
Citrobacter O-Antigens: Structure O-Antigenic Polysaccharide...
8
JOURNAL OF BACTERIOLOGY, May 1978, p. 462-469 0021-9193/78/0134-0462$02.00/0 Copyright i 1978 American Society for Microbiology Vol. 134, No. 2 Printed in U.S.A. Citrobacter O-Antigens: Structure of the O-Antigenic Polysaccharide from Citrobacter sp. 396t BARBARA JANN, PETER PREHM, AND KLAUS JANN* Max-Planck-Institut fur Immunbiologie, Freiburg, Germany Received for publication 14 December 1977 The structure of the 0-specific polysaccharide moiety of the lipopolysaccharide from Citrobacter 396 was elucidated by composition, methylation, and periodate oxidation studies. The repeating unit consists of four 2-linked mannoses and one 3-linked N-acetylglucosamine. One of the mannose units is substituted at C3 with a-glucose, and one is substituted at C3 with a-(2-0-acetyl)-abequose. All the mannosyl linkages appear to have the fl-configuration; the N-acetylglucosaminyl linkage has the a-configuration. In bacterial agglutination and passive hemagglu- tination in some Salmonella antisera, Citrobacter 396 as well as its 0-antigenic lipopolysaccharide expressed the serological factors 5 and 6. In corroboration of our structural studies, this showed the presence of a-(2-0-acetyl)-abequosyl-1,3- mannose (factor 5) and a-glucosyl-1,3-mannose (factor 6). The genus Citrobacter is closely related to Salmonella, and serological cross-reactions have been shown to exist between species of these genera (3, 22). Comparative studies on the chem- ical composition of the cell wall lipopolysaccha- rides (0 antigens) of Citrobacter and Salmo- nella (12, 28) showed that some of the cross- reacting 0 antigens belong to the same chemo- type (i.e., have identical sugar composition), whereas others share only some common sugar constituents. Three serologically distinct Citro- bacter species were shown to contain abequose. Two of them (Citrobacter 22 and 38) cross-react with Salmonella of group B (0 specificities 4, 5, and 12) and belong to the same chemotype. In contrast, Citrobacter 396 exhibits only 04 and 05 specificities, and its lipopolysaccharide lacks rhamnose. In an earlier study on the heteroge- neity of lipopolysaccharides (9), we analyzed the 0-specific polysaccharide of Citrobacter 396. It was found to consist of mannose, glucosamine, glucose, and abequose in the molar ratio of 4:1:1:1. The structure must thus be different from that of the 0-specific polysaccharide of Salmonella group B, which consists of equimo- lar amounts of galactose, rhamnose, mannose, abequose, and glucose. In the present paper we describe the structure and immunochemical properties of the 0-specific polysaccharide from Citrobacter 396. MATERIALS AND METHODS Bacteria. Citrobacter 396 (F7165) was grown on D1.5 agar (20) at 370C in a laboratory fermentor t This paper is dedicated to Otto Westphal on the occasion of his 65th birthday. (Eschweiler Co., Kiel) in a medium containing casein peptone CAT (1.5%), glucose (1%), yeast extract (Difco) (0.5%), Na2HPO4 2H20 (0.8%), NaCI (0.3%), and MgSO4 7H20 (0.02%). Isolation and purification of the lipopolysac- charide. Bacteria were extracted with 45% aqueous phenol at 650C for 10 min, and the lipopolysaccharide was obtained from the aqueous phase by repeated centrifugations at 105,000 x g. The final pellet was dissolved in water and lyophilized. Degradation of the lipopolysaccharide and fractionation of the carbohydrate moiety. Deg- radation was performed in 1% acetic acid at 1000C for 1.5 h. After removal of lipid A by centrifugation, the supernatant was lyophilized, and the material ob- tained was fractionated on Sephadex G-50 columns as described previously (9). Quantitative analyses. The monosaccharides were determined by gas-liquid chromatography of their alditol acetates (8) with xylose as an internal standard. Abequose was determined by the method of Cynkin and Ashwell (1), and glucosamine was deter- mined by the method of Strominger et al. (24). Chromatographic methods. Fractionation of pol- ysaccharides was carried out on Sephadex G-50 and G-200 with pyridine-acetate (10 ml of pyridine and 4 ml of glacial acetic acid per liter) as eluant. Oligosac- charides were fractionated on Sephadex G-25 or on Bio-Gel P2 with water as eluant. Further purification of oligosaccharides was accomplished by preparative paper chromatography on Whatman 3 paper with butanol-pyridine-water (4:6:3) as solvent. Carbohy- drate was detected in the eluants with the phenol- sulfuric acid reagent (2). Methylation analysis. Methylation analysis of the polysaccharide and oligosaccharides was performed as described earlier (6, 19). To establish the linkage to glucosamine, the methylation method described by SteUlner et al. (23) was used. The mixture of methyl- ated sugars was analyzed by gas-liquid chromatogra- 462 on February 1, 2019 by guest http://jb.asm.org/ Downloaded from
Transcript of Citrobacter O-Antigens: Structure O-Antigenic Polysaccharide...

JOURNAL OF BACTERIOLOGY, May 1978, p. 462-4690021-9193/78/0134-0462$02.00/0Copyright i 1978 American Society for Microbiology
Vol. 134, No. 2
Printed in U.S.A.
Citrobacter O-Antigens: Structure of the O-AntigenicPolysaccharide from Citrobacter sp. 396t
BARBARA JANN, PETER PREHM, AND KLAUS JANN*
Max-Planck-Institut fur Immunbiologie, Freiburg, Germany
Received for publication 14 December 1977
The structure of the 0-specific polysaccharide moiety of the lipopolysaccharidefrom Citrobacter 396 was elucidated by composition, methylation, and periodateoxidation studies. The repeating unit consists of four 2-linked mannoses and one
3-linked N-acetylglucosamine. One of the mannose units is substituted at C3 witha-glucose, and one is substituted at C3 with a-(2-0-acetyl)-abequose. All themannosyl linkages appear to have the fl-configuration; the N-acetylglucosaminyllinkage has the a-configuration. In bacterial agglutination and passive hemagglu-tination in some Salmonella antisera, Citrobacter 396 as well as its 0-antigeniclipopolysaccharide expressed the serological factors 5 and 6. In corroboration ofour structural studies, this showed the presence of a-(2-0-acetyl)-abequosyl-1,3-mannose (factor 5) and a-glucosyl-1,3-mannose (factor 6).
The genus Citrobacter is closely related toSalmonella, and serological cross-reactions havebeen shown to exist between species of thesegenera (3, 22). Comparative studies on the chem-ical composition of the cell wall lipopolysaccha-rides (0 antigens) of Citrobacter and Salmo-nella (12, 28) showed that some of the cross-reacting 0 antigens belong to the same chemo-type (i.e., have identical sugar composition),whereas others share only some common sugarconstituents. Three serologically distinct Citro-bacter species were shown to contain abequose.Two of them (Citrobacter 22 and 38) cross-reactwith Salmonella of group B (0 specificities 4, 5,and 12) and belong to the same chemotype. Incontrast, Citrobacter 396 exhibits only 04 and05 specificities, and its lipopolysaccharide lacksrhamnose. In an earlier study on the heteroge-neity of lipopolysaccharides (9), we analyzed the0-specific polysaccharide of Citrobacter 396. Itwas found to consist of mannose, glucosamine,glucose, and abequose in the molar ratio of4:1:1:1. The structure must thus be differentfrom that of the 0-specific polysaccharide ofSalmonella group B, which consists of equimo-lar amounts of galactose, rhamnose, mannose,abequose, and glucose. In the present paper wedescribe the structure and immunochemicalproperties of the 0-specific polysaccharide fromCitrobacter 396.
MATERIALS AND METHODSBacteria. Citrobacter 396 (F7165) was grown on
D1.5 agar (20) at 370C in a laboratory fermentor
t This paper is dedicated to Otto Westphal on the occasionof his 65th birthday.
(Eschweiler Co., Kiel) in a medium containing caseinpeptone CAT (1.5%), glucose (1%), yeast extract(Difco) (0.5%), Na2HPO4 2H20 (0.8%), NaCI (0.3%),and MgSO4 7H20 (0.02%).
Isolation and purification of the lipopolysac-charide. Bacteria were extracted with 45% aqueousphenol at 650C for 10 min, and the lipopolysaccharidewas obtained from the aqueous phase by repeatedcentrifugations at 105,000 x g. The final pellet wasdissolved in water and lyophilized.Degradation of the lipopolysaccharide and
fractionation of the carbohydrate moiety. Deg-radation was performed in 1% acetic acid at 1000C for1.5 h. After removal of lipid A by centrifugation, thesupernatant was lyophilized, and the material ob-tained was fractionated on Sephadex G-50 columns asdescribed previously (9).
Quantitative analyses. The monosaccharideswere determined by gas-liquid chromatography oftheir alditol acetates (8) with xylose as an internalstandard. Abequose was determined by the method ofCynkin and Ashwell (1), and glucosamine was deter-mined by the method of Strominger et al. (24).Chromatographic methods. Fractionation of pol-
ysaccharides was carried out on Sephadex G-50 andG-200 with pyridine-acetate (10 ml of pyridine and 4ml of glacial acetic acid per liter) as eluant. Oligosac-charides were fractionated on Sephadex G-25 or onBio-Gel P2 with water as eluant. Further purificationof oligosaccharides was accomplished by preparativepaper chromatography on Whatman 3 paper withbutanol-pyridine-water (4:6:3) as solvent. Carbohy-drate was detected in the eluants with the phenol-sulfuric acid reagent (2).
Methylation analysis. Methylation analysis of thepolysaccharide and oligosaccharides was performed asdescribed earlier (6, 19). To establish the linkage toglucosamine, the methylation method described bySteUlner et al. (23) was used. The mixture of methyl-ated sugars was analyzed by gas-liquid chromatogra-
462
on February 1, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 134, 1978
phy and mass spectrometry as described earlier (19)with a Finnigan model 3200 chromatograph-spectrom-eter.
Partial acid hydrolysis of the polysaccharide.To obtain oligosaccharides, the polysaccharide was
hydrolyzed with 1 N H2SO4 for 20 min at 100°C,neutralized with Ba(OH)2, lyophilized, and fraction-ated by chromatography. Another method was hy-drolysis with 50% acetic acid at 100°C for 14 h, followedby lyophilization of the hydrolysate.
Periodate oxidation and Smith degradation ofthe lipopolysaccharide. Periodate oxidation andSmith degradation of the lipopolysaccharide were per-
formed as previously described (18). Briefly, the lipo-polysaccharide was oxidized with sodium metaperox-ide for 3 days at 4°C. Excess reagent was then decom-posed with ethylene glycol, and the product was re-
duced with sodium borohydride. The resulting mixturewas dialyzed, and the product was obtained by lyoph-ilization. It was hydrolyzed in 0.25 N sulfuric acid for72 h at room temperature. The neutralized hydrolysatewas chromatographed on Sephadex G-25 with pyridin-ium acetate (pH 5.4) as eluant.Determination ofthe anomeric configurations.
Chromic acid oxidation of the peracetylated polysac-charide as described by Hoffman et al. (7) was usedfor the determination of the anomeric configuration.
Serological methods. The serological techniquesused in this study, i.e., bacterial agglutination andpassive hemagglutination, have been described before(10).
RESULTSIsolation of the polysaccharide. The 0-
specific polysaccharide from Citrobacter 396with the core oligosaccharide at the reducingend (see reference 11) was obtained by gradedhydrolysis, degradation of the lipopolysaccha-ride, and fractionation on Sephadex G-50 as
described previously (9). In this fractionation,the polysaccharide was eluted in an asymmetri-cal peak. The corresponding material could beresolved into two polysaccharides on a SephadexG-200 column. It was shown earlier (9) that thetwo polysaccharides, with molecular weights of16,750 and 53,900, respectively, had the same
sugar composition, the only difference being therelative amount of core sugars in these prepa-rations. For our structural studies we used themixture before fractionation on Sephadex G-200.The sugar composition of the polysaccharide isshown in Table 1. It consists of mannose, glu-cose, glucosamine, abequose, and O-acetyliin themolar ratio of 4:1:1:1:0.8. Besides these sugars,the polysaccharide contains small amounts ofgalactose and heptose, arising from the core
oligosaccharide at its reducing end.Periodate oxidation. When the polysaccha-
ride was treated with sodium metaperiodate for100 h at 4C, 50% of the mannose and practicallyall the glucose were oxidized (Table 1). N-Ace-tylglucosamine and abequose were not de-stroyed. Subsequent reduction of the oxidized
CITROBACTER 396 O-ANTIGEN 463
TABI.F. 1. Composition of the polysaccharide fromCitrobacter 396 before and after periodate oxidation
Polysaccharide Oxidized polysac-Polysaccharidecharide'~Component
% Molar ra- % Molar ra-tio tio
Abequose 9.8 0.9 10.4 0.9Mannose 48.6 4.0 25.2 2.0Glucose 14.5 1.2 0.6Galactose 1.1 0.0Heptose 2.0 1.1 0.5Glucosamine 12.7 1.0 13.8 1.1N-Acetyl 3.2 0.8 NDbO-Acetyl 2.1 0.0
a Before periodate oxidation the polysaccharide wastreated with dilute alkali to remove O-acetyl.
b ND, Not determined.
polysaccharide, followed by total acid hydroly-sis, liberated glycerol. Neither erythritol northreitol was detected. This indicated that noneof the oxidized sugars was substituted at C4.Methylation analysis. For a linkage analy-
sis, the polysaccharide was subjected to meth-ylation with dimethylsulfoxide-sodium hydride-methyliodide as described previously (18). Themethylated polysaccharide was purified by chro-matography on Sephadex LH-20 with ethanol-chloroform (2:1). The product was hydrolyzed,reduced with sodium borodeuteride, acetylatedwith acetic anhydride/pyridine, and the result-ing partially methylated [1-2H]alditol acetateswere then analyzed by gas chromatography andmass spectrometry. The gas chromatogram isshown in Fig. 1. The relative intensities of peaksa, b, and c were 1:2:2, respectively. Mass spectro-metric analysis showed that all peaks were ho-mogeneous. The main signals ofthe mass spectra(primary fragments) together with the retentiontimes of the corresponding peaks are listed inTable 2. By comparison with known data (7),the peaks can be ascribed to the following com-pounds: peak a, 1,5-di-O-acetyl-2,3,4,6-tetra-O-methylglucitol; peak b, 1,2,5-tri-O-acetyl-3,4,6-tri-O-methylmannitol; peak c, 1,2,3,5-tetra-O-acetyl-4,6-di-O-methylmannitol. The results in-dicate that the polysaccharide is branched, glu-cose being at the nonreducing end. Two man-noses were substituted at C2 and C3 and twomannoses at C2 only. These results are in goodagreement with the results of the periodate oxi-dation.To analyze also the linkage to N-acetylglucos-
amine, we methylated the polysaccharide by themethod of Steilner et al. (23). On subsequent gaschromatographic and mass spectrometric anal-ysis, 1,3,5-tri-O-acetyl-4,6-di-O-methyl-2-deoxy-2-(N-methylacetamido)-glucitol was found. Thisshowed that N-acetylglucosamine was substi-tuted at C3.
on February 1, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

464 JANN, PREHM, AND JANN
0
Va
.00
0-I;
A
a
Kb
c0
c
a._0
.S-
ax
0r
C
Time -
B
c
Time -
FIG. 1. Gas chromatographic patterns of the partially methylated alditol acetates obtained from the 0-specific polysaccharide from Citrobacter 396 before (A) and after (B) periodate oxidation. The N-acetylglucosamine residues escaped the methylation analysis under the conditions used. The partially methylatedderivative was decomposed and/or retained on the ion exchange resin during neutralization (see reference23).
TABLE 2. Retention times and mass spectrometric analysis ofpeaks a, b, and c obtained from methylatedpolysaccharide (Fig. IA) andperiodate-oxidized and methylatedpolysaccharide (Fig. IB)
Primary fragmentPeak tria Interpretation
45 89 117 161 189 205 261
a 1.00 x x x x x 1,5-di-O-acetyl-2,3,4,6-tetra-O-methylglucitol
b 1.95 x x x 1,2,5-tri-O-acetyl-3,4,6-tri-0-methylmannitol
c 3.19 x x x 1,2,3,5-tetra-0-acetyl-4,6-di-O-methylmannitol
a t, Retention time relative to 1,5-di-O-acetyl-2,3,4,6-tetra-0-methylglucitol.Isolation ofoligosaccharides. The polysac-
charide was hydrolyzed in 1 N sulfuric acid at1000C for 20 min. After neutralization with bar-ium hydroxide, the barium sulfate formed wasremoved by centrifugation. The supernatant wasapplied to a column (3 by 200 cm) of Bio-Gel P2and eluted with water. The elution pattern isshown in Fig. 2. Peak fractions were pooled, andthe material from fractions III and IV was puri-fied further by preparative paper chromatogra-phy. From fraction III, which eluted in the rangeof a penta- or hexasaccharide, two oligosaccha-rides (B and C) could be purified. From fractionIV, which eluted in the range of a tetrasaccha-ride, one main oligosaccharide (A) was isolatedin a pure form. The sugar analyses ofthe purifiedoligosaccharides are given in Table 3. Oligosac-charide A consists ofmannose only, oligosaccha-ride B contains mannose and glucosamine in a
molar ratio of4:1, and oligosaccharide C containsmannose, glucose, and glucosamine in the ratioof 4:1:1. In oligosaccharides B and C, N-acetyl-glucosamine could be determined (12) in theMorgan-Elson reaction without prior hydrolysis.This showed that glucosamine is at the reducingend of these oligosaccharides. Oligosaccharide Ccould also be obtained in a very good yield froma hydrolysate ofthe polysaccharide in 50% aceticacid. A 100-mg portion of the polysaccharidewas hydrolyzed with 10 ml of 50% acetic acid at1000C for 14 h. The acetic acid was removed bylyophilization. The material obtained was chro-matographed on a Sephadex G-50 column. Themain peak was refractionated on Sephadex G-25. About 50% of the starting material couldthus be obtained as a pure oligosaccharide.Smith degradation ofthe lipopolysaccha-
ride. The periodate oxidation and borohydride
J. BACTERIOL.
on February 1, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
VoiL. 134, 1978
reduction steps of this degradation procedurewere carried out with the complete lipopolysac-charide, because the material could be dialyzedwithout loss. After mild hydrolysis, the productswere fractionated on a Sephadex G-25 column(1.5 by 100 cm). Besides material with highermolecular weight, resulting from incomplete hy-drolysis, two oligosaccharides were obtained.
tIO
0,8
0.6
0.4
0.2
III1- ~~~~~~~IV11
VI I
80. 1 00 10 l 6
so bo Elujte I ml IEtae(ml)
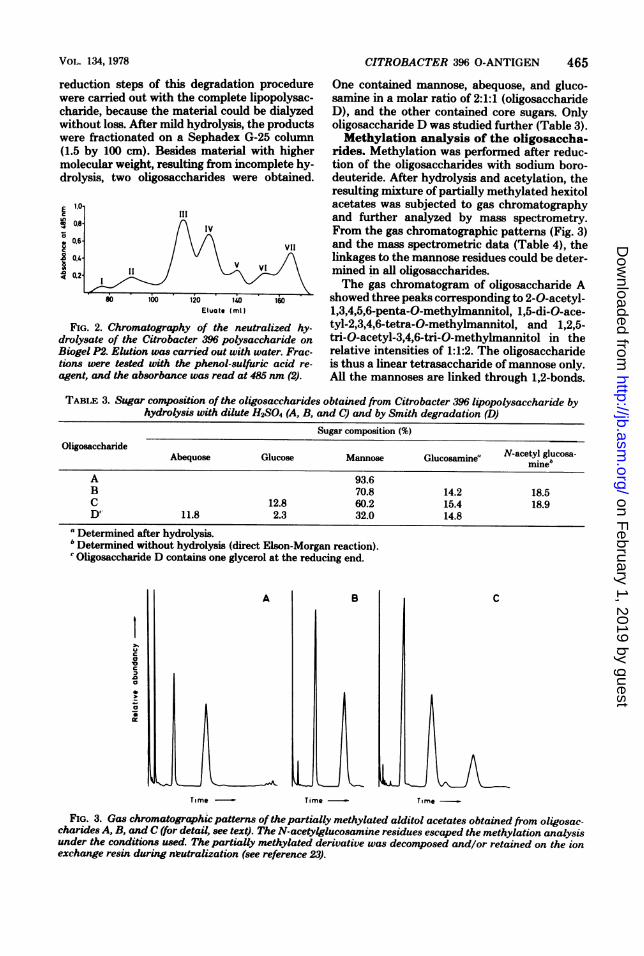
FIG. 2. Chromatography of the neutralized hy-drolysate of the Citrobacter 396 polysaccharide onBiogel P2. Elution was carried out with water. Frac-tions were tested with the phenol-sulfuric acid re-agent, and the absorbance was read at 485 nm (2).
CITROBACTER 396 O-ANTIGEN 465
One contained mannose, abequose, and gluco-samine in a molar ratio of 2:1:1 (oligosaccharideD), and the other contained core sugars. Onlyoligosaccharide D was studied further (Table 3).Methylation analysis of the oligosaccha-
rides. Methylation was performed after reduc-tion of the oligosaccharides with sodium boro-deuteride. After hydrolysis and acetylation, theresulting mixture of partially methylated hexitolacetates was subjected to gas chromatographyand further analyzed by mass spectrometry.From the gas chromatographic patterns (Fig. 3)and the mass spectrometric data (Table 4), thelinkages to the mannose residues could be deter-mined in all oligosaccharides.The gas chromatogram of oligosaccharide A
showed three peaks corresponding to 2-0-acetyl-1,3,4,5,6-penta-0-methylmannitol, 1,5-di-O-ace-tyl-2,3,4,6-tetra-0-methylmannitol, and 1,2,5-tri-O-acetyl-3,4,6-tri-0-methylmannitol in therelative intensities of 1:1:2. The oligosaccharideis thus a linear tetrasaccharide of mannose only.All the mannoses are linked through 1,2-bonds.
TABLE 3. Sugar composition of the oligosaccharides obtained from Citrobacter 396 lipopolysaccharide byhydrolysis with dilute H2SO4 (A, B, and C) and by Smith degradation (D)
Sugar composition (%)Oligosaccharide N-acetyl glucosa-
Abequose Glucose Mannose Glucosamine! mine'
A 93.6B 70.8 14.2 18.5C 12.8 60.2 15.4 18.9D' 11.8 2.3 32.0 14.8
aDetermined after hydrolysis.b Determined without hydrolysis (direct Elson-Morgan reaction).' Oligosaccharide D contains one glycerol at the reducing end.
A
Time
B
Time
C
Time
FIG. 3. Gas chromatographic patterns of the partially methylated alditol acetates obtained from oligosac-charides A, B, and C (for detail, see text). The N-acetylglucosamine residues escaped the methylation analysisunder the conditions used. The partially methylated derivative was decomposed and/or retained on the ionexchange resin during nLeutralization (see reference 23).
Ezcin
a
a
la
c
Dz
a
0
0
on February 1, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

466 JANN, PREHM, AND JANN
TABLE 4. Results ofgas-liquid chromatography and mass spectrometry of the partially methylated alditolacetates obtained from oligosaccharides of Citrobacter 396 0-specific polysaccharide
osa tt,h of Primary fragment Inten- Interpretation
peak 45 89 117 158 161 162 189 202 205 206 261 sity
A 0.26 x x x x 0.7 2-0-acetyl-1,3,4,5,6-penta-0-methylmannitol
1.00 x x x x x 1.0 1,5-di-O-acetyl-2,3,4,6-te-tra-0-methylmannitol
1.9 x x x 2.1 1,2,5-tri-O-acetyl-3,4,6-tri-0-methylmannitol
B 1.00 x x x x x x 1.0 1,5-di-O-acetyl-2,3,4,6-te-tra-0-methylmannitol
1.9 x x x 2.9 1,2,5 tri-O-acetyl-3,4,6-tri-0-methylmannitol
C 1.00 x x x x x 2.0 1,5-di-O-acetyl-2,3,4,6-te-tra-0-methylglucitol +1,5-di-O-acetyl-2,3,4,6-te-tra-0-methylmannitol
1.9 x x x 2.2 1,2,5-tri-O-acetyl-3,4,6-tri-0-methylmannitol
3.2 x x x 0.9 1,2,3,5-tetra-0-acetyl-4,6-10 di-0-methylmannitol
D NCb x x x 1.0 1,2,5-tri-O-acetyl-3,4,6-tri-0-methylmannitol
NC x x x 0.9 1,2,3,5-tetra-0-acetyl-4,6-di-0-methylmannitol
NC x x x x x x 0.8 1,5-di-O-acetyl-3,4,6-tri-0-methyl-2-deoxy-2-N-methylacetamidoglucitol
" OS, Oligosaccharide.b tt, Retention time relative to 1,5-di-O-acetyl-2,3,4,6-tetra-0-acetylglucitol.'NC, Not comparable, because this chromatogram was run with a temperature gradient.
Oligosaccharide B showed two peaks corre-sponding to 1,5-di-0-acetyl-2,3,4,6-tetra-0-methylmannitol and 1,2,5-tri-O-acetyl-3,4,6-tri-0-methylmannitol in the relative intensities of1:3. As demonstrated above, this oligosaccharidehas N-acetylglucosamnine at its reducing end. Itis thus also a linear oligosaccharide of four man-nose units linked to glucosamine. All the man-noses are linked through 1,2-bonds. The gaschromatogram of oligosaccharide C showedthree peaks. The first peak corresponded to 1,5-di-O-acetyl-2,3,4,6-tetra-0-methylmannitol and1,5-di-O-acetyl-2,3,4,6-tetra-0-methylglucitol,which cannot be separated under the conditionsused. The second peak corresponded to 1,2,5-tri-0-acetyl-3,4,6-tri-0-methylmannitol, and thethird peak corresponded to 1,2,3,5-tetra-0-ace-tyl-4,6-di-0-methylmannitol. The reducing endof oligosaccharide C is N-acetylglucosamine.There is one branch mannose, which is substi-tuted by mannose and glucose. OligosaccharideD was methylated by the method of Steilner etal. (23). The gas chromatogram showed threepeaks corresponding to 1,2,5-tri-O-acetyl-3,4,6-
tri-0-methylmannitol, 1,2,3,5-tetra-0-acetyl-4,6-di-0-methylmannitol, and 1,5-di-O-acetyl-4,3,6-tri-0-methyl-2-deoxy-2-(N-methylacetam-ido)-glucitol, respectively.Determination of the anomeric configu-
ration. For the determination of the anomericconfiguration, the method described by Hoffmanet al. (7) was used, in which peracetylated poly-or oligosaccharides are treated with chromiumtrioxide in acetic acid. In this reaction, sugars inequatorial positions are readily oxidized, andsugars in axial positions are stable. The resultsobtained with the polysaccharide from Citro-bacter 396 show that glucose was recovered com-pletely, mannose was destroyed to about 50%,and abequose and N-acetylglucosamine were notdestroyed. This indicated that, in the polysac-charide, glucose and abequose are boundthrough a-glycosidic linkages. An a-N-acetylglu-cosaminyl linkage is also indicated by the lowermolar dextrorotation of oligosaccharide C(+370) as compared with that of the completepolysaccharide (+53°). According to previouslypublished observations (7), all mannoses could
J. BACTERIOL.
on February 1, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

CITROBACTER 396 O-ANTIGEN 467
be bound in ,6-glycosidic linkages, although onlyabout 50% were destroyed by the oxidation.
Serological studies. The serological cross-reaction between Citrobacter 396 and Salmo-nella group B has been reported earlier (3, 22,28). Our structural studies indicated a similarityof the polysaccharide chains of Citrobacter 396and of Salmonella group C (see reference 4). Wetherefore expected serological cross-reaction ofthe latter two species. Using bacterial aggluti-nation as well as passive hemagglutination withthe respective lipopolysaccharides in the anti-sera to Salmonella groups B and C, we wereable to show that these cross-reactions did occur(Table 5).
DISCUSSIONThe genus Citrobacter, which was classified
by Sedlak and Slajsowa (see reference 22), wasfound to contain many strains that cross-reactwith other gram-negative enteric bacteria (3,22). Notably, three strains of Citrobacter, 022,038, and 396, exhibit strong cross-reactivity withfactors 4 (abequose) and 5 (2-0-acetylabequose)of Salmonella group B. Strains 022 and 038 alsocross-react with Salnonella factor 12 (a-gluco-syl-1,4-galactose), and strain 396 does not.
cose and 50% of the mannose and that glucosa-mine and abequose were resistant to oxidation.Methylation analysis showed that two mannoseswere substituted at C2, and two mannoses weresubstituted at C2 and C3, and must thus bebranch points. Glucose and abequose were un-substituted and are thus the sugar branches.Methylation analysis by the method of Steilneret al. (23) and results from oligosaccharide anal-ysis show that glucosamine, linked through its 3position, is part of the main chain. The sequenceof monosaccharide residues follows from theidentification of five oligosaccharides. Four ofthese were obtained by mild acid hydrolysis, andone was obtained after periodate oxidation fol-lowed by Smith degradation. In three oligosac-charides, N-acetylglucosamine is the terminalreducing sugar. This was shown by direct Elson-Morgan-reaction without prior hydrolysis of theoligosaccharide. Sugar analysis and methylationstudies led to the formulation of the oligosac-charide structures given in Fig. 4. From thesepartial structures and from the results of thechromic acid oxidation, we conclude that theCitrobacter 396 polysaccharide has the followingstructure:
2-Oac-Abe Gica 11,3 a 11,3
2 1.2 1,2 1.2 1,32 Man -' Man -. Man -p Man b GlcNac -(p) (1i) (8) (4) a
Whereas the 0-antigenic lipopolysaccharides ofCitrobacter 022 and 038 have the same sugarcomposition as the lipopolysaccharide of S. ty-phimurium (group B) (6), it has been shown (28)that Citrobacter 396 contains neither galactosenor rhamnose in its lipopolysaccharide. There-fore, in the latter strain, abequose cannot beattached to a poly(gal-man-rham) chain as inSalmonella group B, and it was of interest tostudy the structure ofthis 0-specific polysaccha-ride. We previously reported (9) the 0-specificpolysaccharide from Citrobacter 396 to consistof mannose, glucose, glucosamine, and abequosein a molar ratio of 4:1:1:1. In the present inves-tigation we found that periodate destroyed glu-
(Abbreviations: Oac, O-acetyl; Abe, abequose;Glc, glucose; Man, mannose; Nac, N-acetyl; Rha,rhamnose; Gal, galactose.)
The strong reaction of the polysaccharide in anantiserum against the a-2-0-acetylabequosylgroup (factor 5 serum) showed that this group ispresent in the Citrobacter 396 polysaccharide.This finding corroborates the results of thechromic acid oxidation.The structure of the lipopolysaccharide of
Citrobacter 396 combines elements of those ofS. typhimurium (6) and S. montevideo (C. G.Hellerqvist and J. Gmeiner, personal communi-cations).
S. t.phmunurumn2-Oac-Abe
a 11,3Glc
a 11,3
-.Man 1,4 * Rha 'L Gal IO(16) (a) a
Glc1,3 ,/ 1,3
12 Ma 1,2 .2 1,3 1S. montevideo: v- Man -ab Man a,- Man- ' Mant - - GicNac -'
VOL. 134, 1978
on February 1, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
468 JANN, PREHM, AND JANN
TABI.F 5. Reciprocal titers of bacterial agglutination andpassive hemagglutination of Citrobacter 396bacteria before and after heating to 100°C and of isolated lipopolysaccharide (LPS) before and after alkali
treatment (LPS alk)
Titer in antiserum against:
Method Antigen S. typhimu- S. brium Factor 4 Factor 5 (6. 7)"(4, 5, 12)"
Bacterial agglutination Bacteria 320 160 320 640Bacteria 100°C 320 160 320 320
Passive hemagglutination LPS 640 20 320 640LPS alk 320 320 <10 640
"Serological factors.
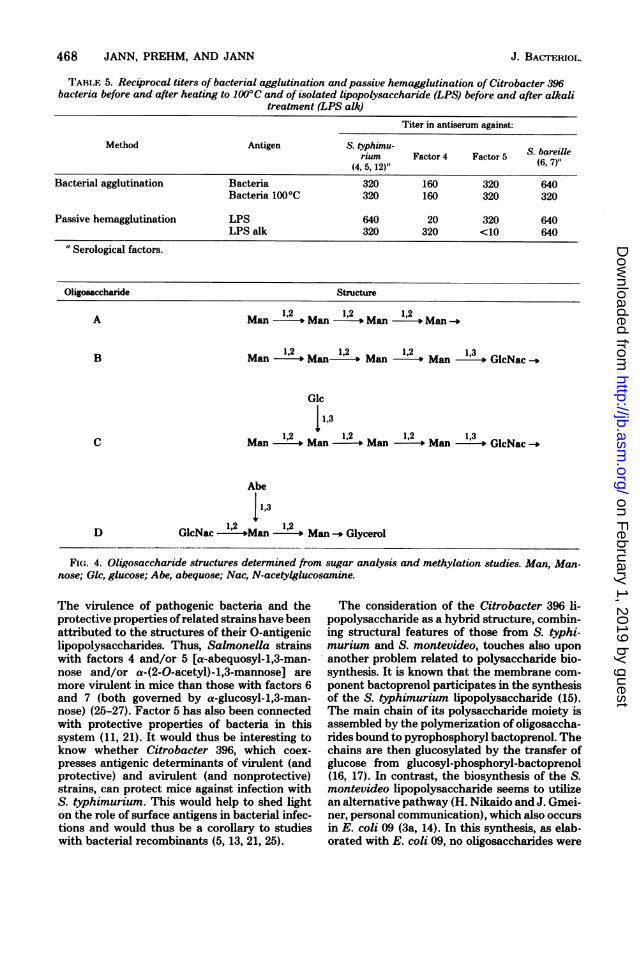
Oligosaccharide Structure
A Man 1-2' Man- 12 ,Man 1,2- Man
B Man 1,2 Manl 1,2 1M 2l Man 1,3
o GlcNac
Glc
11,3C Man 12 Man 12 Man 12 Man 13GcNac
Abe
113D GlcNac '2 S"12Man' *Man Glycerol
FIw. 4. Oligosaccharide structures determined from. sugar analysis and methylation studies. Man, Man-nose; Glc, glucose; Abe, abequose; Nac, N-acetylglucosamine.
The virulence of pathogenic bacteria and theprotective properties ofrelated strains have beenattributed to the structures of their 0-antigeniclipopolysaccharides. Thus, Salmonella strainswith factors 4 and/or 5 [a-abequosyl-1,3-man-nose and/or a-(2-0-acetyl)-1,3-mannose] aremore virulent in mice than those with factors 6and 7 (both governed by a-glucosyl-1,3-man-nose) (26-27). Factor 5 has also been connectedwith protective properties of bacteria in thissystem (11, 21). It would thus be interesting toknow whether Citrobacter 396, which coex-presses antigenic determinants of virulent (andprotective) and avirulent (and nonprotective)strains, can protect mice against infection withS. typhimurium. This would help to shed lighton the role of surface antigens in bacterial infec-tions and would thus be a corollary to studieswith bacterial recombinants (5, 13, 21, 25).
The consideration of the Citrobacter 396 li-popolysaccharide as a hybrid structure, combin-ing structural features of those from S. typhi-murium and S. montevideo, touches also uponanother problem related to polysaccharide bio-synthesis. It is known that the membrane com-ponent bactoprenol participates in the synthesisof the S. typhimurium lipopolysaccharide (15).The main chain of its polysaccharide moiety isassembled by the polymerization of oligosaccha-rides bound to pyrophosphoryl bactoprenol. Thechains are then glucosylated by the transfer ofglucose from glucosyl-phosphoryl-bactoprenol(16, 17). In contrast, the biosynthesis of the S.montevideo lipopolysaccharide seems to utilizean alternative pathway (H. Nikaido and J. Gmei-ner, personal communication), which also occursin E. coli 09 (3a, 14). In this synthesis, as elab-orated with E. coli 09, no oligosaccharides were
J. BACTERIOL,.
on February 1, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 134, 1978
found as intermediates, and the carrier moleculeon which the polysaccharide chain grows is notbactoprenol. In light of the fact that there arealternative synthetic pathways leading to bac-terial cell wall polysaccharides, it would seemthat Citrobacter 396 is a good candidate to usein a comparative study.
ACKNOWLEDGMENTSWe thank S. Schlecht for the cultivation of the bacteria.
We gratefully acknowledge the expert help of C. Heidkamper.
LITERATURE CITED
1. Cynkin, M. A., and G. Ashwell. 1960. Estimation of 3-deoxysugars by means of the malonaldehyde-thiobar-bituric acid reaction. Nature (London) 186:155-156.
2. Dubois, M., K. A. Gilles, J. K. Hamilton, P. A. Rebers,and R. Smith. 1955. Colorimetric method for deter-mination of sugars and related substances. Anal. Chem.28:350-357.
3. Edwards, P. R., and W. H. Ewing. 1966. Identificationof Enterobacteriaceae. Burgers Publishing Co., Minne-apolis, Minn.
3a.Flemming, H. C., and K. Jann. 1978. Biosynthesis ofthe 09 antigen of Escherichia coli. Growth of thepolysaccharide chain. Eur. J. Biochem. 83:47-52.
4. Fuller, N. A., and A. M. Staub. 1968. Immunochemicalstudies on Salmonella. 13. Chemical changes appearingon the specific polysaccharide of S. cholera suis (62, 7)after its conversion by phage 14 (6, 7). Eur. J. Biochem.4:286-300.
5. Gemski, P., Jr., D. G. Sheahan, 0. Washington, andS. B. Formal. 1972. Virulence of Shigella flexnerihybrids expressing Escherichia coli somatic antigens.Infect. Immun. 6:104-111.
6. Hellerqvist, C. G., B. Lindberg, S. Svensson, T.Holme, and A. A. Lindberg. 1968. Structural studieson the 0-specific side-chains of the cell-wall lipopoly-saccharide from Salmonella typhimurium 395MS. Car-bohydr. Res. 8:43-55.
7. Hoffman, J., B. Linberg, and S. Svensson. 1972. De-termination of the anomeric configuration of sugar res-idues in acetylated oligo- and polysaccharides by oxi-dation with chromium trioxide in acetic acid. ActaChem. Scand. 26:661-666.
8. Holme, T., A. A. Lindberg, P. J. Garegg, and T. Onn.1968. Chemical composition of cell-wall polysaccharidesof rough mutants of Salnonella typhimurium. J. Gen.Microbiol. 52:45-54.
9. Jann, B., K. Reske, and K. Jann. 1975. Heterogeneityof lipopolysaccharides. Analysis of polysaccharide chainlengths by sodium dodecylsulfate-polyacrylamide gelelectrophoresis. Eur. J. Biochem. 60:239-246.
10. Jann, K., B. Jann, F. 0rskov, . 0rskov, and 0.Westphal. 1965. Immunchemische Untersuchungen anK-Antigen von Escherichia Coli. II. Das K-Antigen vonE. coli 08:K42(A):H-. Biochem. Z. 342:1-22.
11. Jann, K., and 0. Westphal. 1975. Microbial polysac-charides, p. 1-125. In M. Sela (ed.), The antigens, vol.3. Academic Press Inc., New York.
12. Keleti, J., 0. Luderitz, D. Mlynacik, and J. Sedlak.1971. Immunochemical studies on Citrobacter 0 anti-
CITROBACTER 396 O-ANTIGEN
gens (lipopolysaccharides). Eur. J. Biochem. 20:237-244.
13. Kiefer, W., G. Schmidt, B. Jann, and K. Jann. 1976.Genetic transfer of Salmonella 0 antigens to Esche-richia coli 08. J. Gen. Microbiol. 92:311-324.
14. Kopmann, H. J., and K. Jann. 1975. Biosynthesis of the09 antigen of Escherichia coli. The polysaccharide com-ponent of E. coli 09:K29-. Eur. J. Biochem. 60:587-601.
15. Nikaido, H. 1973. Biosynthesis and assembly of lipopoly-saccharide and the outer membrane layer of gram-neg-ative cell wall, p. 131-208. In L. Leive (ed.), Bacterialmembranes and walls. Microbiology Series, vol. 1. M.Decker, Inc., New York.
16. Nikaido, H., K. Nikaido, and T. Nakae. 1971. Gluco-sylation of lipopolysaccharide in Salmonella: biosyn-thesis of 0 antigen factor 122. I. Over all reactions. J.Biol. Chem. 246:3902-3911.
17. Nikaido, K., and H. Nikaido. 1971. Glucosylation oflipopolysaccharide in Salmonella: biosynthesis of 0antigen factor 122. II. Structure of the lipid intermedi-ate. J. Biol. Chem. 246:3912-3919.
18. Prehm, P., B. Jann, and K. Jann. 1976. The 09 antigenof E. coli. Structure of the polysaccharide chain. Eur.J. Biochem. 67:53-56.
19. Reske, K., and K. Jann. 1972. The 08 antigen of Esch-erichia coli. Structure of the polysaccharide chain. Eur.J. Biochem. 31:320-328.
20. Schlecht, S., and 0. Westphal. 1966. Wachstum undLipopolysaccharid (0-Antigen) Gehalt von Salmonellenbei Zuchtung auf Agarnihrboden. Zentralbl. Bakteriol.Parasitenkd. Infektionskr. Hyg. Abt. 1 Orig.200:241-250.
21. Schmidt, G., and W. Kiefer. 1974. Salmonella-E. coliHybride. Charakterisierung und immunologische Wirk-samkeit. Zentralbl. Bakteriol. Parasitenkd. Infek-tionskr. Hyg. Abt. 1 Orig. 227:257-262.
22. Sedlak, J., and H. Rische. 1961. EnterobacteriaceaeInfektionen. VEB Georg Thieme Verlag, Leipzig.
23. Stelner, K., H. Saito, and S. Hakomori. 1973. Deter-mination of amino-sugar linkages in glycolipids bymethylation. Aminosugar linkages of ceramide penta-saccharides of rabbit erythrocytes and of Forssmanantigen. Arch. Biochem. Biophys. 155:464-472.
24. Strominger, J. L., J. T. Park, and J. Thompson. 1959.Composition of the cell wall of Staphylococcus aureus.Its relation to the mechanism of action of penicillin. J.Biol. Chem. 234:3263-3271.
25. Valtonen, V. V. 1970. Mouse virulence of Salmonellastrains: the effect of different smooth type 0 side chains.J. Gen. Microbiol. 64:255-268.
26. Valtonen, V. V., J. Aird, M. Valtonen, 0. Miikela,and P. H. Miikela. 1971. Mouse virulence of Salmo-nella. Antigen dependent differences are demonstrableafter immunosuppression. Acta Pathol. Microbiol.Scand. Sect. B 79:715-718.
27. Valtonen, V. V., and P. H. Miakela. 1971. The effect oflipopolysaccharide modifications-antigenic factors 1,5, 122 and 27-on the virulence of Salmonella strainsfor mice. J. Gen. Microbiol. 69:107-115.
28. Westphal, O., F. Kauffmann, 0. Luderitz, and H.Stierlin. 1960. Zur Immunchemie der O-Antigene vonEnterobacteriaceae. III. Analyse der Zuckerbausteinekreuzreagierender Salmonella-, Arizona- und Esche-richia-0-Antigene. Zentralbl. Bakteriol. Parasitenkd.Infektionstr. Hyg. Abt. 1 Orig. 179:336-342.
469
on February 1, 2019 by guest
http://jb.asm.org/
Dow
nloaded from


















