23 Splicing occurs in thenucleus and the processed mRNA is stabilized thenexported to the cytoplasm in order to be translated.Modulation of mRNA stability can control geneexpression levels and Plasmodium has such amechanism in place as indicated by the extremelydelayed translation of Pbs21, which occurs only inthe zygote and ookinete stages even though themessage is made much earlier in the gametocytestage.24 A mechanism to rapidly decay nonsensemRNA appears to be more poorly developed, sincetranscripts of pseudogenes are present at easilydetectable levels and their abundance is controlled
in developmental patterns similar to homologous oradjacent genes.7,25,26
Plasmodium motile stages are highly specializedfor infecting the next host cell and to do so utilize anapical complex of organelles.27 Proteins destined forinclusion in the apical organelles are expressed onlyduring the final stages of intracellular developmentand represent a cohort of genes whose expression istightly regulated.5,6,28,29 Despite great differences incell types invaded by the different Plasmodiummotile stages, the basic mechanisms of invasionappear similar in the different stages and species.30
The ebl family of erythrocyte-binding proteinsincludes some of the best characterized malarialligands, such as the Plasmodium vivaxDuffy bindingprotein (DBP) and P. falciparum Erythrocyte BindingAntigen-175 (EBA-175).31 Exon structure, includingthe conserved position of splicing junctions withincodons at the exon boundaries, is an importantcharacteristic of ebl genes.32 maebl is a paralogueof the ebl family, similar except that its two extra-cellular ligand domains have identity to AMA1 andnot the consensus DBL ligand domains of other eblproducts.33 maebl evolved independently prior tospeciation along with the ebl and ama1 in theancestral Plasmodium genome.34 Similar to theconsensus ebl products, MAEBL was identified inPlasmodium yoelii and P. falciparum blood-stageparasites as a minor type I transmembrane proteinwith erythrocyte binding activity.33,35,36 Conven-tional views considered that this type of moleculeevolved to have a stage-specific function in thebiology of malaria parasites. Indeed, proteomicanalysis confirmed that very few proteins areexpressed in all stages and determined that alarge percentage of malarial proteins are stage-specific.28,29 Surprisingly, a number of apical orga-nelle and membrane-associated proteins wereexpressed both in sporozoites and merozoites.28
This supports previous observations that ebl andmaebl products are expressed and can have essentialroles in other stages of development.37–39 Therefore,it is apparent that many parasite ligands withdefined functions are more functionally diversethan originally thought.
In our analysis of maebl expression in differentstages of malaria parasite development we dis-covered multiple transcripts. Alternative splicing ofPlasmodium maebl created different ORF in the 3 0
CDS in all species of malaria parasites examined.We found that mRNA processing and alternativesplicing of maebl was developmentally regulated.These data reveal an efficient conserved mechanismfor alternative splicing among malaria parasites tocreate multiple unique gene products from a singlelocus.
Results
Alternative splicing of the maebl 3 0 exons
The canonical structure of maebl has splicing

Figure 1. Schematic gene structure of the Plasmodiummaebl. The consensus gene structure ofmaebl (top) consistsof five exons, which encode a (1) signal peptide, (2)extracellular domain, (3) transmembrane domain, and(4/5) cytoplasmic domain. Arrows beneath the genestructure display the approximate locations of theprimers used for RT-PCR analysis of the splicing of 3 0
exons. Anti-MAEBL serum used here was preparedagainst the N-terminal ligand domain M1.
RNA Processing in Malaria Parasites 591
junctions within codons at the exon/intron bound-aries in the 3 0 exons identical with other members ofthe ebl family.32,40 All ebl family and maebl productsare deduced as type I membrane proteins and the 3 0
exons encode the putative transmembrane andcytoplasmic domains (Figure 1). In studying maebl
gene expression of P. yoelii sporozoites (S) duringmosquito stage development we expected to detecta single transcript, but instead we discoveredmultiple RT-PCR products when amplifying por-tions of the 3 0 CDS of maebl (Figure 2). We extendedthis transcript analysis to other stages of develop-ment, exoerythrocytic forms (EEF) and blood-stages(BS), with similar results.Clones were isolated out of the total RT-PCR
reaction corresponding to each of the three RT-PCRproducts detected from sporozoite RNA. Oneproduct had the expected canonical splicing pat-tern, but two other products represented noveltranscripts created by alternative splicing of themaebl 3 0 exons (Figure 3(A)). Both alternativelyspliced transcripts, referred to here as ORF2 andORF3, encode putative soluble MAEBL isoforms(Figure 3(B)). Internal alternative cis-splicing ofexon 3 generated the ORF2 transcript by using aright splicing junction (AAG) within intron 2. Thisalternative acceptor splice site is 16 nt before thecanonical junction (YAG) of ORF1 and generates aframe shift to create a new C terminus without atransmembrane domain. The remaining splicingjunctions for ORF2 transcripts remain the same as in
Figure 2. (A) Diagram of the lifecycle of malaria parasites identify-ing the three stages of reproduc-tive development: erythrocytic orblood-stage (BS); sexual includinggametocytes, gametes, and oocyst(O); and exoerythrocytic forms(EEF). Sporozoites (S) are the pro-ducts of sexual reproduction on themosquito midgut then invade thesalivary gland to become infectiveto a new vertebrate host. (B) InitialRT-PCR analysis for alternativesplicing of maebl transcripts insporozoites (S), exoerythrocyticforms at 0 hour and 44 hoursduring development, and blood-stage parasites (BS). Reactionswere done with (C) and without(K) reverse transcriptase. Two pro-ducts (O300 nt) are evident in allsamples except 0 hour EEF, plus athird smaller product visible onlyin the S sample. Sequence analysisof these products identified themas ORF2, ORF1, and ORF3 fromlargest to smallest. Purified game-tocytes or gametes were not avail-able for analysis.

Figure 3. (A) Schematic structures show the canonical (ORF1) and alternative splicing of maebl (ORF2, ORF3). Intronsarranged on top at exon boundaries of the main open reading frame (
V) are removed using splicing junctions that are
homologous to splicing junctions of the paralogous ebl, whereas introns arranged below (W) are removed using
alternative splicing junctions. CDSs that match the consensus MAEBL sequence are shaded in black (&), exons withpossible alternative ORFs are shaded as grey ( ) and untranslated ORFs resulting from alternative splicing are shown asstraight lines (—). (B) Deduced sequences are shown for the consensus and alternative ORF at the C-termini of P. yoeliiMAEBL (AF031886). The nucleotide sequence begins near the end of exon 2 and extends through the final 3 0 codingexons as shown in Figure 1. The dominant alternatively spliced transcript is created by use of an acceptor splicingjunction 16 nucleotides before the ORF1 acceptor junction. Inclusion of this extra sequence creates a shift in the ORF usedin exons 3 and 4, switching deduced amino acid residues from hydrophobic to hydrophilic. ORF3 is created by skippingexons 3 and 4 entirely, connecting exons 2 and 5 together with the consensus donor and acceptor junctions of thecanonical splicing pattern. Internal exon boundaries are identified by a mark (O), donor splicing junctions are shaded,and stop codons are boxed.
592 RNA Processing in Malaria Parasites

Table
1.Speciesan
dstag
e-sp
ecificdetectionofalternativelysp
liced30exonsmaebl
P.yoelii
P.falciparum
P.vivax
YM
265B
Y17
XNL
3D7
ORF
SEEF
BS
SEEF
BS
SEEF
BS
SEEF
BS
SEEF
BS
1C
CC
CC
CC
CC
Cna
Cna
na
C2
CC
CC
CC
CC
CC
na
Cna
na
C3
CK
KC
CK
**
*K
na
Kna
na
C
Constitutiveexpressionin
multiple
dev
elopmen
tstag
estypically
isach
aracteristicofahouse-keepinggen
ean
dso
MAEBLexpressed
inmultiple
parasitedev
elopmen
talstag
essu
ggestsauniversal
functionwithoutstag
e-sp
ecificity.T
hisseem
sunlikelyan
dinsteadMAEBLhas
somedistinct
functional
characteristicsin
each
thedifferentinvasivestag
esbecau
seas
asu
rfaceligan
ditisinteracting
withverydifferenthost
cellsthat
presentquitedistinct
arraysofsu
rfacereceptors.MAEBLav
idly
bindserythrocytes,1isessential
forsp
orozo
iteattach
men
tan
dinvasionofsalivaryglands,2an
dis
importan
tforinfectionofhep
atic
stag
es.3
S,salivaryglandsp
orozo
itestag
e;EEF,
liver-stageexoerythrocyticform
s;BS,blood-stagestage.
C,sp
licingpattern
detected;K,nev
erdetected;*,detectedat
leastin
onesample
preparation;na,
samplesnotav
ailable
forRT-PCRan
alysis.
RNA Processing in Malaria Parasites 593
ORF1, but a stop at the second codon in exon 4results in early termination of the CDS O50 ntbefore the next intron. Exon skipping (3 and 4) is theother type of alternative splicing in the maebl 3 0
exons, which creates the ORF3 transcript with exon2 directly spliced to 5. This third splice variantappears to be only a very minor RT-PCR product asviewed by agarose gel electrophoresis. The codontriplet pattern of the ORF is unchanged but absenceof exon 3 results in omission of the transmembranedomain CDS to create a MAEBL isoform of theextracellular domain with only a short additionalpolypeptide sequence added at the C terminus.
Alternative splicing of 3 0 exons occurs inmultiple stages and across Plasmodium species
Each life cycle stage of the malaria parasiterepresents a unique developmental stage withdifferent types of evolutionary adaptations to thedistinct molecular environments of each host andeach cell infected. Since alternative splicing inmetazoans is often associated with expression ofdistinct functional isoforms in different tissues, wewere interested to determine if there is an analo-gous stage-specific alternative splicing of maebl inother life cycle stages of the protozoan parasite. Weobtained RNA from the sporogonic, exoerythrocyticforms, and blood stages of, P. yoelii (Table 1). Weincluded in our analysis genetically and pheno-typically distinct lines of this rodent malariaparasite: YM, 265BY, and 17XNL. RT-PCR detectedidentical patterns of alternative splicing of the 3 0
maebl exons, except ORF3 that was less consistentlydetected. These results indicate that there is no realdifference in the pattern or level of alternativesplicing in different developmental stages.Because the primary deduced amino acid struc-
ture of maebl is highly conserved across evolution-arily divergent Plasmodium species,34 we thoughtthat the alternative splicing might also be con-served. Analysis of maebl in P. falciparum, Plasmo-dium berghei, Plasmodium knowlesi, and P. vivaxrevealed that the intron 2 nucleotide sequenceinvolved in alternative splicing is highly conservedin each of these species (Figure 4). Although thecanonical acceptor site for ORF1 varies in the firstnucleotide position (TAG/CAG), the acceptor siteinvolved in the ORF2 alternative splicing (AAG) isidentical in all genes. Using RNA isolated fromsporogonic and erythrocytic stages, the same splic-ing patterns of ORF1 and ORF2 were identifiedfrom P. falciparum (NF54) as for P. yoelii. Thealternative splicing pattern for ORF3 was notdetected in any of the P. falciparum stages examined;however, P. vivax had all three splicing patternspresent in erythrocytic stages (other stages notavailable).
Alternative splicing is developmentallyregulated during oocyst development
Occurrence of maebl alternative splicing in all of

Figure 4.Alternatively spliced maebl intron 2 and flanking exon boundaries from evolutionarily divergent Plasmodiumspecies. Nucleotides for exons are capitalized and introns are in lower case with the alternatively spliced region shaded.An asterisk beneath the aligned sequences identifies bases conserved in all species. Portions of the internal intronsequences were deleted and the number is indicated (Xn) (pf, P. falciparum AY042084; pb, P. berghei AF31887; py, P. yoeliiAF031886; pk, P. knowlesi; pv, P. vivax AY042083).
594 RNA Processing in Malaria Parasites
the different developmental stages examined indi-cated that it was not tissue-specific, therefore, wewere interested to determine if alternative splicingwas regulated within a developmental cycle.Sporogonic development of the malaria parasitein the mosquito is slow relative to the otherdevelopmental stages in the vertebrate host and ismore amenable to this type of analysis. Afteringesting a malaria-infected blood meal, oocystsattached to the outside of the mosquito midgutrequire two weeks to develop to the motilesporozoite (Figure 2) compared to the !1-day and!2-day development cycles of the blood stages andexoerythrocytic forms, respectively. In addition,midgut sporozoites must then invade the mosquitosalivary glands in order to complete their develop-ment and for the malaria parasite to be capable ofinfecting a new vertebrate host.
We examined developing oocysts and sporo-zoites of P. yoelii, using semi-quantitative RT-PCRmethods to specifically detect each splicing pattern,and identified developmentally coordinated differ-ences in maebl splicing patterns (Figure 5(A)). Lowlevels of the ORF2 alternatively spliced transcriptsappeared quite early in oocyst development andthen was upregulated in the 6.5 days after ingestingan infected blood meal (Figure 5). In contrast, theORF1 canonical splicing pattern was not detecteduntil 6.5 days in oocyst development and then wasupregulated 24 hours later. Expression of otherapical organelle proteins, identified as important forsporozoite invasion processes (CSP, TRAP/SSP2,AMA1) were more similar to the ORF1 expressionpattern (Figure 5(B)). Surprisingly, we found thatthe only ebl homologue identified in the P. yoeliigenome was not actively transcribed in oocysts orsporozoites. PY-235 demonstrated a much brieferperiod of expression only during the final sporo-zoite differentiation phase at the end of oocystdevelopment. Examination of ARF1 as a marker ofthe secretory pathway, which all of these productswould pass through, indicated that this pathwaywas established early in oocyst development andfurther expanded during sporozoite differentiation.
Western blot analysis, using an antiserum to theN-terminal cysteine-rich ligand domain, firstdetected expression of a single full-length MAEBLproduct early on day 7, approximately 12 hoursafter appearance of the ORF1 transcript (Figure 6).This anti-M1 serum should react to both ORF1 and
ORF2 MAEBL products, but only a single proteinband was evident. Previously, we identified theO200 kDa protein as the transmembrane ORF1MAEBL product.41 Given the small difference insize predicted between ORF1 and ORF2 products(z2 kDa) it is unlikely that the two bands would bedistinguishable on a Western blot. To determine ifORF2 is also expressed, antisera was preparedagainst two different synthetic peptides from theshort unique C terminus of the ORF2 product, butboth of these sera were not reactive to parasiteproteins by Western blots (data not shown). Conse-quently, there are no conclusive data regardingpresence or absence of proteins potentially encodedby the alternatively spliced mRNAs. When themidgut sporozoites matured all full-length MAEBLproduct had undergone post-translational process-ing (Figure 6, 14G). After invasion of the salivaryglands MAEBL was again expressed (Figure 6, 14S).Appearance of the more abundant CSP productcorresponded more closely with appearance of itstranscripts early on day 6 of oocyst development. Theanti-CSP monoclonal identified both the precursorand post-translationally processed forms of CSP inthe midgut and salivary gland sporozoites.42,43
Discussion
Alternative splicing in mammals and othermetazoans is important for creating functionaldiversity, particularly in cell adhesion molecules,through developmentally controlled tissue-specificexpression of distinct isoforms.44 The occurrence ofalternatively spliced transcripts for the ligandMAEBL during the different life cycle stagesidentified a mechanism by which a malaria parasitemight similarly expand the functional diversity ofits genome. Generation of possible soluble isoformsof MAEBL alongwith the canonical transmembraneform of MAEBL appeared to match the paradigm inmetazoan organisms of alternative splicing creatingdistinct protein isoforms. Even if limited in scope,alternative splicing of transcripts may be animportant mechanism to expand the functionaldiversity of the apparent limited genetic infor-mation of the Plasmodium genome. Alternativesplicing has been observed for a few otherPlasmodium genes, still only adenylyl cyclase var-iant isoforms are suggested to have possiblefunctional differences.15
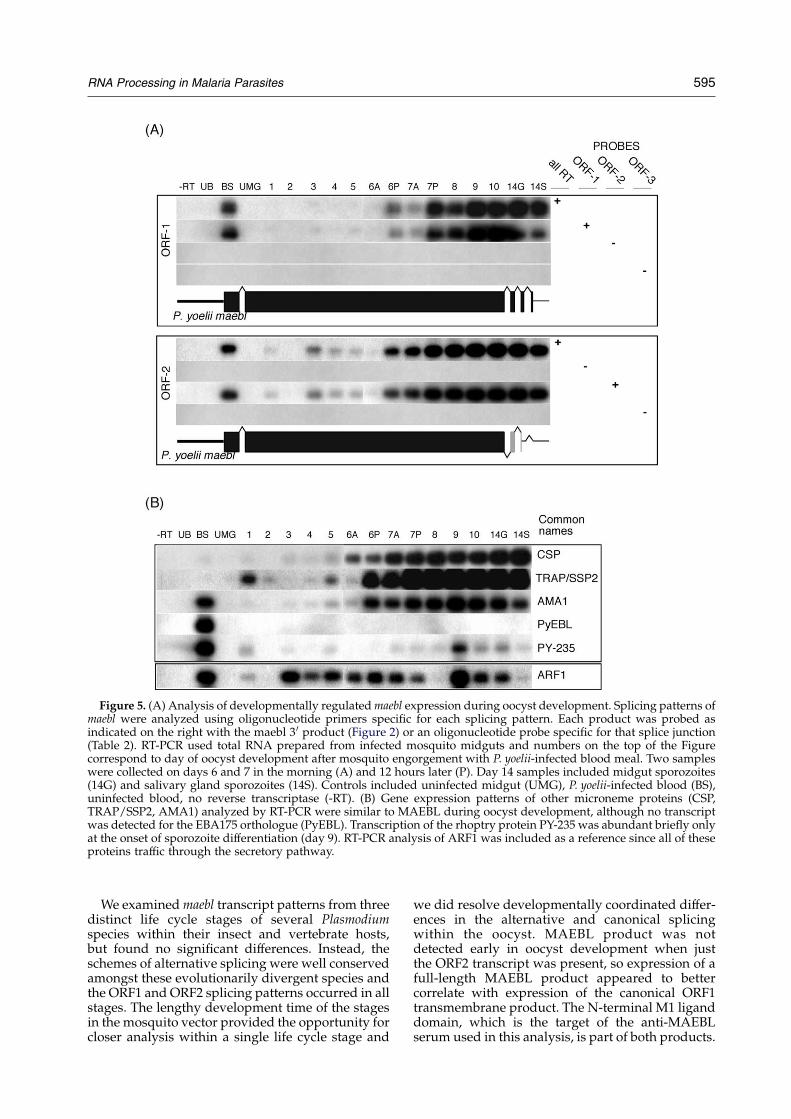
Figure 5. (A) Analysis of developmentally regulatedmaebl expression during oocyst development. Splicing patterns ofmaebl were analyzed using oligonucleotide primers specific for each splicing pattern. Each product was probed asindicated on the right with the maebl 3 0 product (Figure 2) or an oligonucleotide probe specific for that splice junction(Table 2). RT-PCR used total RNA prepared from infected mosquito midguts and numbers on the top of the Figurecorrespond to day of oocyst development after mosquito engorgement with P. yoelii-infected blood meal. Two sampleswere collected on days 6 and 7 in the morning (A) and 12 hours later (P). Day 14 samples included midgut sporozoites(14G) and salivary gland sporozoites (14S). Controls included uninfected midgut (UMG), P. yoelii-infected blood (BS),uninfected blood, no reverse transcriptase (-RT). (B) Gene expression patterns of other microneme proteins (CSP,TRAP/SSP2, AMA1) analyzed by RT-PCR were similar to MAEBL during oocyst development, although no transcriptwas detected for the EBA175 orthologue (PyEBL). Transcription of the rhoptry protein PY-235 was abundant briefly onlyat the onset of sporozoite differentiation (day 9). RT-PCR analysis of ARF1 was included as a reference since all of theseproteins traffic through the secretory pathway.
RNA Processing in Malaria Parasites 595
We examinedmaebl transcript patterns from threedistinct life cycle stages of several Plasmodiumspecies within their insect and vertebrate hosts,but found no significant differences. Instead, theschemes of alternative splicing were well conservedamongst these evolutionarily divergent species andthe ORF1 and ORF2 splicing patterns occurred in allstages. The lengthy development time of the stagesin the mosquito vector provided the opportunity forcloser analysis within a single life cycle stage and
we did resolve developmentally coordinated differ-ences in the alternative and canonical splicingwithin the oocyst. MAEBL product was notdetected early in oocyst development when justthe ORF2 transcript was present, so expression of afull-length MAEBL product appeared to bettercorrelate with expression of the canonical ORF1transmembrane product. The N-terminal M1 liganddomain, which is the target of the anti-MAEBLserum used in this analysis, is part of both products.

Figure 6. (A) Western blot analysis of MAEBLexpression in total protein extracts of midgut oocystsand sporozoites and purified salivary gland sporozoites.Antiserum to the N-terminal M1 ligand domain detecteda O200 kDa protein, corresponding to the full-lengthprecursor protein, was first in the morning sample of day7. The full-length protein was not evident in the maturemidgut sporozoites (14G), but was re-expressed insporozoites after they invaded the mosquito salivaryglands (14S). Uninfected mosquito midgut (UMG) servedas a negative control. (B) Western blot analysis of CSPexpression, using monoclonal antibody NYS1, wasincluded as a positive control for these parasite samples.
596 RNA Processing in Malaria Parasites
In a previous study, we confirmed that this full-length MAEBL product, also expressed in salivarygland sporozoites, is the transmembrane MAEBL.Together these data suggest that alternativelyspliced transcripts may not appear to have import-ance in the expression of different isoforms.
It is suggested that mRNA processing andtranslational control mechanisms play a major rolein regulating protein expression in malaria para-sites, since not many transcriptional regulatoryelements are identified in the P. falciparum gen-ome.45,46 Little is known in Plasmodium abouttranscriptional control of gene expression throughRNA processing,9 although this plays a critical rolein regulating the proteome of higher eukaryoticorganisms.47–49 Suppression of canonical splicingjunctions and exon skipping, like that observed forPlasmodium maebl, are common RNA regulatorymechanisms among multicellular eukaryotic organ-isms.50,51 Several key players in regulated RNAprocessing are readily identifiable in the P. falci-parum genome, including homologues of the SRprotein ASF/SF-2 and the SR-related protein U2AF-65, in accordance with a mechanism of
splicesome assembly on exonic splicing enhancerelements. Separate from creating different isoforms,alternative splicing may also be a mechanismcontrolling Plasmodium gene expression throughintroduction of early termination codons that leadto nonsense mediated decay (NMD) of suchmRNA.In mammals, premature stop codons, those occur-ring O50 nt upstream of the final exon, targettranscripts for decay. The dominant alternativesplicing of maebl does introduce an early stopcodon in the penultimate exon in a pattern con-sistent with NMD. However, surveillance mechan-isms regulating NMD in Plasmodium may be moresimilar to other unicellular eukaryotes like yeast,which utilize other mechanisms.52
Our analysis of maebl transcripts reveals acomplexity of mRNA processing much greaterthan expected for a unicellular eukaryote. Malariaparasites display a high degree of developmentalcontrol of gene expression and yet there is adearth of factors to tightly control transcription.Undoubtedly, this elevates the importance of post-transcriptional control mechanisms to preventtranslation of nonsense proteins or proteins at thewrong stage of development. Identifying how RNAis processed provides important insight into ourgeneral understanding of the gene regulatory net-works of these important unicellular pathogens.
Methods
Parasites
Genomic DNA of P. falciparum (3D7), P. y. yoelii (256 BY,17X NL 1.1, YM lines), and P. vivax (Sal-1) were obtainedby standard extraction protocols.53 P. yoelii 17X wasroutinely maintained in Syrian golden hamsters trans-mitted to three to five day old Anopheles stephensi.54
Mosquitoes weremaintained at 25 8C and 70–80% relativehumidity. The infectivity of mosquitoes was determinedon day 5 post-infective blood meal by counting oocysts indissected midguts stained with mercurochrome andsporozoites on day 14 from salivary glands preparation.The salivary glands and midgut were dissected out frominfected A. stephensi and gently ground in PBS to releaseP. yoelii sporozoites. After the removal of tissue fragmentsby centrifugation at 18g for three minutes, sporozoiteswere collected from the supernatant by centrifugation at5000g for three minutes.
Extraction of RNA and DNA
Infected blood was collected at 10–20% parasitemia,washed in PBS. Leukocytes were removed by filteringblood through Plasmodipur filter (Euro-Diagnostica), andthen the blood pellet resuspended in TRI-reagent (Invi-trogen) for total RNA extraction. Genomic DNA fromblood-stage parasite was extracted by the phenol/chloro-form method. All oocyst RNA samples were collectedfrom infected midguts on day 1, 2, 3, 4, 5, 6/am, 6/pm,7/am, 7/pm, 8, 9, 10 and 14 post-infective bloodmeal andsolubilized in TRI-reagent. Mature sporozoites werecollected on day 14 from infected mosquito salivaryglands and total RNA was extracted with TRI-reagent,according to the manufacturer’s instructions. Total RNA

Table 2. Primer description used in RT-PCR. Each primer pair is listed as sense (S) and antisense (A)
Genes Accession number Primer Primer sequence (5 0-3 0)
maebl 3 0-set 1 AF031886 S ATGCTAAATATAAAACATCACA GAAACAATTATGCTGCAATATCAGATTATT
maebl 3 0-set 2 AF031886 S CAAAGAAGGAATTTTCAGATCCA GCTGTATAAATTTCCGTATG
Pfmaebl 3 0-set 1 AYO42084 S CATTATTAAGTTAATAAACATTGCAAAATTAAGCA ATGTTGTTCCATTTCTGATTTTTGTCT
Pfmaebl 3 0 set 2 AYO42084 S CATTTGATTATGTTTTCTTCCGA GAATTTGCTGATCCTTTATATAG
maebl ORF1 AF031886 S AGTAACGAGGAATATTCAAAAGCA TTGCATAATATTTTATAATTCTTTTAGAATATG
maebl ORF2 AF031886 S AGTAACGAGGAATATTCAATTACA TTGCATAATATTTTATAATTCTTTTAGAATATG
CSP PY03168 S AATGAAGATTCTTATGTCCCAAGCA TACAAATCCTAATGAATTGCTTAC
SSP2 PY03052 S TGGAAAATGGGAAGAATGGAGTGA TGTGGGCAATCACGAACCTTAC
AMA1 PY01581 S AAATTTGGAATCTGGGTTGATGGTA AGTACTAGATAAAGCAGT
EBL PY04764 S TATTATGATTGTATGAATGAAGAGA TACAGGTATATATTCTTGTACACG
Py235 Y11181 S GACGATTTAAAAAATAAAAAACAGGAGA CTATAACTTCATCTTCTCTTTCAAAAAGCG
ARF1 PY04572 S AAAGTCAAATTAGGAGAAGTTGA TGTAGCGCAGGTTGATTGGATG
RNA Processing in Malaria Parasites 597
was also extracted from uninfected blood as well asuninfected midguts for control RT-PCR with the samemethod.
RT-PCR amplification and cloning of maebl
RNA from different stages was extensively treated withDNase I (Amp grade) before use in the reverse-transcrip-tase step, using Superscript (Invitrogen) one-step RT-PCRkit.55 Oligonucleotide primers amplified maebl of P. falci-parum or P. yoelii, at the 3 0-end flanking introns 2–4.Samples from RT-PCR reactions were analyzed byagarose gel electrophoresis. RT-PCR products werepurified, cloned and sequenced as described.7,55
RT-PCR analysis of the maebl 3 0-end of P. yoelii wascarried out with sense (S) and antisense (A) oligonucleo-tide primers in set 1 as indicated in Table 2, usingstandard methods as described.56 Negative samples weresubjected to a nested reaction using set 2. Analysis of theP. falciparum maeblwas identical using Pfmaebl primer set1 and 2. Samples from RT-PCR reactions were analyzedby agarose gel electrophoresis. RT-PCR products werepurified using the QIAquick PCR purification kit (QIA-GEN), cloned and sequenced as described.55
Sequence analysis and Southern blot hybridization
Sequences of cloned DNA were determined by thedideoxy chain termination method. Nucleotide anddeduced amino acid sequences were aligned using theCLUSTAL (MacVector 6.5.3) and adjusted manually. RT-PCR products resolved by agarose gel electrophoresis,denatured and transferred onto positive nylon mem-branes. Blots were hybridized with radiolabeled probes(RT-PCR products or gene-specific oligonucleotide), asindicated in Figure legends, using standard protocols.The oligonucleotide probes for ORF1 was GGAATATTCAAAAGCGGTTTA and ORF2 was GGAATATTCAATTACGTTTTTTT. The hybridization temperature for oligo
probing was 10 8C less than oligo Tm. Hybridized blotswere exposed to X-ray film for variable time periods atK70 8C to obtain clear bands.
Immunoblotting
Infected midguts with oocysts collected on day 5,6/am, 6/pm, 7/am, 7/pm, 8 and 14 as well as maturesalivary gland sporozoites were directly mixed with SDS-PAGE sample buffer for protein analysis by Western blot.Equivalent numbers of uninfected midguts were usedas controls. Western blot analysis was performed withantisera (“C10”) raised against GST fused P. yoeliiMAEBLM1 domain protein in rabbits.4 Anti-CSP (NYS-1)43
murine monoclonal antibody was used as a control.Midgut and sporozoite samples were solubilized in SDS-PAGE sample buffer containing b-mercaptoethanol andheated to 70 8C for five minutes. Equivalent amounts ofuninfected midgut control were also included. Afterelectrophoresis, proteins were electrotransfered ontonitrocellulose membranes (Millipore). Blots were blockedwith 5% non-fat dry milk in PBS-T and washed five timeswith PBS-T. Anti-MAEBL C10 (1:500) and anti-CSP(1:1000) in PBS-T followed by secondary antibody HRP-conjugates were used with an enhanced chemiluminis-cence system (Amersham Biosciences) to detect parasiteantigens.
Acknowledgements
This work was supported by the NationalInstitutes of Health (R01 AI33656) and a BurroughsWellcome Fund travel grant (J.H.A.). Travel (P.R.P.and L.R.) was funded by The Royal Society’s “JointProject under European Science Exchange Pro-gramme” and The British Council’s and French

598 RNA Processing in Malaria Parasites
Ministry of Foreign Affairs “Alliance: Franco-British Partnership Programme”. T.V. held a fellow-ship fromMRT. This work was supported in part byGrants-in-Aid for Scientific Research 13576007(M.T.) from the Ministry of Education, Science,Sports and Culture, Japan. NYS-I was kindlyprovided by Dr Y. Charoenvit.
References
1. Markell, E. K., John, D. T. & Krotoski, W. A. (1998).Medical Parasitology, 8th edit., W. B. Saunders Com-pany, Philadelphia, PA.
2. Gardner, M. J., Tettelin, H., Carucci, D. J., Cummings,L. M., Aravind, L., Koonin, E. V. et al. (1998).Chromosome 2 sequence of the human malariaparasite Plasmodium falciparum. Science, 282, 1126–1132.
3. Bowman, S., Lawson, D., Basham, D., Brown, D.,Chillingworth, T., Churcher, C. M. et al. (1999). Thecomplete nucleotide sequence of chromosome 3 ofPlasmodium falciparum. Nature, 400, 532–538.
4. Noe, A. R., Fishkind, D. J. & Adams, J. H. (2000).Spatial and temporal dynamics of the secretorypathway during differentiation of the Plasmodiumyoelii schizont. Mol. Biochem. Parasitol. 108, 169–185.
5. Bozdech, Z., Llinas, M., Pulliam, B. L., Wong, E. D.,Zhu, J. & Derisi, J. L. (2003). The transcriptome of theintraerythrocytic developmental cycle of Plasmodiumfalciparum. PLoS. Biol. 1, 5.
6. Le Roch, K. G., Zhou, Y., Blair, P. L., Grainger, M.,Moch, J. K., Haynes, J. D. et al. (2003). Discovery ofgene function by expression profiling of the malariaparasite life cycle. Science, 301, 1503–1508.
7. Blair, P. L., Witney, A., Haynes, J. D., Moch, J. K.,Carucci, D. J. & Adams, J. H. (2002). Transcripts ofdevelopmentally regulated Plasmodium falciparumgenes quantified by real-time RT-PCR. Nucl. AcidsRes. 30, 2224–2231.
8. Calderwood, M. S., Gannoun-Zaki, L., Wellems, T. E.& Deitsch, K. W. (2003). Plasmodium falciparum vargenes are regulated by two regions with separatepromoters, one upstream of the coding region and asecond within the intron. J. Biol. Chem. 278, 34125–34132.
9. Horrocks, P., Dechering, K. & Lanzer, M. (1998).Control of gene expression in Plasmodium falciparum.Mol. Biochem. Parasitol. 95, 171–181.
10. van Lin, L. H., Janse, C. J. & Waters, A. P. (2000). Theconserved genome organisation of non-falciparummalaria species: the need to know more. Int.J. Parasitol. 30, 357–370.
11. Crabb, B. S. & Cowman, A. F. (1996). Characterizationof promoters and stable transfection by homologousand nonhomologous recombination in Plasmodiumfalciparum. Proc. Natl Acad. Sci. USA, 93, 7289–7294.
12. Vinkenoog, R., Veldhuisen, B., Speranca, M. A., DelPortillo, H. A., Janse, C. & Waters, A. P. (1995).Comparison of introns in a cdc2-homologous genewithin a number of Plasmodium species. Mol. Biochem.Parasitol. 71, 233–241.
13. Knapp, B., Nau, U., Hundt, E. & Kupper, H. A. (1991).Demonstration of alternative splicing of a pre-mRNAexpressed in the blood stage form of Plasmodiumfalciparum. J. Biol. Chem. 266, 7148–7154.
14. Pace, T., Birago, C., Janse, C. J., Picci, L. & Ponzi, M.(1998). Developmental regulation of a Plasmodium
gene involves the generation of stage-specific 5 0
untranslated sequences. Mol. Biochem. Parasitol. 97,45–53.
15. Muhia, D. K., Swales, C. A., Eckstein-Ludwig, U.,Saran, S., Polley, S. D., Kelly, J. M. et al. (2003). Multiplesplice variants encode a novel adenylyl cyclase ofpossible plastid origin expressed in the sexual stage ofthe malaria parasite Plasmodium falciparum. J. Biol.Chem. 278, 22014–22022.
16. van Lin, L. H., Pace, T., Janse, C. J., Birago, C.,Ramesar, J., Picci, L. et al. (2001). Interspeciesconservation of gene order and intron-exon structurein a genomic locus of high gene density and complex-ity in Plasmodium. Nucl. Acids Res. 29, 2059–2068.
17. Volkman, S. K., Barry, A. E., Lyons, E. J., Nielsen, K. M.,Thomas, S. M., Choi, M. et al. (2001). Recent origin ofPlasmodium falciparum from a single progenitor. Science,293, 482–484.
18. Celotto, A. M. & Graveley, B. R. (2001). Alternativesplicing of the Drosophila Dscam pre-mRNA is bothtemporally and spatially regulated. Genetics, 159, 599–608.
19. Wollerton, M. C., Gooding, C., Robinson, F., Brown,E. C., Jackson, R. J. & Smith, C. W. (2001). Differentialalternative splicing activity of isoforms of polypyri-midine tract binding protein (PTB). RNA, 7, 819–832.
20. Delotto, Y., Smith, C. & Delotto, R. (2001). Multipleisoforms of the Drosophila Spatzle protein areencoded by alternatively spliced maternal mRNAsin the precellular blastoderm embryo.Mol. Gen. Genet.264, 643–652.
21. Smith, C. W. & Valcarcel, J. (2000). Alternative pre-mRNA splicing: the logic of combinatorial control.Trends Biochem. Sci. 25, 381–388.
22. Hillman, R. T., Green, R. E. & Brenner, S. E. (2004). Anunappreciated role for RNA surveillance.Genome Biol.5, R8.
23. Lewis, B. P., Green, R. E. & Brenner, S. E. (2003).Evidence for the widespread coupling of alternativesplicing and nonsense-mediated mRNA decay inhumans. Proc. Natl Acad. Sci. USA, 100, 189–192.
24. Shaw, M. K., Thompson, J. & Sinden, R. E. (1996).Localization of ribosomal RNA and Pbs21-mRNA inthe sexual stages of Plasmodium berghei using electronmicroscope in situ hybridization. Eur. J. Cell Biol. 71,270–276.
25. Triglia, T., Thompson, J. K. & Cowman, A. F. (2001).An EBA175 homologue which is transcribed but nottranslated in erythrocytic stages of Plasmodium falci-parum. Mol. Biochem. Parasitol. 116, 55–63.
26. Taylor, H. M., Triglia, T., Thompson, J., Sajid, M.,Fowler, R., Wickham, M. E. et al. (2001). Plasmodiumfalciparum homologue of the genes for Plasmodiumvivax and Plasmodium yoelii adhesive proteins, whichis transcribed but not translated. Infect. Immun. 69,3635–3645.
27. Preiser, P., Kaviratne, M., Khan, S., Bannister, L. &Jarra, W. (2000). The apical organelles of malariamerozoites: host cell selection, invasion, host immun-ity and immune evasion.Microbes Infect. 2, 1461–1477.
28. Florens, L., Washburn, M. P., Raine, J. D., Anthony,R. M., Grainger, M., Haynes, J. D. et al. (2002). Aproteomic view of the Plasmodium falciparum life cycle.Nature, 419, 520–526.
29. Lasonder, E., Ishihama, Y., Andersen, J. S., Vermunt,A. M., Pain, A., Sauerwein, R. W. et al. (2002). Analysisof the Plasmodium falciparum proteome by high-accuracy mass spectrometry. Nature, 419, 537–542.
30. Dubremetz, J. F., Garcia-Reguet, N., Conseil, V. &

RNA Processing in Malaria Parasites 599
Fourmaux, M. N. (1998). Apical organelles and host-cell invasion by apicomplexa. Int. J. Parasitol. 28, 1007–1013.
31. Adams, J. H., Kaneko, O., Blair, P. L. & Peterson, D. S.(2001). An expanding ebl family of Plasmodiumfalciparum. Trends Parasitol. 17, 297–299.
32. Adams, J. H., Sim, B. K. L., Dolan, S. A., Fang, X.,Kaslow, D. C. & Miller, L. H. (1992). A family oferythrocyte binding proteins of malaria parasites.Proc. Natl Acad. Sci. USA, 89, 7085–7089.
33. Kappe, S. H., Noe, A. R., Fraser, T. S., Blair, P. L. &Adams, J. H. (1998). A family of chimeric erythrocytebinding proteins of malaria parasites. Proc. Natl Acad.Sci. USA, 95, 1230–1235.
34. Michon, P., Stevens, J. R., Kaneko, O. & Adams, J. H.(2002). Evolutionary relationships of conservedcysteine-rich motifs in adhesive molecules of malariaparasites. Mol. Biol. Evol. 19, 1128–1142.
35. Blair, P. L., Kappe, S. H., Maciel, J. E., Balu, B. &Adams, J. H. (2002). Plasmodium falciparum MAEBL isa unique member of the ebl family. Mol. Biochem.Parasitol. 122, 35–44.
36. Ghai, M., Dutta, S., Hall, T., Freilich, D. &Ockenhouse, C. (2002). Identification, expression,and functional characterization of MAEBL, a sporo-zoite and asexual blood stage chimeric erythrocyte-binding protein of Plasmodium falciparum. Mol. Bio-chem. Parasitol. 123, 35.
37. Gruner, A. C., Brahimi, K., Letourneur, F., Renia, L.,Eling, W., Snounou, G. & Druilhe, P. (2001).Expression of the erythrocyte-binding antigen 175 insporozoites and in liver stages of Plasmodium falci-parum. J. Infect. Dis. 184, 892–897.
38. Kappe, S. H., Gardner, M. J., Brown, S. M., Ross, J.,Matuschewski, K., Ribeiro, J. M. et al. (2001). Explor-ing the transcriptome of the malaria sporozoite stage.Proc. Natl Acad. Sci. USA, 98, 9895–9900.
39. Kariu, T., Yuda, M., Yano, K. & Chinzei, Y. (2002).MAEBL is essential for malarial sporozoite infectionof the mosquito salivary gland. J. Expt. Med. 195,1317–1323.
40. Kappe, S. H. I., Noe, A. R., Fraser, T. S., Blair, P. L. &Adams, J. H. (1998). A family of chimeric erythrocytebinding proteins of malaria parasites. Proc. Natl Acad.Sci. USA, 95, 1230–1235.
41. Preiser, P. R., Renia, L., Singh, N., Balu, B., Jarra, W.,Voza, T. et al. (2004). Antibodies against MAEBLligand domains M1 and M2 inhibit sporozoitedevelopment in vitro. Infect. Immun. 72, 3604–3608.
42. Yoshida, N., Potocnjak, P., Nussenzweig, V. &
Nussenzweig, R. S. (1981). Biosynthesis of Pb44, theprotective antigen of sporozoites of Plasmodiumberghei. J. Expt. Med. 154, 1225–1236.
43. Charoenvit, Y., Leef, M. F., Yuan, L. F., Sedegah, M. &Beaudoin, R. L. (1987). Characterization of Plasmo-dium yoelii monoclonal antibodies directed againststage-specific sporozoite antigens. Infect. Immun. 55,604–608.
44. Rowen, L., Young, J., Birditt, B., Kaur, A., Madan, A.,Philipps, D. L. et al. (2002). Analysis of the humanneurexin genes: alternative splicing and the gener-ation of protein diversity. Genomics, 79, 587–597.
45. Gardner, M. J., Hall, N., Fung, E., White, O., Berriman,M., Hyman, R. W. et al. (2002). Genome sequence ofthe human malaria parasite Plasmodium falciparum.Nature, 419, 498–511.
46. Wirth, D. F. (2002). Biological revelations. Nature, 419,495–496.
47. Modrek, B. & Lee, C. (2002). A genomic view ofalternative splicing. Nature Genet. 30, 13–19.
48. Harrison, P. M., Kumar, A., Lang, N., Snyder, M. &Gerstein, M. (2002). A question of size: the eukaryoticproteome and the problems in defining it. Nucl. AcidsRes. 30, 1083–1090.
49. Graveley, B. R. (2001). Alternative splicing: increasingdiversity in the proteomic world. Trends Genet. 17,100–107.
50. Caceres, J. F. & Kornblihtt, A. R. (2002). Alternativesplicing: multiple control mechanisms and involve-ment in human disease. Trends Genet. 18, 186–193.
51. Brett, D., Pospisil, H., Valcarcel, J., Reich, J. & Bork, P.(2002). Alternative splicing and genome complexity.Nature Genet. 30, 29–30.
52. Kebaara, B., Nazarenus, T., Taylor, R., Forch, A. &Atkin, A. L. (2003). The Upf-dependent decay of wild-type PPR1 mRNA depends on its 5 0-UTR and first 92ORF nucleotides. Nucl. Acids Res. 31, 3157–3165.
53. Schlichtherle, M., Wahlgren, M., Perlmann, H. &Scherf, A. (2000).Methods inMalaria Research, 3rd edit.,MR4/ATCC.
54. Singh, N. & Puri, S. K. (1998). Causal prophylacticactivity of antihistaminic agents against Plasmodiumyoelii nigeriensis infection in Swiss mice. Acta Trop. 69,255–260.
55. Preiser, P. R. & Jarra, W. (1998). Plasmodium yoelii:differences in the transcription of the 235-kDa rhoptryprotein multigene family in lethal and nonlethal lines.Expt. Parasitol. 89, 50–57.
56. Preiser, P. R., Jarra, W., Capiod, T. & Snounou, G.(1999). A rhoptry-protein-associated mechanism ofclonal phenotypic variation in rodent malaria. Nature,398, 618–622.
Edited by J. Karn
(Received 13 July 2004; received in revised form 11 August 2004; accepted 13 August 2004)


















