Composición, riqueza de especies y abundancia de … · 222 DE LA VEGA & GREZ INTRODUCCIÓN La...
18
Revista Chilena de Historia Natural 81: 221-238, 2008 Composición, riqueza de especies y abundancia de insectos defoliadores de actividad nocturna asociados a Aristotelia chilensis (maqui) en el bosque maulino fragmentado Composition, species richness and abundance of nocturnal folivorous insects associated with Aristotelia chilensis (maqui) in the fragmented Maulino forest XAVIERA DE LA VEGA 1 & AUDREY A. GREZ 2 * 1 Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile. 16 Casilla 653, Santiago, Chile 2 Departamento de Ciencias Biológicas Animales, Facultad Ciencias Veterinarias y Pecuarias, Universidad de Chile, Casilla 2, Correo 15, La Granja, Santiago, Chile; *e-mail para correspondencia: [email protected] RESUMEN En el bosque maulino, la herbivoría sobre Aristotelia chilensis (maqui) es negativamente afectada por la fragmentación del bosque, siendo mayor en el bosque continuo que en los fragmentos, particularmente a inicios de la temporada de crecimiento. Este fenómeno puede deberse a cambios en la dinámica de las poblaciones de defoliadores, esencialmente insectos. En este trabajo se evaluó la abundancia, riqueza de especies y composición de insectos defoliadores de actividad nocturna presentes en A. chilensis en un bosque continuo (600 ha) y en ocho fragmentos remanentes (0,4-20 ha). Los muestreos se realizaron mensualmente, entre agosto de 2005 y febrero de 2006, en 32 ejemplares adultos de A. chilensis en el bosque continuo y en 32 ejemplares en los fragmentos. Los insectos fueron muestreados durante las cinco primeras horas de la noche. Se recolectaron 890 insectos defoliadores, pertenecientes a 17 familias y 77 especies pertenecientes a los órdenes Coleoptera, Orthoptera y Lepidoptera, siendo todas nativas. La abundancia total no varió según el hábitat. Sin embargo, la fragmentación incrementó o disminuyó la abundancia de algunas especies. La riqueza de especies por árbol tampoco fue afectada por la fragmentación del bosque, aunque el número total de especies fue considerablemente mayor en los fragmentos que en el bosque continuo. La similitud de especies fue mayor dentro del bosque continuo que entre el bosque continuo y los fragmentos o que entre los fragmentos. A principios de la temporada de crecimiento de A. chilensis (septiembre), la abundancia de Sericoides obesa fue significativamente mayor en el bosque continuo que en los fragmentos. Al avanzar en la temporada, Sericoides viridis se hizo más abundante en los fragmentos. Por el tamaño y la voracidad de los insectos del género Sericoides ellos serían los principales responsables de los patrones de defoliación de A. chilensis en el bosque maulino. Palabras clave: insectos defoliadores, bosque maulino, fragmentación del hábitat. ABSTRACT At the Maulino forest, herbivory upon Aristotelia chilensis (maqui) is negatively affected by forest fragmentation, being higher in the continuous forest than in the small remnant fragments, particularly at the beginning of the growing season (spring). This phenomenon may be attributable to changes in the population dynamics of herbivores, mostly insects. In this work, the effect of the Maulino forest fragmentation on the abundance, species richness and composition of nocturnal defoliating insects associated with A. chilensis was evaluated. The insects associated with A. chilensis were sampled monthly, between August 2005 and February 2006, in 32 trees located in a continuous forest (600 ha) and in 32 trees located in eight forest fragments (0.4-20 ha). Insect sampling was carried out during the first five night hours. 890 folivorous insects from 17 families and 77 species of Coleoptera, Orthoptera and Lepidoptera were collected. All species were native. The total abundance did not vary with habitat fragmentation. Nevertheless, some species increased their abundance with fragmentation, while others were depressed. The species richness of folivorous insects per tree did not vary with forest fragmentation, though the total number of species was considerably higher in the forest fragments. The species similarity was higher within the continuous forest than between the continuous forest and fragments or among fragments. At the beginning of the season (September), the abundance of Sericoides obesa was significantly higher in the continuous forest than in the forest fragments. Later in the season, Sericoides viridis became more abundant in the forest fragments. Because of the body size and the voracity of Sericoides sp., these insects seem to be responsible of the herbivory patterns of A. chilensis at the Maulino forest. Key words: folivorous insects, Maulino forest, habitat fragmentation.
Transcript of Composición, riqueza de especies y abundancia de … · 222 DE LA VEGA & GREZ INTRODUCCIÓN La...

221INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADORevista Chilena de Historia Natural81: 221-238, 2008
Composición, riqueza de especies y abundancia de insectos defoliadoresde actividad nocturna asociados a Aristotelia chilensis (maqui) en el
bosque maulino fragmentado
Composition, species richness and abundance of nocturnal folivorous insects associatedwith Aristotelia chilensis (maqui) in the fragmented Maulino forest
XAVIERA DE LA VEGA1 & AUDREY A. GREZ2*
1 Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile. 16 Casilla 653, Santiago, Chile2 Departamento de Ciencias Biológicas Animales, Facultad Ciencias Veterinarias y Pecuarias, Universidad de Chile, Casilla
2, Correo 15, La Granja, Santiago, Chile;*e-mail para correspondencia: [email protected]
RESUMEN
En el bosque maulino, la herbivoría sobre Aristotelia chilensis (maqui) es negativamente afectada por lafragmentación del bosque, siendo mayor en el bosque continuo que en los fragmentos, particularmente ainicios de la temporada de crecimiento. Este fenómeno puede deberse a cambios en la dinámica de laspoblaciones de defoliadores, esencialmente insectos. En este trabajo se evaluó la abundancia, riqueza deespecies y composición de insectos defoliadores de actividad nocturna presentes en A. chilensis en un bosquecontinuo (600 ha) y en ocho fragmentos remanentes (0,4-20 ha). Los muestreos se realizaron mensualmente,entre agosto de 2005 y febrero de 2006, en 32 ejemplares adultos de A. chilensis en el bosque continuo y en32 ejemplares en los fragmentos. Los insectos fueron muestreados durante las cinco primeras horas de lanoche. Se recolectaron 890 insectos defoliadores, pertenecientes a 17 familias y 77 especies pertenecientes alos órdenes Coleoptera, Orthoptera y Lepidoptera, siendo todas nativas. La abundancia total no varió según elhábitat. Sin embargo, la fragmentación incrementó o disminuyó la abundancia de algunas especies. La riquezade especies por árbol tampoco fue afectada por la fragmentación del bosque, aunque el número total deespecies fue considerablemente mayor en los fragmentos que en el bosque continuo. La similitud de especiesfue mayor dentro del bosque continuo que entre el bosque continuo y los fragmentos o que entre losfragmentos. A principios de la temporada de crecimiento de A. chilensis (septiembre), la abundancia deSericoides obesa fue significativamente mayor en el bosque continuo que en los fragmentos. Al avanzar en latemporada, Sericoides viridis se hizo más abundante en los fragmentos. Por el tamaño y la voracidad de losinsectos del género Sericoides ellos serían los principales responsables de los patrones de defoliación de A.chilensis en el bosque maulino.
Palabras clave: insectos defoliadores, bosque maulino, fragmentación del hábitat.
ABSTRACT
At the Maulino forest, herbivory upon Aristotelia chilensis (maqui) is negatively affected by forestfragmentation, being higher in the continuous forest than in the small remnant fragments, particularly at thebeginning of the growing season (spring). This phenomenon may be attributable to changes in the populationdynamics of herbivores, mostly insects. In this work, the effect of the Maulino forest fragmentation on theabundance, species richness and composition of nocturnal defoliating insects associated with A. chilensis wasevaluated. The insects associated with A. chilensis were sampled monthly, between August 2005 and February2006, in 32 trees located in a continuous forest (600 ha) and in 32 trees located in eight forest fragments (0.4-20ha). Insect sampling was carried out during the first five night hours. 890 folivorous insects from 17 families and77 species of Coleoptera, Orthoptera and Lepidoptera were collected. All species were native. The totalabundance did not vary with habitat fragmentation. Nevertheless, some species increased their abundance withfragmentation, while others were depressed. The species richness of folivorous insects per tree did not vary withforest fragmentation, though the total number of species was considerably higher in the forest fragments. Thespecies similarity was higher within the continuous forest than between the continuous forest and fragments oramong fragments. At the beginning of the season (September), the abundance of Sericoides obesa wassignificantly higher in the continuous forest than in the forest fragments. Later in the season, Sericoides viridisbecame more abundant in the forest fragments. Because of the body size and the voracity of Sericoides sp., theseinsects seem to be responsible of the herbivory patterns of A. chilensis at the Maulino forest.
Key words: folivorous insects, Maulino forest, habitat fragmentation.

222 DE LA VEGA & GREZ
INTRODUCCIÓN
La deforestación y fragmentación de losbosques son fenómenos que ocurren a escalaglobal y constituyen problemas ambientalesapremiantes por su impacto sobre labiodiversidad (Myers 1988, Groom &Schumaker 1993, FAO 2002). Los cambios enla ident idad, diversidad o abundanciapoblacional de las especies, producto de lafragmentación, pueden al terar lasinteracciones ecológicas y los procesosecosistémicos, componentes funcionales de labiodiversidad (sensu Noss 1990). Entre lasinteracciones ecológicas afectadas por lafragmentación del hábi tat están lasinteracciones planta-animal , como laherbivoría. Cambios en esta interacción sonrelevantes por cuanto la pérdida de área foliarpuede tener consecuencias en el desempeño delas plantas , ya sea disminuyendo laproducción de frutos (Crawley 1985) oaumentando el aborto de frutos (Hochwenderet al. 2003), conllevando finalmente a cambiosen la estructura del bosque (Ricklefs & Miller1999). Los escasos antecedentes de lal i teratura sugieren que, en general , laherbivoría disminuye con la fragmentación(e.g., Roland 1993, Benítez-Malvido et al.1999, Bresciano et al. 1999, Benítez-Malvido2001, Arnold & Asquith 2002, Vásquez et al.2007). Por ejemplo, en bosques tropicalesexperimentalmente fragmentados en Manaos,la herbivoría sobre plántulas al inicio de latemporada, es más baja en fragmentos debosque que en un bosque continuo, aunquepasado algunos años tal diferencia desaparece(Benítez-Malvido et a l . 1999). Estadisminución en la herbivoría se debería aefectos depresores de la fragmentación sobrela abundancia o r iqueza de especies deherbívoros (Benítez-Malvido et al. 1999).
El bosque maulino, ubicado en la cordillerade la Costa de la Región del Maule, Chile,entre los 35º55’ y 37º20’ S, ha sidointensamente deforestado y fragmentado (Grezet al. 1997) con una reducción del área delbosque original de un 67 % entre los años 1975y 2000, quedando hoy en día distribuido enpequeños fragmentos rodeados por plantacionesde Pinus radiata (Grez et al. 1997, Bustamante& Castor 1998, Echeverría et al. 2006). Estebosque es de gran interés biológico debido al
alto endemismo de su biota y por ser un centrode confluencia de la biota austral y deasociaciones más nortinas como el bosqueesclerófilo (Saavedra & Simonetti 2001),siendo considerado un punto caliente para laconservación de biodiversidad (Myers et al.2000).
La fragmentación ha provocado importantescambios en la biota de este bosque. Así, loscarnívoros y algunas familias de insectosdisminuyen su abundancia en los pequeñosfragmentos de bosque, en cambio, loscoleópteros epígeos, las aves insectívoras, losmamíferos pequeños y el pudú son másabundantes en estos remanentes encomparación con un área extensa (600 ha) debosque continuo (véase Bustamante et al. 2006para una revisión). Estos cambios en lasabundancias poblacionales han implicadoalteraciones en la intensidad de algunasinteracciones ecológicas, como la granivoría,insectivoría y herbivoría (Simonetti et al.2006).
En el bosque maulino, la herbivoría dediferentes especies de plantas, tanto en suestado adulto como juvenil, disminuye en losfragmentos remanentes (Vega 2001, Vásquez etal. 2007, Simonetti et al. en prensa). Enparticular, la remoción de superficie foliar enAristotelia chilensis (Mol.) Stuntz(Elaeocarpaceae), el maqui, essignificativamente menor en los pequeñosfragmentos remanentes que en el bosquecontinuo, particularmente a inicios de latemporada de crecimiento de nuevas hojas.Luego estas diferencias comienzan adesaparecer, siendo al final de la temporada laherbivoría similar en fragmentos de bosque ybosque continuo (Vásquez et al. 2007). Estefenómeno puede deberse a cambios en ladinámica de las poblaciones de defoliadores,esencialmente insectos, debido a que estos sonlos herbívoros más importantes en este bosque(Vega 2001). Sin embargo, esta hipótesis aúnno ha sido puesta a prueba. Observacionespreliminares en maqui sugieren que los insectosherbívoros asociados a esta planta, comoSericoides sp. (Coleoptera: Scarabaeidae),tienen una actividad defoliadora principalmenteen la noche (Artigas 1994) y que los insectosherbívoros diurnos son escasos (Jaña-Prado &Grez 2004). Por ello, un muestreo nocturno esnecesario para evaluar los posibles efectos de la

223INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADO
fragmentación sobre este gremio de insectos. Elobjetivo de este trabajo fue evaluar si lafragmentación del bosque maulino afecta laabundancia, riqueza y composición de insectosdefoliadores de actividad nocturna presentes enA. chilensis y si estas variaciones se relacionancon los patrones de herbivoría descritos porVásquez et al. (2007).
MATERIALES Y MÉTODOS
Lugar del estudio
El estudio se realizó en la Reserva NacionalLos Queules (RNLQ) y en ocho fragmentosremanentes de bosque maulino vecinos a lareserva, pero separados de ella y entre sí por almenos 50 m. La RNLQ (35º41’19” S,72º41’15” O) se encuentra ubicada a 71 km aloeste de Cauquenes, en el sector de Tregualemuque, junto a terrenos vecinos de propiedadprivada, suman un área de 600 ha de bosquemaulino, una de las áreas más extensas de estebosque existente hoy en día. Los fragmentosremanentes de bosque comprenden áreasmenores a 20 ha, y se encuentran aislados entresí y de la reserva por plantaciones de pino (paraun mapa del área véase Donoso et al. 2004).Las especies dominantes son el hualo(Nothofagus glauca) y el roble (Nothofagusobliqua) que coexisten con especies endémicasque se encuentran en peligro de extinción comoel queule (Gomortega keule), ruil (Nothofagusalessandrii) y pitao (Pitavia punctata) (SanMartín & Donoso 1996). En el bosque maulinoA. chilensis es una especie abundante, presentetanto en los fragmentos de bosque como en elbosque continuo y en la matriz de pino,pudiendo en el futuro relegar a las especies quedominan actualmente en los fragmentos (hualoy roble) (Bustamante et al. 2005). De hecho, yahoy la densidad de este árbol essignificativamente mayor en los fragmentosque en el bosque continuo (media ± errorestándar = 43,9 ± 6,18 y 4,2 ± 0,99 árboles por400 m2, respectivamente) (Lagos 2007).
Diseño experimental y muestreo de insectos
Los insectos defoliadores (i.e., Coleoptera,Lepidoptera y Orthoptera) asociados a A.chilensis fueron muestreados mensualmente
desde agosto de 2005 a febrero de 2006, épocade mayor actividad de insectos en Chile centraly en el bosque maulino (Fuentes et al. 1981,Jaña-Prado & Grez 2004) y que coincide con elperiodo de crecimiento vegetativo de A.chilensis en la que se concentra su defoliación(Vásquez et al. 2007). Tanto en el bosquecontinuo como en los 8 fragmentos de bosquese seleccionaron aleatoriamente 32 ejemplaresadultos de A. chilensis (total = 64). En losfragmentos y en el bosque continuo los árbolesseleccionados se hallaban separados por almenos 10 m de distancia y en el caso delbosque continuo, ellos se distribuyeron a lolargo de un transecto de 700 m de largo,aproximadamente.
Los insectos fueron muestreados durantelas cinco primeras horas después delanochecer, a lo largo de tres noches de cadames, bajo condiciones climáticas similares.Cada noche, se incluyeron en el muestreotanto árboles de los fragmentos como delbosque continuo. En cada ejemplar escogidose seleccionaron al azar cinco ramas, cuyashojas fueron observadas por el haz y el envésdurante aproximadamente 10 min por árbol,recolectando los insectos presentes. Esteperiodo de tiempo resultó ser adecuado,incluso superior al necesario para evaluar lafauna presente en cinco ramas, por cuantoantes de este lapso ya no se observan insectosadicionales. Posterior a estas observaciones seutilizó el método de sacudido de follaje sobreparaguas entomológico, sacudiendo dos vecestodo el follaje de cada árbol y colectando losinsectos en un paño blanco de 1 m2 dispuestobajo las ramas.
Las larvas de lepidópteros fueron llevadas auna cámara de crianza, suministrándoles hojasde maqui como alimento hasta que alcanzaranel estado adulto y así poder identificarlas. Entanto, los coleópteros y los ortópteros fueronalmacenados en tubos Eppendorf con alcoholde 70º. Los insectos fueron identificados anivel de morfoespecie o especie cuando fueposible, a través de claves taxonómicas ycomparaciones con colecciones entomológicasdel Museo Nacional de Historia Natural y delMuseo Entomológico Luis Peña de la Facultadde Ciencias Agronómicas de la Universidad deChile. Los insectos fueron consideradosdefoliadores según recomendaciones deespecialistas.

224 DE LA VEGA & GREZ
Análisis estadístico
Debido a que no existe en el área otra unidad debosque continuo de estas dimensiones, lasmuestras de este tipo de bosque fueron tomadasde la misma unidad experimental, lo que podríasignificar que estas muestras no hubiesen sidoestadísticamente independientes. Por ello, paraprobar la independencia espacial de lasmuestras, y usarlas como réplicas en los análisisestadísticos, se aplicó la prueba de Mantel(Fortin & Gurevitch 1993). Esta pruebademostró que las diferencias en abundancia yriqueza de insectos herbívoros entre árbolesdentro del bosque continuo y dentro de losfragmentos fueron espacialmente independientes(abundancia, rs = -0,13; P = 0,76 para bosquecontinuo y rs = -0,13; P = 0,77 para fragmentos;riqueza, rs = 0,01; P = 0,74 para bosque continuoy rs = 0,02; P = 0,56 para fragmentos). Por ello,cada árbol fue considerado como réplica.
La abundancia poblacional (número deindividuos por árbol) y la riqueza de especies(número de especies por árbol) de insectosdefoliadores en bosque continuo y fragmentos secomparó mediante un análisis de varianza paramedidas repetidas (ANDEVAmr), con las fechasde muestreo como medida repetida, usando elprograma computacional Statistica 6.0 (StatsoftInc. 2001). Los datos fueron transformados conlog10 (x + 1), para cumplir con el supuesto denormalidad. Como prueba a posteriori se utilizóla prueba de Tukey con a = 0,05.
La similitud en la composición de insectosherbívoros entre el bosque continuo y losfragmentos (considerando las capturasacumuladas durante todo el estudio) se estimómediante el coeficiente de Sorensen (Krebs1989) y el índice de Morisita-Horn (Moreno2001). Estos índices se calcularon de dosmaneras: primero comparando la totalidad deespecies encontradas en todos los árbolesmuestreados en el bosque continuo y en losocho fragmentos y, segundo, agrupando losdatos en ocho bloques para bosque continuo yocho para fragmentos, de manera de tener unavariabilidad al interior del bosque continuo.Cada bloque estuvo compuesto por cuatroárboles, los que en el bosque continuocorrespondieron a los cuatro árboles máscercanos dentro del transecto y en losfragmentos a los cuatro árboles muestreadosdentro de cada fragmento.
Adicionalmente, para representargráficamente la similitud de especies entre losbloques del bosque continuo y de losfragmentos, se construyó un dendrograma através de un análisis de conglomerados, el queconsideró las abundancias de todas lasespecies, excluyendo a Lepidoptera sp., porcuanto gran parte de este grupo no fueidentificado a nivel de especie (i .e. ,lepidópteros criados y que no llegaron aadultos). El análisis se realizó mediante elmétodo de agrupamiento Encadenamientopromedio o UPGMA, usando el programaInfoStat (InfoStat 2006). En el análisis, ladistancia util izada fue la euclidiana . Lasignificancia estadística de las agrupacionesobservadas se determinó a partir de una pruebade aleatorización, utilizando el programaPopTools 2.7.5 (Poptools 2006).
RESULTADOS
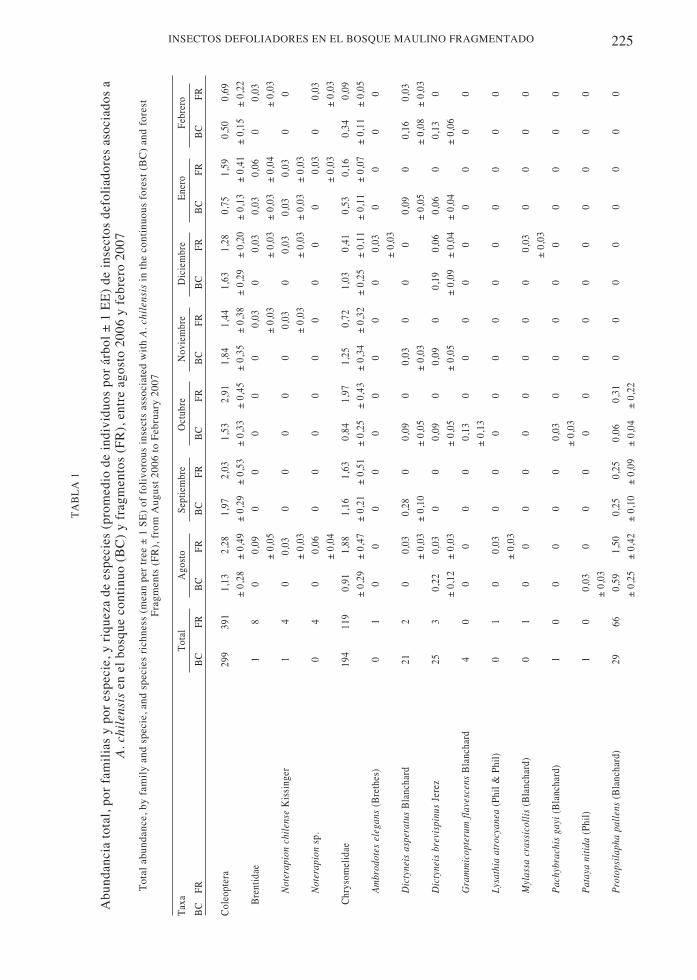
A lo largo del estudio se colectaron 890individuos de insectos defoliadorespertenecientes a Coleoptera, Orthoptera yLepidoptera, de 17 familias y 77 especies(Tabla 1). El 93,5 % de las especies fueronidentificadas a nivel de especie o morfo-especie, siendo todas nativas.
En general Coleoptera fue el orden con mayorabundancia, representando el 77,5 % del total deindividuos capturados, seguido por Lepidopteracon 12,4 % y Orthoptera con 10,1 % del total deindividuos. Las familias más comunes fueron:Chrysomelidae, Scarabaeidae y Elateridae, todaspertenecientes a Coleoptera.
Desde agosto a diciembre la familia másabundante fue Chrysomelidae, siendo en estosmeses sus especies más representadas:Protopsilapha pallens (Blanchard) ,Protopsilapha pyrroptera (Phil & Phil) ,Psathyrocerus sp. 1 y Psathyrocerus fulvipesBlanchard. En enero, en cambio, la familia conmás individuos fue Scarabaeidae y la especiemás habitual fue Sericoides viridis (Solier). Enfebrero, Chrysomelidae volvió a ser la familiamás abundante pero S. viridis, al igual que en elmes anterior, fue la especie más abundante.Además de P. pallens, P. pyrroptera y P.fulvipes, durante todo el periodo de muestreoDeromecus pallipes (Solier) (Coleoptera:Elateridae) fue otra especie abundante.
225INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADO
TA
BL
A 1
Abu
ndan
cia
tota
l, p
or f
amil
ias
y po
r es
peci
e, y
riq
ueza
de
espe
cies
(pr
omed
io d
e in
divi
duos
por
árb
ol ±
1 E
E)
de i
nsec
tos
defo
liad
ores
aso
ciad
os a
A. c
hile
nsis
en
el b
osqu
e co
ntin
uo (
BC
) y
frag
men
tos
(FR
), e
ntre
ago
sto
2006
y f
ebre
ro 2
007
Tot
al a
bund
ance
, by
fam
ily
and
spec
ie,
and
spec
ies
rich
ness
(m
ean
per
tree
± 1
SE
) of
fol
ivor
ous
inse
cts
asso
ciat
ed w
ith
A.
chil
ensi
s in
the
con
tinu
ous
fore
st (
BC
) an
d fo
rest
Fra
gmen
ts (
FR
), f
rom
Aug
ust
2006
to
Feb
ruar
y 20
07
Tax
aT
otal
Ago
sto
Sept
iem
bre
Oct
ubre
Nov
iem
bre
Dic
iem
bre
Ene
roFe
brer
o
BC
FRB
CFR
BC
FRB
CFR
BC
FRB
CFR
BC
FRB
CFR
BC
FR
Col
eopt
era
299
391
1,1
3 2
,28
1,9
7 2
,03
1,5
3 2
,91
1,8
4 1
,44
1,6
3 1
,28
0,7
5 1
,59
0,5
0 0
,69
± 0,
28±
0,49
± 0,
29±
0,53
± 0,
33±
0,45
± 0,
35±
0,38
± 0,
29±
0,20
± 0,
13±
0,41
± 0,
15±
0,22
Bre
ntid
ae1
80
0,0
90
00
00
0,0
30
0,0
3 0
,03
0,0
60
0,0
3
± 0,
05±
0,03
± 0,
03±
0,03
± 0,
04±
0,03
Not
erap
ion
chil
ense
Kis
sing
er1
40
0,03
00
00
00,
030
0,03
0,03
0,0
30
0
± 0,
03±
0,03
± 0,
03±
0,03
± 0,
03
Not
erap
ion
sp.
04
0 0
,06
00
00
00
00
0
0,03
0 0
,03
± 0,
04±
0,03
± 0,
03
Chr
ysom
e lid
a e19
411
9 0
,91
1,8
8 1
,16
1,6
3 0
,84
1,9
7 1
,25
0,7
2 1
,03
0,4
1 0
,53
0,1
6 0
,34
0,0
9
± 0,
29±
0,47
± 0,
21±
0,51
± 0,
25±
0,43
± 0,
34±
0,32
± 0,
25±
0,11
± 0,
11±
0,07
± 0,
11±
0,05
Am
brod
ote s
ele
gans
(B
reth
e s)
01
00
00
00
00
0 0
,03
00
00
± 0,
03
Dic
tyne
is a
spe r
atus
Bla
ncha
rd21
20
0,0
3 0
,28
0 0
,09
0 0
,03
00
0 0
,09
0 0
,16
0,0
3
± 0,
03±
0,10
± 0,
05±
0,03
± 0,
05±
0,08
± 0,
03
Dic
tyne
is b
rev i
spin
us J
e re z
253
0,2
2 0
,03
00
0,0
90
0,0
90
0,1
9 0
,06
0,0
60
0,1
30
± 0,
12±
0,03
± 0,
05±
0,05
± 0,
09±
0,04
± 0,
04±
0,06
Gra
mm
icop
teru
m f
lav e
sce n
s B
lanc
hard
40
00
00
0,1
30
00
00
00
00
± 0,
13
Lysa
thia
atr
ocy a
nea
(Phi
l & P
hil)
01
0 0
,03
00
00
00
00
00
00
± 0,
03
My l
assa
cra
ssic
olli
s (B
lanc
hard
)0
10
00
00
00
00
0,0
30
00
0
± 0,
03
Pac
hybr
achi
s ga
y i (
Bla
ncha
rd)
10
00
00
0,0
30
00
00
00
00
± 0,
03
Pat
aya
niti
da (
Phil)
10
0,0
30
00
00
00
00
00
00
± 0,
03
Pro
tops
ilap
ha p
alle
ns (
Bla
ncha
rd)
2966
0,5
9 1
,50
0,2
5 0
,25
0,06
0,3
10
00
00
00
0
± 0,
25±
0,42
± 0,
10±
0,09
± 0,
04±
0,22

226 DE LA VEGA & GREZ
TA
BL
A 1
(co
ntin
uaci
ón)
Tax
aT
otal
Ago
sto
Sept
iem
bre
Oct
ubre
Nov
iem
bre
Dic
iem
bre
Ene
roFe
brer
o
BC
FRB
CFR
BC
FRB
CFR
BC
FRB
CFR
BC
FRB
CFR
Pro
tops
ilap
ha p
yrro
pter
a (P
hil &
Phi
l)7
770
0,0
9 0
,06
1,0
6 0
,16
1,1
30
0,1
30
00
00
0
± 0,
07±
0,04
± 0,
45±
0,10
± 0,
42±
0,07
Pro
tops
ilap
ha s
igna
ta (
Bla
ncha
rd)
2619
0,0
6 0
,19
0,5
6 0
,28
0,1
9 0
,13
00
00
00
00
± 0,
04±
0,07
± 0,
15±
0,09
± 0,
10±
0,06
Pro
tops
ilap
ha s
p. 1
01
00
00
00
00
00
0 0
,03
00
± 0,
03
Psa
thyr
ocer
us f
ulvi
pes
Bla
ncha
rd40
270
00
0 0
,06
0,1
6 0
,47
0,3
2 0
,44
0,1
9 0
,22
0,1
3 0
,06
0,0
6
± 0,
04±
0,08
± 0,
18±
0,17
± 0,
13±
0,08
± 0
,07
± 0,
06±
0,06
± 0,
04
Psa
thyr
ocer
us s
p.40
210
00
0,0
3 0
,03
0,2
5 0
,66
0,2
8 0
,41
0,0
9 0
,16
00
0
± 0,
03±
0,03
± 0,
08±
0,20
± 0,
12±
0,14
± 0,
05±
0,07
Cur
culio
nida
e26
35 0
,06
0,1
6 0
,22
0,1
3 0
,16
0,3
4 0
,09
0,0
9 0
,06
0,1
3 0
,13
0,1
6 0
,09
0,0
9
± 0,
04±
0,08
± 0,
09±
0,07
± 0,
07±
0,10
± 0,
05±
0,05
± 0,
04±
0,07
± 0,
06±
0,13
± 0,
05±
0,05
Aca
lles
fus
ensc
ens
Bla
ncha
rd1
00
00
0 0
,03
00
00
00
00
0
± 0,
03
Aca
lle s
lin
e ole
tus
Bla
ncha
rd0
10
00
00
00
00
0,0
30
00
0
± 0,
03
Aca
lle s
sp.
10
10
0,0
30
00
00
00
00
00
0
± 0,
03
Aca
lle s
sp.
27
10
0 0
,16
00
0,0
3 0
,03
0 0
,03
00
00
0
± 0,
07±
0,03
± 0,
03±
0,03
Aca
lle s
sp.
37
2 0
,03
0 0
,03
0,0
3 0
,03
0,0
3 0
,03
00
0 0
,03
0 0
,06
0
± 0,
03±
0,03
± 0,
03±
0,03
± 0,
03±
0,03
± 0,
03±
0,04
Aca
lle s
sp.
41
10
00
00
00
00
0 0
,03
0,0
30
0
± 0,
03±
0,03
Aeg
orhi
nus
supe
rcil
iosu
s (G
uéri
n)0
10
00
00
00
00
00
00
0,0
3
± 0,
03
Ana
ball
us c
rist
atig
e r B
lanc
hard
01
00
00
0 0
,03
00
00
00
00
± 0,
03
Ana
ball
us p
lagi
atus
Bla
ncha
rd1
00
0 0
,03
00
00
00
00
00
0
± 0,
03
Be r
beri
dic o
la a
ter
(Phi
l & P
hil)
10
0,0
30
00
00
00
00
00
00
± 0,
03
Euc
alus
te s
sell
atus
Bla
ncha
rd2
00
00
0 0
,06
00
00
00
00
0
± 0,
04

227INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADOG
ayus
ele
gans
(B
lanc
hard
)0
20
0,0
30
00
0,0
30
00
00
00
0
± 0,
03±
0,03
Hib
reol
epto
ps s
p.4
80
0,0
60
0 0
,03
0,0
9 0
,03
00
0,0
3 0
,03
0 0
,03
0,0
6
± 0,
04±
0,03
± 0,
05±
0,03
± 0,
03±
0,03
± 0,
03±
0,04
Meg
alom
etis
spi
nife
ra B
oh1
00
00
00
00
00
0 0
,03
00
0
± 0,
03
Om
oide
s fl
avip
es (
Bla
ncha
rd)
16
00
00
00
00
0,0
3 0
,06
0 0
,13
00
± 0,
03±
0,06
± 0,
13
Pol
ydro
sus
noth
ofag
i K
usch
el0
60
00
00
0,0
90
0,0
90
00
00
0
± 0,
05±
0,05
Pol
ydro
sus
rose
us B
lanc
hard
03
00
0 0
,09
00
00
00
00
00
± 0,
07
Spha
erio
poeu
s fa
ber
Kus
hel
02
0 0
,03
00
0 0
,03
00
00
00
00
± 0,
03±
0,03
Ela
teri
dae
2456
00
0,0
6 0
,19
0,3
1 0
,47
0,1
9 0
,38
0,1
6 0
,41
0,0
3 0
,25
0 0
,06
± 0,
04±
0,08
± 0,
16±
0,19
± 0,
07±
0,13
± 0,
08±
0,11
± 0,
03±
0,08
± 0,
04
Agr
iote
s ge
rmai
ni F
leut
01
00
00
00
00
00
0 0
,03
00
± 0,
03
Der
omec
us f
ilic
orni
s So
lier
01
00
00
00
00
0 0
,03
00
00
± 0,
03
De r
ome c
us p
alli
pes
(Sol
ier)
2348
00
0,0
6 0
,19
0,3
1 0
,47
0,1
9 0
,34
0,1
3 0
,31
0,0
3 0
,19
00
± 0,
04±
0,08
± 0,
16±
0,19
± 0,
07±
0,12
± 0,
07±
0,09
± 0,
03±
0,07
De r
ome c
us s
p. 1
01
00
00
00
00
0 0
,03
00
00
0±
0,03
De r
ome c
us s
p. 2
01
00
00
00
00
0 0
,03
00
00
± 0,
03
De r
ome c
us v
ulga
ris
Solie
r0
10
00
00
00
00
00
00
0,0
3
± 0,
03
Ov i
palp
us p
ube s
c ens
Sol
ier
01
00
00
00
00
00
0 0
,03
00
± 0,
03
Ela
teri
dae
sp.1
11
00
00
00
00
0,0
3 0
,03
00
00
± 0,
03±
0,03
Ela
teri
dae
sp. 2
01
00
00
00
00
0 0
,03
00
00
± 0,
03
Nem
onyc
hida
e1
0 0
,03
00
00
00
00
00
00
0
± 0,
03

228 DE LA VEGA & GREZ
TA
BL
A 1
(co
ntin
uaci
ón)
Tax
aT
otal
Ago
sto
Sept
iem
bre
Oct
ubre
Nov
iem
bre
Dic
iem
bre
Ene
roFe
brer
o
BC
FRB
CFR
BC
FRB
CFR
BC
FRB
CFR
BC
FRB
CFR
Rhy
nchi
tom
acer
api
onoi
des
Kus
ch1
0 0
,03
00
00
00
00
00
00
0
± 0,
03
Scar
abae
idae
4364
0,1
3 0
,16
0,4
4 0
,09
0,2
2 0
,13
0,3
1 0
,19
0,2
2 0
,19
0,0
3 0
,88
0 0
,38
± 0,
06±
0,08
± 0,
11±
0,05
± 0,
09±
0,06
± 0,
12±
0,09
± 0,
11±
0,09
± 0,
03±
0,26
± 0,
19
Phy
tola
ema
mut
abil
is S
olie
r1
10
00
0 0
,03
0,0
30
00
00
00
0
± 0,
03±
0,03
Seri
coid
es o
besa
(G
erm
ain)
419
0,1
3 0
,13
0,4
4 0
,09
0,1
90
0,3
1 0
,06
0,2
20
00
00
± 0,
06±
0,07
± 0,
11±
0,05
± 0,
08±
0,12
± 0,
04±
0,11
Seri
coid
es s
p. 1
01
00
00
0 0
,03
00
00
00
00
± 0,
03
Seri
coid
es s
p. 2
01
0 0
,03
00
00
00
00
00
00
± 0,
03
Seri
coid
es s
p. 3
07
00
00
00
00
00
0 0
,22
00
± 0,
09
Seri
c oid
e s v
arie
gata
(G
e rm
a in)
07
00
00
0 0
,06
0 0
,13
0 0
,03
00
00
± 0,
04±
0,09
± 0,
03
Seri
c oid
e s v
irid
is (
Solie
r)1
380
00
00
00
00
0,1
6 0
,03
0,6
60
0,3
8
± 0,
08±
0,03
± 0,
24±
0,19
Ten
ebri
onid
a e10
90
0 0
,09
00
00
0,0
3 0
,16
0,1
3 0
,03
0,0
9 0
,03
0,0
3
± 0,
07±
0,03
± 0,
07±
0,07
± 0,
03±
0,05
± 0,
03±
0,03
All
e cul
a se
mio
bcur
a (C
ambe
ll)0
10
00
00
00
00
00
0,0
30
0
± 0,
03
Euc
olis
a Sa
ngui
nic o
llis
F.G
01
00
00
00
00
0 0
,03
00
00
± 0,
03
He l
iofu
gus
are n
osus
Gué
rin-
Mén
evill
e6
20
0 0
,06
00
00
0 0
,09
0,0
6 0
,03
00
0
±0,
06±
0,05
± 0,
06±
0,03
He l
iofu
gus
impr
e ssu
s G
uéri
n-M
énev
ille
45
00
0,0
30
00
0 0
,03
0,0
6 0
,03
0 0
,06
0,0
3 0
,03
± 0,
03±
0,03
± 0,
04±
0,03
± 0,
04±
0,03
± 0,
03
Lep
idop
tera
5258
0,0
3 0
,03
0,1
9 0
,25
0,2
2 0
,47
0,3
8 0
,34
0,2
5 0
,25
0,3
4 0
,28
0,
22 0
,19
± 0,
03±
0,03
± 0,
08±
0,10
± 0,
07±
0,16
± 0,
09±
0,10
± 0,
08±
0,09
± 0,
10±
0,08
± 0,
07±
0,07
Geo
me t
rida
e2
50
00
00
0 0
,03
0,0
6 0
,03
0,0
90
00
0
± 0,
03±
0,04
± 0,
03±
0,07
Mal
lec o
sp.
02
00
00
00
0 0
,03
0 0
,03
00
00
± 0,
03±
0,03

229INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADOSy
ncir
sode
s pr
imat
a W
alke
r2
30
00
00
0 0
,03
0,0
3 0
,03
0,0
60
00
0
± 0,
03±
0,03
± 0,
03±
0,06
Las
ioca
mpi
dae
03
00
0 0
,03
0 0
,06
00
00
00
00
± 0,
03±
0,04
Mac
rom
phal
ia a
ffin
is F
eish
amel
02
00
0 0
,03
0 0
,03
00
00
00
00
± 0,
03±
0,03
Mac
rom
phal
ia s
p.0
10
00
00
0 ,0
30
00
00
00
0
± 0,
03
Oec
opho
rida
e6
40
00
00
00
00
0 0
,16
0,0
6 0
,03
0,0
6
± 0,
07±
0,04
± 0,
03±
0,04
Dit
a sp
.1
00
00
00
00
00
0 0
,03
00
0
± 0,
03
Oec
opho
rida
e sp
.4
40
00
00
00
00
0 0
,09
0,0
6 0
,03
0,0
6
± 0,
05±
0,04
± 0,
03±
0,04
Osm
aria
arg
illa
Cla
rke
10
00
00
00
00
00
0,0
30
00
± 0,
03
Pyra
lidae
20
00
00
00
00
00
0,0
60
00
± 0,
04
Pyra
lidae
sp.
20
00
00
00
00
00
0,0
60
00
± 0,
04
Satu
rniid
a e1
10
00
00
0 0
,03
0,0
30
00
00
0
± 0,
03±
0,03
Orm
isc o
des
rufo
sign
ata
Bla
ncha
rd1
10
00
00
0 0
,03
0,0
30
00
00
0
± 0,
03±
0,03
Tor
tric
ida e
01
00
00
00
00
00
00
0 0
,03
± 0,
03
Hae
mat
e uli
a ba
rrig
ana
Ra z
owsk
i y G
onz á
lez
01
00
00
00
00
00
00
0 0
,03
± 0,
03
Lep
idop
tera
sp.
4144
0,0
3 0
,03
0,1
9 0
,22
0,2
2 0
,41
0,3
1 0
,25
0,2
2 0
,16
0,1
3 0
,22
0,1
9 0
,09
± 0,
03±
0,03
± 0,
08±
0,09
± 0,
07±
0,15
± 0,
08±
0,09
± 0,
07±
0,07
± 0,
06±
0,07
± 0,
07±
0,05
Ort
hopt
e ra
5634
0,0
9 0
,16
0,4
1 0
,22
0,3
1 0
,13
0,0
3 0
,13
0,2
5 0
,25
0,4
4 0
,09
0,2
2 0
,09
± 0,
05±
0,08
± 0,
15±
0,11
± 0,
10±
0,06
± 0,
03±
0,06
± 0,
08±
0,13
± 0,
11±
0,05
± 0,
09±
0,05
Pse u
doph
a sm
a tid
a e3
00
0 0
,03
00
00
00
0 0
,03
0 0
,03
0
± 0,
03±
0,03
± 0,
03
Bac
uncu
lus
phy l
lopu
s (G
ray)
30
00
0,0
30
00
00
00
0,0
30
0,0
30
± 0,
03±
0,03
± 0,
03
Rha
phid
opho
rida
e21
19 0
,09
0,1
3 0
,19
0,1
3 0
,16
0,0
6 0
,03
0,0
9 0
,06
0,1
3 0
,09
0 0
,03
0,0
6
± 0,
05±
0,07
± 0,
09±
0,09
± 0,
07±
0,04
± 0,
03±
0,05
± 0,
04±
0,06
± 0,
05±
0,03
± 0,
04
230 DE LA VEGA & GREZ
TA
BL
A 1
(co
ntin
uaci
ón)
Tax
aT
otal
Ago
sto
Sept
iem
bre
Oct
ubre
Nov
iem
bre
Dic
iem
bre
Ene
roFe
brer
o
BC
FRB
CFR
BC
FRB
CFR
BC
FRB
CFR
BC
FRB
CFR
Het
erom
allu
s sp
. 10
10
00
00
00
0,0
30
00
00
0
± 0,
03
Het
erom
allu
s sp
. 22
10
00
00
00
0,
03 0
,03
00
0 0
,03
0
± 0,
03±
0,03
± 0,
03
Het
erom
allu
s sp
. 314
5 0
,03
0 0
,16
0 0
,13
0 0
,03
0,0
30
0,0
9 0
,09
00
0,0
3
± 0,
03±
0,08
± 0,
06±
0,03
± 0,
03±
0,05
± 0,
05±
0,03
Ude
nus
fasc
iatu
s A
rder
512
0,0
6 0
,13
0,0
3 0
,13
0,0
3 0
,06
00
0,0
3 0
,03
00
0
0,03
± 0,
04±
0,07
± 0,
03±
0,09
± 0,
03±
0,04
± 0,
03±
0,03
± 0,
03
Tet
tigon
idae
3011
0 0
,03
0,1
6 0
,09
0,1
6 0
,06
0 0
,03
0,1
6 0
,03
0,3
1 0
,09
0,1
60
± 0,
03±
0,08
± 0,
07±
0,07
± 0,
04±
0,03
± 0,
07±
0,03
± 0,
09±
0,05
± 0,
07
Aph
ract
us a
cum
inat
us (
Bru
ner
von
Wat
tenw
yl)
01
0 0
,03
00
00
00
00
00
00
± 0,
03
Cos
mop
hyll
um c
f. ol
ivac
eum
Bla
ncha
rd4
10
00
0 0
,03
00
00
0 0
,09
0,0
30
0
± 0,
03±
0,07
± 0,
03
Par
aphr
actu
s ab
bre v
iatu
s (B
rune
r vo
n W
a tte
nwyl
)14
30
0 0
,03
00
00
0 0
,09
0,0
3 0
,19
0,0
6 0
,13
0
± 0,
03±
0,05
± 0,
03±
0,07
± 0,
04±
0,06
Pol
y cle
ptis
sp.
20
00
00
00
00
0,0
30
0,0
30
00
± 0,
03±
0,03
Ste n
ophy
llia
mod
e sta
(B
lanc
hard
)10
60
0 0
,13
0,0
9 0
,13
0,0
60
0,0
30
00
00
0
± 0,
06±
0,07
± 0,
06±
0,04
± 0,
03
Tri
ste r
ida e
24
00
0,0
30
00
00
0,0
3 0
,09
00
0 0
,03
± 0,
03±
0,03
± 0,
07±
0,03
Ely
siac
ris
e nsi
c orn
is C
iglia
no2
40
0 0
,03
00
00
0 0
,03
0,0
90
00
0,0
3
± 0,
03±
0,03
± 0,
07±
0,03
Indi
vidu
os p
or á
rbol
12,
72 1
5,09
1,2
5 2
,63
2,56
2
,50
2,0
63,
52,
25 1
,91
2,1
3 1
,78
1,5
3 1
,97
0,9
4 0
,97
± 1,
03±
1,39
± 0,
28±
0,50
± 0,
34±
0,57
± 0
,33
± 0,
52±
0,36
± 0,
38±
0,32
± 0,
26±
0,22
± 0,
43±
0,20
± 0,
23
Tot
a l in
divi
duos
407
483
4079
8280
6611
272
6168
5749
6330
31
Esp
e cie
s po
r á r
bol
11,9
411
,84
1,25
1,67
2,16
1,69
1,88
2,19
1,84
1,63
1,75
1,78
1,75
1,78
1,3
1
1,22
± 0,
59±
0,63
± 0,
08±
0,16
± 0,
25±
0,20
± 0,
18±
0,26
± 0,
18±
0,17
± 0,
20±
0,18
± 0,
17±
0,23
± 0,
12±
0,09
Tot
a l e
spe c
ies
4663
1117
1812
2121
1318
1825
2117
1215

231INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADO
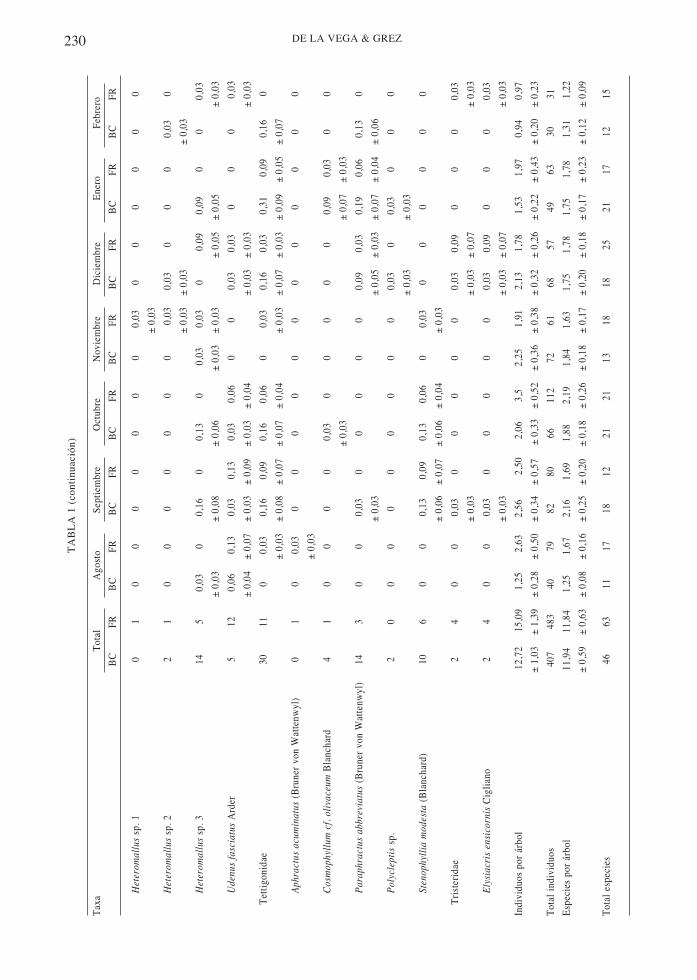
En cuanto al número de especies dentro decada orden, de las 77 especies colectadas,Coleoptera acumula el 72,7 % (56 especies),seguida por Orthoptera con un 14,3 % (11) ypor Lepidoptera con un 13 % (10 especies) deltotal de especies. Las familias con mayorriqueza fueron, en Coleoptera, Curculionidae(18 especies), Chrysomelidae (14 especies) yElateridae (nueve especies); en Lepidoptera,Oecophoridae (tres especies) y en Orthoptera,Tettigonidae (cinco especies, Tabla 1).
Abundancia de insectos defoliadores asociadosa A. chilensis en fragmentos y bosque continuo
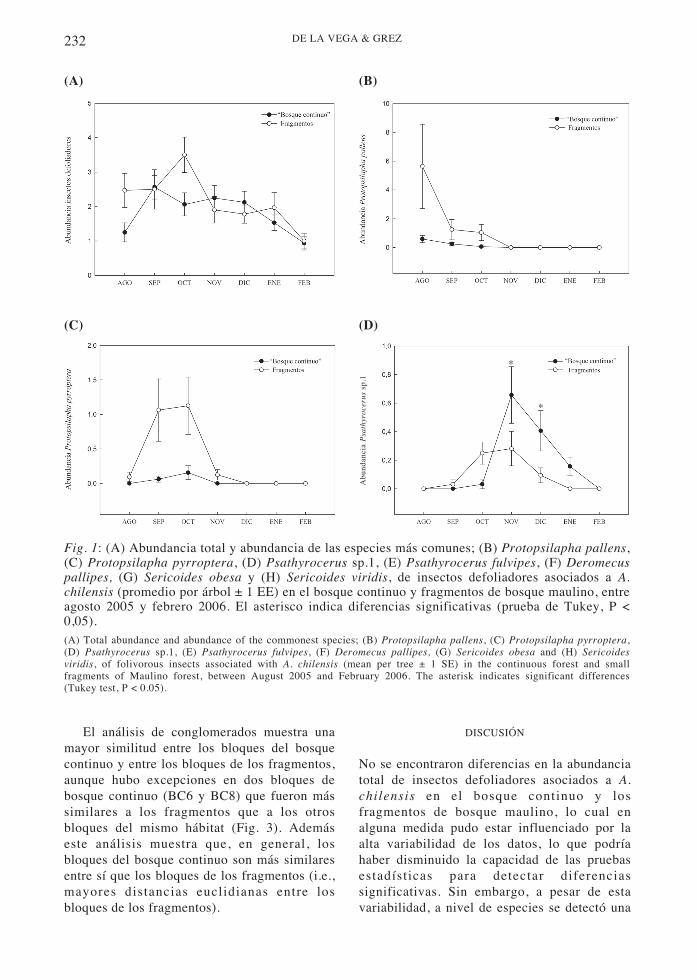
Considerando todas las fechas de muestreo, laabundancia total de insectos defoliadoresasociados a A. chilensis no varió entre bosquecontinuo y fragmentos (ANDEVAmr, efectohábitat, F1,62 = 0,30; P = 0,59). Si bien en agostoy octubre la abundancia total fue levementesuperior en los fragmentos que en el bosquecontinuo, estas diferencias no fueronestadísticamente significativas (ANDEVAmr,efecto hábitat*fecha, F6,372 = 2,06; P = 0,06).Desde finales de primavera (noviembre) hastafinales de verano (febrero) la abundancia totalde insectos defoliadores fue disminuyendo,manteniéndose similar entre los dos tipos hábitat(Fig. 1A). A nivel de especies, entre losChrysomelidae la abundancia de P. pallens fuemayor en los fragmentos que en el bosquecontinuo (ANDEVAmr, efecto hábitat, F1,62 =3,84; P = 0,05). Esta diferencia ocurrióparticularmente en agosto, cuando se capturó lamayor cantidad de individuos, en tanto laabundancia en el bosque continuo siempre fuebaja (Fig. 1B). Protopsilapha pyrroptera en elbosque continuo solo fue capturada enseptiembre y octubre, en cambio, en losfragmentos estuvo presente de agosto anoviembre. La abundancia de esta especie fueafectada por el tipo de hábitat, siendo siempremás abundante en los fragmentos (ANDEVAmr,efecto hábitat, F1,62 = 9,73; P = 0,003; Fig. 1C).La abundancia de Psathyrocerus sp. 1 no fueafectada por el tipo de hábitat (ANDEVAmr,efecto hábitat, F1,62 = 2,31; P = 0,13). Sinembargo, en noviembre y diciembre esta morfo-especie fue significativamente más abundante enel bosque continuo que en los fragmentos(ANDEVAmr, efecto hábitat*fecha, F4,248 =4,21; P = 0,003; Fig. 1D). La abundancia de P.
fulvipes nunca fue afectada por el tipo de hábitat(ANDEVAmr, efecto hábitat, F1,62 = 1,44; P =0,23, Fig. 1E).
Entre los Elateridae, D. pallipes fue siempremás abundante en los fragmentos de bosqueque en el bosque continuo (ANDEVAmr, efectohábitat, F1,62 = 4,30; P = 0,04; Fig. 1F), entanto entre los Scarabaeidae, S. obesa fuesiempre más abundante en el bosque continuoque en los fragmentos (ANDEVAmr, efectohábitat, F1,62 = 11,44; P < 0,001; Fig. 1G). Enel bosque continuo esta especie se capturó entreagosto y diciembre, en cambio, en losfragmentos se capturó entre agosto ynoviembre, pero en muy bajas abundancias. Porotra parte, S. viridis fue significativamente másabundante en los fragmentos que en bosquecontinuo (ANDEVAmr, efecto hábitat, F1,62 =9,14; P < 0,001), particularmente entrediciembre y febrero. Solo un individuo de S.viridis fue observado en el bosque continuo, locual ocurrió en enero (Fig. 1H).
Riqueza de especies y composición de insectosdefoliadores asociados a A. chilensis en frag-mentos y bosque continuo
La riqueza de especies de insectos defoliadorespor árbol no fue afectada por la fragmentacióndel bosque maulino (ANDEVAmr, efectohábitat, F1,62 = 0,16; P = 0,69; Fig. 2). Sinembargo, en total se capturaron más especies enlos fragmentos que en el bosque continuo, con63 y 46 especies, respectivamente (Tabla 1).
Considerando todo el periodo de estudio, 14especies fueron capturadas solo en el bosquecontinuo, 32 solo en los fragmentos y 30 fueroncompartidas por ambos hábitat. La similitud enla composición de especies entre bosquecontinuo y fragmentos según el índice deSorensen fue de 0,59 y según el índice deMorsita-Horn fue de 0,60. Al hacer el análisispor bloques (ocho del bosque continuo y ocho delos fragmentos), en promedio la similitud segúnel Índice de Sorensen fue 0,77 entre los bloquesdel bosque continuo y 0,73 entre los bloques delos fragmentos, en tanto entre los bloques delbosque continuo y de los fragmentos fue 0,74.Según el índice de Morisita-Horn, la similitud delos bloques al interior del bosque continuo fue0,17, de los bloques de los fragmentos fue 0,09 yentre los bloques del bosque continuo y de losfragmentos fue 0,13.
232 DE LA VEGA & GREZ
Fig. 1: (A) Abundancia total y abundancia de las especies más comunes; (B) Protopsilapha pallens,(C) Protopsilapha pyrroptera, (D) Psathyrocerus sp.1, (E) Psathyrocerus fulvipes, (F) Deromecuspallipes, (G) Sericoides obesa y (H) Sericoides viridis, de insectos defoliadores asociados a A.chilensis (promedio por árbol ± 1 EE) en el bosque continuo y fragmentos de bosque maulino, entreagosto 2005 y febrero 2006. El asterisco indica diferencias significativas (prueba de Tukey, P <0,05).(A) Total abundance and abundance of the commonest species; (B) Protopsilapha pallens, (C) Protopsilapha pyrroptera,(D) Psathyrocerus sp.1, (E) Psathyrocerus fulvipes, (F) Deromecus pallipes, (G) Sericoides obesa and (H) Sericoidesviridis, of folivorous insects associated with A. chilensis (mean per tree ± 1 SE) in the continuous forest and smallfragments of Maulino forest, between August 2005 and February 2006. The asterisk indicates significant differences(Tukey test, P < 0.05).
(A) (B)
(C) (D)
El análisis de conglomerados muestra unamayor similitud entre los bloques del bosquecontinuo y entre los bloques de los fragmentos,aunque hubo excepciones en dos bloques debosque continuo (BC6 y BC8) que fueron mássimilares a los fragmentos que a los otrosbloques del mismo hábitat (Fig. 3). Ademáseste análisis muestra que, en general, losbloques del bosque continuo son más similaresentre sí que los bloques de los fragmentos (i.e.,mayores distancias euclidianas entre losbloques de los fragmentos).
DISCUSIÓN
No se encontraron diferencias en la abundanciatotal de insectos defoliadores asociados a A.chilensis en el bosque continuo y losfragmentos de bosque maulino, lo cual enalguna medida pudo estar influenciado por laalta variabilidad de los datos, lo que podríahaber disminuido la capacidad de las pruebasestadísticas para detectar diferenciassignificativas. Sin embargo, a pesar de estavariabilidad, a nivel de especies se detectó una
Abu
ndan
cia
Psa
thy r
oce r
us s
p.1
*
*

233INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADO
Fig. 2: Riqueza de especies de insectos defoliadores asociados a A. chilensis (promedio por árbol ±1 EE) en el bosque continuo y fragmentos de bosque maulino, entre agosto 2005 y febrero 2006.Species richness of folivorous insects associated with A. chilensis (mean per tree ± 1 SE) in the continuous forest and smallfragments of Maulino forest, between August 2005 and February 2006.
(E) (F)
(G) (H)

234 DE LA VEGA & GREZ
Fig. 3: Dendrograma de similitud, considerando la abundancia de las diferentes especies de insectosdefoliadores asociados a A. chilensis en los 16 bloques (ocho del bosque continuo, BC, y ocho delos fragmentos, F). La línea punteada perpendicular al eje de similitud representa el valor crítico(para a = 0,05) obtenido a partir de la distribución nula (n = 100 iteraciones).Similarity dendrogram, considering the abundance of the different species of folivorous insects associated with A. chilensisin the 16 blocks (eight from the continuous forest, BC, and eight from small fragments, F). The perpendicular dotted lineon the similarity axis represents the critical value (for a = 0.05) obtained from the null distribution (n = 100 iterations).
respuesta diferencial, siendo algunas másabundantes en el bosque continuo y otras en losfragmentos. Incluso, dentro del mismo género,algunas especies respondieron de maneradiferente a la fragmentación del bosquemaulino. Por ejemplo, entre los Scarabaeidae,S. obesa fue más abundante en el bosquecontinuo en tanto S. viridis lo fue en losfragmentos; entre los Chrysomelidae,Psathyrocerus sp. 1 fue más abundante en elbosque continuo en tanto P. fulvipes obtuvoabundancias similares en ambos hábitat. Otrasespecies del mismo género como P. pallens yP. pyrroptera fueron más abundantes en losfragmentos de bosque, así como también elElateridae D. pallipes.
La riqueza de especies por árbol tampocofue afectada por la fragmentación del bosque
maulino, aunque en total se encontraron másespecies en el conjunto de fragmentos que enbosque continuo, con 63 y 46 especies,respectivamente. Sin embargo, la composiciónde especies de insectos sí cambió con lafragmentación del bosque. De las 77 especiescapturadas durante el estudio, solo 30 fueroncompartidas por el bosque continuo y losfragmentos, 32 fueron halladas solo en losfragmentos y 14 solo en el bosque continuo.Cuando se analizó la similitud de especiesentre bloques al interior del bosque continuo,entre fragmentos y entre los bloques del bosquecontinuo y fragmentos, según el índice desimilitud Sorensen, que incluye solo lapresencia de especies, la fauna de insectosdefoliadores fue parecida en todas lascomparaciones, compartiéndose en promedio
Encadenamiento promedioDistancia: (euclidiana)

235INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADO
cerca del 70 % de las especies. En tanto segúnel índice de Morisita Horn, que incorporaademás las abundancias relativas de lasespecies, la similitud de la fauna de insectosdefoliadores en general disminuyó en relaciónal índice de Sorensen, sugiriendo que si bien secomparte un alto número de especies, susabundancias relativas varían de un bloque aotro. Además, según este índice, la similitudentre bloques del bosque continuo fue superior(0,17) que cuando estos se comparan con losfragmentos (0,13) y que la similitud entre losfragmentos (0,09). Al hacer el análisis deconglomerados se observa que los bloques BC6y BC8 son muy disímiles al resto de losbloques del bosque continuo. Ello posiblementese deba a la heterogeneidad de hábitat que esposible encontrar en el bosque continuo, consitios más abiertos e iluminados (más parecidosa los fragmentos) que otros. En efecto, elbloque BC8 se encuentra en el borde delbosque continuo.
La menor similitud de especies entre losfragmentos de bosque es consistente con elmayor número total de especies capturadas enel conjunto de los fragmentos, en comparacióncon el bosque continuo, a pesar de que ladiversidad por árbol no varió. Ello sugiere unamayor diversidad gamma (a nivel de paisaje) enlos paisajes fragmentados. Esto se puede deberal arreglo espacial de los fragmentos pequeñosde bosque nativo, los que al estar separados ycubrir un área geográfica mayor que el bosquecontinuo capturan una mayor heterogeneidad dehábitat en el paisaje, mecanismo propuesto porTscharntke et al. (2002). Debido a que pormotivos logísticos en el bosque continuo no fueposible muestrear árboles en un área similar ala abarcada por el conjunto de los fragmentos,no es posible descartar que la mayor diversidadgamma se deba al mayor espaciamiento (yheterogeneidad de hábitat) de los fragmentos, yno consecuencia de la fragmentación.
Los resultados obtenidos en este estudio, enconjunto con otros realizados en la misma zona(Grez et al. 2003, Bustamante-Sánchez et al.2004, Jaña-Prado & Grez 2004), así como losrealizados en otros paisajes fragmentados,indican que los efectos de la fragmentaciónpueden ser variados. La riqueza o abundanciapuede disminuir, mantenerse o inclusoaumentar a medida que el tamaño delfragmento se reduce (Kruess & Tscharntke
2000). Por ejemplo, en Alemania la riqueza deespecies de mariposas se correlacionapositivamente con el tamaño del fragmento(Tscharntke et al. 2002); lo mismo ocurre conla riqueza y abundancia de un ensamble decoleópteros en el bosque fragmentado de FrayJorge, Chile (Barbosa & Marquet 2002), asícomo también en coleópteros carroñeros enbosques fragmentados de la Amazonia y deNueva York (Klein 1989, Gibbs & Stanton2001). Otras evidencias, sin embargo, muestranel efecto contrario, con un aumento en laabundancia y riqueza de especies con lafragmentación. Tal es el caso de lo que ocurreen fragmentos de bosque en Hungría y Ucrania,donde el número total de especies de carábidos(Coleoptera) aumenta en fragmentos máspequeños (Magura et al . 2001). Enagroecosistemas también se ha observado unincremento de la densidad poblacional yriqueza de especies de coleópteros enambientes más fragmentados (Grez et al.2004a, 2004b). Revisiones recientes sobre elefecto de la fragmentación del hábitat muestranque las especies responden de manera diferentey con distinta intensidad a la fragmentación delhábitat debido a sus diferentes estrategias dehistoria de vida (Didham 1997, Ewers &Didham 2006). Especies de niveles tróficossuperiores, especialistas de hábitat, especiescon tamaños corporales grandes y con pocacapacidad de dispersión son más susceptibles ala fragmentación del hábitat y a la extinciónEwers & Didham 2006). Por ello, paracomprender los efectos de la fragmentación delbosque maulino es necesario conocer la historianatural de las especies que habitan este bosquey su importancia en el funcionamientoecosistémico (Jaña-Prado & Grez 2004).
La baja abundancia de insectos defoliadores(con un promedio de solo 13,91 ± 0,87individuos por árbol) coincide con los bajosniveles de herbivoría (12 % de defoliación detejido vegetal) registrados en el bosque maulinoen adultos de A. chilensis (Vásquez et al. 2007)y en plántulas de Cryptocarya alba (Vega2001). Además, el patrón de abundancia de losinsectos herbívoros de más alta representaciónen el bosque maulino es coherente con elpatrón de defoliación descrito para A. chilensisen este bosque. En las primeras cohortes dehojas, al principio de la temporada deexpansión foliar (septiembre), la herbivoría es

236 DE LA VEGA & GREZ
mayor en el bosque continuo que en losfragmentos (Vásquez et al. 2007). Al mismotiempo, según los resultados de este estudio, laabundancia de S. obesa, insecto de gran tamaño(1 cm de largo aprox.) y que remueve grandescantidades de superficie foliar, essignificativamente mayor en el bosque continuoque en los fragmentos. Al avanzar en latemporada, las diferencias en herbivoría entrehábitat desaparecen, junto con el incremento dela abundancia en los fragmentos de insectoscomo los Chrysomelidae y S. viridis. Por ello esmuy probable que estos insectos sean losprincipales responsables de los patrones dedefoliación de A. chilensis en el bosquemaulino. Mediciones posteriores a este estudioconfirman estos patrones de herbivoría yabundancia de insectos herbívoros en el bosquecontinuo y fragmentos de bosque Maulino (X.De la Vega, resultados no publicados).
Todos los insectos defoliadores encontradostanto en el bosque continuo como en losfragmentos corresponden a especies nativas. Lomismo se observó en la fauna de coleópterosasociados a la descomposición de heces deganado bovino (Bustamante-Sánchez et al.2004), como también en los insectos herbívorosasociados al follaje de cuatro especies arbóreas(Jaña-Prado & Grez 2004). En otro trabajorealizado en la zona, solamente dos especies decoleópteros epigeos introducidos, asociados aconíferas exóticas, fueron recolectadas en lasplantaciones de pino en muy baja abundancia ynunca fueron capturadas en los fragmentos delbosque maulino (Grez et al . 2003). Ladominancia de especies nativas y la raraocurrencia de especies de insectos exóticossugiere que, a pesar de la fragmentación delbosque maulino y de la gran extensión ocupadapor las plantaciones de pino en la zona, no haocurrido una llegada masiva y establecimientode especies invasoras y que la entomofaunanativa estaría relativamente bien conservada. Eltipo de matriz que rodea a los remanentes delbosque maulino sin duda ha ayudado a mantenerla fauna. Las plantaciones de pino que están enla zona de la Reserva Nacional Los Queulestienen un manejo que permite el establecimientode un abundante sotobosque mayoritariamentecompuesto por especies nativas, incluida A.chilensis, la que alcanza altas densidades allí.Desde el punto de vista de esta especie, elpaisaje puede ser considerado un continuo. Sin
embargo, a pesar de esta continuidad en ladistribución del maqui, algunos insectosdefoliadores cambian su abundancia en elpaisaje. Esto podría deberse a cambios en lascondiciones abióticas y bióticas producto de lafragmentación, que afectan al maqui y/o a losinsectos defoliadores. Así por ejemplo, se sabeque con la fragmentación del bosque maulino,incrementa la luminosidad y decrece lahumedad del suelo (Henríquez 2002, Burgos etal. 2008), incrementa la densidad de individuos(Lagos 2007), el contenido de nitrógeno y elgrosor de las hojas, y disminuye el área foliarde A. chilensis (Repetto et al . 2007),incrementa la abundancia de aves insectívorasy la insectivoría (González-Gómez et al.2006), todos factores que podrían serrelevantes para las poblaciones de insectosdefoliadores asociados a A. chilensis. Además,con la fragmentación de este bosque cambianlas abundancias relativas de otras especiesarbóreas (Bustamante et al. 2005), lo quetambién podría ser relevante para estosinsectos, considerando que no necesariamenteserían especialistas del maqui.
En general, la fragmentación y pérdida delhábitat son consideradas las mayores amenazaspara la biodiversidad (Myers 1988, Groom &Schumaker 1993, FAO 2002). Estasperturbaciones son cada vez más frecuentes ymuchas veces irreversibles, si no se tomanmedidas para restaurar el paisaje. Por ello, losfragmentos pequeños de bosque son unarealidad. En este estudio mostramos que lafauna de insectos defoliadores asociados a losfragmentos remanentes del bosque maulino nosufre grandes cambios respecto al bosquecontinuo. Además, el 41,5 % de las especies deinsectos defoliadores está solo presente en losfragmentos, e incluso, el conjunto defragmentos soporta una mayor diversidad deespecies nativas que el bosque continuo. Otrosgrupos de insectos nativos, aves, roedorestambién desarrollan abundantes poblaciones enestos fragmentos. Esto sugiere que losfragmentos del bosque maulino, a pesar de sutamaño o aislamiento, son un importantereservorio de biodiversidad nativa y deben serconservados (Grez 2005). Ellos son depropiedad privada, lo que hace necesario unplan de manejo público-privado para asegurarla conservación de este bosque (Simonetti etal. 2002, Simonetti 2006).

237INSECTOS DEFOLIADORES EN EL BOSQUE MAULINO FRAGMENTADO
AGRADECIMIENTOS
Agradecemos a Yuri Zúñiga por su ayuda enterreno, a Mario Elgueta (Museo Nacional deHistoria Natural), Danilo Cepeda (MuseoEntomológico Luis Peña de la Facultad deCiencias Agronómicas de la Universidad deChile) y Viviane Jerez (Universidad deConcepción) en la ayuda prestada en laidentificación de los insectos. Agradecemostambién a Masisa S.A. y CONAF VII Regiónpor permitirnos trabajar en sus terrenos. Estetrabajo fue financiado por FONDECYT1050745.
LITERATURA CITADA
ARNOLD AE & NM ASQUITH (2002) Herbivory in afragmented tropical forest: patterns from islands atLago Gatun, Panama. Biodiversi ty andConservation 11: 1663-1680.
ARTIGAS JN (1994) Entomología económica. Insectos deinterés agrícola, forestal, médico y veterinario(nativos, introducidos y susceptibles de serintroducidos). Editorial Aníbal Pinto, Concepción,Chile. 943 pp.
BARBOSA O & PA MARQUET (2002) Effects of forestfragmentation on the beetle assemblage at the relictforest of Fray Jorge, Chile. Oecologia 132: 296-306.
BENÍTEZ-MALVIDO J, G GARCÍA-GUZMÁN & IDKOSSMANN-FERRAZ (1999) Leaf-fungalincidence and herbivory on tree seedlings intropical rainforest fragments: an experimentalstudy. Biological Conservation 91: 143-150.
BENÍTEZ-MALVIDO J (2001) Regeneration in tropicalrainforest fragments. En: Bierregaard Jr. RO, CGascon, TE Lovejoy & R Mesquita (eds) Lessonsfrom Amazonia: the ecology and conservation of afragmented forest: 136-145. Yale University Press,New Haven, Connecticut, USA.
BRESCIANO D, JA SIMONETTI & AA GREZ (1999)Edge effects in a Mediterranean woodland ofcentral Chile. Journal of Mediterranean Ecology 1:35-40.
BURGOS A, AA GREZ & RO BUSTAMANTE (2008)Seed production, pre-dispersal seed predation andgermination of Nothofagus glauca (Nothofagaceae)in a temperate fragmented forest in Chile. ForestEcology and Management 255: 1226-1233.
BUSTAMANTE RO & C CASTOR (1998) The decline ofan endangered temperate ecosystem: the ruil(Nothofagus alessandrii) forest in central Chile.Biodiversity and Conservation 7: 1607-1626.
BUSTAMANTE RO, JA SIMONETTI, AA GREZ & JSAN MARTÍN (2005) La fragmentación del bosquemaulino y su dinámica regeneracional: diagnósticoactual y perspectivas futuras, pp. 555-564. En:Smith C, JJ Armesto & C Valdovinos (eds)Historia, biodiversidad y ecología de los bosques dela cordillera de la Costa. Editorial Universitaria,Santiago, Chile.
BUSTAMANTE RO, AA GREZ & JA SIMONETTI(2006) Efectos de la fragmentación del bosque
maulino sobre la abundancia y diversidad deespecies nativas. En: Grez AA, JA Simonetti & ROBustamante (eds) Biodiversidad en ambientesfragmentados de Chile: patrones y procesos adiferentes escalas: 83-97. Editorial Universitaria,Santiago, Chile.
BUSTAMANTE-SÁNCHEZ MA, AA GREZ & JASIMONETTI (2004) Dung decomposition andassociated beetles in a fragmented temperate forest.Revista Chilena de Historia Natural 77: 107-120.
CRAWLEY MJ (1985) Reduction of oak fecundity by low-density herbivore populations. Nature 314: 163-164.
DIDHAM RK (1997) An overview of invertebrateresponses to forest fragmentation. En: Watt AD, NEStork & MD Hunter (eds) Forests and insects: 303-320. Chapman and Hall, London, United.Kingdom.
DONOSO DS, AA GREZ & JA SIMONETTI (2004)Effect of forest fragmentation on the granivory ofdifferently-sized seeds. Biological Conservation115: 63-70.
ECHEVERRÍA C, D COOMES, J SALAS, JM REY-BENAYAS, A LARA & A NEWTON (2006) Rapiddeforestat ion and fragmentation of Chileantemperate forests. Biological Conservation 130:481-494.
EWERS R & R DIDHAM (2006) Confounding factors inthe detection of species responses to habitatfragmentation. Biological Reviews 81: 117-142.
FAO (FOOD AND AGRICULTURE ORGANIZATIONOF THE UNITED NATIONS) (2002) Evaluaciónde los recursos forestales mundiales 2000. Roma,Italia. 367 pp.
FORTIN MJ & J GUREVITCH (1993) Mantel tests:spatial structure in field experiments. En: ScheinerSM & J Gurevitch (eds) Design and analysis ofecological experiments: 342-359. Chapman & Hall,New York, USA.
FUENTES ER, J ETCHEGARAY, ME ALJARO & GMONTENEGRO (1981) Shrub defoliation bymatorral insects. En: Di Castri F, DW Goodlall &RL Specht (eds) Mediterranean-type shrublands:345-359. Elsevier, Amsterdam, Holland.
GIBBS JP & EJ STANTON (2001) Habitat fragmentationand arthropod community change: carrion beetles,phoretic mites and flies. Ecological Applications11: 79-85.
GONZÁLEZ-GÓMEZ P, CF ESTADES & JASIMONETTI (2006) Strengthened insectivory in atemperate fragmented forest. Oecologia 148: 137-143.
GREZ AA (2005) El valor de los fragmentos pequeños delbosque maulino en la conservación de la fauna decoleópteros epigeos. En: Smith-Ramírez C, JArmesto & C Valdovinos (eds) Biodiversidad yecología de los bosques de la cordillera de la Costade Chile: 565-572. Editorial Universi taria,Santiago, Chile.
GREZ AA, RO BUSTAMANTE, JA SIMONETTI & LFAHRIG (1997) Landscape ecology, deforestationand habitat fragmentation: the case of the ruil forestin Chile. En: Salinas-Chávez E & J Middleton.(eds) Landscape ecology as a tool for sustainabledevelopment in Latin America. Disponible en:www.brocku.ca/epi/lebk/lebk.html. [consulta: 15/05/2007].
GREZ AA, P MORENO & M ELGUETA (2003)Coleópteros (Insecta, Coleoptera) epigeos asociadosal bosque maulino y plantaciones de pino aledañas.Revista Chilena de Entomología 29: 9-18.

238 DE LA VEGA & GREZ
GREZ AA, T ZAVIEZO & S REYES (2004a) Short-termeffects of habitat fragmentation on the abundanceand species richness of beetles in experimentalalfalfa micro-landscapes. Revista Chilena deHistoria Natural 77: 547-558.
GREZ AA, T ZAVIEZO, L TISCHENDORF & L FAHRIG(2004b) A transient, positive effect of habitatfragmentation on insect population densities.Oecologia 141: 444-451.
GROOM MJ & N SCHUMAKER (1993) Evaluatinglandscape change: pattern of worldwidedeforestation and local fragmentation. En: KareivaPM, JG Kingsolver & RB Huey (eds) Bioticinteractions and global change: 24-44. Sinauer,Sunderland, Massachusetts, USA.
HENRÍQUEZ C (2002) El dilema de Lapageria rosea enbosques fragmentados: ¿cantidad o calidad de laprogenie? Tesis Doctoral, Facultad de Ciencias,Universidad de Chile, Chile. 133 pp.
HOCHWENDER CG, VL SORK & RJ MARQUIS (2003)Fitness consequences of herbivory on Quercus alba.American Midland Naturalist 150: 246-253.
INFOSTAT (2003) InfoStat versión 1.5. Grupo InfoStat,FCA, Universidad Nacional de Córdoba, Argentina.
JAÑA-PRADO R & AA GREZ (2004) Insectos herbívorosen el bosque maulino: un ecosistema fragmentado.Revista Chilena de Entomología 30: 27-43.
KLEIN BC (1989) Effects of forest fragmentation on dungand carrion beetle communities in centralAmazonia. Ecology 70: 1715-1725.
KREBS CJ (1989) Ecological methodology. Harper andRow, New York, New York, USA. 654 pp.
KRUESS A & T TSCHARNTKE (2000) Species richnessand parasit ism in a fragmented landscape:experiments and field studies with insects on Viciasepium. Oecologia 122: 129-137.
LAGOS N (2007) Distribución de especies arbóreas en unbosque maulino fragmentado. Memoria de Título,Facultad de Ciencias Agronómicas, Universidad deChile, Santiago, Chile. 53 pp.
MAGURA T, V KODOBOCZ & B TÓTHMÉRÉSZ (2001)Effects of habitat fragmentation on carabids inforest patches. Journal of Biogeography 28: 129-138.
MORENO CE (2001) Métodos para medir labiodiversidad. M&T- Manuales y tesis SEA,Volumen 1. España. 84 pp.
MYERS N (1988) Tropical forest and their species. Going,going…? En: Wilson EO (ed) Biodiversity: 28-35.Academic Press, New York, New York, USA.
MYERS N, RA MITTERMEIER, CG MITTERMEIER,GAB DA FONSECA & J KENT (2000)Biodiversity hotspots for conservation priorities.Nature 403: 853-858.
NOSS RF (1990) Indicators for monitoring biodiversity: ahierarchical approach. Conservation Biology 4:355-364.
POPTOOLS (2006) Versión 2.7.5, Austral ia:www.cse.csiro.au/poptools/
REPETTO-GIAVELLI F, LA CAVIERES & JASIMONETTI (2007) Respuestas foliares de
Aristotelia chilensis a la fragmentación del bosquemaulino. Revista Chilena de Historia Natural 80:469-477.
RICKLEFS R & G MILLER (1999) Ecology. Fourthedition. Freeman and Company, New York, NewYork, USA. 481 pp.
ROLAND J (1993) Large-scale forest fragmentationincreases the duration of tent caterpillar outbreak.Oecologia 93: 25-30.
SAAVEDRA B & JA SIMONETTI (2001) New records ofDromiciops gliroides (Marsupialia:Microbiotheri idae) and Geoxus valdivianus(Rodentia: Muridae) in central Chile: theirimplications for biogeography and conservation.Mammalia 65: 96-100.
SAN MARTÍN J & C DONOSO (1996) Estructuraflorística e impacto antrópico en el bosque maulinoy plantaciones de pino aledañas. En: Armesto J, CVillagrán & M Arroyo (eds) Ecología de losbosques nativos de Chile: 153-168. EditorialUniversitaria, Santiago, Chile.
SIMONETTI JA (2006) Conservación de biodiversidad enambientes fragmentados: el caso del bosquemaulino. En: Grez AA, JA Simonetti & ROBustamante (eds) Biodiversidad en ambientesfragmentados de Chile: patrones y procesos adiferentes escalas: 215-229. Editorial Universitaria,Santiago, Chile.
SIMONETTI JA, AA GREZ & RO BUSTAMANTE(2002) El valor de la matriz en la conservaciónambiental. Ambiente y Desarrollo (Chile) 18: 116-118.
SIMONETTI JA, AA GREZ & RO BUSTAMANTE(2006) Interacciones y procesos en el bosquemaulino fragmentado. En: Grez AA, Simonetti JA& RO Bustamante (eds). Biodiversidad enambientes fragmentados de Chile: patrones yprocesos a diferentes escalas: 99-114. EditorialUniversitaria, Santiago, Chile.
SIMONETTI JA, AA GREZ, JL CELIS-DIEZ & ROBUSTAMANTE (2007) Herbivory and seedlingperformance in a fragmented temperate forest ofChile. Acta Oecologica 32: 312-318.
STATSOFT (2001) Statistica data analysis softwaresystem, version 6., Statsoft. Inc., Tulsa, Oklahoma,USA; <www.statsoft.com> [consulta: 08/11/2006].
TSCHARNTKE T, I STEFFAN-DEWENTER, A KRUESS& C THIES (2002) Contribution of small habitatfragments to conservation of insects communities ofgrassland-cropland landscapes. EcologicalApplications 12: 354-363.
VÁSQUEZ PA, AA GREZ, RO BUSTAMANTE & JASIMONETTI (2007) Herbivory, foliar survival andshoot growth in fragmented populations ofAristotelia chilensis. Acta Oecologica 31: 48-53.
VEGA MI (2001) Efecto de la fragmentación del bosquemaulino sobre la herbivoría en plántulas deCryptocarya alba. Memoria de Título, MedicinaVeterinaria, Facultad de Ciencias Veterinarias yPecuarias, Universidad de Chile, Santiago, Chile.79 pp.
Editor Asociado: Julio GutiérrezRecibido el 19 de abril de 2007; aceptado el 10 de enero de 2008