Characterization of SoybeanEndopeptidase Activity ...SOYBEAN ENDOPEPTIDASES t A 6 'RI O1 30 60 90...
6
Plant Physiol. (1983) 72, 345-350 0032-0889/83/72/0345/06/$00.50/0 Characterization of Soybean Endopeptidase Activity Using Exogenous and Endogenous Substrates' Received for publication October 21, 1982 and in revised form January 19, 1983 HEATHER M. BOND AND DIANNA J. BOWLES Department of Biochemistry, University of Leeds, Leeds, LS2 9JT United Kingdom ABSTRACT Endopeptidase activity in mature soybean seeds (Glycine max), has been measured using an exogenous substrate, 1125lliodoinsulin B chain. On the basis of pH optimum and the use of specific proteinase inhibitors, two distinct endopeptidase activities can be identified in both the embryonic axis and the cotyledons. One activity is characteristic of a neutral/alkaline metalloendopeptidase(s) and the other of an acidic carboxylendopepti- dase(s). Neither activity is membrane-bound. The metalloendopeptidase(s), most probably working with neutral expopeptidases also present in the tissues, is capable of degrading certain subunits of the storage proteins. The a subunit of conglycinin and additional seed polypeptides remain resistant to degradation. The carboxylpeptidase activity displayed a differ- ent specificity towards endogenous substrates; in particular, an acid-soluble polypeptide of apparent molecular weight 30,000 appeared to be the principal substrate for limited proteolytic degradation by the proteinase(s). Soybean agglutinin remained resistant to degradation by either class of endopeptidases. Storage reserves in soybeans are composed of specific polypep- tides localized in the cotyledons and axes of the mature seeds. Much recent work has focused on the characterization of the seed storage proteins, including the regulation of their synthesis during seed maturation and the processes that lead to their eventual deposition in the protein bodies (14, 16, 19-21). In contrast, very little information is available concerning the mechanism of soy- bean storage protein degradation and the events that regulate germination and mobilization of reserves during the early stages of seedling growth (9). As a first step into this complexity, we have now characterized the endopeptidase activity that exists in cotyledons and axes of mature soybean seeds. We present evidence to indicate there are two distinct classes of endopeptidases, one with an acidic pH optimum and characteristics of carboxylpeptidase activity, the other with a neutral/alkaline pH optimum and typical of metal- loendopeptidase activity. The neutral metalloendopeptidase activ- ity, working most probably in conjunction with exopeptidases also present in the tissue, is able to degrade the storage polypeptides in vitro with the exception of the ,8-subunit of conglycinin and the basic subunits of glycinin. MATERIALS AND METHODS Soybean seeds (Glycine max var. Williams) were provided by the US Intsoy program 1979, with a germination efficiency of 'This work was financially supported by an Science Research Council Grant A 97783 and B 73088 to Dr Dianna J. Bowles. 100%. I251-labeled B chain of insulin was a gift from Dr. A. J. Kenny. All chemicals, unless otherwise stated, were from Sigma. Soybean agglutinin was prepared from affinity chromatography as described in Allen and Neuberger (2). Preparation of Soybean Seed Extracts. For determination of endopeptidase activity in dry seeds, cotyledons were dissected and ground first to a fine powder in a coffee grinder. Otherwise, soybeans were soaked for 4 h in H20 at room temperature and dissected into cotyledons and axes. The material from either dry or imbibed seeds was then homogenized in a pestle and mortar at the fresh weight: buffer ratio of 1 g: 10 ml. The buffers used were PBS,2 100 mm Hepes/NaOH pH 8.0, or 100 mm NaCH3COO/ CH3COOH, pH 4.0. The homogenates were sonicated on ice (4 x 10 s, using a microprobe fitted to a Dawe Sonicator), centrifuged for 5 min on a bench centrifuge to provide a debris-free superna- tant which was used to assay for protein and endopeptidase activity. Endopeptidase Assay. [125JIlodoinsulin B chain is hydrolyzed to release TCA-soluble radioactive peptides. The assay was carried out essentially as described in Barrett (4) and George and Kenny (13). For determination of pH optima, the following buffers were used: pH 2.3, 3.0, and 4.0, 100 mm NaCH3COO/CH3COOH; pH 5 and 6, 100 mm Mes/HCl; pH 7 and 8, 100 mm Hepes/NaOH; pH 9 and 10, 100 mm Tris/HCl. The assay contained 200 pl of appropriate buffer, 50 ,l of extract, and H20 and/or potential inhibitors to a final volume of 275 1d. The reaction was started with 25 ,ul 1251-labeled B chain at 5.7 nmol/ml. After the incubations at temperature and time specified, the reaction was terminated by the addition of 500 ,ul TCA (25% w/v in H20) and 250 id casein (2% w/v in H20). After centrifugation at l0,000g for 3 min (Eppendorf Microfuge), 500-Al aliquots were removed and counted in a y-counter and the per cent degradation of '25I-B chain was measured. After obtaining total radioactivity present and TCA-soluble radioactivity, in the absence of enzyme/ extract, nmol 125I-B chain degraded was calculated as described in George and Kenny (13). When measuring the effect of potential inhibitors on endopep- tidase activity, the extract, appropriate buffer, and inhibitor were preincubated for 2 h at 4°C prior to addition of 1251I-B chain and start of assay. Aminopeptidase Assay. Activity was measured fluorometrically using AMC derivatives of amino acids as substrates in an assay essentially the same as that used for 2-naphthylamide derivatives (4). The fluorimeter cell contained 1.9 ml of buffer substrate mixture equilibrated at 37°C. Buffer substrates used were: Ala- AMC, 0.2 mm in 100 mm Tris/HCl, pH 7.0; Gly-Pro-AMC, 0.2 2Abbreviations: PBS, phosphate-buffered saline (10 mm KH2PO4/ K2HPO4, 145 mm NaCl, pH 7.4), pCMB, p-chloromercuribenzoic acid; PMSF, phenylmethylsulfonyl fluoride; AMC, 7-amino-4-methylcoumarin; STI, soybean trypsin inhibitor, SBA, soybean agglutinin; PAGE, poly- acrylamide gel electrophoresis. 345 https://plantphysiol.org Downloaded on March 30, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
Transcript of Characterization of SoybeanEndopeptidase Activity ...SOYBEAN ENDOPEPTIDASES t A 6 'RI O1 30 60 90...
-
Plant Physiol. (1983) 72, 345-3500032-0889/83/72/0345/06/$00.50/0
Characterization of Soybean Endopeptidase Activity UsingExogenous and Endogenous Substrates'
Received for publication October 21, 1982 and in revised form January 19, 1983
HEATHER M. BOND AND DIANNA J. BOWLESDepartment of Biochemistry, University of Leeds, Leeds, LS2 9JT United Kingdom
ABSTRACT
Endopeptidase activity in mature soybean seeds (Glycine max), has beenmeasured using an exogenous substrate, 1125lliodoinsulin B chain. On thebasis of pH optimum and the use of specific proteinase inhibitors, twodistinct endopeptidase activities can be identified in both the embryonicaxis and the cotyledons. One activity is characteristic of a neutral/alkalinemetalloendopeptidase(s) and the other of an acidic carboxylendopepti-dase(s). Neither activity is membrane-bound. The metalloendopeptidase(s),most probably working with neutral expopeptidases also present in thetissues, is capable of degrading certain subunits of the storage proteins.The a subunit of conglycinin and additional seed polypeptides remainresistant to degradation. The carboxylpeptidase activity displayed a differ-ent specificity towards endogenous substrates; in particular, an acid-solublepolypeptide of apparent molecular weight 30,000 appeared to be theprincipal substrate for limited proteolytic degradation by the proteinase(s).Soybean agglutinin remained resistant to degradation by either class ofendopeptidases.
Storage reserves in soybeans are composed of specific polypep-tides localized in the cotyledons and axes of the mature seeds.Much recent work has focused on the characterization of the seedstorage proteins, including the regulation of their synthesis duringseed maturation and the processes that lead to their eventualdeposition in the protein bodies (14, 16, 19-21). In contrast, verylittle information is available concerning the mechanism of soy-bean storage protein degradation and the events that regulategermination and mobilization of reserves during the early stagesof seedling growth (9).As a first step into this complexity, we have now characterized
the endopeptidase activity that exists in cotyledons and axes ofmature soybean seeds. We present evidence to indicate there aretwo distinct classes of endopeptidases, one with an acidic pHoptimum and characteristics of carboxylpeptidase activity, theother with a neutral/alkaline pH optimum and typical of metal-loendopeptidase activity. The neutral metalloendopeptidase activ-ity, working most probably in conjunction with exopeptidases alsopresent in the tissue, is able to degrade the storage polypeptides invitro with the exception of the ,8-subunit of conglycinin and thebasic subunits of glycinin.
MATERIALS AND METHODS
Soybean seeds (Glycine max var. Williams) were provided bythe US Intsoy program 1979, with a germination efficiency of
'This work was financially supported by an Science Research CouncilGrant A 97783 and B 73088 to Dr Dianna J. Bowles.
100%. I251-labeled B chain of insulin was a gift from Dr. A. J.Kenny. All chemicals, unless otherwise stated, were from Sigma.Soybean agglutinin was prepared from affinity chromatographyas described in Allen and Neuberger (2).
Preparation of Soybean Seed Extracts. For determination ofendopeptidase activity in dry seeds, cotyledons were dissected andground first to a fine powder in a coffee grinder. Otherwise,soybeans were soaked for 4 h in H20 at room temperature anddissected into cotyledons and axes. The material from either dryor imbibed seeds was then homogenized in a pestle and mortar atthe fresh weight: buffer ratio of 1 g:10 ml. The buffers used werePBS,2 100 mm Hepes/NaOH pH 8.0, or 100 mm NaCH3COO/CH3COOH, pH 4.0. The homogenates were sonicated on ice (4x 10 s, using a microprobe fitted to a Dawe Sonicator), centrifugedfor 5 min on a bench centrifuge to provide a debris-free superna-tant which was used to assay for protein and endopeptidaseactivity.
Endopeptidase Assay. [125JIlodoinsulin B chain is hydrolyzed torelease TCA-soluble radioactive peptides. The assay was carriedout essentially as described in Barrett (4) and George and Kenny(13). For determination of pH optima, the following buffers wereused: pH 2.3, 3.0, and 4.0, 100 mm NaCH3COO/CH3COOH; pH5 and 6, 100 mm Mes/HCl; pH 7 and 8, 100 mm Hepes/NaOH;pH 9 and 10, 100 mm Tris/HCl. The assay contained 200 pl ofappropriate buffer, 50 ,l of extract, and H20 and/or potentialinhibitors to a final volume of 275 1d. The reaction was startedwith 25 ,ul 1251-labeled B chain at 5.7 nmol/ml.
After the incubations at temperature and time specified, thereaction was terminated by the addition of 500 ,ul TCA (25% w/vin H20) and 250 id casein (2% w/v in H20). After centrifugationat l0,000g for 3 min (Eppendorf Microfuge), 500-Al aliquots wereremoved and counted in a y-counter and the per cent degradationof '25I-B chain was measured. After obtaining total radioactivitypresent and TCA-soluble radioactivity, in the absence ofenzyme/extract, nmol 125I-B chain degraded was calculated as described inGeorge and Kenny (13).When measuring the effect of potential inhibitors on endopep-
tidase activity, the extract, appropriate buffer, and inhibitor werepreincubated for 2 h at 4°C prior to addition of 1251I-B chain andstart of assay.
Aminopeptidase Assay. Activity was measured fluorometricallyusing AMC derivatives of amino acids as substrates in an assayessentially the same as that used for 2-naphthylamide derivatives(4). The fluorimeter cell contained 1.9 ml of buffer substratemixture equilibrated at 37°C. Buffer substrates used were: Ala-AMC, 0.2 mm in 100 mm Tris/HCl, pH 7.0; Gly-Pro-AMC, 0.2
2Abbreviations: PBS, phosphate-buffered saline (10 mm KH2PO4/K2HPO4, 145 mm NaCl, pH 7.4), pCMB, p-chloromercuribenzoic acid;PMSF, phenylmethylsulfonyl fluoride; AMC, 7-amino-4-methylcoumarin;STI, soybean trypsin inhibitor, SBA, soybean agglutinin; PAGE, poly-acrylamide gel electrophoresis.
345 https://plantphysiol.orgDownloaded on March 30, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
https://plantphysiol.org
-
Plant Physiol. Vol. 72, 1983
mm in 100 mm Tris/HCl, pH 8.0; Glu-AMC, 0.2 mm Tris/HCl,pH 7.0, containing 1.25 mm CaCl2. The reaction was started byaddition of 100 I1 of soybean extract (made in Hepes/NaOH, pH8.0, at a protein concentration of 10 mg/ml). The increase inemission was measured over 3 min in the cell maintained at 37°C(excitation, 370 nm, slit 0.5; emission, 442, slit 2).
General Methods. Protein was determined using the Coomassiedye method (26). For analysis of polypeptides using polyacryl-amide electrophoresis in the presence of SDS, samples wereprecipitated using 12.5% (w/v) TCA (final concentration) andresolubilized in sample buffer and processed using 10 to 15% (w/v) gradient gels as described in Bowles and Marcus (7). Ultracen-trifugation of the extracts was performed using an SW 50.1 rotortype in a Kontron ultracentrifuge at 4°C for 30 min at 100,000g.Hemagglutination assays using trypsin-treated rabbit erythrocytes,were performed as described in Bowles and Marcus (7).
RESULTS
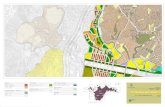
Characterization of Endopeptidase Activity Using an ExogenousSubstrate. The initial experiments were based on the ability of atissue extract to hydrolyze an exogenous 125I-labeled substrate ina standard endopeptidase assay. Using this approach, severalparameters of endopeptidase activity in soybeans were examined.Figure 1 shows the pH optima of activity in extracts preparedfrom axes (A) and cotyledons (B) of 4-h-imbibed seeds. For bothtissues, two distinct optima exist, one at pH 4.0 and the other atpH 8.0. At either pH, the specific activity is higher in the axis thanin the cotyledon. A time course for degradation of the exogenoussubstrate is shown in Figure 2. Hydrolysis of 1251-B chain insulinis linear up to 50 min at 37°C.A range of additives were included in the standard assay to gain
an insight into the class of endopeptidase activity present in thetissues. Table I shows the results of the additions to tissue extractsprepared at pH 4.0 and 8.0. Cotyledon and axis extracts respondin a similar manner to all the additives. However, a major distinc-tion exists in the response to the activity measured at pH 4.0compared with that measured at pH 8.0. Endopeptidase activityat pH 8.0 is inhibited by a range of chelators and metal ionsincluding 1,10-phenanthroline, EDTA, Cu2+, and Zn2+. This pat-tern of inhibition is typical of that shown by metalloendopepti-dases which generally exhibit a pH optimum in the range of 7 to9 (4). The inhibition by pCMB and DTT also indicates theinvolvement of thiol groups in the activity. PMSF interacts bothwith thiol groups and serine proteases (4). Since other serineprotease inhibitors were without effect on the activity at pH 8.0,PMSF inhibition may also be related to the involvement of thiolgroups. Certain metal ions including Ca2" and Mn2+ stimulatedactivity. However, since the tissue extracts most probably con-tained endogenous metal ions and chelators, the specific interac-tion between the enzyme and metal ions must await the purifica-tion and detailed analysis of the enzyme(s). In contrast to thepattern of inhibition at pH 8.0, the activity measured at pH 4.0was unaffected by metal ions and chelators but was abolished bypepstatin A. The pH optimum and specific inhibition are typicalof carboxylpeptidases that generally have a pH range of 2 to 5.Aprotinin was also able to inhibit the activity measured at pH 4.0,although it is generally assumed to be specific for trypsin andserine proteases. However, other serine proteinase inhibitors in-cluding benzamidine were without effect on the activity. Neitherthe presence of excess SBA nor its removal by passage of theextracts through an affinity matrix affected endopeptidase activityat either pH optimum. It is therefore unlikely that the lectin isdirectly involved through its carbohydrate-binding site with theregulation of endopeptidase activity.
Effect of Endopeptidase Activity on Endogenous Substrates. Itwas of interest to determine the effect of endopeptidase activityon soybean proteins. Extracts prepared from cotyledons and axes
.' 0
150
.1
S so
Co
50-
c 44
IC.15
R la
*e5
gopH
B
I I I I I I I I2 3 4 5 6 7 8 9 10
pHFIG. 1. pH optima of endopeptidase activity in extracts of soybean
axes and cotyledons. Extracts were prepared from the embryonic axis (A)and the cotyledon tissue (B) in PBS as in "Materials and Methods" andassayed at 37°C for 15 min, using ['"Iliodoinsulin B chain as a substrateover the pH range 2.3 to 10.0.
were incubated for 24 h at 30°C at either pH 4.0 or 8.0 in thepresence/absence of inhibitors of endopeptidase activity. Usingthe exogenous "2I-labeled substrate, it was shown that both classesof endopeptidase activity were stable for 24 h at 30°C and theeffect of the inhibitors was maintained throughout the incubation(results not shown). After the incubation period, samples wereanalyzed by polyacrylamide gel electrophoresis in the presence ofSDS and compared with the polypeptide profile of the tissueextracts at time 0. The results for cotyledon extracts are shown inFigure 3 and those for embryonic axis extracts are in Figure 4.The extract of the cotyledons prepared at pH 8.0 and time 0 (Fig.3, lane A) contains the subunits of the major storage polypeptides:glycinin (llS: A-4, acidic, and basic subunits) and conglycinin(7S: a'-, a-, and fl-subunits). Extraction of the cotyledons at pH
I I I
346 BOND AND BOWLES
https://plantphysiol.orgDownloaded on March 30, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
https://plantphysiol.org
-
SOYBEAN ENDOPEPTIDASES
A
t 6
'RIO1
30 60 90 120Minutes
B
5
.a 4 ~ 03
30 60 90 120Minutes
FIG. 2. Time course of endopeptidase activity in extracts of soybeanaxes and cotyledons. Extracts were prepared from the axis (A), andcotyledon (B) at pH 8.0 or 4.0 and as described in "Materials and Methods"were assayed for endopeptidase activity using ['25Iliodoinsulin B chain asa substrate for times up to 2 h at 370C.
4.0, time 0, (Fig. 3, lane E) causes the selective precipitation of thestorage proteins and much other material (-75% of the totalprotein) resulting in an enrichment in certain acid-soluble poly-peptides including urease.
Incubation of the cotyledon extract at pH 8.0 for 24 h in theabsence of endopeptidase inhibitors causes a massive alteration tothe polypeptide profile (Fig. 3, lane B). The A-4 and acidicglycinin subunits and the a'- and a-subunits of conglycinin areabsent. Many new low mol wt bands appear, including prominent
Table I. Effects ofAdditives on Endopeptidase Activity in ExtractsPreparedfrom Soybean Cotyledons and Axes
Extracts prepared at pH 4.0 or 8.0 were preincubated at 4°C for 2 h inthe presence of different additives, before inclusion of ['25Iiodoinsulin Bchain. Endopeptidase activity was then measured over a 30-min period asdescribed in "Materials and Methods." TIU, trypsin-inhibiting units.
Cotyledon Embryonic Axis
Additive Concn. of Ho saInhibitor pH of assay
4.0 8.0 4.0 8.0
% activity remainingNo addition 100 100 100 100CuCl2 2 mM 102.3 0 69.1 0ZnCl2 2 mm 93.9 68.6 77.4 38.0MnCl2 2 mM 127.0 183.0 81.3 115.0MgCl2 2 mM 96.5 103.1 65.4 116.0CaCl2 2 mm 110.6 176.0 60.9 92.9EDTA 5 mM 100.7 10.7 79.7 62.4DTT 2 mM 100.5 12.5 79.4 45.71,10-Phenanthroline 5 mm 45.2 0 70.7 0pCMB I mM 87.0 0 63.5 0lodoacetimide 2 mm 106.2 95.5 69.8 108.0PMSF I mm 49.4 52.0 39.3 84.9STI 250 yg/ml 109.4 107.8 96.2 94.0Benzamidine I mm 100.3 99.6 113.1 102.0Aprotinin 2.83 TIU/ml 10.4 63.9 0 103.9Pepstatin A 5 ,tg/ml 2.5 109.0 0 123.7SBA 250 tig/ml 109.4 107.8 96.2 94.2Minus SBA 89.8 109.9 100.6 91.3
ones at apparent mol wt 48,000, 33,000, 29,000 and 26,000. Theresults suggest that the fl-subunit of conglycinin is unaffected bythe incubation as are the basic subunits of glycinin. However, thepossibility cannot be ruled out that degradation products of iden-tical apparent mol wt to these subunits are formed and cannot bedistinguished on a one-dimensional gradient gel. Inclusion ofeither 1 mM pCMB (Fig. 3, lane C) or 5 mm 1,10-phenanthroline(Fig. 3, lane D) completely prevents degradation and the polypep-tide profiles of the incubations with either of these inhibitorspresent remain unchanged from that at time 0.The effect of a 24-h incubation of the cotyledon extract at pH
4.0, in the absence ofendopeptidase inhibitors, is far less dramatic(Fig. 3, lane F). The apparent mol wt of urease may be slightlyreduced, but certain other unidentified polypeptides disappearcompletely, including a prominent band at apparent mol wt 30,000and several minor ones. A new lower apparent mol wt bandappears at 23,400. In the presence of either 5 jg/ml pepstatin or10%o (v/v) aprotinin, the polypeptide at apparent mol wt 30,000 ismaintained and the main degradation product is absent. The effectof the two inhibitors seems to be similar: neither is able to preventdegradation of several minor polypeptides in the extract. Thepolypeptide in Figure 3, lane G, of apparent mol wt 6,500 isaprotinin.As shown in Figure 3, the storage proteins precipitated in 100
mm acetate buffer at pH 4.0 and were generally removed bycentrifugation before assaying the effect of the acidic carboxyl-peptidase activity. In an attempt to determine whether the acidicendopeptidase could also degrade the storage proteins, a cotyledonextract prepared under neutral conditions was reduced to pH 4.0and the suspension was incubated with shaking at 30°C for 24 h.No significant alteration to the polypeptide profile occurred (re-sults not shown).The data therefore suggest that degradation of the storage
proteins of the cotyledons can occur in vitro at pH 8.0 and beprevented by inhibitors of the neutral metalloendopeptidase activ-
347
https://plantphysiol.orgDownloaded on March 30, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
https://plantphysiol.org
-
Plant Physiol. Vol. 72, 1983
-40
FiG. 3. SDS-PAGE of soybean cotyledon extracts incubated at 300C for 24 h in the presence/absence of endopeptidase inhibitors. Cotyledonextracts were prepared at pH 8.0 (A-D) and 4.0 (E-H). Time 0, A and E; 24-h incubation in the absence of inhibitors, B and F; 24 h in the presence ofinhibitors, C, D, G, and H. C, 1 mM pCMB; D, 5 mM 1,10-phenanthroline; G, 10% (v/v) aprotinin; and H, 5 pg/ml pepstatin A.
ity. However, it was possible that degradation of the polypeptidesalso involved the action ofexopeptidases which exhibited a similarpH optimum and characteristics of inhibition. As a preliminarystep into testing this possibility, several synthetic AMC-aminoacid derivatives were used to assay for aminopeptidases and theirsusceptibility to pCMB and 1, l0-phenanthroline. The results areshown in Table II and indicate that soybean cotyledon extracts docontain substantial levels of aminopeptidases which are inhibitedby 1, l0-phenanthroline and to a lesser extent by pCMB.
Figure 4 shows the effect of endopeptidase activity on embry-onic axis polypeptides. Extraction of the axis at pH 8.0 (time 0,Fig. 4, lane A) revealed a markedly different profile to that of thecotyledon. Although bands corresponding in apparent size to themajor storage proteins, glycinin and conglycinin, were present,their amounts in the axis were much lower than in the cotyledons.As before, extraction of the tissue at pH 4.0 (time 0, Fig. 4, laneE) caused precipitation of many polypeptides.The effect of a 24-h incubation of the axis extracts in the
absence of proteinase inhibitors was less dramatic than for thecotyledons but the overall pattern of change remained similar. AtpH 8.0, a- and a-subunits ofconglycinin and glycinin disappeared(Fig. 4, lane B) and were retained in the presence of proteinaseinhibitors (Fig. 4, lanes C and D). At pH 4.0, the major substratefor proteolytic activity might again be a polypeptide at apparentmol wt 30,000, inasmuch as the most notable change in the extractwas the appearance ofa probable degradation product at apparentmol wt 23,400 (Fig. 4, lane F) and its absence if the incubationcontained inhibitors (Fig. 4, lanes G and H).SDS-PAGE analysis provided an overall insight into the sus-
ceptibility of abundant proteins to degradation by endogenousproteinases. However, SBA which constitutes only
-
SOYBEAN ENDOPEPTIDASES
A B C D E F G Hw _ WIW
Table III. The Effect of Ultracentrifugation on the Solubility ofEndopeptidase Activities in Extracts Preparedfrom Soybean CotyledonsCotyledons were homogenized at pH 4.0 or 8.0 and centrifuged at low
speed to obtain a debris-free extract as described in "Materials andMethods." This extract was then further centrifuged at l00,000g for 30min at 4VC. Endopeptidase activities before and after ultracentrifugationwere assayed using I'25lliodoinsulin B chain as a substrate.
Precentrifugation Exract PostcentrifugationCotyledon Supernatant
pH of Preparation B Chain o B ChainProtein Hydrolyzed Protei Hydrolyzed
mglml nmoi/min.mg mg/ml nmol/min mgprotein protein8.0 7.6 0.126 6.2 0.1544.0 2.4 0.562 1.6 0.5 12
i
FIG. 4. SDS-PAGE of soybean axes extracts incubated at 30'C for 24hours in the presence/absence of endopeptidase inhibitors. Axes extractswere prepared at pH 8.0 (A-D) and 4.0 (E-H). Time 0, A and E; 24-hincubation in the absence of inhibitors, B and F; 24 h in the presence ofinhibitors, C, D, G, and H. C, I mm pCMB; D, 5 mM 1,10-phenanthroline;G, 10%1b (v/v) aprotinin; and H, 5 jig/ml pepstatin A.
Table II. Aminopeptidase Activities in Extracts Preparedfrom SoybeanCotyledons
Exogenous 7-amino-4-methylcoumarin amino acid derivatives wereused as substrates and assayed fluorimetrically using a soluble extractprepared from 4-h-imbibed seeds homogenized in 0.1 M Hepes, pH 8.0, asdescribed in "Materials and Methods."
Substrate Inhibitor AMC Hydrolyzed
mM nmol/min .mgproteinAla-AMC 6.28
l,1O-Phenanthroline (5) 0.40pCMB (1) 3.00
Gly-Pro-AMC 0.2531, 10-Phenanthroline (5) 0.16pCMB (1) 0.216
a-Glu-AMC 0.0841, 10-Phenanthroline (5) 0.028pCMB (1) 0.022
change in TCA solubility of iodinated residues (13). The natureof the substrate is such that it is specific for endo- rather thanexopeptidase activity and has been used widely to characterizeproteinases of mammalian systems (4). Endopeptidases are clas-sified into distinct groups on the basis of their pH optima and,more definitively, on the effect of specific substances to inhibittheir activity (4). Using these criteria, cotyledons and axes ofsoybean contain neutral metalloendopeptidase activity and acidiccarboxylpeptidase activity.
Previously, when Na-benzoyl-DL-arginine-p-nitronilide was
used as exogenous substrate, only a single neutral (pH 8.2-8.5)peptidase activity was found (8, 15). In earlier studies (19, 22), theuse of casein as substrate indicated a heterogeneous population ofpeptidase activities, all with pH optima in the range 5.0 to 6.0 (17,27, 31). Given the limitations of these substrates and assays tospecifically detect endopeptidase activity and the absence of anydetailed inhibitor data in previous investigations, it is unclear towhat extent the present data correspond with earlier findings.A major weakness in using any exogenous substrate in vitro is
that it might not reflect proteolytic activity of relevance in vivo.Therefore, as a preliminary to enzyme purification, the inhibitorsof the metallo- and carboxylendopeptidases were used to studythe effect of the proteinases on soybean proteins. The resultsindicate that the two proteolytic activities are each able to degradespecific seed proteins in vitro. Under neutral pH conditions, inwhich the major storage proteins are soluble and the metalloen-dopeptidase(s) is active, certain storage polypeptides are com-pletely degraded whereas others remain unaltered. Thus, the a'-,a-, A4, and acidic subunits of conglycinin and glycinin, respec-tively, are degraded, whereas the ,B-subunit of conglycinin and thebasic subunits of glycinin remain intact. We suggest the differentsubunits of the storage reserves may possess distinct physiologicalfunctions in vivo. Interestingly, the ,B-subunit of conglycinin ap-pears later in seed maturation (22 d after flowering) than the a'-and a-subunits (15-17 d after flowering) (14). Although formed atseparate stages of embryogenesis, the different subunits still asso-ciate to form seven naturally occurring isomeric multimers in themature seed (27). Multimer formation may, however, only reflectan efficiency in packaging the reserves into protein bodies forstorage until germination. Evidence suggests that the carboxylpep-tidase may not be involved in the degradation of storage reserves,and, at least in the early stages of germination, some mobilizationto amino acids could occur through only the combined action ofthe metalloendopeptidase(s) and exopeptidases. The principal sub-strate for the carboxylpeptidases(s) is a prominent unidentifiedpolypeptide of mol wt 30,000 which occurs in both the axis andcotyledons. Proteolysis of the polypeptide appears to be limited inthat its disappearance coincides with the appearance of a newband of lower mol wt which could represent one of the fragmentsof the original substrate. However, tryptic maps would be neces-sary to confirm their relatedness. The carbohydrate-binding activ-ity of SBA is not affected by endopeptidase activity at either pH8.0 or 4.0. This suggests the lectin is not used as an immediatesource of nitrogen for the growing axis and confirms the proba-bility that lectins cannot be classified strictly as storage reserves inthe seed (3, 6, 11, 31).Our results indicate that the specific activity of the proteinases
in extracts prepared from axes and measured using iodoinsulin Bchain is greater than in cotyledon extracts. This seems surprising
349
https://plantphysiol.orgDownloaded on March 30, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
https://plantphysiol.org
-
Plant Physiol. Vol. 72, 1983
since the cotyledons are envisaged as the main autolytic centersduring germination and early stages of seedling growth. Onepossible explanation could be competition from endogenous sub-strates which might be at a higher level in the cotyledons than inthe axes. A comparison of activities using iodoinsulin B chainafter a 24-h incubation in vitro, during which depletion of endog-enous substrates had occurred, still revealed the axis was higherthan the cotyledon. However, it is probable that neither class ofendopeptidase is functioning optimally in the tissue extracts dueto the presence/absence ofinhibitors/cofactors, etc. and, therefore,no idea of the full proteolytic potential of the tissues will beobtained until the enzymes have been purified and antisera areavailable.The use of tissue extracts can only provide an indication of the
probable events that occur in vivo. When polypeptide profiles ofcotyledon and axis extracts prepared from 24-h-imbibed seedswere investigated by SDS-PAGE, essentially no alteration inprofiles from time 0 had occurred (results not shown). This mayreflect an acceleration in degradation brought about by the con-ditions of the in vitro incubation. Alternatively, it is possible thatneither endopeptidase activity present in mature soybeans is re-sponsible for the physiological mobilization of reserves duringgermination. For example, in the two well characterized systemsof mung bean and castor bean, the proteinases responsible formobilization of storage reserves are only synthesized 2 to 3 d afterseed germination (1, 5, 10, 23, 24, 28, 30). However, our dataindicate that proteinases which have been synthesized duringsoybean maturation have at least the potential to degrade specificstorage polypeptides. As yet, we have no evidence whether theprimary function ofthese proteinases is in storage protein synthesisand processing (12) or reserve mobilization.
LITERATURE CITED
1. ALPI A, H BEEVERS 1981 Protelnases and enzyme stability in crude extracts ofcaster bean endosperm. Plant Physiol 67: 499-502
2. ALLEN AK, A NEUBERGER 1975 A simple method for the preparation of anaffinity absorbent for soybean agglutinin using galactosamine and CH-Seph-arose. FEBS Lett 50: 362-363
3. BARONDES SH 1981 Lectins: their multiple endogenous cellular functions. AnnuRev Biochem 50: 207-231
4. BARRr AJ (ed) 1977 Proteinases in mammalian cells and tissues. In ResearchMonographs in Cell and Tissue Physiology, Vol 2. Elsevier/North-HollandBiomedical Press, pp 1-52 (Barrett) and pp 436-440 (AJ Kenny)
5. BAUMGARTNER B, MJ CHRISPELS 1977 Purification and characterization ofvicilin peptidohydrolase the major endopeptidase in the cotyledons of mungbean seedlings. Eur J Biochem 77: 223-233
6. BowLEs DJ 1982 The enigma of seed lectins. Recherche 13: 784-785
7. BowLEs DJ, S MARCUS 1981 Characterization of receptors for the endogenouslectins of soybean and jackbean seeds. FEBS Lett 129: 135-138
8. CATSIMPOOLAS N, SK FUNK, J WANG, J KENNY 1972 Isoelectric fractionationand some properties of a proteases from soybean seeds. J Sci Food Agric 22:79-82
9. CATSIMPOOLAS N, TG CAMPBELL, EW MEYER 1968 Immunochemical study ofchanges in reserve proteins of germinating soybean seeds. Plant Physiol 43:799-805
10. CHRIsPEELs MJ, B BAUMGARTNER, N HARRIS 1976 Regulation of reserve proteinmetabolism in the cotyledons of mung bean seedlings. Proc Natl Acad SciUSA 73: 3168-3 172
11. DALKIN K, S MARCUS, DJ BowLEs 1983 Endopeptidase activity in jackbeansand its effect on Concanavalin A. Planta 157. In press .
12. EREKEN-TuM[ER N, JD RICHTER, NC NIELSEN 1982 Structural characterizationof the glycinin precursors. J Biol Chem 257: 4016-4018
13. GEORGE SG, AJ KENNY 1973 Studies on the enzymology ofpurified preparationsof brush border from rabbit kidney. Biochem J 134: 43-57
14. GAYLER KR, GE SYKEs 1981 ,B-Conglycinins in developing soybean seeds. PlantPhysiol 67: 958-961
15. GRAF G, RE HOAGLAND 1969 Partial purification and characterization of anamidohydrolase from soybean. Phytochemistry 8: 827-830
16. HILL JE, RW BRIDENBACH 1974 Proteins of soybean seeds. Plant Physiol 53:742-746
17. INAGAKI T, M KATAYAmA, K MIuRA 1979 Partial purification and properties ofproteases from defatted soybean flour. J Nutr Sci 25: 333-342
18. LAUFER S, H TAUBER, CP DAvIS 1944 The amylolytic and proteolytic activity ofsoybean seeds. Cereal Chem 21: 267-274
19. MOREIA MA, MA HERmODSON, BA LARNS, NC NIELSEN 1979 Partial charac-terization of the acidic and basic polypeptides of glycinin. J Biol Chem 254:9921-9926
20. MOREIA MA, MA HERMODSON, BA LARKINS, NC NIELSEN 1981 Comparison ofthe primary structure of the acidic polypeptides of glycinin. Arch BiochemBiophys 210: 633-642
21. MORI T, Y WAHABAYASHI, S TAKAGI 1978 Occurrence of mRNA for storageprotein in dry soybean seeds. J Biochem 84: 1103-1111
22. OFELT CW, AK SMITH, JM MILLS 1955 Proteases of the soybean. Cereal Chem32: 53-63
23. NISHIMURA M, H BEEVERS 1978 Hydrolases in vacuoles from castor beanendosperm. Plant Physiol 62: 4448
24. NISHIMURA M, H BEEVERS 1979 Hydrolysis of protein in vacuoles isolated fromhigher plant tissue. Nature 277: 412413
25. PINSKY A, S GROSSMAN 1969 Proteases of soybean. II. Specificity of the activefractions. J Sci Fd Agric 20: 374-376
26. SEDMARK JJ, SE GROSSBERG 1977 A rapid, sensitive, and versatile assay forprotein using Coomassie brilliant blue G250. Anal Biochem 79: 544-552
27. SYKEs EK, KR GAYLER 1981 Detection and characterization of a new fi-congly-cinin from soybean seeds. Arch Biochem Biophys 210: 525-530
28. TULLY RE, H BEEVERS 1978 Proteases and peptidases of castor bean endosperm:enzyme characterization and changes during germination. Plant Physiol 62:746-750
29. WEIL J, A PINSKY, S GROSSMAN 1966 The proteases of soybean. Cereal Chem43: 392-399
30. WILDEN WV, NR GILKES, MJ CHRISPEELS 1980 The endoplasmic reticulum ofmung bean cotyledons role in the accumulation of hydrolases in protein bodiesduring seedling growth. Plant Physiol 66: 390-394
31. YOULE RJ, AHC HUANG 1976 Protein bodies from the endosperm of castorbean. Plant Physiol 58: 703-709
350 BOND AND BOWLES
https://plantphysiol.orgDownloaded on March 30, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
https://plantphysiol.org



![special stroller · 2020. 6. 9. · RACER ® special stroller ... X Backrest tilt angle [°] 90–140 90–135 90–140 90–140 Z Footplate tilt angle [°] 100–120 100–125 100–120](https://static.fdocuments.in/doc/165x107/5fe3fef5034a18344b68cb0d/special-stroller-2020-6-9-racer-special-stroller-x-backrest-tilt-angle.jpg)