
Characterization of Pulmonary T Cell Response to Helper ... · Characterization of Pulmonary T Cell...
12
of December 1, 2018. This information is current as Vectors following Intranasal Delivery Response to Helper-Dependent Adenoviral Characterization of Pulmonary T Cell Rahul Kushwah, Huibi Cao and Jim Hu http://www.jimmunol.org/content/180/6/4098 doi: 10.4049/jimmunol.180.6.4098 2008; 180:4098-4108; ; J Immunol References http://www.jimmunol.org/content/180/6/4098.full#ref-list-1 , 17 of which you can access for free at: cites 46 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2008 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on December 1, 2018 http://www.jimmunol.org/ Downloaded from by guest on December 1, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of Characterization of Pulmonary T Cell Response to Helper ... · Characterization of Pulmonary T Cell...

of December 1, 2018.This information is current as
Vectors following Intranasal DeliveryResponse to Helper-Dependent Adenoviral Characterization of Pulmonary T Cell
Rahul Kushwah, Huibi Cao and Jim Hu
http://www.jimmunol.org/content/180/6/4098doi: 10.4049/jimmunol.180.6.4098
2008; 180:4098-4108; ;J Immunol
Referenceshttp://www.jimmunol.org/content/180/6/4098.full#ref-list-1
, 17 of which you can access for free at: cites 46 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2008 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on D
ecember 1, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Characterization of Pulmonary T Cell Response toHelper-Dependent Adenoviral Vectors followingIntranasal Delivery1
Rahul Kushwah,*† Huibi Cao,* and Jim Hu2*†
In spite of the extensive research in the field of gene therapy, host immune responses continue to be the major barrier intranslating basic research to clinical practice. Helper-dependent adenoviral (HD-Ad) vectors show great potential for pul-monary gene therapy, but the knowledge of pulmonary immune responses toward these vectors is very limited. In this study,we show that HD-Ad vectors are potent stimulators of dendritic cell (DC) maturation, thus leading to stimulation of T cellproliferation with �6% of naive CD4� T cells from pulmonary mediastinal lymph node responding to HD-Ad-treated DCs.In contrast to the belief that HD-Ad vectors are unable to prime adaptive immune response, we show for the first time,through in vivo pulmonary studies in mice, that HD-Ad vectors can prime CD4� and CD8� T cell responses in the lung athigh and substantially low doses. This indicates cross-presentation of HD-Ad-derived epitopes by DCs to prime CD8� T cellresponses. To assess the basis of pulmonary T cell response against HD-Ad vectors, we examined the response of conventionalDCs (cDCs) and plasmacytoid DCs (pDCs) in the lung. In response to HD-Ad delivery, there is induction of maturation inboth cDC and pDC subsets, but it is the cDCs, not pDCs, that migrate rapidly to draining lymph nodes within the first 2 daysafter vector delivery to prime adaptive immune response against these vectors. These findings have implications for devel-opment of strategies to prevent adaptive immune responses against gene therapy vectors. The Journal of Immunology, 2008,180: 4098 – 4108.
T he ability of adenoviral-derived vectors to transduce awide variety of cells, both dividing and nondividing, hasled to their development as an efficient system for pul-
monary gene transfer (1). However, the problems of host adaptiveand innate immune responses have limited the use of adenoviralvectors for gene therapy, because of concern for their safety andefficacy in vivo. Over the last decade, immense research has beenconducted on improving the vector design, which has resulted indevelopment of helper-dependent adenoviral (HD-Ad)3 vectors,which do not encode any viral genes. The only viral sequencespresent are the terminal repeats along with the packaging signal(2). This has resulted in HD-Ad vectors being selected as the vec-
tor of choice for pulmonary gene therapy for diseases such ascystic fibrosis (3–5).
Due to the lack of viral coding sequences, HD-Ad vectors arethought to be less immunogenic than the first generation vec-tors, and it is widely believed that HD-Ad vectors do not inducean adaptive immune response (6 – 8). However, other studieshave shown that with readministration of HD-Ad vector parti-cles, there is a decrease in transgene expression correlating withan increasing Ab titer against the virus, indicating that theremay be an adaptive response being mounted against HD-Adparticles (9). Nevertheless, none of the studies to date haveassessed the ability of HD-Ad particles to induce T cell immuneresponse upon airway delivery (9, 10). Among gene therapyvectors, adeno-associated virus (AAV)-derived vectors arethought to be the least immunogenic (11). However, in recentclinical trials using AAV vectors, there has been appearance ofcytotoxic CD8� T cells upon vector delivery (12). These ob-servations clearly highlight the importance of studying adaptiveimmune responses to the so-called “less-immunogenic” vectorssuch as HD-Ad vectors. The ability to develop safe and effec-tive gene therapy therapeutics depends on having a better un-derstanding of the immunological processes in the lung, fortargeting/inhibition of these processes will help in increasingthe efficacy and persistence of gene therapy vectors.
Dendritic cells (DCs) are potent APCs and lung dendriticcells are ideally positioned in the airway epithelium network toperform surveillance of the inhaled Ags (13–15). Upon uptakeof foreign Ags, airway DCs migrate to the T cell zone of thedraining lymph nodes, in particular the mediastinal lymph node(MLN), where they interact with naive T cells and prime adap-tive immune response (16, 17). Despite the critical role of air-way DCs in modulating immune responses to inhaled Ags, toour best knowledge, studies have not examined the effects ofHD-Ad vectors on pulmonary DCs. Moreover, pulmonary DCs
*Physiology and Experimental Medicine Research Program, Hospital for Sick Chil-dren and †Department of Laboratory Medicine and Pathobiology, University of To-ronto, Toronto, Ontario, Canada
Received for publication September 26, 2007. Accepted for publication January4, 2008.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by Operating Grants from the Canadian Institutesof Health Research, the Canadian Cystic Fibrosis Foundation, and the FoundationFighting Blindness-Canada (to J.H.). J.H. was a Canadian Cystic Fibrosis FoundationScholar and recipient of the Canadian Cystic Fibrosis Foundation Zellers Senior Sci-entist Award, and holds a Premier’s Research Excellence Award of Ontario, Canada.R.K. is a recipient of Restracomp Studentship Award from the Hospital for SickChildren.2 Address correspondence and reprint requests to Dr. Jim Hu, Physiology and Experi-mental Medicine Research Program, Hospital for Sick Children, 555 University Avenue,Toronto, Ontario, Canada M5G 1X8. E-mail address: [email protected] Abbreviations used in this paper: HD-Ad, helper-dependent adenoviral; DC, den-dritic cell; cDC, conventional DC; pDC, plasmacytoid DC; AAV, adeno-associatedvirus; MLN, mediastinal lymph node; BMDC, bone marrow-derived dendritic cell;BALF, bronchoalveolar lavage fluid; MOI, multiplicity of infection.
Copyright © 2008 by The American Association of Immunologists, Inc. 0022-1767/08/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

include both the conventional (cDC) and plasmacytoid DC(pDC) subsets, and the relative contribution of the two subsetsand their response to HD-Ad vectors has not been investigatedto date. The understanding of the pulmonary adaptive immuneresponse to HD-Ad can hold the key to development of strate-gies to prevent immune responses against vectors, and may holdthe key to understanding the reason behind sudden appearanceof unwanted immune response in many gene therapy clinicaltrials.
In this study, we assessed the pulmonary adaptive immune re-sponses to HD-Ad vectors. Because incorporation of a transgenecan further elicit/potentiate immune responses, we chose to assessimmune responses to an empty HD-Ad vector that did not encodeany transgene. We show that in contrast to the commonly heldbelief, HD-Ad vectors do have the ability to potentiate CD4� andCD8� T cell response upon pulmonary delivery at high and sub-stantially low doses. Furthermore, we also show that HD-Ad vec-tors are potent stimulators of DC activation in vitro and in vivowith cDCs playing the major role in priming T cells in the drainingMLN. Moreover, HD-Ad vector delivery also resulted in matura-tion of CD8� DCs within the draining MLN, indicating that thisparticular subset of DC may play a role in cross-presentation ofHD-Ad-derived Ags and inducing CD8� T cell proliferation. Incontrast, though pulmonary pDCs do mature upon HD-Ad deliv-ery, they do not migrate to draining MLN, perhaps acting as localsource of IFN-� to prime innate and adaptive immune responseagainst HD-Ad vectors.
Materials and MethodsAbs and other reagents
All the Abs used were directed against mouse Ags. The following Abswere purchased from eBioscience: CD86 PECy5, CD80 PE, MHC II PE,CCR7 PE, BrdU FITC, CD11c PE, mPDCA1 PE, and the following fromBD Biosciences: CD11c FITC, CD4 PECy7, CD8PECy7, CD8�PECy7,and CD3 PE. CD11b Ab was a gift from Dr. Jim Xiang (SaskatoonCancer Center, Saskatchewan). Isotype control IgGs were obtainedfrom eBiosciences and/or Serotec. CFSE was obtained from MolecularProbes and BrdU, DEAE-dextran, heparin and FITC-dextran from Sig-ma-Aldrich. GM-CSF was obtained from R&D Systems. Cell prolifer-ation ELISA based on BrdU incorporation and chemiluminescent de-tection and collagenase D was obtained from Roche. Aerrane wasobtained from Baxter.
Mice and vector delivery
C57BL/6 mice were purchased from Charles River Laboratories and main-tained as per guidelines of SickKids animal facilities. All the animal studieswere reviewed and approved by the SickKids Institutional committee forhumane use of laboratory animals. Mice that were 8–11 wks of age werelightly anesthetized by Aerrane inhalation, and a highly purified batch ofHD-Ad vector particles purified via cesium chloride density gradient cen-trifugation were delivered intranasally in a volume of 50 �l with 5 � 109
or 1 � 1011 particles in complex with DEAE-dextran for efficient delivery,as described previously (4).
Generation of bone marrow-derived dendritic cells (BMDC)and HD-Ad induced maturation
Bone marrow cells were isolated from the tibia and femur of adult mice andcultured in the presence of GM-CSF as described previously (18). On day7, weakly adherent cells were isolated, and 85–90% of the cells was con-firmed to be CD11c� DCs via FACS analysis. Preparation of “empty”HD-Ad vector particles was performed in our laboratory as described pre-viously (19). CD11c� DCs were treated with HD-Ad vectors at differentmultiplicities of infection (MOI) and after 24 h marker expression wasassessed via FACS analysis.
In vitro T cell proliferation
HD-Ad-treated CD11c� DCs were exposed to single dose of 20 gray of60Co gamma irradiation and then cultured with CD4�CD25� T cells,isolated from draining MLN via cell sorting for a period of 72 h. BrdU
solution was added during the last 12 h to label the cells, after whichproliferation was measured using BrdU cell proliferation, chemilumi-nescent ELISA, according to the manufacturer’s instructions (Roche).To calculate precursor frequencies, HD-Ad-treated DCs were culturedwith CFSE-labeled naive CD4� T cells isolated from draining MLN fora period of 7 days, after which FACS analysis was performed to assessCFSE dilution. T cell precursor frequencies were calculated from CFSEdilution data as described previously (20).
Preparation of tissue lymphocytes
At different time points after HD-Ad vector delivery, mice were sacrificedby i.p. injection of Euthanyl (Bimeda-MTC). To collect bronchoalveolarlavage fluid (BALF), mouse lungs were lavaged as described previously(9). After performing lavage, lungs were perfused with 10 ml of PBS con-taining 10 U/ml heparin via the right ventricle of the heart to remove bloodcells from the lung vasculature. Perfusion was performed until the lungsturned completely white in color. Lungs were dissected out, and after re-moval of draining MLN, lungs were minced and digested for 25 min at37°C using 250 U/ml collagenase D solution, with the addition of EDTA(10 mM final) during the last 5 min of incubation. Fragments of digestedlungs were passed through a 100-�m cell strainer (BD Biosciences), andhypotonic lysis was used to remove erythrocytes. Cells were then countedand resuspended at appropriate concentrations for different experiments.Similarly, draining MLN were digested, followed by suspension at appro-priate concentrations.
BrdU labeling
At different time points after vector delivery, mice were anesthetized usingAerrane inhalation and then BrdU was administered intranasally in a vol-ume of 50 �l at 16 mg/ml concentration. Mice were killed 24 h after BrdUdelivery, and lungs/draining MLN were isolated.
Ex vivo assessment of T cell specificity upon HD-Ad delivery
Six days after delivery of HD-Ad vector particles, mice were sacrificed andMLN were isolated. T cells isolated from MLN by nylon-wool enrichmentwere cocultured with saline/HD-Ad-treated irradiated BMDCs for a periodof 72 h. BrdU solution was added during the last 12 h to label the cells,after which proliferation was measured using BrdU cell proliferationELISA according to the manufacturer’s instructions.
In vivo labeling of DCs
Mice were anesthetized using Aerrane inhalation and 50 �l of 1 mg/mlFITC-dextran or 50 �l of 5 mM CFSE was delivered intranasally, 2 hbefore viral delivery. At indicated time points, mice were killed and drain-ing MLN were isolated.
Measurement of absolute DC counts
Lung or draining MLN were isolated and single cell suspension was pre-pared as described above. Cell suspensions were stained for DC-specificmarkers, and absolute counts were assessed in the lung or draining MLNfor each mouse separately.
Ab labeling and flow cytometry
For in vitro BMDC experiments, DCs were labeled at 4°C with CD11c andmaturation markers. To assess pulmonary T cell proliferation, cells fromBALF/lung or MLN single cell suspensions were stained for CD3 andCD4/CD8 with/without BrdU staining as described previously (21). Mi-gration of FITC-dextran-labeled DCs was assessed by labeling MLN singlecell suspensions with CD11c. DCs in the airways were identified by stain-ing single cell lung suspensions with CD11c and CD11b Abs for cDCs andwith mPDCA1 Ab for pDCs. Maturation of DCs in vivo was assessed bystaining with CD86 Ab as marker for DC maturation. Maturation of CD8�DCs was assessed by gating on CD11c�CD8�� cells from the draininglymph nodes and staining with CD86 Ab as a maturation marker. Cellswere gated according to their forward scatter vs side scatter characteristicsto discriminate highly autofluorescent macrophages from DCs as describedpreviously (22). Flow cytometry data was acquired for each of the exper-iments using a BD FACSCalibur (BD Biosciences) at the SickKids-Uni-versity Health Network Flow Cytometry Facility and was analyzed usingFlowJo flow cytometry analysis software (Tree Star).
Statistical analysis
Student’s t test was used to assess statistical significance between means.Significance was set at p � 0.05. All the data is presented as mean � SD.
4099The Journal of Immunology
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
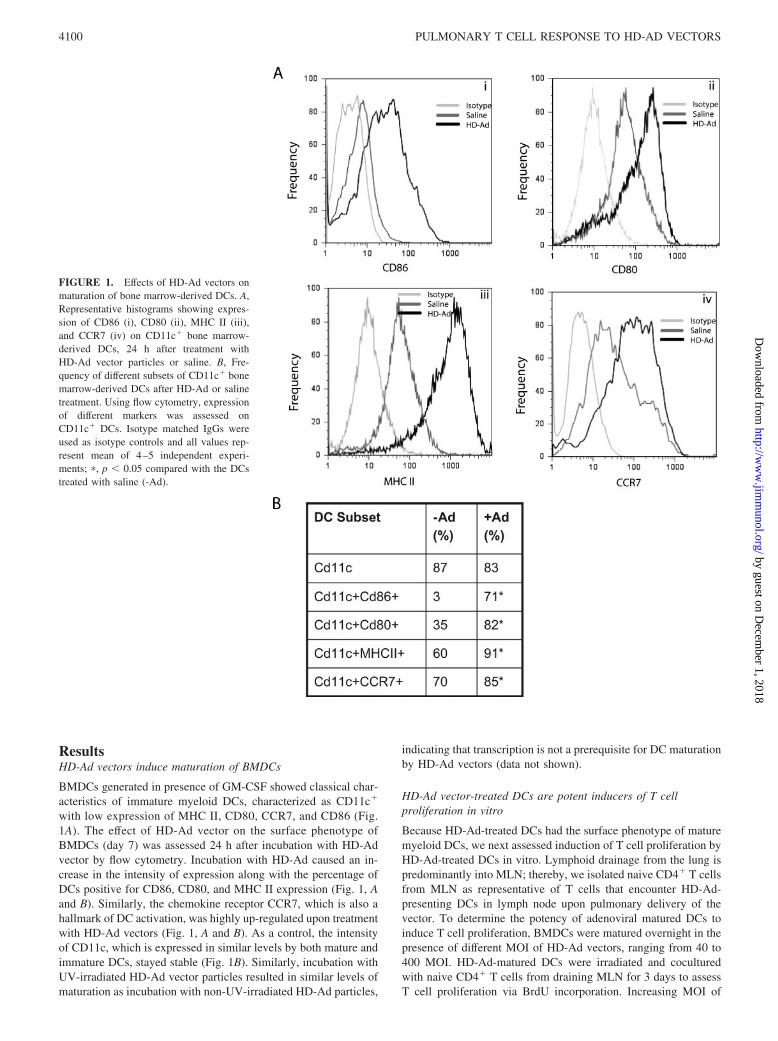
ResultsHD-Ad vectors induce maturation of BMDCs
BMDCs generated in presence of GM-CSF showed classical char-acteristics of immature myeloid DCs, characterized as CD11c�
with low expression of MHC II, CD80, CCR7, and CD86 (Fig.1A). The effect of HD-Ad vector on the surface phenotype ofBMDCs (day 7) was assessed 24 h after incubation with HD-Advector by flow cytometry. Incubation with HD-Ad caused an in-crease in the intensity of expression along with the percentage ofDCs positive for CD86, CD80, and MHC II expression (Fig. 1, Aand B). Similarly, the chemokine receptor CCR7, which is also ahallmark of DC activation, was highly up-regulated upon treatmentwith HD-Ad vectors (Fig. 1, A and B). As a control, the intensityof CD11c, which is expressed in similar levels by both mature andimmature DCs, stayed stable (Fig. 1B). Similarly, incubation withUV-irradiated HD-Ad vector particles resulted in similar levels ofmaturation as incubation with non-UV-irradiated HD-Ad particles,
indicating that transcription is not a prerequisite for DC maturationby HD-Ad vectors (data not shown).
HD-Ad vector-treated DCs are potent inducers of T cellproliferation in vitro
Because HD-Ad-treated DCs had the surface phenotype of maturemyeloid DCs, we next assessed induction of T cell proliferation byHD-Ad-treated DCs in vitro. Lymphoid drainage from the lung ispredominantly into MLN; thereby, we isolated naive CD4� T cellsfrom MLN as representative of T cells that encounter HD-Ad-presenting DCs in lymph node upon pulmonary delivery of thevector. To determine the potency of adenoviral matured DCs toinduce T cell proliferation, BMDCs were matured overnight in thepresence of different MOI of HD-Ad vectors, ranging from 40 to400 MOI. HD-Ad-matured DCs were irradiated and coculturedwith naive CD4� T cells from draining MLN for 3 days to assessT cell proliferation via BrdU incorporation. Increasing MOI of
FIGURE 1. Effects of HD-Ad vectors onmaturation of bone marrow-derived DCs. A,Representative histograms showing expres-sion of CD86 (i), CD80 (ii), MHC II (iii),and CCR7 (iv) on CD11c� bone marrow-derived DCs, 24 h after treatment withHD-Ad vector particles or saline. B, Fre-quency of different subsets of CD11c� bonemarrow-derived DCs after HD-Ad or salinetreatment. Using flow cytometry, expressionof different markers was assessed onCD11c� DCs. Isotype matched IgGs wereused as isotype controls and all values rep-resent mean of 4–5 independent experi-ments; �, p � 0.05 compared with the DCstreated with saline (-Ad).
4100 PULMONARY T CELL RESPONSE TO HD-AD VECTORS
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

HD-Ad used to treat DCs led to a parallel increase in levels of Tcell proliferation, which plateaued around 80–200 MOI, indicatingthat this particular MOI was inducing maximal T cell proliferation(Fig. 2A). Therefore, MOI of 100 was used for all the experiments,and T cell proliferation in response to HD-Ad vectors was alsoconfirmed using CFSE dilution analysis (Fig. 2B). Furthermore,we wanted to estimate the frequency of naive T cells from drainingMLN, responding to adenoviral vectors. To do that, we assessed Tcell proliferation in a coculture of DCs with CFSE-labeled naiveCD4� T cells. As a CFSE-labeled cell undergoes division, theCFSE is halved and every round of division gives a different peakdue to the varying CFSE intensity upon FACS analysis. Eventuallythe frequency of responsive T cells can be calculated. Therefore,naive CD4� T cells from draining MLN labeled with CFSEwere cultured with HD-Ad-matured DCs for a period of 7 days,after which CFSE dye dilution was assessed via flow cytometry(Fig. 2B). The results indicated �9% of naive CD4� T cellsfrom draining MLN to be responding to HD-Ad epitopes pre-sented by DCs (Fig. 2C). As a parallel control, we used cocul-ture of CFSE-labeled naive T cells with DCs without additionof any HD-Ad vectors for maturation, because this would in-dicate the percentage of nonspecific T cell proliferation. Non-specific proliferation was determined to be around 3% by CFSEdilution analysis (Fig. 2C). Therefore, upon accounting for non-specific proliferation, results indicate that �6% of naive T cellsunderwent proliferation in response to HD-Ad-derived epitopespresented by HD-Ad-treated DCs.
HD-Ad vectors induce T cell response upon pulmonary delivery
HD-Ad vectors are regularly used for pulmonary gene therapy un-der the assumption that they do not induce adaptive immune re-sponse in vivo. We next wanted to explore the ability of HD-Advectors to induce T cell responses upon pulmonary delivery. Anal-ysis of immunological responses was performed at two differentdoses. The first dose being 1 � 1011 vector particles/mouse as ahigh dose, which has been shown to result in therapeutic effects in
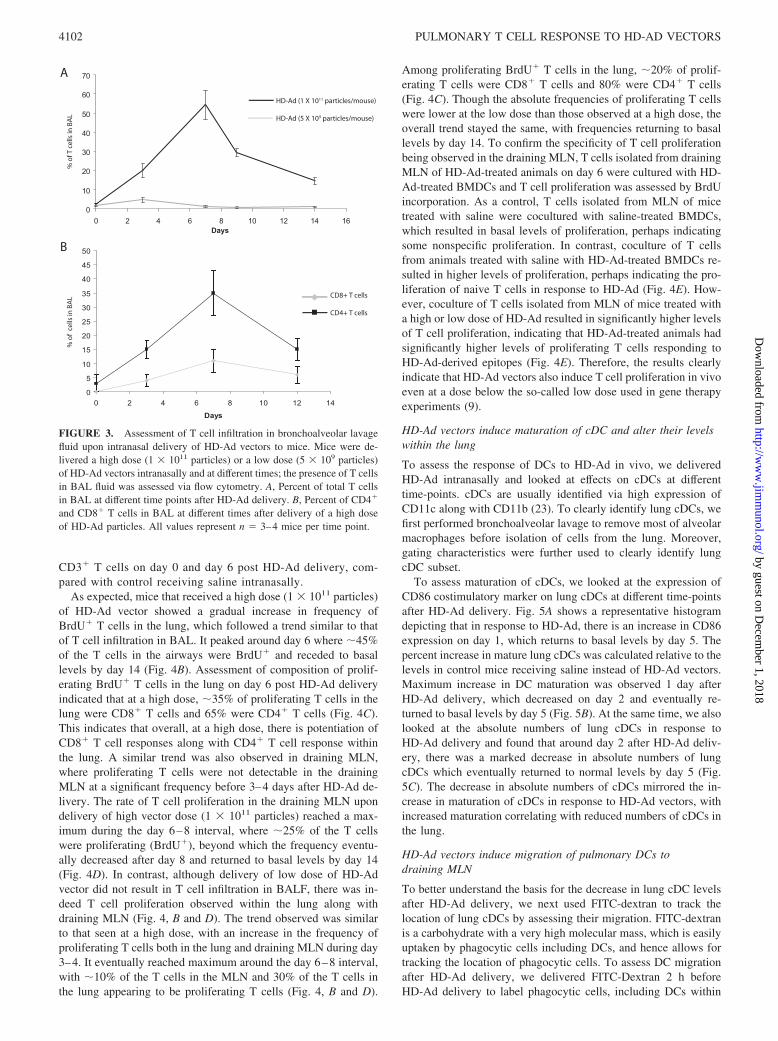
CFTR knockout mice (19). The second dose was a low dose of 5 �109 vector particles/mouse and is 3-fold lower than the low dosenormally used for HD-Ad pulmonary delivery (9, 19). At first, welooked at the infiltration of T cells in the BALF of mice at differenttime points after vector delivery. Under basal conditions in theabsence of any inflammatory response, and before delivery ofHD-Ad vectors, BALF is primarily composed of macrophages,and only 2–3% of the cells can be identified as T lymphocytes (Fig.3A). However, upon delivery of a high dose of HD-Ad vector (1 �1011 vector particles), there was a gradual recruitment of T lym-phocytes in BALF, which peaked around day 7, when approxi-mately half of the cells in BALF were T lymphocytes. The infil-tration gradually receded and was close to basal levels by day 14(Fig. 3A). Further assessment of the phenotype of T cells indicatedthat there was infiltration of both CD4� and CD8� T cells, whichfollowed a similar trend as overall T cell responses, peakingaround day 7 with �10–15% of the cells in BALF being CD8� Tcells and 30–40% being CD4� T cells (Fig. 3B). In contrast tohigh dose, delivery of a low dose of HD-Ad vector (5 � 109
particles), which is even lower than the low dose used in genetherapy experiments, did not result in any significant changes tothe composition of the BALF (Fig. 3A).
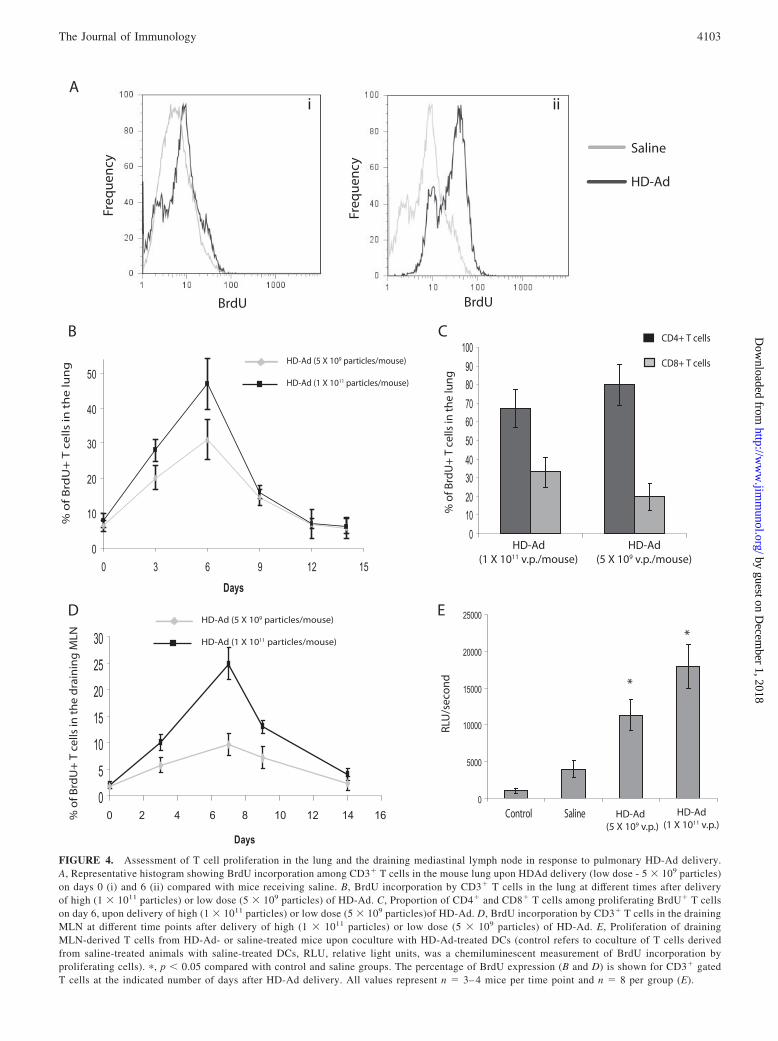
Because the presence of T cells in BALF is indicative ofextensive inflammation, and the absence of T cells in BALFdoes not indicate absence of T cell proliferation in response toHD-Ad, we also assessed T cell proliferation within the airwaysalong with the draining MLN. To directly estimate the fre-quency of proliferating T cells responding to intranasal deliv-ery, we used the strategy of in vivo uptake of the thymidineanalog BrdU with a flow cytometry-based analysis to identifyBrdU� proliferating cells within the airways and draininglymph nodes. BrdU was delivered 24 h before sacrificing mice,and single cell suspensions of lung and lymph node cells wereanalyzed via flow cytometry to look at BrdU� T cells. Fig. 4Ashows representative histograms indicating BrdU staining on
FIGURE 2. Induction of CD4� T cellproliferation in vitro by HD-Ad-treatedBMDCs. A, Assessment of CD4� T cell pro-liferation using BrdU incorporation 3 daysafter a coculture of naive CD4� T cellsisolated from draining MLN with DCstreated with HD-Ad at different MOI (RLU,relative light units, was a chemiluminescentmeasurement of BrdU incorporation by pro-liferating cells). B, Representative histogramdepicting CFSE dilution profile of CFSE�
naive CD4� T cells which were culturedwith HDAd-treated DCs or medium-treatedDCs for a period of 7 days. C, Frequency ofT cells proliferating in coculture of naiveCD4� T cells (CFSE�) with HD-Ad/media-treated DCs, calculated from CFSE dilutionanalysis. �, p � 0.05, compared with precur-sor frequency of T cells cultured in presenceof media-treated DCs. Results for panel Awere normalized to control measuring T cellproliferation in naive CD4� T cell and DCcoculture, where DCs were not treated withHD-Ad vectors results are representative ofthree independent experiments.
4101The Journal of Immunology
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
CD3� T cells on day 0 and day 6 post HD-Ad delivery, com-pared with control receiving saline intranasally.
As expected, mice that received a high dose (1 � 1011 particles)of HD-Ad vector showed a gradual increase in frequency ofBrdU� T cells in the lung, which followed a trend similar to thatof T cell infiltration in BAL. It peaked around day 6 where �45%of the T cells in the airways were BrdU� and receded to basallevels by day 14 (Fig. 4B). Assessment of composition of prolif-erating BrdU� T cells in the lung on day 6 post HD-Ad deliveryindicated that at a high dose, �35% of proliferating T cells in thelung were CD8� T cells and 65% were CD4� T cells (Fig. 4C).This indicates that overall, at a high dose, there is potentiation ofCD8� T cell responses along with CD4� T cell response withinthe lung. A similar trend was also observed in draining MLN,where proliferating T cells were not detectable in the drainingMLN at a significant frequency before 3–4 days after HD-Ad de-livery. The rate of T cell proliferation in the draining MLN upondelivery of high vector dose (1 � 1011 particles) reached a max-imum during the day 6–8 interval, where �25% of the T cellswere proliferating (BrdU�), beyond which the frequency eventu-ally decreased after day 8 and returned to basal levels by day 14(Fig. 4D). In contrast, although delivery of low dose of HD-Advector did not result in T cell infiltration in BALF, there was in-deed T cell proliferation observed within the lung along withdraining MLN (Fig. 4, B and D). The trend observed was similarto that seen at a high dose, with an increase in the frequency ofproliferating T cells both in the lung and draining MLN during day3–4. It eventually reached maximum around the day 6–8 interval,with �10% of the T cells in the MLN and 30% of the T cells inthe lung appearing to be proliferating T cells (Fig. 4, B and D).
Among proliferating BrdU� T cells in the lung, �20% of prolif-erating T cells were CD8� T cells and 80% were CD4� T cells(Fig. 4C). Though the absolute frequencies of proliferating T cellswere lower at the low dose than those observed at a high dose, theoverall trend stayed the same, with frequencies returning to basallevels by day 14. To confirm the specificity of T cell proliferationbeing observed in the draining MLN, T cells isolated from drainingMLN of HD-Ad-treated animals on day 6 were cultured with HD-Ad-treated BMDCs and T cell proliferation was assessed by BrdUincorporation. As a control, T cells isolated from MLN of micetreated with saline were cocultured with saline-treated BMDCs,which resulted in basal levels of proliferation, perhaps indicatingsome nonspecific proliferation. In contrast, coculture of T cellsfrom animals treated with saline with HD-Ad-treated BMDCs re-sulted in higher levels of proliferation, perhaps indicating the pro-liferation of naive T cells in response to HD-Ad (Fig. 4E). How-ever, coculture of T cells isolated from MLN of mice treated witha high or low dose of HD-Ad resulted in significantly higher levelsof T cell proliferation, indicating that HD-Ad-treated animals hadsignificantly higher levels of proliferating T cells responding toHD-Ad-derived epitopes (Fig. 4E). Therefore, the results clearlyindicate that HD-Ad vectors also induce T cell proliferation in vivoeven at a dose below the so-called low dose used in gene therapyexperiments (9).
HD-Ad vectors induce maturation of cDC and alter their levelswithin the lung
To assess the response of DCs to HD-Ad in vivo, we deliveredHD-Ad intranasally and looked at effects on cDCs at differenttime-points. cDCs are usually identified via high expression ofCD11c along with CD11b (23). To clearly identify lung cDCs, wefirst performed bronchoalveolar lavage to remove most of alveolarmacrophages before isolation of cells from the lung. Moreover,gating characteristics were further used to clearly identify lungcDC subset.
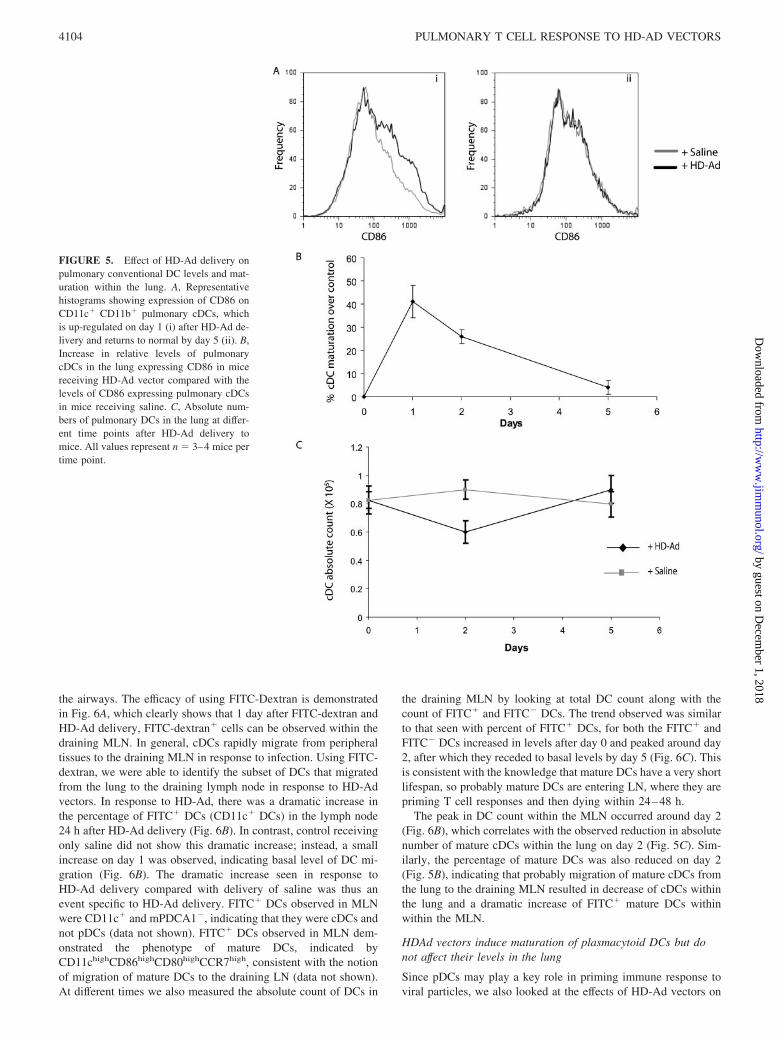
To assess maturation of cDCs, we looked at the expression ofCD86 costimulatory marker on lung cDCs at different time-pointsafter HD-Ad delivery. Fig. 5A shows a representative histogramdepicting that in response to HD-Ad, there is an increase in CD86expression on day 1, which returns to basal levels by day 5. Thepercent increase in mature lung cDCs was calculated relative to thelevels in control mice receiving saline instead of HD-Ad vectors.Maximum increase in DC maturation was observed 1 day afterHD-Ad delivery, which decreased on day 2 and eventually re-turned to basal levels by day 5 (Fig. 5B). At the same time, we alsolooked at the absolute numbers of lung cDCs in response toHD-Ad delivery and found that around day 2 after HD-Ad deliv-ery, there was a marked decrease in absolute numbers of lungcDCs which eventually returned to normal levels by day 5 (Fig.5C). The decrease in absolute numbers of cDCs mirrored the in-crease in maturation of cDCs in response to HD-Ad vectors, withincreased maturation correlating with reduced numbers of cDCs inthe lung.
HD-Ad vectors induce migration of pulmonary DCs todraining MLN
To better understand the basis for the decrease in lung cDC levelsafter HD-Ad delivery, we next used FITC-dextran to track thelocation of lung cDCs by assessing their migration. FITC-dextranis a carbohydrate with a very high molecular mass, which is easilyuptaken by phagocytic cells including DCs, and hence allows fortracking the location of phagocytic cells. To assess DC migrationafter HD-Ad delivery, we delivered FITC-Dextran 2 h beforeHD-Ad delivery to label phagocytic cells, including DCs within
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16
% o
f T c
ells
in B
AL
HD-Ad (1 X 1011 particles/mouse)
HD-Ad (5 X 109 particles/mouse)
0
5
10
15
20
25
30
35
40
45
50
0 2 4 6 8 10 12 14
Days
% o
f ce
lls in
BA
L CD8+ T cells
CD4+ T cells
Days
A
B
FIGURE 3. Assessment of T cell infiltration in bronchoalveolar lavagefluid upon intranasal delivery of HD-Ad vectors to mice. Mice were de-livered a high dose (1 � 1011 particles) or a low dose (5 � 109 particles)of HD-Ad vectors intranasally and at different times; the presence of T cellsin BAL fluid was assessed via flow cytometry. A, Percent of total T cellsin BAL at different time points after HD-Ad delivery. B, Percent of CD4�
and CD8� T cells in BAL at different times after delivery of a high doseof HD-Ad particles. All values represent n � 3–4 mice per time point.
4102 PULMONARY T CELL RESPONSE TO HD-AD VECTORS
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
BrdU BrdU
Freq
uen
cy
Freq
uen
cy
Saline
HD-Ad
0
10
20
30
40
50
0 3 6 9 12 15
Days
% o
f B
rdU
+ T
ce
lls
in t
he
lun
g HD-Ad (5 X 109 particles/mouse)
HD-Ad (1 X 1011 particles/mouse)
0
5
10
15
20
25
30
% o
f Brd
U+
T c
ells
in t
he
dra
inin
g M
LN
0 2 4 6 8 10 12 14 16
Days
HD-Ad (5 X 109 particles/mouse)
HD-Ad (1 X 1011 particles/mouse)
iiiA
B
D
0102030405060708090
100
21
% o
f Brd
U+
T c
ells
in t
he
lun
g
HD-Ad(1 X 1011 v.p./mouse)
HD-Ad(5 X 109 v.p./mouse)
CD4+ T cells
CD8+ T cells
C
0
5000
10000
15000
20000
25000
Control Saline HD-Ad(5 X 109 v.p.)
E
*
*
RLU
/sec
on
d
HD-Ad(1 X 1011 v.p.)
FIGURE 4. Assessment of T cell proliferation in the lung and the draining mediastinal lymph node in response to pulmonary HD-Ad delivery.A, Representative histogram showing BrdU incorporation among CD3� T cells in the mouse lung upon HDAd delivery (low dose - 5 � 109 particles)on days 0 (i) and 6 (ii) compared with mice receiving saline. B, BrdU incorporation by CD3� T cells in the lung at different times after deliveryof high (1 � 1011 particles) or low dose (5 � 109 particles) of HD-Ad. C, Proportion of CD4� and CD8� T cells among proliferating BrdU� T cellson day 6, upon delivery of high (1 � 1011 particles) or low dose (5 � 109 particles)of HD-Ad. D, BrdU incorporation by CD3� T cells in the drainingMLN at different time points after delivery of high (1 � 1011 particles) or low dose (5 � 109 particles) of HD-Ad. E, Proliferation of drainingMLN-derived T cells from HD-Ad- or saline-treated mice upon coculture with HD-Ad-treated DCs (control refers to coculture of T cells derivedfrom saline-treated animals with saline-treated DCs, RLU, relative light units, was a chemiluminescent measurement of BrdU incorporation byproliferating cells). �, p � 0.05 compared with control and saline groups. The percentage of BrdU expression (B and D) is shown for CD3� gatedT cells at the indicated number of days after HD-Ad delivery. All values represent n � 3– 4 mice per time point and n � 8 per group (E).
4103The Journal of Immunology
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
the airways. The efficacy of using FITC-Dextran is demonstratedin Fig. 6A, which clearly shows that 1 day after FITC-dextran andHD-Ad delivery, FITC-dextran� cells can be observed within thedraining MLN. In general, cDCs rapidly migrate from peripheraltissues to the draining MLN in response to infection. Using FITC-dextran, we were able to identify the subset of DCs that migratedfrom the lung to the draining lymph node in response to HD-Advectors. In response to HD-Ad, there was a dramatic increase inthe percentage of FITC� DCs (CD11c� DCs) in the lymph node24 h after HD-Ad delivery (Fig. 6B). In contrast, control receivingonly saline did not show this dramatic increase; instead, a smallincrease on day 1 was observed, indicating basal level of DC mi-gration (Fig. 6B). The dramatic increase seen in response toHD-Ad delivery compared with delivery of saline was thus anevent specific to HD-Ad delivery. FITC� DCs observed in MLNwere CD11c� and mPDCA1�, indicating that they were cDCs andnot pDCs (data not shown). FITC� DCs observed in MLN dem-onstrated the phenotype of mature DCs, indicated byCD11chighCD86highCD80highCCR7high, consistent with the notionof migration of mature DCs to the draining LN (data not shown).At different times we also measured the absolute count of DCs in
the draining MLN by looking at total DC count along with thecount of FITC� and FITC� DCs. The trend observed was similarto that seen with percent of FITC� DCs, for both the FITC� andFITC� DCs increased in levels after day 0 and peaked around day2, after which they receded to basal levels by day 5 (Fig. 6C). Thisis consistent with the knowledge that mature DCs have a very shortlifespan, so probably mature DCs are entering LN, where they arepriming T cell responses and then dying within 24–48 h.
The peak in DC count within the MLN occurred around day 2(Fig. 6B), which correlates with the observed reduction in absolutenumber of mature cDCs within the lung on day 2 (Fig. 5C). Sim-ilarly, the percentage of mature DCs was also reduced on day 2(Fig. 5B), indicating that probably migration of mature cDCs fromthe lung to the draining MLN resulted in decrease of cDCs withinthe lung and a dramatic increase of FITC� mature DCs withinwithin the MLN.
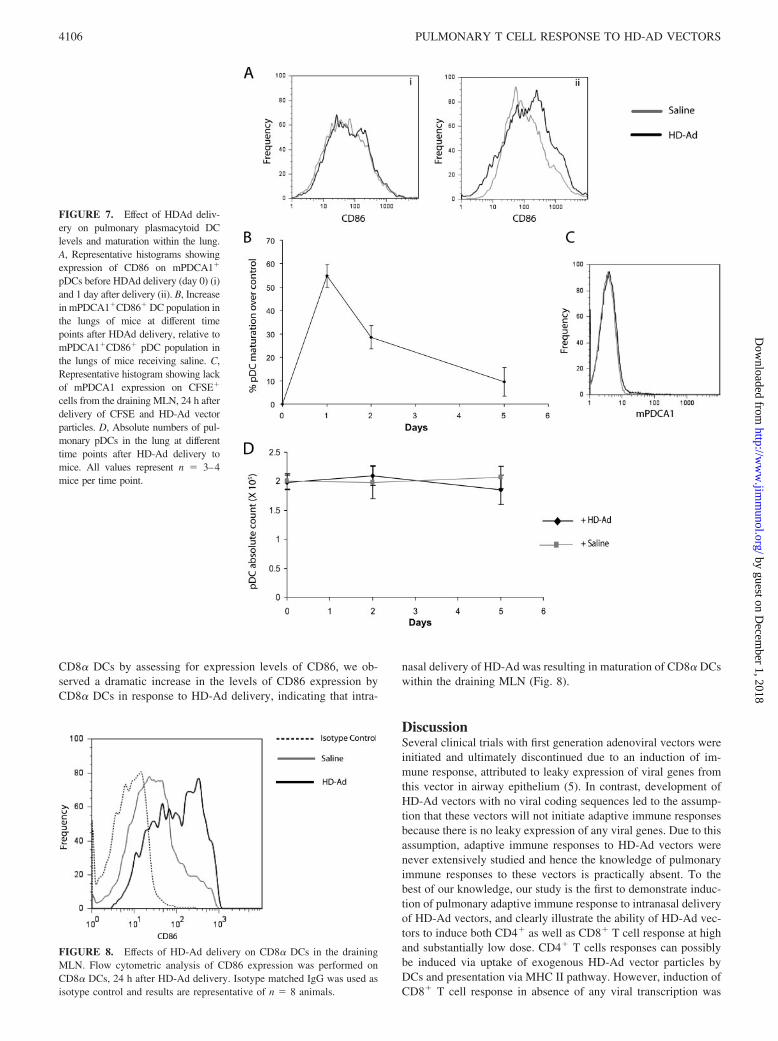
HDAd vectors induce maturation of plasmacytoid DCs but donot affect their levels in the lung
Since pDCs may play a key role in priming immune response toviral particles, we also looked at the effects of HD-Ad vectors on
FIGURE 5. Effect of HD-Ad delivery onpulmonary conventional DC levels and mat-uration within the lung. A, Representativehistograms showing expression of CD86 onCD11c� CD11b� pulmonary cDCs, whichis up-regulated on day 1 (i) after HD-Ad de-livery and returns to normal by day 5 (ii). B,Increase in relative levels of pulmonarycDCs in the lung expressing CD86 in micereceiving HD-Ad vector compared with thelevels of CD86 expressing pulmonary cDCsin mice receiving saline. C, Absolute num-bers of pulmonary DCs in the lung at differ-ent time points after HD-Ad delivery tomice. All values represent n � 3–4 mice pertime point.
4104 PULMONARY T CELL RESPONSE TO HD-AD VECTORS
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

pDCs within the airways (24). First, we assessed the effects ofHD-Ad on maturation of pDC. Fig. 7A shows representative his-tograms showing CD86 expression on lung pDCs at day 0 and day1 after HD-Ad vector delivery. We observed that upon HD-Addelivery pDCs matured rapidly, with maximum maturation beingobserved around day 1 and eventually receding by day 2 and re-turning to basal levels by day 5 (Fig. 7, A and B). Thus, the trendwas similar to that observed with cDCs. Since the trend observedwith cDCs was associated with migration to the draining LN, wenext assessed the migration of pDCs from the lung to draining LNin response to HD-Ad. It has been reported that FITC-Dextrancannot be used for identifying pDCs, because pDCs do not readilyuptake FITC-Dextran. CFSE has been used before to track pDCs,and therefore we used intranasal delivery of CFSE to track move-ment of pDCs from the lung to the draining LN (25). CFSE wasdelivered 2 h before viral delivery to label all the cells in theairways. At different times after HD-Ad delivery, we assessed thepresence of CFSE� DCs within the lymph nodes and found resultssimilar to that observed with FITC-Dextran, with percentage ofCFSE� DCs peaking around day 2 and then receding to basal
levels by day 5 (data not shown). However, we did not detectpresence of mPDCA1� cells among CFSE� population within thelymph nodes, indicating that there was no migration of pDCs in-duced in response to HD-Ad vectors (Fig. 7C). At the same time,we also measured the absolute numbers of pDC within the lung atdifferent times and observed that upon HD-Ad vector delivery, thenumber of pDC in the lung stayed very consistent, further con-firming the lack of pDC migration from the lung (Fig. 7D).
HD-Ad vectors induce maturation of CD8� DCs in the drainingMLN
CD8� DCs are thought to play a key role in cross-presentation ofAgs, and because we observed induction of a CD8� T cell prolif-eration even without viral transcription upon HD-Ad delivery, weassessed the effects of HD-Ad delivery on CD8� DCs to identifywhether CD8� DCs can play a potential role in induction of CD8�
T cell proliferation in response to intranasal delivery of HD-Advectors. Because CD8� DCs are known to be lymphoid resident,we did not observe recruitment of CD8� DCs in the lungs. How-ever, upon assessing for maturation of draining MLN resident
0
1
2
0 1 2 3 4 5 6
Days
Total DC count
FITC - DC count
FITC+ DC count
DC
ab
solu
te n
um
ber
(X 1
03 ) in
dra
inin
g M
LN
0
3
6
9
12
15
18
21
24
0 1 2 3 4 5 6Days
% o
f FIT
C+
DC
s in
th
e d
rain
ing
MLN
Saline
HD-Ad
A
B
C
FITC-Dextran
Saline
HD-Ad
Freq
uen
cy
FITC-Dextran
FIGURE 6. Effect of HD-Ad delivery onDC migration from the lung to the drainingMLN. A, Representative histogram, 1 day af-ter HD-Ad delivery showing that FITC�
DCs can be identified in draining MLN(FITC expression by CD11c� DCs in MLN).B, Relative levels of FITC� CD11c� DCs inMLN at different days after intranasal deliv-ery of HDAd or saline, 2 h after FITC-Dex-tran delivery. C, Absolute number of DCs,both FITC� and FITC� along with total DCsin MLN were quantified at different timepoints after HD-Ad delivery via expressionof CD11c. All values represent n � 3–4mice per condition.
4105The Journal of Immunology
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
CD8� DCs by assessing for expression levels of CD86, we ob-served a dramatic increase in the levels of CD86 expression byCD8� DCs in response to HD-Ad delivery, indicating that intra-
nasal delivery of HD-Ad was resulting in maturation of CD8� DCswithin the draining MLN (Fig. 8).
DiscussionSeveral clinical trials with first generation adenoviral vectors wereinitiated and ultimately discontinued due to an induction of im-mune response, attributed to leaky expression of viral genes fromthis vector in airway epithelium (5). In contrast, development ofHD-Ad vectors with no viral coding sequences led to the assump-tion that these vectors will not initiate adaptive immune responsesbecause there is no leaky expression of any viral genes. Due to thisassumption, adaptive immune responses to HD-Ad vectors werenever extensively studied and hence the knowledge of pulmonaryimmune responses to these vectors is practically absent. To thebest of our knowledge, our study is the first to demonstrate induc-tion of pulmonary adaptive immune response to intranasal deliveryof HD-Ad vectors, and clearly illustrate the ability of HD-Ad vec-tors to induce both CD4� as well as CD8� T cell response at highand substantially low dose. CD4� T cells responses can possiblybe induced via uptake of exogenous HD-Ad vector particles byDCs and presentation via MHC II pathway. However, induction ofCD8� T cell response in absence of any viral transcription was
FIGURE 7. Effect of HDAd deliv-ery on pulmonary plasmacytoid DClevels and maturation within the lung.A, Representative histograms showingexpression of CD86 on mPDCA1�
pDCs before HDAd delivery (day 0) (i)and 1 day after delivery (ii). B, Increasein mPDCA1�CD86� DC population inthe lungs of mice at different timepoints after HDAd delivery, relative tomPDCA1�CD86� pDC population inthe lungs of mice receiving saline. C,Representative histogram showing lackof mPDCA1 expression on CFSE�
cells from the draining MLN, 24 h afterdelivery of CFSE and HD-Ad vectorparticles. D, Absolute numbers of pul-monary pDCs in the lung at differenttime points after HD-Ad delivery tomice. All values represent n � 3–4mice per time point.
FIGURE 8. Effects of HD-Ad delivery on CD8� DCs in the drainingMLN. Flow cytometric analysis of CD86 expression was performed onCD8� DCs, 24 h after HD-Ad delivery. Isotype matched IgG was used asisotype control and results are representative of n � 8 animals.
4106 PULMONARY T CELL RESPONSE TO HD-AD VECTORS
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

unexpected, because viral gene expression is usually needed forAgs to be shuttled through the MHC I pathway.
In two recent gene therapy clinical trials using AAV serotype 2vectors, which are thought to be the least immunogenic, there werereports of apparent immune responses with appearance of CD8�
effector T cells (26). This gave rise to the so-called capsid T cellhypothesis, which postulates that exogenous vector proteins maybe shuttled into the MHC class I pathway for T cell priming (12).Our results demonstrate similar phenomena occurring upon pul-monary delivery of HD-Ad vectors, whereby we are seeing pro-liferation of CD8� T cells along with CD4� T cells. This obser-vation indicates that HD-Ad-derived peptides are being shuttled toMHC I pathway and cross-presented in context with MHC class Ito induce CD8� T cell response along with presentation with MHCclass II to induce CD4� T cell response. Five dominant CD8� Tcell epitopes on the capsid hexon protein of adenovirus have beenidentified and because HD-Ad vector does indeed has the capsidproteins, it is possible that perhaps cross-presentation of theseepitopes may play a role in inducing CD8� T cell responses (27).Our results indicate that perhaps CD8� DCs may play a role incross-presenting HD-Ad-derived epitopes, because this subset ofDC undergoes maturation within the draining MLN upon HD-Addelivery. We did not observe recruitment of CD8� DCs in thelungs as expected, because these DCs are thought to be nonmigra-tory. However, maturation of CD8� DCs indicates that perhapscDCs are migrating to the draining MLN and transferring the Agsto CD8� DCs, which results in maturation of CD8� DCs. MatureCD8� DCs may then go on to induce CD8� T cell response, as hasbeen observed with lung and s.c. infection using HSV (28, 29).Moreover, CD103� DCs with the ability to cross-present inhaledAgs and migrate to draining MLN have also been identified in themouse lung (30). Therefore, there may be a specialized populationof DCs that may be initiating CD8� T cell responses upon HD-Addelivery via cross-presentation, which may include CD103� DCsand CD8� DCs, which we have shown to mature in draining MLNin response to HD-Ad. MHC:peptide complexes of immunodom-inant epitopes can have a half-life of �7 days (31, 32). Therefore,HD-Ad transduced epithelial cells may present vector-derivedepitopes for substantial amounts of time to allow for responding Tcells to migrate to the airways and gradually target/eliminate sometransduced cells. Furthermore, upon readministration, memory Tcells are probably activated which are recruited to the site withinthe first 3 days and mediate gradual clearing of transduced cells,and hence may account for observed loss in transgene expressionupon vector readministration (9, 33). Moreover, the extent of bothCD4� and CD8� proliferation in the lung and draining MLNpeaked around day 6–7, which is similar to the timeline of pul-monary T cell responses associated with other pulmonary infec-tions such as influenza (34, 35).
We assessed the response of pulmonary DCs to HD-Ad to gainan insight behind priming of T cell response to HD-Ad vectorsupon pulmonary delivery. Our results demonstrated that early onafter HD-Ad delivery, there is rapid maturation of both cDC alongwith pDC, which peaks around day 1. As DCs mature, there ismigration of only cDCs to the lymph nodes, resulting in reductionof percent of mature cells, as well as reduction in absolute count ofcDC in the lung by day 2 after HD-Ad vector delivery. During thefirst 48 h after delivery, there is massive increase in the numbersof lung-derived DCs in the draining MLN, which eventually re-cedes to basal levels by day 5. In contrast to cDCs, pDCs do notmigrate and probably act as local sites of IFN-� secretion to primeinnate and adaptive immune responses. Ex vivo studies have fur-ther demonstrated that adenovirus can induce IFN-� secretion bypDCs via TLR-MyD88 pathway (36). The observation of priming
of DC maturation and migration within the first 48 h is consistentwith the previous findings involving other respiratory viruses suchas paramoxyviruses (22). This may in part explain the role of clin-ically used mucolytic agent Nacystelyn, which can inhibit DC mat-uration, in enhancing HD-Ad mediated gene transfer to mouseairways (4, 37). Thus, cDCs seem to play the central role in prim-ing pulmonary immune responses against HD-Ad vectors, andhence intervention within the first 24 h of vector delivery to pre-vent DC maturation/migration may help in mitigating the adaptiveimmune responses induced by HD-Ad vectors. In contrast to ourfindings that HD-Ad vectors act as potent stimulators of pulmo-nary DC maturation and pulmonary T cell response, it was recentlyreported that infection with adenovirus can suppress lung DC func-tion, and such suppression was related to adenovirus transcription(38). We reason that the report’s conclusion may be instead attrib-uted to gene products encoded by E3 region, which function tomodulate host-immune response (39). In particular, E3gp19K pro-tein encoded by E3 region of the adenoviruses genome has beenshown to sequester MHC I complexes in endoplasmic reticulum,thereby suppressing CD8� T cell responses (40). Since HD-Adparticles do not encode any viral genes including E3 region, thesevectors cannot suppress pulmonary DC responses and are ratherinitiators of adaptive immune response.
In conclusion, our data demonstrate that pulmonary delivery ofHD-Ad results in induction of CD4� as well as CD8� T cell re-sponses that peak around day 5–7. These responses are primarilyorchestrated by cDCs that mature and migrate to draining MLNwithin the first 48 h after HD-Ad delivery. At the same time, pDCsalso mature but do not migrate to MLN, and hence may act as localsource of IFN-� to prime innate and adaptive immune response.Therefore, pulmonary delivery of known DC maturation inhibitorssuch as prostaglandin E2, Bortezomib, FK778, a derivative of theactive leflunomide-metabolite, within the first 24 h after HD-Advector delivery, may inhibit priming of pulmonary adaptive im-mune responses by preventing maturation and thereby migration ofcDCs (41–43). Moreover, because T cell proliferation peakedaround a week after HD-Ad delivery, transient immunosuppres-sion within a week after HD-Ad vector delivery using cyclosporineA or CTLA4Ig to inhibit CD28/B7 pathway or inhibition of CD40/CD40L pathway via CD40LIg administration will prevent T cellproliferation, and hence suppress adaptive immune response toHD-Ad vectors (44–46). Strategies to inhibit adaptive immuneresponses will inhibit induction of memory T cells. This may go onto prevent the observed loss of transgene expression and anti-HD-Ad Ab titer, observed in mice upon subsequent re-administra-tions. Identification of adaptive immune responses initiated uponpulmonary delivery of HD-Ad vectors indicates that strategies tar-geted to improve vector delivery and persistence need to targetboth adaptive and innate immune responses. Moreover, identifica-tion of the time-line of these responses may help in identifying thetime-point for intervention to prevent subsequent unwanted immu-nological responses.
DisclosuresThe authors have no financial conflict of interest.
References1. Campos, S. K., and M. A. Barry. 2007. Current advances and future challenges
in Adenoviral vector biology and targeting. Curr. Gene Ther. 7: 189–204.2. Flotte, T. R., P. Ng, D. E. Dylla, P. B. McCray, Jr., G. Wang, J. K. Kolls, and
J. Hu. 2007. Viral vector-mediated and cell-based therapies for treatment of cys-tic fibrosis. Mol. Ther. 15: 229–241.
3. Brunetti-Pierri, N., and P. Ng. 2006. Progress towards the clinical application ofhelper-dependent adenoviral vectors for liver and lung gene therapy. Curr. Opin.Mol. Ther. 8: 446–454.
4107The Journal of Immunology
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

4. Kushwah, R., J. R. Oliver, H. Cao, and J. Hu. 2007. Nacystelyn enhances ad-enoviral vector-mediated gene delivery to mouse airways. Gene Ther. 14:1243–1248.
5. Kushwah, R., H. Cao, and J. Hu. 2007. Potential of helper-dependent adenoviralvectors in modulating airway innate immunity. Cell. Mol. Immunol. 4: 81–89.
6. Alba, R., A. Bosch, and M. Chillon. 2005. Gutless adenovirus: last-generationadenovirus for gene therapy. Gene Ther. 12 (Suppl. 1): S18–S27.
7. Muruve, D. A. 2004. The innate immune response to adenovirus vectors. Hum.Gene Ther. 15: 1157–1166.
8. Liu, Q., and D. A. Muruve. 2003. Molecular basis of the inflammatory responseto adenovirus vectors. Gene Ther. 10: 935–940.
9. Koehler, D. R., B. Martin, M. Corey, D. Palmer, P. Ng, A. K. Tanswell, andJ. Hu. 2006. Readministration of helper-dependent adenovirus to mouse lung.Gene Ther. 13: 773–780.
10. Morral, N., W. O’Neal, K. Rice, M. Leland, J. Kaplan, P. A. Piedra, H. Zhou,R. J. Parks, R. Velji, E. Aguilar-Cordova, et al. 1999. Administration of helper-dependent adenoviral vectors and sequential delivery of different vector serotypefor long-term liver-directed gene transfer in baboons. Proc. Natl. Acad. Sci. USA96: 12816–12821.
11. Zaiss, A. K., and D. A. Muruve. 2005. Immune responses to adeno-associatedvirus vectors. Curr. Gene Ther. 5: 323–331.
12. Wilson, J. 2007. Humility and clinical trials. Mol. Ther. 15: 1571–1573.13. Banchereau, J., and R. M. Steinman. 1998. Dendritic cells and the control of
immunity. Nature 392: 245–252.14. Lambrecht, B. N., B. Salomon, D. Klatzmann, and R. A. Pauwels. 1998. Den-
dritic cells are required for the development of chronic eosinophilic airway in-flammation in response to inhaled antigen in sensitized mice. J. Immunol. 160:4090–4097.
15. Upham, J. W. 2003. The role of dendritic cells in immune regulation and allergicairway inflammation. Respirology 8: 140–148.
16. Lambrecht, B. N., R. A. Pauwels, and B. Fazekas De St. Groth. 2000. Inductionof rapid T cell activation, division, and recirculation by intratracheal injection ofdendritic cells in a TCR transgenic model. J. Immunol. 164: 2937–2946.
17. Lambrecht, B. N., M. De Veerman, A. J. Coyle, J. C. Gutierrez-Ramos,K. Thielemans, and R. A. Pauwels. 2000. Myeloid dendritic cells induce Th2responses to inhaled antigen, leading to eosinophilic airway inflammation.J. Clin. Invest. 106: 551–559.
18. Lutz, M. B., N. Kukutsch, A. L. Ogilvie, S. Rossner, F. Koch, N. Romani, andG. Schuler. 1999. An advanced culture method for generating large quantities ofhighly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 223:77–92.
19. Koehler, D. R., U. Sajjan, Y. H. Chow, B. Martin, G. Kent, A. K. Tanswell,C. McKerlie, J. F. Forstner, and J. Hu. 2003. Protection of Cftr knockout micefrom acute lung infection by a helper-dependent adenoviral vector expressingCftr in airway epithelia. Proc. Natl. Acad. Sci. USA 100: 15364–15369.
20. Chen, J. C., M. L. Chang, and M. O. Muench. 2003. A kinetic study of the murinemixed lymphocyte reaction by 5,6-carboxyfluorescein diacetate succinimidyl es-ter labeling. J. Immunol. Methods 279: 123–133.
21. Carayon, P., and A. Bord. 1992. Identification of DNA-replicating lymphocytesubsets using a new method to label the bromo-deoxyuridine incorporated intothe DNA. J. Immunol. Methods 147: 225–230.
22. Grayson, M. H., M. S. Ramos, M. M. Rohlfing, R. Kitchens, H. D. Wang,A. Gould, E. Agapov, and M. J. Holtzman. 2007. Controls for lung dendritic cellmaturation and migration during respiratory viral infection. J. Immunol. 179:1438–1448.
23. Smit, J. J., B. D. Rudd, and N. W. Lukacs. 2006. Plasmacytoid dendritic cellsinhibit pulmonary immunopathology and promote clearance of respiratory syn-cytial virus. J. Exp. Med. 203: 1153–1159.
24. Santini, S. M., T. Di Pucchio, C. Lapenta, S. Parlato, M. Logozzi, andF. Belardelli. 2002. The natural alliance between type I interferon and dendriticcells and its role in linking innate and adaptive immunity. J. Interferon CytokineRes. 22: 1071–1080.
25. Legge, K. L., and T. J. Braciale. 2003. Accelerated migration of respiratorydendritic cells to the regional lymph nodes is limited to the early phase of pul-monary infection. Immunity 18: 265–277.
26. Manno, C. S., G. F. Pierce, V. R. Arruda, B. Glader, M. Ragni, J. J. Rasko,M. C. Ozelo, K. Hoots, P. Blatt, B. Konkle, et al. 2006. Successful transductionof liver in hemophilia by AAV-Factor IX and limitations imposed by the hostimmune response. Nat. Med. 12: 342–347.
27. Leen, A. M., U. Sili, E. F. Vanin, A. M. Jewell, W. Xie, D. Vignali, P. A. Piedra,M. K. Brenner, and C. M. Rooney. 2004. Conserved CTL epitopes on the ade-
novirus hexon protein expand subgroup cross-reactive and subgroup-specificCD8� T cells. Blood 104: 2432–2440.
28. Belz, G. T., C. M. Smith, L. Kleinert, P. Reading, A. Brooks, K. Shortman,F. R. Carbone, and W. R. Heath. 2004. Distinct migrating and nonmigratingdendritic cell populations are involved in MHC class I-restricted antigen presen-tation after lung infection with virus. Proc. Natl. Acad. Sci. USA 101:8670–8675.
29. Allan, R. S., J. Waithman, S. Bedoui, C. M. Jones, J. A. Villadangos, Y. Zhan,A. M. Lew, K. Shortman, W. R. Heath, and F. R. Carbone. 2006. Migratorydendritic cells transfer antigen to a lymph node-resident dendritic cell populationfor efficient CTL priming. Immunity 25: 153–162.
30. del Rio, M. L., J. I. Rodriguez-Barbosa, E. Kremmer, and R. Forster. 2007.CD103� and CD103� bronchial lymph node dendritic cells are specialized inpresenting and cross-presenting innocuous antigen to CD4� and CD8� T cells.J. Immunol. 178: 6861–6866.
31. Sant, A. J., F. A. Chaves, S. A. Jenks, K. A. Richards, P. Menges, J. M. Weaver,and C. A. Lazarski. 2005. The relationship between immunodominance, DMediting, and the kinetic stability of MHC class II:peptide complexes. Immunol.Rev. 207: 261–278.
32. Lazarski, C. A., F. A. Chaves, S. A. Jenks, S. Wu, K. A. Richards, J. M. Weaver,and A. J. Sant. 2005. The kinetic stability of MHC class II:peptide complexes isa key parameter that dictates immunodominance. Immunity 23: 29–40.
33. Hikono, H., J. E. Kohlmeier, K. H. Ely, I. Scott, A. D. Roberts, M. A. Blackman,and D. L. Woodland. 2006. T-cell memory and recall responses to respiratoryvirus infections. Immunol. Rev. 211: 119–132.
34. Lawrence, C. W., R. M. Ream, and T. J. Braciale. 2005. Frequency, specificity,and sites of expansion of CD8� T cells during primary pulmonary influenza virusinfection. J. Immunol. 174: 5332–5340.
35. Lawrence, C. W., and T. J. Braciale. 2004. Activation, differentiation, and mi-gration of naive virus-specific CD8� T cells during pulmonary influenza virusinfection. J. Immunol. 173: 1209–1218.
36. Zhu, J., X. Huang, and Y. Yang. 2007. Innate immune response to adenoviralvectors is mediated by both Toll-like receptor-dependent and -independent path-ways. J. Virol. 81: 3170–3180.
37. Vosters, O., J. Neve, D. De Wit, F. Willems, M. Goldman, and V. Verhasselt.2003. Dendritic cells exposed to nacystelyn are refractory to maturation andpromote the emergence of alloreactive regulatory t cells. Transplantation 75:383–389.
38. Thiele, A. T., T. L. Sumpter, J. A. Walker, Q. Xu, C. H. Chang, R. L. Bacallao,R. Kher, and D. S. Wilkes. 2006. Pulmonary immunity to viral infection: ade-novirus infection of lung dendritic cells renders T cells nonresponsive to inter-leukin-2. J. Virol. 80: 1826–1836.
39. Rawle, F. C., A. E. Tollefson, W. S. Wold, and L. R. Gooding. 1989. Mouseanti-adenovirus cytotoxic T lymphocytes: inhibition of lysis by E3 gp19K but notE3 14.7K. J. Immunol. 143: 2031–2037.
40. Sparer, T. E., R. A. Tripp, D. L. Dillehay, T. W. Hermiston, W. S. Wold, andL. R. Gooding. 1996. The role of human adenovirus early region 3 proteins(gp19K, 10.4K, 14.5K, and 14.7K) in a murine pneumonia model. J. Virol. 70:2431–2439.
41. Sa-Nunes, A., A. Bafica, D. A. Lucas, T. P. Conrads, T. D. Veenstra,J. F. Andersen, T. N. Mather, J. M. Ribeiro, and I. M. Francischetti. 2007. Pros-taglandin E2 is a major inhibitor of dendritic cell maturation and function inIxodes scapularis saliva. J. Immunol. 179: 1497–1505.
42. Straube, C., R. Wehner, M. Wendisch, M. Bornhauser, M. Bachmann,E. P. Rieber, and M. Schmitz. 2007. Bortezomib significantly impairs the immu-nostimulatory capacity of human myeloid blood dendritic cells. Leukemia 21:1464–1471.
43. Zeyda, M., B. M. Kirsch, R. Geyeregger, K. M. Stuhlmeier, G. J. Zlabinger,W. H. Horl, M. D. Saemann, and T. M. Stulnig. 2005. Inhibition of humandendritic cell maturation and function by the novel immunosuppressant FK778.Transplantation 80: 1105–1111.
44. Nishiyama, S., N. Manabe, Y. Kubota, H. Ohnishi, A. Kitanaka, M. Tokuda,T. Taminato, T. Ishida, J. Takahara, and T. Tanaka. 2005. Cyclosporin A inhibitsthe early phase of NF-�B/RelA activation induced by CD28 costimulatory sig-naling to reduce the IL-2 expression in human peripheral T cells. Int. Immuno-pharmacol. 5: 699–710.
45. Jooss, K., L. A. Turka, and J. M. Wilson. 1998. Blunting of immune responsesto adenoviral vectors in mouse liver and lung with CTLA4Ig. Gene Ther. 5:309–319.
46. Yang, Y., Q. Su, I. S. Grewal, R. Schilz, R. A. Flavell, and J. M. Wilson. 1996.Transient subversion of CD40 ligand function diminishes immune responses toadenovirus vectors in mouse liver and lung tissues. J. Virol. 70: 6370–6377.
4108 PULMONARY T CELL RESPONSE TO HD-AD VECTORS
by guest on Decem
ber 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from


















