
Characterization New Type Sulfite Reductase Present ...jb.asm.org/content/153/3/1211.full.pdf ·...
10
Vol. 153. No. 3 JOURNAL OF BACTERIOLOGY, Mar. 1983, P. 1211-1220 0021-9193/83/031211-10$02.00/0 Copyright © 1983, American Society for Microbiology Characterization of a New Type of Dissimilatory Sulfite Reductase Present in Thermodesulfobacterium commune E. C. HATCHIKIAN1* AND J. G. ZEIKUS2 Laboratoire de Chimie Bacterienne-CNRS, 13277 Marseille Cedex 9, France'; and Department of Bacteriology, University of Wisconsin, Madison, Wisc onsin 537062 Received 23 September 1982/Accepted 15 December 1982 A new type of dissimilatory bisulfite reductase, desulfofuscidin, was isolated from the nonsporeforming thermophilic sulfate-reducing microorganism Thermo- desulfobacterium commune. The molecular weight of the enzyme was estimated at 167,000 by sedimentation equilibrium, and the protein was pure by both disc electrophoresis and ultracentrifugation. The bisulfite reductase was a tetramer and had two types of subunits with an a232 structure and an individual molecular weight of 47,000. The enzyme exhibited absorption maxima at 576, 389, and 279 nm, with a weak band at 693 nm. Upon the addition of dithionite, the absorption maxima at 576 and 693 nm were weakened, and a new band appeared at 605 nm. The protein reacted with CO in the presence of dithionite to give a complex with absorption peaks at 593, 548, and 395 nm. The extinction coefficients of the purified enzyme at 576, 389, and 279 nm were 89,000, 310,000, and 663,000 M- cm-, respectively. Siroheme was detected as the prosthetic group. The protein contains 20 to 21 nonheme iron atoms and 16 to 17 acid-labile sulfur groups per molecule. The data suggest the presence of four sirohemes and probably four (4Fe-4S) centers per molecule by comparison with desulfoviridin, the dissimila- tory sulfite reductase from Desulfovibrio species. The protein contains 36 cysteine residues and is high in acidic and aromatic amino acids. The N-terminal amino acids of the a and ,B subunits were threonine and serine, respectively. With reduced methyl viologen as electron donor, the major product of sulfite reduction was trithionate, and the pH optimum for activity was 6.0. The enzyme was stable to 70°C and denatured rapidly above this temperature. The dependence of T. commune bisulfite reductase activity on temperature was linear between 35 and 65°C, and the Qio values observed were above 3. The presence of this new type of dissimilatory bisulfite reductase in T. commune is discussed in terms of taxonomic significance. Three different types of dissimilatory sulfite reductases, desulfoviridin (21, 25), desulforubi- din (26), and P582 (54), have been isolated from the sulfate-reducing bacteria. The microorga- nisms belonging to the genus Desulfovibrio con- tain either desulfoviridin or desulforubidin. The former (16, 39), which is the green pigment of taxonomic significance for the genus Desulfovi- brio (40), was later identified as sulfite reductase (21, 25), whereas the reddish-brown protein de- sulforubidin (26) was isolated from an anoma- lous strain of Desulfovibrio desulfuricans lack- ing desulfoviridin (32). On the other hand, the microorganisms included in the genus Desulfo- tomaculum contain a characteristic pigment (P582) implicated in the reduction of sulfite (54, 55). These enzymes, termed bisulfite reductases, have been reported to reduce bisulfite (HSO3) mainly to trithionate, which appears to be an intermediary compound in the respiratory path- way of sulfite reduction to sulfide (10, 20, 25, 51). However, variable amounts of thiosulfate and sulfide are detected as products, depending on the concentrations of substrate and electron donor (9, 17, 19). The described sulfite reductases contain siro- hemes and iron-sulfur centers but can be readily differentiated on the basis of their spectral and redox properties (27, 34, 36). The physicochemi- cal properties of the three enzymes and the fact that the chromophore from desulfoviridin is ex- tracted as siroporphyrin indicate that the mem- bers of the iron tetrahydroporphyrin prosthetic group exist in very different environments with- in their respective proteins (36). A new sulfate-reducing bacterium, designated Thermodesulfobacterium commune, was recent- ly isolated (J. G. Zeikus, M. A. Dawson, T. E. Thompson, K. Ingvorsen, and E. C. Hatchikian, 1211 on September 7, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript of Characterization New Type Sulfite Reductase Present ...jb.asm.org/content/153/3/1211.full.pdf ·...

Vol. 153. No. 3JOURNAL OF BACTERIOLOGY, Mar. 1983, P. 1211-12200021-9193/83/031211-10$02.00/0Copyright © 1983, American Society for Microbiology
Characterization of a New Type of Dissimilatory SulfiteReductase Present in Thermodesulfobacterium commune
E. C. HATCHIKIAN1* AND J. G. ZEIKUS2
Laboratoire de Chimie Bacterienne-CNRS, 13277 Marseille Cedex 9, France'; and Department ofBacteriology, University of Wisconsin, Madison, Wisc onsin 537062
Received 23 September 1982/Accepted 15 December 1982
A new type of dissimilatory bisulfite reductase, desulfofuscidin, was isolatedfrom the nonsporeforming thermophilic sulfate-reducing microorganism Thermo-desulfobacterium commune. The molecular weight of the enzyme was estimatedat 167,000 by sedimentation equilibrium, and the protein was pure by both discelectrophoresis and ultracentrifugation. The bisulfite reductase was a tetramerand had two types of subunits with an a232 structure and an individual molecularweight of 47,000. The enzyme exhibited absorption maxima at 576, 389, and 279nm, with a weak band at 693 nm. Upon the addition of dithionite, the absorptionmaxima at 576 and 693 nm were weakened, and a new band appeared at 605 nm.
The protein reacted with CO in the presence of dithionite to give a complex withabsorption peaks at 593, 548, and 395 nm. The extinction coefficients of thepurified enzyme at 576, 389, and 279 nm were 89,000, 310,000, and 663,000 M-cm-, respectively. Siroheme was detected as the prosthetic group. The proteincontains 20 to 21 nonheme iron atoms and 16 to 17 acid-labile sulfur groups per
molecule. The data suggest the presence of four sirohemes and probably four(4Fe-4S) centers per molecule by comparison with desulfoviridin, the dissimila-tory sulfite reductase from Desulfovibrio species. The protein contains 36 cysteineresidues and is high in acidic and aromatic amino acids. The N-terminal aminoacids of the a and ,B subunits were threonine and serine, respectively. Withreduced methyl viologen as electron donor, the major product of sulfite reductionwas trithionate, and the pH optimum for activity was 6.0. The enzyme was stableto 70°C and denatured rapidly above this temperature. The dependence of T.commune bisulfite reductase activity on temperature was linear between 35 and65°C, and the Qio values observed were above 3. The presence of this new type ofdissimilatory bisulfite reductase in T. commune is discussed in terms of taxonomicsignificance.
Three different types of dissimilatory sulfitereductases, desulfoviridin (21, 25), desulforubi-din (26), and P582 (54), have been isolated fromthe sulfate-reducing bacteria. The microorga-nisms belonging to the genus Desulfovibrio con-tain either desulfoviridin or desulforubidin. Theformer (16, 39), which is the green pigment oftaxonomic significance for the genus Desulfovi-brio (40), was later identified as sulfite reductase(21, 25), whereas the reddish-brown protein de-sulforubidin (26) was isolated from an anoma-lous strain of Desulfovibrio desulfuricans lack-ing desulfoviridin (32). On the other hand, themicroorganisms included in the genus Desulfo-tomaculum contain a characteristic pigment(P582) implicated in the reduction of sulfite (54,55).These enzymes, termed bisulfite reductases,
have been reported to reduce bisulfite (HSO3)mainly to trithionate, which appears to be an
intermediary compound in the respiratory path-way of sulfite reduction to sulfide (10, 20, 25,51). However, variable amounts of thiosulfateand sulfide are detected as products, dependingon the concentrations of substrate and electrondonor (9, 17, 19).The described sulfite reductases contain siro-
hemes and iron-sulfur centers but can be readilydifferentiated on the basis of their spectral andredox properties (27, 34, 36). The physicochemi-cal properties of the three enzymes and the factthat the chromophore from desulfoviridin is ex-tracted as siroporphyrin indicate that the mem-bers of the iron tetrahydroporphyrin prostheticgroup exist in very different environments with-in their respective proteins (36).A new sulfate-reducing bacterium, designated
Thermodesulfobacterium commune, was recent-ly isolated (J. G. Zeikus, M. A. Dawson, T. E.Thompson, K. Ingvorsen, and E. C. Hatchikian,
1211
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

1212 HATCHIKIAN AND ZEIKUS
J. Gen Microbiol., in press). This extreme ther-mophilic microorganism grows above 70°C anddiffers from all of the bacteria described so farbecause it contains nonisopranoid branchedglycerol diethers and mono-ethers (T. A. Lange-worthy et al., submitted for publication). Wewere previously unable to detect desulfoviridinin this species, and we report here that it con-tains a unique sulfite reductase.
MATERIALS AND METHODSChemicals and enzymes. Potassium trithionate was
synthesized as described by Stamm and Goehring (50)and was a generous gift of A. Le Faou (Institut deBacteriologie, Strasbourg, France). Hydrogenase waspurified from Desulfovibrio gigas as reported previ-ously (14). Desulforubidin used for comparative stud-ies was purified from D. desulfuricans (Norway 4) bythe procedure of Lee et al. (26). All other chemicalswere of reagent grade quality.Organism and grpwth conditions. T. commune type
strain YSRA-1 (DSM 2178, ATCC 33708) was grownat 65°C in the low-phosphate-buffered basal mediumdescribed by Zeikus et al. (60) with 0.1% yeast extract,0.4% Na2SO4, and 0.4% sodium lactate.
Preparation of extracts. Unless otherwise noted, allbuffers were at pH 7.6, and all operations were per-formed aerobically at 4°C. Bisulfite reductase in theextracts was monitored by its enzymatic activity andits absorption at 576 nm. The bacteria were suspendedin 20 mM Tris-hydrochloride buffer (pH 7.6) contain-ing 100 ,uM DNase and were passed twice through aFrench pressure cell at 15,000 lb/in2. The extract wascentrifuged at 30,000 x g for 30 min, and the superna-tant fluid was then centrifuged for 2 h at 198,000 x g ina Spinco model L3-50 ultracentrifuge. The pellet waswashed once with 20 mM Tris-hydrochloride buffer(pH 7.6) and centrifuged in the same conditions. Thecrude soluble extract constituted by the mixture of thetwo 198,000 x g supernatants was utilized for thepurification of the enzyme.Enzyme assays. Bisulfite reductase activity was de-
termined manometrically as hydrogen uptake in asystem containing hydrogenase, sulfite reductase,methyl viologen (MV), and sulfite, as previously re-ported (25). The main compartment of the manometricvessel contained 150 ,umol of phosphate buffer (pH6.0), 2.2 ,umol of MV, 3 mg of bovine serum albumin,50 p.g of pure D. gigas hydrogenase with a specificactivity of 180 U/mg (14), and variable amounts ofenzyme. The sidearm contained 50 ,umol of sodiumsulfite freshly prepared, and the center well received0.1 ml of 10 N NaOH. The enzymatic activity wasroutinely measured at 45°C, and the final volume was 3ml. After preincubation under hydrogen for 30 min,the reaction was started by tipping in sulfite from thesidearm.The effect of temperature on enzymatic activity was
investigated by using a modified Warburg apparatus,allowing measurements at high temperatures.
Thiosulfate reductase activity present in the extractswas determined as previously reported (13). Trithion-ate reductase activity was measured manometricallyas hydrogen utilization by reaction mixtures contain-ing 150 ,umol of phosphate buffer (pH 7.0), 2.2 ,umol ofMV, 50 ,ug of D. gigas pure hydrogenase, 3 mg of
bovine serum albumin, and enzymatic extract. Thereaction was started by tipping in 50 ,umol of trithion-ate from the sidearm. The other assay conditions wereas reported for the bisulfite reductase activity.
Analytical determinations. Trithionate and thiosul-fate were estimated from the reaction mixtures accord-ing to the procedure of Kelly et al. (18). Trithionateformation was confirmed by thin-layer chromatogra-phy on silica gel, using a solvent phase of glycerol-n-butanol-acetone-water-30% NH40H (5:35:30:2.5)(42). Protein was determined according to the proce-dure of Lowry et al. (30), using bovine serum albuminas standard. The iron content of bisulfite reductasewas determined by atomic absorption spectrometry,using a Unicam model SP 1900 spectrometer. Inorgan-ic sulfide was estimated by the colorimetric method ofFogo and Popowsky (11) directly or as modified byLovenberg et al. (29) for protein.
Analytical gel electrophoresis was performed ac-cording to the method of Davis (7) on 7% polyacryl-amide gels at pH 8.8. Bisulfite reductase activity wasalso determined directly in the gel as the sulfite-dependent oxidation of reduced MV. The gels, afterelectrophoresis, were stained for 30 min in 10 ml of 100mM phosphate buffer (pH 6) containing 12 mM MVpreviously reduced by hydrogenase (250 ,ug of protein)under H2 and then were dipped into a solution contain-ing 20 mM sodium sulfite in the same buffer main-tained under argon. Sodium dodecyl sulfate (SDS)electrophoresis was performed according to Weberand Osbom (56), using a 10% acrylamide concentra-tion.Molecular weight determination. The molecular
weight of purified bisulfite reductase was determinedby analytical ultracentrifugation with a Beckman mod-el E analytical ultracentrifuge. It was estimated atequilibrium sedimentation by the meniscus depletionmethod as developed by Yphantis (58). Subunit molec-ular weights of the protein were estimated by the sameprocedure after extensive dialysis of the enzymeagainst 4 M guanidine-hydrochloride containing 0.1 M2-mercaptoethanol. Subunit molecular weight was alsodetermined by the method of Weber and Osborn (56),using protein samples in 10 mM phosphate buffer (pH7.0) containing 1% SDS and heated for 1 min in aboiling bath. The following protein standards wereused: bovine serum albumin, ovalbumin, chymotryp-sinogen A, soybean trypsin inhibitor, and cytochromeC.Anino acid analyses. Amino acid analyses were
performed on bisulfite reductase samples hydrolyzedfor 24, 48, and 72 h at 110°C in 6 M HCI in evacuated,sealed tubes according to the method of Moore andStein (33). The average was calculated from severaldeterminations. The amino acid composition was de-termined with a Beckman 121 M amino acid analyzer.Cysteine and methionine were analyzed after perfor-mic acid oxidation as cysteic acid and methioninesulfone, respectively, according to Hirs (15).
Determination of the N-terminal sequence. The N-terminal sequence of bisulfite reductase was estab-lished by automatic Edman degradation of the apopro-tein in a Beckman sequencer model 890 C, using aQuadrol (N-N-N'-N' tetrakis 2-hydroxypropyl ethy-lenediamine) 0.1 M Program. The quantitative deter-mination of the phenylthiohydantoin derivatives wasdone relative to known amounts of the appropriate
J. BACTERIOL.
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 153, 1983
stawdards by high-pressure liquid chromatography(waters Associates). The apoprotein was obtained byutatment of 25 nmol of bisulfite reductase with ace-t0n0ehydrochloride according to the procedure ofMurphy ct al. (35).Spectrophotometrk studies. Absorption spectra
were measured on a Kontron spectrophotometer mod-el Uvikon 810. Molar extinction coefficients of theprotein were obtained by measuring the values of theoptical densities of its absorption maxima, using asolution of known protein concentrations calculatedfrom amino acid analysis. The reduction of the bisul-fite reductase was performed under anaerobic condi-tions, using cuvettes sealed with rubber stoppers.
purifIction of bisulfite reductase. Freshly thawedcells of T. commune (430 g wet weight) were suspend-ed in 250 ml of 20 mM Tris-hydrochloride buffer. Asettled volume of Silica Gel (Merck type 60) equal to70 ml was added to the soluble extract (380 ml, 45 mg/ml), and the mixture was stirred for 3 h. This suspen-sion was subsequently poured into a column, and theunadsorbed proteins were collected. The unadsorbedproteins were then treated with DEAE-celiulose by abatchwise technique. A settled volume of DEAE-cellulose (100 ml) was added to the extract, and themixture was stirred for S h. The adsorbed acidicproteins were eluted with 1 M Tris-hydrochloridebuffer. The acidic extract (300 ml, 29 mg/ml) wasdialyzed overnight against 10 liters of distilled water,and the dialyzed proteins were loaded onto a DEAE-cellulose column (4.2 by 16 cm) previously equilibrat-ed with 20 mM Tris-hydrochloride buffer. The columnwas washed with 80 ml of the same buffer, and theadsorbed proteins were cluted by using a discontinu-ous gadient (800 ml) from 50 to 500 mM Tris-hydro-chloride buffer. The bisulfite reductase exhibited abrown color and was eluted with 300 mM Tris-hydro-chloride buffer, whereas the acidic electron carriers(fiavodoxin and ferredoxin) were eluted with 400 and500mM buffers, respectively. At this stage, the extractcontaining the enzyme (180 ml, 21 mg/nl) was dividedinto three parts of equal volume, and each fraction waspurified separately. The bisulfite reductase was passedthrough an Ultrogel AcA 34 column (5 by 100 cm) andequilibrated with 20 mM Tris-hydrochloride, and 8-mlfrctions were collected. The tubes, which had anabsorbancy ratio (A27,./Am,) lower than 2.7 were com-bined, and the subsequent fraction was adsorbed on aDEAE-cellulose column (3 by 12 cm) equilibrated with20 mM Tris-hydrochloride. The enzyme was eluted bya discontinuous gradient (400 ml) from 50 to 350 mMand was subsequently filtered through an UltrogelAcA 34 column (5 by 100 cm). Finally, the protein wasadsorbed onto a hydroxylapatite column (2.8 by 8 cm)equilibrated with 10 mM Tris-hydrochloride buffer andwas cluted by using a discontinuous gradient (600 ml)of phosphate buffer from 2 to 100 mM. The dark-brown-colored bisulfite reductase was judged to bepure from both polyacrylamide gel analysis and itsabsorption spectrum (A279IA3", 2.14). The purifiedenzyme exhibited a specific activity of0.20 pmol of H2utilized per min per mg of protein at 45°C. The yieldwas 117 mg of pure bisulfite reductase. This punrfica-tion procedure represents a fivefold purification ascompared with the specific activity of the acidic pro-tein fraction obtained after the batch treatment withDEAE-celiulose. The high specific activity of the
DESULFOFUSCIDIN OF T. COMMUNE 1213
crude extract in terms of hydrogen uptake (0.24 I.molof H2 utilized per mn per mg of protein) is due to thereduction of trithionate, the product of bisulfite reduc-tase, to sulfide by the other enzymes in the pathway asobserved previously with desulfoviindin (25). After thebatch tratment with DEAE-celulose, trithionate andthiosulfate reductase activities were separated fromthe bisulfite reductase, as shown by the presence ofthese activities in the unadsorbed protein fraction.
RESULTSPurity of the enzyme. After disc electrophore-
sis of the bisulfite reductase, one brownish bandwas observed in gels, and staining with Coomas-sic blue gave a single band with the same Rf asthe brownish pigment. The protein band in thegels directly catalyzed the sulfite-dependent oxi-dation of reduced MV. Furthermore, when thepurified enzyme was subjected to ultracentrifu-gation in 20 mM Tris-hydrochloride buffer (pH7.6) containing 0.1 M NaCI, a single brown-colored symmetrical peak was observed.
Molecular weight of native enzyme and sub-units. The molecular weight of bisulfite reduc-tase-determined by analytical ultracentrifuga-tion at equilibrium sedimentation-wasestimated to be 167,000, using a partial specificvolume (0.736) derived from amino acid analysis(6).The molecular weights of the subunits of
bisulfite reductase were determined by sedimen-tation equilibrium centrifugation of the enzyme,which had been dialyzed extensively against 4 Mguanidine-hydrochloride containing 0.1 M 2-mercaptoethanol. A molecular weight of 47,000was obtained, assuming that the native protein iscomposed of four subunits of equal molecularweight. The molecular weight of the subunitswas also estimated from the results of SDS-gelelectrophoresis. This procedure revealed thepresence of a single band which exhibited amolecular weight of 48,000 when compared withprotein standards. However, the determinationof the N-terminal sequence of the protein clearlyindicated the presence of two different polypep-tidic chains in the molecule. Therefore the na-tive protein is a tetramer exhibiting a quaternarystucture of an x2132 type. These different sub-units are not distinguishable by SDS-gel electro-phoresis.Absorptin spectra. The absorption spectra of
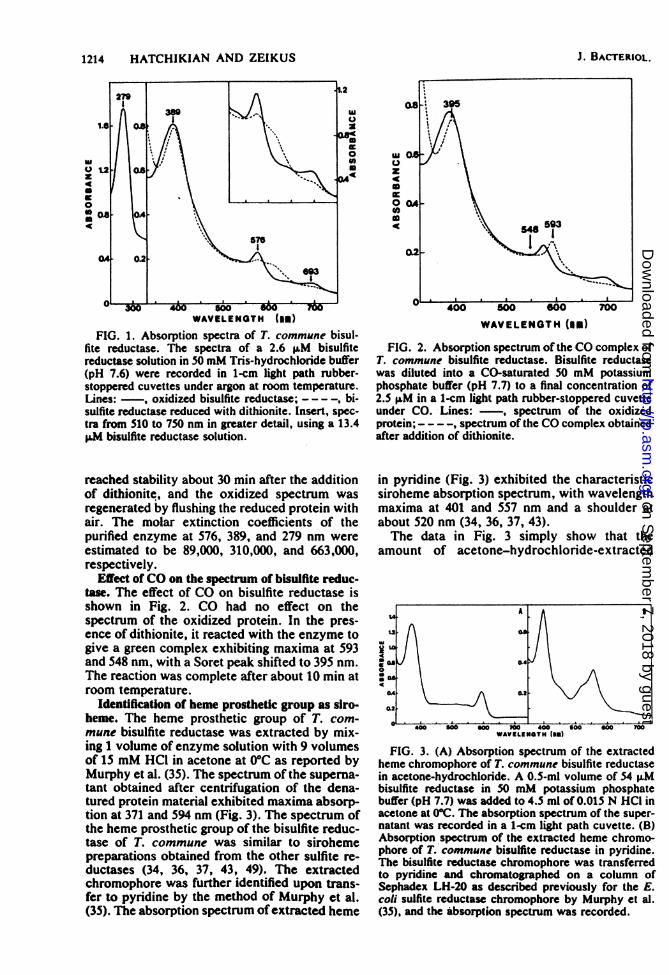
oxidized and dithionite-reduced bisulfite reduc-tase are shown in Fig. 1. The oxidized formexhibited major peaks at 576, 389, and 279 nm,with a weak absorption maximum at 693 nm anda shoulder at 532 nm. After addition of dithion-ite, the peaks at 576 and 693 nm decreased, and anew absorption band appeared around 605 nm.Furthermore, the Soret peak shifted from 389 to391 nm, and a shoulder appeared in the 420-nmregion. At room temperature, the spectrum
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
1214 HATCHIKIAN AND ZEIKUS
ou1.2X
30 4044o o
0
0
S.8 0.4-
693
WAVELENGTH (mm)FIG. 1. Absorption spectra of T. commune bisul-
fite reductase. The spectra of a 2.6 ,LM bisulfitereductase solution in 50 mM Tris-hydrochloride buffer(pH 7.6) were recorded in 1-cm light path rubber-stoppered cuvettes under argon at room temperature.Lines: -, oxidized bisulfite reductase; - - - -, bi-sulfite reductase reduced with dithionite. Insert, spec-tra from 510 to 750 nm in greater detail, using a 13.4LM bisulfite reductase solution.
reached stability about 30 min after the additionof dithionite, and the oxidized spectmm wasregenerated by flushing the reduced protein withair. The molar extinction coefficients of thepurified enzyme at 576, 389, and 279 nm wereestimated to be 89,000, 310,000, and 663,000,respectively.
Effect ofCO on the spectrum of bisulfite reduc-tse. The effect of CO on bisulfite reductase isshown in Fig. 2. CO had no effect on thespectrum of the oxidized protein. In the pres-ence of dithionite, it reacted with the enzyme togive a green complex exhibiting maxima at 593and 548 nm, with a Soret peak shifted to 395 nm.The reaction was complete after about 10 min atroom temperature.
Identificaton of henm prosthetic group as siro-heme. The heme prosthetic group of T. com-mune bisulfite reductase was extracted by mix-ing 1 volume of enzyme solution with 9 volumesof 15 mM HCI in acetone at O'C as reported byMurphy et al. (35). The spectrum of the superna-tant obtained after centrifugation of the dena-tured protein material exhibited maxima absorp-tion at 371 and 594 nm (Fig. 3). The spectrum ofthe heme prosthetic group of the bisulfite reduc-tase of T. commune was similar to sirohemepreparations obtained from the other sulfite re-ductases (34, 36, 37, 43, 49). The extractedchromophore was further identified upon trans-fer to pyridine by the method of Murphy et al.(35). The absorption spectrum ofextracted heme
us 0*
z4
A
coSc
400 No 500 700
WAVELENGTH (NM)
FIG. 2. Absorption spectrum of the CO complex ofT. commune bisulfite reductase. Bisulfite reductasewas diluted into a CO-saturated 50 mM potassiumphosphate buffer (pH 7.7) to a final concentration of2.5 g&M in a 1-cm light path rubber-stoppered cuvetteunder CO. Lines: -, spectrum of the oxidizedprotein; -, spectrum of the CO complex obtained
after addition of dithionite.
in pyridine (Fig. 3) exhibited the characteristicsiroheme absorption spectrum, with wavelengthmaxima at 401 and 557 nm and a shoulder atabout 520 nm (34, 36, 37, 43).The data in Fig. 3 simply show that the
amount of acetone-hydrochloride-extracted
AI
Iit
0S49
DA 0.2
400 0 00 100 400 400 400700ooooWAVELENGTH (em)
FIG. 3. (A) Absorption spectrum of the extractedheme chromophore of T. commune bisulfite reductasein acetone-hydrochloride. A 0.5-ml volume of 54 IMbisulfite reductase in 50 mM potassium phosphatebuffer (pH 7.7) was added to 4.5 ml of 0.015 N HCI inacetone at 0°C. The absorption spectrum of the super-natant was recorded in a 1-cm light path cuvette. (B)Absorption spectrum of the extracted heme chromo-phore of T. commune bisulfite reductase in pyridine.The bisulfite reductase chromophore was transferredto pyridine and chromatographed on a column ofSephadex LH-20 as described previously for the E.coli sulfite reductase chromophore by Murphy et al.(35), and the absorption spectrum was recorded.
-6 395
\1 54
iI.
J. BACTERIOL.
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 153, 1983
TABLE 1. Comparison of the amino acidcompositions of T. commune, D. gigas, and D.
vulgaris bisulfite reductases
Desulfo- DesuffoviridinAndnoacid fusckdinino acid fu(T. (D. (D.
commune) gigasj vulganis)
Aspartic acid 160 152 134Threonine 92 103 57Scrine 60 110 64Glutamic acid 200 202 136Proline 120 102 90Cystine (half) 36 NDd NDdGlycine 144 180 120Alanine 100 244 100Valine 108 134 86Methioninea 36 38 35isoleucine 136 103 82Leucine 104 140 92Tyrosine 68 50 47Phenylalanine 72 69 64Histidine 60 43 42Lysime 128 104 88Arginine 84 100 68
Cysteine and methionine were determined as cys-teic acid and methionine sulfone after performateoxidation of the protein according to the method ofHirs (1S).
* From Hall and Prince (12).¢ From Seki et al. (44)."ND, Not deternined.
beme from T. commune bisulfite reductase,based on the absorbance of the a peak (594 nm),is twice that of the heme prosthetic group ob-tained from desulforubidin under the same con-ditions (36).Content of Iron and aid-abile sulfur. The
concentration of bisulfite reductase was calcu-lated from amino acid analysis. The averagevalue of iron content of the enzyme estimatedfrom several analyses was 20 to 21 atoms permolecule of protein. The complete release oflabile sulfide from bisulfite reductase required 4h of incubation of the protein sample with thealkaline zinc reagent used by Suhara et al. (52).After these procedures, the presence of 16 to 17atoms of acid-labile sulfur per molecule wasestimated. Preliminary data of electron pam-magnetic resonance studies performed on thebisulfite reductase showed resonances consis-tent with the presence of high-spin ferric hemeand (4Fe-4S) type cluster in the protein (R. C.Cammack and E. C. Hatchikian, unpublisheddata). These results suggest that the native pro-tein contains four sirohemes and four (4Fe-4S)clusters per molecule.Amino ad compodtion. The amino acid com-
position of bisulfite reductase from T. communeis compared with those of Desu(fovibrio speciesin Table 1. The acidic properties of bisulfite
DESULFOFUSCIDIN OF T. COMMUNE 1215
reductase are explained by the preponderance ofacidic amino acids. The ratio (Asx + Glx)/(Lys+ Arg) is about 1.7. The enzyme exhibited ahigh content of the aromatic residues, tyrosineand phenylalanine. It contained 36 cysteine resi-dues, which is higher than the number requiredto link four (4Fe-4S) clusters. When comparedwith the amino acid composition of desulfoviri-din from D. gigas (12) and from D. vulgaris (44),T. commune bisulfite reductase shows a highcontent of isoleucine, a hydrophobic residue. Inaddition, it had higher amounts ofglutamic acid,lysine, and tyrosine, whereas the numbers ofserine, alanine, and leucine residues were lower.
N-terminal sequence of bisulfite reductase. TheN-teminal sequence of bisulfite reductase es-tablished by automatic Edman degradation ofthe apoprotein was determined up to 10 resi-dues. The results are shown in Table 2. The N-terminal sequencing data indicated the presenceof two distinct polypeptidic chains in the pro-tein, with threonine and scrine as N-terminalresidues, respectively. Quantitative determina-tions on the stable residues isoleucine and glu-tamic acid (step 2 of Edman degadation) indi-cated the presence of similar concentration ofeach residue (molar ratio of Glu to lieu = 0.85).
Catalytic roperties and stoichiometry of sulfitereduction. T. commune sulfite reductase re-duced sulfite but not thiosulfate, trithionate, ortetrathionate. Enzymatic activity was obtainedwith methyl or benzyl viologen. The optimumpH was in the acid region, at about 5.5 to 6.0,indicating that bisulfite was probably the actualionic species of sulfite functioning as substrate(51). At pH 6.0, trithionate was the major prod-uct ofthe reaction (Table 3). An average value of88% of sulfite sulfur appeared as trithionate,whereas trace amounts of thiosulfate and sulfidewere produced. At its optimum pH, T. communebisulfite reductase catalyzes the reduction of
TABLE 2. N-terminal sequencing data of T.commune bisulfite reductase4
ResiduesStep identified
1 Thr, Ser2 Glu, lle3 Val, Glu4 Lys, Lys5 Phe, Lys6 Lys, Lys7 Glu, Asp8 Leu, Thr9 Asp, Asp10 Pro, Lys
A The results indicate the two amino acids identi-fied in each step of Edman degradation and corre-sponding to the a and p chains of the protein.
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

1216 HATCHIKIAN AND ZEIKUS
TABLE 3. Products of bisulfite reduction catalyzedby bisulfite reductase of T. communeu
S032- Hz consumed Amt of product (4mol)(Imol) (S0mo)S,O2 0 H.S22 8.1 7.0 0.30 0.1 9810 3.4 2.8 0.20 0.11 92
3.8 2.9 0.21 0.14 934 1.8 1.1 0.15 0.09 92a Bisulfite reductase activity was measured by the
manometric assay after hydrogen uptake. The reactionemployed 2.2 mg of pure enzyme at 45C, and theincubation time was 45 min. When no more hydrogenwas consumed by the reaction, the products weredetermined colorimetrically.
sulfite mainly to trithionate, according to thestoichiometry reported by Lee and Peck (25)with desulfoviridin; however, sulfide and thio-sulfate slightly increased when low sulfite con-centrations were used.
It was observed that T. commune bisulfitereductase could also catalyze the reduction ofnitrite and hydroxylamine to NH3 as reportedwith other sulfite reductases (24, 46).
Effect of temperature on bisulfite reductaseactivity. The effect of temperature on bisulfitereductase activity was determined in phosphatebuffer (pH 6.0). Figure 4 shows an Arrhenius
cdD
.0
0~~~~~~~
E\
J
28 2.9 3 3.1 32 3.3
x 103T
FIG. 4. Arrhenius plot of T. commune bisulfitereductase. Temperature dependence of bisulfite reduc-tase activity was measured after hydrogen consump-tion, as reported in the text.
J. BACTERIOL.*
plot that summarizes the results. The enzymaticactivity was enhanced from 35 to nearly 700Cand no deflection point was observed in theslope of the curve. The maximum activity oc-curred between 65 and 70°C. No change in theproducts of the reaction was observed between35 and 70°C. The effect of temperature on bisul-fite reductase activity was associated with un-usually high Qlo values of 3.6 and 3.2 in the 35 to500C and 50 to 65°C temperature ranges, respec.tively. The value of the energy of activationcalculated from the slope of the data for the Vn,xplots between 35 and 65°C was 23.8 kcal/mol(99,580 J/mol). The enzyme exhibited a very lowactivity below 35°C. On the other hand, theactivity becomes less dependent on temperatureabove 65°C.
DISCUSSIONThe physicochemical and catalytic properties
of T. commune bisulfite reductase reported inthis paper are summarized in Table 4 and com-
TABLE 4. Comparison of molecular and catalyticproperties of T. commune and D. gigas bisulfite
reductases
Property Desulfofuscidin Desulfoviridin'Mol wt of native enzyme 167,000 200,000Mol wt of subunits 2a = 47,000 2a = 44,600
2(3 = 47,000 2(3 = 61,000Subunit composition cx2(32 a2(32Absorption maxima (nm) 279/663 279
extinction coefficient 389/310 390(mM' cm-') 576/89 408/200
693 580628
CO complex formation + -
Iron content (Fe per en- 20-21 10zyme molecule)b
Acid-labile sulfur (S per 16 8enzyme molecule)b
Cysteine residues per 36 NDmolecule
Most probable composi- 4 Sirohemes 2 Sirohemestion 4 x (4Fe-4S) 2 x (4Fe-4S)
Sp actc 2d 0.63Major product of sulfite S3062- S3062-
reductionChromophore extracted Siroheme Siroporphyrin
with acetone-hydro-chloride
Optimum pH 5.5-6.0 6.0Unstability temp 70°C NDI
a Data of D. gigas sulfite reductase were taken fromLee and Peck (25), Hall and Prince (12), and Murphyet al. (36).
b Atoms per enzyme molecule.' Specific activity is expressed as micromoles of H2
consumed per minute per milligram of protein.'d Specific activity determined at the optimum tem-
perature (65°C).' ND, Not determined.
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

FrOL. 153, 1983
d with those of the mesophilic bisulfiteeductase (desulfoviridin) from D. gigas. Al-
tough the two enzymes have similar molecularwcights and quarternary structure, they differ bytheir spectral characteristics and their content ofprosthetic groups. T. commune bisulfite reduc-tase is a tetramer which appears to be constitut-ed by two different types of subunits exhibitingthe same molecular weight. The enzyme fromthe therlmophilic sulfate-reducing microorgan-ism differs also from desulfoviridin by the natureofthe acetone-hydrochloride-extracted chromo-phore (see Table 4).The absorption spectrum of T. commune bi-
sulfite reductase shows maximum absorptionpeaks at 279, 389, and 576 nm, with a weakabsorption band at 693 nm. It differs both fromdesulfoviridin and desulforubidin, although itexhibits a spectrum of a hemoprotein with aSoret peak centered around 390 nm common tothe other bisulfite reductases. It lacks the pro-nounced absorption peak at 628 nm and the splitSoret peak present in desulfoviridin (25) as wellas the maximum absorption at 545 nm shown bydesulforubidin (26). It appears to be similar tothe CO-reacting pigment, P582, which is thedissimilatory sulfite reductase in the thermophil-ic sporeforming sulfate reducer D. nigrjficans(54). However, the long wavelength absorptionmaxima of T. commune bisulfite reductase (693and 576 nm) are shifted to the short wavelengthof approximately 6 nm when compared with thespectral characteristics of P582. In the presenceof dithionite, T. commune bisulfite reductaseformed a complex with CO characterized by ana peak at 593 nm. The name desulfofuscidin issuggested for the well-characterized thermal sta-ble bisulfite reductase of T. commune.The value of the extinction coefficient of T.
cormnfuqe bisulfite reductase at the Soret peak9cm = 310,000), which is twice that of D.
vulgaris desulfoviridin (24), is in good agreementwith the presence of four sirohemes per mole-cule, using the value reported recently by Lan-caster et al. (23). These data are confirmed bythe amount of acetone-hydrochloride-extractedheme, which is twice that obtained with desul-forubidin in the same conditions (36). This latterbisulfite reductase has been reported to containtwo sirohemes per molecule (27). Furthermore.the content of iron and acid-labile sulfur sug-gests that the native protein contains four siro-hemes and four (4Fe-4S) centers per moleculeinstead of two sirohemes and two (4Fe-4S) clus-ters in D. gigas desulfoviridin (12).
T. commune bisulfite reductase is distin-guished from the dissimilatory sulfite reductasesof mesophilic bacteria (12, 28) by its high con-tent of siroheme. The lack of available data onthe siroheme content of P582 from the thermo-
DESULFOFUSCIDIN OF T. COMMUNE 1217
philic bacterium D. nigr(ficans (34, 54, 55) doesnot allow us to conclude whether it differs fromT. commune sulfite reductase, although P582from the mesophilic sporeforming strain D. ru-minis has been reported to contain only twosirohemes per molecule (27). It is to be notedthat the increase of the number of active sitesper molecule of enzyme has been recently de-scribed with an alcohol dehydrogenase isolatedfrom a thermophilic microorganism (4).
T. commune bisulfite reductase shows higherthermostability than desulforubidin, the homolo-gous protein from the mesophilic sulfate-reduc-ing bacterium D. desulfuricans (Norway) (un-published data). With the T. commune enzyme,in contrast to desulforubidin, no absorbancechange of the chromophore occurred until 70°C.From these data, it can be postulated that siro-hemes are more strongly bound to the apopro-tein in the enzyme of the thermophilic organism.The greater thermostability of desulfofuscidinmay be related to higher amounts of hydropho-bic residues (e.g., isoleucine) in the thermophilicenzyme. This has been previously suggested toaccount for the generaily increased stability ofthermophilic enzymes (59). Another point tonotice is the discrepancy existing between theunstabiity temperature of the pure bisulfite re-ductase (approximately 70°C) and the maximumgrowth temperature of T. commune, which hasbeen reported to be in the vicinity of 80°C(Zeikus et al., in press). This could be attributedto the higher stabilization of this cytoplasmicprotein within the cell.The activity of T. commune bisulfite reduc-
tase appears to be highly dependent on tempera-ture, since Qlo values higher than 3 were ob-served from 40 to 65C. The enzyme activity atthe optimum temperature (650C) was threefoldhigher than that of desulfoviridin (Table 4). Incontrast to some enzymes from thermophilicorganisms (22, 28), it does not show a biphasicArrhenius plot.
In our experimental conditions, the mainproduct of sulfite reduction catalyzed by T.commune bisulfite reductase was trithionate, asreported with most dissimilatory sulfite reduc-tases (21, 24-26, 44). It is to be noted that duringthe last decade, two mechanisms of dissimila-tory bisulfite reduction to sulfide have beenpostulated. The first mechanism assumes thatbisulfite is reduced to hydrogen sulfide via oneenzyme, bisulfite reductase, without any freeintermediates (5, 46, 54, 55) as was demonstratedwith the assimilatory type sulfite reductases (2,48, 58). The second mechanism of bisulfite re-duction assumes that sulfite is reduced to sulfidevia three enzymes, bisulfite reductase, trithion-ate reductase, and thiosulfate reductase, withtrithionate and thiosulfate as free intermediates
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

1218 HATCHIKIAN AND ZEIKUS
(1, 8, 19, 20, 51). It was further demonstratedthat the products of sulfite reduction are depen-dent on the experimental conditions, includingthe pH of the reaction, the concentration ofreduced MV, and the concentration of sulfite (8,17, 19, 31). Siegel (46, 47) has subsequentlyreported that trithionate and thiosulfate are by-products of sulfite reduction which appear whenhigh concentrations of sulfite are used, and hehas suggested that the dissimilatory sulfite re-ductase reaction proceeds in classical one- ortwo-electron stages, with intermediates normal-ly remaining enzyme bound. One has also topoint out that siroheme itself exhibits sulfitereductase activity, as demonstrated recently(45), and that thiosulfate, which is the mainreduction product, could be considered an inter-mediate state compared with the productsformed by dissimilatory-type (S3062-) and byassimilatory-type (H2S) sulfite reductases. Aspostulated by Seki et al. (45), the function ofsiroheme in sulfite reductases could be modifiedby interactions with the protein moiety or othercofactors as iron-sulfur clusters. Thus, the ques-tion of whether additional free intermediatesoccur between sulfite and sulfide is still a matterofcontroversy, and, as reported by Thauer et al.(53), the definitive answer to this question mustawait the discovery of the physiological electrondonor of the dissimilatory bisulfite reductaseand further data about the intracellular pH andsulfite concentration.
Thiosulfate and trithionate reductase activi-ties appear to be present in the extract of T.commune; however, our data did not distinguishwhether these two activities were exhibited bythiosulfate reductase itself, as reported by Olt-mann and Stouthamer (38) with the thiosulfatereductase of Proteus mirabilis. These observa-tions could be indicative of the presence of thetrithionate pathway of bisulfite reduction in thismicroorganism.
Desulfoviridin was initially used as a biochem-ical criterion to distinguish the microorganismsbelonging to the genus Desulfovibrio (40, 41)from the sporeforming sulfate-reducing bacteriaof the genus Desulfotomaculum, which weresubsequently shown to contain the brown pig-ment P582 (27, 54). In addition, several anoma-lous strains of the genus Desulfovibrio, includingD. desulfuricans (Norway 4), contain anothertype of sulfite reductase (desulforubidin) (26)and have been shown to differ from most otherDesulfovibrio species by their morphologicaland physiological characters (3, 32). The type ofdissimilatory bisulfite reductase appears there-fore to be a valuable criterion for the taxonomyof the sulfate-reducing microorganisms. Thepresent work confirms the taxonomic signifi-cance of this criterion, since we have demon-
strated that the nonsporeforming thermophilicsulfate-reducing bacterium T. commune, whichwas classified in a new genus (Langeworthy etal., submitted for publication; Zeikus et al., inpress), contains a novel type of bisulfite reduc-tase differing clearly from the homologous en-zymes by its absorption spectrum, content ofsiroheme, and thermostability.
ACKNOWLEDGMENTSWe thank N. Foret for skillful technical assistace and P.
Sauve for the determination of the molecular weight of thebisulfite reductase by analytical ultracentrifugation. Thanksare also due to D. Moinier (Laboratoire d'Immunologie.Universitd d'Aix-Maseille 11) for the amino acid analysis andto J. Bonicel and M. Bruschi for the determination of the N-terminal sequence of bisulfite reductase. We are indebted toM. Scandellari and R. Bourrelli for providing mass cultures ofT. mmune.This investigation was supported by a grant from PIRSEM
to E.C.H. and by grat EY79-S42-2161 from the U.S. Depart-ment of Energy to J.G.Z.
LITERATURE CITED1. Akagl, J. M., M. Chan, and V. Adams. 1974. Observations
on the bisulfite reductase (PS82) isolated from Desum(foto-maculm nigr(ficans. J. Bacteriol. 128:240-244.
2. Asad, K., G. Ta_ua, ad R. S. Id kl. 1969. Methylviologeninked sulfite reductawse from spinach leaves. J.Biol. Chem. 244:4904-4915.
3. BebM, H., and N. Pfanig. 1977. Growth of sulfate-reduc-ing bacteria with sulfur as electron acceptor. Arch. Micro-biol. 112:115-117.
4. Brat, F., and L. G. LJnngdahl 1981. Characterization ofan alcohol dehydrogenase from Thermoanaerobacter eth-anolicas active with ethanol and secondary alcohols.Biochem. Biophys. Res. Commun. 100:793-799.
S. Chambers, L. A., and P. A. Trudlger. 1975. Are thiosul^fate and trithionate intermediates in dissimilatory sulfatereduction? J. Bacteriol. 123:36-40.
6. Cohn, E. J., and J. T. Edsall. 1943. Protein, amino acidand peptides. Reinhold, New York, p. 373.
7. Davs, B. J. 1964. Disc electrophoresis. 11. Method andapplication to human serum proteins. Ann. N.Y. Acad.Sci. 121:404-427.
8. Drake, RL L., and J. M. Akal. 1976. Product analysis ofbisulfite reductase activity isolated from Desulfovibriovwlgaris. J. Bacteriol. 126:733-738.
9. Drake, H. L, and J. M. Akag. 1977. Bisulfite reductase ofDesu#fovibrio vulgaris: explanation for product formation.J. Bacteriol. 132:139-143.
10. Drake, H. L., ad J. M. Akag. 1978. Dissimilatoryreduction of bisulfite by Desu4fovibrio vulgaris. J. Bacte-riol. 136:916-923.
11. Fop, J. K., and M. Pupo . 1949. Spectrophotometricdetermination of hydrogen sulfide. Anal. Chem. 21:732-734.
12. Hall, M. H., and R. H. PrInce. 1981. Iron-containingreductases: investigation of sirohaem-dependent reac-tions involving sulphur oxyanions in desulphoviridin fromDesuMfovibrio gigas. I. Characterization of physical andenzymic properties of desulphoviridin. J. Inorg. Nucl.Chem. 43:815-823.
13. Haclklan, E C. 1975. Purification and properties ofthiosulfate reductase from Desuffovibrio gigas. Arch.Microbiol. 105:249-256.
14. H a, E. C., M. Bdal, and J. LeGall. 1978.Charcterization of the periplamic hydrogenase fromDesuLfovibrio gi,gas. Biochem. Biophys. Res. Commun.82:451-461.
15. Hirs, C. H. W. 1967. Determination of cystine as cystek
J. BACTERtIOL.
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VoL. 153, 1983
acid. Methods Enzymol. 11:59-62.16. IshImot, M., J. Koyan`, and Y. NagaL 1954. Biochemi-
cal studes on sulfate-reducing bacteria. III. The cyto-chrome system of sulfte-reducing bacteria. J. Biochem.(Tokyo) 41:763-770.
17. J mes H. E., and G. W. Skyrlig. 1975. Effect of enzymica y conditions on sulfite reduction catalyzed by desul-fovirdn from Desdfovibrio gigas. Biochim. Biophys.Acta 37752-60.
is. Kel, D. P., L. A. Ch s,a P. A. Trudigr. 1969.Cyanolysis and spectrophotometric estimation oftrithion-ate in mixture with thiosulfate and tetrathionate. Anal.Chem. 41:896-901.
19. Kbyabl, K., V. SekI, and M. Iblmote. 1974. Biochemi-cal studies on sulfate-reducing bacteria. XIII. Sulfitereductase from Desu(fovibrio vulgarls-mechanism oftrithionate, thiosulfate and sulfide formation and enzy-maic properties. J. Biochem. (Tokyo) 75:519-529.
20. Koba l, K., S. Taclbana, and M. Ish1moto. 1969.Intermediary formation of trithionate in sulfite rductionby a sulfate-reducing bacterium. 3. Biochem. (Tokyo)65:155-157.
21. Koby"yahl, K., E. Takahasl, and M. Is_moto. 1972.Biochemical studies on sulfite-reducing bacteria. Xl. Puri-fication and some properties of sulfite reductase, desulfo-viridin. J. Biochem. (Tokyo) h8727487.
22. Lamed, R. J., and J. G. Zeiku. 1981. Novel NADP-linkedalcohol-aldehyd/cketone oxidoeductase in thermophilicethanokgenic bacteria. Biochem. J. 196:183-190.
23. Lace, J. R., J. M. Veg, H. Kami, N. R. OrmeJohon, W. H. Orm-Jokma,4 R. J. Krug, and L. M.Sege. 1979. Identification of the iron-sulfur center ofspinach ferredoxin-nitrite reductase as a tetranuclear cen-ter, and preliminary EPR studies of mechanism. J. Biol.Chem. 254:1268-1272.
24. Lee, J.-P., J. LeGall, and H. D. Pck, Jr. 1973. Isolation ofassimilatory-and dissimilatory-type sulfite reductasesfrom Desu(fovibrio vulgaris. J. Bacteriol. 115529-542.
25. Lee, J. P., and H. D. Pck, Jr. 1971. Purification of theenzyme reducing bisulfite to trithionate from Desulfovi-brio gigas and its identification as desulfoviridin. Bio-chem. Biophys. Res. Commun. 45:583-9.
26. Le, J.-P., C.-S. YI, 2. LeGall, and H D. , Jr. 1973.Isolation of a new pimnt, desulforubidin, from DesuLfo-vibro desuifuricans (Norway strain) and its role in sulfitereduction. J. Bacteriol. 115:453-455.
27. Liu, C. L., D. V. Der Valan, and H. D. Peck, Jr. 1979.On the redox properties of three bisulfite reductases fromthe sulfate-reducing bacteria. Biochem. Biophys. Res.Commun. 91:962-970.
28. Ljungdhl, L. G., and D. Sbard. 1976. Proteins fromthermophilic microorganisms, p. 147-187. In M. R. Hein-rieb (ed.), Extreme environments, mechanisms of mi-crobial adaptation. Academic Press, Inc., New York.
29. Leeberg, W., S. B. Buha , and J. C. Rabnwwb.1963. Studies on the chemical nature of clostridial ferre-doxins. J. Biol. Chem. 238:3899-3913.
30. Lawry, 0. H., N J. RoleWmg, A. L. Far, ad R. J.Randal. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
31. Maybew, S. G., R. Abels, and R. Pl . 1977. Theproduction of dithionite and SO2- by chemical reaction of(bi)sulphite with methyl viologen semiquinone. Biochem.Biophys. Res. Commun. 77:1397-1403.
32. Mer, J. D. A., and A. M. Salek. 1964. A sulphate-rducing bacterium containig cytochrome c3 but lackingdesulfoviridin. J. Gen. Microbiol. 37:419-423.
33. More, S., and W. H. Sen. 1963. Chronatoxraphicdetermination of ainuo acids by the use of automticrcording equipment. Methods Enzymol. 6:819-831.
34. Muphy, M. J., and L M. Segl 1973. Siroem andsirohydrocha. The basis for a new type of porphyrin-reled prosthec group common to both asimilatory uddisiniatory sulfite reductases. J. Biol. Cbat. 248-6911-6919.
DESULFOFUSCIDIN OF T. COMMUNE 1219
35. Murphy, M. J., L. M. Siegd, and H. Kamin. 1973.Reduced nicotinamide adenine dinucleotide phosphate-slte SCducta of enterobactena. 11. Identification of anw class of hem prothetic oup: an iron-tetrahydro-porphyrin (isobacteriochlorin type) with eight carboxylicacid gups. J. Biol. Chem. 246:2801-2814.
36. Murphy, M. 2.L M.SOIgel, H. Kamla, D. V. De nIa, J. P. Lee, J. LeGal, and H. D. Peck, Jr. 1973. An irontetrahydroporphyrin prosthetic group common to bothassimilatory and dissiimiatory sulfite reductases. Bio-chem. Biophys. Res. Commun. 54:82-88.
37. Mwphy, M. J., L. M. Sigd, S. R. Tove, and H. Kamin.1974. Siroheme: a new prosthetic group participating insix electron reduction reactions catalyzed by both sulfiteand nitrite reductases. Proc. Natl. Acad. Sci. U.S.A.71:612-616.
38. Oltmana, L. F., ad A. H. _butamr. 1975. Reduction oftetrathionate, trithionate and thiosulfate and oxidation ofsulphide in Proteus mirabilis. Arch. Microbiol. 105:135-142.
39. t J. IL 1956. Cytochrome c3 and desulphoviridin;pigments of the anaeobe Desulphovibrio desulphuricans.1. Gen. Microbiol. 14:545-572.
40. Podlgat, J. R. 1959. A diagnstic reaction of Desulphovi.brio desulphuricans. Nature (London) 183:481-482.
41. Pesgpte J. R., ad L L. Ca l. 1966. Classification ofDesutfovlbrio species, the nonsporulating sulfate-reducingbacteria. Bacteriol. Rev. 30:732-738.
42. Roy, A. B., and P. A. Trnge. 1970. The biochemistryof compounds of sulfur, p. 79. CambridgeUniversity Prss, London.
43. Schedd, M., an H. G. Truper. 1979. Purification ofThiobacllus denitrificans siroheme sulfite reductase andinvestigation of soe molecular and catalyti properties.Biochim. Biophys. Acta 566:454-467.
44. Seki, Y., K. Kobayadb, ad M. Is . 1979. Biochemi-cal studies on sulate-reducing bacteria. XV. Separationand comparison of two foms of desulfoviridin. J. Bio-chem. (Tokyo) 85:705-711.
45. Sdd, Y., N. Sopw and M. Ishlmoto. 1981. Siroheme asan active catalyst in sulfite reduction. J. Biochem. (To-kyo) 9.1487-1492.
46. Sigel, L. M. 1975. Biochemistry of the sulfur cycle, p.217-286. In D. M. Greenberg (ed.), Metabolic pathways.Metabolism of sulfur compounds, vol. 7. Academic Press.Inc., New York.
47. Slegd, L. M. 1978. Structure and function of siroheme andthe siroheme enzymes, p. 201-214. In T. P. Singer andR. N. Ondarza (ed.), Mechanisms of oxidizing enzymes.Elsevier/North-Holland Publishing Co., Amsterdam.
48. Sgel, L. M., P. S. Davs, an H. KaInC. 1974. Reducednicotinamide adenine dinucleotide phosphate sulfite re-ductase of enterobacteria. III. The Escherlchia coli hemo-havoprotein: catalytic parameters and the sequence ofelectron flow. J. Biol. Chem. 249:1572-1586.
49. Sbel, L. M., M. J. Murphy, and H. Kamla. 1973.Reduced icinamide adenine dinuclootide phosphate-sulfite reductase of enterobacteria. I. The Escherichia colihemoflavoprotein: molecular parameters and prostheticgroups. J. Biol. Chem. 24:251-264.
30. Stama, H., and M. Gobriln. 1942. Zur kenntnis derpolythionsaurn und ihrer bildung; VI. Neye verfagrebzyr darstekkybg vib jakuyntrutguibat ybd vib jakuynte-tratgibat. Z. Anorg. Ails. Chem. 250:226-228.
51. Sb, B.* and 2. M. AkagL 1969. Formation of thiosulfatefrom sulfite by Desa4fovlbrio vulgaris. J. Bactenol.9.210-215.
52. Suhd, K., K. Kanyama, S. Takemorl, ad M. Katagr.1974. Bovine adrenal steroid hydroxylae system. III.Reconstitution of adrenal Iron-sulfur protein. Biochim.Biophys. Acta 336:309-317.
53. Thno, R. K., K. J_u-e n, and K. Decker. 1977.Energy conservation in chemotrophic anaerobic bacteria.Bacteriol. Rev. 41:100-180.
54. Trudlier, P. A. 1970. Carbon-monoxide-acting pig-
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

1220 HATCHIKIAN AND ZEIKUS
ment from Desulfolomacdlum ngricans and its possiblermevance to sulfite reduction. J. Bacteriol. 10:158-170.
55. Trudhger, P. A., and L A.C . 1973. Reaction ofPM" from Desu(fotomaculum nigrificans with substrates.reducing agents and carbon monoxide. Biochim. Biophys.Acta 293:26-35.
56. Weber, K., and M. Osbw. 1969. he rliability ofmolecular weight determinations by dodecyl sulfate-poly-acrylamie gel clectrophoresis. J. Biol. Chem. 244:4406-4412.
57. Yoshmoto, A., and R. Sato. 1968. Studies on yeast sulfite
J. BACTERIOL,.
reducase. 1. Purifiation and characterization. Biochim.Biophys. Acta 153:555-575.
5. Ypbms, D. A. 1964. Equilibrium ultracentrifuption ofdilute solutions. Biochemistry 3:297-317.
59. Zelkus, J. G. 1979. Thermophilic bacteria: ecology, physi.ology and technology. Enzyme Microb. Technol. 1:243.252.
60. Zellks, J. G., P. W. Hge, and M. A. Andersen. 1979.Thermoanaerobium brockii gen. nov. and sp. nov., a newchemoorganotrophic. caldoactive, anaerobic bacterium.Arch. Microbiol. 122:41-8.
on Septem
ber 7, 2018 by guesthttp://jb.asm
.org/D
ownloaded from


















