COM PAC TION-RE LATED STYLE OF …asgp.pl/sites/default/files/volumes/83_4_317_327.pdfAnnales...
11
Annales Societatis Geologorum Poloniae (2013), vol. 83: 317–327. COMPACTION-RELATED STYLE OF RUSOPHYCUS PRESERVATION FROM FURONGIAN (UPPER CAMBRIAN) OF HOLY CROSS MOUNTAINS (POLAND) Grzegorz SADLOK Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, PL- 00-818 Warszawa, Poland; address for correspondence: 77 Leadside Road, Aberdeen AB251RX, Aberdeenshire, Scotland, United Kingdom e-mail: [email protected] Sadlok, G., 2013. Compaction-related style of Rusophycus preservation from Furongian (Upper Cambrian) of Holy Cross Mountains (Poland). Annales Societatis Geologorum Poloniae, 83: 317–327. Abstract: In general, the trace fossil Rusophycus, preserved as a concave-upward structure on the top of a bed, is considered to be a fossilized marking, made by a trace maker. The structures described from the Cambrian (Furon- gian) of central Poland are genetically related to Rusophycus. However, despite their occurrence on the tops of beds, they are not fossilized traces, but compaction-related features, resulting from differential sandstone and mudstone compaction with possible mediation by organic-rich, heterolithic sediments. The preservation of these structures probably was influenced by biofilms or biomats. Key words: Rusophycus, trace fossil, preservation, compaction, Cambrian, Furongian, Poland. Manuscript received 14 November 2013, accepted 27 December 2013 INTRODUCTION Rusophycus is a resting trace fossil (cubichnion; Brom- ley, 1996; Seilacher, 2007). It is most commonly attributed to the life activity of arthropods. Palaeozoic examples occur mainly in marine strata and have been ascribed to trilobites (Crimes, 1970a, b; Seilacher, 1970). Mesozoic, non-marine forms of Rusophycus have been attributed to crustaceans (Bromley and Asgaard, 1972; Pollard, 1985; Schlirf et al., 2001). Small ichnospecies of Rusophycus, made by repre- sentatives of both of these arthropod groups, are morpho- logically indistinguishable (Bromley and Asgaard, 1979). Previously, concave, epichnial Rusophycus – a trace fossil on the upper surface of a sedimentary bed (Martins- son, 1970) – was interpreted to be actually a true trace, a de- pression in bottom sediment, excavated by a trace maker. There are two hypotheses on the mode of formation of the concave Rusophycus: (1) concave Rusophycus is a fossil- ized epifaunal trace, produced at the sediment-water inter- face (Crimes, 1975), and (2) Rusophycus is a fossilized infaunal trace, created at the sand-mud interface within the substrate (Seilacher, 1970, 1985). Both of these scenarios assume that the depressions made by trace makers were subsequently filled with sedi- ment, usually sand. The timing of the addition of the fill is the main difference between these scenarios. The epifaunal trace would be filled just after the biological activity or with a delay, after some period of non-deposition and possible erosion. In the case of infaunal traces, filling occurred im- mediately after the burrowing activity and this “instant cast- ing” mechanism is considered as leading to high-resolution preservation of the shallow traces (Seilacher, 1970, 1985). The sand-filled epifaunal or infaunal true traces are preserved as hypichnia that are natural casts of these traces, occurring on the lower surfaces of a sand bed (Martinsson, 1970). Hypichnia constitute the most commonly encountered mode of preservation of Rusophycus in the fossil record. According to this widely accepted interpretation, all Rusophycus-shaped structures occurring on bed tops in the Upper Cambrian Wiœniówka Sandstone Formation previou- sly were interpreted as the true traces of arthropods, namely of trilobites (Radwañski and Roniewicz, 1963; ¯yliñska and Radwañski, 2008). The results of this study indicate that at least some of these structures, occurring on tops of beds/la- minae, appear to be compaction-related phenomena, result- ing from differential compaction involving mud and sand (Baldwin, 1971; Nadon and Issler, 1997). MATERIAL AND METHODS The material studied comes from the Wiœniówka Sand- stone Formation (Fig. 1). The observations and conclusions presented in this paper are based on numerous field observa- tions and studies of trace fos sils from the collec tion ZPAL Tf. 4. The collection comprises material collected by the Author
Transcript of COM PAC TION-RE LATED STYLE OF …asgp.pl/sites/default/files/volumes/83_4_317_327.pdfAnnales...

Annales Societatis Geologorum Poloniae (2013), vol. 83: 317–327.
COM PAC TION-RE LATED STYLE OF RUSOPHYCUSPRES ER VA TION FROM FURONGIAN (UP PER CAM BRIAN)
OF HOLY CROSS MOUN TAINS (PO LAND)
Grzegorz SADLOK
Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, PL- 00-818 Warszawa, Poland;address for correspondence: 77 Leadside Road, Aberdeen AB251RX, Aberdeenshire, Scot land, United Kingdom
e-mail: [email protected]
Sadlok, G., 2013. Com pac tion-re lated style of Rusophycus pres er va tion from Furongian (Up per Cam brian) of HolyCross Moun tains (Po land). Annales Societatis Geologorum Poloniae, 83: 317–327.
Ab stract: In gen eral, the trace fos sil Rusophycus, pre served as a con cave-up ward struc ture on the top of a bed, iscon sid ered to be a fos sil ized mark ing, made by a trace maker. The struc tures de scribed from the Cam brian (Furon- gian) of cen tral Po land are ge net i cally re lated to Rusophycus. How ever, de spite their oc cur rence on the tops ofbeds, they are not fos sil ized traces, but com pac tion-re lated fea tures, re sult ing from dif fer en tial sand stone andmudstone com pac tion with pos si ble me di a tion by or ganic-rich, heterolithic sed i ments. The pres er va tion of thesestruc tures prob a bly was in flu enced by biofilms or biomats.
Key words: Rusophycus, trace fos sil, pres er va tion, com pac tion, Cam brian, Furongian, Po land.
Manu script re ceived 14 November 2013, ac cepted 27 December 2013
IN TRO DUC TION
Rusophycus is a rest ing trace fos sil (cubichnion; Brom-ley, 1996; Seilacher, 2007). It is most com monly at trib utedto the life ac tiv ity of ar thro pods. Palaeozoic ex am ples oc cur mainly in ma rine strata and have been as cribed to trilobites(Crimes, 1970a, b; Seilacher, 1970). Me so zoic, non-ma rineforms of Rusophycus have been at trib uted to crus ta ceans(Bromley and Asgaard, 1972; Pol lard, 1985; Schlirf et al.,2001). Small ichnospecies of Rusophycus, made by rep re -sen ta tives of both of these ar thro pod groups, are mor pho -log i cally in dis tin guish able (Bromley and Asgaard, 1979).
Pre vi ously, con cave, epichnial Rusophycus – a tracefossil on the up per sur face of a sed i men tary bed (Mar tins-son, 1970) – was in ter preted to be ac tu ally a true trace, a de -pres sion in bot tom sed i ment, ex ca vated by a trace maker.There are two hy poth e ses on the mode of for ma tion of thecon cave Rusophycus: (1) con cave Rusophycus is a fos sil -ized epifaunal trace, pro duced at the sed i ment-wa ter in ter -face (Crimes, 1975), and (2) Rusophycus is a fos sil izedinfaunal trace, cre ated at the sand-mud in ter face within thesub strate (Seilacher, 1970, 1985).
Both of these sce nar ios as sume that the de pres sionsmade by trace mak ers were sub se quently filled with sed i -ment, usu ally sand. The tim ing of the ad di tion of the fill isthe main dif fer ence be tween these sce nar ios. The epifaunaltrace would be filled just af ter the bi o log i cal ac tiv ity or with a de lay, af ter some pe riod of non-de po si tion and pos si ble
ero sion. In the case of infaunal traces, fill ing oc curred im -me di ately af ter the bur row ing ac tiv ity and this “in stant cast -ing” mech a nism is con sid ered as lead ing to high-res o lu tionpres er va tion of the shal low traces (Seilacher, 1970, 1985).The sand-filled epifaunal or infaunal true traces are pre served as hypichnia that are nat u ral casts of these traces, oc cur ringon the lower sur faces of a sand bed (Martinsson, 1970).Hypichnia con sti tute the most com monly en coun tered modeof pres er va tion of Rusophycus in the fos sil re cord.
Ac cord ing to this widely ac cepted in ter pre ta tion, allRusophycus-shaped struc tures oc cur ring on bed tops in theUp per Cam brian Wiœniówka Sand stone For ma tion previou- sly were in ter preted as the true traces of ar thro pods, namelyof trilobites (Radwañski and Roniewicz, 1963; ̄ yliñska and Radwañski, 2008). The re sults of this study in di cate that atleast some of these struc tures, oc cur ring on tops of beds/la-minae, ap pear to be com pac tion-re lated phe nom ena, re sult -ing from dif fer en tial com pac tion in volv ing mud and sand(Baldwin, 1971; Nadon and Issler, 1997).
MA TE RIAL AND METH ODS
The ma te rial stud ied co mes from the Wiœniówka Sand-stone For ma tion (Fig. 1). The ob ser va tions and con clu sionspre sented in this pa per are based on nu mer ous field observa-tions and stud ies of trace fos sils from the col lec tion ZPAL Tf. 4.The col lec tion com prises ma te rial col lected by the Au thor

(80%) and by Marcin Machalski (20%) from the In sti tute ofPaleobiology of Pol ish Acad emy of Sci ences, War saw.Thefol low ing spec i mens from the col lec tion ZPAL Tf. 4 bestdoc u ment the type of toponomy and pres er va tion de scribed:192, 513, 518, 600, 791, 1151, 1238, 1468, 1471, 1472,1473.
The heterolithic in ter vals of the Wiœniówka Sand stoneFor ma tion were split with a scis sor knife to search lamina-by-lamina for trace fos sils. Both the top and bot tom sur faces of each lamina were in ves ti gated. This mode of bed ex am i -na tion al lowed di rect ob ser va tion of the laminae sur round -ing the hypichnial Rusophycus, in clud ing their re la tion shipwith the hypichnial trace fos sil.
LO CA TION AND GEO LOG I CALSET TINGS
The ma te rial stud ied was col lected from the Cam brian(Furongian) Wiœniówka Sand stone For ma tion (Or³owski,1992a, 1992a; ¯yliñska et al., 2006). The siliciclastic depo-sits of this unit com prise quartz-dom i nated sand stones,mudstones and heterolithic in ter vals and are ex posed best at a few quar ries in wes tern most part of the Holy Cross Moun -tains (Wiœniówka Wielka, Wiœniówka Ma³a and Podwiœ-niówka), as well as in the Opatów area (Kowalczewski etal., 2006 and ref er ences therein). The es ti mates for thethick ness of the Wiœniówka Sand stone For ma tion rangefrom 80–200 m, up to 400–1400 m, de pend ing on the tec -tonic model ap plied (Kowalczewski et al. 2006 and ref er -ences therein). The pres ent study was con ducted in the Wiœ-
niówka Wielka Quarry, in the wes tern most part of the nor-thern Holy Cross Moun tains, Po land (Fig. 1).
The Wiœniówka Sand stone For ma tion is poor in in dexbody fos sils (¯yliñska et al., 2006). The unit was dated as“Mid dle to Up per Cam brian” by Or³owski (1992a, b), who(Or³owski, 1992b) based his con clu sions on “Cruziana stra -tig ra phy“, an ichnostratigraphical scheme ap plied to LowerPalaeozoic non-fossiliferous ma rine siliciclastics (Seila-cher, 1970, 1994, 2007; MacNaughton, 2007). ¯yliñska etal. (2006), study ing acritarchs and rare tri lo bite body fos -sils, con cluded that the unit is lower Furongian.
Sed i men tary en vi ron ment and diagenetic his tory
The Wiœniówka Sand stone For ma tion is com posed ofthick, amal gam ated quartz ite sand stone beds (quartz-areni-tes sensu Nagtegaal, 1978), mudstones and heterolithic in -ter vals char ac ter ized by flaser, wavy and len tic u lar bed ding(Studencki, 1994; Jaworowski and Sikorska, 2006; ¯y-liñska et al., 2006). Studencki (1994) in ter preted the sand -stone in ter vals as de pos ited on the prox i mal in ner shelf, andthe heterolithic sed i ments as de pos ited on the dis tal in nershelf. Jaworowski and Sikorska (2006) in ter preted this unitin a sim i lar way, as, storm-af fected de pos its on a shelf, withthe sands as tidal sand ridges or tidal sand waves and theheterolithic in ter vals as tran si tional to shelf muds. How ever, D¿u³yñski and ¯ak (1960) noted in their sedimentologicalstudy of the Wiœniówka Sand stone For ma tion that a shiftfrom mud- to sand-dom i nated sed i men ta tion could havetaken place in the same depth zone, as a re sult of dy namicchanges in the bot tom to pog ra phy and vari a tion in the cur -
318 G. SADLOK
Fig. 1. Lo ca tion of study area. A. Po si tion of Po land in Eu rope. B. Map of Po land, show ing lo ca tion of Holy Cross Moun tains. C. HolyCross Moun tains (Palaeozoic core). D. Plan of Wiœniówka Wielka Quarry (stra tig ra phy based on ¯yliñska et al., 2006)

rent paths, typ i cal for shal low sub aque ous en vi ron ments.Nu mer ous and var i ous rip ple types, pre served very well inthe heterolithic in ter vals, also were taken as in di cat ing shal -low- to very shal low-wa ter con di tions (D¿u³yñski and ¯ak,1960; Radwañski and Roniewicz, 1960).
The Wiœniówka Sand stone For ma tion hosts nu mer ousstruc tures that may have orig i nated as a re sult of mi cro bialbind ing of the sed i ment, the so-called microbially in ducedsed i men tary struc tures (MISS; Noffke et al. 2001). Theseoc cur on bed ding planes and ap pear to be ge net i cally di -verse (see the clas si fi ca tion in Eriksson et al., 2007). Thesestruc tures were ob served in the same in ter vals that yieldedthe trace fos sils stud ied. The most char ac ter is tic of these arewrin kle marks (Fig. 2A–C), cracks de vel oped within thesand stone lithologies (Fig. 2D), in ter fer ence rip ples (Fig.2E) and “sand chips” (Fig. 2F), that is, oval and flat clasts of sand stone, oc cur ring within sand stone or mudstone litho-logies. This as so ci a tion of struc tures is a strong in di ca tionof the pres ence of biofilms and/or mi cro bial mats within thesed i men tary en vi ron ment (cf. Hagadorn and Bottjer, 1997;Pflüger, 1999; Bottjer and Hagadorn, 2007; Eriksson et al.,2007; Porada and Bouougri, 2007; Porada et al., 2008). Allof these MISS re sulted from the pres ence of a microbiallypro duced extracellular poly meric sub stance that makes thesandy sub strate atyp i cally co he sive and re sis tant to ero sion(Bottjer and Hagadorn, 2007).
Re cent mi cro bial mats typ i cally oc cur within tidal set -tings, where spe cific pre req ui sites as to sub strate tex ture,high mois ture, low hy dro dy namic en ergy and low metazoan ac tiv ity are met (Porada and Bouougri, 2007). MISS in thefos sil re cord are also most com monly en coun tered in shal -low ma rine con di tions (intertidal to supratidal; e.g.Carmona et al., 2012). How ever, mi cro bial mats are by nomeans re stricted to such set tings (Schieber et al., 2007) andin the Pre cam brian and Lower Palaeozoic mi cro bial matswere much more wide spread. Some work ers even pos tu latea Pre cam brian–Cam brian shift from “matground” (mi cro -bial mats) to a “mixground” (mi cro bial cov ers around in di -vid ual grains) type of sub strate struc ture and link this sig nif -i cant event to the in creas ing ac tiv ity of bioturbators (Seila-cher and Pflüger, 1994). Pflüger (1999) showed a wide en -vi ron men tal range for the MISS that he de scribed from theSi lu rian Tanezzuft Shale and the Acacus Sand stone in Li-bya. In his ex am ples, the MISS oc curred in strata, char ac ter -ized by ichnofossils typ i cal of the Skolithos and Cruzianaichnofacies (Pflüger, 1999, fig, 7). There fore, the oc cur -rence of MISS in strata, char ac ter ized by the Cruziana ich-nofacies (in clud ing stud ied Rusophycus) as in the case ofthe Wiœniówka Sand stone For ma tion, is not un usual for theLower Palaeozoic.
The sig nif i cance of mi cro bial ac tiv ity for the pres er va -tion of trace fos sils is well known (e.g. Carmona et al.,2012). How ever, in the pres ent pa per, a po ten tially new as -pect of the taphonomical sig nif i cance of mi cro bial mats isbriefly dis cussed.
The postdepositional his tory of the Wiœniówka Sand -stone For ma tion is not known ex actly, es pe cially for post-Si -lu rian time (Sikorska, 2000). How ever, it has been pos tu lated that the Wiœniówka Sand stone For ma tion ex pe ri enced deepburial within the Cam brian to Si lu rian time in ter val, reach ing
a burial depth of about 1,700 m in the Si lu rian (Sikorska,2000). Late diagenetic silification oc curred at that time un dercon di tions of high tem per a ture and pres sure, re sult ing in theoblit er a tion of the orig i nal grain boundaries by the ex ten sivefor ma tion of quartz overgrowths (see Sikorska, 2000).
RE SULTS
Ob ser va tions
The in di vid ual sand stone and mudstone laminae can beas thin as 1 mm in these in ter vals and are usu ally 1–2 mmthick. Rusophycus is an abun dant trace fos sil in the het ero-lithic in ter vals stud ied. Hypichnial Rusophycus is em bed -ded in the sur round ing mudstone laminae; mudstone lami-nae run be low and above the Rusophycus, which in cross-sec tion take the form of sand stone lenses (Fig. 3A–E). Thetops of these lenses are more or less flat (Fig. 3E). Lo cally,ar thro pod trackways oc cur on the up per sur faces of the len-ses. The sand stone bod ies in the form of lens-like hypich-nial Rusophycus de form the muddy laminae be low andabove. The laminae above are slightly con vex-up ward (Fig.3E) and those be low mimic the mor phol ogy of the lowersur face of the hypichnial Rusophycus (Fig. 3F): the laminaeare con cave-up ward be low the endopodal lobes and con -vex-up ward be tween the lobes (Fig. 3C, D).
Typ i cally, no in ter rup tion of the lower muddy laminaewas ob served. There fore the cross-sec tions of these laminae mostly are not ob served on the lat eral mar gins of the bed-top Rusophycus-shaped struc ture, ex cept for rare, thin cutlaminae, as so ci ated with hypichnial Rusophycus (Fig. 5D).The laminae within the Rusophycus-shaped fill ing are typ i -cally con tin u ous (Fig. 4A, C–F). Some dis con ti nu ities or“win dows” in the lower muddy laminae were ob served, butthese re sult mainly from weath er ing and/or dam age dur ingsam pling (Fig. 4B). Fi nally, bed-top Rusophycus dis playsfewer mor pho log i cal de tails (e.g. sculp tur ing of the lobes),than does the coun ter part, hypichnial Rusophycus (Figs 4E,F, 5A, B). In the sam ple stud ied, the re verse sit u a tion wasnot ob served, i.e. more de tails were seen on the bed-topRusophycus-shaped struc ture.
In ter pre ta tion
Pres er va tion of hypichnial RusophycusThe sandy lens-like oc cur rences of Rusophycus are typ -
i cal hypichnia (see Martinsson, 1965, 1970), with a lat er ally re stricted ex tent of the cast ing me dium (a lens-like ap pear -ance). This mode of pres er va tion may be ex plained as anex am ple of con cealed bed-junc tion pres er va tion, a spe cialtype of trace-fos sil pres er va tion, in which the fill ing mate-rial dif fers in tex ture sig nif i cantly from the host sed i ment and has no con nec tion with sim i lar lay ers above, e.g. a sandy fill -ing of bur rows within a mudstone (Simpson, 1957; Jensen,1997; Jensen et al., 2005). Hypichnial Rusophycus in theform of sand stone lenses are the re sults of pref er en tial pres er -va tion of trace-in fill ing sandy ma te rial, de pos ited in a shel -tered de pres sion (the ac tual trace made by the or gan ism).
RUSOPHYCUS PRES ER VA TION FROM FURONGIAN 319

320 G. SADLOK
Fig. 2. Microbially in duced sed i men tary struc tures from Wiœniówka Sand stone For ma tion. A. Top sur face of sand stone bed with wrin -kle marks (marked with ar row and let ter K), note that struc ture oc curs at sand stone-sand stone con tact (ar rowed as s-s) with no clay or siltin ter ca la tion be tween sand stone strata (field pho to graph; com pare Bottjer and Hagadorn, 2007). B. Wrin kle marks with clearly vis i ble re -lief (bed top). C. Thin sec tion of wrin kle marks with vis i ble flat crest and trough (ar rowed) typ i cal fea tures, known from sim i lar struc tures(com pare Calner and Eriksson, 2011; pic ture taken in plane-po lar ized light close to bed top). D. Si nu soi dal crack within sand stone with no clay or silt in ter ca la tion at part ing sur face. E. In ter fer ence rip ples (field pho to graph), pres er va tion of older set might be re lated to mi cro -bial sta bi li za tion (bed top). F. “Sand chips”, rounded and flat sand stone intraclasts, pre served on top of an other sand stone bed (com pareBottjer and Hagadorn, 2007). These dis play lo cally char ac ter is tic prop er ties, as they seem to be glued to gether (cen tral part of the pic ture;field pho to graph)
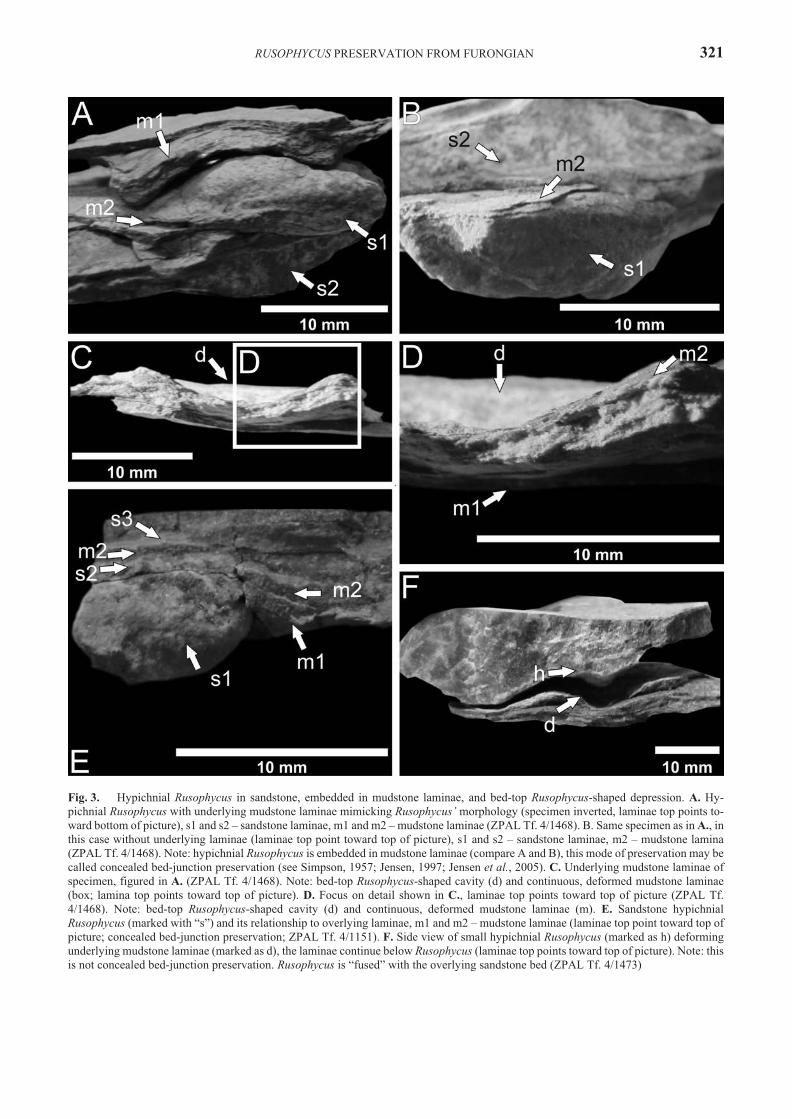
RUSOPHYCUS PRES ER VA TION FROM FURONGIAN 321
Fig. 3. Hypichnial Rusophycus in sand stone, em bed ded in mudstone laminae, and bed-top Rusophycus-shaped de pres sion. A. Hy-pichnial Rusophycus with un der ly ing mudstone laminae mim ick ing Rusophycus’ mor phol ogy (spec i men in verted, laminae top points to -ward bot tom of pic ture), s1 and s2 – sand stone laminae, m1 and m2 – mudstone laminae (ZPAL Tf. 4/1468). B. Same spec i men as in A., in this case with out un der ly ing laminae (laminae top point to ward top of pic ture), s1 and s2 – sand stone laminae, m2 – mudstone lamina(ZPAL Tf. 4/1468). Note: hypichnial Rusophycus is em bed ded in mudstone laminae (com pare A and B), this mode of pres er va tion may be called con cealed bed-junc tion pres er va tion (see Simpson, 1957; Jensen, 1997; Jensen et al., 2005). C. Un der ly ing mudstone laminae ofspec i men, fig ured in A. (ZPAL Tf. 4/1468). Note: bed-top Rusophycus-shaped cav ity (d) and con tin u ous, de formed mudstone laminae(box; lamina top points to ward top of pic ture). D. Fo cus on de tail shown in C., laminae top points to ward top of pic ture (ZPAL Tf.4/1468). Note: bed-top Rusophycus-shaped cav ity (d) and con tin u ous, de formed mudstone laminae (m). E. Sand stone hypichnialRusophycus (marked with “s”) and its re la tion ship to over ly ing laminae, m1 and m2 – mudstone laminae (laminae top point to ward top ofpic ture; con cealed bed-junc tion pres er va tion; ZPAL Tf. 4/1151). F. Side view of small hypichnial Rusophycus (marked as h) de form ingun der ly ing mudstone laminae (marked as d), the laminae con tinue be low Rusophycus (laminae top points to ward top of pic ture). Note: this is not con cealed bed-junc tion pres er va tion. Rusophycus is “fused” with the over ly ing sand stone bed (ZPAL Tf. 4/1473)

322 G. SADLOK
Fig. 4. Bed-top Rusophycus-shaped struc tures and laminae con ti nu ity in side struc ture. A. Bed-top Rusophycus-shaped struc tures incon tin u ous lamina (edges of lamina shown by ar rows; ZPAL Tf. 4/192). B. Hypichnial sand stone Rusophycus (marked with “s”) de form -ing un der ly ing mudstone lamina (marked with m; ZPAL Tf. 4/1470). C. Bed-top Rusophycus-shaped struc ture de form ing con tin u ouslamina (the edge of lamina is marked with box; ZPAL Tf. 4/600). D. Close-up of de tail shown in C, edge (marked with l) of con tin u ouslamina de formed by bed-top Rusophycus-shaped struc ture (ZPAL Tf. 4/600). E. Hypichnial Rusophycus (on right) and bed-top Ru-sophycus-shaped struc ture (on left), m – mudstone lamina, s – sand stone lamina (ZPAL Tf. 4/1471). Note: to tal lack of sculp ture onepichnial struc ture. F. Close-up of de tail shown in E, bed-top Rusophycus-shaped struc ture with no sculp ture and with cross-sec tion ofmudstone laminae vis i ble out side struc ture (ar rowed; ZPAL Tf. 4/1471)

For ma tion and pres er va tion of bed-topRusophycus-shaped struc tures
The bur row ing ac tiv ity by the Rusophycus trace makershould have re moved the de posit from be low and dis placedit (Fig. 5C), and this ac tion should have pro duced a cav ity in the un der ly ing bed (com pare Seilacher, 1970, 1985). There -fore, the laminae in side the cav ity should be dis con tin u ous
and the cross-sec tional views of these laminae should be ex -posed at the cav ity side mar gins. On the con trary, the sand -stone laminae un der neath the sand stone Rusophycus typ i -cally have not been in ter rupted by the trace maker and in -stead are con tin u ous. This type of laminae mod i fi ca tion,with out any cut ting of laminae, in di cates that the Rusophy-cus-shaped bed-top cav i ties are the re sult of de for ma tion
RUSOPHYCUS PRES ER VA TION FROM FURONGIAN 323
Fig. 5. Bed-top Rusophycus-shaped struc ture, epichnial Rusophycus and mud laminae com pac tion. A. Hypichnial Rusophycus withwell de fined sculp ture on its lobes (ZPAL Tf. 4/1238). B. Bed-top Rusophycus-shaped struc ture with con tin u ous laminae in side cav ity,l1-l2 – laminae (ZPAL Tf. 4/1238). C. Epichnial Rusophycus (true trace or undertrace) with clearly vis i ble dis place ment of ma te rial(ZPAL Tf. 4/513). D. Hypichnial sand stone Rusophycus (marked with “s”) and sin gle muddy lamina it cuts through (marked with “m”;(ZPAL Tf. 4/1468). E, F. Top of rip pled sand stone bed bear ing ar thro pod track or undertrack with 1 mm thick set of mudstone laminae onits top (E), and same bed with out over ly ing set of muddy laminae (F) (ZPAL Tf. 4/1472). Note: in E there are ar thro pod tracks (ar rowed)and in F there are cor re spond ing undertracks (ar rowed)

that af fected the laminae. The mode of de for ma tion in di -cates that the laminae were not de formed by the trace pro -ducer it self, but rather by pres sure re lated to the pro cess ofcom pac tion (Fig. 6), caused by an in crease in the load dur -ing later burial.
The hypichnial Rusophycus stud ied dis plays more mor -pho log i cal de tails, such as sculp ture of the lobes, than thecoun ter parts on the bed tops do (Fig. 5A, B). This phe nom e -non is dif fi cult to ex plain in a typ i cal epifaunal or infaunalsce nario, in which con cave struc tures are the ac tual epich-nial traces. In such a case, the bed-top struc tures (epichnia)should dis play more de tails of mor phol ogy. The model of acom pac tion-re lated or i gin for the bed-top Rusophycus-shaped struc tures, de scribed in this pa per, would ex plain the dif fer ences in pres er va tion res o lu tion. The loss of mor pho -log i cal de tails could be ex plained by the “copy ef fect”,where each copy loses the def i ni tion of some qual ity. In thecom pac tion-re lated model, the con cave bed-top struc turesare de tail-de pleted cop ies of the hypichnial Rusophycus(sandstone lens-like Rusophycus), whereas the hypichniathem selves are cop ies of epifaunal/infaunal traces.
These ob ser va tions lead to the fi nal con clu sion that thebed-top Rusophycus-shaped struc tures de scribed are not
epifaunal or infaunal traces. The two sub strates were orig i -nally sep a rated. Dur ing diagenesis, the hypichnial Rusophy- cus fill in the up per layer acted as an in denter that shapedthe sur face of the sandy laminae be neath. The ex act matchbe tween the hypichnial Rusophycus and its im pres sion onthe bed top be low in di cates that ver ti cally act ing pres surewas in volved, rather than lat eral bed slides and stress, where more elon gated grooves would be ex pected. This is how thehypichnial Rusophycus made the bed-top “im print mark” on the top of the bed be low. In this sce nario, the orig i nally bur -rowed sub strate, in which a trace orig i nally was pro duced,is not pre served in the fos sil state and there fore it is pos tu -lated that its orig i nal com po si tion was very sen si tive todiagenetic al ter ation. There fore, the or ganic con tent of thissub strate was likely to be high. The pres ence of microbiallyin duced sed i men tary struc tures in di cates that mi cro bialbiofilms and mats played a role in the sed i men tary en vi ron -ment of the strata. Thus it is pos si ble that the or ganic-en -riched ma te rial, oc cur ring be tween the laminae and tar geted by the trace pro ducer, was a mi cro bial mat or biofilm.
If the or i gin of the bed-top Rusophycus-shaped struc -tures sig nif i cantly post dates (pos si bly by mil lions of years?) the for ma tion of the hypichnial Rusophycus, then the pro -
324 G. SADLOK
Fig. 6. Sche matic draw ing to ex plain or i gins of bed-top com pac tion-me di ated Rusophycus-shaped struc tures. A. The tri lo bite digs in amuddy sub strate. B. The trace is filled with sand. C. The trace and fill ing sandy ma te rial are bur ied. Note that the sand-filled Rusophycusap pears to be em bed ded in muddy sed i ment; this is con cealed bed-junc tion pres er va tion (see text for ref er ences). D. Be gin ning of com pac -tion. The de for ma tion of the un der ly ing laminae oc curs af ter the ac tiv ity of the trace-maker and this dif fer en ti ates these struc tures fromcleav age-re lief pres er va tion (“undertrack”; see Frey and Pem ber ton, 1985). Let ters “m” and “s” re fer to muddy and sandy lithologies, re -spec tively, whereas the num bers in di cate rel a tive ages of laminae (start ing from 1 as the old est). Ow ing to com pac tion, the mor phol ogy ofhypichnial Rusophycus is im printed on the top of the lower lamina and the up per laminae also are af fected. The de gree of de for ma tion de -creases away from the hypichnial Rusophycus

posed mode of the for ma tion of these struc tures is dif fer entfrom that of cleav age-re lief or undertrack pres er va tion (Fig. 5E, F), in which subsurface laminae are de formed dur ingtrace-maker ac tiv ity at the sur face (cf. Lessertisseur, 1955;Heyler and Lessertisseur, 1963; Goldring and Seilacher,1971; Frey and Pem ber ton, 1985). The trace fos sil is a fos -sil ized trace, which is a re sult of sub strate-or gan ism in ter ac -tion (Bertling et al., 2006). How ever, the tran si tion from the orig i nal trace into the fos sil re cord (trace fos sil) is com -monly as so ci ated with abiotic pro cesses, such as the for ma -tion of hypichnial casts, in which no bi o log i cal ac tiv ity oc -curs. The bed-top struc tures de scribed are not fos sil izedepichnial traces, but a new type of pres er va tion of them, inwhich a diagenetic pro cess (com pac tion) played a ma jorrole. The new term “compactional epichnia” is pro posedhere to cover this type of trace fos sil toponomy and pres er -va tion. The fol low ing model is pro posed for the or i gin ofthe struc tures de scribed: 1) an or gan ism bur rows the sub -strate, which prob a bly was cov ered by a mi cro bial mat ormi cro bial film; 2) the trace is filled with sand; 3) the bur -rowed sub strate and the sandy fill ing are bur ied; 4) com pac -tion starts and cre ates a compactional epichnion (Fig. 6).
DIS CUS SION
Be cause sands and muds (a mix ture of silt and clay) dif -fer in orig i nal po ros ity, the de pos its rep re sented by theselithologies will tend to com pact dif fer en tially. Sand has alower po ros ity than freshly de pos ited mud and hence, sandtends to com pact less than mud (Baldwin, 1971; Perrier andQuiblier, 1974; Nadon and Issler, 1997). More over, in mud- stones, the grow ing pres sure in creases sil ica dis so lu tion, but at the same time the pre cip i ta tion of sil ica in the same bedsis in hib ited (Mullis, 1992). This means that the vol ume ofmud de creases, ow ing to a de ple tion in sil ica. On the otherhand, in sands sil ica is pre cip i tated freely as inter-gran u larce ment (Mullis, 1992). Thus, the pre cip i tated sil ica, trans -ported by pore flu ids from the neigh bour ing muds, can in -crease the re sis tance of the sand stone to com pac tion (Ste -phenson et al., 1992). Sim i lar pro cesses prob a bly oc curredin the strata stud ied, as sand-dom i nated beds dis play per va -sive sili ci fi ca tion, in ad di tion to pres sure so lu tion (see Si-korska, 2000). This is why com pac tion-re lated de for ma -tions are so well pre served in the mudstone laminae, sur -round ing Rusophycus (com pare Nadon and Issler, 1997, fig. 4 therein). As in di cated, the dif fer ence in pres er va tion ofde tail, ob served in the bed-top and hypichnion struc turecou plets, can be ex plained by the “copy ef fect”, with twoorig i nally sep a rate sub strates com ing into di rect con tactdur ing diagenesis.
The bur rowed sub strate ap pears to be vir tu ally miss ingin the cases stud ied as cut laminae are mostly not ob served(spec i mens show no mac ro scop i cally vis i ble in ter ca la tionsbe tween the sandy laminae (Fig. 5A, B)). Dur ing dif fer en -tial com pac tion, beds may ex hibit sig nif i cant re duc tion ofthick ness, but typ i cally their con ti nu ity is not dis rupted andat least a thin rem nant of the orig i nal lamina is pre served(Wetzel and Reisdorf, 2007). There fore, it is pos tu lated here that the bur rowed sub strate had a high con tent of or ganic
mat ter and a sig nif i cant re duc tion of its thick ness wascaused by de com po si tion pro cesses that could have takenplace be fore dif fer en tial com pac tion be gan and dur ing thecom pac tion the thick ness of the laminae was fur ther re -duced (see Wetzel and Reisdorf, 2007). The sub strate couldrep re sent a liv ing or de cay ing biofilm or mi cro bial mat. Thepres ence of mi cro bial biofilms or mats in the sed i men taryen vi ron ment of the Wiœniówka Sand stone For ma tion isindicated by microbially in duced sed i men tary struc tures, asdocumented in this pa per. The mi cro bial com po nent of theseinterbeds could have been the real tar get of the dig ging ar -thro pods, namely trilobites feed ing on the biomat.
CON CLU SIONS
The Rusophycus-shaped struc tures, oc cur ring on thetops of beds in the Cam brian (Furongian) Wiœniówka Sand -stone For ma tion of the Holy Cross Moun tains (Po land) re -sem ble true epifaunal or infaunal trace fos sils. How ever, ade tailed study of the muddy laminae as so ci ated with thesand stones shows that the laminae un der ly ing Rusophycus(hypichnion) were not in ter rupted by the ac tiv i ties of thetrace maker, but are con tin u ous. This mode of mod i fi ca tionof laminae in di cates a com pac tion-re lated or i gin with pos si -ble me di a tion by mi cro bial biofilms or biomats, occurringbetween them.
Ac knowl edg ments
This study is a part of a Ph.D. pro ject su per vised by MarcinMachalski at the In sti tute of Paleobiology of Pol ish Acad emy ofSci ences. It was fi nanced by a grant from the Min is try of Sci enceand Higher Ed u ca tion of Po land in years 2010 to 2012 (no. NN307 243539) to the Au thor and, par tially, by the In sti tute of Pa-leobiology PAS. I thank the of fi cials and mem bers of staff work -ing on the Wiœniówka Wielka quarry for their help, as sis tance anden thu si asm. I am grate ful to work ers and stu dents in the Fac ulty ofEarth Sci ences of the Uni ver sity of Silesia for lo gis tic help andgrant ing ac cess to petrographic equip ment. Fi nally, I would like to thank An drew K. Rindsberg (Uni ver sity of Al a bama, Living stone, USA) for lin guis tic cor rec tions and crit i cal re marks. I am grate fulto two ASGP re view ers, Andreas Wetzel (Uni ver sity of Basel,Swit zer land) and one anon y mous re viewer, for their con struc tivecrit i cism of the manu script. I thank also Al fred Uchman (Jagiel-lonian Uni ver sity, Kraków) for his con struc tive crit i cism andFrank Simpson (Wind sor, Can ada) for help, re marks and lin guis tic cor rects.
REF ER ENCES
Baldwin, B., 1971. Ways of de ci pher ing com pacted sed i ments.Jour nal of Sed i men tary Re search, 41: 293–301.
Bertling, M., Braddy, S. J., Bromley, R. G., Demathieu, G. R.,Genise, J., Mikuláš, R., Niel sen, J. K., Niel sen, K. S. S.,Rindsberg, A. K., Schlirf, M. & Uchman, A., 2006. Names for trace fos sils: a uni form ap proach. Lethaia, 39: 265–286.
Bottjer, D. & Hagadorn, J. W., 2007. Mat growth fea tures, In:Schieber, J., Bose, P. K., Eriksson, P. G., Banerjee, S., Sarkar, S., Altermann, W. & Catuneau, O. (eds), At las of Mi cro bialMat Fea tures Pre served within the Clastic Rock Re cord.
RUSOPHYCUS PRES ER VA TION FROM FURONGIAN 325

Elsevier, Am ster dam, pp. 53–71.Bromley, R. G., 1996. Trace Fos sils: Bi ol ogy, Taphonomy and
Ap pli ca tions. Chap man & Hall, Routledge, 386 pp.Bromley, R. G. & Asgaard, U., 1972. Notes on Green land trace
fos sils. I. Fresh wa ter Cruziana from the Up per Tri as sic ofJameson Land, East Green land. GrÝnlands Geologiske Un -dersÝgelse, 49: 7 13.
Bromley, R. G. & Asgaard, U., 1979. Tri as sic fresh wa ter ichno-coenoses from Carlsberg Fjord, East Green land. Palaeo ge -ogra phy, Palaeoclimatology, Palaeo ec ol ogy, 28: 39–80.
Calner, M. & Eriksson, M. E., 2011. The re cord of microbially in -duced sed i men tary struc tures (MISS) in the Swed ish Pa leo -zoic. SEPM So ci ety for Sed i men tary Ge ol ogy Spe cial Publi-cat ion, 101: 29–35.
Carmona, N. B., Ponce, J. J., Wetzel, A., Bournod, C. N. &Cuadrado, D. G., 2012. Microbially in duced sed i men tarystructures in Neo gene tidal flats from Ar gen tina: Paleoen vi -ron men tal, strati graphic and taphonomic im pli ca tions. Pala-eogeography, Palaeoclimatology, Palaeo ec ol ogy, 353–355:1–9.
Crimes, T. P., 1975. The pro duc tion and pres er va tion of tri lo biterest ing and fur row ing traces Lethaia, 8: 35–48.
Crimes, T. P., 1970a. The sig nif i cance of trace fos sils in sedimen-tology, stra tig ra phy and palaeo ec ol ogy with ex am ples fromLower Palaeozoic strata. In: Crimes, T. P. & Harper, J. C.(eds), Trace Fos sils. Geo log i cal Jour nal Spe cial Is sue, 3:101–126.
Crimes, T. P., 1970b. Tri lo bite tracks and other trace fos sils fromthe Up per Cam brian of North Wales. Geo log i cal Jour nal, 7:47–68.
D¿u³yñski, S. & ¯ak, C., 1960. Sed i men tary en vi ron ment of theCam brian quartzites in the Holy Cross Moun tains, Cen tralPo land, and their re la tion ship to the flysch fa cies. RocznikPolskiego Towarzystwa Geologicznego, 30: 213–241.
Eriksson, P. G., Schieber, J., Bouougri, E., Gerdes, G., Porada, H.,Banerjee, S., Bose, P. K. & Sarkar, S., 2007. Clas si fi ca tion ofstruc tures left by mi cro bial mats in their host sed i ments. In:Schieber, J., Bose, P. K., Eriksson, P. G., Banerjee, S., Sarkar, S., Altermann, W. & Catuneau, O. (eds), At las of Mi cro bialMats Fea tures Pre served within the Clastic Rocks Re cord.Elsevier, Am ster dam, pp. 39–52.
Frey, R. W. & Pem ber ton, S. G., 1985. Biogenic struc tures in out -crops and cores. I Ap proaches to ichnology. Bul le tin of Ca na -dian Pe tro leum Ge ol ogy, 33: 72–115.
Goldring, R. & Seilacher, A., 1971. Limulid undertracks and theirsedimentological im pli ca tions. Neues Jahrbuch für Geologieund Paläontologie, Abhandlungen, 31: 422–442.
Hagadorn, J. W. & Bottjer, D. J., 1997. Wrin kle struc tures: Mi cro-bially me di ated sed i men tary struc tures com mon in subtidalsiliciclastic set tings at the Pro tero zoic–Phanerozoic tran si -tion. Ge ol ogy, 25: 1047–1050.
Heyler, D. & Lessertisseur, J., 1963. Pistes de tétrapodes permiens dans la re gion de LodÀve (Hérault). Mémoires du Muséumd’Histoire Naturelle, Nou velle Série, Série C, Sci ences de laTerre, 11: 125–221.
Jaworowski, K. & Sikorska, M., 2006. £ysogóry Unit (Cen tral Po -land) ver sus East Eu ro pean Craton – ap pli ca tion of sedimen-tological data from Cam brian siliciclastic as so ci a tion. Geo -log i cal Quar terly, 50: 77–88.
Jensen, S., 1997. Trace fos sils from the Lower Cam brian Mick-witzia sand stone, south-cen tral Swe den. Fos sils and Strata,42: 1–111.
Jensen, S., Droser, M. L. & Gehling, J. G., 2005. Trace fos sil pres -er va tion and the early evo lu tion of an i mals. Palaeo ge ogra -phy, Palaeoclimatology, Palaeo ec ol ogy, 220: 19–29.
Kowalczewski, Z., ¯yliñska, A. & Szczepanik, Z., 2006. Kambr w Górach Œwiêtokrzyskich. In: Skompski, S. & ¯yliñska, A.(eds), Procesy i zdarzenia w historii geologicznej Gór Œwiêto- krzyskich, LXXVII Zjazd Naukowy Polskiego TowarzystwaGeologicznego, Ameliówka k. Kielc 28-30 czerwca 2006 r.,Materia³y Konferencyjne. Polskie Towarzystwo Geologi-czne, Pañstwowy Instytut Geologiczny, Wydzia³ GeologiiUniwersytetu Warszawskiego, Warszawa, pp. 14–27.
Lessertisseur, J., 1955. Traces fossiles d’activité animale et leursig ni fi ca tion paléobiologique. Mémoires de la Société Géolo-gique de la France, Nou velle Série, 34:. 1–150.
MacNaughton, R. B., 2007. The ap pli ca tion of trace fos sils tobiostratigraphy. In: Miller, W., (ed.), Trace Fos sils: Con -cepts, Prob lems, Pros pects. Elsevier, Am ster dam, pp. 135–148.
Martinsson, A., 1965, As pects of a Mid dle Cam brian thanatotopeon Öland. GFF, 87: 181–230.
Martinsson, A., 1970. Toponomy of trace fos sils. In: Crimes, T. P.& Harper, J. C. (eds), Trace Fos sils. Geo log i cal Jour nal Spe -cial Is sue, 3: 323–330.
Mullis, A. M., 1992. A nu mer i cal model for po ros ity mod i fi ca tionat a sand stone-mudstone bound ary by quartz pres sure dis so -lu tion and dif fu sive mass trans fer. Sedimentology, 39: 99–107.
Nadon, G. C. & Issler, D. R., 1997. The com pac tion of floodplainsed i ments: tim ing, mag ni tude and im pli ca tions. GeoscienceCan ada, 24: 37–43.
Nagtegaal, P. J. C. 1978. Sand stone-frame work in sta bil ity as afunc tion of burial diagenesis. Jour nal of the Geo log i cal So ci -ety, 135: 101–105.
Noffke, N., Gerdes, G., Klenke, T. & Krumbein, W. E., 2001.Microbially in duced sed i men tary struc tures: A new cat e gorywithin the clas si fi ca tion of pri mary sed i men tary struc tures.Jour nal of Sed i men tary Re search, 71: 649–656.
Or³owski, S., 1992a. Cam brian stra tig ra phy and stage sub di vi sionin the Holy Cross Moun tains, Po land. Geo log i cal Mag a zine,129: 471–474.
Or³owski, S., 1992b. Tri lo bite trace fos sils and their strati graphi cal sig nif i cance in the Cam brian se quence of the Holy CrossMoun tains, Po land. Geo log i cal Jour nal, 27: 15–34.
Perrier, R. & Quiblier, J., 1974. Thick ness changes in sed i men tarylay ers dur ing com pac tion his tory; meth ods for quan ti ta tiveeval u a tion. Amer i can As so ci a tion of Pe tro leum Ge ol o gistsBul le tin, 58: 507–520.
Pflüger, F., 1999. Matground struc tures and re dox fa cies. Ge ol ogy, 14: 25–39.
Pol lard, J. E., 1985. Isopodichnus, re lated ar thro pod trace fos silsand notostracans from Tri as sic flu vial sed i ments. Trans ac -tions of the Royal So ci ety of Ed in burgh, Earth Sci ences, 76:272–285.
Porada, H. & Bouougri, E. H., 2007. Wrin kle struc ture – crit i calre view. Earth-Sci ence Re views, 81: 199–215.
Porada, H., Ghergut, J. & Bouougri, E. H., 2008. Kinneyia-typewrin kle struc tures – crit i cal re view and model of for ma tion.Palaios, 23: 65–77.
Radwañski, A. & Roniewicz, P., 1960. Rip ple marks and othersed i men tary struc tures of the Up per Cam brian at Wielka Wiœ- niówka (Holy Cross Mts.). Acta Geologica Polonica, 10:371–397.
Radwañski, A. & Roniewicz, P., 1963. Up per Cam brian tri lo biteichnocoenosis from Wielka Wiœniówka (Holy Cross Moun -tains, Po land). Acta Palaeontologica Polonica, 8: 259–280.
Schieber, J., Bose, P. K., Eriksson, P. G. & Sarkar, S., 2007.Paleogeography of mi cro bial mats in terrigenous clastics –en vi ron men tal dis tri bu tion of as so ci ated sed i men tary fea tures
326 G. SADLOK

and the role of geo logic time. In: Schieber, J., Bose, P. K.,Eriksson, P. G., Banerjee, S., Sarkar, S., Altermann, W. &Catuneau, O. (eds), At las of Mi cro bial Mats Fea tures Pre -served within the Clastic Rocks Re cord. Elsevier, Am ster -dam, pp. 267–275.
Schlirf, M., Uchman, A. & Kümmel, M., 2001. Up per Tri as sic(Keuper) non-ma rine trace fos sils from the Haßberge area(Franconia, south-east ern Ger many). Paläontologische Zeit-schrift, 75: 71–96.
Seilacher, A., 1970. Cruziana stra tig ra phy of “non-fossiliferous”Palaeozoic sand stones. In: Crimes, T. P. & Harper, J. C. (eds), Trace Fos sils. Geo log i cal Jour nal Spe cial Is sue, 3: 447–475.
Seilacher, A., 1994, How valid is Cruziana stra tig ra phy? Geolo-gische Rundschau, 83: 752–758.
Seilacher, A., 2007. Trace Fos sil Anal y sis. Springer, Berlin, 238pp.
Seilacher, A., 1985. Tri lo bite palaeo bi ol ogy and sub strate re la -tion ships. Trans ac tions of the Royal So ci ety of Ed in burgh.Earth Sci ences, 76: 231–237.
Seilacher, A. & Pflüger, F., 1994. From biomats to ben thic ag ri -cul ture: a biohistoric rev o lu tion. In: Krumbein, W. E., Pat er -son, D. M. & Stal, L. J. (eds), Biostabilization of Sed i ments.Bibliotheks und Informationssystem der Carl von OssietzkyUniversität Oldenburg, pp. 97–105.
Sikorska, M., 2000. Silification his tory of Cam brian sand stones inthe Wiœniówka area, Holy Cross Mts (Cen tral Po land). Prze-gl¹d Geologiczny, 48: 251–258. [In Pol ish, Eng lish ab stract].
Simpson, S., 1957. On the trace-fos sil Chondrites. Geo log i cal So -ci ety of Lon don, Quar terly Jour nal, 112: 475–499.
Stephenson, L. P., Plumley, W. J. & Palciauskas, V. V., 1992. Amodel for sand stone com pac tion by grain interpenetration.Jour nal of Sed i men tary Re search, 62: 11–22.
Studencki, M., 1994. Wiœniówka Du¿a quarry. In: Kowalczewski,Z., Szulczewski, M., Migaszewski, Z. & Jarecka-Strycz, K.(eds), Europrobe Trans-Eu ro pean Su ture Zone Work shop,Ex cur sion Guide book, the Holy Cross Moun tains, Pol ishGeolog i cal In sti tute, Kielce, 51–57 pp.
Wetzel, A., & Reisdorf, A. G., 2007. Ichnofabrics elu ci date the ac -cu mu la tion his tory of a con densed in ter val con tain ing a ver ti -cally emplaced ich thyo saur skull. In: Bromley, R. G., Bua-tois, L. A., Mángano, M. G., Genise, J. F. & Melchor, R. N.(eds), Sed i ment-Or gan ism In ter ac tions: A Mul ti fac eted Ich-nology. SEPM Spe cial Pub li ca tions, 88: 241–251.
¯yliñska, A. & Radwañski, A., 2008. Stop 2 – Wiœniówka Du¿a,Up per Cam brian. In: Pieñkowski, G. & Uchman, A. (eds),Ichnologial Sites of Po land: The Holy Cross Moun tains andthe Carpathian Flysch. The Pre-Con gress and Post-Con gress Field Trip Guide book. Pol ish Geo log i cal In sti tute, Warszawa, pp. 37–46.
¯yliñska, A., Szczepanik, Z. & Salwa, S., 2006. Cam brian of theHoly Cross Moun tains, Po land; biostratigraphy of the Wiœ-niówka Hill suc ces sion. Acta Geologica Polonica, 56: 443–461.
RUSOPHYCUS PRES ER VA TION FROM FURONGIAN 327