
Conformational flexibility in a small globular hormone: X-ray analysis of avian pancreatic...
12
Conformational Flexibility in a Small Globular Hormone: X-Ray Analysis of Avian Pancreatic Polypeptide at 0.98-A Resolution IAN GLOVER, ILLYAS HANEEF, JIM PITTS, STEVE WOOD, DAVID MOSS, IAN TICKLE, and TOM BLUNDELL, Laboratory of Molecular Biology, Department of Crystallography, Birkbeck College, University of London, London WClE 7HX, United Kingdom Synopsis The 36-amino acid avian pancreatic polypeptide has been studied by x-ray analysis at 0.98-A resolution and refined using a restrained least-squares technique to an agreement factor of 15.6%. The polypeptide, which has a compact globular structure with a hydrophobic core, comprises a polyproline-like helix (residues 2-8) and an a-helix (residues 14-32). The molecule forms symmetrical dimers linked through zinc atoms in the crystal lattice. The high-resolution analysis defines sequence-dependent distortions in the a-helical parameters due to hydrogen bonding of water molecules and side chains. The thermal parameters indicate an increased flexibility of the main chain at the turn between the helices and in the C-terminal residues. For the first time, six-parameter anisotropic thermal ellipsoids have been refined for each atom; these define the directions of the molecular motions in the polypeptide, indi- cating concerted vibrations. The physiological roles of conformation, flexibility, and dynamics of this polypeptide hormone are discussed. INTRODUCTION Pancreatic polypeptide is a 36-amino acid hormone (Fig. 1) synthesized in the endocrine pancreas as part of a larger precurs0r.l The molecule has a wide range of physiological activities (for review, see Ref. 2), including effects on gut motility and gastrin-like actions3 Injection into hypergly- cemic oblob mice appears to decrease food intake and lead to a loss of body weight4; it may therefore act as a satiety factor. Homologous peptides have recently been identified in the upper intestinal tissue5 and in the braih,6 indicating the existence of a family of related hormones and neuro- transmitters. The three-dimensional structure of avian pancreatic polypeptide (aPP) has been defined by x-ray analysis at 1.4-w res~lution.~ The molecule has a globular structure with residues 2-8 forming a polyproline 11-like helix that is closely packed through hydrophobic interactions against an a-helix formed from residues 14-32. Recent CD experimentss show that these interactions are maintained by the monomer in solution (J. E. Pitts, I. D. Glover, W. Strassburger, and A. Wollmer, unpublished results, 1982),unlike those of crystalline pancreatic glucagon, which appears to have a flexible Biopolymers, Vol. 22, 293-304 (1983) 0 1983 John Wiley & Sons, Inc. CCC 0006-3525/83/010293-12$02.20
-
Upload
ian-glover -
Category
Documents
-
view
214 -
download
0
Transcript of Conformational flexibility in a small globular hormone: X-ray analysis of avian pancreatic...

Conformational Flexibility in a Small Globular Hormone: X-Ray Analysis of Avian Pancreatic
Polypeptide at 0.98-A Resolution
IAN GLOVER, ILLYAS HANEEF, JIM PITTS, STEVE WOOD, DAVID MOSS, IAN TICKLE, and TOM BLUNDELL, Laboratory of Molecular Biology, Department of Crystallography, Birkbeck College,
University of London, London W C l E 7 H X , United Kingdom
Synopsis
The 36-amino acid avian pancreatic polypeptide has been studied by x-ray analysis a t 0.98-A resolution and refined using a restrained least-squares technique to an agreement factor of 15.6%. The polypeptide, which has a compact globular structure with a hydrophobic core, comprises a polyproline-like helix (residues 2-8) and an a-helix (residues 14-32). The molecule forms symmetrical dimers linked through zinc atoms in the crystal lattice. The high-resolution analysis defines sequence-dependent distortions in the a-helical parameters due to hydrogen bonding of water molecules and side chains. The thermal parameters indicate an increased flexibility of the main chain at the turn between the helices and in the C-terminal residues. For the first time, six-parameter anisotropic thermal ellipsoids have been refined for each atom; these define the directions of the molecular motions in the polypeptide, indi- cating concerted vibrations. The physiological roles of conformation, flexibility, and dynamics of this polypeptide hormone are discussed.
INTRODUCTION
Pancreatic polypeptide is a 36-amino acid hormone (Fig. 1) synthesized in the endocrine pancreas as part of a larger precurs0r.l The molecule has a wide range of physiological activities (for review, see Ref. 2), including effects on gut motility and gastrin-like actions3 Injection into hypergly- cemic oblob mice appears to decrease food intake and lead to a loss of body weight4; it may therefore act as a satiety factor. Homologous peptides have recently been identified in the upper intestinal tissue5 and in the braih,6 indicating the existence of a family of related hormones and neuro- transmitters.
The three-dimensional structure of avian pancreatic polypeptide (aPP) has been defined by x-ray analysis a t 1.4-w res~lu t ion .~ The molecule has a globular structure with residues 2-8 forming a polyproline 11-like helix that is closely packed through hydrophobic interactions against an a-helix formed from residues 14-32. Recent CD experimentss show that these interactions are maintained by the monomer in solution (J. E. Pitts, I. D. Glover, W. Strassburger, and A. Wollmer, unpublished results, 1982), unlike those of crystalline pancreatic glucagon, which appears to have a flexible
Biopolymers, Vol. 22, 293-304 (1983) 0 1983 John Wiley & Sons, Inc. CCC 0006-3525/83/010293-12$02.20

1 5
10
15
20
25
30
35
NP
Y
-__
__
Y
PS
KP
DN
P~
~D
~~
AE
DL
AR
YV
SA
CR
HY
~N
L I
TR
QR
Y~
PY
Y
--
Y P
AK
PE
A P
G E
D AS P
EE
L S
R Y Y A
SLRHxLHCV T
R Q
R xa
AP
L E
PV
YP
GD
DA
T P
~O
M&
o~
A
A E
AE
RV
LN
M L
zE
PE
ya
P
PP
BP
P
AP
L E
PE
YP
GD
NA
TP
EO
MA
QY
AA
EL
ER
IL
EM
LK
EP
EE
a
Fy D
DL
Q
Ox
L nv
VL
RH
RE
a
AP
P
Gp
SO
PT
Y EGO D
A P
V
I
Fig.
1.
Sequ
ence
s of p
ancr
eatic
pol
ypep
tides
fro
m p
orci
ne, P
PP
(R
ef. 1
8); b
ovin
e, B
PP (R
ef. 1
8); a
nd a
vian
, APP
(Ref
. 17)
sour
ces a
nd re
late
d pe
ptid
es
NPY
fro
m b
rain
(R
ef. 6
) and
PY
Y fr
om u
pper
inte
stin
al ti
ssue
(Ref
. 5).
T
he o
ne-l
ette
r cod
e us
ed is
A =
Ala
, D =
Asp
, E =
Glu
, F =
Phe
, G =
Gly
, H =
His
, I =
Ile,
K =
Lys
, L =
Leu
, M =
Met
, N =
Asn
, P =
Pro
, Q =
Gln
, R =
Arg
, S =
Ser
, T =
Thr
, V =
Val
, W =
Trp
, Y =
Tyr
. T
he su
pers
crip
t “a”
den
otes
a c-
te
rmin
al a
mid
e gro
up.

CONFORMATION OF SMALL GLOBULAR HORMONE 295
structure in dilute aqueous solution.9 Nevertheless, the C-terminal resi- dues (35-36) are relatively flexible in aPP even in the crystals. aPP forms symmetrical dimers mainly through hydrophobic interactions; and in the crystals, aPP dimers are cross-linked in the lattice by zinc ions, each of which coordinate to the ligands of three polypeptides.
We have recently extended the resolution of our x-ray analysis to 0.98-w resolution. This allows refinement with individual anisotropic thermal factors and a precise definition of bond lengths and angles. In this paper we discuss the conformational flexibility with particular attention to se- quence-dependant deviations from perfect helix structure and to variations in the anisotropic thermal motions that may be correlated with both sec- ondary and tertiary interactions.
EXPERIMENTAL
Purification and crystallization were carried out as described before.1° Crystals were of space group C2, with dimensions a = 34.18 A, b = 32.92 A, c = 28.45 A, and 0 = 105.26'. Data were collected from six crystals using a four-circle diffractometer using a helium path and the scan technique of Tickle." We collected 53,000 reflections, merging them to give 17,058 independent reflections with an agreement value, RM = 6.0%. The model, refined to 21% a t 1.4-A resolution, was used as a starting point for the re- strained least-squares refinement a t 0.98-A resolution using the RESTRAIN program of Morffew and Moss.12 Bond lengths, bond angles, and planarity of aromatic rings and peptides were restrained to target values defined from analyses of smaller peptides and amino acids. The planarity was main- tained by minimizing the square of the variance-covariance matrix of the atoms in the plane. Anisotropic thermal ellipsoids were defined by six parameters. The Gauss-Seidel iterative method was used for the solution of the normal equations. A t present, hydrogen atoms have not been in- cluded. There were 300 protein Zn, C, N, and 0 atoms and 60 water mol- ecules included in the refinement, each defined by three positional and six anisotropic thermal parameters. All 17,058 data points in the 10-0.98 A resolution range and 800 restraints were used, giving an observab1es:pa- rameters ratio of -4:l. The program was run on a Science and Engineering Research Council Cray 1 computer, taking 40 s per cycle with isotropic and 70 s per cycle with anisotropic thermal parameters. The course of the re- finement is shown in Fig. 2. The initial model gave an R factor of 29% and the present R value is 15.6%.
RESULTS AND DISCUSSION
Figure 3 shows a stereo view of the aPP protomer. The torsional angles for the residues of the polyproline-like helix are shown in Table I. The mean values of 4 = -72(8)O and 1c, = 140(14)O are closest to those of poly-
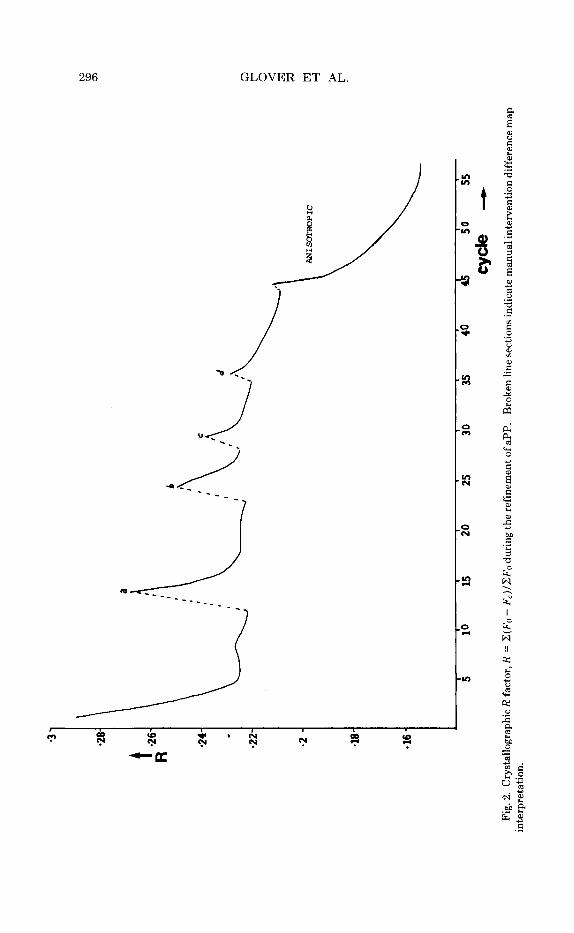
63 *28
-26
R *2
4
.22 *2
-18 - 16
I I
I I
I 4!i
I
5 10
l5
20
25
30
35
40
50
55
cy
cle -
Fig.
2.
Cry
stal
logr
aphi
c R
fact
or, R
= Z(F0 -
F,)
/ZFo
dur
ing
the
refi
nem
ent o
f aP
P.
Bro
ken
line
sect
ions
indi
cate
man
ual i
nter
vent
ion
diff
eren
ce m
ap
inte
rpre
tati
on.
s 0
C M z M 4

CONFORMATION OF SMALL GLOBULAR HORMONE 297
Fig. 3. Stereo view of aPP monomer, showing the polyproline and a-helical segments of the molecule. The C-terminal section is oriented away from the body of the molecule.
proline 11,13 but there is a wide distribution. The a-helix of residues 14-32 is relatively more regular with respect to helical parameters with 4 and I) as - 6 4 ( 5 ) O and -41(4)', respectively (Table 11). These values are close to those reported for other proteins refined at a fairly high resolution, as
TABLE I Main-Chain Torsional Angles G, 6 of the Polyproline Helix of aPP Compared with
Collagen and Polvwoline Helices
Peptide 6 (deg) G (degi
aPPa Pro 2 -66 150
Gln 4 -68 126 Pro 5 -63 150 Thr 6 -85 127
Pro 8 -81
Ser 3 -74 158
Tyr 7 -64 128
Mean -72(8) 140(14)
Polyproline I1
Collagen a b
I
C
-78 -83 -51 -76 -45
149 158 153 127 148
a Resolution: 0.98 8.

298 GLOVER ET AL.
TABLE I1 Main-Chain Torsional Angles $, qb for a P P compared with Actinidin and Erythrocruorin
Peptide qb (deg) $ (deg)
aPPa 14 - 54 -36 15 -61 -41 16 -73 -34 17 -65 -38 18 -62 -45 19 -62 -41 20 -68 -42 21 -55 -48 22 -65 -39 23 -70 -35 24 -69 -40 25 -57 -47 26 -63 -44 27 - 60 -49 28 -63 -39 29 -64 -36 30 -71 -44 31 -63 -37
Mean -64(5) -41(4) Actinidinb -63 -40 ErythrocruorinC -64 -41
a Resolution, 0.98 A. From Ref. 15 From Ref. 16.
N18-014-Cl4 = 164'
(a) (b)
Fig. 4. (a) Hydrogen-bonding geometry a t Asp 23 carbonyl group, showing hydrogen- bonding distances and angle in this exposed a-helical region compared to (b) hydrogen bonding in a hydrophobic region.

CONFORMATION OF SMALL GLOBULAR HORMONE
- -
-50-
-48-
- 46-
i -44-
-42-
-40-
-38-
-36-
-34-
-50
299
x2?
x 21 25
Xl8
0 26
x20
6 15 24 x 28
0 2 2 'x 17
x w 0" 0%
I 1 1 I I I I I I I 1 -52 -54 -56 -58 -60 -62 -64 -66 -68 -70 -72 -74
shown in Table 11. These results appear to indicate that the values ac- cepted for the classical a-helix, $J = -57" and + = -47",14 are not a good description of the a-helix found in globular proteins. The geometry of the a-helix found here leads to an orientation of the C-0 vector of the peptide away from the direction of the helix axis.
It appears that this distortion is not regular. The helix is amphipathic, having one hydrophilic side exposed to the aqueous environment. Most carbonyl oxygens on this side are also hydrogen-bonded to a water molecule (residues 15, 16, 19, 22, 23, and 26) or a hydrophilic side chain-see, for example, Fig. 4. There is little apparent effect on the 4L, GL angles of any single residue, but it can be seen in the two torsion angles defining the peptide orientation rc/l and $Jlfl, as shown in Fig. 5. The effect of hydrogen bonding appears to be to make +L less negative, whereas $ 1 + 1 is more neg- ative. Mean values of GG, &+I, are -39", -68" for hydrogen-bonded groups and -44O, -61" for groups lying against the hydrophobic interior.

300
% 0.8-
0.7-
0.6-
0 5 -
0.4 -
GLOVER ET AL.
POCYPROLINE HELIX
Angle (9 h 0 Ci- 0,- N,.+ 140 15
142
e 2 i
fa *17
154
1%
158 '25 8 26
x n x
28 %9 30 3.1
distance (n) 0,. Ni++
Helix 14 32 aPP
Fig. 6. Plot of distance against hydrogen bond angle for the a-helical section of aPP. Hydrophilic residues, whose carbonyls are hydrogen-bonded to a water molecule or side chain, are circled and show greater distortions from C, - 0, - Ht+4 of 180' and longer bond lengths in comparison with those in a hydrophobic environment.
The same hydrogen bonding can be seen in the geometry at the carbonyl groups. Figure 6 shows that the distance Oi - Ni+4 is increased and the angle at the oxygen (Ci - Oi - Ni+4) is further from 180" than for peptides in a hydrophobic environment. These small distortions show a regular
X
O(-HELIX
n
0.1
1 5 0 1 ~ 1 , U I . . . I , . , I , . , I . . , I , . r
GPSAPT YPGDDAPVAOL I R FYDNLOAVLNVVTRHR
Fig. 7. Individual isotropic thermal parameters plotted as a function of chain sequence. C'" atom thermal parameters are plotted as m; side-chain atoms, X ( C T ) and 0 (Cr).
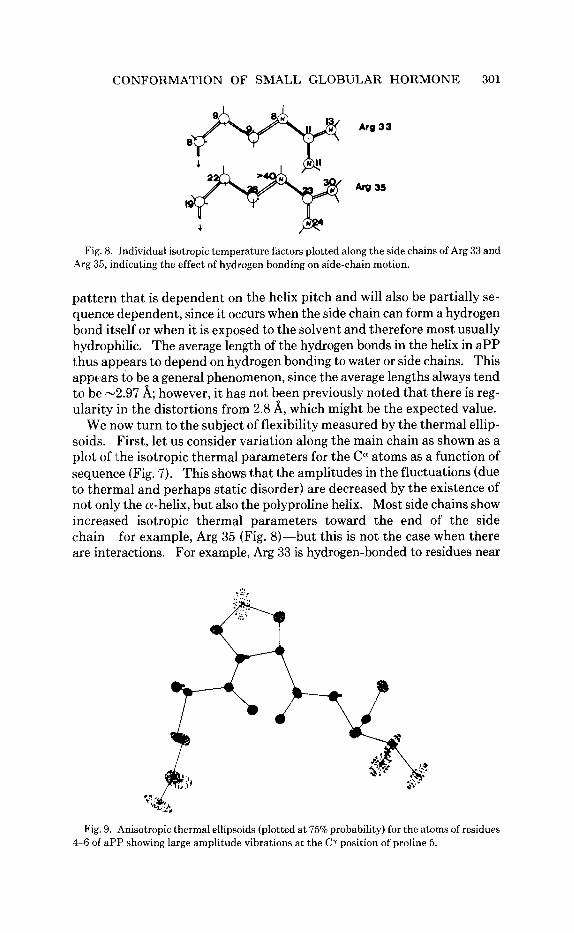
CONFORMATION OF SMALL GLOBULAR HORMONE 301
Arg 33
Fig. 8. Individual isotropic temperature factors plotted along the side chains of Arg 33 and Arg 35, indicating the effect of hydrogen bonding on side-chain motion.
pattern that is dependent on the helix pitch and will also be partially se- quence dependent, since it occurs when the side chain can form a hydrogen bond itself or when it is exposed to the solvent and therefore most usually hydrophilic. The average length of the hydrogen bonds in the helix in aPP thus appears to depend on hydrogen bonding to water or side chains. This appears to be a general phenomenon, since the average lengths always tend to be -2.97 A; however, it has not been previously noted that there is reg- ularity in the distortions from 2.8 A, which might be the expected value.
We now turn to the subject of flexibility measured by the thermal ellip- soids. First, let us consider variation along the main chain as shown as a plot of ithe isotropic thermal parameters for the Ccy atoms as a function of sequence (Fig. 7). This shows that the amplitudes in the fluctuations (due to thermal and perhaps static disorder) are decreased by the existence of not only the a-helix, but also the polyproline helix. Most side chains show increased isotropic thermal parameters toward the end of the side chain-for example, Arg 35 (Fig. 8)-but this is not the case when there are interactions. For example, Arg 33 is hydrogen-bonded to residues near
> :::! .. ... ,
Fig. 9. Anisotropic thermal ellipsoids (plotted a t 75% probability) for the atoms of residues 4-6 of aPP showing large amplitude vibrations a t the Cr position of proline 5.

302 GLOVER ET AL.
a Fig. 10. Individual anisotropic thermal ellipsoids plotted at residues Phe 20, Tyr 21 showing
some evidence of concerted motion perpendicular to the ring planes.
the N-terminus (Gln 4, Pro 5) and has decreased thermal parameters a t the guanidinium group. A similar effect is seen in Asp 23, which is coor- dinated to a zinc ion.
The prolines a t positions 2 , 5 , and 8 have large anisotropic vibrations at the C' atom, as shown, for example, in Fig. 9. It appears that the Cr may be flipping between two different puckered positions with the atom above or below the plane defined by CP, C", N, and C6. If so, the density distri- bution will not be properly described by a thermal ellipsoid and might show two peaks of electron density in a more detailed analysis.
The 07 of Ser 3 also has two orientations. This appears to result from the juxtaposition of two serine residues in dimer formation, giving rise to two possible orientations and a local loss of twofold symmetry. The side
Fig. 11. Individual anisotropic thermal ellipsoids plotted a t residues 7-9 indicating some correlation in the amplitudes of vibration of the side chain of tyrosine 7 , which stacks above the peptide plane of glycine 9.

CONFORMATION OF SMALL GLOBULAR HORMONE 303
chain of residue 15 also appears to be disordered in at least two orientations. Finally, the C-terminus is also disordered, evidenced by difference electron density maps where this residue has been omitted from the calculation and that appear to indicate two orientations for the side chains.
Examples of anisotropic vibrations in the aromatic rings of tyrosine and phenylalanine side chains are shown in Fig. 10. These often appear to oscillate in directions perpendicular to the rings, as in Tyr 21. An inter- esting concerted motion appears to occur in Tyr 7 and the peptide of residue 9 against which it is stacked (Fig. 11).
CONCLUSIONS
The molecule of avian pancreatic polypeptide exhibits considerable flexibility, particularly in the C-terminal region and some acidic side chains (Asp 15, Glu 26), and in the &turn region, the amplitudes of motion within the helical segments being considerably lower. aPP does, however, maintain, economically and simply, a compact tertiary structure both in the monomer and dimeric state, brought about by the packing of hydro- phobic surfaces to form a stable globular core. The maintenance of this integral three-dimensional structure is probably essential for correct binding at the hormone receptor (although this has not yet been charac- terized) by the presentation of a complementary surface. This is likely to be mediated through recognition of and binding to the largely hydrophobic surface involved in dimerization. The apparent flexibility of the C-ter- minal region may also be important and play a role in the triggering or ac- tivation of a biological response in a separate binding event at the re- ceptor.
The extent of the crystallographic data permits the inclusion of aniso- tropic temperature factors into a refinement procedure. This more realistic representation of atomic motion allows an interpretation of both extent and direction of motion. An analysis of these parameters gives some evi- dence of concerted motion within the molecule. This is apparent in two classes: first, aromatic residue side chains appear to vibrate perpendicular to the ring plane, although this is not the case in prolyl residues where flipping between two different puckering states may be taking place; second, in regions of close packing within the core of the molecule, amplitudes and directions of motion do appear to have some correlation.
Small distortions from ideal geometry are apparent in the molecule. These are mainly confined to the exposed, hydrophilic regions of the mol- ecule and are due to the effect of solvent molecules. The surrounding water molecules act in hydrogen bonding both to side chains and also to carbonyl atoms along the helix backbone. The effect of water participating in the helix hydrogen bonding alters both the hydrogen-bonding distance and geometry a t the carbonyl atom. This distortion is reflected clearly in the main-chain torsional angles, introducing a partially sequence-dependent distortion of geometry in the a-helical section of aPP.

304 GLOVER ET AL.
These effects will also be present in the solution state, where a t physio- logical concentrations aPP is likely to be monomeric, but a compact and stable globular structure is maintained despite the small size of the protein, indicating the importance of the hydrophobic interactions in the core of the molecule, offsetting the tendency of water to distort and possibly de- stabilize the helical regions.
A full account of the refinement method and water structure will be published on completion of the refinement.
We are grateful to many members of the Department, particularly Dr. W. C. Wu, for help and assistance during the course of this work. The work was supported financially by the Science and Engineering Research Council.
References
1. Schwarz, T. W. & Tager, H. S. (1981) Nature 294,589-591. 2. Lonovics, J., Devitt, P., Watson, L. C., Rayford., P. L. & Thompson, J. C. (1981) Arc.
3. Lim, T-M. & Chance, R. E. (1974) in Endocrinology ofthe Gut, Chey, W. Y. &Brooks,
4. Gates, R. J. & Lazarus, N. R. (1977) Horm. Res. 8,189-202. 5. Tatemoto, K. & Mutt, V. (1980) Nature 285,417-418. 6. Tatemoto, K. (1982) Proc. Natl. Acad. Sci. USA (in press). 7. Blundell, T. L., Pitts, J. E., Tickle, I. J., Wood, S. P. & Wu, C-W. (1981) Proc. Natl.
8. Strassburger, W., Glatter, U., Wollmer, A,, Fleischhauer, J., Mercola, D. A., Blundell,
9. Boesch, C., Bundi, A,, Oppliger, M. & Wuthrich, K. (1978) Eur. J . Biochem. 91,209-
10. Wood, S. P., Pitts, J. E., Blundell, T. L., Tickle, I. J . & Jenkins, J . A. (1977) Eur. J .
11. Tickle, I. J. (1975) Acta Crystallogr., Sect. B 31,329-333. 12. Morffew, A. & Moss, D. S. (1982) Comput. Chem. 6,l-3. 13. Ramachandran, G. N. & Sasisekharan, V. (1968) Adu. Protein Chem. 23,283-437. 14. Amott, S. & Dover, S. D. (1967) J . Mol. Eiol. 30,209-212. 15. Baker, E. N. (1980) J. Mol. Biol. 141,441-484. 16. Steigemann, W. & Weber, E. (1979) J . Mol. E d . 127,309-338. 17. Kimmel, J . R., Hayden, L. J. & Pollock, H. G. (1975) J. Eiol. Chem. 250,9369-9376. 18. Floyd, J . C., Fayans, S. S., Pek, S. & Chance, R. E. (1977) Rec. Prog. Horm. Res. 33,
Surg. 116,1256-1264.
P. F., Eds., Charles B. Slack, New Jersey, pp. 143-145.
Acad. Sci. USA 78,4175-4179.
T . L., Glover, I., Pitts, J . E., Tickle, I. J. & Wood, S. P. (1982) FEES Lett. 139,295-299.
211.
Biochem. 78,119-126.
519-570.
Received June 20,1982 Accepted September 14,1982


















