
Compartmentation of oxidative stress defense in...
56
Compartmentation of oxidative stress defense in mitochondria: Implications of drug toxicity Dean P. Jones, Ph.D. Department of Medicine/Division of Pulmonary, Allergy and Critical Care Medicine Emory University, Atlanta Emory Clinical Biomarkers Laboratory EMORY SCHOOL OF MEDICINE
Transcript of Compartmentation of oxidative stress defense in...

Compartmentation of oxidative stress
defense in mitochondria:
Implications of drug toxicity
Dean P. Jones, Ph.D.Department of Medicine/Division of Pulmonary, Allergy
and Critical Care Medicine
Emory University, Atlanta
Emory Clinical Biomarkers Laboratory
EMORYSCHOOL OF
MEDICINE

What actually
happens?What is possible?
Redox Systems Biology applies quantitative principles to the
possibilities within the context of the entire biologic system
Critical transition for mechanistic toxicology:
translation of possibilities (done in vitro in a short time)
into realities (what happens in humans over decades)

Critical transition for mechanistic toxicology: translation of possibilities (done in vitro in a short time)into realities (what happens in humans over decades)

A small fraction of the mitochondrial electron currency
causes toxicity
DP Jones, Chem-Biol Interact 2006
NADH
CoQ
Cyt c
O2
-400
-200
0
+200
+400
+600
Metabolic Redox Circuits
ATP production
Eh
Pyr
Mal
Succinate
MnSOD
Radical-mediated
Macromolecular
damage

A small fraction of the mitochondrial electron currency
causes toxicity
DP Jones, Chem-Biol Interact 2006
NADH
CoQ
Cyt c
O2
-400
-200
0
+200
+400
+600
Metabolic Redox Circuits
ATP production
Eh
Pyr
Mal
Succinate
MnSOD
Radical-mediated
Macromolecular
damage
Concept of ROS as
unavoidable consequence
of aerobic life

A small fraction of the mitochondrial electron currency is
used for signaling and control functions
DP Jones, Chem-Biol Interact 2006
NADH
CoQ
Cyt c
O2
GR
GSH
-400
-200
0
+200
+400
+600
Redox Signaling and Control Circuits
(low flux)
Metabolic Redox Circuits
(high flux)
ATP production
Eh
Pyr
Mal
SuccinateMPT
TR2
Trx2
O2
NADPH
GPx Prx3
H2O2
PrSSGGrx2
Metabolic
substrates
ASK1
MnSOD
Non-Radical
mechanisms of
redox signaling
and control

A small fraction of the mitochondrial electron currency is
used for signaling and control functions
DP Jones, Chem-Biol Interact 2006
NADH
CoQ
Cyt c
O2
GR
GSH
-400
-200
0
+200
+400
+600
Redox Signaling and Control Circuits
(low flux)
Metabolic Redox Circuits
(high flux)
ATP production
Eh
Pyr
Mal
SuccinateMPT
TR2
Trx2
O2
NADPH
GPx Prx3
H2O2
PrSSGGrx2
Metabolic
substrates
ASK1
MnSOD
Non-Radical
mechanisms of
redox signaling
and control
ROS are products of
respiration which
function in signaling
and control

Imbalance in prooxidant and
antioxidant reactions
Macromolecular
damageDisruption of thiol redox
signaling and control
Free Radicals Non-radical Oxidants
Free radical mechanisms
of oxidative stressNon-radical mechanisms
of oxidative stress
1000
900100
9991
99
Efficient
scavenging
Jones 2008 “Radical-Free Biology of Oxidative Stress” AJP Cell Physiol 295:849-868

H3C COOH
H3C
H3C
H3C CO
H3C
H3C
+ CH3
C = O
H3C
H3C
H3C C OH
H3C
H3C
Acetone
t-Butanol
t-Butylhydroperoxide
Methyl radical
2 e-
1 e-
99.98%
0.02%
Tribble et al, Mol Pharm 1988
TBH toxicity involves non-radical mechanisms affecting
mitochondria

Redox Signaling
(Physiologic)
Redox Sensor
Sulfur Switches
ROS/RNS
Redox Messenger
Macromolecular
Targets
ROS/RNS
Toxic Species
Imbalance between
Oxidants/Antioxidants
NADPH oxidase
NO synthase
Oxidative Stress
(Toxicologic)
PhysiologicToxicologic
Thiol redox sensors are sensitive targets of non-radical oxidants

Redox Signaling
(Physiologic)
Redox Sensor
Sulfur Switches
ROS/RNS
Redox Messenger
Macromolecular
Targets
ROS/RNS
Toxic Species
Imbalance between
Oxidants/Antioxidants
NADPH oxidase
NO synthase
Oxidative Stress
(Toxicologic)
PhysiologicToxicologic
Thiol redox sensors are sensitive targets of non-radical oxidants
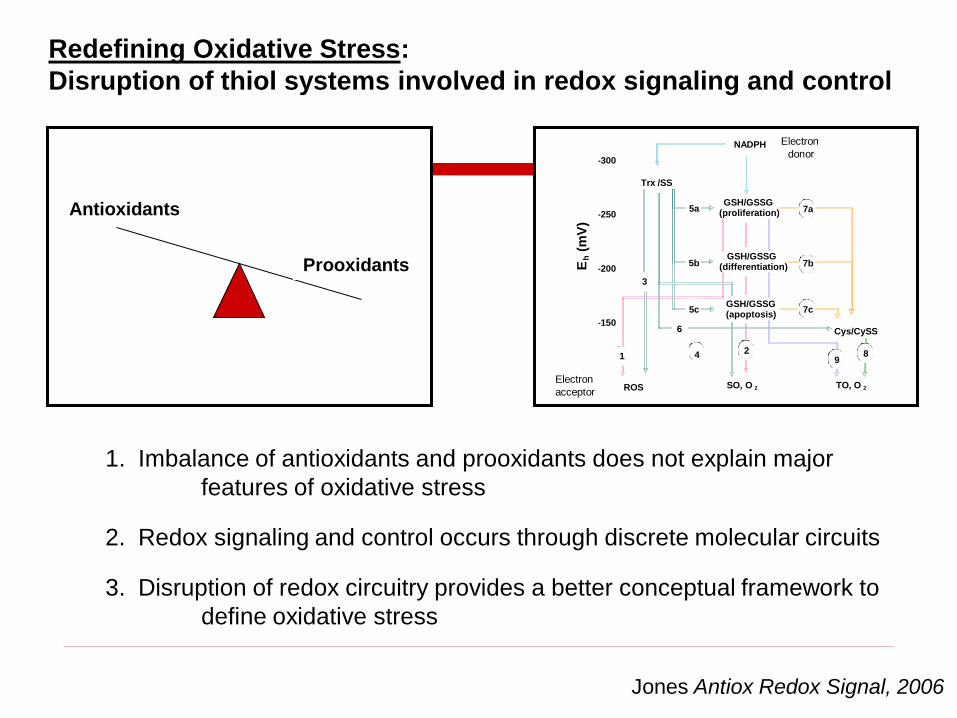
Prooxidants
Antioxidants
1. Imbalance of antioxidants and prooxidants does not explain major
features of oxidative stress
Jones Antiox Redox Signal, 2006
2. Redox signaling and control occurs through discrete molecular circuits
3. Disruption of redox circuitry provides a better conceptual framework to
define oxidative stress
5c
GSH/GSSG(differentiation)
12
9
7a
7b
7c
8
Eh
(mV
)
GSH/GSSG(apoptosis)
4
GSH/GSSG(proliferation)
Trx /SS
-300
-200
-150
-250
-300
-200
-150
-250
Cys/CySS
ROSROS SO, O 2 TO, O 2
3
6
5a
5b
5c
GSH/GSSG(differentiation)
112
9
7a7a
7b7b
7c7c
8
h(m
V)
GSH/GSSG(apoptosis)
4
Electron
donor
Electron
acceptor
NADPH
Redefining Oxidative Stress:
Disruption of thiol systems involved in redox signaling and control

Jones Meth Enzymol 2002
GSHGSSG
Redox states of different couples can be compared in terms of
Nernst redox potentials, i.e., balance can be measured
Eh = Eo + * ln[Ox]_
[Red]
RT
nF
DTT H2O2 0 24 48 h
Ox1
Ox2
Red
Go et al 2007
Trx-1
Calculate from GSH and GSSG, Cys and CySS or redox western
blots of proteins (thioredoxins, PDI, etc)

Biologic Redox Potentials: Steady-state Eh
values from about -400 to +600 mV
-400 0 +200 +600
Most reduced Most oxidized
Mitochondrial ET chain

Biologic Redox Potentials: Steady-state Eh
values from about -400 to +600 mV
-400 0 +200 +600
Most reduced Most oxidized
Mitochondrial ET chain
GSH/GSSG

Biologic Redox Potentials: Steady-state Eh
values from about -400 to +600 mV
-400 0 +200 +600
Most reduced Most oxidized
Mitochondrial ET chain
Pr-SH
GSH/GSSG

Biologic Redox Potentials: Steady-state Eh
values from about -400 to +600 mV
-400 0 +200 +600
Most reduced Most oxidized
Mitochondrial ET chain
Pr-SH
GSH/GSSG
NA
DP
H

NADPH/NADP+
mtGSSG reductase
Prx5 Ask1Prx3 PT pore,
other proteinsGrx2mtGpx1 mtGpx4
NDH, other
proteins
Cyt c
ROOH ROOH ROOHROOH ROOH
GSSGGSH
Trx2(SH)2 Trx2(SS)
TrxR2(red) TrxR2(ox)
Mitochondrial Thiol/Disulfide Redox Pathways

Reduced f
orm
(%
)
100
50
0
TR
2
Trx
2
Prx
3
NA
DP
H
-Glc,-Gln
0 24 h
TrxR2Red
Prx3Red
Trx2Red
Trx2Ox
Reduction
by TCEPTrxR2Red
Vs.TrxR2Red
/totTrxR2
Trx2Red/Trx2Ox
Reduction
by TCEPPrx3Red
Prx3Red
/totPrx3Vs.
Quantitation by redox western blot
100
50
0
TR
2
Trx
2
Prx
3
NA
DP
H
-Glc,-Gln
Reduced f
orm
(%
)
0 24h
Thiol/disulfide redox control pathway
NADPH TrxR2 Trx2 Prx3 H2O2
Redox states of components in pathway provide
information on sites of rate limitation

Reduced f
orm
(%
)
100
50
0
TR
2
Trx
2
Prx
3
NA
DP
H
-Glc,-Gln
0 24 h
TrxR2Red
Prx3Red
Trx2Red
Trx2Ox
Reduction
by TCEPTrxR2Red
Vs.TrxR2Red
/totTrxR2
Trx2Red/Trx2Ox
Reduction
by TCEPPrx3Red
Prx3Red
/totPrx3Vs.
Quantitation by redox western blot
100
50
0
TR
2
Trx
2
Prx
3
NA
DP
H
-Glc,-Gln
Reduced f
orm
(%
)
0 24h
Thiol/disulfide redox control pathway
NADPH TrxR2 Trx2 Prx3 H2O2
v2 v3 v4 v5
Rate
limitation
v1

Trx2 and GSH support parallel, non-redundant functions
in mitochondria
Via
bili
ty (
%)
Mock WT C93S
tBH
Mock WT C93S Mock WT C93S*
TNF TNF +BSO
**
0
20
40
60
80
*
*
*
Trx2C93S to inhibit Trx2, BSO to deplete GSH

Nuc Trx1
Oxidized
Mitochondrial are the most
reduced compartment in cells;
most susceptible to oxidation
Ste
ady-
Sta
te R
edox P
ote
ntial
(Eh
, m
V)
Reduced
-210
-180
-150
-120
-90
-60
-330
-300
-270
-240
-360Mito Trx2
Mito GSH/GSSG
Cyto GSH/GSSG
Cyto Trx1
Cyto Cys/CySS
Plasma GSH/GSSG
Plasma Cys/CySS
ER GSH/GSSG
Chen et al 2006
Kemp et al, 2008; Jones, 2008
µM tBH0 50
Trx1R
Trx1O
Trx2R
Trx2O
Y Chen et al FEBS Lett 2006
Mito
Cyto
143B cells
+ t-Butyl Hydroperoxide

Nuc Trx1
Oxidized
Mitochondrial are the most
reduced compartment in cells;
most susceptible to oxidation
Ste
ady-
Sta
te R
edox P
ote
ntial
(Eh
, m
V)
Reduced
-210
-180
-150
-120
-90
-60
-330
-300
-270
-240
-360Mito Trx2
Mito GSH/GSSG
Cyto GSH/GSSG
Cyto Trx1
Cyto Cys/CySS
Plasma GSH/GSSG
Plasma Cys/CySS
ER GSH/GSSG
Trx-1
(Cyto)
xTrx-2
(Cyto)
Trx-2
(Mito)
H2O2 ( M)
0 20 50 100 200 300
Chen et al 2006
Kemp et al, 2008; Jones, 2008
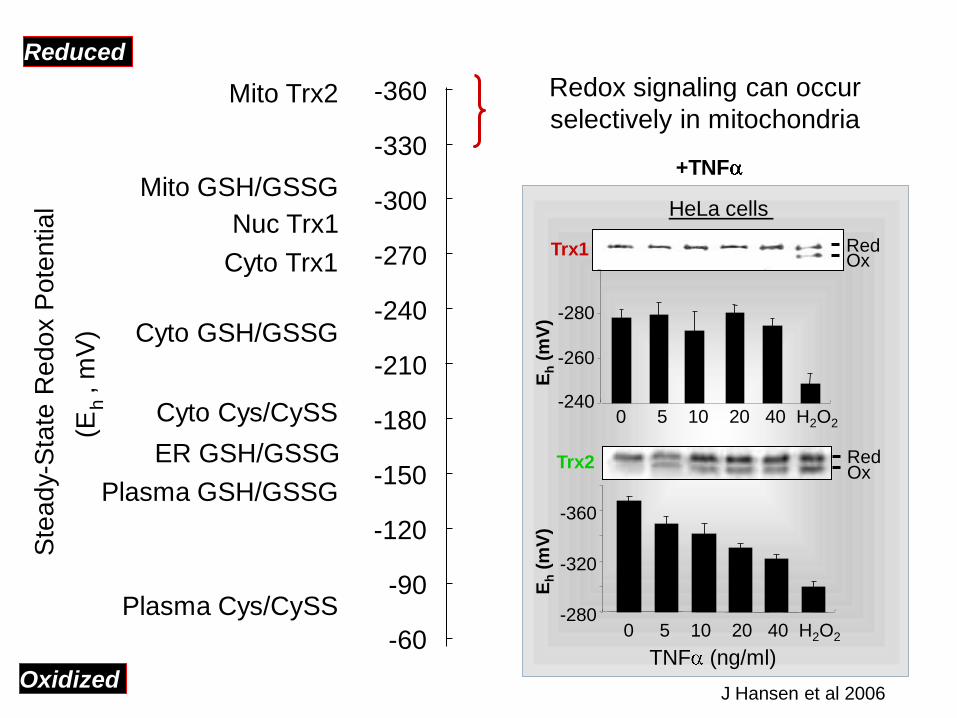
Nuc Trx1
Oxidized
Redox signaling can occur
selectively in mitochondria
Ste
ady-
Sta
te R
edox P
ote
ntial
(Eh
, m
V)
Reduced
-210
-180
-150
-120
-90
-60
-330
-300
-270
-240
-360Mito Trx2
Mito GSH/GSSG
Cyto GSH/GSSG
Cyto Trx1
Cyto Cys/CySS
Plasma GSH/GSSG
Plasma Cys/CySS
ER GSH/GSSG
TNF (ng/ml)
HeLa cells
Trx1
-280
-260
-240
Eh
(mV
)
Red
0 5 10 20 40 H2O2
Ox
Trx2
0 5 10 20 40 H2O2
-360
-320
-280
RedOx
Eh
(mV
)
J Hansen et al 2006
+TNF

Mitochondrial oxidative stress: Disruption of thiol signaling
and control functions by non-radical oxidants
DP Jones, Chem-Biol Interact 2006
NADH
CoQ
Cyt c
O2
GR
GSH
-400
-200
0
+200
+400
+600
Redox Signaling and Control Circuits
(low flux)
Metabolic Redox Circuits
(high flux)
ATP production
Eh
Pyr
Mal
SuccinateMPT
TR2
Trx2
O2
NADPH
GPx Prx3
H2O2
PrSSGGrx2
Metabolic
substrates
ASK1
MnSOD
Radical-mediated
Macromolecular
damage
Non-Radical
mechanisms:
Altered signaling
and control
Minor
Major

tBH None Mal
Red
OxTrx2
Deficient precursors for NADPH supply causes greater
oxidation of Trx2 than mitochondrial respiratory inhibitors
None AA
Trx
2, E
h (m
V)
Media deficient in
Energy precursors
None AA
-360
-320
-280
-240
Control media
Isolated
Mitochondria
Cells

DP Jones, Chem-Biol Interact 2006
NADH
CoQ
Cyt c
O2
GR
GSH
-400
-200
0
+200
+400
+600
Redox Signaling and Control Circuits
(low flux)
Metabolic Redox Circuits
(high flux)
ATP production
Eh
Pyr
Mal
SuccinateMPT
TR2
Trx2
O2
NADPH
GPx Prx3
H2O2
PrSSGGrx2
Metabolic
substrates
ASK1
MnSOD
Radical-mediated
Macromolecular
damage
Metabolic
substrate
availability
Minor
Major
Inhibition of
respiratory
chain
X
X
Deficient precursors for NADPH supply may be more
critical for toxicity than inhibition of respiration

N Adimora et al (2010) ARS in press
Computational model for metabolism of exogenously added H2O2
100 µM
Jurkat cells, 28 kinetic parameters, 24 molecular species
Catalase
<1%90 nM
Gpx
44%
Prx
32%
Protein
oxidation
23%
% Flux of peroxide
GS-
<1%
-SS-
22%

Redox Signaling
(Physiologic)
Redox Sensor
Sulfur Switches
ROS/RNS
Redox Messenger
Macromolecular
Targets
ROS/RNS
Toxic Species
Imbalance between
Oxidants/Antioxidants
NADPH oxidase
NO synthase
Oxidative Stress
(Toxicologic)
PhysiologicToxicologic
The human genome encodes 214,000 distinct Cys residues
Jones 2008 “Radical-Free Biology of Oxidative Stress” AJP Cell Physiol 295:849-868
Estimated 2000 function in redox signaling and control

Modify
Thiols-L
Redox Proteomics using ICAT-LC/MSMS
Thiol/disulfide
mixtureModify
Thiols-H
Reduce
disulfides
Trypsin
cleavage
Mass spectrometry
m/z
Inte
nsity H
L

Extracellular Eh
Protein Name Peptide Sequence -150 mV 0 mV
Protein metabolism Elongation factor (EF)1- SGDAAIVDOVPGKPMCVESFSDYPPLGR 0.2741 0.1903
DGNASGTTLLEALDCILPPTRPTDKPLR 0.8248 0.3356
NOITGTSQADCAVLIVAAGVGEFEAGISK 1.5961 0.8464
EF-2 ETVSEESNVLCLSK 1.4843 0.9473
EGALCEENOR 1.4196 1.2931
Glu-tRNA synthetase EAPCVLIYIPDGHTK 0.5893 0.3745
Ubiquitin-activating E1 DNPGVVTCLDEAR 0.3381 0.4276
Cofilin HELQANCYEEVK 0.6281 0.2798
Calregulin HEQNIDCGGGIVK 1.6932 1.0689
Signal transduction Tyrosine phosphatase YPLNCSDPTSER 0.2983 0.2843
14-3-3 protein YLAEVACGDDR 0.8484 0.6631
Detoxification Peroxiredoxin 6 DFTPVCTTELGR 0.6783 0.5088
GSH-S-transferase CLDEFPNLK 0.6678 0.4968
Stress Response Hsp 60 AAVEEGIVLGGGCALLR 0.3321 0.2663
CEFQDAYVLLSEK 0.8389 0.2601
CIPALDSLTPANEDQK 0.7340 0.3750
Hsp 90 GFEVVYOTEPIDEYCVOOLK 0.5092 0.3261
Cell Structure/Motility Tubulin EIVHIQAGQCGNQIGAK 0.5351 0.2362
NOOAACDPR 0.6334 0.3395
-tubulin AYHEQLSVAEITNACFEPANQMVK 0.7390 0.3242
ICAT-MS/MS redox proteomics gives ability to measure
percent reduction of 1% of Cys proteome
Y.-M.-Go

Studies of Mitochondrial Redox Proteome
using redox ICAT-mass spectrometry
What happens to mitochondrial proteins with steady-state
generation of H2O2 at low rates in culture medium?
What happens to mitochondrial proteins with
oxidized extracellular Cys/CySS redox potential?

Percent oxidation was determined for peptides representing over 1000
proteins
Total cell protein peptidyl Cys oxidation was measured by Redox ICAT; for
2D LC-MSMS methods, see Go and Jones, 2008; Go and Jones 2009
HT 29 cells were treated with Glc oxidase in culture media to generate
H2O2 at a rate about 1 to 10% of the cellular O2 consumption
Mitochondrial proteins are relatively sensitive to oxidation
Y-M Go and DP Jones, unpublished
Proteins were sorted for subcellular localization, with over 500 proteins
assigned to either cytoplasm, nuclei, mitochondria or plasma membrane
Average mitochondrial protein oxidation was 2-fold
No significant average oxidation occurred in other compartments

Studies of Mitochondrial Redox Proteome
using redox ICAT-mass spectrometry
What happens to mitochondrial proteins with steady-state
generation of H2O2 at low rates in culture medium?
What happens to mitochondrial proteins with oxidized
extracellular Cys/CySS redox potential?

Extracellular Eh Cys/CySS is regulated by human
cells in culture to the value found in human plasma
Jonas et al, FRBM 2002
Extr
acellu
lar
Eh
(Cys/C
yS
S)
(mV
)
0 4 8 12 16 20 24
Time (h)
-120
-100
-80
-60
-40
-20
+200 M Cysteine
+100 M Cystine
CaCo2 cells
Reduced
Oxidized
Go and Jones, Circulation 2005
0-50-100-150-200
120
80
40
0
Eh (mV)
Eh, Cys/CySS
Mean=-72.4
Eh, GSH/GSSG
Mean=-130.9
Fre
quency
Plasma, 740 subjects
Young healthy in RED
OxidizedReduced

Plasma ECys in humans is oxidized in association
with aging, oxidative stress and disease
-400 +600 mV
Most reduced Most oxidized
Aging
Oxidative stress
Chronic disease
ECys = -80 mV
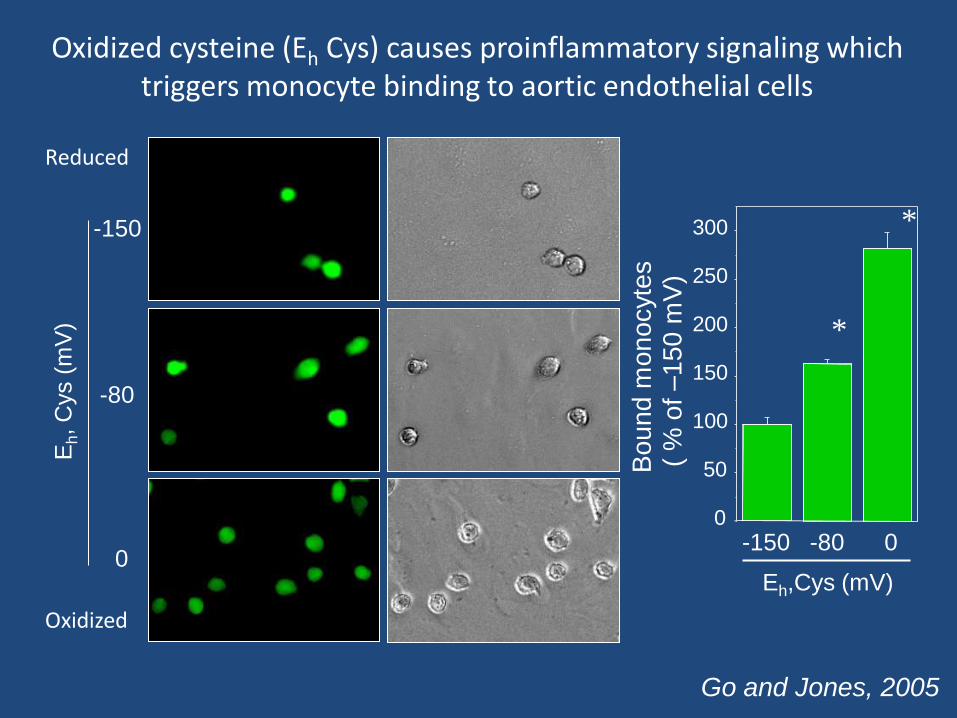
-150
-80
0
Eh, C
ys (
mV
)
Eh,Cys (mV)
-150 -80 0
Bound m
onocyte
s(
% o
f –150 m
V)
0
50
100
150
200
250
300
*
*
Oxidized cysteine (Eh Cys) causes proinflammatory signaling which triggers monocyte binding to aortic endothelial cells
Reduced
Oxidized
Go and Jones, 2005

-150 -80 0 mV
DC
F f
luo
resce
nce
(% o
f –
15
0 m
V)
0
50
100
150
200
250
300
*
Eh, Cys
*
Go and Jones, Circulation 2005
Oxidized extracellular Cys/CySS redox
stimulates H2O2* generation*Signal is blocked by catalase

EhCys/CySS
HS-Pr/
(SS)-PrKinase/
PhosphataseH2O2
IκB
NF-κBNF-κB
DNA binding
Redox
sensor
Transcription
Translation
Protein processing
In secretory pathwayCAMs
Cytoskeletal/Plasma
membrane InteractionsIntegrins
NucleusCytoplasmPlasma
Plasma Cys/CySS redox potential signals H2O2 production
and downstream mechanisms for monocyte adhesion
Go and Jones, Circulation 2005
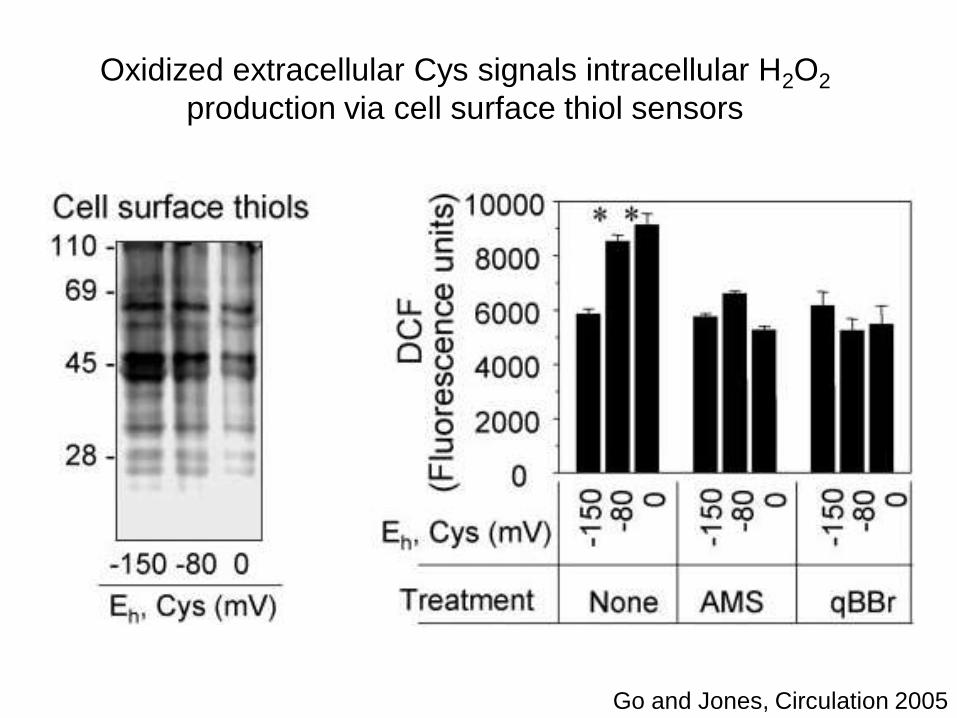
Oxidized extracellular Cys signals intracellular H2O2
production via cell surface thiol sensors
Go and Jones, Circulation 2005
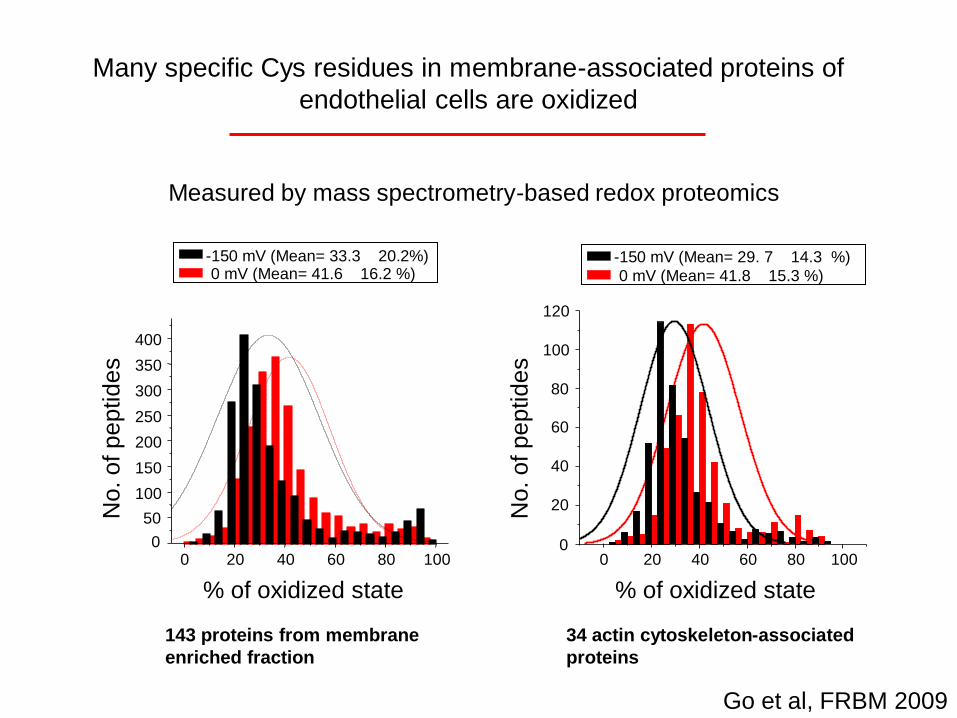
No. o
f p
ep
tid
es
% of oxidized state
A
143 proteins from membrane
enriched fraction
0 mV (Mean= 41.6 16.2 %)-150 mV (Mean= 33.3 20.2%)
0
50
100
150
200
250
300
350
400
0 20 40 60 80 1000
20
40
60
80
100
120
0 20 40 60 80 100
% of oxidized state
No. o
f p
ep
tid
es
34 actin cytoskeleton-associated
proteins
0 mV (Mean= 41.8 15.3 %)
-150 mV (Mean= 29. 7 14.3 %)
B
Go et al, FRBM 2009
Many specific Cys residues in membrane-associated proteins of
endothelial cells are oxidized
Measured by mass spectrometry-based redox proteomicsMeasured by mass spectrometry-based redox proteomics

-150 -80 0 DTT H2O2
EhCys (mV)
WTTrx2Red
Trx2Ox
Go et al, FRBM 2009
Trx2 is oxidized by
extracellular Eh at 0 mV
NADPH TR Trx2 Prx3 H2O2 metabolismNo effect on cytoplasmic GSH
EhCys/CySS
HS-Pr/
(SS)-PrKinase/
PhosphataseMito
Redox
sensor
Oxidation
Glu
tath
ione
Eh
(mV
)
(2G
SH
/GS
SG
)
-150
-200
-250
-300
EhCys (mV) :
WT
-150
-80 0
0.0
0.4
0.88
12
16
18
(mM
)

-150 -80 0 DTT H2O2
EhCys (mV)
WT
Tg
Trx2Red
Trx2Ox
Trx2Red
Trx2Ox
Trx1Red
NLS-Trx1Red
NLS-Trx1Ox
-150 -80 0 DTT H2O2
EhCys (mV)
Trx1Ox
Glu
tath
ione
Eh
(mV
)
(2G
SH
/GS
SG
)
-150
-200
-250
-300
EhCys (mV) :
WT Tg
-150
-80 0
-150
-80 0
GSHGSSG
0.0
0.4
0.88
12
16
18
(mM
)
WT TgTg
MAECTail
V5-hTrx2
mTrx2
mTrx1
WT Tg
PCR product
of v5-hTrx2
V5-hTrx2
Tg
WT
Go et al, FRBM 2009
Trx2 is oxidized by
extracellular Eh at 0 mV
NADPH TR Trx2 Prx3 H2O2 metabolism
Signaling is blocked in
cells from Trx2 Tg mice

EhCys/CySS
HS-Pr/
(SS)-PrKinase/
PhosphataseH2O2Mito
IκB
NF-κBNF-κB
DNA binding
Redox
sensor
Transcription
Translation
Protein processing
In secretory pathwayCAMs
Cytoskeletal/Plasma
membrane InteractionsIntegrins
NucleusCytoplasmPlasma
Plasma Cys/CySS redox potential signals mitochondrial H2O2
production and downstream mechanisms for monocyte adhesion

A B
EhCys (mV) :
WT Tg
MitoS
OX
(% o
f m
inim
um
)
-150
-80 0
-150
-80 0
1
2
3
50
75
100
125
150
MitoS
OX
(% o
f m
inim
um
)
EhCys (mV) :
-150
-80 0
-150
-80 0
-150
-80 0
None qBBr AMS
C 4
5
6
4
56
*
75
100
125
150
175
200
1
2
3
**
*
N.S
80
100
120
140**
*D
CF
(% o
f m
inim
um
)
EhCys (mV) : -150 -80 0 -150 -80 0
WT Tg
Go et al, FRBM 2009

EhCySS-Dependent Redox Changes in Mitochondrial Proteins are Involved in
Amino Acid Metabolism, Molecular Transport, and Small Molecule Biochemistry
Network 1
Mitochondria proteins from MAEC treated with EhCySS, 0 mV

EhCySS-dependent redox changes in Mitochondrial Proteins are involved in
Cellular Assembly and Organization, Cell Death, and Liver Necrosis
Network 2
Mitochondria proteins from MAEC treated with EhCySS, 0 mV

EhCySS-dependent redox changes in Mitochondrial Proteins are involved in
Genetic Disorder, Metabolic Disease, and Free Radical Scavenging
Network 3
Mitochondria proteins from MAEC treated with EhCySS, 0 mV

EhCys
O2∸
H2O2
Trx2 oxidation
PM
Cell adhesion molecules
Cytokine (IL-1 )
Inflammation, vascular disease
OxidationReduction
-150 mV 0 mV
H2O2
Nrf2
H2O2
NF- B
Antioxidants
(NQO1, MAF, FTH1)
Pr-SH Pr-SSPr-SH
MAPK PDGF
Early growth response genes
Eukaryotic translational initiation factor
Cell growth, proliferation Detoxification
Combination of Gene Expression Array and Proteomic Analyses
Extracellular EhCys-dependent cell signaling and control
Go et al Tox Sci 2009

EhCys/CySS
HS-Pr/
(SS)-PrKinase/
PhosphataseH2O2Mito
IκB
NF-κBNF-κB
DNA binding
Redox
sensor
Transcription
Translation
Protein processing
In secretory pathwayCAMs
Cytoskeletal/Plasma
membrane InteractionsIntegrins
NucleusCytoplasmPlasma
Plasma Cys/CySS redox potential signals mitochondrial H2O2
production and downstream mechanisms for monocyte adhesion
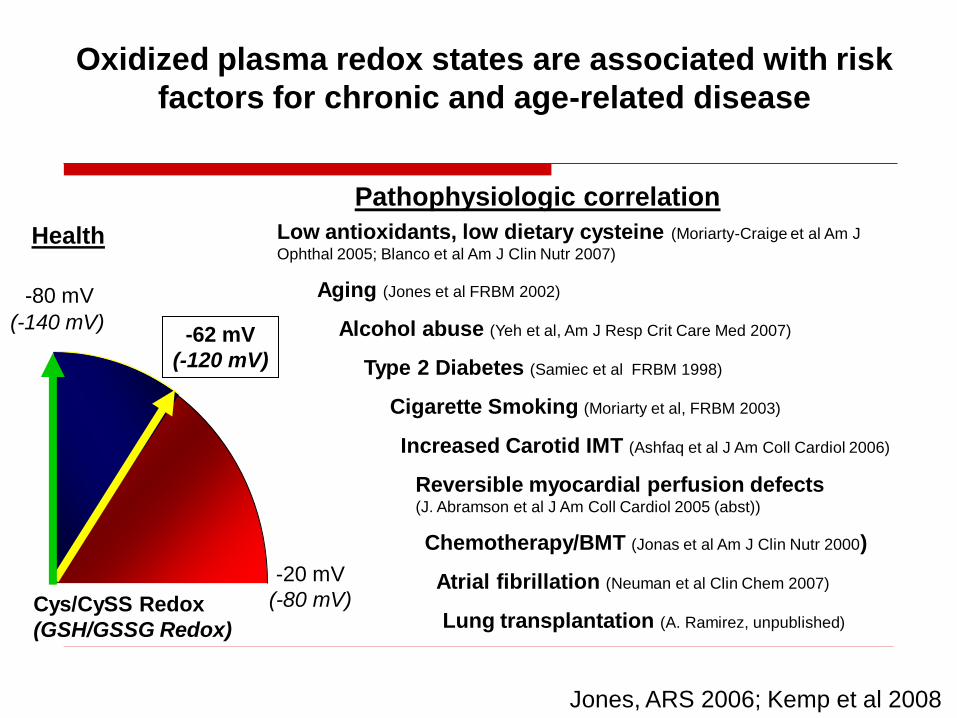
Increased Carotid IMT (Ashfaq et al J Am Coll Cardiol 2006)
Chemotherapy/BMT (Jonas et al Am J Clin Nutr 2000)
Cigarette Smoking (Moriarty et al, FRBM 2003)
Type 2 Diabetes (Samiec et al FRBM 1998)
Reversible myocardial perfusion defects (J. Abramson et al J Am Coll Cardiol 2005 (abst))
Pathophysiologic correlation
Low antioxidants, low dietary cysteine (Moriarty-Craige et al Am J
Ophthal 2005; Blanco et al Am J Clin Nutr 2007)
Jones, ARS 2006; Kemp et al 2008
Atrial fibrillation (Neuman et al Clin Chem 2007)
Alcohol abuse (Yeh et al, Am J Resp Crit Care Med 2007)
Aging (Jones et al FRBM 2002)
Oxidized plasma redox states are associated with risk
factors for chronic and age-related disease
Health
-80 mV
-20 mV
(-80 mV)Cys/CySS Redox
(GSH/GSSG Redox)
(-140 mV)-62 mV
(-120 mV)
Lung transplantation (A. Ramirez, unpublished)

Oxidized Eh for GSH and Cys are associated
with persistent atrial fibrillation
Neuman et al , 2007
Odds ratio
Eh GSH*
Eh Cys*
dROMs*
hsCRP
TNFα
IL-1β
IL-6
1 10 100 1000
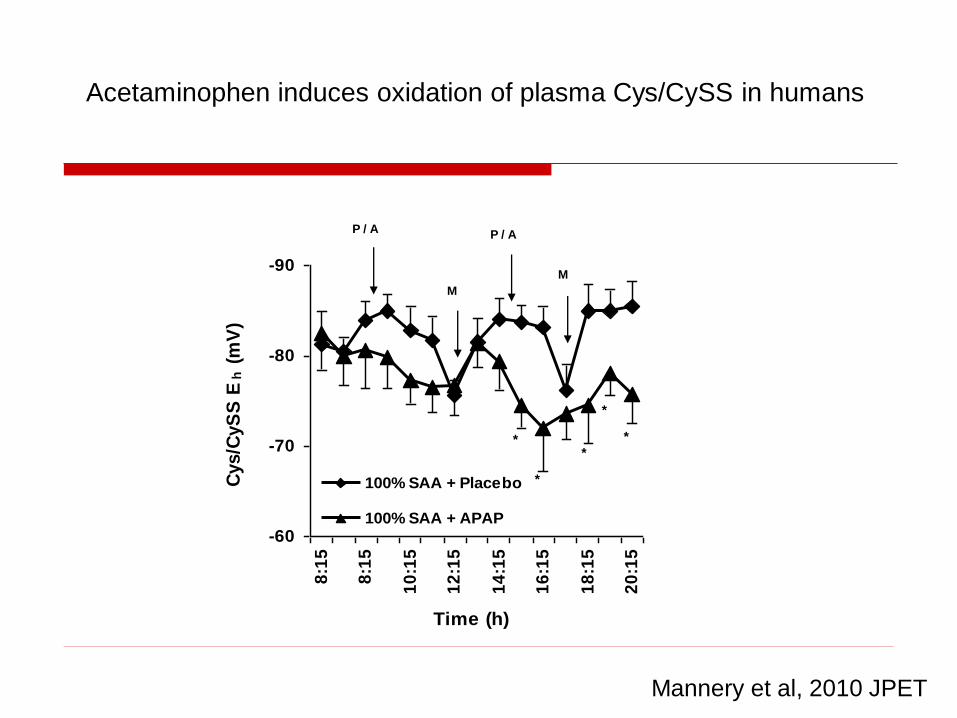
-90
-80
-70
-60
8:1
5
8:1
5
10
:15
12
:15
14
:15
16
:15
18
:15
20
:15
Time (h)
Cys/C
yS
S E
h (
mV
)
100% SAA + Placebo
100% SAA + APAP
P / A P / A
M
M
*
**
*
*
Mannery et al, 2010 JPET
Acetaminophen induces oxidation of plasma Cys/CySS in humans

2-Day sulfur amino acid insufficiency results in oxidation of plasma
GSH/GSSG in humans taking therapeutic dose of acetaminophen
Mannery et al, 2010 JPET
-150
-140
-130
-120
-110
-100
8:1
5
8:1
5
10
:15
12
:15
14
:15
16
:15
18
:15
20
:15
Time (h)
GS
H/G
SS
G E
h (
mV
)100% SAA + APAP
0% SAA + APAP
A
M
M A
**

Perturbation of the low flux mitochondrial redox circuits activates
cell death machinery and probably underlies mitochondrial
toxicities
Mitochondrial redox proteome is highly responsive to oxidative
conditions—reflects central role of redox mechanisms in regulation
of mitochondrial and cellular functions
Summary: Compartmentation of oxidative stress defense in mitochondria
Implications of drug toxicity
Low flux electron transfer pathways control thiol/disulfide systems
in mitochondria

Acknowledgements: NIEHS
Redox Proteomics
Young-Mi Go
Jan Pohl
Junmin Peng
Cell and molecular
Yan Chen
Smita Iyer
Siobhan Craige
Jason Hansen
Lou Ann Brown
Bill Liang
Hong Zhang
James Roede
Transgenic mice
Jiyang Cai
Redox modeling
Nnenna Adimora
Melissa Kemp


















