
CHF1/Hey2 Plays a Pivotal Role in Left Ventricular...
12
CHF1/Hey2 Plays a Pivotal Role in Left Ventricular Maturation Through Suppression of Ectopic Atrial Gene Expression Nobutaka Koibuchi, Michael T. Chin Abstract—We previously reported that mice lacking the hairy-related basic helix–loop– helix (bHLH) transcription factor CHF1/Hey2 develop a thin-walled left ventricle. To explore the basis for this phenotype, we examined regional gene expression patterns in the developing myocardium. We found that atrial natriuretic factor (ANF), which is normally expressed in the atria and trabeculae and is restricted from the developing compact myocardium beginning at embryonic day 13.5, is persistently expressed in the left ventricular compact myocardium of the knockout animals. We also examined the expression pattern of the T-box transcription factor Tbx5, a known regulator of ANF, and an additional Tbx5-dependent gene, connexin 40 (Cx40), both of which share a similar expression pattern to ANF during development. Tbx5 and Cx40 were similarly expressed ectopically in the compact myocardium of the CHF1/Hey2 knockout mouse. The atrial contractile genes mlc1a and mlc2a were also expressed ectopically in the left ventricular compact myocardium, providing evidence for a general dysregulation of atrial gene expression. Crossing of a myocardial-specific CHF1/Hey2 transgenic mouse with the knockouts led to rescue of the thin-walled myocardial phenotype and restoration of the normal patterns of gene expression. Myocardial cell proliferation, which has been shown previously to be suppressed by Tbx5, was also decreased in the knockout mice and rescued by the transgene. Our findings suggest that CHF1/Hey2 suppresses atrial identity in the left ventricular compact myocardium, facilitates myocardial proliferation by suppressing Tbx5, and thereby promotes proper ventricular myocardial maturation. (Circ Res. 2007;100:850-855.) Key Words: CHF1/Hey2 Tbx5 cardiac development knockout mouse W e and others have previously identified a family of cardiovascular restricted, basic helix–loop– helix (bHLH) transcription factors related to hairy, a drosophila bHLH gene involved in Notch signaling and patterning of the peripheral nervous system. 1–6 This family has received various names, including Hey, Hesr, HRT, CHF, gridlock, and HERP. In zebrafish, gridlock has been found to play an important role in development of the aorta and in arterial versus venous cell fate decisions. 5,7 Numerous studies have postulated roles for these genes in Notch signaling, 6,8,9 endothelial differentiation, 10,11 skeletal muscle differentiation, 12 transcriptional repression, 2,12,13 smooth muscle function, 9,14 –16 morphogenesis of the heart, 17–20 and cardiac hypertrophy. 21 CHF1/Hey2 (also called Hesr-2, HRT2, gridlock, and HERP1) was originally described as a bHLH protein ex- pressed primarily in the developing ventricular myocardium and vasculature. 2–4 Loss-of-function studies in zebrafish ini- tially suggested that the primary effect of CHF1/Hey2 is on vascular development. 5,7 Loss-of-function studies in mice, how- ever, revealed primarily cardiac abnormalities. 17–20,22 The spec- trum of defects varies, with reports of cardiomyopathy, 17,19 ventricular septal defects, 18,19 valvular abnormalities, 18,20,22 vas- cular abnormalities, 16,20 and a thin-walled myocardium. 20 The basis for this variation is primarily genetic, as the phenotype varies significantly by genetic background, although there is some variation even when the background is inbred. 20 Our previous work on the phenotype of the CHF1/Hey2 knockout mice inbred to the C57BL/6 background has re- vealed the importance of CHF1/Hey2 in the developing myocardium. In these mice, the myocardium is thin walled, because of a marked decrease in the thickness of the compact myocardium, 20 and this phenotype is completely penetrant. The importance of CHF1/Hey2 in the myocardium is further underscored by our finding that a transgenic mouse express- ing CHF1/Hey2 in the myocardium can not only rescue the myocardial phenotype in the knockout mice but can also partially rescue the septal and valvular phenotypes. 20 To identify potential mechanisms by which CHF1/Hey2 affects myocardial development, we performed a systematic analysis of regional myocardial gene expression, using mark- ers of the atria, trabeculae, and ventricles, and also assessed for an alteration in myocardial cell proliferation. We have Original received October 13, 2006; revision received January 17, 2007; accepted February 19, 2007. From the Vascular Medicine Research, Cardiovascular Division, Department of Medicine, Brigham and Women’s Hospital and Harvard Medical School, Boston, Mass. Correspondence to Center for Cardiovascular Biology, University of Washington, 815 Mercer St, Rm 353, Seattle, WA 98109. E-mail mtchin@ u.washington.edu © 2007 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000261693.13269.bf 850 by guest on July 9, 2018 http://circres.ahajournals.org/ Downloaded from by guest on July 9, 2018 http://circres.ahajournals.org/ Downloaded from by guest on July 9, 2018 http://circres.ahajournals.org/ Downloaded from by guest on July 9, 2018 http://circres.ahajournals.org/ Downloaded from by guest on July 9, 2018 http://circres.ahajournals.org/ Downloaded from by guest on July 9, 2018 http://circres.ahajournals.org/ Downloaded from by guest on July 9, 2018 http://circres.ahajournals.org/ Downloaded from
Transcript of CHF1/Hey2 Plays a Pivotal Role in Left Ventricular...

CHF1/Hey2 Plays a Pivotal Role in Left VentricularMaturation Through Suppression of Ectopic Atrial
Gene ExpressionNobutaka Koibuchi, Michael T. Chin
Abstract—We previously reported that mice lacking the hairy-related basic helix–loop–helix (bHLH) transcription factorCHF1/Hey2 develop a thin-walled left ventricle. To explore the basis for this phenotype, we examined regional geneexpression patterns in the developing myocardium. We found that atrial natriuretic factor (ANF), which is normallyexpressed in the atria and trabeculae and is restricted from the developing compact myocardium beginning at embryonicday 13.5, is persistently expressed in the left ventricular compact myocardium of the knockout animals. We alsoexamined the expression pattern of the T-box transcription factor Tbx5, a known regulator of ANF, and an additionalTbx5-dependent gene, connexin 40 (Cx40), both of which share a similar expression pattern to ANF during development.Tbx5 and Cx40 were similarly expressed ectopically in the compact myocardium of the CHF1/Hey2 knockout mouse.The atrial contractile genes mlc1a and mlc2a were also expressed ectopically in the left ventricular compactmyocardium, providing evidence for a general dysregulation of atrial gene expression. Crossing of a myocardial-specificCHF1/Hey2 transgenic mouse with the knockouts led to rescue of the thin-walled myocardial phenotype and restorationof the normal patterns of gene expression. Myocardial cell proliferation, which has been shown previously to besuppressed by Tbx5, was also decreased in the knockout mice and rescued by the transgene. Our findings suggest thatCHF1/Hey2 suppresses atrial identity in the left ventricular compact myocardium, facilitates myocardial proliferationby suppressing Tbx5, and thereby promotes proper ventricular myocardial maturation. (Circ Res. 2007;100:850-855.)
Key Words: CHF1/Hey2 � Tbx5 � cardiac development � knockout mouse
We and others have previously identified a family ofcardiovascular restricted, basic helix–loop–helix (bHLH)
transcription factors related to hairy, a drosophila bHLH geneinvolved in Notch signaling and patterning of the peripheralnervous system.1–6 This family has received various names,including Hey, Hesr, HRT, CHF, gridlock, and HERP. Inzebrafish, gridlock has been found to play an important role indevelopment of the aorta and in arterial versus venous cell fatedecisions.5,7 Numerous studies have postulated roles for thesegenes in Notch signaling,6,8,9 endothelial differentiation,10,11
skeletal muscle differentiation,12 transcriptional repression,2,12,13
smooth muscle function,9,14–16 morphogenesis of the heart,17–20
and cardiac hypertrophy.21
CHF1/Hey2 (also called Hesr-2, HRT2, gridlock, andHERP1) was originally described as a bHLH protein ex-pressed primarily in the developing ventricular myocardiumand vasculature.2–4 Loss-of-function studies in zebrafish ini-tially suggested that the primary effect of CHF1/Hey2 is onvascular development.5,7 Loss-of-function studies in mice, how-ever, revealed primarily cardiac abnormalities.17–20,22 The spec-trum of defects varies, with reports of cardiomyopathy,17,19
ventricular septal defects,18,19 valvular abnormalities,18,20,22 vas-cular abnormalities,16,20 and a thin-walled myocardium.20 Thebasis for this variation is primarily genetic, as the phenotypevaries significantly by genetic background, although there issome variation even when the background is inbred.20
Our previous work on the phenotype of the CHF1/Hey2knockout mice inbred to the C57BL/6 background has re-vealed the importance of CHF1/Hey2 in the developingmyocardium. In these mice, the myocardium is thin walled,because of a marked decrease in the thickness of the compactmyocardium,20 and this phenotype is completely penetrant.The importance of CHF1/Hey2 in the myocardium is furtherunderscored by our finding that a transgenic mouse express-ing CHF1/Hey2 in the myocardium can not only rescue themyocardial phenotype in the knockout mice but can alsopartially rescue the septal and valvular phenotypes.20
To identify potential mechanisms by which CHF1/Hey2affects myocardial development, we performed a systematicanalysis of regional myocardial gene expression, using mark-ers of the atria, trabeculae, and ventricles, and also assessedfor an alteration in myocardial cell proliferation. We have
Original received October 13, 2006; revision received January 17, 2007; accepted February 19, 2007.From the Vascular Medicine Research, Cardiovascular Division, Department of Medicine, Brigham and Women’s Hospital and Harvard Medical
School, Boston, Mass.Correspondence to Center for Cardiovascular Biology, University of Washington, 815 Mercer St, Rm 353, Seattle, WA 98109. E-mail mtchin@
u.washington.edu© 2007 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000261693.13269.bf
850
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from

found that loss of CHF1/Hey2 leads to ectopic expression ofatrial genes in the left ventricular compact myocardium,including the transcription factor Tbx5 and its target genesANF and Cx40, and a profound decrease in myocardial cellproliferation. Our findings suggest an important mechanismby which CHF1/Hey2 regulates ventricular maturation.
Materials and MethodsAnimalsThe mice lacking CHF1/Hey2 and expressing CHF1/Hey2 under thecontrol of mlc2v promoter on a C57BL/6 background were generatedand bred as described previously.20 All animals were managedaccording to guidelines published in the Use and Care of LaboratoryAnimals and in accordance with IACUC approved experimentalprotocols.
In Situ Hybridization and ImmunohistochemistryIn situ hybridization was performed as described previously,20,23,24
with slight modifications. The embryos were fixed (4% formalde-hyde, 0.1 mol/L 4-morpholinepropanesulfonic acid, 2 mmol/LEGTA, 1 mmol/L MgSO4) for 4 hours at room temperature andstored at �20°C in methanol. The embryos were bleached with 30%H2O2 in methanol at 4°C for 1 hour and embedded in paraplast(Fisher HealthCare). Deparaffinized 15-�m-thick sections werebriefly rinsed in PBS, followed by 10-minute treatment with 5�L/mL acetic anhydride in 0.1 mol/L triethanolamine and 15-minutetreatment with 2 mg/mL glycine in PBS/1% Tween 20. The sectionswere prehybridized with hybridization solution24 at 70°C for 1 hourand hybridized with hybridization solution23 containing 0.5 �g/mLdigoxigenin-labeled riboprobe at 70°C for 16 hours. The sectionswere washed with 50% formamide, 2� SSC at 70°C for 2 hours andrinsed in PBS at room temperature. After blocking and reaction withalkaline phosphatase-conjugated anti-Dig antibody (1:2000 Boeh-ringer) at 4°C for 16 hours, color was developed using BM purple(Boehringer).
The cDNAs used for generation of Digoxigenin-labeled mouseriboprobes were ANF (nucleotides 173 to 744; GenBank no. NM_008725), �-MHC (nucleotides 7 to 237; GenBank no. NM_080728),BMP10 (nucleotides 86 to 1222; GenBank no. NM_009756), CHF1/Hey2,20 Cx40 (nucleotides 144 to 2127; GenBank no. NM_008121),GATA4 (nucleotides 1413 to 3272; GenBank no. NM_008092),mlc1a (nucleotides 46 to 339; GenBank no. NM_010858), mlc2a(nucleotides 65 to 529; GenBank no. NM_022879), mlc2v,25 Nkx2.5(nucleotides 231 to 1181; GenBank no. NM_008700), N-myc (nu-cleotides 525 to 1762; GenBank no. NM_008709), and Tbx5(nucleotides 360 to 1950; GenBank no. NM_011537). At least 3embryos were examined for each gene.
Cell ProliferationSections (5-�m thick) were stained immunohistochemically usingrabbit anti–phospho-histone H3 (Cell Signaling Technology) at 1:50dilution. Alkaline phosphatase-conjugated anti-rabbit IgG (Chemi-con) was used at 1:1000 dilution for the second antibody. The totalnumber of positive cells in the left ventricular compact myocardiumwas recorded, and then sections were counterstained with hematox-ylin to count total cell number, to derive the percentage of positivecells. We examined hearts from 2 embryos of each genotype andsampled 7 sections from each heart. Statistical comparison was doneby an unpaired Student’s t test.
ResultsCHF1/Hey2 Is Expressed in the Compact but NotTrabecular MyocardiumPrevious reports have demonstrated that CHF1/Hey2 is ex-pressed primarily in the developing ventricle through whole-mount and radioactive in situ hybridization.2–4,26 To examine
the expression pattern in pure C57BL/6 background mice indetail, we performed nonradioactive section in situ hybrid-ization for CHF1/Hey2 as previously described,2,20 withslight modifications. As shown in Figure 1, CHF1/Hey2 isexpressed predominantly in the compact myocardium andinterventricular septum at all time points examined, fromembryonic day 11.5 (E11.5) through E15.5. Expression isalso seen in the epithelial cells lining the endocardial cush-ions, but is not seen in the trabecular myocardium.
ANF Is Expressed Ectopically in the LeftVentricular Compact Myocardium of CHF1/Hey2Knockout HeartsWe and others have previously shown that CHF1/Hey2 caninhibit expression of the ANF promoter in vitro,27 duringdevelopment,26 and during development of cardiac hypertro-phy,21 through an interaction with the transcription factorGATA4. To assess for alteration in ANF expression in ourC57BL/6 CHF1/Hey2 knockout mice, we performed in situhybridization for ANF expression at E12.5 through E15.5. Asshown in Figure 2, ANF is normally expressed in the atria,trabeculae, and left ventricular compact myocardium atE12.5, but becomes restricted from the left ventricular com-pact myocardium beginning at E13.5, while persisting in theatrial and trabecular myocardium. In the CHF1/Hey2 knock-out embryos, however, ANF is expressed continuously in theleft ventricular compact myocardium after E13.5 throughE15.5. This pattern of ectopic expression is consistent with aprevious study,26 occurs in a domain where CHF1/Hey2 isnormally expressed, and is suggestive of derepression of ANFafter deletion of CHF1/Hey2. Derepression of ANF throughincreased GATA4 activity alone, however, cannot explain theectopic expression of ANF limited to the left ventricularcompact myocardium, as GATA4 and CHF1/Hey2 are bothexpressed in the left and the right ventricle (Figure 1 andFigure I in the online data supplement, available athttp://circres.ahajournals.org).
Tbx5, an Upstream Regulator of ANF, and ItsTarget Gene, Cx40, Are Also ExpressedEctopically in the Left Ventricular CompactMyocardium of the Knockout HeartsThe ANF promoter has been studied extensively in vitro andis positively regulated by the cardiac transcription factors
Figure 1. Expression pattern of CHF1/Hey2 during heart devel-opment. A, E11.5. B, E12.5. C, E13.5. D, E14.5. E, E15.5. Redarrowheads indicate the epithelial cells lining the endocardialcushions.
Koibuchi and Chin CHF1/Hey2 Suppresses Atrial Gene Expression 851
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from

GATA4, Nkx2.5, and Tbx5.28–30 To determine the potentialcontribution of these factors, we investigated their expressionpatterns by in situ hybridization. As shown in supplementalFigures I and II, the expression patterns of GATA4 and Nkx2.5are indistinguishable in wild-type and knockout embryos.Tbx5 has previously been reported to be expressed weakly inthe developing left ventricle at E13.5.31 As shown in Figure 3,we observe weak expression of Tbx5 in the developingventricle at E13.5; however, this expression is decreased byE15.5 in wild-type hearts. The expression pattern for Tbx5 inthe knockout embryos, however, is similar to that observedfor ANF, in that it is persistently expressed in the leftventricular compact myocardium through E15.5 (Figure 3).These findings suggest that ectopic expression of ANF resultsfrom persistent expression of Tbx5.
To verify that persistent expression of Tbx5 is sufficient topromote expression of downstream target genes, we assessedexpression of another Tbx5-dependent gene, connexin40(Cx40).29 Cx40 is normally expressed in the atrial andtrabecular myocardium and is also expressed in the vascula-ture at later stages in development. As shown in Figure 4,Cx40 is expressed normally in both wild-type and knockoutembryos at E12.5 but is expressed ectopically in the leftventricular compact myocardium of the CHF1/Hey2 knock-out embryos beginning at E13.5. The punctate pattern seen inboth the wild-type and knockout heart compact myocardiumbeginning at E14.5 reflects expression in the vasculature, asCx40 is known to be expressed in both the myocardium andthe vasculature at later stages of development.32 The myocar-dial expression pattern of Cx40 is similar to that of Tbx5 andANF in the knockout embryos. These findings indicate thatTbx5 is a downstream target of CHF1/Hey2 and that CHF1/Hey2 normally suppresses Tbx5 expression in the left ven-tricular compact myocardium, resulting in suppression ofANF and Cx40.
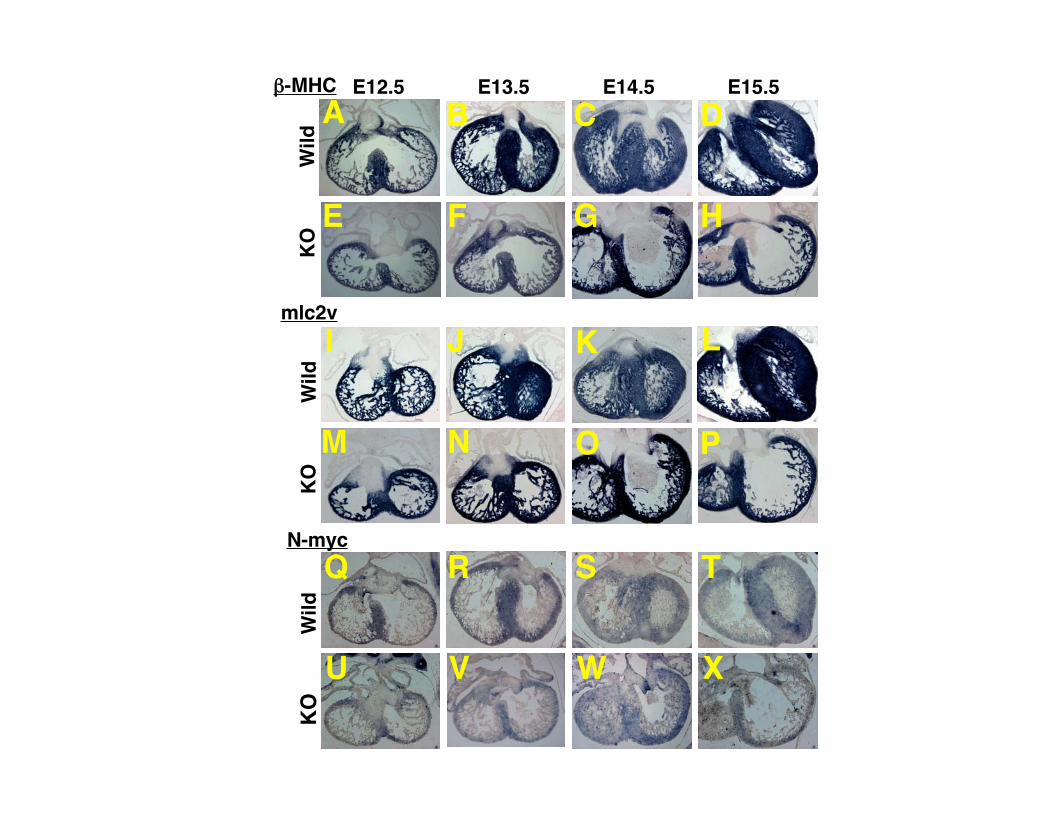
Atrial Contractile Markers mlc1a and mlc2a AreAlso Expressed Ectopically in the Left VentricularCompact Myocardium, Suggesting Perturbation inVentricular MaturationANF, Tbx5, and Cx40 are normally expressed in the atria andtrabeculae. Dysregulation of ANF, Tbx5, and Cx40 withectopic expression in the left ventricular compact myocardi-um suggested that the left ventricular compact myocardialcells might have atrial or trabecular cell characteristicsinstead of or in addition to ventricular characteristics. Toexplore these possibilities, we examined the expression pat-tern of atrial, ventricular and trabecular markers. We firstexamined the expression of the ventricular markers mlc2v,�-MHC, and N-myc, to determine whether ventricular cellidentity was altered in the compact myocardium. As shown insupplemental Figure III, there is no difference in regionalexpression of these markers in the knockout embryos, dem-onstrating that the left ventricular compact myocardial cellshave ventricular cell characteristics. Overall N-myc expres-sion may be decreased in the knockout ventricles, however,which may contribute in part to decreased myocardial prolif-eration, as described below.
We next examined the expression of the atrial and trabec-ular markers mlc1a and mlc2a. Neither is known to bedependent on Tbx5. As shown in Figure 5, both mlc1a andmlc2a show ectopic expression in the left ventricular compactmyocardium of the CHF1/Hey2 knockout embryos afterE13.5, providing further evidence that ventricular maturationis dysregulated and that the left ventricular myocardium hascharacteristics not only of ventricular cells but also of atrial ortrabecular cells.
Because ANF, Tbx5, Cx40, mlc1a, and mlc2a are alsoexpressed in the trabeculae, we considered the possibility thatthe cells ectopically expressing these genes may have trabec-ular rather than atrial characteristics. To assess this possibil-
Figure 2. Expression pattern of ANF during heartdevelopment. A through D, Hearts of wild-typemice. E through H, Hearts of CHF1/Hey2 knockoutmice. A and E, E12.5. B and F, E13.5. C and G,E14.5. D and H, E15.5. Red arrowheads indicateectopic expression of ANF in the left compactmyocardium. RA indicates right atrium; LA, leftatrium; RV, right ventricle; LV, left ventricle.
Figure 3. Expression pattern of Tbx5 during heartdevelopment. A through D, Hearts of wild-typemice. E through H, Hearts of CHF1/Hey2 knockoutmice. A and E, E12.5. B and F, E13.5. C and G,E14.5. D and H, E15.5. Red arrowheads indicateectopic expression of Tbx5 in the left compactmyocardium.
852 Circulation Research March 30, 2007
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from

ity, we examined expression of the trabecular marker BMP10.BMP10 is expressed predominantly in the trabecular myocar-dium, and mice lacking BMP10 demonstrate decreased tra-beculation and a thin-walled myocardium associated withdecreased myocyte proliferation.33 As shown in Figure 6,BMP10 is expressed normally in both wild-type and knockoutembryos, suggesting that the knockout left ventricular com-pact myocardial cells are more atrial in character thantrabecular. Overall, these findings suggest that the compactmyocardial cells have both atrial and ventricular characteris-tics, which is reminiscent of undifferentiated cardiac myo-cytes. These findings also suggest a role for CHF1/Hey2 inpromoting proper myocardial maturation beyond E13.5.
A CHF1/Hey2 Transgene Expressed Diffusely inthe Myocardium Suppresses Ectopic, but NotNormal, Expression of Atrial GenesWe have previously reported that an mlc2v-driven CHF1/Hey2 transgene expressed diffusely in all 4 cardiac chamberscan rescue the myocardial and partially rescue the valvularand septation phenotypes in the knockout mice and improvesurvival.20 To assess whether this diffusely expressed trans-
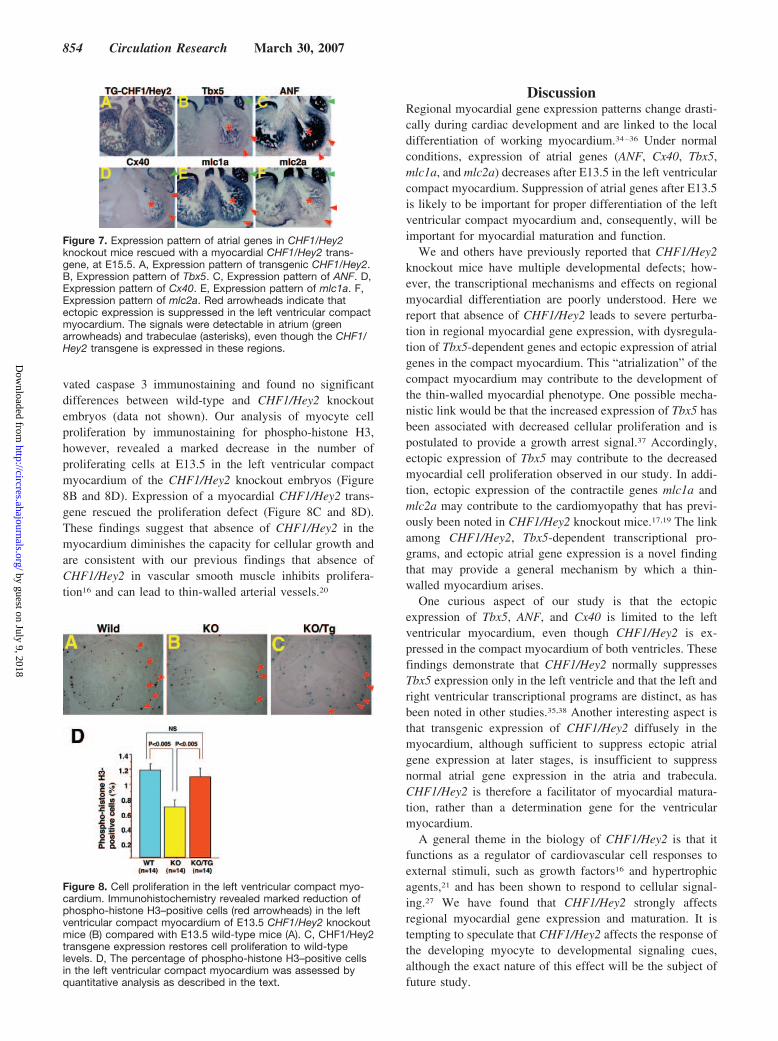
gene can suppress ectopic gene expression, we performed insitu hybridization on CHF1/Hey2 knockout embryos carryingthis transgene at E12.5 and E15.5. As shown in Figure 7 andsupplemental Figure IV, the transgene is expressed diffuselyin all 4 chambers and in both the trabecular and compactmyocardium of the ventricles. Knockout embryos carryingthe transgene did not express ANF, Tbx5, Cx40, mlc1a, ormlc2a in the compact myocardium at E15.5, demonstratingthat CHF1/Hey2 does indeed rescue gene expression patternsby suppressing ectopic expression. Interestingly, expressionof the CHF1/Hey2 transgene in the trabecular myocardiumand atria was not sufficient to suppress the normal expressionof these genes in the atria and trabeculae, or suppressexpression at E12.5. Our findings suggest that CHF1/Hey2 islikely a facilitator of normal ventricular maturation, but is notsufficient to suppress atrial gene expression and act as adetermination gene.
Myocyte Proliferation Is Decreased inCHF1/Hey2-Null EmbryosTo investigate the mechanistic basis for the observed thin-walled myocardium, we hypothesized that the thin myocar-dium may result from either increased myocyte apoptosis orfrom decreased myocyte proliferation. We analyzed apoptosisat various developmental stages by both TUNEL and acti-
Figure 6. Expression pattern of BMP10 during heart develop-ment. A through D and I, Hearts of wild-type mice. E through H,Hearts of CHF1/Hey2 knockout mice. A and E, E12.5. B and F,E13.5. C and G, E14.5. D, H, and I, E15.5. I, Low magnification.The signal of BMP10 was detectable in atrial and ventricular tra-beculae and right atrial compact myocardium but not ventricularand left atrial compact myocardium. No marked differenceswere observed between CHF1/Hey2 knockout and wild-typemice. RA indicates right atrium; LA, left atrium; RV, right ventri-cle; LV, left ventricle.
Figure 4. Expression pattern of Cx40 during heartdevelopment. A through D, Hearts of wild-typemice. E through H, Hearts of CHF1/Hey2 knockoutmice. A and E, E12.5. B and F, E13.5. C and G,E14.5. D and H, E15.5. Green arrowheads indicatevasculature. Red arrowheads indicate ectopicexpression of Cx40 in the left compactmyocardium.
Figure 5. Expression patterns of mlc1a and mlc2a during heartdevelopment. A through H, Expression pattern of mlc1a. Ithrough P, Expression pattern of mlc2a. A through D and Ithrough L, Hearts of wild-type mice. E through H and M throughP, Hearts of CHF1/Hey2 knockout mice. A, E, I, and M, E12.5.B, F, J, and N, E13.5. C, G, K, and O, E14.5. D, H, L, and P,E15.5. Red arrowheads indicate ectopic expression of mlc1aand mlc2a in the left compact myocardium.
Koibuchi and Chin CHF1/Hey2 Suppresses Atrial Gene Expression 853
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from
vated caspase 3 immunostaining and found no significantdifferences between wild-type and CHF1/Hey2 knockoutembryos (data not shown). Our analysis of myocyte cellproliferation by immunostaining for phospho-histone H3,however, revealed a marked decrease in the number ofproliferating cells at E13.5 in the left ventricular compactmyocardium of the CHF1/Hey2 knockout embryos (Figure8B and 8D). Expression of a myocardial CHF1/Hey2 trans-gene rescued the proliferation defect (Figure 8C and 8D).These findings suggest that absence of CHF1/Hey2 in themyocardium diminishes the capacity for cellular growth andare consistent with our previous findings that absence ofCHF1/Hey2 in vascular smooth muscle inhibits prolifera-tion16 and can lead to thin-walled arterial vessels.20
DiscussionRegional myocardial gene expression patterns change drasti-cally during cardiac development and are linked to the localdifferentiation of working myocardium.34–36 Under normalconditions, expression of atrial genes (ANF, Cx40, Tbx5,mlc1a, and mlc2a) decreases after E13.5 in the left ventricularcompact myocardium. Suppression of atrial genes after E13.5is likely to be important for proper differentiation of the leftventricular compact myocardium and, consequently, will beimportant for myocardial maturation and function.
We and others have previously reported that CHF1/Hey2knockout mice have multiple developmental defects; how-ever, the transcriptional mechanisms and effects on regionalmyocardial differentiation are poorly understood. Here wereport that absence of CHF1/Hey2 leads to severe perturba-tion in regional myocardial gene expression, with dysregula-tion of Tbx5-dependent genes and ectopic expression of atrialgenes in the compact myocardium. This “atrialization” of thecompact myocardium may contribute to the development ofthe thin-walled myocardial phenotype. One possible mecha-nistic link would be that the increased expression of Tbx5 hasbeen associated with decreased cellular proliferation and ispostulated to provide a growth arrest signal.37 Accordingly,ectopic expression of Tbx5 may contribute to the decreasedmyocardial cell proliferation observed in our study. In addi-tion, ectopic expression of the contractile genes mlc1a andmlc2a may contribute to the cardiomyopathy that has previ-ously been noted in CHF1/Hey2 knockout mice.17,19 The linkamong CHF1/Hey2, Tbx5-dependent transcriptional pro-grams, and ectopic atrial gene expression is a novel findingthat may provide a general mechanism by which a thin-walled myocardium arises.
One curious aspect of our study is that the ectopicexpression of Tbx5, ANF, and Cx40 is limited to the leftventricular myocardium, even though CHF1/Hey2 is ex-pressed in the compact myocardium of both ventricles. Thesefindings demonstrate that CHF1/Hey2 normally suppressesTbx5 expression only in the left ventricle and that the left andright ventricular transcriptional programs are distinct, as hasbeen noted in other studies.35,38 Another interesting aspect isthat transgenic expression of CHF1/Hey2 diffusely in themyocardium, although sufficient to suppress ectopic atrialgene expression at later stages, is insufficient to suppressnormal atrial gene expression in the atria and trabecula.CHF1/Hey2 is therefore a facilitator of myocardial matura-tion, rather than a determination gene for the ventricularmyocardium.
A general theme in the biology of CHF1/Hey2 is that itfunctions as a regulator of cardiovascular cell responses toexternal stimuli, such as growth factors16 and hypertrophicagents,21 and has been shown to respond to cellular signal-ing.27 We have found that CHF1/Hey2 strongly affectsregional myocardial gene expression and maturation. It istempting to speculate that CHF1/Hey2 affects the response ofthe developing myocyte to developmental signaling cues,although the exact nature of this effect will be the subject offuture study.
Figure 7. Expression pattern of atrial genes in CHF1/Hey2knockout mice rescued with a myocardial CHF1/Hey2 trans-gene, at E15.5. A, Expression pattern of transgenic CHF1/Hey2.B, Expression pattern of Tbx5. C, Expression pattern of ANF. D,Expression pattern of Cx40. E, Expression pattern of mlc1a. F,Expression pattern of mlc2a. Red arrowheads indicate thatectopic expression is suppressed in the left ventricular compactmyocardium. The signals were detectable in atrium (greenarrowheads) and trabeculae (asterisks), even though the CHF1/Hey2 transgene is expressed in these regions.
Figure 8. Cell proliferation in the left ventricular compact myo-cardium. Immunohistochemistry revealed marked reduction ofphospho-histone H3–positive cells (red arrowheads) in the leftventricular compact myocardium of E13.5 CHF1/Hey2 knockoutmice (B) compared with E13.5 wild-type mice (A). C, CHF1/Hey2transgene expression restores cell proliferation to wild-typelevels. D, The percentage of phospho-histone H3–positive cellsin the left ventricular compact myocardium was assessed byquantitative analysis as described in the text.
854 Circulation Research March 30, 2007
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from

Sources of FundingThis work was supported by NIH grants HL076232 and HL081088(to M.T.C.) and a Japan Heart Foundation fellowship (to N.K.).
DisclosuresNone.
References1. Kokubo H, Lun Y, Johnson RL. Identification and expression of a novel
family of bHLH cDNAs related to drosophila hairy and enhancer of split.Biochem Biophys Res Commun. 1999;260:459–465.
2. Chin MT, Maemura K, Fukumoto S, Jain MK, Layne MD, Watanabe M,Hsieh C-M, Lee M-E. Cardiovascular basic helix loop helix factor 1, anovel transcriptional repressor expressed preferentially in the developingand adult cardiovascular system. J Biol Chem. 2000;275:6381–6387.
3. Nakagawa O, Nakagawa M, Richardson JA, Olson EN, Srivastava D.HRT1, HRT2, and HRT3: a new subclass of bHLH transcription factorsmarking specific cardiac, somitic, and pharyngeal arch segments. DevBiol. 1999;216:72–84.
4. Leimeister C, Externbrink A, Klamt B, Gessler M. Hey genes: a novelsubfamily of hairy- and Enhancer of split related genes specificallyexpressed during mouse embryogenesis. Mech Dev. 1999;85:173–177.
5. Zhong TP, Rosenberg M, Mohideen M-APK, Weinstein B, Fishman MC.gridlock, an HLH gene required for assembly of the aorta in zebrafish.Science. 2000;287:1820–1824.
6. Iso T, Sartorelli V, Chung G, Shichinohe T, Kedes L, Hamamori Y.HERP, a new primary target of Notch regulated by ligand binding. MolCell Biol. 2001;21:6071–6079.
7. Zhong TP, Childs S, Leu JP, Fishman MC. Gridlock signalling pathwayfashions the first embryonic artery. Nature. 2001;414:216–220.
8. Nakagawa O, McFadden DG, Nakagawa M, Yanagisawa H, Hu TH,Srivastava D, Olson EN. Members of the HRT family of basic helix-loop-helix proteins act as transcriptional repressors downstream of Notchsignaling. Proc Natl Acad Sci U S A. 2000;97:13655–13660.
9. Wang W, Campos AH, Prince CZ, Mou Y, Pollman MJ. CoordinateNotch3-hairy-related transcription factor pathway regulation in responseto arterial injury. Mediator role of platelet-derived growth factor andERK. J Biol Chem. 2002;277:23165–23171.
10. Henderson AM, Wang SJ, Taylor AC, Aitkenhead M, Hughes CC. Thebasic helix-loop-helix transcription factor HESR1 regulates endothelialcell tube formation. J Biol Chem. 2001;276:6169–6176.
11. Fischer A, Schumacher N, Maier M, Sendtner M, Gessler M. The Notchtarget genes Hey1 and Hey2 are required for embryonic vascular devel-opment. Genes Dev. 2004;18:901–911.
12. Sun JX, Kamei CN, Layne MD, Jain MK, Liao JK, Lee ME, Chin MT.Regulation of myogenic terminal differentiation by the hairy-related tran-scription factor CHF2. J Biol Chem. 2001;276:18591–18596.
13. Iso T, Sartorelli V, Poizat C, Iezzi S, Wu HY, Chung G, Kedes L,Hamamori Y. HERP, a novel heterodimer partner of HES/E(spl) in Notchsignaling. Mol Cell Biol. 2001;21:6080–6089.
14. Campos AH, Wang W, Pollman MJ, Gibbons GH. Determinants ofNotch-3 receptor expression and signaling in vascular smooth musclecells: implications in cell-cycle regulation. Circ Res. 2002;91:999–1006.
15. Wang W, Prince CZ, Hu X, Pollman MJ. HRT1 modulates vascularsmooth muscle cell proliferation and apoptosis. Biochem Biophys ResCommun. 2003;308:596–601.
16. Sakata Y, Xiang F, Chen Z, Kiriyama Y, Kamei CN, Simon DI, Chin MT.Transcription factor CHF1/Hey2 regulates neointimal formation in vivoand vascular smooth muscle proliferation and migration in vitro. Arte-rioscler Thromb Vasc Biol. 2004;24:2069–2074.
17. Gessler M, Knobeloch K-P, Helisch A, Amann K, Schumacher N, RohdeE, Fischer A, Leimeister C. Mouse gridlock: no aortic coarctation ordeficiency, but fatal cardiac defects in Hey2 -/- mice. Curr Biol. 2002;12:1601–1604.
18. Donovan J, Kordylewska A, Jan YN, Utset MF. Tetralogy of Fallot andother congenital heart defects in Hey2 mutant mice. Curr Biol. 2002;12:1605–1610.
19. Sakata Y, Kamei CN, Nakagami H, Bronson R, Liao JK, Chin MT.Ventricular septal defect and cardiomyopathy in mice lacking the tran-
scription factor CHF1/Hey2. Proc Natl Acad Sci U S A. 2002;99:16197–16202.
20. Sakata Y, Koibuchi N, Xiang F, Youngblood JM, Kamei CN, Chin MT.The spectrum of cardiovascular anomalies in CHF1/Hey2 deficient micereveals roles in endocardial cushion, myocardial and vascular maturation.J Mol Cell Cardiol. 2006;40:267–273.
21. Xiang F, Sakata Y, Cui L, Youngblood JM, Nakagami H, Liao JK, LiaoR, Chin MT. Transcription factor CHF1/Hey2 suppresses cardiac hyper-trophy through an inhibitory interaction with GATA4. Am J PhysiolHeart Circ Physiol. 2006;290:H1997–H2006.
22. Kokubo H, Miyagawa-Tomita S, Tomimatsu H, Nakashima Y, NakazawaM, Saga Y, Johnson RL. Targeted disruption of hesr2 results in atrioven-tricular valve anomalies that lead to heart dysfunction. Circ Res. 2004;95:540–547.
23. Harland RM. In situ hybridization: an improved whole-mount method forXenopus embryos. Methods Cell Biol. 1991;36:685–695.
24. Shain DH, Zuber MX. Sodium dodecyl sulfate (SDS)-based whole-mountin situ hybridization of Xenopus laevis embryos. J Biochem BiophysMethods. 1996;31:185–188.
25. O’Brien TX, Lee KJ, Chien KR. Positional specification of ventricularmyosin light chain 2 expression in the primitive murine heart tube. ProcNatl Acad Sci U S A. 1993;90:5157–5161.
26. Fischer A, Klattig J, Kneitz B, Diez H, Maier M, Holtmann B, Englert C,Gessler M. Hey basic helix-loop-helix transcription factors are repressorsof GATA4 and GATA6 and restrict expression of the GATA target geneANF in fetal hearts. Mol Cell Biol. 2005;25:8960–8970.
27. Kathiriya IS, King IN, Murakami M, Nakagawa M, Astle JM, GardnerKA, Gerard RD, Olson EN, Srivastava D, Nakagawa O. Hairy-relatedtranscription factors inhibit GATA-dependent cardiac gene expressionthrough a signal-responsive mechanism. J Biol Chem. 2004;279:54937–54943.
28. Durocher D, Charron F, Warren R, Schwartz RJ, Nemer M. The cardiactranscription factors Nkx2-5 and GATA-4 are mutual cofactors. EMBO J.1997;16:5687–5696.
29. Bruneau BG, Nemer G, Schmitt JP, Charron F, Robitaille L, Caron S,Conner DA, Gessler M, Nemer M, Seidman CE, Seidman JG. A murinemodel of Holt-Oram syndrome defines roles of the T-box transcriptionfactor Tbx5 in cardiogenesis and disease. Cell. 2001;106:709–721.
30. Hiroi Y, Kudoh S, Monzen K, Ikeda Y, Yazaki Y, Nagai R, Komuro I.Tbx5 associates with Nkx2-5 and synergistically promotes cardiomyocytedifferentiation. Nat Genet. 2001;28:276–280.
31. Bruneau BG, Logan M, Davis N, Levi T, Tabin CJ, Seidman JG, SeidmanCE. Chamber-specific cardiac expression of Tbx5 and heart defects inHolt-Oram syndrome. Dev Biol. 1999;211:100–108.
32. Delorme B, Dahl E, Jarry-Guichard T, Briand JP, Willecke K, Gros D,Theveniau-Ruissy M. Expression pattern of connexin gene products at theearly developmental stages of the mouse cardiovascular system. Circ Res.1997;81:423–437.
33. Chen H, Shi S, Acosta L, Li W, Lu J, Bao S, Chen Z, Yang Z, SchneiderMD, Chien KR, Conway SJ, Yoder MC, Haneline LS, Franco D, Shou W.BMP10 is essential for maintaining cardiac growth during murine car-diogenesis. Development. 2004;131:2219–2231.
34. Lyons GE, Schiaffino S, Sassoon D, Barton P, Buckingham M. Devel-opmental regulation of myosin gene expression in mouse cardiac muscle.J Cell Biol. 1990;111:2427–2436.
35. Zammit PS, Kelly RG, Franco D, Brown N, Moorman AF, BuckinghamME. Suppression of atrial myosin gene expression occurs independentlyin the left and right ventricles of the developing mouse heart. Dev Dyn.2000;217:75–85.
36. Christoffels VM, Habets PE, Franco D, Campione M, de Jong F, LamersWH, Bao ZZ, Palmer S, Biben C, Harvey RP, Moorman AF. Chamberformation and morphogenesis in the developing mammalian heart. DevBiol. 2000;223:266–278.
37. Hatcher CJ, Kim MS, Mah CS, Goldstein MM, Wong B, Mikawa T,Basson CT. TBX5 transcription factor regulates cell proliferation duringcardiogenesis. Dev Biol. 2001;230:177–188.
38. Kelly RG, Zammit PS, Mouly V, Butler-Browne G, Buckingham ME.Dynamic left/right regionalisation of endogenous myosin light chain 3Ftranscripts in the developing mouse heart. J Mol Cell Cardiol. 1998;30:1067–1081.
Koibuchi and Chin CHF1/Hey2 Suppresses Atrial Gene Expression 855
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from

Nobutaka Koibuchi and Michael T. ChinEctopic Atrial Gene Expression
CHF1/Hey2 Plays a Pivotal Role in Left Ventricular Maturation Through Suppression of
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2007 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000261693.13269.bf2007;100:850-855; originally published online March 1, 2007;Circ Res.
http://circres.ahajournals.org/content/100/6/850World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2007/03/02/01.RES.0000261693.13269.bf.DC1Data Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 9, 2018http://circres.ahajournals.org/
Dow
nloaded from

Online Supplement Koibuchi and Chin
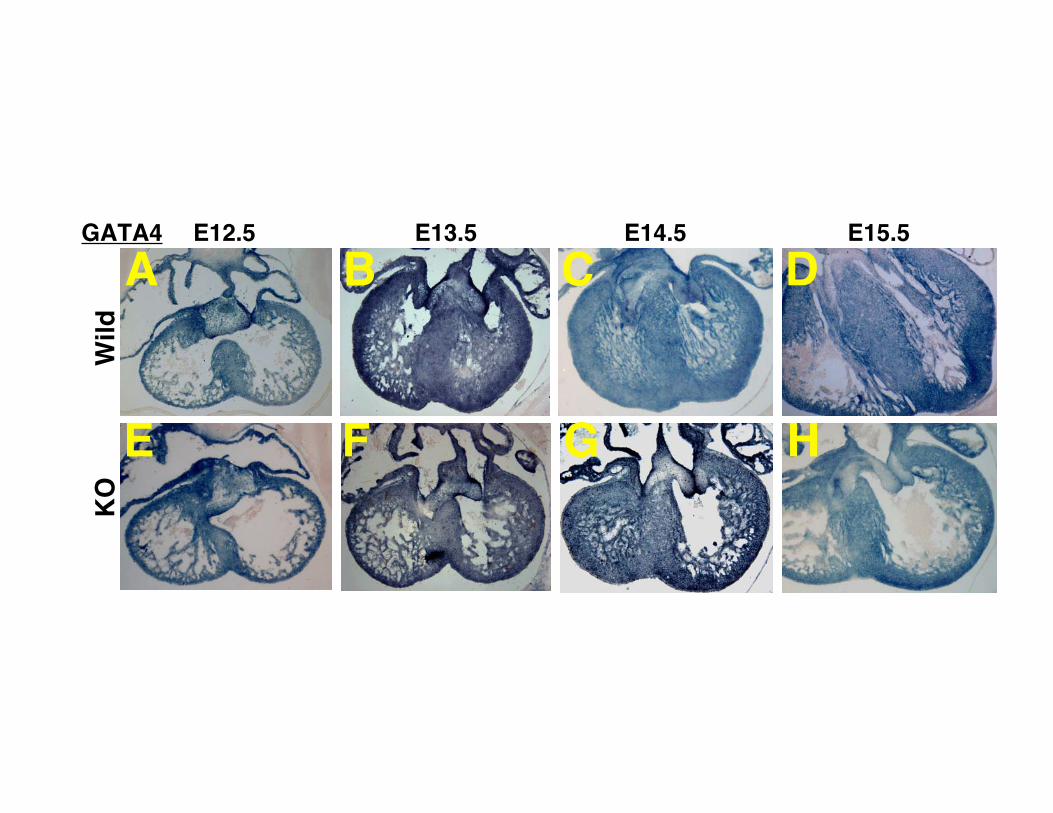
Supplemental Figure I. Expression pattern of GATA4 during heart development. (A-D) The heart of wild type mice. (E-H) The heart of CHF1/Hey2 knockout mice. (A, E) E12.5. (B, F) E13.5. (C, G) E14.5. (D, H) E15.5. No marked differences were observed between CHF1/Hey2 KO and wild type mice. Supplemental Figure II. Expression pattern of Nkx2.5 during heart development. (A-D) The heart of wild type mice. (E-H) The heart of CHF1/Hey2 knockout mice. (A, E) E12.5. (B, F) E13.5. (C, G) E14.5. (D, H) E15.5. No marked differences were observed between CHF1/Hey2 KO and wild type mice. Supplemental Figure III. Expression pattern of β-MHC, mlc2v and N-myc during heart development. (A-H) Expression pattern of β-MHC. (I-P) Expression pattern of mlc2v. (Q-X) Expression pattern of N-myc. (A-D, I-L and Q-T) The heart of wild type mice. (E-H, M-P and U-X) The heart of CHF1/Hey2 knockout mice. (A, E, I, M, Q and U) E12.5. (B, F, J, N, R and V) E13.5. (C, G, K, O, S and W) E14.5. (D, H, L, P, T and X) E15.5. No marked differences in regional expression patterns were observed between CHF1/Hey2 KO and wild type mice. Supplemental Figure IV. Expression pattern of atrial genes in CHF1/Hey2 knockout mice rescued with a myocardial CHF1/Hey2 transgene, at E12.5. (A) Expression pattern of transgenic CHF1/Hey2. (B) Expression pattern of Tbx5. (C) Expression pattern of ANF (D) Expression pattern of Cx40. (E) Expression pattern of mlc1a. (F) Expression pattern of mlc2a. Red arrowheads indicate that expression is present in the left ventricular compact myocardium.
B C D
E G H
A
F
KO
Wild
E13.5 E14.5 E15.5GATA4 E12.5
michelle
Text Box
Supplemental Figure I

B C D
E G H
A
F
KO
Wild
E12.5 E13.5 E14.5 E15.5Nkx2-5
michelle
Text Box
Supplemental Figure II
KO
Wild
E12.5 E13.5 E14.5 E15.5
N-myc
mlc2v
-MHC
KO
Wild
KO
Wild
B C D
E G H
A
F
J K LI
N O PM
R S TQ
V W XU
michelle
Text Box
Supplemental Figure III
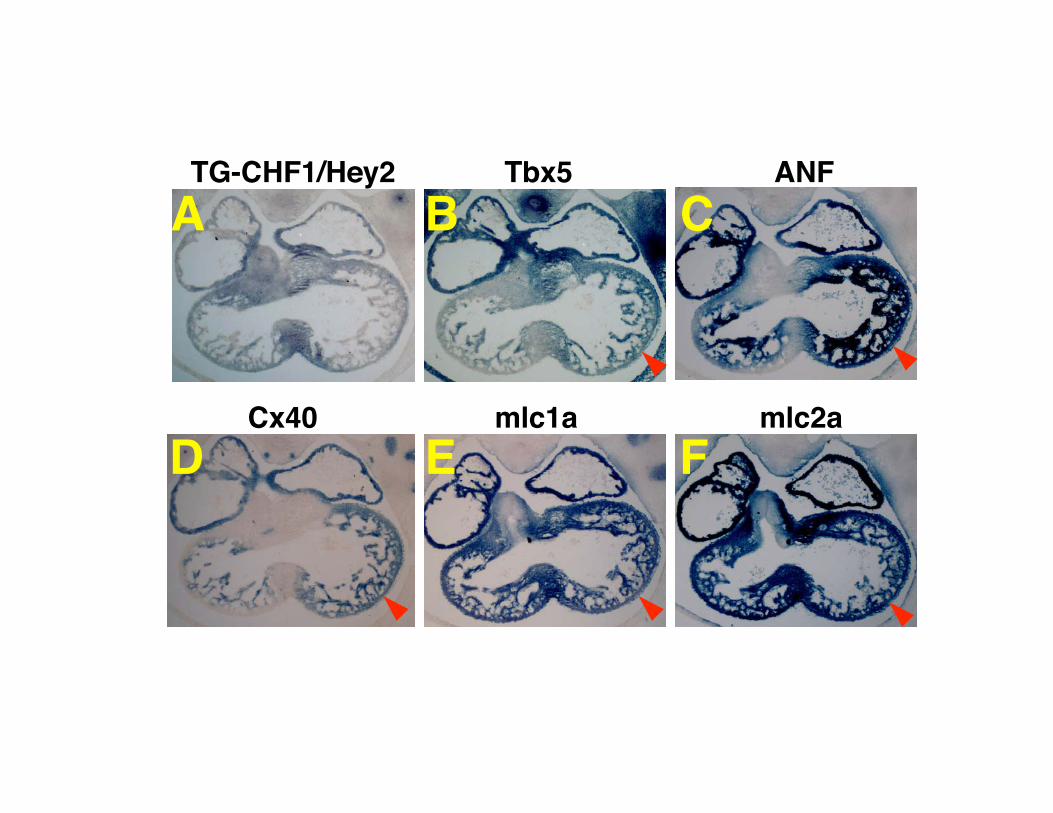
TG-CHF1/Hey2 ANF
Cx40 mlc1a mlc2a
Tbx5
B C
D E F
A
michelle
Text Box
Supplemental Figure IV


















