Characterization of Foam Cells in Experimental Atherosclerosis
8
Acta Med Scand (Suppl) 642: 151-158, 1980 Characterization of Foam Cells in Experimental Atherosclerosis Stanley Fowler From the Rockefeller University, New York, New York, USA ABSTRACT. A series of biochemical and morphological studies has focused on the properties and origins of lipid laden foam cells in experimentally induced atherosclero- sis in rabbits. Lipid inclusions present in these cells occupy half or more of the cyto- plasmic volume and are of two kinds: cyto- plasmic lipid droplets composed predomi- nantly of cholesteryl esters and lysosomes in which substantial quantities of free choles- terol have accumulated. The foam cells ex- hibit some properties of macrophages but not others. They possess high levels of acid hy- drolases and catalase and Fc membrane re- ceptors can be detected on their surface. Only about one third of the foam cells, however, exhibit C3 receptors and few if any of the cells appear to contain or to secrete lysozyme. Key words: Atherosclerosis, cholesterol, Fc re- ceptors, foam cells, lysosomes, macrophages, smooth muscle cells Foam cells are a characteristic cellular component of atherosclerosis, whether the disease develops spontaneously or is induced experimentally. The cells are filled with lipid and by light microscopy exhibit a “foamy” appearance from which they draw their name. Although the presence of foam cells in atherosclerotic lesions has been re- cognized for many decades, many relevant aspects of their cell biology have not been well studied. Little is known, for example, about the nature or mode of formation of the lipid inclusions in these cells and even the cellular origin of the foam cells is not estab- lished. Information about foam cells has been difficult to obtain in the past because of the inaccessability of these cells within the arte- rial wall. Recent advances in cell biology, in particular the use of enzymic procedures for the isolation of cells directly from tissues, now permit a detailed study of various tissue cell populations by sensitive biochemical and morphological tools. This report will describe the use of such techniques for the study of C 0 .- + E: c C 0) 0 C 0 0 O)‘ > 0 0, .- c - %Volume Fig. 1. Separation of isolated rabbit athero- matous aortic cells by Metrizamide density gradent centrifugation. The graph shows distri- butions of cells as represented by DNA, and their content of cholesterol and two acid hydrolases, N-acetyl-P-glucosaminidase and p-galactosidase. The ))staircase)) on top indicates the densities of individual fractions. The two shaded areas re- present the sub-populations of low density and high density cells selected for subsequent bio- chemical and morphologic analyses. For methodo- logical details, see ref 9.
-
Upload
stanley-fowler -
Category
Documents
-
view
212 -
download
0
Transcript of Characterization of Foam Cells in Experimental Atherosclerosis

Acta Med Scand (Suppl) 642: 151-158, 1980
Characterization of Foam Cells in Experimental Atherosclerosis
Stanley Fowler
From the Rockefeller University, New York, New York, U S A
ABSTRACT. A series of biochemical and morphological studies has focused on the properties and origins of lipid laden foam cells in experimentally induced atherosclero- sis in rabbits. Lipid inclusions present in these cells occupy half or more of the cyto- plasmic volume and are of two kinds: cyto- plasmic lipid droplets composed predomi- nantly of cholesteryl esters and lysosomes in which substantial quantities of free choles- terol have accumulated. The foam cells ex- hibit some properties of macrophages but not others. They possess high levels of acid hy- drolases and catalase and Fc membrane re- ceptors can be detected on their surface. Only about one third of the foam cells, however, exhibit C3 receptors and few if any of the cells appear to contain or to secrete lysozyme.
Key words: Atherosclerosis, cholesterol, Fc re- ceptors, foam cells, lysosomes, macrophages, smooth muscle cells
Foam cells are a characteristic cellular component of atherosclerosis, whether the disease develops spontaneously or is induced experimentally. The cells are filled with lipid and by light microscopy exhibit a “foamy” appearance from which they draw their name. Although the presence of foam cells in atherosclerotic lesions has been re- cognized for many decades, many relevant aspects of their cell biology have not been well studied. Little is known, for example, about the nature or mode of formation of the lipid inclusions in these cells and even the cellular origin of the foam cells is not estab-
lished. Information about foam cells has been difficult to obtain in the past because of the inaccessability of these cells within the arte- rial wall. Recent advances in cell biology, in particular the use of enzymic procedures for the isolation of cells directly from tissues, now permit a detailed study of various tissue cell populations by sensitive biochemical and morphological tools. This report will describe the use of such techniques for the study of
C 0 .- + E : c C 0) 0 C 0 0
O ) ‘ > 0 0,
.- c -
%Volume Fig. 1. Separation of isolated rabbit athero- matous aortic cells by Metrizamide density gradent centrifugation. The graph shows distri- butions of cells as represented by DNA, and their content of cholesterol and two acid hydrolases, N-acetyl-P-glucosaminidase and p-galactosidase. The ))staircase)) on top indicates the densities of individual fractions. The two shaded areas re- present the sub-populations of low density and high density cells selected for subsequent bio- chemical and morphologic analyses. For methodo- logical details, see ref 9.

152 S . Fowler
foam cells in experimental atherosclerosis, focusing on characteristic properties of the cells and eslpecially on the nature of the lipid deposits within them.
Isolation of lipid laden aortic cell populations
Large numbers of cells filled with lipids ap- pear in the aortas of rabbits fed for several months on a diet enriched in cholesterol. These cells can be isolated from the rabbit aortas by collagenase and elastase digestion which breaks down the extracellular tissue matrix in which the cells are embedded (9). The cells which are released form a mor- phologically and biochemically heterogeneous mixture, showing varying degrees of intra- cellular lipid deposition. These cells can be
separated, depending on their bouyant den- sity, by means of Metrizamide density gra- dient centrifugation. The separation achieved is illustrated in Fig. 1. As shown by the dis- tribution of DNA, the cells are spread throughout the entire Metrizamide density gradient. Many cells, however, equilibrate in the density range of 1.10-1.14. When viewed by electron microscopy most of these cells contain deposits of lipid and can be identified as arterial smooth muscle cells. A second, smaller population of cells equilibrates a t a much lower density (1.03-1.07). These cells are greatly enriched in cholesterol and cho- lesteryl esters and in lysosomal enzymes. Morphologically, they are seen to be heavily laden with lipid and appear as typical arterial foam cells (Fig. 2). For the biochemical and
Fig. 2. Portion of rabbit low density foam cell filled with lipid. Lipid deposition appears in two forms, as electron translucent cytoplasmic droplets (D) and as polymorphic vacuoles (Ly) containing amorphous debris and bounded by a membrane. The latter structures can be identified as lyso- somes by acid phosphatase cytochemistry. N = nucleus (X 14,500).

Characterization of foam cells 153
morphological studies to be described next, the cells in gradient fractions of densities 1.02-1.07 and 1.09-1.14 were pooled sepa- rately as indicated by the shaded areas of Fig. 1. These cell preparations will be re- ferred to as low density foam cells and as high density atheromatous cells, respectively.
Analysis of lipid inclusions in aortic cells
Sub-cellular fractionation studies employing sucrose density gradient centrifugation tech- niques show that the atheromatous aortic cells, and in particular the low density foam cells, possess lysosomes that equilibrate a t much lower densities than normal (9, 11). Their unusual bouyancy suggested that such lysosomes might contain lipid. Electron mi- croscopy of the atheromatous cells confirmed this notion, revealing that lipid deposition in these cells occurs in two distinct morphologic forms (Fig. 2): as polymorphic membrane- bounded structures filled with lipid which can be identified as lysosomes by acid phos- phatase staining, and as lipid droplets present free in the cytoplasm (13, 14). Morphometric analyses of electron micrographs of the low density foam cells (Table I) demonstrate that the lipid laden lysosomes and cytoplasmic lipid droplets together occupy 40-60 O / o of the cytoplasm. The lysosomes account for half or more of the lipid inclusion volume in these cells (14). Lipid deposits in the high density atheromatous cells represent a much smaller proportion (< 20 O i o ) of the cyto- plasmic volume, and the cytoplasmic lipid droplets are the predominant storage form.
Since cholesterol and cholesteryl esters are the lipids accumulating in greatest amounts in the foam cells, we attempted to establish in which of the lipid inclusions they occur (15). Taking advantage of the well known reaction of digitonin with unesterified sterols to form insoluble precipitates, we examined the cytoplasm of the foam cells by electron microscopy after aldehyde fixation in the
Table 1. Morphometric analysis of lipid in- clusions in rabbit atheromatous cells (mean f SD)”
Percent cytoplasmic volume
Cell type Experiment 1 Experiment 2 (N = 30 cells) (N = 28 cells)
Low density foam cells
lipid laden lysosomes 22.6 f 5 40.4 f 6 cytoplasmic lipid
High density athero- matous cells
lipid laden lysosomes 1.8 f 1’2 5.1 f 9 cytoplasmic lipid droplets 4.9 k a 12.9 2 7
droplets 26.3 f 5 21.2 f 4
a : Derived from Shio et al. (ref. 14)
presence of digitonin. As shown in Fig. 3, tubular structures called “spicules”, charac- teristic of lipid-digitonide complexes, were observed in discrete sites of the cell cyto- plasm, distinct from the lipid droplets. If the foam cells were first stained cytochemically for acid phosphatase and then treated with digitonin-aldehyde fixative, enzyme reaction product was found associated with the spic- ules (Inset, Fig. 3) indicating that the lyso- somes of the foam cells possess digitonin- reactive lipid, presumably cholesterol.
Analyses of sub-cellular fractions obtained after isopyknic centrifugation of homogenates of low density foam cells demonstrated co- equilibration of most of the intracellular un- esterified cholesterol with low density lyso- somes. Cholesteryl esters were also associated with low density lysosomes, although most were found in a lipid droplet fraction of very low density. Taken together, the morpho- logical and biochemical studies indicate that in rabbit foam cells, lysosomes are the site of accumulation of free cholesterol and that both cytoplasmic lipid droplets and lysosomes are depot sites for cholesteryl esters.

154 S. Fowler
Fig 3. Appearance of cytoplasm of rabbit foam cell after treatment with digitonin containing fixative. Digitonin-induced spicules (*) appear in discrete areas of the cytoplasm, separate from electron translucent regions (D) which are presumed to be lipid droplets. The association of acid phosphatase reaction product with the spicules (inset) indicates digitonin reactive lipid is in lysosomes. For ex- perimental details, see ref 15. N = nucleus (X 24,000;'nset X 30,000).
Studies on the origin of the low density foam cells
Controversy exists over the origin of foam cells in atherosclerosis (7). Some authors con- tend that arterial smooth muscle cells are gradually transformed into foam cells through the intracellular accumulation of lipids that are only slowly metabolized and disposed of by the cells. Others propose that foam cells are wandering macrophages that have entered the arterial wall, engulfed lipid containing debris, and thereby turned into foam cells. Differences in opinion arise mainly because of the inadequacy of cell identification by morphological criteria alone, especially since during lipid overloading, characteristic ultrastructural features are lost in foam cells. As already mentioned, the high density atheromatous cell population consists
of a mixture of smooth muscle cells in varying stages of lipid accumulation. The ap- pearance of these cells suggests the possibility of their progressive evolution into low den- sity foam cells. The alternative, that some or all of the low density foam cells are derived instead from wandering macrophages cannot, of course, be excluded. To differentiate be- tween these two possibilities, we applied sev- eral biochemical and immunological tests to determine if the foam cells possess properties characteristic of macrophages, such as high levels of certain enzymes, the presence of typical surface membrane receptors, and the ability to ingest particles (4). Alveolar macro- phages, isolated from the same animals, were also analyzed for purposes of comparison 'n these studies.
In a survey of their content of various en- zymes, we found both the low density foam
Characterization of foam cells 155
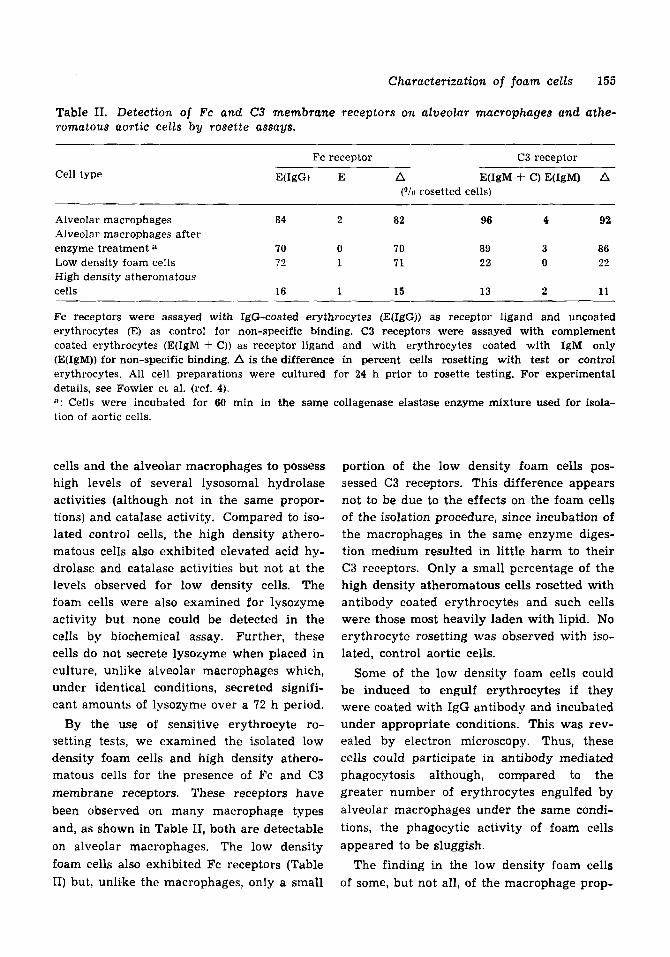
Table 11. Detection of Fc and C3 membrane receptors on alveolar macrophages and athe- romatous aortic cells by rosette assays.
Cell type Fc receptor C3 receptor
E(IgG) E A E(1gM + C) E(IgM) A ("/u rosetted cells)
Alveolar macrophages 84 2 82 w 4 92 Alveolar macrophages after enzyme treatment a 70 0 70 89 3 86 Low density foam cells 72 1 71 22 0 22 High density atheromatous cells 16 1 15 13 2 11
Fc receptors were assayed with IgG-coated erythrocytes (E(1gG)) as receptor ligand and uncoated erythrocytes (E) as control for non-specific binding. C3 receptors were assayed with complement coated erythrocytes (E(1gM f C)) as receptor ligand and with erythrocytes coated with IgM only (E(1gM)) for non-specific binding. A is the difference in percent cells rosetting with test or control erythrocytes. All cell preparations were cultured for 24 h prior to rosette testing. For experimental details, see Fowler et al. (ref. 4). a: Cells were incubated for &o min in the same collagenase elastase enzyme mixture used for isola- tion of aortic cells.
cells and the alveolar macrophages to possess high levels of several lysosomal hydrolase activities (although not in the same propor- tions) and catalase activity. Compared to iso- lated control cells, the high density athero- matous cells also exhibited elevated acid hy- drolase and catalase activities but not a t the levels observed for low density cells. The foam cells were also examined for lysozyme activity but none could be detected in the cells by biochemical assay. Further, these cells do not secrete lysozyme when placed in culture, unlike alveolar macrophages which, under identical conditions, secreted signifi- cant amounts of lysozyme over a 72 h period.
By the use of sensitive erythrocyte ro- setting tests, we examined the isolated low density foam cells and high density athero- matous cells for the presence of Fc and C3 membrane receptors. These receptors have been observed on many macrophage types and, as shown in Table 11, both are detectable on alveolar macrophages. The low density foam cells also exhibited Fc receptors (Table 11) but, unlike the macrophages, only a small
portion of the low density foam cells pos- sessed C3 receptors. This difference appears not to be due to the effects on the foam cells of the isolation procedure, since incubation of the macrophages in the same enzyme diges- tion medium resulted in little harm to their C3 receptors. Only a small percentage of the high density atheromatous cells rosetted with antibody coated erythrocytes and such cells were those most heavily laden with lipid. No erythrocyte rosetting was observed with iso- lated, control aortic cells.
Some of the low density foam cells could be induced to engulf erythrocytes if they were coated with IgG antibody and incubated under appropriate conditions. This was rev- ealed by electron microscopy. Thus, these cells could participate in antibody mediated phagocytosis although, comlpared to the greater number of erythrocytes engulfed by alveolar macrophages under the same condi- tions, the phagocytic activity of foam cells appeared to be sluggish.
The finding in the low density foam cells of some, but not all, of the macrophage prop-

156 S. Fowler
erties tested is puzzling. A possible explana- tion for these apparent discrepancies may be found in the studies of Zucker-Franklin et al. (19) who recently described the transforma- tion of monocytes into giant fat cells when blood monocytes were cultured in soft agar. Following the transformation, Fc receptors were retained, complement receptors were lost and lysozyme secretion was not detected. The evidence we have presented, therefore, would seem to point to a macrophage origin of the low density foam cells.
This is not the only interpretation possible, however. Fc membrane receptors, for exam- ple, are not unique to macrophages alone. These receptors are found on lymphoid cells and various types of leukocytes, on cultured fibroblasts infected with Herpes virus (lo), and even on vascular endothelial cells (12). It is conceivable therefore that during fatty transformation, vascular smooth muscle cells (high density cells) could acquire the Fc re- ceptors we have observed. Phagocytosis is also not a unique property of macrophages. There are several reports describing particle engulfment by vascular smooth muscle cells both in vivo and in culture (1, 6, 16, 17). The high levels of lysosomal enzyme activities characteristic of the low density foam cell, t should be noted, are also typical of proli- ferating smooth muscle cells (5) as well as of macrophages.
Since only about one third of the low den- sity foam cells exhibited both Fc and comple- ment receptors and a similar fraction engaged in receptor mediated phagocytosis, it is also possible that the rabbit low density foam cell preparation contains cells derived both from macrophages and from arterial smooth mus- cle cells. Our results are thus somewhat am- biguous. We believe that before the ques- tion of the origin@) of foam cells can be fully resolved, pre-labelling of macrophages and smooth muscle cells will be required and their movements into arterial lesions followed.
Significance of the findings
We have applied several of the methods de- scribed here for the study of rabbit foam cells to an analysis of foam cells in experimental atherosclerosis in monkeys (3) and to the in- vestigation of similar cells in spontaneous human lesions (2). Although studied in less detail, many qualitative similarities in the foam cells are found, including the presence of both lipid laden lysosomes and cytoplasmic lipid droplets. Associated with them were elevated cellular levels of acid hydrolase and catalase activities. The cellular mechanisms by which lipids accumulate in foam cells are under active investigation in a number of laboratories and it is thought that uptake of lipid by endocytosis of lipoproteins plays an important role in lipid overloading (8, 18).
Less well understood is the contribution foam cells make to the formation or progres- sion of arterial lesions. Do the lipids depos- ited in foam cells account for a significant portion of the lipid accumulating in athero- matous plaques? Are the hydrolases present in foam cells responsible for destruction of the matrix components of the arterial wall? Do foam cells interact with neighbouring cells, for example by secretion of growth promoting agents? These questions will no doubt be topics of future research.
ACKNOWLEDGEMENTS
I am indebted to my co-workers, Dr N. J. Haley and Ms H. Shio for their important contributions to the studies described. I am also grateful for the assistance and discus- sions given by Drs P. A. Berberian, A. Yau- Young and also by Dr H. Wolinsky of the Albert Einstein College of Medicine. I also wish to thank Dr C. de Duve for his active interest and continued encouragement in this work.

Characterization of foam cells 157
This work was supported in part by U S Public Health Service Grant HL-18157 from the National Heart, Lung, and Blood Institute and in part by a grant-in-aid from the New York Heart Association. Dr Fowler is an Otto G. Storm Established Investigator of the American Heart Association.
DISCUSSION
Professor Ross (Seattle) said that it was reasonably well established that macrophages in most tissue were monocyte derived. When a monocyte entered a tissue it took on the characteristics of that tissue. Unfiortunately they did not know how entry into the aorta would alter the monocytes' presentation. He thought it possible that there might be some heterogeneity in Dr Fowler's high density and low density populations both perhaps being unequal mixtures of monocytes which had become macrophages and which had taken on particular characteristics. The critical point was to identify such changes so that there still remained some means of re- cognising these cells.
Several years previously his group had described how activated monocytes could be converted to macrophages that secreted a very potent mitogen in culture. He did not know whether such a mitogen was found in vivo although there were suggestions that it was, and if so, the presence of large numbers of macrophages in the lesions could provide an opportunity for stimulating the prolifera- tive process. It was also important to enquire how the cell population in the fatty streak, which appeared to be enriched with poten- tially monocyte derived cells, differed from the later lesions which contained other mix- tures of these cell populations. Comparison was certainlly required of earlier versus later lesions in terms of the cell populations
Dr Fowler said it was certainly important to establish whether macrophages did secrete mitogens. He had referred to this problem in his presentation. As to the question of whether there miglht be macrophages present in the high density cell population, they had searched for them with the electron micro- scope but found none among those cell types that they recognized. But there were other cells present containing large quantities of lipid and the possibility that they were atypical macrophages could not be overlodked.
Professor Nordey (Tromso) asked how one could be sure that the macrophages appearing in rabbits on a cholesterol diet were similar to those found in the human body.
Dr Fowler said that unfortunately one had to rely so far on morphological criteria, i.e. the two cell populations looked the same. But having worked out the technique in the rabbit, he now hoped to apply it to human beings.
Professor Ross (Seattle) had some observa- tions relevant to this question. He had grown cells in culture from human lesions and was able to identify two clear cell populations. One of them did not divide but did contain Fc receptors and were clearly macrophages. The others did divide and were smooth muscle cells. He had been surprised at the initially high percentage of macrophages in the lesions but there was no doubt that they were present.
Dr Fowler said that one had to be careful in assuming that the presence of Fc receptors was indicative of macrophages. Fibroblasts in culture inoculated with virus produced such receptors and endothelial cells were also re- ported to have them. The potential for their production was in the genome and a number of stimulants would cause it to be expressed. Unfortunately there was no single specific criterion that could be used to label a cell as being a macrophage.
Professor Ross (Seattle) said that Brown and Goldstein had recently suggested that macro-
present. phages had receptors for acetylated LDL and

158 S. Fowler
smooth muscle cells d i d not. That would represent a clear way of separating the two cell types.
Professor Woolf (London) said that a time sequence study on proliferating smooth muscle cells following balloon i n j u r y might also be of value. One could see whether the cell population altered any characteristics and accumulated l ipid on feeding with cholesterol.
Dr Fowler had appl ied t h i s technique to smooth muscle cells in culture and had not observed the formation of any Fc receptors. However, he had not so far used LDL to in- duce l ipid accumulat ion.
REFERENCES
1.
2.
3.
4.
5.
6.
7.
Coltoff-Schiller, B., Goldfischer, S., Adamany, A. M. & Wolinsky, H.: Endocytosis by vas- cular smooth muscle cells in vivo and in vitro. Am J Pathol 83: 45, 1976. Fowler, S., Berberian, P. A., Haley, N. J. & Shio, H. : Lysosomes, vascular intracellular lipid accumulation, and atherosclerosis. In: Proceedings of the International Conference on Atherosclerosis, Milan, 1977 (ed. J. Carlson, R. Paoletti, C. Sirtori, & G. Weber) p. 19. Raven Press, New York 1978. Fowler, S., Berberian, P. A., Shio, H., Gold- fischer, S. & Wolinsky, H.: Characterization of cell populations isolated from aortas of Rhesus monkeys with experimental atherosclerosis. Circ Res 46: 520, 1980. Fowler, S., Shio, H. & Haley, N. J.: Characteri- zation of lipid-laden aortic cells from choles- terol-fed rabbits. IV. Investigation of macro- phage-like properties or aortic cell popula- tions. Lab Invest 41: 372, 1979. Fowler, S. , Shio, H. & Wolinsky, H.: Sub- cellular fractionation and morphology of calf aortic smooth muscle cells. Studies on whole aorta, aortic explants, and subcultures grown under different conditions. J Cell Biol 75: 166, 1977. Garfield, R. E., Chacko, S. & Blose, S . : Phago- cytosis by muscle cells. Lab Invest 33: 418, 1975. Geer, J. C. & Haust, D. A.: Smooth muscle
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
cells in atherosclerosis. In: Monographs on Atherosclerosis (ed. 0. S. Pollack, H. S. Simms & J. E. Kirk) Vol. 2. S. Karger, New York 1972. Goldstein, J. L. & Brown, M. S.: The low density lipoprotein paithway and its relation to atherosclerosis. Ann Rev Biochem 46: 897, 1977. Haley, N. J., Shio, H. & Fowler, S.: Charac- terization of lipid laden aortic cells from cholesterol-fed rabbits. I. Resolution of aortic cell population by Metrizamide density gradient centrifugation. Lab Invest 37: 287, 1977. Kerbel, R. S.: Herpes virus induction of Fc receptors. Nature (Lond) 263: 192, 1976. Peters, T. J. and de Duve, C.: Lysosomes of the arterial wall. 11. Subcellular fractiona- tion of aortic cells from rabbits with experi- mental atheroma. Exp Mol Pathol 20:228, 1974. Shadforth, M. F., Cunningham, P. H., Andrews, B. S.: The demonstration of complement Clq and Fc-IgG receptors on the surface of human endothelial cells. Fed Proc 38: 1075, 1979. Shio, H., Farquhar, M. G. & de Duve, C.: Lysosomes of the arterial wall. IV. Cytochemi- cal localization of acid phosphatase and catalase in smooth muscle cells and foam cells from rabbit atheromatous aorta. Am J Path 76: 1, 1974. Shio, H., Haley, N. J. & Fowler, S.: Charac- terization of lipid-laden aortic cells from cholesterol-fed rabbits. 11. Morphometric analysis of lipid-filled lysosomes and lipid droplets in aortic cell populations. Lab Invest 39: 390, 1978. iShio, H., Haley, N. J. & Fowler, S.: Charac- terization of lipid-laden aortic cells from cholesterol-fed rabbits. 111. Intracellular localization of cholesterol and cholesteryl ester. Lab Invest 41: 160, 1979. Sirnipson, C. F.: Phagocytosis by aortic modi- fied smooth muscle cells. Artery 3: 210, 1977. Weidenbach, H. & Massmann, J.: Erybhrocyte phagocytosis by arterial smooth muscle cells in experimental atherosclerosis. Exp Pathol (Jena) 15: 111, 1978. Wolinsky, H. & Fowler, S.: Participation of lysosomes in atherosclerosis. N Engl J Med 299: 1173, 1978. Zucker-Franklin, D., Grusky, G. & Marcus, A.: Transformation of monocytes into ),fat,, cells. Lab Invest 38: 620, 1978.