
Cognition enhancement by the acetylcholine releaser DuP 996
14
Drug Development Research 19:301-314 (1990) Cognition Enhancement by the Acetylcholine Releaser DuP 996 Leonard Cook, Victor J. Nickolson, George F. Steinfels, Kenneth W. Rohrbach, and Victor J. DeNoble Medical Products Department, E. 1. du pont de Nemours & Co., Inc., Wilmington, Delaware ABSTRACT Cook, L., V.J. Nickolson, G.F. Steinfels, K.W. Rohrbach, and V.J. DeNoble: Cognition enhancement by the acetylcholine releaser DuP 996. Drug Dev. Res. 19:301-314, 1990. DuP 996, 3,3-Bis(4-pyridinylmethyl)-l -phenylindolin-2-one, a potent in vitro and in vivo releaser of acetylcholine (ACh), dopamine (DA), and serotonin (5HT) in rat brain, signifi- cantly enhanced the performance of rats and mice in several behavioraltest procedures. At doses of 0.01-0.1 mgikg S.C. DuP 996 protected against a hypoxia-inducedpassive avoid- ance deficit in rats. In active avoidance procedures, DuP 996 enhanced acquisition of responses: in rats, at doses between 0.085 and 0.85 mgikg S.C. and 0.25 and 0.85 mgikg p.0.; in mice, at doses between 0.85 and 2.5 mgikg S.C. These effects occurred without any alteration of sensitivity to foot-shock. In addition, DuP 996 prevented a C0,-induced reten- tion deficit of a passive avoidance response when administered prior to acquisition testing. In a test for acquisition of lever pressingfor food in the rat, DuP 996 increasedthe proportion of animals acquiring this response. Thus, DuP 996 was active in both the shock- and appetitive-motivatedprocedures and was shown to enhance performance levels when ad- ministered post-training as well as before training trials. These results suggest that DuP 996 may be useful in the treatment of cognition dysfunction. Key words: Alzheimer’s disease, dementia, cholinergic, learning and memory INTRODUCTION Several lines of converging evidence suggest that the cerebral cholinergic system is involved in cognitive processes. For example, studies have shown that cognitive performance Received final version October 26, 1989; accepted November 10, 1989 Address reprint requests to Leonard Cook, Ph.D., E. I. du Pont de Nemours & Co., Inc., Experimental Station, P. 0. Box 80400, Wilmington, DE 19880-0400. 0 1990 Wiley-Liss, Inc.
-
Upload
leonard-cook -
Category
Documents
-
view
212 -
download
0
Transcript of Cognition enhancement by the acetylcholine releaser DuP 996

Drug Development Research 19:301-314 (1990)
Cognition Enhancement by the Acetylcholine Releaser DuP 996 Leonard Cook, Victor J. Nickolson, George F. Steinfels, Kenneth W. Rohrbach, and Victor J. DeNoble
Medical Products Department, E. 1. du pont de Nemours & Co., Inc., Wilmington, Delaware
ABSTRACT
Cook, L., V.J. Nickolson, G.F. Steinfels, K.W. Rohrbach, and V.J. DeNoble: Cognition enhancement by the acetylcholine releaser DuP 996. Drug Dev. Res. 19:301-314, 1990.
DuP 996, 3,3-Bis(4-pyridinylmethyl)-l -phenylindolin-2-one, a potent in vitro and in vivo releaser of acetylcholine (ACh), dopamine (DA), and serotonin (5HT) in rat brain, signifi- cantly enhanced the performance of rats and mice in several behavioral test procedures. At doses of 0.01-0.1 mgikg S.C. DuP 996 protected against a hypoxia-induced passive avoid- ance deficit in rats. In active avoidance procedures, DuP 996 enhanced acquisition of responses: in rats, at doses between 0.085 and 0.85 mgikg S.C. and 0.25 and 0.85 mgikg p.0.; in mice, at doses between 0.85 and 2.5 mgikg S.C. These effects occurred without any alteration of sensitivity to foot-shock. In addition, DuP 996 prevented a C0,-induced reten- tion deficit of a passive avoidance response when administered prior to acquisition testing. In a test for acquisition of lever pressing for food in the rat, DuP 996 increased the proportion of animals acquiring this response. Thus, DuP 996 was active in both the shock- and appetitive-motivated procedures and was shown to enhance performance levels when ad- ministered post-training as well as before training trials. These results suggest that DuP 996 may be useful in the treatment of cognition dysfunction.
Key words: Alzheimer’s disease, dementia, cholinergic, learning and memory
INTRODUCTION
Several lines of converging evidence suggest that the cerebral cholinergic system is involved in cognitive processes. For example, studies have shown that cognitive performance
Received final version October 26, 1989; accepted November 10, 1989
Address reprint requests to Leonard Cook, Ph.D., E. I. du Pont de Nemours & Co., Inc., Experimental Station, P. 0. Box 80400, Wilmington, DE 19880-0400.
0 1990 Wiley-Liss, Inc.

302 Cook et al.
is enhanced under conditions of cholinesterase inhibition, which increases the availability of acetylcholine (ACh) [Bartus, 1978, 1979; Deutsch, 19831. Similarly, direct nicotinic and muscarinic cholinergic agonists such as nicotine, arecoline, and oxotremorine have been shown to improve cognitive performance [Bartus et al., 19821, while anticholinergic agents such as scopolamine and hemicholinium impair learning and memory processes in both man and experimental animals [Bartus, 1978; Bohdanechy and Jarvik, 1967; Caulfield et al., 1981; Drachman, 19781.
The progressive dementia of Alzheimer’s disease (AD) appears to be associated with changes in several neurotransmitter systems, especially the neocortical cholinergic pathway. The extent of the loss of cortical cholinergic integrity is highly correlated with the severity of dementia [Davies and Maloney, 1976; Reisine et al., 1978; Whitehouse et al., 1981, 1982; Coyle et a]., 19831. For example, studies of neocortical biopsy samples from demented patients have shown that histological evidence of AD (senile plaques and tangles) was ac- companied by markedly reduced ACh synthesis, choline uptake, and choline acetyltransferase (CAT) activity [Sims et al., 19831. Further, several investigations have shown that the loss of cortical cholinergic innervation in AD can be traced to a degeneration in the nucleus basalis of Meynert [NBM; Price et al., 1982; Whitehouse et al., 19821. Animal studies confirm that loss of the cholinergic neurons that arise from the NBM and innervate cerebral neocortex produces significant behavioral impairments that are directly related to cortical cholinergic dysfunction [Loconte et a]., 1982; Bartus et al., 1985; Irle and Markowitsch, 19871.
Recently, reports of the amelioration of behavioral deficits and dementia symptoms in AD by the use of acetylcholinesterase (AChE) inhibitors such as physostigmine [Davis and Mohs, 1979, 19821 and tetrahydroaminoacridine [THA; Sitaram, 1984; Summers et al., 19861 have renewed interest in the search for cholinomimetic agents which could be of value in the treatment of AD. The AChE inhibitors apparently have narrow therapeutic windows and severe adverse side effects [Becker and Giacobini, 19881, and direct cholinergic receptor agonists, such as oxotremorine or carbachol, have been employed with only limited success thus far in the pharmacotherapeutic treatment of AD; the latter requires an intracerebral mode of administration [Harbaugh et al., 19841.
It has been suggested that drugs which enhance stimulation-induced Ach release in the brain would have advantages over both AChE inhibitors and receptor agonists [Nickolson et al., 19891. In the preceding paper in this journal, DuP 996, 3,3-Bis(4-pyridinylmethyl)- 1 -phenylindolin-2-one, was reported to have such ACh release-enhancing activity as well as enhancement of stimulated release of dopamine (DA) and serotonin (5HT). In the present experiments, DuP 996 was profiled with regard to its efficacy in several tests of cognitive performance.
MATERIALS AND METHODS Subjects
For all rat behavioral experiments, except where indicated, naive male Sprague-Dawley rats (Charles River Farms) were housed three or four to a cage for at least 7 days prior to testing. Rat weights ranged from 150 to 215 g. They had ad lib access to standard rat chow (Purim) and water.
Hypoxia-Induced Passive Avoidance Deficit in Rats
The experimental sessions were conducted in a two-compartment shuttlebox: one com- partment, made of clear plastic, measured 21(L) x 24.5(H) X 17(W) cm and had a perfo- rated, clear plastic floor; the other compartment, made of black plastic, measured 30.5(L) X
20.3(H) x 21.5(W) cm and had a floor made of 4 mm stainless steel rods spaced 1.2 cm apart. These steel rods were connected to a Coulbourn Instruments Grid Floor Shocker which provided a scrambled footshock. The clear compartment was illuminated with a lamp con-

DuP 996 Cognition Enhancement 303
taining a 60 W bulb placed 36 cm above the clear Compartment. The two compartments were separated by a solenoid-operated slide door (Lafayette Instrument Co., Lafayette, IN). Open- ing or closing of the slide door triggered a Coulbourn Instruments Electronic Counter to record acquisition and retention latencies.
Rats were exposed to an hypoxic environment before passive avoidance training. The hypoxia chamber was constructed of clear plastic, measured 32.5(L) X 22.5(H) X 23(W) cm, and was continuously perfused with a gas mixture of oxygen and nitrogen. The flow rate was adjusted such that the gas turnover in the chamber was 15 liters per min.
Passive avoidance training began by placing the rat into the clear compartment, and following a 10 sec delay, the slide door was raised. Once the rat moved completely into the dark compartment (all four paws on the shock grid floor), the slide door was lowered and after a 10 sec delay, a 1.5 mA shock was applied to the grid floor for three sec which was followed by a 10 sec delay and another three sec shock. The rats were immediately removed from the dark compartment after receiving the second shock and returned to their home cage. Vehicle or test compounds were administered 1 min after PA training. Rats not entering the dark compartment within 40 sec were removed from the study. A retention test was given 4 hr later. It proceeded in the same manner as the training session except no shock was applied to the grid floor and the rats were provided a maximum entry latency to the dark compartment of 300 sec. Memory disruption was induced by exposing the rats to a gas mixture containing percentages of oxygen supplemented with nitrogen for 30 min immediately prior to passive avoidance training. Median entry latencies during the retention tests were compared for treated and vehicle control groups (N = 18 minimum per group) using a Mann-Whitney U-test.
C0,-Induced Passive Avoidance Deficit in Rats
Testing was carried out in a modified Coulbourn E10-10 modular test cage equipped with a removable grid floor. The inside of the cage was painted flat-black (dark compartment). A 16.9 cm X 5.9 cm metal grid platform was mounted outside the front center of the cage. A black plastic slide door separated the platform from the cage. A microswitch mounted above the door was activated when the door was raised and it started a timer which recorded a rat’s latency to enter the dark Compartment.
The rats were injected with DuP 996 s.c., and 30 min later, testing began. A rat was first placed on the platform so that his head faced away from the slide door. Next, the slide door was opened and any rat that remained on the platform for more than 20 sec was excluded from the experiment. When the rat entered the cage (all four paws on the grid floor inside the dark compartment), the door closed, which automatically stopped the timer and switched on a 0.9 mA footshock for 3 sec. The rat was then taken from the test cage and placed in a chamber containing air or 80% CO2/2O% 0,. Rats exposed to C02/02 became unconscious after approximately 15 sec and remained in the chamber for a total of 2 min, following which they were returned to their home cage. Retention testing was performed 24 hr later. Retention trials were conducted in the same way as acquisition trials, except that footshock was not given. Rats were allowed a maximum of 180 sec to enter the dark compartment.
The percentage of animals reaching the retention criterion (1 80 sec) was calculated for each treatment group and differences between groups were tested with the chi-square test.
Rat Active Avoidance
Testing was carried out in a Coulbourn model ElO-10 large modular test cage equipped with a removable shock grid floor and a houselight mounted at the top center of one wall. A 27 cm wooden pole (1.9 cm diam) with parallel diagonal notches was mounted perpendicular to the ceiling. Footshock was supplied by a Coulbourn model E13-08 grid floor shocker. Timing and control of trials were programmed with Coulbourn equipment interfaced to a Commodore 64 computer. Each rat received two blocks of five trials (one A M and one P M , 4 hr apart) daily for 5 days. At the beginning of the experiment the rat was first placed on the

304 Cook et al.
pole and then removed from the test cage 45 sec later. During the test trials the rats were placed on the floor of the cage, facing the pole, and the cage top was closed. A trial consisted of a 10 sec presentation of the CS (illumination of the house light) followed by delivery of a 1 mA footshock for 10 sec. At the end of each trial, the houselight and footshock were terminated and the rat was removed from the test chamber. An avoidance response was defined as a pole climb prior to the onset of shock. An escape was defined as a pole climb during the footshock.
In the first and second experiment, rats were treated with DuP 996 30 min before the first trial of each block. In the third experiment, rats were treated with DuP 996 immediately after the last trial of each block.
The cumulative number of avoidances was calculated for each treatment group. The slope of a regression curve was calculated (cumulative avoidances per block versus the block number) and this was used as the measure of performance. The performances of the drug- treated groups and the corresponding vehicle-treated group were compared using the Student’s t-test . Footshock Sensitivity Testing
Rats were injected with DuP 996, s.c., 30 min prior to testing, and then tested indi- vidually in a Coulbourn E10-10 modular test cage equipped with a removable grid floor. A rat was permitted to acclimate to the test cage for 60 sec; then footshock was applied for 1.5 sec with a Coulbourn E13-08 Grid Floor Shocker and the rat was scored for vocalization, running, jumping or flinching. The order of presentation of the shocks were 0.7,0.6,0.5,0.4,0.3,0.2, 0.1, and 0 mA, with 20 t 3 sec between shocks. The lowest current level at which a reaction was elicited was considered to be the footshock sensitivity threshold for a particular drug dose. Shock levels at each dose were compared with ANOVA.
Mouse Active Avoidance
Unfasted male CRiC57BW6 mice (Charles River) were used in the active avoidance experiment. They weighed 17-20 g, were housed four to a cage, and food and deionized water were available ad lib during testing.
Testing was carried out in modified Coulbourn model E10-11 small test cages set within sound-attenuated cubicles. The cages were equipped with stationary shock grid floors and shock was supplied by an E13-08 grid floor shocker. A Coulbourn motor-driven retractable lever was located at the bottom center of one wall. Photobeams mounted behind the lever detected lever presses. A houselight located in the center of the ceiling provided the cage illumination. Timing and data acquisition were carried out by Commodore 64 computers interfaced with the Coulbourn equipment.
Prior to the first trial, all mice were given a 60 sec habituation period during which the houselight was on and the lever extended. After this pre-exposure, the houselight was extin- guished and the lever retracted for 5 sec (Inter-Trial-Interval: ITI). After the IT1 the light was illuminated and the lever was extended for 5 sec (conditioned stimulus-CS) after which a 0.5 mA scrambled footshock (unconditioned stimulus-US) was delivered. Footshock was pre- sented for a maximum of 10 sec or until a lever press was made. At the termination of the footshock, the lever was retracted and the houselight extinguished.
A lever press made during the CS period was considered an avoidance and resulted in retracting the lever, cancelling the footshock and extinguishing the houselight. A lever press made during the 10 sec US period was considered an escape and resulted in retracting the lever, terminating the footshock, and extinguishing the houselight.
A session consisted of 50 trials and the mice were dosed (s.c.) 30 min before the session. The data from the last 25 trials were used in the statistical analysis. Mice that failed to escape 10 or more times during the session were excluded from the data analysis. In each treatment group, the mouse with the greatest number of avoidances and the mouse with the fewest avoidances also were excluded from the analysis. Data was analyzed by ANOVA.

DuP 996 Cognition Enhancement 305
Acquisition of Lever Pressing for Food in Rats
Male CDF rats (Charles River) weighing 150-180 g were used. Animals underwent a 4 day regimen of food deprivation (with water continuously available) and adaptation to the test chamber. This procedure was as follows. Food was removed on day one. On day two the animals were given 5 g of food. On days 3 and 4 the animals were adapted to the operant chamber using a program of automatic food delivery (45 mg food pellet, Bioserve, Inc.) every minute for 30 min (this is the only food the animals received on these days). Food delivery was accompanied by a magazine light (2 sec) and feeder click. On day 5 the adapted animals were exposed to the autoshape paradigm which consisted of alternating fixed ratio 1 and random interval 45 sec timeout (FRURI45-TO) schedule of food reward. A similar procedure has been described previously by Messing and Sparber [1983, 19851. The lever was active for 7.5 sec (cued with the houselight) on the FR1 schedule and each lever press yielded a 45 mg food pellet after an 8 sec delay. This alternated with a timeout (TO) period, with a random length (25-65 sec) for each trial and averaged 45 sec during which responding had no consequence. The procedure ran for 8 hr. In order to maintain the activity level of the rats for that time, a free pellet was delivered every 15 min.
The animals were adapted to the operant chambers between 3:30 and 4:30 P.M. on days 3 and 4. The autoshape procedure on day 5 begins at approximately 5:OO P.M. and ran for 8 hr.
The number of correct responses was recorded for every 15 min period and the criteria for acquisition was 5 reinforced responses in a 15 min period. The percentage of animals acquiring the lever press response (e.g. reaching criteria) during the 8 hr procedure were compared with vehicle treated controls using the Fisher exact test for individual comparisons.
Electroencephalographic (EEG) Effects in Rats
In the EEG studies adult, male Fischer F-344 rats (Charles River, Kingston, NY) weighing 225-275 g at the time of surgery were used. Animals were fed Wayne Research Rodent Blocks (Wayne Co.) and water ab libitum. They were maintained on a reverse light- dark cycle with lights on 2030-0830 hr and lights off 0830-2030 hr.
Prior to surgery, rats were first anesthetized with a combination of ketamine (105 mg/kg, i.m.) and xylazine (10 mgikg, i.m.). Stainless steel screws (#O-80) served as cortical elec- trodes and were implanted ipsilaterally over the frontal cortex at the following coordinates: 2 mm anteriori2 mm lateral to bregma, and 2 mm posterior/3 mm lateral to bregma. Wire leads from the EEG electrodes were soldered into a miniature connector which was secured to the skull with dental acrylic. Rats were allowed at least 1 week recovery from surgery before experiments were begun.
The cortical EEG was recorded bipolar (right hemisphere) with a ground electrode located on the occipital cortex. EEG activity was recorded on a Grass Model 7D polygraph with the 0.5 A high and low bandpass filters set at 1 Hz and 75 Hz, respectively. EEG data was digitized online (Nicolet Pathfinder I1 computer) at a rate of 125fsec. Power spectral epochs were derived from an average of fifty 4 sec samples, and spectral smoothing was accomplished using a 3-point Hanning window. Spectral power was calculated for the fol- lowing bandwidths (1.0-5.0 Hz, delta), 5.25-9.0 Hz, theta), (9.25-13.00 Hz, alpha), 13.25- 25.00 Hz, beta-1), and (25.25-60.00 Hz, beta-2).
The experiments were performed in a darkened room and rats were individually housed in chambers measuring 14 cm X 14 cm X 28 cm. To permit unrestrained movement of the rat during EEG recordings, each cage was equipped with a swivel connector with concentric mercury pools which served as noise-free contacts.
Vehicle or drug was administered and EEG was sampled at 15, 30, and 45 min post- injection. The vigilence peak effect for each rat was defined as the maximum percentage change in beta activity compared to vehicle control. The peak effect, independent of time, was
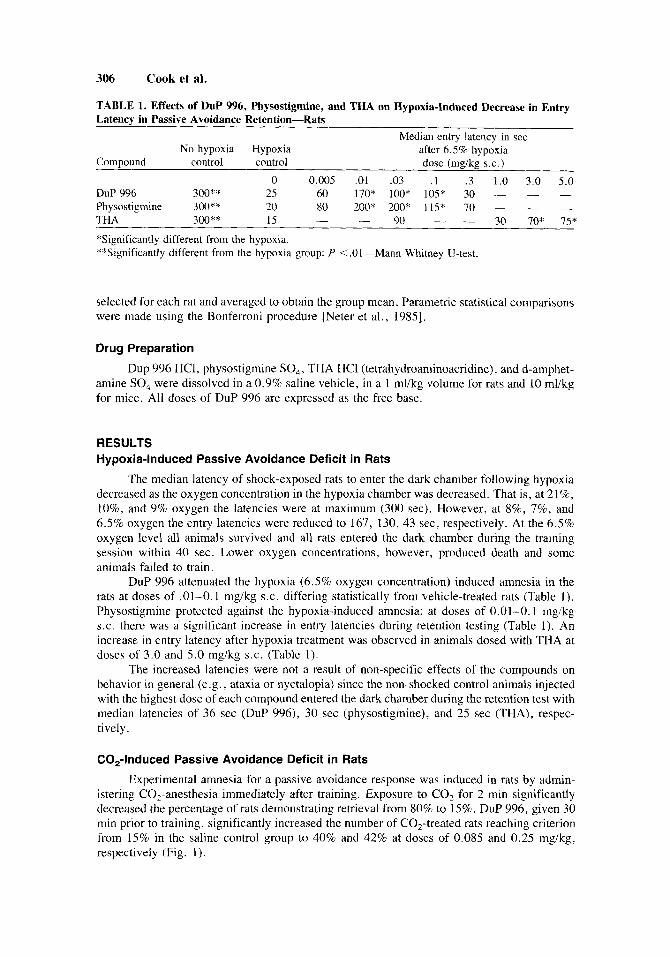
306 Cook et al.
TABLE 1. Effects of DuP 996, Physostigmine, and THA on Hypoxia-Induced Decrease in Entry Latency in Passive Avoidance Retention-Rats
Median entry latency in sec No hypoxia Hypoxia after 6.5% hypoxia
Compound control control dose (mg/kg s.c.) 0 0.005 .O1 .03 . I . 3 1.0 3.0 5.0
DuP 996 300** 25 60 170* 100" 105" 30 - - - Physostigmine 300** 20 80 200* 200* 115* 70 - - - THA 300** 15
"Significantly different from the hypoxia. **Significantly different from the hypoxia group: P <.Ol-Mann-Whitney U-test.
90 - - 30 70" 15" - -
selected for each rat and averaged to obtain the group mean. Parametric statistical comparisons were made using the Bonferroni procedure [Neter et al., 19851.
Drug Preparation
Dup 996 HCI, physostigmine SO,, THA HCI (tetrahydroaminoacridine), and d-amphet- amine SO, were dissolved in a 0.9% saline vehicle, in a 1 mlikg volume for rats and 10 mlikg for mice. All doses of DuP 996 are expressed as the free base.
RESULTS Hypoxia-Induced Passive Avoidance Deficit in Rats
The median latency of shock-exposed rats to enter the dark chamber following hypoxia decreased as the oxygen concentration in the hypoxia chamber was decreased. That is, at 2 1 %, lo%, and 9% oxygen the latencies were at maximum (300 sec). However, at 8%, 7%, and 6.5% oxygen the entry latencies were reduced to 167, 130, 43 sec, respectively. At the 6.5% oxygen level all animals survived and all rats entered the dark chamber during the training session within 40 sec. Lower oxygen concentrations, however, produced death and some animals failed to train.
DuP 996 attenuated the hypoxia (6.5% oxygen concentration) induced amnesia in the rats at doses of .01-0.1 mgikg S.C. differing statistically from vehicle-treated rats (Table 1). Physostigmine protected against the hypoxia-induced amnesia; at doses of 0.01-0.1 mgikg S.C. there was a significant increase in entry latencies during retention testing (Table 1). An increase in entry latency after hypoxia treatment was observed in animals dosed with THA at doses of 3.0 and 5.0 mgikg S.C. (Table 1).
The increased latencies were not a result of non-specific effects of the compounds on behavior in general (e.g., ataxia or nyctalopia) since the non-shocked control animals injected with the highest dose of each compound entered the dark chamber during the retention test with median latencies of 36 sec (DuP 996), 30 sec (physostigmine), and 25 sec (THA), respec- tively.
C0,-Induced Passive Avoidance Deficit in Rats
Experimental amnesia for a passive avoidance response was induced in rats by admin- istering C0,-anesthesia immediately after training. Exposure to CO, for 2 min significantly decreased the percentage of rats demonstrating retrieval from 80% to 15%. DuP 996, given 30 min prior to training, significantly increased the number of C0,-treated rats reaching criterion from 15% in the saline control group to 40% and 42% at doses of 0.085 and 0.25 mg/kg, respectively (Fig. 1) .

DuP 996 Cognition Enhancement 307
= 100 0 z .- L a,
!I 5 80
v)
a LI
C a,
a,
c
+ 20
2 L O
3 ***
Sal Sal 0.025 0.085 0.25 0.85 S.C.
Air c02
Fig. 1. Effect of DuP 996 on the percentage of rats showing (180 sec) retention following exposure to CO, amnesia. Each bar is a mean of 8 rats and asterisks show the doses that were significantly different from the vehicle non-CO, group. ***Significantly different from SallCO,, P < 0.001. *Significantly different from SallCO,, P < 0.05.
Rat Active Avoidance
The effect of DuP 996 on active avoidance was tested in rats. DuP 996, given either subcutaneously at doses of 0.085 to 0.85 mg/kg or orally at 0.25 and 0.85 mg/kg 30 min before testing increased the number of pole-jump avoidances (Figs. 2, 3). It is interesting to note that the increase in the number of avoidances was not restricted to pre-test administration since DuP 996 given S.C. (0.25 and 0.85 mgikg) immediately after training also increased the number of pole-jump avoidance responses (Fig. 4).
Footshock Sensitivity in Rats
Studies were carried out to evaluate whether the improvements in footshock-motivated behavior observed with DuP 996 were due to a drug-induced increase in footshock sensitivity. These studies demonstrated that DuP 996 did not significantly alter the average threshold at which rats reacted to noxious footshock (SAL, 0.28 mA; 0.25 mgikg s.c., 0.22 mA; 0.85 mgikg s.c., 0.22 mA; 2.5 mg/kg s.c., 0.20 mA).
Mouse Active Avoidance
As shown in Table 2, DuP 996, given S.C. 30 min before training, significantly enhanced the performance of mice on a lever press footshock-motivated active avoidance task over a broad range of doses (0.085-2.5 rngikg s.c.). Physostigmine also enhanced performance at 0.03 mgikg with lower doses showing inactivity (Table 2) . In contrast, d-amphetamine (. 1-1 .O mgikg s.c.) was inactive in this test (Table 2).
Acquisition of Lever Press for Food in Rats The proportion of animals acquiring the lever press response over the 8 hr session is low,
22-33%, due to the lirnited access to the lever and to the long time interval (8 seconds) between response and its reinforcement. DuP 996 significantly increased the percent

308 Cook et al.
$ 4
Y O 3
.- c Lo - 0 0 - m L a Q 2 v) a 0 t m E l 2 a S m
Sal 0.085 0.25 0.85 2.5
Dose of DuP 996 (mg/kg s.c.)
$ 0
Fig. 2. The effect of DuP 996 administered S.C. 30 min before each block of 5 trials on rat pole-jump active avoidance. The mean number of avoidances per treatment group (N =6 per group) from the last 25 trials of the 50-trial test were cumulated and compared to the vehicle group. Asterisks indicate doses that are significantly different from vehicle, P < 0.05.
v)
.- a 3 c Lo
0 Y 0
c
- 0 2 rn L a Q 0 a 0
2 ’ 9 a n
c m $ 0
Sal 0.25 0.85 2.5 Dose of DuP 996 (mg/kg p.0.)
Fig. 3. The effect of DuP 996 administered orally 30 min before each block of 5 trials on rat pole-jump active avoidance. The mean number of avoidances per treatment group (N = 9 per group) from the last 25 trials of the 50-trial test were cumulated and compared to the vehicle group. Asterisks indicate doses that are significantly different from vehicle, P < 0.05.

h
In
0
0 0
c
Y 3
m 2) -
0 . 2 v) e, 0 C ca .z 1 9 a C ca
Sal 0.085 0.25 0.85 2.5 g o
Dose of DuP 996 (mg/kg s.c.)
Fig. 4. The effect of DuP 996 administered S.C. immediately after each block of 5 trials on rat pole-jump active avoidance. The mean number of avoidances per treatment group (N = 6 per group) from the last 25 trials of the 50-trial test were cumulated and compared to the vehicle group. Asterisks indicate doses that are significantly different from vehicle, P < 0.05.
of animals acquiring the response at doses of 0.03 and 0.1 mg/kg. Neither physostigmine or THA significantly improved acquisition of the lever press response (Fig. 5).
EEG Effects in Rats
The results for DuP 996 (Fig. 6) show significant increases in the faster frequency bands, alpha and beta, and concommitant decreases in the slower frequency bands delta and theta. This pattern was seen for all tested doses (0.3, 1 .O, and 3.0 mgikg s.c.) and a maximal effect occurred at 1 .O mgikg. Similar trends of increased fast frequency activity were seen with physostigmine and amphetamine. In contrast, the lower doses of amphetamine tended to increase delta and decrease theta activity, whereas the highest tested dose decreased delta and increased theta activity.
DISCUSSION
DuP 996 enhanced behavioral performance under a variety of experimental conditions, suggesting that its pharmacological effects result in an improvement of cognitive function and are not due to any experimental treatment artifacts that might have been present in any individual test. In different tests DuP 996 enhanced performance whether treatment was given before or after acquisition. Post-training treatment with DuP 996, physostigmine and THA attenuated the hypoxia-induced passive avoidance retention deficit. While the effect of cho- linergic drugs on hypoxia-induced behavioral deficits (tight rope test scores) has been well documented [Gibson et al., 19831, this is the first reporting of cholinergically acting drugs preventing an hypoxia effect on a single trial passive avoidance test. When DuP 996 was administered acutely before training, as in the mouse active avoidance test, it increased the number of avoidance responses in a dose-dependent manner, indicating improved task acqui- sition. In the rat active avoidance procedure, DuP 996 increased the number of avoidances when administered either pre-trial or post-training suggesting that DuP 996 was effective both

310 Cook et al.
Ph y sostigmine
TABLE 2. Effect of DuP 996, Physostigmine, d-Amphetamine, and Piracetam on Mouse Lever-Press Active Avoidance Responding"
DOSE COMPOUND (mgikg, s.c.) MEAN S.E.M. N DuP 996 0.0 7.5 .61 62
0.025 8. 1 .9 8 0.085 12.0* 1 .o 11 0.25 12.3" 1.4 4 0.85 14.8* 3.3 4 2.5 12.3" 2.4 6 0.0 9.8 2.34 10 0.003 9.0 1.3 7 0.01 7.7 1.3 7 0.03 13.0* 2.0 9
0.1 8.0 I .9 7 0.3 8.5 1.3 6 1 .o 6.5 2.9 2 h
d- Amphetamine 0 8.3 1.5 9
"The data are expressed as the mean t number of avoidance responses made during the final 25 trials of testing. bBecause of locomotor stimulation ifiduced by the high dose of am- phetamine, many mice in this group were excluded for scoring 10 or more escape failures. *P < 0.05.
in the acquisition and consolidation of information. DuP 996 given immediately before train- ing of passive avoidance behavior also protected against C0,-induced retention deficits.
In the acquisition of lever press for food paradigm, no negative reinforcement (e.g., footshock) was involved; neither was there any type of hypoxia or cholinergic disruption employed. Performance levels (i.e., lever pressing for food) was low by arranging a weak association between response and reinforcement. The acetylcholine release enhancer DuP 996 improved the acquisition of the lever press response under these conditions, whereas the cholinesterase inhibitors physostigmine and THA did not. This suggests a qualitative different spectrum of effect for DuP 996 compared to cholinesterase inhibitors on certain cognitive functions.
Considerable literature supports the role of the central cholinergic system as being intimately involved in mediating critical cognitive processes [Bartus et al., 1982; Deutsch, 19831. For example, the anticholinergic drug scopolamine impairs learning and memory, but the quaternary scopolamine methylbromide, which is only active peripherally, has no effects on cognition [Drachman, 19771. Conversely, the AChE inhibitor physostigmine, can reverse scopolamine-induced deficits [Bartus, 19781, enhance suboptimal performance in young and aged primates [Bartus, 19791, and improve performance of rats with NBM lesions [Haroutu- nian et al., 19851.
DuP 996 is a potent enhancer of stimulation-induced release of ACh in vitro and in vivo [Nickolson et a]., 19891; however, it does not increase the baseline level of ACh release. DuP 996 is active in tests where physostigmine is also active, suggesting an enhancement of performance due to cholinomimetic properties for both compounds, although due to different mechanisms. Side effects do not appear to account for the effects of the enhanced performance of DuP 996, physostigmine, or THA. Data from our laboratory studies in rats show that physostigmine produced tremors at 0.3 mgikg s.c.; THA produced tremors at 6.0 mgikg s.c.; and DuP 996 produced tremors at 10.0 mgikg S.C. [DeNoble, unpublished results]. There was a comparatively larger margin of safety, in this regard, with DuP 996.

DuP 996 Cognition Enhancement 311
DuP 996 * * 300 i
0 .03 . I .3 1 DuP 996 (rng/kg p.0.)
P h ysost ig mine
c
150 c
5 100
50
0 L
0 0.00030.001 0.003 0.01 0.03 0.1
Physostigmine (rng/kg p.0.)
THA
0 0.6 1.3 2 3 THA (mg/kg p.0.)
Fig. 5 . The effect of DuP 996. physostigmine. and THA on the 8 hr acquisition of lewr press for food paradigm. DLIP 996 significantly increases the percentage of animals acquiring the task, while physo- stigmine and THA do not. Astcrisks indicate doses that are significantly different from vehicle (0). P < 0.05.
It is important to note that DuP 996 is also a dopamine and serotonin release enhancer [Nickolson et a l . , I9891 and it is possible that these effects may also play some role in its behavioral enhancing properties. It is unlikely that any non-specific stirnulant-like effect influenced the enhanced behavioral measures; for example, studies in our laboratory showed

312 Cook et al.
DuP 996 60 40
20 0 0.3 0 1.0
0 3.0
-20
-40 I, DELTA THETA ALPHA BETA
- 2 * t Physostig m ine
0.01 a 0.03 LBI 0.1
y -40 ; 1 I I i DELTA THETA ALPHA BETA a,
a, a 2
Amphetamine 60 1 40
20 P 0.1
0 IB1 1.0 m 0.3
-20
-40 DELTA THETA ALPHA BETA
Band
Fig. 6. phetamine. *P < 0.10; **P < 0.05.
The effects of DuP 996 on rat EEG power spectra in comparison to physostigmine and am-
that DuP 996 did not produce any increase in spontaneous locomotor activity [Johnson, unpublished results]. In this regard, DuP 996 enhanced activity in the mouse lever-press active avoidance task whereas amphetamine was inactive.
Age related changes in the human electroencephalogram (EEG) have been described by several laboratories. The general trend in the human EEG is a slowing of the background activity with increasing age [Matejcek and Devos, 1976; Duffy et al., 19841. Specifically, the higher the age, the slower the dominant frequency bands designated delta and theta. These changes in EEG activity are believed to indicate a deterioration in vigilance and may be due

DuP 996 Cognition Enhancement 313
to deficits in the regulatory systems for vigilance and cognition. Further, pathologic states associated with deficits, such as Alzheimer’s disease, are strongly correlated with increased delta activity and decreased beta (highest frequencies) activity [Saletu and Grunberger, 19851. Investigators in the field of pharmacogeriatrics have postulated that a drug that reverses age-related EEG changes should also produce improvement in the clinical status of the geri- atric patients. The EEG profile produced by DuP 996 suggests vigilance enhancing properties, a finding which also has been confirmed in Phase I clinical trials [Saletu et al., 19891.
A consideration of the biochemical, behavioral, and EEG results described suggests that DuP 996 may have utility in the treatment of cognition deficits resulting from cholinergic hypofunction.
REFERENCES
Bartus, R.T.: Evidence for a direct cholinergic involvement in the scopolamine-induced amnesia in monkeys: Effects of concurrent administration of physostigmine and methylphenidate with sco- polamine. Pharmacol. Biochem. Behav. 9:833-836, 1978.
Bartus, R.T.: Physostigmine and recent memory: Effects in young and aged non-human primates. Science
Bartus, R.T., Dean, R.L., Beer, B., and Lippa, A.S.: The cholinergic hypothesis of geriatric memory dysfunction. Science 217:408-417, 1982.
Bartus, R.T., Flicker, C., Dean, R.L., Pontecorvo, M., Figueiredo, J.C., and Fisher, S .K.: Selective memory loss following nucleus basalis lesions: long term behavioral recovery despite persistent cholinergic deficiencies. Pharmacol. Biochem. Behav. (23) 1: 125-135, 1985.
Becker, R.E. and Giacobini, E.: Mechanisms of cholinesterase inhibition in senile dementia of the Alzheimer type: clinical pharmacological and therapeutic aspects. Drug Dev. Res. 12: 163-195, 1988.
Bohdanechy, Z. and Jarvik M.E.: Impairment of one trial passive avoidance learning in mice by sco- polamine methylbromide and physostigmine. Int. J . Neuropharmacol. 6:217-222, 1967.
Caulfield, M.P., Fortune, D.H., Roberts, P.M., and Stubley, J.K.: Intracerebroventricular hemicholin- ium-3 (HC-3) impairs learning of a passive avoidance task in mice. Br. J . Pharmacol. 74:865, 198 1.
Coyle, J.T., Price, D., and DeLong, M.: Alzheimer’s Disease: A disorder of cortical cholinergic inner- vation. Science 219:1184-1190, 1983.
Davies, P. and Maloney, A.J.F.: Selective loss of central cholinergic neurons in Alzheimer’s disease. Lancet Ik1403, 1976.
Davis, K. and Mohs, R.C.: Enhancement of memory by physostigmine. N. Eng. J . Med. 301:946, 1979. Davis, K.L. and Mohs, R.C.: Enhancement of memory processes in Alzheimer’s Disease with multiple-
dose intravenous physostigmine. Am. J . Psychiatry 139:1421-1424, 1982. Deutsch, J.A.: The cholinergic synapse and the site of memory. In Deutsch, J.A. (ed.): “The Physio-
logical Basis of Memory,” 2nd edition. New York: Academic Press, 1983, pp. 367-386. Domjan, M. and Burkhard, B.: “The Principles of Learning and Behavior.” Monterey, CA: Brooks/Cole
Publishing Company, 1982. Drachman, D.: Memory and cognitive function in man: does the cholinergic system have a specific role?
Neurology 27:783-790, 1977. Drachman, D.A.: Memory, dementia and the cholinergic system. In Katzman, R., Terry, R.D., and
Bick, K.L. (eds.): “Alzheimer’s Disease: Senile Dementia and Related Disorders,” Volume 7. New York: Raven Press, 1978, pp. 141-148.
Duffy, F.H., Albert, M.S. and McAnulty, G.: Brain electrical activity in patients with presenile and senile dementia of the Alzheimer type. Ann. Neurol 16:439-448, 1984.
Gibson, G.E., Pelmas, C.J. and Peterson, C.: Cholinergic drugs and 4-aminopyridine alter hypoxia- induced behavioral deficits. Pharm. Biochem. Behav 18:909-916, 1983
Haroutunian, V., Kanof, P., and Davis, K.L.: Pharmacological alleviation of cholinergic lesion induced memory deficits in rats. Life Sci. 37(10):945-952, 1985.
Harbaugh, R.E., Roberts, D.W., Coombs, D.W., Saunders, R.L., and Reeder, T.M.: Preliminary Report: Intracranial cholinergic drug infusion in patients with Alzheimer’s Disease. Neurosurgery
206: 1087-1089, 1979.
15:514-518, 1984.

314 Cook et al.
Hepler, D.J., Wenk, G.L., Cribbs, B.L., Olton, D.S., and Coyle J.T.: Memory impairments following basal forebrain lesions. Brain Res. 34653-14, 1985.
Irle, E. and Markowitsch, H.J.: Basal forebrain-lesioned monkeys are severely impaired in tasks of association and recognition memory. Ann. Neurol. (22):735-743, 1987.
Kuhar, M.J., Sethy, M.J., Roth, R.H., and Aghajanian, G.K.: Choline: Selective accumulation by central cholinergic neurons. J. Neurochem. 20581-593, 1973.
Lerer, B.E., Warner, J., Friedman, E., Vincent, G., and Gamzu, E. Cortical cholinergic impairment and behavioral deficits produced by kainic acid lesions of rat magnocellular basal forebrain. Behav. Neurosci. 99(4):661-677, 1985.
Lerer, B .E. and Nickolson, V.J.: A longitudinal examination of the behavioral and biochemical deficits produced by ibotenate lesions of the rat nucleus basalis magnocellularis. Soc. Neurosci. Abs. 12:553, 1986.
Loconte, G., Bartolini, L., Casamenti, F., Marloconcini-Pepeu, and Pepeu, G.: Lesions of cholinergic forebrain nuclei: Changes in avoidance behavior and scopolamine actions. Pharmacol. Biochem. Behav. 17(5):933-937, 1982.
Lowry. O.H., Rosebrough, J.J., Farr, A.L., and Randall, R.J.: Protein measurement with the folin phenol reagent. J. Biol. Chem. 193:265-275, 1951.
Matejcek, M. and Devos, J.E.: Selected methods of quantitative EEG analysis and their applications in psychotropic drug research. In Kellaway, P. and Petersen, I. (eds) Quantitative Analytic Studies in Epilepsy, New York, Raven Press, 1976, pp. 183-205.
Messing, R.B. and Sparber, S.B .: Des-Gly-Vasopressin improves acquisition and slows extinction of autoshaped behavior. Eur. J . Pharmacol. 89:43-51, 1983.
Messing, R.B. and Sparber, S.B.: Greater task difficulty amplifies the facilitatory effect of des-glycinam- ide arginine vasopressin on appetitively motivated learning. Behav. Neurosci. 99.1 114-1 119, 1985.
Neter, J., Wasserman, W., and Kutner: Applied linear statistical models. Homewood, Illinois: Richard E. Irwin, Inc., 1985.
Nickolson, V.J., Tam, S.W., Myers, M.J., and Cook, L.: DuP 996 (3,3-bis(4-pyridinylmethyl)-l- phenylindolin-2-one) enhances the stimulus-induced release of acetylcholine in rat brain in vitro and in vivo. Drug Dev. Res. 19:285-300, 1990.
Quartermain, D. and Altman, H. J.: Facilitation of retrieval by d-amphetamine following anisomycin- induced amnesia. Physiol. Psychol. 10(3):283-292, 1982.
Reisine, T., Yamamura, H.1., Bird, E.D., Spokes, E., and Enna, S.J.: Pre-and post-synaptic neuro- chemical alteration in Alzheimer’s disease. Brain Res. 159:477-481, 1978.
Saletu, B. and Grunberger, J.: Memory dysfunction and vigilance: neurophysiological and psychophar- macological aspects. In Olton, D.S., Gamzu, E. , and Corkin, S. (eds.) Memory Dysfunctions: An Integration of Animal and Human research From Preclinical and Clinical Perspectives, Annals of New York Academy of Sciences, vol. 444, 1985, pp.406-427.
Saletu, B., Darragh, A , , Salmon, P. and Coen, R.: EEG brain mapping in evaluating the time course of the central action of DuP 996-a new acetylcholine releasing drug. Br. J . Clin. Pharmacol.
Sims, N.R., Bowen, D.M., Allen, S.J., Smith, C.C.T., Neary, D., Thomas, D.J., and Davison, A.N.: Presynaptic cholinergic dysfunction in patients with dementia. 3 . Neurochem. 40503-509, 1983.
Sitaram, N.: Cholinergic hypothesis of human memory: Review of basic and clinical studies. Drug Develop. Res. 4:481-488, 1984.
Summers, W .K., Majovski, L.V., Marsh, G.M., Tachiki, K. , and Kling, A,: Oral tetrahydroaminoacri- dine in long-term treatment of senile dementia, Alzheimer type. N. Eng. J. Med. 315:1241-1245, 1986.
Whitehouse, P.J., Price, D.L., Clark, A.W., Coyle, J.T., and DeLong, M.R.: Alzheimer disease: Evidence for selective loss of cholinergic neurons in the nucleus basalis. Ann. Neurol. 10:
Whitehouse, P.J., Price, D.L., Clark, A.W., Coyle, J.T., and DeLong, M.R.: Alzheimer’s disease and
28~1-16, 1989.
122-126, 1981.
senile dementia: Lass of neurons in the basal forebrain. Science 215: 1237-1 239, 1982.


















