
Chemokine IL-8, and RANTES in MERS-CoV-Infected
32
Page 1/32 Elevated Expression Levels of Lung Complement Anaphylatoxin, Neutrophil Chemoattractant Chemokine IL-8, and RANTES in MERS-CoV-Infected Patients: Predictive Biomarkers for Disease Severity and Mortality Maaweya Awadalla King Saud University https://orcid.org/0000-0002-1270-2216 Asif Naeem King Fahad Medical City Haitham Alkadi King Fahad Medical City Aref A. Alamri Ministry of Health Ahmad S. AlYami King Fahad Medical City Abdullah AlJuryyan King Fahad Medical City Wael Alturaiki Majmaah University Mushira Enani King Fahad Medical City Samia Towfeek Al-Shouli King Saud University Abdullah M. Assiri Ministry of Health Bandar Alosaimi ( [email protected] ) King Fahad Medical City Research Article Keywords: MERS-CoV infection, complement system, immunopathology, lung, cytokine, chemokine Posted Date: March 15th, 2021
Transcript of Chemokine IL-8, and RANTES in MERS-CoV-Infected

Page 1/32
Elevated Expression Levels of Lung ComplementAnaphylatoxin, Neutrophil ChemoattractantChemokine IL-8, and RANTES in MERS-CoV-InfectedPatients: Predictive Biomarkers for Disease Severityand MortalityMaaweya Awadalla
King Saud University https://orcid.org/0000-0002-1270-2216Asif Naeem
King Fahad Medical CityHaitham Alkadi
King Fahad Medical CityAref A. Alamri
Ministry of HealthAhmad S. AlYami
King Fahad Medical CityAbdullah AlJuryyan
King Fahad Medical CityWael Alturaiki
Majmaah UniversityMushira Enani
King Fahad Medical CitySamia Towfeek Al-Shouli
King Saud UniversityAbdullah M. Assiri
Ministry of HealthBandar Alosaimi ( [email protected] )
King Fahad Medical City
Research Article
Keywords: MERS-CoV infection, complement system, immunopathology, lung, cytokine, chemokine
Posted Date: March 15th, 2021
Page 2/32
DOI: https://doi.org/10.21203/rs.3.rs-285036/v1
License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License
Version of Record: A version of this preprint was published at Journal of Clinical Immunology on July 7th,2021. See the published version at https://doi.org/10.1007/s10875-021-01061-z.

Page 3/32
AbstractThe complement system represents an innate immune response consisting of a protein network. Over-activation of the complement system plays an important role in in�ammation, tissue damage, andinfectious disease severity. The prevalence of MERS-CoV in Saudi Arabia remains signi�cant and casesare still being reported. The role of complement in Middle East Respiratory Syndrome coronavirus (MERS-CoV) pathogenesis and complement‐modulating treatment strategies has received limited attention, andstudies involving MERS-CoV-infected patients have not been reported. This study offers the �rst insightinto the pulmonary expression pro�le including Seven complement proteins including complementregulatory factors during MERS-CoV infection. We also measured the expression of lung neutrophilchemoattractant chemokine IL-8 (CXCL8) and RANTES (CCL5). Our results signi�cantly indicate highexpression levels of complement anaphylatoxins (C3a and C5a), IL-8, and RANTES in the lungs of MERS-CoV-infected patients. The upregulation of lung complement anaphylatoxins, C5a and C3a, waspositively correlated with IL-8, RANTES and the fatality rate. Our results also showed upregulation of thepositive regulatory complement factor P (properdin), suggesting positive regulation of the complementduring MERS-CoV infection. In addition, we also demonstrated that a high viral load in all patients withMERS-CoV correlated with C5a and C3a levels. Pulmonary complement mediators, disease severity, andan increased fatality rate may be linked to the degree of complement activation against MERS-CoV. Highlevels of lung C5a, C3a, factor P, IL-8 and RANTES may contribute to the immunopathology, diseaseseverity, ARDS development, and a higher fatality rate in MERS-CoV-infected patients. These �ndingshighlight the potential prognostic utility of C5a, C3a, IL-8 and RANTES as biomarkers for MERS-CoVdisease severity and mortality. To further explore the functional partners (protiens) prediction of highlyexpressed proteins (C5a, C3a, factor P, IL-8 and RANTES), the computational protein–protein interaction(PPI) network was constructed, and six proteins (hub nodes) were identi�ed.
IntroductionThe COVID-19 pandemic caused by severe respiratory syndrome coronavirus 2 (SARS-CoV-2) has gainedsigni�cant attention in the medical and scienti�c communities. The Middle East Respiratory Syndromecoronavirus (MERS-CoV) has caused considerable medical and health issues in many countries,particularly in Saudi Arabia. The prevalence of MERS-CoV in the Kingdom of Saudi Arabia (KSA) remainssigni�cant. MERS-CoV cases are still being reported in Saudi Arabia, and a high prevalence of MERS-CoVin dromedary camels and direct contact with infected camels have been linked to human infections(Alosaimi et al., 2020; Assiri et al., 2016; Azhar et al., 2014; Rabaan et al., 2021; WHO, 2021). MERS-CoV isa single-standard RNA virus of the Betacoronavirus genus. It was �rst reported in the KSA (Jeddah City) in2012. As of December 27, 2020, 2564 laboratory-confirmed cases and 881 associated deaths (case-fatality ratio, 34.4%) were reported in 27 countries worldwide, of which 2121 cases (82.7%) were reportedin Saudi Arabia. The majority of the fatalities (37.1%, 788 deaths) also occurred in Saudi Arabia (WHO,2021). An excessive in�ammatory response is a prominent phenotype associated with MERS-CoVinfection, which leads to lung immunopathology, disease progression, and poor clinical outcome. MERS-

Page 4/32
CoV infections are characterized by dysregulation in both the innate and adaptive immune systems [5],[6]. Several in�ammatory cytokines and chemokines (IL-1β, IL-2, IL-6, IL-7, IL-8, IL-10, G-CSF, GM-CSF, IP-10, MCP-1, MIP-1α, IFN-γ, TNF-α, CCL2, and CCL3) are signi�cantly associated with severe MERS-CoVinfection and higher fatality rates [5], [7], [8]. Elevated in�ammatory cytokine and chemokine levels duringSARS‐CoV-1, MERS‐CoV, and SARS‐CoV‐2 infections are signi�cantly associated with massive in�ltrationof immune cells into the lungs and poor disease outcome [5], [8]–[10]. The complement system consistsof a multiprotein network belonging to both the innate and adoptive immune systems [11]. Depending onthe manner of activation, the complement cascade operates by three pathways: the classical pathway,the lectin pathway, and the alternative pathway [12], [13]. Cross talk between the complement andcoagulation systems plays a crucial role in vascular endothelial damage and thromboin�ammation [14].A number of viral infections are associated with complement activation and coagulation dysfunction [11],[15], [16]. Over-activation of pulmonary and systemic complement plays a key role in in�ammation,endothelial cell damage, thrombus formation, and intravascular coagulation, which results in multipleorgan failure and eventually death [12], [15], [17]. This over-stimulation leads to the formation of thecomplement anaphylatoxins, C3a and C5a. C5a is a chemoattractant for neutrophils, monocytes,eosinophils, and T cells [15], [18]. Following infection, complement anaphylatoxins stimulate phagocyticcells and enhance the production of TNF-α, IL‐1β, IL‐6, IL‐8, granular enzymes, and free radicals. Thesemediators promote vascular dysfunction, �brinolysis, and microvascular thrombosis formation [11], [12],[14], [15], [17], [18]. C5a also plays an important role in stimulating the expression of P‐selectin,intercellular adhesion molecule‐1, �bronectin, and �brinogen. The upregulation of adhesion moleculesactivates different cell signaling and pro‐in�ammatory pathways [15], [16].
The role of complement in MERS-CoV disease immunopathology and complement‐modulating treatmentstrategies during MERS-CoV infection has received limited attention. There are many importantunanswered questions regarding complement, MERS-CoV interactions, and disease outcome. In addition,little is known regarding pulmonary complement activation during MERS-CoV infection, the manner inwhich complement activation affects disease severity, or the association of complement response withviral load and mortality.
In this study, we performed a comprehensive investigation of the pulmonary complement proteins, IL-8(CXCL8) and CCL5 (RANTES) expression in MERS-CoV-infected patients in addition to viral loaddetermination. We also assessed the correlation between these factors and the fatality rate. To ourknowledge, this is the �rst study demonstrating a relationship between lung complement proteins andcomplement regulatory factors in MERS-CoV-infected patients.
Materials And MethodsPatient Selection, Sample Collection, and Preparation and Analysis
A total of 31 MERS-CoV-positive patients and 15 healthy matched controls were enrolled in this study.Lower respiratory samples (bronchoalveolar lavage (BAL) or tracheal aspirate (TA)) were collected. To

Page 5/32
exclude the effects of antiviral therapy on the expression of complement proteins, IL-8 (CXCL8) and CCL5(RANTES), all samples were collected before the administration of any antiviral treatment. Samples werecentrifuged at 1000 rpm at 4°C for 5–10 min. The cell-free supernatants were used for the analysis ofcomplement proteins, in�ammatory chemokines, RANTES (CCL5), and MERS-CoV viral load. Theexclusion criteria were as follows: 1) patients coinfected with other respiratory pathogens, 2)immunocompromised patients, 3) patients under treatment with anti-in�ammatory and/orimmunosuppressive drugs, and 4) patients with preexisting autoimmune disease. These exclusion criteriawere selected to exclude any possible effects on the expression of the clinicopathological factors listedabove. This study was reviewed and approved by the Institutional Review Board at King Fahad MedicalCity (IRB register number 019-053).
Measurement of Complement In�ammatory MediatorsAnaphylatoxins (C3a/C3b and C5a) and Classical PathwayComplement Component (C1q) and C2 LevelsPulmonary levels of human complement C3 and C5 fragments, as well as C3b, C1q, and C2, weremeasured using ELISA kits (HCA39-K01-Eagle Biosciences, Inc., Columbia, USA; ab193695, Abcam,Cambridge, UK; ab195461, Abcam, Cambridge, UK; ab170246, Abcam, Cambridge, UK; ab154132, Abcam,Cambridge, UK) following the manufacturer’s instructions. The concentrations were calculated usingstandard curves.
Quantitation of Serum Regulatory Complement Component (Factor) Levels
The concentrations of four human complement regulatory factors, factors P (properdin), I, C4-bindingprotein (C4-BP), and H, were quanti�ed in the lung using ELISA kits (ab222864, Abcam, Cambridge, UK;ab195460, Abcam, Cambridge, UK; ab222866, Abcam, Cambridge, UK; HK342 Hycult Biotech, Uden,Netherlands). The ELISAs were done following the manufacturer’s instructions. The concentrations ofeach factor were calculated using standard curves.
Quanti�cation of Serum Chemokine RANTES (CCL5) LevelsWe determined the local RANTES chemokine levels in MERS-CoV-infected patients (n = 30) and healthyindividuals (n = 18). Lung RANTES levels were quanti�ed using the human RANTES ELISA Kit (R&DSystems, Minneapolis, USA) following the manufacturer’s protocol. RANTES concentrations werecalculated using standard curves, and the results were expressed as pg/ml.
Measurement of Pulmonary Pro-In�ammatory Cytokine andChemokine Pro�les Using ELISArray

Page 6/32
The concentrations of major human pro-in�ammatory cytokines and chemokines were measured in therespiratory samples of 30 MERS-CoV-infected patients and 18 healthy individuals using the multi-analyteELISArray (Qiagen, Germantown, MD, USA) following the manufacturer’s protocol. The absorbance of theELISArray was measured at 450 nm. The concentrations were calculated using a standard curve.Cytokine/chemokine levels were expressed as pg/ml.
Protein-protein interaction (PPI) Network Construction andIdenti�cation of Hub ProteinsTo further explore the potential interplay among the differentially expressed proteins (C3a, C5a, factor P(properdin) IL-8 and RANTES) in the lung of MERS-CoV infected patients with the potential interactors.Protein-protein interaction networks was constructed using bioinformatics resources of the Search Toolfor the Retrieval of Interacting Genes/Proteins database (STRING version 11.0). This bioinformaticsresources provides known and predicted protein-protein interactions networks. We used multiple proteins(C3a, C5a, factor P (properdin) IL-8 and RANTES) as an input (seed proteins). Active interaction sources,includes; databases, co-occurrence of genes, homology of proteins, experiments (biochemical/geneticdata), co-expression of gene and text mining as well as species limited to “Homo sapiens”, con�dencescore > 0.4 and maximum interactors (= 20) were used to construct the STRING networks. The STRINGresource is available online at https://string-db.org/. The constructed STRING PPI network was exportedto Cytoscape software (version 3.8.2 http://apps.cytoscape.org/apps/mcode) to visualize and additionalanalysis of functional protein-protein interaction.
Statistical AnalysisStatistical analyses were performed using the GraphPad 5.0 software (GraphPad Software, San Diego,CA, USA). Data were assessed using a t-test. The correlations between complement proteins,in�ammatory factors, and chemokines were assessed using the Pearson’s correlation test. The resultswere presented as means ± standard deviation (SD), unless otherwise speci�ed. A p-value of <0.05 wasconsidered statistically signi�cant.
Results3.1 Basic Patient Characteristics
A total of 31 MERS-CoV-infected patients (22 male and 9 female) and 15 healthy controls (9 male and 6female) were included in this study (Figure 1). The ages ranged from 31 to 100 years. The overall meanand median age were 68.26 ± 16.04 and 73 years, respectively. The mean ages in the non-survival andsurvival patients were 74.5 ± 11.3 and 53± 15.8 years, respectively. Twenty-two (70.97%) patients died, 9recovered (29.03%), and 25 (80.6 %) required intensive care unit (ICU) admission (Fig 1).

Page 7/32
3.2 Activation of Complement Is Associated with MERS-CoV Infection
3.2.1 Complement Anaphylatoxin Expression Levels Are Signi�cantlyIncreased in MERS-CoV-Infected Patients Compared with HealthyControlsOver-activation of the complement system is associated with immunopathology and tissue damage.Therefore, we hypothesized that the elevated lung complement protein levels occurring during MERS-CoVinfection are involved in the massive in�ltration of immune cells into the lungs and result in poor diseaseoutcome. To assess whether lung complement anaphylatoxins (C3a and C5a) were increased in MERS-CoV-infected patients, we examined complement components in lower respiratory lung samples usingELISA. As shown in (Fig 2), the levels of complement anaphylatoxins (C3a and C5a) and C1q weresigni�cantly higher in the lungs of MERS-CoV-infected patients compared with healthy controls (Fig 2A,2B, and 2G). Together, these results show that the high levels of complement anaphylatoxins in the lungsof MERS-CoV-infected patients are signi�cantly increased, suggesting a signi�cant pulmonarycomplement activation. In addition, the elevated levels of lung complement anaphylatoxins suggest arole for these factors in lung tissue damage, immunopathology, ARDS development, and mortality ofMERS-CoV-infected patients.
3.2.2 MERS-CoV Infection Is Associated with Positive Regulation of the Complement Response
Elevated levels of complement anaphylatoxins prompted us to evaluate the changes in otherimmunoregulatory proteins. Complement regulatory proteins function by inhibiting complement over-activation to avoid in�ammation and tissue damage. We quanti�ed factor P (properdin) levels in the lungof MERS-CoV-infected patients. As shown in (Fig 2C), the levels of positive regulatory factor P (properdin)were signi�cantly higher in the MERS-CoV-infected patients compared with the healthy control group,suggesting that the levels of factor P may enhance complement activation during MERS-CoV infection.Factor P measurement in serum may provide evidence for the involvement of the alternative complementpathway since factor P (properdin) represents an important factor in the activation of the alternativepathway.
3.2.3 MERS-CoV Infection Is Associated with Distinct Levels of Negative Regulatory Complement Proteins
The measurement of negative regulatory proteins in the lung of MERS-CoV-infected patients may provideevidence for complement system regulation. The quantitation of several complement negative regulatoryfactors (factor I, C4-BP, and factor H) revealed that the levels of factors I and H were increased in thelower lung respiratory samples (Fig 2D and 2E). These negative regulatory proteins play an important rolein complement system regulation. In contrast, the levels of C4-BP were unchanged, and there was nostatistical signi�cance between the levels in the MERS-CoV-infected patients compared with healthycontrols (Fig 2F). This result indicates that MERS-CoV may suppress and inhibit the function of C4-B.

Page 8/32
3.3 Pulmonary Neutrophil Chemoattractant Chemokine IL-8 (CXCL8) and RANTES (CCL5) Levels AreElevated in MERS-CoV-Infected Patients
To examine the levels of chemokines in the lung during MERS-CoV infection, we quanti�ed IL-8 andRANTES in the lower respiratory tract of MERS-CoV-infected patients and the healthy control group. Asshown in Figure 2, IL-8 and RANTES levels were signi�cantly higher in the lungs of MERS-CoV-infectedpatients compared with that in the healthy control group (Fig 2H and 2I).
3.2 Activation of Complement Is Associated with MERS-CoV Infection
3.2.1 Complement Anaphylatoxin Expression Levels Are Signi�cantlyIncreased in MERS-CoV-Infected Patients Compared with HealthyControlsOver-activation of the complement system is associated with immunopathology and tissue damage.Therefore, we hypothesized that the elevated lung complement protein levels occurring during MERS-CoVinfection are involved in the massive in�ltration of immune cells into the lungs and result in poor diseaseoutcome. To assess whether lung complement anaphylatoxins (C3a and C5a) were increased in MERS-CoV-infected patients, we examined complement components in lower respiratory lung samples usingELISA. As shown in (Fig 2), the levels of complement anaphylatoxins (C3a and C5a) and C1q weresigni�cantly higher in the lungs of MERS-CoV-infected patients compared with healthy controls (Fig 2A,2B, and 2G). Together, these results show that the high levels of complement anaphylatoxins in the lungsof MERS-CoV-infected patients are signi�cantly increased, suggesting a signi�cant pulmonarycomplement activation. In addition, the elevated levels of lung complement anaphylatoxins suggest arole for these factors in lung tissue damage, immunopathology, ARDS development, and mortality ofMERS-CoV-infected patients.
3.2.2 MERS-CoV Infection Is Associated with Positive Regulation of the Complement Response
Elevated levels of complement anaphylatoxins prompted us to evaluate the changes in otherimmunoregulatory proteins. Complement regulatory proteins function by inhibiting complement over-activation to avoid in�ammation and tissue damage. We quanti�ed factor P (properdin) levels in the lungof MERS-CoV-infected patients. As shown in (Fig 2C), the levels of positive regulatory factor P (properdin)were signi�cantly higher in the MERS-CoV-infected patients compared with the healthy control group,suggesting that the levels of factor P may enhance complement activation during MERS-CoV infection.Factor P measurement in serum may provide evidence for the involvement of the alternative complementpathway since factor P (properdin) represents an important factor in the activation of the alternativepathway.
3.2.3 MERS-CoV Infection Is Associated with Distinct Levels of Negative Regulatory Complement Proteins

Page 9/32
The measurement of negative regulatory proteins in the lung of MERS-CoV-infected patients may provideevidence for complement system regulation. The quantitation of several complement negative regulatoryfactors (factor I, C4-BP, and factor H) revealed that the levels of factors I and H were increased in thelower lung respiratory samples (Fig 2D and 2E). These negative regulatory proteins play an important rolein complement system regulation. In contrast, the levels of C4-BP were unchanged, and there was nostatistical signi�cance between the levels in the MERS-CoV-infected patients compared with healthycontrols (Fig 2F). This result indicates that MERS-CoV may suppress and inhibit the function of C4-B.
3.3 Pulmonary Neutrophil Chemoattractant Chemokine IL-8 (CXCL8) and RANTES (CCL5) Levels AreElevated in MERS-CoV-Infected Patients
To examine the levels of chemokines in the lung during MERS-CoV infection, we quanti�ed IL-8 andRANTES in the lower respiratory tract of MERS-CoV-infected patients and the healthy control group. Asshown in Figure 2, IL-8 and RANTES levels were signi�cantly higher in the lungs of MERS-CoV-infectedpatients compared with that in the healthy control group (Fig 2H and 2I).
3.5 Lung Complement Anaphylatoxins (C3a and C5a), IL-8, and RANTES Levels Are Associated withARDS Development and the Fatality Rate
The Pearson correlation analysis revealed a signi�cant association of C3a, C5a, IL-8, and RANTES withARDS development and a higher fatality rate (Table 1). Higher levels of these mediators were signi�cantlypositively correlated with ARDS and a higher fatality rate. These immune mediators increase the risk ofdeveloping ARDS and mortality in MERS-CoV-infected patients. Elevation of lung C3a, C5a, IL-8, andRANTES may represent biomarkers for ARDS development and a predictor for inhospital mortality.
Table 1. Correlation between lung complement C3a, C5a, IL-8, and RANTES with ARDS development andin hospital mortality
Variable Pearson (r) P-value
C3a Death (r = 0.5249)
ARDS (r = 0.5249)
0.0024
0.0024
C5a Death (r = 0.6845)
ARDS (r = 0.6845)
<0.0001
<0.0001
RANTES Death (r = 0.7392)
ARDS (r = 0.7392)
<0.0001
<0.0001
IL-8 Death (r = 0.7836)
ARDS (r = 0.7836)
<0.0001
<0.0001

Page 10/32
3.6 Lung Complement Anaphylatoxins (C3a and C5a) and Their Correlation with Factor P, IL-8, andRANTES
We further examined whether there were correlations between lung complement anaphylatoxins (C3a andC5a), factor P, IL-8, and RANTES. The results indicated that lung complement C5a was positivelycorrelated with factor P, IL-8, and RANTES, whereas complement C3a was positively correlated with IL-8and RANTES (Table 2). A positive correlation between RANTES, IL-8, and complement factor P was alsoobserved (Table 2). Similarly, IL-8 was positively correlated with complement factor P (Table 2).
Table 2. Lung complement anaphylatoxins (C3a and C5a) and theircorrelation with factor P, IL-8, and RANTES
Variable Pearson (r) P-value
C3a RANTES (r = 0.3806)
IL-8 (r = 0. 0.4710)
0.0347
0.0075
C5a RANTES (r = 0.5486)
IL-8 (r = 0.3762)
Factor P (r = − 0.3949)
0.0014
0.0370
0.0279
RANTES Factor P (r = 0.5563)
IL-8 (r = 0.5141)
0.0012
0.0031
IL-8 Factor P (r = 0.5939) 0.0004
3.7 Protein-protein interaction (PPI) Network Analysis and Hub Proteins
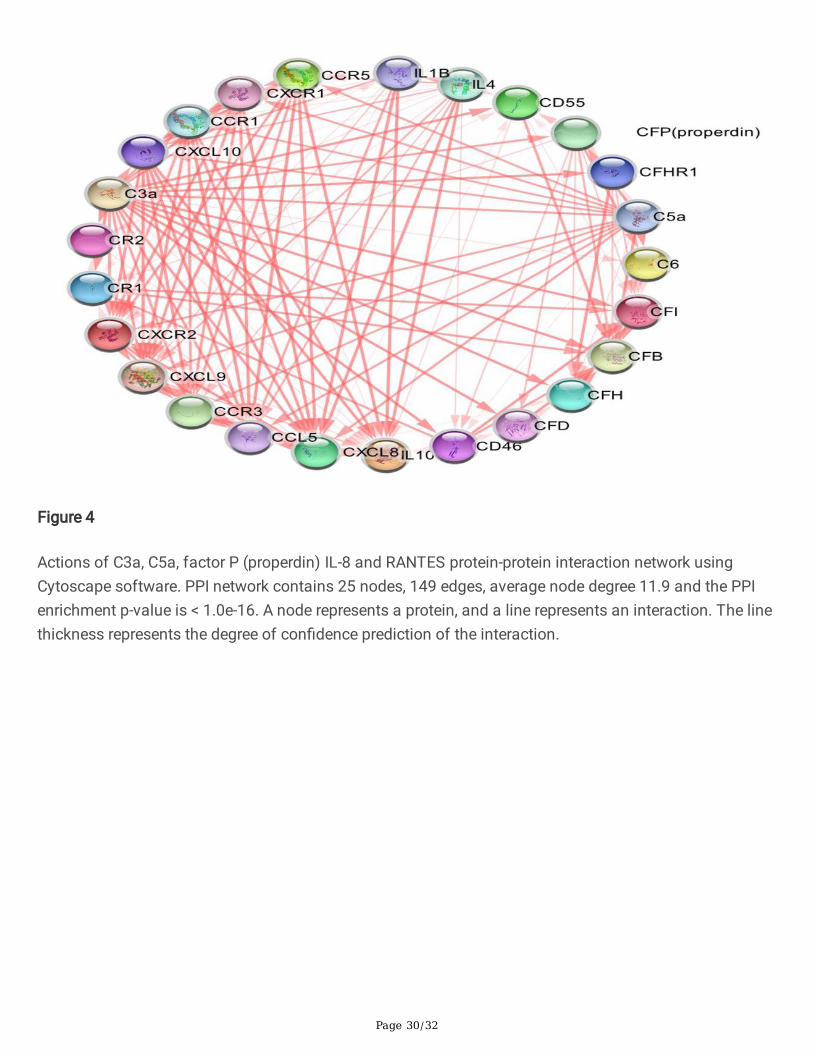
To understand the protein-protein interaction between the complement proteins andcytokines/chemokines and predicted interactors, we performed data mining and curation to constructand map the network of protein-protein interactions using the STRING databases. C3a, C5a, factor P(properdin) IL-8 and RANTES were used as seeds for network analysis. The PPI network showed 20proteins that have the highest interaction scores with C3a, C5a, factor P (properdin), IL-8 and RANTES(Fig 4).For C3a, C5a, IL-8 and RANTES, individual protein-protein interaction was also created (Fig 5,6 and7). The name of proteins, predicted functional partners and actions are shown in (Table S1). This networkcomprises several types of interactions including; gene neighborhood, gene fusions, co-occurrence, co-expression, text-mining, protein homology and experiments (biochemical/genetic data). The physical(direct) or functional (indirect) interactions also was shown. The constructed network contained 25nodes, 149 edges, average node degree 11.9 and the PPI enrichment p-value was < 1.0e-16. Cytoscapeanalysis were shown in the (Table S2). CytoHubba analysis identi�ed top 6 proteins (hub nodes) basedon Betweenness, Closeness, Degree, EcCentricity, EPC, MCC and MNC calculation methods (Table S3 and

Page 11/32
S4). These hub nodes are involved in in�ammatory response and molecular binding interaction as well asligand-receptor interaction. These results indicate that predicted proteins (interactors) and hub proteinsmay also have a role in MERS-CoV disease severity.
DiscussionAn excessive in�ammatory response is a major characteristic of MERS-CoV infections and results indisease progression and poor clinical outcomes. MERS-CoV infections are characterized by dysregulationof the innate and adaptive immunity systems. Several in�ammatory cytokines/chemokines and over-activation of complement proteins are signi�cantly associated with severe MERS-CoV infections and ahigher fatality rate [5], [6], [8]. In this study, we sought to determine the levels of pulmonary complementproteins, neutrophil chemoattractant chemokine IL-8 (CXCL8), and RANTES (CCL5), as well as viral load,in MERS-CoV-infected patients and assessed their association with mortality. This study is the �rst toevaluate the pulmonary complement protein expression pro�le during MERS-CoV infection. Variousinnate immune markers in respiratory samples collected from MERS-CoV-infected patients and healthysubjects were evaluated. The results showed high levels of complement anaphylatoxins (C3a and C5a),positive complement regulatory proteins, neutrophil chemoattractant chemokine IL-8 (CXCL8), RANTES(CCL5), and viral load (data not shown) in MERS-CoV-infected patients. A number of studies have foundthat high viral load, duration of viral shedding, and direct cytopathic effects were associated with severecomplications during SARS-CoV-1, SARS-CoV-2, and MERS-CoV infections [19]–[22]. MERS-CoV infectionis characterized by persistent viral load, and in severe cases of MERS-CoV infection, viral shedding isdetected beyond 21 days [23]. Our results showed that all MERS-CoV-infected patients exhibited high viralloads (Ct values). Further studies revealed that MERS-CoV-infected patients requiring ICU admission hadhigh MERS-CoV RNA levels [24]. In this study, 70% of the MERS-CoV-infected patients developedpneumonia and required ICU admission.
Over-activation of the immune response, including the complement system, is believed to be an importantfactor for the high fatality rate of in�uenza pandemic (1918); H1N1, H5N1, and H7N9; SARS-CoV-1,MERS-CoV; Ebola infection; and, most recently, COVID-19 (SARS-CoV-2) infection [5], [10], [25]–[32].Complement activation leads to the production of several effector pro-in�ammatory molecules, includinganaphylatoxins C3a and C5a. Over-stimulation of the complement system or inadequate inhibitioncauses tissue damage [33] and formation of high levels of anaphylatoxins C3a and C5a. C5a is achemoattractant for neutrophils, monocytes, eosinophils, and other in�ammatory cells. C5a and C3a alsoplay important roles in in�ammatory cascades [34], [35]. Following infection, complement anaphylatoxinsstimulate phagocytic cells and the production of high levels of in�ammatory cytokines (cytokine storm),granular enzymes, and free radicals. These mediators may eventually contribute to vascular dysfunction,�brinolysis, microvascular thrombosis formation, or tissue damage [11], [12], [14], [15], [17], [18]. C5a is akey complement protein in the activation and bridging of the innate and adaptive immune responses [35].In this study, the expression levels of the pulmonary neutrophil chemoattractant chemokine, IL-8 (CXCL8),C5a, and C3a were increased in MERS-CoV-infected patients and associated with a higher fatality rate. Anumber of viral infections are associated with complement activation and coagulation dysfunction [11],

Page 12/32
[15], [16]. Our results are consistent with previous findings [36], [37] showing excessive complementactivation, particularly anaphylatoxins C3a, C5a, and C5b-9, during MERS-CoV and SARS-CoV-1 infection,which, in turn, contribute to lung tissue damage, a hyper-in�ammatory response, and severecomplications from infection. Furthermore, several studies have revealed that C5a was associated withacute lung diseases, severe pneumonia, and immunopathology during highly pathogenic viral infectionwith H1N1, H5N1, H7N9, and SARS-CoV-1 [31], [32], [36], [38]. Similar to SARS-CoV-1 and MERS-CoV,patients with COVID-19 (SARS-CoV-2) are characterized by complement activation with high levels ofcomplement protein [11], [26], [39], [40]. Moreover, one study found that high levels of C3a and C5a inlower respiratory tract samples were associated with severe complications from H5N1 infection [41]. Thisresult con�rms that the pulmonary complement is activated and anaphylatoxins C3a and C5a arereleased upon infection with in�uenza. Many studies have reported high levels of pulmonary andsystematic C5a in highly pathogenic in�uenza-infected patients [16], [31]. Likewise, respiratory samplesfrom mice infected with the highly pathogenic avian in�uenza, H5N1, exhibited signi�cant increases ofC5a [42]. A rapid elevation of C3a variants (C3a, C3b, iC3b, C3c, C3d) was observed in mice infected withSARS-CoV-1, which contributed to the systemic pro-in�ammatory response and lung injury [36]. C3activation has also been shown to be elevated in cells infected with SARS-CoV-2 [43]. In addition, in vitroand in vivo experiments on human respiratory syncytial viral infections showed complement-mediatedlung damage induced by high levels of C3a [44]. These �ndings clearly demonstrate that C5a and C3arecruit and activate in�ammatory immune cells and play a central role in lung injury andimmunopathology during respiratory viral infection. In contrast, complement C5a and C3a are involved inARDS pathogenesis, neutrophil recruitment and activation, and lung endothelial and epithelial injury [35],[45]. Previous studies have demonstrated that highly pathogenic viral infections are characterized by arapid progression to ARDS [46]–[51]. In addition, MERS-CoV infection is associated with a more severepneumonia compared with SARS-CoV-1 infection [47]. ARDS was shown to be the main cause ofmortality among patients infected with MERS-CoV, SARS-CoV-1, and highly pathogenic in�uenza virus[46], [52], [53]. In this study, we observed that almost all of the MERS-CoV-infected patients rapidlydeveloped ARDS and required ICU admission. Therefore, we hypothesize that complement-induced ARDSmay contribute to the disease severity and high mortality rate of MERS-CoV-infected patients observed inthis study.
We observed that the expression levels of pulmonary neutrophil chemoattractant chemokine IL-8 (CXCL8)and C5a were signi�cantly higher in MERS-CoV-infected patients compared with the healthy controlgroup. C5a contributes to the formation of neutrophil extracellular traps (NETosis) and in�ammatorycytokine induction. IL-8 and C5a are important chemoattractants for neutrophil recruitment, activation,and neutrophil accumulation, and both induce NETosis. Excess NETosis contributes to the in�ammation,pathological cellular damage, and acute lung injury in mice infected by in�uenza virus [54]–[56].Similarly, high numbers of neutrophils and NETs were induced by C5a and contributed to theimmunopathology, alveolar damage, and acute lung injury during in�uenza A H1N1 infection [54]. Anumber of in vitro and in vivo studies have found that C5a was associated with macrophage andendothelial cell activation, endothelial damage, NETosis, and increases in alveolar-capillary barrier

Page 13/32
permeability, as well as vascular leakage [34], [57]. Thus, we hypothesize that high C5a, C3a, and IL-8levels may play a vital role in lung tissue damage, immunopathology, ARDS development, ICU admission,and mortality in MERS-CoV-infected patients.
During SARS-CoV-1 infection, C5a induced several pro-in�ammatory cytokines and chemokines includingIL-8 [58]. C5a can also establish a positive feedback loop consisting of IL-8 induction, which results infurther IL-8 production. Thus, a loop of continued IL-8 production may result in further in�ammatory cellrecruitment to the lung and subsequently contribute to lung damage and pathological changes [59]. Wefound that high levels of the lung complement anaphylatoxins, C5a and C3a, were closely associatedwith the overexpression of pulmonary IL-8 in MERS-CoV-infected patients. There was a signi�cantcorrelation between C5a, C3a, and IL-8. The overproduction of C5a and IL-8 may be responsible for moredamage to the host tissue compared with MERS-CoV. Previous studies have also shown that the levels ofIL‐8 signi�cantly correlate with neutrophil numbers and airway in�ammation, as well as lung dysfunction,in patients with chronic obstructive pulmonary disease and asthma [18], [60]. Therefore, we conclude thatthe high levels of C5a might be a direct mediator of neutrophil chemoattraction or an indirect inducer ofIL-8 production during MERS-CoV infection.
C5a is also known to activate in�ammatory cells to release reactive oxygen species (ROS). ROSoverproduction leads to oxidative stress that subsequently contributes to airway and lung damage [61]–[63]. A previous in vivo study demonstrated that ROS are strongly associated with lung damage andpneumonia in mice infected with in�uenza virus [64]. Mice treated with an antioxidant resulted insigni�cantly reduced mortality, lung damage, and pathogenesis following in�uenza virus infection [65].Several studies have observed that anti-C5a and a C5aR receptor antagonist signi�cantly blocked andinhibited neutrophil oxidative burst formation [18], [66], [67]. These studies suggest a critical role of theC5a/C5aR/ROS axis in lung pathology and virus-induced immunopathology. We hypothesize that highlevels of C5a induce ROS, which results in a more severe MERS-CoV infection, ARDS, andimmunopathology.
Several studies have shown that targeted therapy with C5a and C5aR represents a signi�cant anti-in�ammatory approach to control in�ammatory cell recruitment, immunopathology, and acute lungdamage induced by highly pathogenic viral infections [31], [68], [69]. One study showed that the blockadeof the C5a–C5aR axis in mice infected with MERS-CoV markedly reduced pathological changes in lungand spleen tissue, viral load, lymphopenia, and systemic and pulmonary in�ammatory responses [37].Another study showed that complement-de�cient mice infected with SARS-CoV-1 were associated withdecreased lung neutrophilia, a pro-in�ammatory response, and viral replication [36]. Likewise, micede�cient in C3aR infected with HRSV exhibited reduced airway hyper-responsiveness; mucus secretion;and reduced lymphocyte, neutrophil, macrophage, and eosinophil in�ltration [44]. A recent study indicatedthat high levels of C3 mediated periodontal in�ammation and an increased risk for periodontitis inpatients with diabetes [70]. The same study found that the C3 knockout mouse model inhibitedin�ammatory cytokines in diabetic mice and reduced the risk of periodontal disease.

Page 14/32
Blocking and targeting C5a/C5aR and C3a/C3aR during respiratory viral infections signi�cantly reducedand controlled pathology, lung damage, and mortality and inhibited in�ammatory cell in�ltration in thelung, T-lymphocyte apoptosis, and NETosis [31], [71]. In addition, in vivo experiments con�rmed thatantihuman C5a antibody treatment in H5N1- and H7N9-infected African green monkeys resulted inreduced in�ltration of in�ammatory cells in the lung and decreased levels of pro-in�ammatory cytokinesand chemokines [32], [71]. Furthermore, studies have shown similar associations between the inhibitionof the complement system during highly pathogenic viral infection and decreased acute lung injury and arobust immune response [12], [36], [71], [72].
A recent study demonstrated that severe COVID-19 cases are associated with high levels of plasma C5aand soluble membrane attack complex (sC5b-9), signifying the C5a blockade as a potential interventionstrategy [73], [74]. Several randomized controlled trials showed that anti-C5 therapy increased survival insevere COVID-19 cases and signi�cantly reduced in�ammatory marker production [73], [75]–[77]. Thesestudies con�rmed that the inhibition of an over-activated complement response and its signalingpathways signi�cantly affected immunopathology, respiratory disease severity, and viral replication.Accordingly, it is reasonable to speculate that targeting the C5a/C5aR and C3a/C3aR axes may beeffective at controlling MERS-CoV infection-induced immunopathology as MERS-CoV infection similarlyleads to acute lung injury.
Over-activation of the complement system is controlled by a series of proteins known as the regulators ofcomplement activation. These proteins play a key role in preventing complement-mediated tissue and celldamage and dysregulation of one or more of the complement regulatory proteins, which results in tissueinjury, immunopathology, and in�ammation-associated disease [78], [79]. The complement system isnegatively regulated by several complement proteins, including factor I, C1-inhibitor (CI-INH), factor H, andC4-BP [80]–[83]. In contrast, factor P (properdin) is a positive regulatory complement protein. In this study,the levels of positive regulatory factor P (properdin) were signi�cantly higher in the lungs of MERS-CoV-infected patients compared with healthy controls, suggesting that the levels of factor P may positivelyregulate complement activation during MERS-CoV infection [81]. Factor P (properdin) has beenassociated with complement-mediated organ injury in various human diseases. Therapy targeting factorP (properdin) showed bene�cial outcomes and prevented complement-mediated tissue damage [84]–[90].The measurement of factor P may provide evidence for the involvement of the alternative complementpathway since factor P (properdin) is an important factor in alternative pathway activation [91]. A recentstudy demonstrated that SARS-CoV-2 can directly activate the alternative complement pathway [74]. Incontrast, the levels of negative regulatory proteins, factor I and C4-BP, were decreased in the lung ofMERS-CoV-infected patients when compared with healthy subjects. These negative regulatory proteinsplay an important role in regulating the complement system. The low levels of factor I and C4-BP suggestthat patients with MERS-CoV have a reduced capacity to control and regulate complement activation.These results indicate that MERS-CoV somehow suppressed and inhibited negative regulatorycomplement proteins during infection [80], [81], [92], [93]. A number of viral infection-mediated chronicin�ammatory and autoimmunity diseases are associated with complement regulatory proteindysfunction [79], [81], [94], [95]. Over-activation of pulmonary and systemic complement plays a key role

Page 15/32
in in�ammation, endothelial cell damage, thrombus formation, and intravascular coagulation andultimately leads to multiple-organ failure and death [12], [18]. In this study, high levels of pulmonarycomplement mediators, disease severity, and increased mortality appear to be linked to the degree ofcomplement activation against MERS-CoV. Complement C3a and C5a may be an independent risk factorfor death in MERS-CoV-infected patients.
RANTES (CCL5) is a key proin�ammatory chemokine produced during respiratory viral infection. Virus-infected lung and epithelial cells secret high amount of RANTES (in addition to immune cells). Also,RANTES (CCL5) has a critical role in the platelet activation and initiation of the coagulation cascade [96].In this study, we detected high levels of RANTES (CCL5), this in�ammatory chemokine was signi�cantlycorrelated with death and ARDS among MERS-CoV infected patients. In an in vitro experiment, infectionof human monocyte-derived macrophages (MDMs) and dendritic cells (MDDCs) with MERS-CoV showeda high levels and upregulation of RANTES (CCL5) and other in�ammatory chemokines such as MIP-1α,IP-10 and IL-8 [97]. Recent studies showed that the sever and mild COVID-19 patients had elevated levelsof RANTES (CCL5) [97]–[99]. Recent data form SARAS-CoV studies suggested targeting CCR5 could be atherapeutic strategy for COVID-19 [99]–[101].Previous studies also showed increased levels of RANTES(CCL5) contributed to exacerbation of allergic airway in�ammation and severe human respiratorysyncytial infection [102]–[104].
Studying the protein-protein interaction networks and the predicted protein interactors provides asigni�cance data to explore the functions of proteins [105], [106]. In PPI networks, proteins that havedirect interaction with several other proteins are called hub proteins (hub nodes). Proteins with moreinteraction partners may become targets for follow-up investigation [107]. In this study, the constructedPPI network (25 nodes and 149 edges) demonstrated that C3, C5, CCL5, CR1, CXCL8, IL10, IL-4 andCXCR1 were the hub proteins (nodes). Most of these hub proteins are involved in the in�ammatoryresponse and molecular binding interaction as well as ligand-receptor interaction. These results indicatethat predicted proteins (interactors) may also have a role in MERS-CoV disease severity. The function andexact role of hub proteins identi�ed in our PPI network in MERS-CoV infection require furtherinvestigation.
Here, we propose a mechanism to explain the role of pulmonary complement anaphylatoxins (C3a andC5a) IL-8 and RANTES (CCL5) in severe MERS-CoV infection. High levels of complement anaphylatoxinsC5a and IL-8 recruit and activate neutrophils. Subsequently, activated neutrophils undergo NETosis andROS production, which results in oxidative stress. Increased levels of C5a and IL-8 may cause anin�ammatory loop that contributes to extensive cellular damage and pathological changes. Also,elevation of complement anaphylatoxins C5a and C3a may induce cytokine storm and in�ammatorycascades. On the other hand, RANTES (CCL5) recruit and activate macrophage while, C3a recruit, activeand degranulate mast cells and eosinophils which results in airway smooth muscle contraction. Thesemeditators eventually contribute to airway and lung damage, more severe MERS-CoV infection, ARDS,and immunopathology. The overexpression of lung C5a during MERS-CoV infection may establish anampli�cation loop of IL-8 induction. Thus, a loop of continued IL-8 production leads to increased

Page 16/32
in�ammatory cell activation and recruitment to the lung, subsequently contributing to the lungpathological features. Our results and hypothesis may explain the events regulating complementactivation during MERS-CoV infection.
The main limitation of this study is that, we measured complement protiens, IL-8 (CXCL8), RANTES(CCL5) levels and viral load at a single time point; measuring this mediator expression levels and viralload at different time points to create a kinetic pro�le might provide additional information.
We conclude that the high levels of complement anaphylatoxins, C5a and C3a; positive regulatorycomplement protein (factor P); IL-8 (CXCL8); and CCL5 in the lower respiratory tracts of MERS-CoV-infected patients are associated with immunopathology, higher fatality rates, more severe disease, andARDS development. High levels of complement mediators, disease severity, and increased mortalityappear to be linked to the degree of complement activation against MERS-CoV. Furthermore, the levels ofC3a, C5a, IL-8 and CCL5 in the lung may be useful biomarkers to predict a more severe MERS-CoVinfection and mortality. targeting any of these mediators may offer an effective treatment for MERS-CoV.As such, future large studies characterizing components of the complement system at different stages ofMERS-CoV infection may offer an effective immunotherapeutic strategy.
DeclarationsAcknowledgments
The authors thank the Deanship of Scienti�c Research and RSSU at King Saud University for theirtechnical support.
Funding
The authors thank the Research Center at King Fahad Medical City for funding this study (Grant No 019-053).
Ethics approval
This study was reviewed and approved by the Institutional Review Board at King Fahad Medical City (IRBregister number 019-053). This study procedures were in accordance with the 1964 Helsinki Declarationand later versions.
Declaration of Competing Interest
The authors declare that they have no con�ict of interest.
Availability of data and materials
The datasets generated during and/or analysed during the current study are available from thecorresponding author on reasonable request.

Page 17/32
Con�ict of Interest
The authors declare that they have no competing interests
References[1] E. I. Azhar et al., “Evidence for camel-to-human transmission of MERS coronavirus.,” The New Englandjournal of medicine, vol. 370, no. 26, pp. 2499–505, Jun. 2014, doi: 10.1056/NEJMoa1401505.
[2] A. Assiri et al., “Multifacility Outbreak of Middle East Respiratory Syndrome in Taif, Saudi Arabia,”Emerging infectious diseases, vol. 22, no. 1, pp. 32–40, Jan. 2016, doi: 10.3201/eid2201.151370.
[3] A. A. Rabaan et al., “MERS-CoV: epidemiology, molecular dynamics, therapeutics, and futurechallenges,” Annals of Clinical Microbiology and Antimicrobials, vol. 20, no. 1, p. 8, 2021, doi:10.1186/s12941-020-00414-7.
[4] “WHO 2021.” www.emro.who.int/health-topics/mers-cov/mers-outbreaks.html%0D (accessed Jan. 28,2021).
[5] B. Alosaimi et al., “MERS-CoV infection is associated with downregulation of genes encoding Th1 andTh2 cytokines/chemokines and elevated in�ammatory innate immune response in the lower respiratorytract,” Cytokine, vol. 126, Feb. 2020, doi: 10.1016/j.cyto.2019.154895.
[6] J. Tynell et al., “Middle east respiratory syndrome coronavirus shows poor replication but signi�cantinduction of antiviral responses in human monocyte-derived macrophages and dendritic cells,” Journal ofGeneral Virology, vol. 97, no. 2, pp. 344–355, 2016, doi: 10.1099/jgv.0.000351.
[7] W. H. Mahallawi, O. F. Khabour, Q. Zhang, H. M. Makhdoum, and B. A. Suliman, “Cytokine MERS-CoVinfection in humans is associated with a pro-in � ammatory Th1 and Th17 cytokine pro � le,” vol. 104, no.November 2017, pp. 8–13, 2018, doi: 10.1016/j.cyto.2018.01.025.
[8] H.-S. Shin et al., “Immune Responses to Middle East Respiratory Syndrome Coronavirus During theAcute and Convalescent Phases of Human Infection.,” Clinical infectious diseases : an o�cial publicationof the Infectious Diseases Society of America, vol. 68, no. 6, pp. 984–992, Mar. 2019, doi:10.1093/cid/ciy595.
[9] R. Reghunathan et al., “Expression pro�le of immune response genes in patients with severe acuterespiratory syndrome,” BMC Immunology, vol. 6, 2005, doi: 10.1186/1471-2172-6-2.
[10] R. Channappanavar and S. Perlman, “Pathogenic human coronavirus infections: causes andconsequences of cytokine storm and immunopathology,” Seminars in Immunopathology, vol. 39, no. 5,pp. 529–539, 2017, doi: 10.1007/s00281-017-0629-x.

Page 18/32
[11] M. Marchetti, “COVID-19-driven endothelial damage: complement, HIF-1, and ABL2 are potentialpathways of damage and targets for cure,” Annals of Hematology, vol. 99, no. 8, pp. 1701–1707, 2020,doi: 10.1007/s00277-020-04138-8.
[12] M. Noris and G. Remuzzi, “Overview of complement activation and regulation,” Seminars innephrology, vol. 33, no. 6, pp. 479–492, Nov. 2013, doi: 10.1016/j.semnephrol.2013.08.001.
[13] N. S. Merle, R. Noe, L. Halbwachs-Mecarelli, V. Fremeaux-Bacchi, and L. T. Roumenina, “Complementsystem part II: Role in immunity,” Frontiers in Immunology, vol. 6, no. MAY, pp. 1–26, 2015, doi:10.3389/�mmu.2015.00257.
[14] M. Riedl et al., “Spectrum of complement-mediated thrombotic microangiopathies: pathogeneticinsights identifying novel treatment approaches.,” Seminars in thrombosis and hemostasis, vol. 40, no. 4,pp. 444–464, Jun. 2014, doi: 10.1055/s-0034-1376153.
[15] R. Wang, H. Xiao, R. Guo, Y. Li, and B. Shen, “The role of C5a in acute lung injury induced by highlypathogenic viral infections,” Emerging Microbes and Infections, vol. 4, no. January, pp. 1–7, 2015, doi:10.1038/emi.2015.28.
[16] R. Ohta, Y. Torii, M. Imai, H. Kimura, N. Okada, and Y. Ito, “Serum concentrations of complementanaphylatoxins and proin�ammatory mediators in patients with 2009 H1N1 in�uenza.,” Microbiology andimmunology, vol. 55, no. 3, pp. 191–198, Mar. 2011, doi: 10.1111/j.1348-0421.2011.00309.x.
[17] C. Magro et al., “Complement associated microvascular injury and thrombosis in the pathogenesis ofsevere COVID-19 infection: A report of �ve cases,” Translational research : the journal of laboratory andclinical medicine, vol. 220, pp. 1–13, Jun. 2020, doi: 10.1016/j.trsl.2020.04.007.
[18] R. Wang, H. Xiao, R. Guo, Y. Li, and B. Shen, “The role of C5a in acute lung injury induced by highlypathogenic viral infections,” Emerging microbes & infections, vol. 4, no. 5, pp. e28–e28, May 2015, doi:10.1038/emi.2015.28.
[19] K. V Argyropoulos et al., “Association of Initial Viral Load in Severe Acute Respiratory SyndromeCoronavirus 2 (SARS-CoV-2) Patients with Outcome and Symptoms,” The American Journal of Pathology,vol. 190, no. 9, pp. 1881–1887, 2020, doi: https://doi.org/10.1016/j.ajpath.2020.07.001.
[20] M. Cevik, M. Tate, O. Lloyd, A. E. Maraolo, J. Schafers, and A. Ho, “SARS-CoV-2, SARS-CoV-1 andMERS-CoV viral load dynamics, duration of viral shedding and infectiousness: a living systematic reviewand meta-analysis.” medRxiv, 2020, doi: 10.1101/2020.07.25.20162107.
[21] J. A. Al-Taw�q, “Viral loads of SARS-CoV, MERS-CoV and SARS-CoV-2 in respiratory specimens: Whathave we learned?,” Travel medicine and infectious disease, vol. 34, p. 101629, 2020, doi:10.1016/j.tmaid.2020.101629.

Page 19/32
[22] I. F. N. Hung et al., “Viral loads in clinical specimens and SARS manifestations,” Emerging infectiousdiseases, vol. 10, no. 9, pp. 1550–1557, Sep. 2004, doi: 10.3201/eid1009.040058.
[23] M.-D. Oh et al., “Viral Load Kinetics of MERS Coronavirus Infection.,” The New England journal ofmedicine, vol. 375, no. 13. United States, pp. 1303–1305, Sep. 2016, doi: 10.1056/NEJMc1511695.
[24] H. M. Al-Abdely et al., “Middle East Respiratory Syndrome Coronavirus Infection Dynamics andAntibody Responses among Clinically Diverse Patients, Saudi Arabia.,” Emerging infectious diseases, vol.25, no. 4, pp. 753–766, Apr. 2019, doi: 10.3201/eid2504.181595.
[25] J. Zhou et al., “Active replication of Middle East respiratory syndrome coronavirus and aberrantinduction of in�ammatory cytokines and chemokines in human macrophages: implications forpathogenesis.,” The Journal of infectious diseases, vol. 209, no. 9, pp. 1331–1342, May 2014, doi:10.1093/infdis/jit504.
[26] M. W. Lo, C. Kemper, and T. M. Woodruff, “COVID-19: Complement, Coagulation, and CollateralDamage,” The Journal of Immunology, vol. 205, no. 6, pp. 1488 LP – 1495, Sep. 2020, doi:10.4049/jimmunol.2000644.
[27] A. A. Ansari, “Clinical features and pathobiology of Ebolavirus infection.,” Journal of autoimmunity,vol. 55, pp. 1–9, Dec. 2014, doi: 10.1016/j.jaut.2014.09.001.
[28] A. Allegra, M. Di Gioacchino, A. Tonacci, C. Musolino, and S. Gangemi, “Immunopathology of SARS-CoV-2 Infection: Immune Cells and Mediators, Prognostic Factors, and Immune-Therapeutic Implications,”International journal of molecular sciences, vol. 21, no. 13, p. 4782, Jul. 2020, doi:10.3390/ijms21134782.
[29] A. Haque, D. Hober, and L. H. Kasper, “Confronting potential in�uenza A (H5N1) pandemic with bettervaccines.,” Emerging infectious diseases, vol. 13, no. 10. pp. 1512–1518, Oct. 2007, doi:10.3201/eid1310.061262.
[30] K.-J. Huang et al., “An interferon-γ-related cytokine storm in SARS patients,” Journal of MedicalVirology, vol. 75, no. 2, pp. 185–194, Feb. 2005, doi: 10.1002/jmv.20255.
[31] C. C. Garcia et al., “Complement C5 Activation during In�uenza A Infection in Mice Contributes toNeutrophil Recruitment and Lung Injury,” PLOS ONE, vol. 8, no. 5, p. e64443, May 2013, [Online]. Available:https://doi.org/10.1371/journal.pone.0064443.
[32] S. Sun et al., “Treatment With Anti-C5a Antibody Improves the Outcome of H7N9 Virus Infection inAfrican Green Monkeys,” Clinical Infectious Diseases, vol. 60, no. 4, pp. 586–595, Feb. 2015, doi:10.1093/cid/ciu887.
[33] D. M. Eldewi et al., “Expression levels of complement regulatory proteins (CD35, CD55 and CD59) onperipheral blood cells of patients with chronic kidney disease,” International Journal of General Medicine,

Page 20/32
vol. 12, pp. 343–351, Sep. 2019, doi: 10.2147/IJGM.S216989.
[34] R.-F. Guo and P. A. Ward, “ROLE OF C5A IN INFLAMMATORY RESPONSES,” Annual Review ofImmunology, vol. 23, no. 1, pp. 821–852, Sep. 2004, doi: 10.1146/annurev.immunol.23.021704.115835.
[35] R. Wang, H. Xiao, R. Guo, Y. Li, and B. Shen, “The role of C5a in acute lung injury induced by highlypathogenic viral infections,” Emerging Microbes and Infections, vol. 4, 2015, doi: 10.1038/emi.2015.28.
[36] L. E. Gralinski et al., “Complement Activation Contributes to Severe Acute Respiratory SyndromeCoronavirus Pathogenesis.,” mBio, vol. 9, no. 5, Oct. 2018, doi: 10.1128/mBio.01753-18.
[37] Y. Jiang et al., “Blockade of the C5a–C5aR axis alleviates lung damage in hDPP4 -transgenic miceinfected with MERS-CoV,” Emerging Microbes & Infections, vol. 7, no. 1, pp. 1–12, Dec. 2018, doi:10.1038/s41426-018-0063-8.
[38] S. Sun et al., “Inhibition of complement activation alleviates acute lung injury induced by highlypathogenic avian in�uenza H5N1 virus infection,” American Journal of Respiratory Cell and MolecularBiology, vol. 49, no. 2, pp. 221–230, 2013, doi: 10.1165/rcmb.2012-0428OC.
[39] J. L. Huang et al., “Th2 predominance and CD8+ memory T cell depletion in patients with severeacute respiratory syndrome,” Microbes and Infection, vol. 7, no. 3, pp. 427–436, 2005, doi:10.1016/j.micinf.2004.11.017.
[40] J.-H. Chen et al., “Plasma proteome of severe acute respiratory syndrome analyzed by two-dimensional gel electrophoresis and mass spectrometry,” Proceedings of the National Academy ofSciences of the United States of America, vol. 101, no. 49, pp. 17039 LP – 17044, Dec. 2004, doi:10.1073/pnas.0407992101.
[41] D. van Riel et al., “H5N1 Virus Attachment to Lower Respiratory Tract,” Science, vol. 312, no. 5772, pp.399 LP – 399, Apr. 2006, doi: 10.1126/science.1125548.
[42] K. B. O’Brien, T. E. Morrison, D. Y. Dundore, M. T. Heise, and S. Schultz-Cherry, “A Protective Role forComplement C3 Protein during Pandemic 2009 H1N1 and H5N1 In�uenza A Virus Infection,” PLOS ONE,vol. 6, no. 3, p. e17377, Mar. 2011, [Online]. Available: https://doi.org/10.1371/journal.pone.0017377.
[43] T. Gao et al., “Highly pathogenic coronavirus N protein aggravates lung injury by MASP-2-mediatedcomplement over-activation,” medRxiv, p. 2020.03.29.20041962, Jan. 2020, doi:10.1101/2020.03.29.20041962.
[44] M. M. Bera et al., “Th17 Cytokines Are Critical for Respiratory Syncytial Virus-Associated AirwayHyperreponsiveness through Regulation by Complement C3a and Tachykinins,” The Journal ofImmunology, vol. 187, no. 8, pp. 4245 LP – 4255, Oct. 2011, doi: 10.4049/jimmunol.1101789.

Page 21/32
[45] D. Rittirsch, H. Redl, and M. Huber-Lang, “Role of Complement in Multiorgan Failure,” Clinical andDevelopmental Immunology, vol. 2012, p. 962927, 2012, doi: 10.1155/2012/962927.
[46] K. J. Szretter et al., “Role of Host Cytokine Responses in the Pathogenesis of Avian H5N1 In�uenzaViruses in Mice,” Journal of Virology, vol. 81, no. 6, pp. 2736 LP – 2744, Mar. 2007, doi:10.1128/JVI.02336-06.
[47] T. Chotpitayasunondh et al., “Human disease from in�uenza A (H5N1), Thailand, 2004,” Emerginginfectious diseases, vol. 11, no. 2, pp. 201–209, Feb. 2005, doi: 10.3201/eid1102.041061.
[48] L. Yu et al., “Clinical, Virological, and Histopathological Manifestations of Fatal Human Infections byAvian In�uenza A(H7N9) Virus,” Clinical Infectious Diseases, vol. 57, no. 10, pp. 1449–1457, Nov. 2013,doi: 10.1093/cid/cit541.
[49] K. K. W. To, J. F. W. Chan, H. Chen, L. Li, and K.-Y. Yuen, “The emergence of in�uenza A H7N9 inhuman beings 16 years after in�uenza A H5N1: a tale of two cities,” The Lancet Infectious Diseases, vol.13, no. 9, pp. 809–821, 2013, doi: https://doi.org/10.1016/S1473-3099(13)70167-1.
[50] W.-F. Ng, K.-F. To, W. W. L. Lam, T.-K. Ng, and K.-C. Lee, “The comparative pathology of severe acuterespiratory syndrome and avian in�uenza A subtype H5N1—a review,” Human Pathology, vol. 37, no. 4,pp. 381–390, 2006, doi: https://doi.org/10.1016/j.humpath.2006.01.015.
[51] V. A. Meliopoulos et al., “Human H7N9 and H5N1 In�uenza Viruses Differ in Induction of Cytokinesand Tissue Tropism,” Journal of Virology, vol. 88, no. 22, pp. 12982 LP – 12991, Nov. 2014, doi:10.1128/JVI.01571-14.
[52] L. E. Gralinski et al., “Mechanisms of severe acute respiratory syndrome coronavirus-induced acutelung injury.,” mBio, vol. 4, no. 4, Aug. 2013, doi: 10.1128/mBio.00271-13.
[53] R. Channappanavar and S. Perlman, “Pathogenic human coronavirus infections: causes andconsequences of cytokine storm and immunopathology,” Seminars in Immunopathology, vol. 39, no. 5,pp. 529–539, 2017, doi: 10.1007/s00281-017-0629-x.
[54] T. Narasaraju et al., “Excessive neutrophils and neutrophil extracellular traps contribute to acute lunginjury of in�uenza pneumonitis,” American Journal of Pathology, vol. 179, no. 1, pp. 199–210, 2011, doi:10.1016/j.ajpath.2011.03.013.
[55] O. Z. Cheng and N. Palaniyar, “NET balancing: A problem in in�ammatory lung diseases,” Frontiers inImmunology, vol. 4, no. JAN, pp. 1–13, 2013, doi: 10.3389/�mmu.2013.00001.
[56] H. H. Ng, T. Narasaraju, M. C. Phoon, M. K. Sim, J. E. Seet, and V. T. Chow, “Doxycycline treatmentattenuates acute lung injury in mice infected with virulent in�uenza H3N2 virus: Involvement of matrixmetalloproteinases,” Experimental and Molecular Pathology, vol. 92, no. 3, pp. 287–295, 2012, doi:https://doi.org/10.1016/j.yexmp.2012.03.003.

Page 22/32
[57] S. Youse�, C. Mihalache, E. Kozlowski, I. Schmid, and H. U. Simon, “Viable neutrophils releasemitochondrial DNA to form neutrophil extracellular traps,” Cell Death & Differentiation, vol. 16, no. 11, pp.1438–1444, 2009, doi: 10.1038/cdd.2009.96.
[58] S. L. Smits et al., “Distinct severe acute respiratory syndrome coronavirus-induced acute lung injurypathways in two different nonhuman primate species.,” Journal of virology, vol. 85, no. 9, pp. 4234–4245,May 2011, doi: 10.1128/JVI.02395-10.
[59] L. Wang et al., “Regulation of IL-8 production by complement-activated product, C5a, in vitro and invivo during sepsis,” Clinical Immunology, vol. 137, no. 1, pp. 157–165, 2010, doi:https://doi.org/10.1016/j.clim.2010.05.012.
[60] M. M. Marc et al., “Complement Factors C3a, C4a, and C5a in Chronic Obstructive Pulmonary Diseaseand Asthma,” American Journal of Respiratory Cell and Molecular Biology, vol. 31, no. 2, pp. 216–219,Aug. 2004, doi: 10.1165/rcmb.2003-0394OC.
[61] D. P. Rosanna and C. Salvatore, “Reactive oxygen species, in�ammation, and lung diseases.,” Currentpharmaceutical design, vol. 18, no. 26, pp. 3889–3900, 2012, doi: 10.2174/138161212802083716.
[62] I.-T. Lee and C.-M. Yang, “Role of NADPH oxidase/ROS in pro-in�ammatory mediators-induced airwayand pulmonary diseases.,” Biochemical pharmacology, vol. 84, no. 5, pp. 581–590, Sep. 2012, doi:10.1016/j.bcp.2012.05.005.
[63] J. F.-W. Chan, K. K.-W. To, H. Tse, D.-Y. Jin, and K.-Y. Yuen, “Interspecies transmission and emergenceof novel viruses: lessons from bats and birds.,” Trends in microbiology, vol. 21, no. 10, pp. 544–555, Oct.2013, doi: 10.1016/j.tim.2013.05.005.
[64] W. Domej, K. Oettl, and W. Renner, “Oxidative stress and free radicals in COPD--implications andrelevance for treatment.,” International journal of chronic obstructive pulmonary disease, vol. 9, pp. 1207–1224, 2014, doi: 10.2147/COPD.S51226.
[65] T. Akaike et al., “Dependence on O2- generation by xanthine oxidase of pathogenesis of in�uenzavirus infection in mice.,” The Journal of clinical investigation, vol. 85, no. 3, pp. 739–745, Mar. 1990, doi:10.1172/JCI114499.
[66] T. Sprong et al., “Inhibition of C5a-induced in�ammation with preserved C5b-9-mediated bactericidalactivity in a human whole blood model of meningococcal sepsis.,” Blood, vol. 102, no. 10, pp. 3702–3710, Nov. 2003, doi: 10.1182/blood-2003-03-0703.
[67] T. E. Mollnes et al., “Essential role of the C5a receptor in E coli-induced oxidative burst andphagocytosis revealed by a novel lepirudin-based human whole blood model of in�ammation.,” Blood,vol. 100, no. 5, pp. 1869–1877, Sep. 2002.

Page 23/32
[68] M. Bosmann and P. A. Ward, “Protein-based therapies for acute lung injury: targeting neutrophilextracellular traps.,” Expert opinion on therapeutic targets, vol. 18, no. 6, pp. 703–714, Jun. 2014, doi:10.1517/14728222.2014.902938.
[69] L. Sun, R.-F. Guo, H. Gao, J. V. Sarma, F. S. Zetoune, and P. A. Ward, “Attenuation of IgG immunecomplex-induced acute lung injury by silencing C5aR in lung epithelial cells.,” FASEB journal : o�cialpublication of the Federation of American Societies for Experimental Biology, vol. 23, no. 11, pp. 3808–3818, Nov. 2009, doi: 10.1096/fj.09-133694.
[70] Y. Li et al., “Complement 3 mediates periodontal destruction in patients with type 2 diabetes byregulating macrophage polarization in periodontal tissues.,” Cell proliferation, vol. 53, no. 10, p. e12886,Oct. 2020, doi: 10.1111/cpr.12886.
[71] S. Sun et al., “Inhibition of complement activation alleviates acute lung injury induced by highlypathogenic avian in�uenza H5N1 virus infection.,” American journal of respiratory cell and molecularbiology, vol. 49, no. 2, pp. 221–230, Aug. 2013, doi: 10.1165/rcmb.2012-0428OC.
[72] A. Rattan et al., “Synergy between the classical and alternative pathways of complement is essentialfor conferring effective protection against the pandemic in�uenza A(H1N1) 2009 virus infection,” PLOSPathogens, vol. 13, no. 3, p. e1006248, Mar. 2017, [Online]. Available:https://doi.org/10.1371/journal.ppat.1006248.
[73] M. Cugno et al., “Complement activation in patients with COVID-19: A novel therapeutic target.,” TheJournal of allergy and clinical immunology, vol. 146, no. 1. pp. 215–217, Jul. 2020, doi:10.1016/j.jaci.2020.05.006.
[74] J. Yu, X. Yuan, H. Chen, S. Chaturvedi, E. M. Braunstein, and R. A. Brodsky, “Direct activation of thealternative complement pathway by SARS-CoV-2 spike proteins is blocked by factor D inhibition,” Blood,vol. 136, no. 18, pp. 2080–2089, Oct. 2020, doi: 10.1182/blood.2020008248.
[75] P. F. Stahel and S. R. Barnum, “Complement Inhibition in Coronavirus Disease (COVID)-19: ANeglected Therapeutic Option ,” Frontiers in Immunology , vol. 11. p. 1661, 2020, [Online]. Available:https://www.frontiersin.org/article/10.3389/�mmu.2020.01661.
[76] J. Carvelli et al., “Association of COVID-19 in�ammation with activation of the C5a–C5aR1 axis,”Nature, vol. 588, no. 7836, pp. 146–150, 2020, doi: 10.1038/s41586-020-2600-6.
[77] F. Diurno et al., “Eculizumab treatment in patients with COVID-19: preliminary results from real lifeASL Napoli 2 Nord experience.,” European review for medical and pharmacological sciences, vol. 24, no.7, pp. 4040–4047, Apr. 2020, doi: 10.26355/eurrev_202004_20875.
[78] S. Meri and H. Jarva, “Complement Regulatory Proteins and Related Diseases,” eLS. Jun. 13, 2013,doi: https://doi.org/10.1002/9780470015902.a0001434.pub3.

Page 24/32
[79] P. Ghosh, R. Sahoo, A. Vaidya, M. Chorev, and J. A. Halperin, “Role of Complement and ComplementRegulatory Proteins in the Complications of Diabetes,” Endocrine Reviews, vol. 36, no. 3, pp. 272–288,Jun. 2015, doi: 10.1210/er.2014-1099.
[80] J. M. Thurman and B. Renner, “Dynamic control of the complement system by modulated expressionof regulatory proteins,” Laboratory Investigation, vol. 91, no. 1, pp. 4–11, 2011, doi:10.1038/labinvest.2010.173.
[81] P. Agrawal, R. Nawadkar, H. Ojha, J. Kumar, and A. Sahu, “Complement Evasion Strategies of Viruses:An Overview,” Frontiers in microbiology, vol. 8, p. 1117, Jun. 2017, doi: 10.3389/fmicb.2017.01117.
[82] A. Fletcher-Sandersjöö and B.-M. Bellander, “Is COVID-19 associated thrombosis caused byoveractivation of the complement cascade? A literature review,” Thrombosis research, vol. 194, pp. 36–41, Oct. 2020, doi: 10.1016/j.thromres.2020.06.027.
[83] V. Afshar-Kharghan, “The role of the complement system in cancer,” The Journal of clinicalinvestigation, vol. 127, no. 3, pp. 780–789, Mar. 2017, doi: 10.1172/JCI90962.
[84] F. Corvillo et al., “Serum properdin consumption as a biomarker of C5 convertase dysregulation in C3glomerulopathy.,” Clinical and experimental immunology, vol. 184, no. 1, pp. 118–125, Apr. 2016, doi:10.1111/cei.12754.
[85] P. Dimitrova, N. Ivanovska, W. Schwaeble, V. Gyurkovska, and C. Stover, “The role of properdin inmurine zymosan-induced arthritis.,” Molecular immunology, vol. 47, no. 7–8, pp. 1458–1466, Apr. 2010,doi: 10.1016/j.molimm.2010.02.007.
[86] Y. Wang et al., “Properdin Contributes to Allergic Airway In�ammation through Local C3aGeneration.,” Journal of immunology (Baltimore, Md. : 1950), vol. 195, no. 3, pp. 1171–1181, Aug. 2015,doi: 10.4049/jimmunol.1401819.
[87] D. Pauly et al., “A novel antibody against human properdin inhibits the alternative complementsystem and speci�cally detects properdin from blood samples.,” PloS one, vol. 9, no. 5, p. e96371, 2014,doi: 10.1371/journal.pone.0096371.
[88] Y. Ueda et al., “Blocking Properdin Prevents Complement-Mediated Hemolytic Uremic Syndrome andSystemic Thrombophilia.,” Journal of the American Society of Nephrology : JASN, vol. 29, no. 7, pp.1928–1937, Jul. 2018, doi: 10.1681/ASN.2017121244.
[89] M. Noris and G. Remuzzi, “Glomerular Diseases Dependent on Complement Activation, IncludingAtypical Hemolytic Uremic Syndrome, Membranoproliferative Glomerulonephritis, and C3Glomerulopathy: Core Curriculum 2015.,” American journal of kidney diseases : the o�cial journal of theNational Kidney Foundation, vol. 66, no. 2, pp. 359–375, Aug. 2015, doi: 10.1053/j.ajkd.2015.03.040.

Page 25/32
[90] P. F. Zipfel et al., “The role of complement in C3 glomerulopathy.,” Molecular immunology, vol. 67, no.1, pp. 21–30, Sep. 2015, doi: 10.1016/j.molimm.2015.03.012.
[91] M. A. H. M. Michels, E. B. Volokhina, N. C. A. J. van de Kar, and L. P. W. J. van den Heuvel, “The role ofproperdin in complement-mediated renal diseases: a new player in complement-inhibiting therapy?,”Pediatric Nephrology, vol. 34, no. 8, pp. 1349–1367, 2019, doi: 10.1007/s00467-018-4042-z.
[92] J. C. Holter et al., “Systemic complement activation is associated with respiratory failure in COVID-19hospitalized patients,” Proceedings of the National Academy of Sciences, vol. 117, no. 40, pp. 25018 LP –25025, Oct. 2020, doi: 10.1073/pnas.2010540117.
[93] C. M. Wilk, “Coronaviruses hijack the complement system,” Nature Reviews Immunology, vol. 20, no.6, p. 350, 2020, doi: 10.1038/s41577-020-0314-5.
[94] M. Nangaku, “Complement Regulatory Proteins: Are They Important in Disease?,” Journal of theAmerican Society of Nephrology, vol. 14, no. 9, pp. 2411 LP – 2413, Sep. 2003, doi:10.1097/01.ASN.0000088010.15313.A1.
[95] M. Kawano, “Complement Regulatory Proteins and Autoimmunity BT - Autoimmunity,” A. Górski, H.Krotkiewski, and M. Zimecki, Eds. Dordrecht: Springer Netherlands, 2001, pp. 73–82.
[96] K. R. Machlus et al., “CCL5 derived from platelets increases megakaryocyte proplatelet formation,”Blood, vol. 127, no. 7, pp. 921–926, Feb. 2016, doi: 10.1182/blood-2015-05-644583.
[97] Y. Cong et al., “MERS-CoV pathogenesis and antiviral e�cacy of licensed drugs in human monocyte-derived antigen-presenting cells,” PloS one, vol. 13, no. 3, pp. e0194868–e0194868, Mar. 2018, doi:10.1371/journal.pone.0194868.
[98] S. Li et al., “Clinical and pathological investigation of patients with severe COVID-19,” JCI insight, vol.5, no. 12, p. e138070, Jun. 2020, doi: 10.1172/jci.insight.138070.
[99] Y. Zhao et al., “Longitudinal COVID-19 pro�ling associates IL-1RA and IL-10 with disease severity andRANTES with mild disease,” JCI insight, vol. 5, no. 13, p. e139834, Jul. 2020, doi:10.1172/jci.insight.139834.
[100] R. L. Chua et al., “COVID-19 severity correlates with airway epithelium–immune cell interactionsidenti�ed by single-cell analysis,” Nature Biotechnology, vol. 38, no. 8, pp. 970–979, 2020, doi:10.1038/s41587-020-0602-4.
[101] B. K. Patterson et al., “CCR5 inhibition in critical COVID-19 patients decreases in�ammatorycytokines, increases CD8 T-cells, and decreases SARS-CoV2 RNA in plasma by day 14,” InternationalJournal of Infectious Diseases, vol. 103, pp. 25–32, 2021, doi: https://doi.org/10.1016/j.ijid.2020.10.101.

Page 26/32
[102] F. J. Culley et al., “Role of CCL5 (RANTES) in Viral Lung Disease,” Journal of Virology, vol. 80, no. 16,pp. 8151 LP – 8157, Aug. 2006, doi: 10.1128/JVI.00496-06.
[103] M. Schaller, C. M. Hogaboam, N. Lukacs, and S. L. Kunkel, “Respiratory viral infections drivechemokine expression and exacerbate the asthmatic response,” Journal of Allergy and ClinicalImmunology, vol. 118, no. 2, pp. 295–302, Aug. 2006, doi: 10.1016/j.jaci.2006.05.025.
[104] A. E. John, A. A. Berlin, and N. W. Lukacs, “Respiratory syncytial virus-induced CCL5/RANTEScontributes to exacerbation of allergic airway in�ammation.,” European journal of immunology, vol. 33,no. 6, pp. 1677–1685, Jun. 2003, doi: 10.1002/eji.200323930.
[105] N. Bostanci et al., “Targeted Proteomics Guided by Label-free Quantitative Proteome Analysis inSaliva Reveal Transition Signatures from Health to Periodontal Disease*,” Molecular & CellularProteomics, vol. 17, no. 7, pp. 1392–1409, 2018, doi: https://doi.org/10.1074/mcp.RA118.000718.
[106] V. Taraslia et al., “A High-Resolution Proteomic Landscaping of Primary Human Dental Stem Cells:Identi�cation of SHED- and PDLSC-Speci�c Biomarkers,” International Journal of Molecular Sciences ,vol. 19, no. 1. 2018, doi: 10.3390/ijms19010158.
[107] X. Xiao et al., “Characterization of Odontogenic Differentiation from Human Dental Pulp Stem CellsUsing TMT-Based Proteomic Analysis,” BioMed Research International, vol. 2020, p. 3871496, 2020, doi:10.1155/2020/3871496.
Figures

Page 27/32
Figure 1
Characteristics and classi�cation of MERS-CoV -infected patients and clinical disease outcomes
Page 28/32
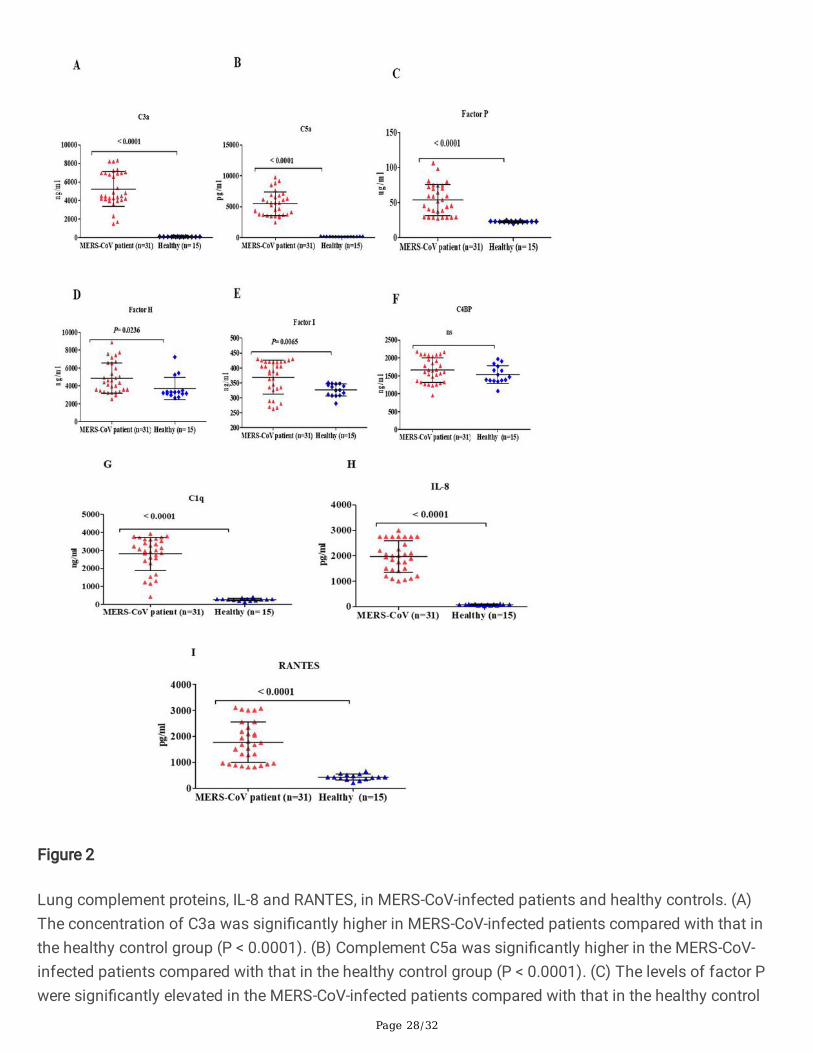
Figure 2
Lung complement proteins, IL-8 and RANTES, in MERS-CoV-infected patients and healthy controls. (A)The concentration of C3a was signi�cantly higher in MERS-CoV-infected patients compared with that inthe healthy control group (P < 0.0001). (B) Complement C5a was signi�cantly higher in the MERS-CoV-infected patients compared with that in the healthy control group (P < 0.0001). (C) The levels of factor Pwere signi�cantly elevated in the MERS-CoV-infected patients compared with that in the healthy control

Page 29/32
group (P < 0.0001). (D) The levels of factor H were elevated in the MERS-CoV-infected patients comparedwith that in the healthy control group (P < 0.0236). (E) Complement factor I was elevated in the MERS-CoV-infected patients compared with that in the healthy control group (P < 0.0065). (F) There was nostatistical signi�cance (ns) in the lung concentrations of C4BP in MERS-CoV-infected patients comparedwith that in the healthy control group. (G) The levels of C1q were signi�cantly elevated in the MERS-CoV-infected patients compared with that in the healthy control group (P < 0.0001). (H) The concentrations ofIL-8 were signi�cantly higher in the MERS-CoV-infected patients compared with that in the healthy controlgroup (P < 0.0001). (I) The levels of RANTES were signi�cantly elevated in the MERS-CoV-infectedpatients compared with that in the healthy control group (P < 0.0001).
Figure 3
Lung complement proteins, IL-8 and RANTES, in MERS-CoV-infected patients. Patients were divided intonon-survival (n = 22) and survival (n = 9) groups. (A) The concentration of C3a was elevated in the non-survival group compared with that in the survival group (P < 0.0024). (B) Complement C5a was higher inthe non-survival group compared with that in the survival group (P < 0.0001). (C) The levels of factor Pwere signi�cantly elevated in the non-survival group compared with that in the survival group (P <0.0001). (D) The levels of C4BP were signi�cantly elevated in the survival group compared with that inthe non-survival group (P < 0.0113). (E) The concentration of IL-8 was signi�cantly higher in the non-survival group compared with that in the survival group (P < 0.0001). (F) The levels of RANTES weresigni�cantly elevated in the non-survival group compared with that in the survival group (P < 0.0001).
Page 30/32
Figure 4
Actions of C3a, C5a, factor P (properdin) IL-8 and RANTES protein-protein interaction network usingCytoscape software. PPI network contains 25 nodes, 149 edges, average node degree 11.9 and the PPIenrichment p-value is < 1.0e-16. A node represents a protein, and a line represents an interaction. The linethickness represents the degree of con�dence prediction of the interaction.
Page 31/32
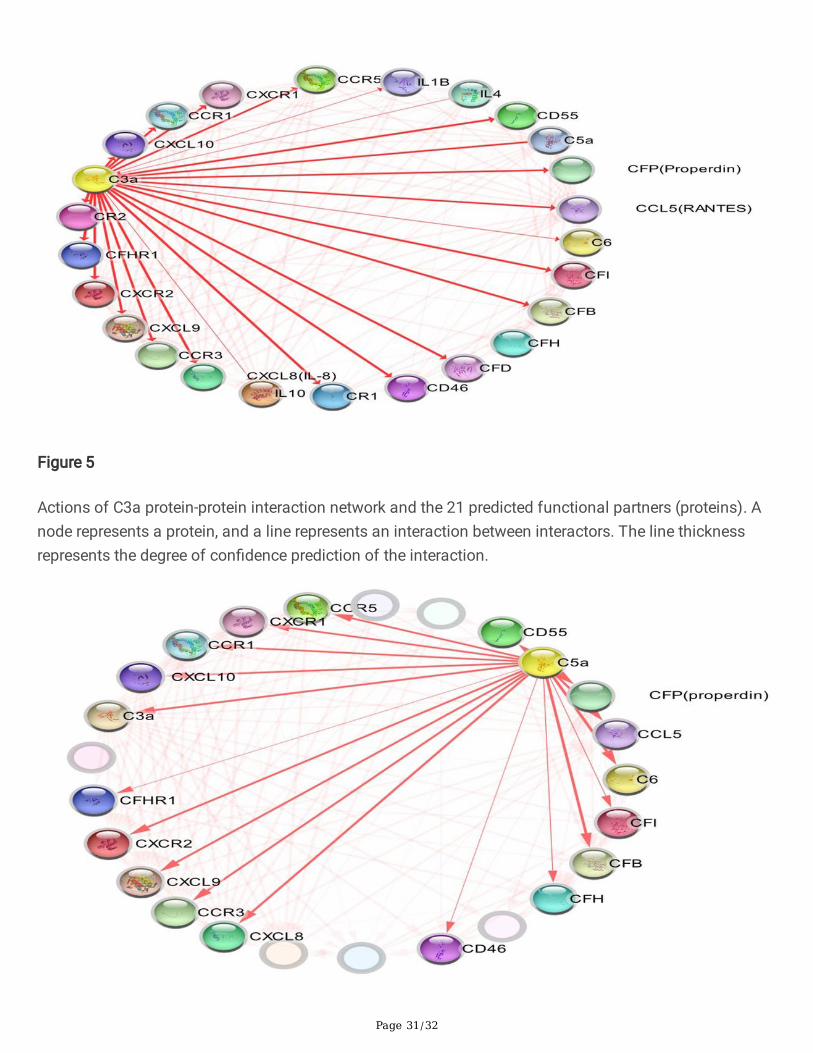
Figure 5
Actions of C3a protein-protein interaction network and the 21 predicted functional partners (proteins). Anode represents a protein, and a line represents an interaction between interactors. The line thicknessrepresents the degree of con�dence prediction of the interaction.

Page 32/32
Figure 6
Actions of C5a protein-protein interaction network and the 18 predicted functional partners (proteins). Anode represents a protein, and a line represents an interaction between interactors. The line thicknessrepresents the degree of con�dence prediction of the interaction.
Figure 7
Actions of IL-8 protein-protein interaction network and the 8 predicted functional partners (proteins). Anode represents a protein, and a line represents an interaction between interactors. The line thicknessrepresents the degree of con�dence prediction of the interaction.
Supplementary Files
This is a list of supplementary �les associated with this preprint. Click to download.
graphicsabstract.jpg










![chemokine/chemokine receptor pair ccL20/ccR6 in human ... · pancreas, stomach, prostate, testis, uterine cervix and skin[11]. The chemokine receptor CCR6 was originally described](https://static.fdocuments.in/doc/165x107/5f9ac7b0798b75658905651c/chemokinechemokine-receptor-pair-ccl20ccr6-in-human-pancreas-stomach-prostate.jpg)







