
Cloning and expression of a transcription factor activator...
6
Short communication Cloning and expression of a transcription factor activator protein-1 (AP-1) member identified from manila clam Venerupis philippinarum Luning Wu a,b,1 , Lei Zhang b,1 , Jianmin Zhao b, ⁎, Xuanxuan Ning c , Changkao Mu a , Chunlin Wang a, ⁎⁎ a School of Marine Science of Ningbo University, Zhejiang, Ningbo 315211, PR China b Key Laboratory of Coastal Zone Environmental Processes and Ecological Remediation, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai 264003, PR China c Yantai Oceanic Environmental Monitoring Central Station, State Oceanic Administration, Yantai 264006, PR China abstract article info Article history: Received 14 September 2014 Received in revised form 12 December 2014 Accepted 13 December 2014 Available online 16 December 2014 Keywords: Transcription factor activator protein-1 Venerupis philippinarum Immune response Bacterial challenge The transcription factor activator protein-1 (AP-1) proteins are implicated to play a major role in the regulation of numerous genes involved in the function and development of the immune system, cell differentiation, prolifer- ation, apoptosis, etc. It can bind to promoter of its target genes in a sequence-specific manner to transactivate or repress them. In this study, the full-length cDNA of an AP-1 was identified from Venerupis philippinarum (denoted as VpAP-1) by EST analysis and RACE approaches. Phylogenetic analysis indicated that VpAP-1 had higher evolutional conservation to invertebrate than vertebrate counterparts and should be a new member of the AP- 1 protein family. Spatial expression analysis found that VpAP-1 transcript was most abundantly expressed in the hemocytes and hepatopancreas, weakly expressed in the tissues of the gills, mantle and muscle. After Vibrio anguillarum challenge, the expression of VpAP-1 transcript in overall hemocyte population was up-regulated in the first 6 h, and then decreased to 1.5-fold of the control group at 12 h. As time progressed, a second peak of VpAP-1 expression was detected at 24 h post-infection, which was 5-fold compared with that of the control group (P b 0.01). After that, the expression level was sharply decreased and dropped to 0.5-fold of the control at 96 h. The above results indicated that VpAP-1 was perhaps involved in the immune responses against microbe infection and might be contributed to the clearance of bacterial pathogens. © 2014 Published by Elsevier B.V. 1. Introduction The regulation of transcriptional activation or control is an essential and critical point in gene expression, which is mainly governed by a group of regulatory proteins, transcription factors (Moye-Rowley et al., 1988; Sun and Oberley, 1996). They bind to specific DNA sequence in the promoter region of the downstream target genes to regulate gene expression at both transcriptional and post-translational levels (Baeuerle, 1991). As one of the most important transcription factors identified from mammals, activator protein (AP-1) consists of Jun (c-Jun, JunB, JunD), Fos (c-Fos, Fra-1, Fra-2 and Fos B) and Atf (activat- ing transcription factor) sub-families (Karin and Shaulian, 2001). In ad- dition, AP-1 is composed of homo- or heterodimers of Jun-Jun, Jun-Fos, or Jun-Atf and categorized into the class of basic leucine zipper (bZIP) transcription factors (Vesely et al., 2009; Shaulian, 2010; Halazonetis et al., 1988). AP-1 has been demonstrated to be involved in the regulation of host immune responses and plays a major role in the activation of numerous genes involved in the immune responses, cell differentiation, prolifera- tion, apoptosis, etc. (Karin and Shaulian, 2001; Vesely et al., 2009; Angel and Karin, 1991). Moreover, AP-1 can regulate gene expression in response to a variety of stimuli, including cytokines, growth factors, stresses, and bacterial or viral infections. To our knowledge, most stud- ies on AP-1 are focused on mammals and the model organisms, while the molecular features and expression pattern of AP-1 remain poorly in- vestigated in mollusk. Molluscan AP-1 was identified from abalone Haliotis discus discus (De Zoysa et al., 2010) and Hong Kong oyster Crassostrea hongkongensis (Xiang et al., 2014). Manila clam, Venerupis philippinarum, is an important marine bi- valve for commercial fisheries, accounting for about 80% of mudflat fish- ery production in China. However, with the rapid development of intensive culture and environmental deterioration, clams are continu- ously confronted with infectious microorganisms, which have resulted in enormous economic losses to the clam aquaculture (Munroe and McKinley, 2007). Lacking the adaptive immune system, clams are sup- posed to depend solely on the innate immune system to defend invad- ing microorganisms. AP-1 functions as a central transcription factor and link with various immune functional genes. The characterization of such immune regulatory components is helpful to better understand the Gene 557 (2015) 106–111 Abbreviations: AP, transcription factor activator protein‐1; V. philippinarum, Venerupis philippinarum. ⁎ Correspondence to: J. Zhao, Yantai Coastal Zone Research Institute, Chinese Academy of Sciences, 17 Chunhui Road, Laishan District, 264003 Yantai, PR China. ⁎⁎ Correspondence to: C. Wang, 818 Fenghua Rd, Ningbo, 315211 Zhejiang, PR China. E-mail addresses: [email protected] (J. Zhao), [email protected] (C. Wang). 1 These authors contributed equally to this article. http://dx.doi.org/10.1016/j.gene.2014.12.027 0378-1119/© 2014 Published by Elsevier B.V. Contents lists available at ScienceDirect Gene journal homepage: www.elsevier.com/locate/gene
Transcript of Cloning and expression of a transcription factor activator...

Gene 557 (2015) 106–111
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
Short communication
Cloning and expression of a transcription factor activator protein-1(AP-1) member identified from manila clam Venerupis philippinarum
Luning Wu a,b,1, Lei Zhang b,1, Jianmin Zhao b,⁎, Xuanxuan Ning c, Changkao Mu a, Chunlin Wang a,⁎⁎a School of Marine Science of Ningbo University, Zhejiang, Ningbo 315211, PR Chinab Key Laboratory of Coastal Zone Environmental Processes and Ecological Remediation, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai 264003, PR Chinac Yantai Oceanic Environmental Monitoring Central Station, State Oceanic Administration, Yantai 264006, PR China
Abbreviations:AP, transcription factor activator proteiphilippinarum.⁎ Correspondence to: J. Zhao, Yantai Coastal Zone Resea
of Sciences, 17 Chunhui Road, Laishan District, 264003 Ya⁎⁎ Correspondence to: C. Wang, 818 Fenghua Rd, Ningb
E-mail addresses: [email protected] (J. Zhao), wangch1 These authors contributed equally to this article.
http://dx.doi.org/10.1016/j.gene.2014.12.0270378-1119/© 2014 Published by Elsevier B.V.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 September 2014Received in revised form 12 December 2014Accepted 13 December 2014Available online 16 December 2014
Keywords:Transcription factor activator protein-1Venerupis philippinarumImmune responseBacterial challenge
The transcription factor activator protein-1 (AP-1) proteins are implicated to play amajor role in the regulation ofnumerous genes involved in the function and development of the immune system, cell differentiation, prolifer-ation, apoptosis, etc. It can bind to promoter of its target genes in a sequence-specific manner to transactivate orrepress them. In this study, the full-length cDNAof anAP-1was identified from Venerupis philippinarum (denotedas VpAP-1) by EST analysis and RACE approaches. Phylogenetic analysis indicated that VpAP-1 had higherevolutional conservation to invertebrate than vertebrate counterparts and should be a new member of the AP-1 protein family. Spatial expression analysis found that VpAP-1 transcript was most abundantly expressed inthe hemocytes and hepatopancreas, weakly expressed in the tissues of the gills, mantle and muscle. After Vibrioanguillarum challenge, the expression of VpAP-1 transcript in overall hemocyte population was up-regulated inthe first 6 h, and then decreased to 1.5-fold of the control group at 12 h. As time progressed, a second peak ofVpAP-1 expression was detected at 24 h post-infection, which was 5-fold compared with that of the controlgroup (P b 0.01). After that, the expression level was sharply decreased and dropped to 0.5-fold of the controlat 96 h. The above results indicated that VpAP-1was perhaps involved in the immune responses against microbeinfection and might be contributed to the clearance of bacterial pathogens.
© 2014 Published by Elsevier B.V.
1. Introduction
The regulation of transcriptional activation or control is an essentialand critical point in gene expression, which is mainly governed by agroup of regulatory proteins, transcription factors (Moye-Rowleyet al., 1988; Sun andOberley, 1996). They bind to specific DNA sequencein the promoter region of the downstream target genes to regulategene expression at both transcriptional and post-translational levels(Baeuerle, 1991). As one of the most important transcription factorsidentified from mammals, activator protein (AP-1) consists of Jun(c-Jun, JunB, JunD), Fos (c-Fos, Fra-1, Fra-2 and Fos B) and Atf (activat-ing transcription factor) sub-families (Karin and Shaulian, 2001). In ad-dition, AP-1 is composed of homo- or heterodimers of Jun-Jun, Jun-Fos,or Jun-Atf and categorized into the class of basic leucine zipper (bZIP)transcription factors (Vesely et al., 2009; Shaulian, 2010; Halazonetiset al., 1988).
n‐1; V. philippinarum, Venerupis
rch Institute, Chinese Academyntai, PR China.o, 315211 Zhejiang, PR [email protected] (C. Wang).
AP-1 has been demonstrated to be involved in the regulation of hostimmune responses and plays amajor role in the activation of numerousgenes involved in the immune responses, cell differentiation, prolifera-tion, apoptosis, etc. (Karin and Shaulian, 2001; Vesely et al., 2009; Angeland Karin, 1991). Moreover, AP-1 can regulate gene expression inresponse to a variety of stimuli, including cytokines, growth factors,stresses, and bacterial or viral infections. To our knowledge, most stud-ies on AP-1 are focused on mammals and the model organisms, whilethemolecular features and expression pattern of AP-1 remain poorly in-vestigated in mollusk. Molluscan AP-1 was identified from abaloneHaliotis discus discus (De Zoysa et al., 2010) and Hong Kong oysterCrassostrea hongkongensis (Xiang et al., 2014).
Manila clam, Venerupis philippinarum, is an important marine bi-valve for commercialfisheries, accounting for about 80% ofmudflatfish-ery production in China. However, with the rapid development ofintensive culture and environmental deterioration, clams are continu-ously confronted with infectious microorganisms, which have resultedin enormous economic losses to the clam aquaculture (Munroe andMcKinley, 2007). Lacking the adaptive immune system, clams are sup-posed to depend solely on the innate immune system to defend invad-ingmicroorganisms. AP-1 functions as a central transcription factor andlinkwith various immune functional genes. The characterization of suchimmune regulatory components is helpful to better understand the

Table 1PCR primers used in this study.
Primers Sequence (5′–3′) Sequence information
P1 ACTAAACCCAGGGACATA 3′-RACE primerP2 TCACCTCCGCCATCTCCA 3′-RACE primerT7 GTAATACGACTCACTATAGGGC 3′-RACE primerP3 CCTGCTCATCCGTCACAA 5′-RACE primerP4 TTCCGTTTGCTTGAATGA 5′-RACE primerOligo(dG)-adaptor GGCACGCGTCGACTAGTACG10 5′-RACE primerP5 AATCATTACTAAACCCAGGGAC Real time VpAP-1 primerP6 TTGGAGATGGCGGAGGTGA Real time VpAP-1 primerP7 CTCCCTTGAGAAGAGCTACGA Real time actin primerP8 GATACCAGCAGATTCCATACCC Real time actin primerM13-47 CGCCAGGGTTTTCCAGTCACGAC Sequencing primerRV-M GAGCGGATAACAATTTCACACAGG Sequencing primer
Fig. 1. The nucleotide sequence (below) and deduced amino acid sequence (above) ofVpAP-1. The nucleotide and amino acid positions were numbered on the left. The startand stop codon were in bold and the canonical polyadenylation signal sequence (ATTAA)was double-underlined. The shaded amino acids indicated the predicted Jun transcriptionfactor domain. The basic leucine zipper (bZIP) domain profile was underlined.
107L. Wu et al. / Gene 557 (2015) 106–111
innate immune defense mechanisms of mollusk and to give new in-sights to health management and disease control of mollusk aqua-culture. In the present study, the full-length cDNA of AP-1 wascloned from manila clam V. philippinarum, and the spatial and tem-poral expression patterns postVibrio anguillarum challengewere inves-tigated in order to provide clues on the immune regulation functions ofVpAP-1.
2. Materials and methods
2.1. Clams and bacterial challenge
The clams V. philippinarum (shell length: 3.0–4.0 cm) were pur-chased from a local farm, Yantai of Shandong Province and main-tained in aerated tanks at 20–22 °C for ten days before processing.The seawater (salinity 30‰) was changed 100% daily and the clamswere fed with Isochrysis galbana and Platymonas helgolandica duringthe experiment.
V. anguillarum was inoculated in marine broth 2216E at 28 °C toOD600 = 0.6, and then centrifuged at 3000 g for 10 min to harvest thebacteria. The pellet was re-suspended in filtered seawater and adjustedto 107 CFU ml−1, which did not caused mortalities immediately, butinduced significant immune responses in clam V. philippinarum. Forthe bacterial challenge experiment, 300 clams were employed andkept in six aerated tanks (50 individuals in each tank). One tankwith untreated clams served as control, the other five tanks were ex-posed to V. anguillarum with final concentration of 107 CFU ml−1.Five individualswere randomly sampled at 6, 12, 24, 48, 72 and 96 h, re-spectively. 0.5 ml of hemolymphs from each individual (control andinfected groups) was collected using a syringe and centrifuged at1000 g, 4 °C for 10 min to harvest the hemocytes. The hemocyte pel-let from each sample was immediately subjected to RNA extractionseparately.
2.2. RNA isolation and cDNA synthesis
Total RNA was extracted using Trizol reagent according to themanufacturer's protocol (Invitrogen). The cDNA first-strand synthesiswas carried out based on Promega M-MLV RT Usage information(Promega) using total RNA treatedwith DNase Ι (Promega) as template.Single-strand cDNA was synthesized from 1 μg of total RNA in a finalvolume of 20 μl containing 50 pmol of random hexamers, 50 mmol/lTris–HCl, pH 8.3, 75 mmol/l KCl, 3 mmol/l MgCl2, 50 mmol/l DTT,0.75 U of RNasin, 0.2 mmol/l each of dATP, dCTP, dGTP, and dTTP, and200 U of MMLV reverse transcriptase (Promega). Reactions were incu-bated at 37 °C for 1 h, terminated by heating the mixture at 95 °C for5 min, and subsequently stored at −80 °C. cDNA mix was diluted to1:50 for subsequent experiment.
2.3. Cloning the full length cDNA of VpAP-1 gene
A cDNA library was constructed from the hemocytes of manila clamchallenged with V. anguillarum, using the ZAP-cDNA synthesis kitand ZAP-cDNA Gigapack III Gold cloning kit (Stratagene). Random se-quencing of the library using T3 primer yielded 3226 successful se-quencing reactions (Li et al., 2010). BLAST analysis of the ESTsequences revealed that an EST of 498 bp was highly similar to theknown AP-1 sequences. This sequence was then selected for furthercloning of full-length cDNA of AP-1 gene from V. philippinarum (desig-nated VpAP-1).
Based on the above EST sequence, the 3′ and 5′ends of VpAP-1 cDNAwere obtained by RACE approaches. Four specific primers, senseprimers P1 and P2, and reverse primers P3 and P4 (Table 1) were de-signed based on the EST sequence, the nested PCR strategy was appliedto clone the 3′ end of VpAP-1 using sense primers P1 and P2 and reverseprimer T7, while sense primer oligo (dG)-adaptor and reverse primers
P3 and P4 were used to get the 5′ end of VpAP-1. The PCR temperatureprofile was 94 °C for 5 min followed by 32 cycles of 94 °C for 30 s, 60 °Cfor 30 s, 72 °C for 1 min, and the final extension step at 72 °C for

108 L. Wu et al. / Gene 557 (2015) 106–111
10 min. The PCR products were cloned into the pMD18-T simple vec-tor (TaKaRa) and sequenced bi-directionally with primers M13–47 andRV-M (Table 1). The sequencing results were verified and subjected tocluster analysis.
V.philippinarum - - - - - - - M E T T M Y N D D G A G R E N - N T F G SH.discus - - - - - - - M E T N F Y E T D D A A R E M R Q A M K RR.norvegicus - - - M T A K M E T T F Y D D A L N A S F L Q S E S G AM.musculus - - - M T A K M E T T F Y D D A L N A S F L Q S E S G AB.taurus - - - M T A K M E T T F Y D D A L N A S F L Q S E S G AC.jacchus - - - M T A K M E T T F Y D D A L N A S F L Q S E S G AH.sapiens - - - M T A K M E T T F Y D D A L N A S F L P S E S G PG.gallus - - - - - - - M E P T F Y E D A L N A S F A P P E S G GS.salar - - - M S R K M E T T F Y D D S L N - S F S Q H E K P DO.mykiss - - - M S T K M E T T F Y D D S H N - A F S Q H Q N A GD.rerio - - - M S T K M E T T F Y D D S L N S A F S Q H D S A TS.kowalevskii - - - - M M T M E A Q L Y D D Q P N P W L D K - - - - -H.saltator - M V R S L A M E Q T F Y E D - A N A Y G A M N - R E NC.floridanus - M V R S L T M E P T F Y E N - A S V Y G T V N G R E NI.scapularis - - - - - - - - - - - - - - - - - - - - - - - - - - - -
V.philippinarum - G I S S A K R Q K I Q T A - - - - - - - - - - - L L QH.discus - D F N S E G S K K S K F S - - - - - - - - - - - L L QR.norvegicus - G N L K P H L R A K N S D - - - - - - - - - - - L L TM.musculus - G S L K P H L R A K N S D - - - - - - - - - - - L L TB.taurus - G S L K P H L R A K N S D - - - - - - - - - - - L L TC.jacchus - G S L K P H L R A K N S D - - - - - - - - - - - L L TH.sapiens - G S L K P H L R A K N S D - - - - - - - - - - - L L TG.gallus - S S L K P H L R N K N A D - - - - - - - - - - - I L TS.salar - S T L K P H L R A K A S D - - - - - - - - - - - I L TO.mykiss - I T L K P H L Q A K A S D - - - - - - - - - - - I L TD.rerio - G N L K P H L R A K A S D - - - - - - - - - - - I L TS.kowalevskii - - - - - - - - T K K S Y A - - - - - - - - - - - V L SH.saltator - Q R Q G P Q A K R P R L G P L P P T L N S V T P I L AC.floridanus - Q R Q G P Q V K R P R L G P L P P A L N N V T P I L SI.scapularis - G R - - P E R K A A R F S - - - - - - - - - - S L L T
V.philippinarum - T Q I L F P K F V T D E Q E A Y A Q G F V A A L A E LH.discus - P Q F I F P K F V T E E Q E A Y A R G F V D A L A E LR.norvegicus - T Q F L C P K N V T D E Q E G F A E G F V R A L A E LM.musculus - T Q F L C P K N V T D E Q E G F A E G F V R A L A E LB.taurus - T Q F L C P K N V T D E Q E G F A E G F V R A L A E LC.jacchus - T Q F L C P K N V T D E Q E G F A E G F V R A L A E LH.sapiens - T Q F L C P K N V T D E Q E G F A E G F V R A L A E LG.gallus - T Q F L C P K N V T D E Q E G F A E G F V R A L A E LS.salar - T Q F L C P R N V T D E Q E G F A E G F V R A L A E LO.mykiss - T Q F L C P R N V T D E Q E G F A E G F V R A L A E LD.rerio - T Q F L C P K N V T D E Q E G F A E G F V R A L A E L
S.kowalevskii - T S I L C P K N V T L E Q E Q F A E G F V Q A L S Q LH.saltator - T Q I L F P K T V T E A Q E L Y A R G F V D A L N E LC.floridanus - T Q I L F P K T V T E D Q E L Y A R G F I D A L N E LI.scapularis - T Q Y L F T K T A T E E Q E Q Y A R G F V D A L A Q L
V.philippinarum - - - - P N S S S I S I N D A L K N I - - - F T T T T SH.discus - L F A P N S N S V S S F N S V P T T N Q G L T T T T SR.norvegicus - M V A P A V A S V A G A G G G G G Y S A S L H S E P PM.musculus - M V A P A V A S V A G A G G G G G Y S A S L H S E P PB.taurus - - - - - - - - - - - - - - - - - - - - - - - - - - - PC.jacchus - M V A P A V A S V A G G S G S G G F S A S L H S E P PH.sapiens - M V A P A V A S V S G G S G S G G F S A S L H S E P PG.gallus - - - - - - M A P V S S M A G G G S F N T S L H S E P PS.salar - T I P P P V S A L T V S S V Y N N N - I T M R S E S PO.mykiss - T T L P P V C S V A G S T V Y N N - - V A M R S E S PD.rerio - S S M A P V S S I A G G A V Y S S - - - A M R A D P PS.kowalevskii - G T L H Y P S Q A S V T T T T T S L P G S A S V D Q PH.saltator - - - - Q E P G S I H G - - - - - - - - - - - A T Y T TC.floridanus - - - - Q E P G S V H G - - - - - - - - - - - A T Y T TI.scapularis - - - - P S E T T L S A - - - - - - - - - - - A E A S D
V.philippinarum - - - - - - - - - - - - - - - - - - - - - G T Y P M V TH.discus - - - - - - - - - - - - - - - - - - - - - N D I L L P RR.norvegicus - Q Q Q Q Q - P P Q P P H H L P Q Q I P V Q H P R L Q AM.musculus - Q Q Q Q Q - P P Q P P H H L P Q Q I P V Q H P R L Q AB.taurus - Q Q Q Q Q Q P P Q P P H H L P Q Q I P V Q H P R L Q AC.jacchus - Q Q Q Q Q - P P Q P P H H L P Q Q M P V Q H P R L Q AH.sapiens - Q Q Q Q Q - - - - P P H H L P Q Q M P V Q H P R L Q AG.gallus - - - - - - - - - - P Q H H I N P Q M P V Q H P R L Q AS.salar - - - - - - - - - - P Q L P V Y G Q Q S H P H P R L T AO.mykiss - - - - - - - - - - P Q L P I Y G Q Q S H P H P R L T AD.rerio - - - - - - - - - - P Q L P V Q - - - - - - H P R L Q AS.kowalevskii - - - - - - - - - - G T H D Y I G T I A T V G V Q R I KH.saltator - - - - - - - - - - - - - - - - - - - - - - - G G L L HC.floridanus - - - - - - - - - - - - - - - - - - - - - - - P N L V HI.scapularis - - - - - - - - - - - - - - - - - - - - - L G G G D G H
IV.philippinarum - E R K R A R N R V A A R K C R T R K L E R I A R L E DH.discus - E R K R A R N R V A A R K C R T R K L E R I S R L E DR.norvegicus - E R K R M R N R I A A S K C R K R K L E R I A R L E EM.musculus - E R K R M R N R I A A S K C R K R K L E R I A R L E EB.taurus - E R K R M R N R I A A S K C R K R K L E R I A R L E EC.jacchus - E R K R M R N R I A A S K C R K R K L E R I A R L E EH.sapiens - E R K R M R N R I A A S K C R K R K L E R I A R L E EG.gallus - E R K R M R N R I A A S K C R K R K L E R I A R L E ES.salar - E R K R M R N R V A A S K C R K R K L E R I S R L E D
O.mykiss - E R K R M R N R V A A S K C R K R K L E R I S R L E DD.rerio - E R K R M R N R I A A S K C R K R K L E R I S R L E DS.kowalevskii - E R K R L R N R I A A S K C R K R K L E R I S R L E EH.saltator - E R K R Q R N R V A A S K C R R R K L E R I S R L E DC.floridanus - E R K R Q R N R V A A S K C R R R K L E R I S R L E DI.scapularis - E R K R L R N R I A A S K C R K R K L E R I S R L E D
IV.philippinarum - V N S G C Q I M M S Q N - - I N F -H.discus - V N S G C Q I M M T Q N - - I S F -R.norvegicus - V N S G C Q L M L T Q Q L Q T F - -M.musculus - V N S G C Q L M L T Q Q L Q T F - -B.taurus - V N S G C Q L M L T Q Q L Q T F - -C.jacchus - V N S G C Q L M L T Q Q L Q T F - -H.sapiens - V N S G C Q L M L T Q Q L Q T F - -G.gallus - V N S G C Q L M L T Q Q L Q T F - -S.salar - V N S G C Q L M L T Q Q L Q T F - -O.mykiss - V N S G C Q L M L T Q Q L Q T F - -D.rerio - V N S G C Q L M L T Q Q L Q T F - -S.kowalevskii - V K S G C Q V M L A Q Q L A F - - -H.saltator - V H S G C A I M I N E H G S Q Q F -C.floridanus - V N A G C H I V P - - - - - G Q F -I.scapularis - V K Q G C Q I M M S T A Y G A T T AI
Fig. 2.Multiple alignments of theVpAP-1with other registered counterparts fromHaliotis discusmusculus (NP_034721), Bos taurus (NP_001071295), Callithrix jacchus (XP_002750926),Homo smykiss (NP_001117883),Danio rerio (NP_956281), Saccoglossus kowalevskii (NP_001161502),Cregions indicated identical amino acids among the different sequences, while the gray shaded re(*). Gaps were indicated by dashes to improve the alignment.
2.4. Sequence analysis of VpAP-1
The VpAP-1 cDNA sequencewas analyzed by the BLAST algorithm atthe National Center for Biotechnology Information (http://www.ncbi.
K L N - - - - S M K R A M T - - - - - - - - - - - - L D F N S G N - 37K I N - - - - S S L G L F S N S G S G D R D A V N R L K Q N M K L - 50Y G Y S N P K I L K Q S M T - - - - - - - - - - - - L N L A D P V - 46Y G Y S N P K I L K Q S M T - - - - - - - - - - - - L N L A D P V - 46Y G Y S N P K I L K Q S M T - - - - - - - - - - - - L N L A D P V - 46Y G Y S N P K I L K Q S M T - - - - - - - - - - - - L N L A D P V - 46Y G Y S N P K I L K Q S M T - - - - - - - - - - - - L N L A D P V - 46Y G Y N N A K V L K Q S M T - - - - - - - - - - - - L N L S D A A - 42Y G Y - N P K A L K H S M T - - - - - - - - - - - - L N L A D P T - 44Y G Y - N P K A L K H S M T - - - - - - - - - - - - L N L A D P T - 44F G Y - N H K A L K H N M T - - - - - - - - - - - - L N L S D P A - 45- - - - - - - - - K K S M K - - - - - - - - - - - - L D L C T Q P - 31N V G - - - - Q L K R N L T - - - - - - - - - - - - L D L N - G C - 41S M G - - - - Q L K R N L T - - - - - - - - - - - - L D L N - G C - 42- - - - - - - - - - - - M T - - - - - - - - - - - - L D L N - S A - 8
S P D L N M L K L A S P E L E K M I I Q A N - G M V T T T P - T P - 84S P D L N M L K L A S P E L E K M I I Q A N - G M V T T T P - P T - 97S P D V G L L K L A S P E L E R L I I Q S S N G H I T T T P - T P - 94S P D V G L L K L A S P E L E R L I I Q S S N G H I T T T P - T P - 94S P D V G L L K L A S P E L E R L I I Q S S N G H I T T T P - T P - 94S P D V G L L K L A S P E L E R L I I Q S S N G H I T T T P - T P - 94S P D V G L L K L A S P E L E R L I I Q S S N G H I T T T P - T P - 94S P D V G L L K L A S P E L E R L I I Q S S N G L I T T T P - T P - 90S P D V Q L L K L A S P E L E R L I I Q S S N G L I T T T P - T P - 92S P D V Q L L K L A S P E L E R L I I Q S S N G L I T T T P - T P - 92S P D V G L L K L A S P E L E R L I I Q S S N G M I T T T P - T P - 93T P D V Q M L K L A S P E L E K L I M S Q - Q G T V I T T P - T P - 71S P D L N M L K L G S P E L E K L I M A Q Q D N L V T S L P T P T - 101S P D L N M L K L A S P E L E K F I M T Q S D S L V T G L P T P T - 102S P D L N M L Q L A S P E L E R L I I A H N - G L V T T T P T P T - 55
H S K P E E E C V D I - - - - - - - - - - - - - - - - - - - - - - - 122H D P N Q T E N I P I T I Q G K G V V S S Q S L P Q T S K G C V A - 157H S Q N T L P S - - - - - - - - - - - - - V T S A A Q P V S G A G - 141H S Q N T L P S - - - - - - - - - - - - - V T S A A Q P V S G A G - 141H S Q N T L P S - - - - - - - - - - - - - V T S A A Q - - - - - - - 135H S Q N T H P S - - - - - - - - - - - - - V T S A A Q P V S G A G - 141H S Q N T L P S - - - - - - - - - - - - - V T S A A Q P V N G A G - 141H N Q N T L P S - - - - - - - - - - - - - V T S A A Q P V S G G - - 136H H Q H V L P N A P G G - - - - - - - - P V T S A G Q T S I N N P - 144H H Q H V L P N A P G V - - - - - - - - P V T S T G Q T R I N N S - 144H H Q H M P N - - - - - - - - - - - - - - V T S A P Q T T I N - - - 137
H G G - - V P S - - - - - - - - - - - - - S T S A T T T L P S S S - 116H H S D S S - - - - - - - - - - - - - - - - - - - - - - - - - - - - 134H H S D S S - - - - - - - - - - - - - - - - - - - - - - - - - - - - 135H Q T A G G - - - - - - - - - - - - - - - - - - - - - - - - - - - - 88
L P G G I V P S S S L P S K S L L N P - - - - - - - - - - - - - - - 162L P G G L T V S S A S R S L K S V S N - - - - - - - - - - - - - - - 203V Y A N L S N F N P G A L S S G G G A P S Y G A T G L A F P S Q P - 201V Y A N L S N F N P G A L S S G G G A P S Y G A A G L A F P S Q P - 201V Y A N L S N F N P G S L S S G G G A P S Y G A A G L A F P A Q P - 169V Y A N L S N F N P G A L S S G G G A P S Y G A A G L A F P A Q P - 201V Y A N L S N F N P G A L S S G G G A P S Y G A A G L A F P A Q P - 201V Y A N L S N F N P N A L N S - - - A P N Y N A N G M G Y A - - - - 185V Y E D L N T F T P A I T T N S A L G Y T T S A P A M S F P S A P - 203V Y E N L N T F T P A V T T N S A P G Y T T S V P T M S F P S A P - 202V Y A D L N T F N P A I S S S S A N - - - - - P A A M S F P S A P - 189M Q P V S S T A S T C V T S P R R N N - - - - - - - - - - - - - - - 162L E P P G S V Q T S T D S S T M S Q - - - - - - - - - - - - - - - - 165L E P P G S V Q S T - - E S T M S N - - - - - - - - - - - - - - - - 164S G A S T S S D S F I L P S S S E H S - - - - - - - - - - - - - - - 120
V K E E P Q - T V P C G L N - - S P P P S P I D M A N Q E V I K L - 199L K E E P Q T T V P C I E N - - T P P L S P I N M E N Q E V I K L - 241L K E E P Q - T V P E M P G - E T P P L S P I D M E S Q E R I K A - 258L K E E P Q - T V P E M P G - E T P P L S P I D M E S Q E R I K A - 258L K E E P Q - T V P E M P G - E T P P L S P I D M E S Q E R I K A - 227L K E E P Q - T V P E M P G - E T P P L S P I D M E S Q E R I K A - 258L K E E P Q - T V P E M P G - E T P P L S P I D M E S Q E R I K A - 255L K E E P Q - T V P E M P G - E T P P L S P I D M E S Q E R I K A - 234L K E E P Q - T V P E M P G - E T P P L S P I D M E N Q E R I K A - 252L K E E P Q - T V P E M L G - E T P P L S P I D M E N Q E R I K A - 251L K E E P Q - T V P E M P G - E T P P L S P I D M E S Q E R I K A - 232Q E E A P Q - T V P V F L A K G T P P T S P I D M E M Q E R F K A - 212V K D E P Q - T V P S V S S - - S P P M S P I D M E N Q E K I K L - 200V K D E P Q - T V P S V S S - - T P P M S P I D M E N Q E K I K L - 199V K D E P Q - T V P N L G A - - T P P L S P I D M R D Q E R I K L - 157
*K V A D L K G Q N N D L S S Q A S K L R D E V C K L K Q T I I E H - 259R V K D L K G Q N A D L V T T A T T L R D Q V C K L K R Q I I D H - 301K V K T L K A Q N S E L A S T A N M L R E Q V A Q L K Q K V M H - 318K V K T L K A Q N S E L A S T A N M L R E Q V A Q L K Q K V M N H - 318K V K T L K A Q N S E L A S T A N M L R E Q V A Q L K Q K V M N H - 287K V K T L K A Q N S E L A S T A N M L R E Q V A Q L K Q K V M N H - 318K V K T L K A Q N S E L A S T A N M L R E Q V A Q L K Q K V M N H - 315K V K T L K A Q N S E L A S T A N M L R E Q V A Q L K Q K V M N H - 294K V K N L K T Q N S D L A S T A N M L R E Q V A Q L K Q K V M N H - 312
K V K N L K T Q N S D L A S T A N M L R E Q V A Q L K Q K V M N H - 311K V K T L K S Q N S E L A S T A N M L R E Q V A Q L K Q K V M N H - 292K V D D L K N Q N S D L S L S A T Q L R E Q V C K L K Q Q V M E H - 272K V K V L K G E N H E L S A V V V R L K E H V C R L K E Q V M D H - 260K V K L L K G E N T E L S G I V H K L K E H V C R L K E Q V M D H - 259K V H A L K T E N S E L G S V V S S L R D Q V C R L K Q E V M M H - 217
- 274- 316- 334- 334- 303- 334- 331- 310- 328- 327- 308- 287- 277- 271- 235
discus (ACJ65689),Harpegnathos saltator (EFN76477),Rattus norvegicus (NP_068607),Musapiens (CAG46525),Gallus gallus (NP_001026460), Salmo salar (ACN11435),Oncorhynchusamponotus floridanus (EFN66194) and Ixodes scapularis (XP_002404571). The black shadedgions represented conservative replacements. The bZIP domainwas indicated by asterisks

109L. Wu et al. / Gene 557 (2015) 106–111
nlm.nih.gov/blast) and the deduced amino acid sequence was analyzedwith the Expert Protein Analysis System (http://www.expasy.org/).Characteristic domains or motifs were identified using the PROSITEprofile database (Bairoch et al., 1997). Identity, similarity and gap per-centages were calculated using FASTA program (Pearson, 1990). TheClustalW program (http://www.ebi.ac.uk/clustalw/) was used for mul-tiple alignment of VpAP-1, and the Signal P 3.0 software (http://www.cbs.dtu.dk/services/SignalP/) was employed for the prediction of signalpeptide (Thompson et al., 1994). An unrooted phylogenetic tree wasconstructed according to amino acid sequences of the selected AP-1 byusing the neighbor-joining (NJ) algorithm embedded in Mega 3.1 pro-gram (http://www.megasoftware.net/). The bootstrap trials were repli-cated 1000 times to derive the confidence value for the phylogenyanalysis.
2.5. Tissue distribution and temporal expression profile of VpAP-1 transcriptpost Vibrio challenge
Total RNA was extracted from various tissues of five randomlyhealthy clams, including the muscle, gill, hemocytes, hepatopancreasand mantle. RNA isolation and cDNA synthesis were carried out as de-scribed above. cDNA mix was diluted to 1:100 and stored at −80 °Cfor subsequent SYBR Green assay.
The tissue distribution and temporal expression of VpAP-1 transcriptin the hemocytes of clams challenged with V. anguillarum were deter-mined by quantitative real-time PCR (Q-PCR) as previously described(Zhang et al., 2011; Zhao et al., 2007). Two VpAP-1 gene-specificprimers P5 and P6 (Table 1) were used to amplify a product of 96 bpfrom cDNA, and the PCR product was sequenced to verify the specificityof RT-PCR. Twoβ-actin primers, P7 and P8 (Table 1)were used to ampli-fy a 121 bp fragment as internal control. The comparative Ct methodwas used to analyze the relative expression level of VpAP-1. The Ct forthe target amplified products of VpAP-1 and internal control β-actinwere determined for each sample. Difference in the Ct for the targetand the internal control, called ΔCt, was calculated to normalize thedifferences in the amount of template and the efficiency of the RT-PCR. TheΔCt for each samplewas subtracted from theΔCt of the calibra-tor; the difference was called ΔΔCt. The expression level of VpAP-1 wascalculated by 2−ΔΔCt, and the value stood for an n-fold differencerelative to the calibrator (Livak and Schmittgen, 2001). For tissuedistribution, VpAP-1 transcript levels in the hemocyte, mantle, gill andhepatopancreas were normalized to that of muscle. As concerned totemporal expression of VpAP-1, the hemocytes from the clams in con-trol group were used as the reference sample.
Fig. 3. A phylogenetic tree constructed by the neighbor-joining method. The common names a(AHF51977), Haliotis discus discus (ADQ43242), Zootermopsis nevadensis (KDR21486), CrassoDanio rerio (NP_956281), Xenopus (Silurana) tropicalis (XP_002931634) and Pelodiscus sinensisbranch in the phylogenetic tree nodes from 1000 replicates.
Real-time PCR amplificationwas carried out in anApplied Biosystem7500 fast real-time PCR system. Dissociation curve analysis of ampli-fication was performed at the end of each PCR reaction to confirmthat only one PCR product was amplified and detected. After thePCR program, data were analyzed using SDS software V2.01 (AppliedBiosystems). To maintain consistency, the baseline was set automat-ically by the software. All data were given in terms of relative mRNAexpressed as mean ± S.E. (N = 5). One-way analysis of variance(one-way ANOVA)was performed on all data using SPSS 13.0 statisticalsoftware, and P values less than 0.05 were considered statisticallysignificant.
3. Results and discussion
3.1. cDNA cloning and structural analysis of VpAP-1
The full sequence of VpAP-1 cDNA consisted of 1692 bp (GenBankaccession no. HQ918289), encoding a polypeptide of 274 amino acidswith the predicted molecular mass of 30.12 kDa (Fig. 1). No signal pep-tidewas identified from the deduced amino acid of VpAP-1, which indi-cated that VpAP-1 was a non-secreted protein as the other AP-1s. Junproteins (components of AP-1) were found to contain an N-terminaltranscriptional activation domain, a basic domain for DNA binding,and a C-terminal leucine zipper domain for dimerization (Sun andOberley, 1996). SMART analysis revealed that the characteristic Juntranscription factor domain (1–185 aa) and the basic leucine zipper(bZIP) region (194–258 aa) existed in the VpAP-1 amino acid se-quence. In addition, the intercrine alpha family or small cytokine CXC(SCY) motif (170–217 aa), interleukin 4 and 13 domains (123–244 aa)and helix–loop–helix (HLH) domain (205–258 aa) were also identifiedin the deduced amino acid sequence of VpAP-1. All these conservedcharacteristics indicated that VpAP-1 should be a new member ofAP-1 family.
ClustalW analysis revealed that the deduced amino acid sequence ofVpAP-1 shared high identity with previously identified AP-1s (Fig. 2).For example, VpAP-1 shared 75% identity to H. discus discus AP-1(ACJ65689), 63% identity to Salmo salar AP-1 (ACN11435), 62% identityto Homo sapiens AP-1 (CAG46525), 45% identity to Saccoglossuskowalevskii AP-1 (NP_001161502) and 43% identity to Oncorhynchusmykiss AP-1 (NP_001117883). Multiple alignments clearly showedthat the C-terminus of VpAP-1 was highly conserved, while the N-terminal domain was considerably variable (Fig. 2). Although the Jundomain was not well conserved at the N-terminus, the bZIP domainstill exhibited relatively high similarity among all the sequences.
Charadrius vociferus
Pelodiscus sinensis
Homo sapiens
Xenopus (Silurana) tropicalis
Salmo salar
Danio rerio
Zootermopsis nevadensis
Ruditapes philippinarum
Haliotis discus discus
Crassostrea hongkongensis
Crassostrea gigas
vertebrate
invertebrate
nd the GenBank accession numbers were the same as follows: Crassostrea hongkongensisstrea gigas (EKC41210), Homo sapiens (NP_002219), Charadrius vociferous (KGL92897),(XP_006133565). Numbers next to the branches indicated bootstrap value of each internal
3500
4000
4500c
500
1000
1500
2000
2500
3000
3500
AP
-1 m
RN
A
b b b
4
6
8
re
ssio
n le
ve
l o
f A
0
2
4
Exp
r
MU GI HE HP MA
a
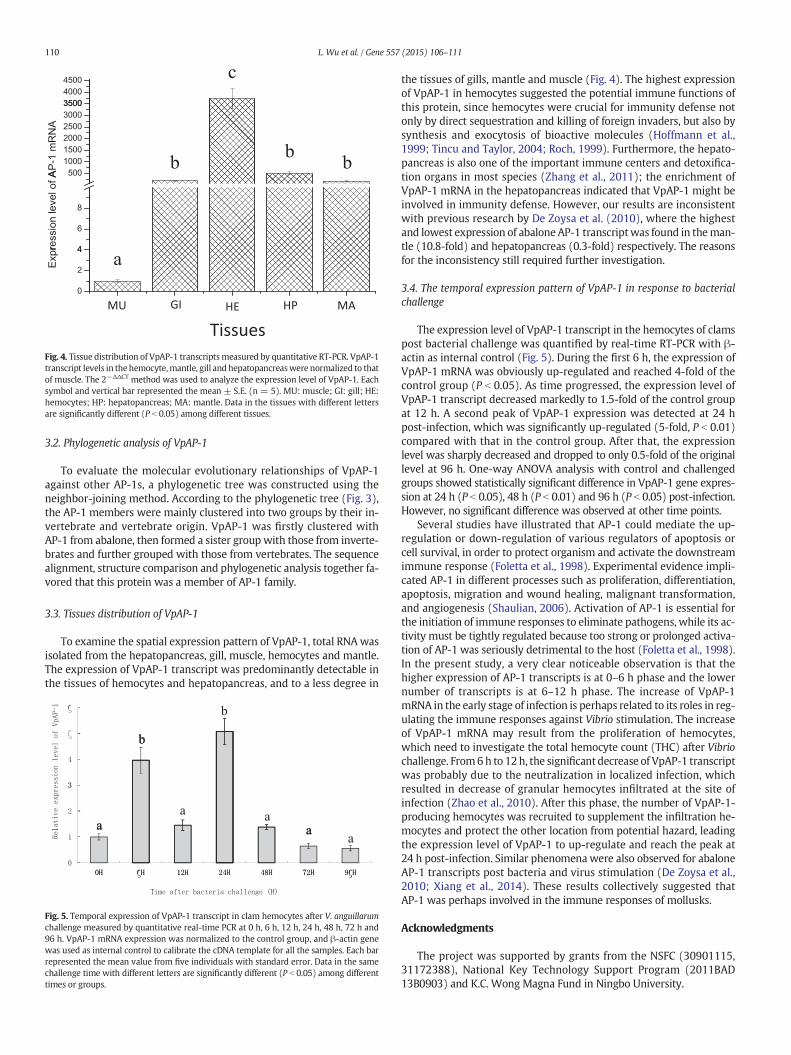
TissuesFig. 4. Tissue distribution of VpAP-1 transcriptsmeasured by quantitative RT-PCR. VpAP-1transcript levels in thehemocyte,mantle, gill and hepatopancreaswere normalized to thatof muscle. The 2−ΔΔCT method was used to analyze the expression level of VpAP-1. Eachsymbol and vertical bar represented the mean ± S.E. (n = 5). MU: muscle; GI: gill; HE:hemocytes; HP: hepatopancreas; MA: mantle. Data in the tissues with different lettersare significantly different (P b 0.05) among different tissues.
110 L. Wu et al. / Gene 557 (2015) 106–111
3.2. Phylogenetic analysis of VpAP-1
To evaluate the molecular evolutionary relationships of VpAP-1against other AP-1s, a phylogenetic tree was constructed using theneighbor-joining method. According to the phylogenetic tree (Fig. 3),the AP-1 members were mainly clustered into two groups by their in-vertebrate and vertebrate origin. VpAP-1 was firstly clustered withAP-1 from abalone, then formed a sister group with those from inverte-brates and further grouped with those from vertebrates. The sequencealignment, structure comparison and phylogenetic analysis together fa-vored that this protein was a member of AP-1 family.
3.3. Tissues distribution of VpAP-1
To examine the spatial expression pattern of VpAP-1, total RNAwasisolated from the hepatopancreas, gill, muscle, hemocytes and mantle.The expression of VpAP-1 transcript was predominantly detectable inthe tissues of hemocytes and hepatopancreas, and to a less degree in
b
b
b
aa a
aa a a
Fig. 5. Temporal expression of VpAP-1 transcript in clam hemocytes after V. anguillarumchallenge measured by quantitative real-time PCR at 0 h, 6 h, 12 h, 24 h, 48 h, 72 h and96 h. VpAP-1 mRNA expression was normalized to the control group, and β-actin genewas used as internal control to calibrate the cDNA template for all the samples. Each barrepresented the mean value from five individuals with standard error. Data in the samechallenge time with different letters are significantly different (P b 0.05) among differenttimes or groups.
the tissues of gills, mantle and muscle (Fig. 4). The highest expressionof VpAP-1 in hemocytes suggested the potential immune functions ofthis protein, since hemocytes were crucial for immunity defense notonly by direct sequestration and killing of foreign invaders, but also bysynthesis and exocytosis of bioactive molecules (Hoffmann et al.,1999; Tincu and Taylor, 2004; Roch, 1999). Furthermore, the hepato-pancreas is also one of the important immune centers and detoxifica-tion organs in most species (Zhang et al., 2011); the enrichment ofVpAP-1 mRNA in the hepatopancreas indicated that VpAP-1 might beinvolved in immunity defense. However, our results are inconsistentwith previous research by De Zoysa et al. (2010), where the highestand lowest expression of abaloneAP-1 transcriptwas found in theman-tle (10.8-fold) and hepatopancreas (0.3-fold) respectively. The reasonsfor the inconsistency still required further investigation.
3.4. The temporal expression pattern of VpAP-1 in response to bacterialchallenge
The expression level of VpAP-1 transcript in the hemocytes of clamspost bacterial challenge was quantified by real-time RT-PCR with β-actin as internal control (Fig. 5). During the first 6 h, the expression ofVpAP-1 mRNA was obviously up-regulated and reached 4-fold of thecontrol group (P b 0.05). As time progressed, the expression level ofVpAP-1 transcript decreased markedly to 1.5-fold of the control groupat 12 h. A second peak of VpAP-1 expression was detected at 24 hpost-infection, which was significantly up-regulated (5-fold, P b 0.01)compared with that in the control group. After that, the expressionlevel was sharply decreased and dropped to only 0.5-fold of the originallevel at 96 h. One-way ANOVA analysis with control and challengedgroups showed statistically significant difference in VpAP-1 gene expres-sion at 24 h (P b 0.05), 48 h (P b 0.01) and 96 h (P b 0.05) post-infection.However, no significant difference was observed at other time points.
Several studies have illustrated that AP-1 could mediate the up-regulation or down-regulation of various regulators of apoptosis orcell survival, in order to protect organism and activate the downstreamimmune response (Foletta et al., 1998). Experimental evidence impli-cated AP-1 in different processes such as proliferation, differentiation,apoptosis, migration and wound healing, malignant transformation,and angiogenesis (Shaulian, 2006). Activation of AP-1 is essential forthe initiation of immune responses to eliminate pathogens, while its ac-tivity must be tightly regulated because too strong or prolonged activa-tion of AP-1 was seriously detrimental to the host (Foletta et al., 1998).In the present study, a very clear noticeable observation is that thehigher expression of AP-1 transcripts is at 0–6 h phase and the lowernumber of transcripts is at 6–12 h phase. The increase of VpAP-1mRNA in the early stage of infection is perhaps related to its roles in reg-ulating the immune responses against Vibrio stimulation. The increaseof VpAP-1 mRNA may result from the proliferation of hemocytes,which need to investigate the total hemocyte count (THC) after Vibriochallenge. From6h to 12h, the significant decrease of VpAP-1 transcriptwas probably due to the neutralization in localized infection, whichresulted in decrease of granular hemocytes infiltrated at the site ofinfection (Zhao et al., 2010). After this phase, the number of VpAP-1-producing hemocytes was recruited to supplement the infiltration he-mocytes and protect the other location from potential hazard, leadingthe expression level of VpAP-1 to up-regulate and reach the peak at24 h post-infection. Similar phenomenawere also observed for abaloneAP-1 transcripts post bacteria and virus stimulation (De Zoysa et al.,2010; Xiang et al., 2014). These results collectively suggested thatAP-1 was perhaps involved in the immune responses of mollusks.
Acknowledgments
The project was supported by grants from the NSFC (30901115,31172388), National Key Technology Support Program (2011BAD13B0903) and K.C. Wong Magna Fund in Ningbo University.

111L. Wu et al. / Gene 557 (2015) 106–111
References
Angel, P., Karin, M., 1991. The role of Jun, Fos and the AP-1 complex in cell-proliferationand transformation. Biochim. Biophys. Acta 1072, 129.
Baeuerle, P.A., 1991. The inducible transcription activator NF-kappa-B-regulation by dis-tinct protein subunits. Biochim. Biophys. Acta 1072, 63–80.
Bairoch, A., Bucher, P., Hofman, K., 1997. The PROSITE database, its status in 1997. NucleicAcids Res. 25, 217.
De Zoysa, M., Nikapitiya, C., Lee, Y., Lee, S., Oh, C., Whang, I., Yeo, S.Y., Choi, C.Y., Lee, J.,2010. First molluscan transcription factor activator protein-1 (Ap-1) member fromdisk abalone and its expression profiling against immune challenge and tissue injury.Fish Shellfish Immunol. 29, 1028–1036.
Foletta, V.C., Segal, D.H., Cohen, D.R., 1998. Transcriptional regulation in the immune sys-tem: all roads lead to AP-1. J. Leukoc. Biol. 63, 139–152.
Halazonetis, T.D., Georgopoulos, K., Greenberg, M.E., Leder, P., 1988. c-Jun dimerizes withitself and with c-Fos, forming complexes of different DNA binding affinities. Cell 55,917–924.
Hoffmann, J.A., Kafatos, F.C., Janeway, C.A., Ezekowitz, R.A., 1999. Phylogenetic perspec-tives in innate immunity. Science 284, 1313–1318.
Karin, M., Shaulian, E., 2001. AP-1: linking hydrogen peroxide and oxidative stress to thecontrol of cell proliferation and death. IUBMB Life 52, 17–24.
Li, C., Qiu, L., Ning, X., Chen, A., Qin, S., Wu, H., Zhao, J., 2010. The first molluscan TCTP inVenerupis philippinarum: molecular cloning and expression analysis. Fish ShellfishImmunol. 29, 530–533.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−delta delta c(T)) method. Methods 25, 402–408.
Moye-Rowley, W., Harshman, K., Parker, C., 1988. YAP1 Encodes a Yeast Homolog ofMammalian Transcription Factor AP-1. Cold Spring Harbor Laboratory Press, p. 711.
Munroe, D., McKinley, R.S., 2007. Commercial Manila clam (Tapes philippinarum) culturein British Columbia, Canada: the effects of predator netting on intertidal sedimentcharacteristics. Estuar. Coast. Shelf Sci. 72, 319–328.
Pearson, W.R., 1990. Rapid and sensitive sequence comparison with FASTP and FASTA.Method Enzymol. 183, 63–98.
Roch, P., 1999. Defense mechanisms and disease prevention in farmed marine inverte-brates. Aquaculture 172, 125–145.
Shaulian, E., 2006. Transcription factors: AP-1. In: Shapiro, G.J.L.D. (Ed.), Encyclopedia ofRespiratory Medicine. Academic Press, Oxford, pp. 251–257.
Shaulian, E., 2010. AP-1—the Jun proteins: oncogenes or tumor suppressors in disguise?Cell. Signal. 22, 894–899.
Sun, Y., Oberley, L.W., 1996. Redox regulation of transcriptional activators. Free Radic. Biol.Med. 21, 335–348.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving the sensitivity ofprogressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acid Res. 22, 4673.
Tincu, J.A., Taylor, S.W., 2004. Antimicrobial peptides from marine invertebrates.Antimicrob. Agents Chemother. 48, 3645–3654.
Vesely, P.W., Staber, P.B., Hoefler, G., Kenner, L., 2009. Translational regulation mecha-nisms of AP-1 proteins. Mutat. Res. Rev. Mutat. 682, 7–12.
Xiang, Z.M., Qu, F., Li, J., Qi, L., Yang, Z., Kong, X., Xiang, Z.M., Qu, F., Li, J., Qi, L., Yang, Z.,Kong, X., Yu, Z., 2014. Activator protein-1 (AP-1) and response to pathogen infectionin the Hong Kong oyster (Crassostrea hongkongensis). Fish Shellfish Immunol. 36,83–89.
Zhang, L., Zhao, J., Li, C., Su, X., Chen, A., Li, T., Qin, S., 2011. Cloning and characterization ofallograft inflammatory factor-1 (AIF-1) from manila clam Venerupis philippinarum.Fish Shellfish Immunol. 30, 148–153.
Zhao, J., Song, L., Li, C., Ni, D., Wu, L., 2007. Molecular cloning, expression of a big defensingene from bay scallop Argopecten irradians and the antimicrobial activity of its recom-binant protein. Mol. Immunol. 44, 360–368.
Zhao, J., Qiu, L., Ning, X., Chen, A., Wu, H., Li, C., 2010. Cloning and characterization of aninvertebrate type lysozyme from Venerupis philippinarum. Comp. Biochem. Phys. B156, 56–60.


















