
Characterization and immunolocalization of the protein ... fileof Sarcocystis muris cystozoites...
7
Parasitol Res (1989) 75:199-205 Parasitnlngy Research Springer-Verlag 1989 Characterization and immunolocalization of the protein contents of micronemes of Sarcocystis muris cystozoites (Protozoa, Apicomplexa) U. PohP, J.F. Dubremetz 2, and R. Entzeroth 1 1 Zoologisches Institut der Universitfit, Poppelsdorfer Schloss, D-5300 Bonn 1, Federal Republic of Germany 2 INSERM, U42, 369, Rue Jules - Guesde, F-59650 Villeneuve-D'Ascq, France Abstract. Affinity-purified polyclonal antibodies generated against micronemes obtained by the sub- cellular fractionation of Sarcocystis muris contain two major proteins of 16 and 17 kDa. Antibodies cross-react with microneme antigen of S. tenella and S. cuniculi but not with the S. sp. of wild boar (Sus scrofa). Microneme antigens could be traced by immunoelectron microscopy in metro- cytes as early as 47 days postinfection and in ga- monts of S. muris in cell cultures up to 4 h after inoculation. Sarcocystis muris (Coccidia, Apicomplexa) is an obligate intracellular and two-host parasite that uses cats (gamogony) and mice (merogony) to complete its life cycle. After the ingestion of an infected mouse, the motile merogonic forms of the parasite, called zoites, invade enterocytes of the cat's small intestine, undergo gamogony, and give rise to sporulated oocysts containing two sporo- cysts, each with four sporozoites. The zoites pos- sess an apical complex with characteristic vesicular organelles: rhoptries, dense granules, and micro- nemes. Rhoptries are involved in the early phase of invasion (Entzeroth 1985). At 30-60 min after zoite invasion, the contents of the dense granules are exocytosed into a newly formed secondary par- asitophorous vacuole (Entzeroth etal. 1985), which appears to be involved in adapting the para- site to the host cell. The role of micronemes has not been elucidated. Within a few hours after in- vading the host cell, the parasite rounds and its micronemes organize underneath the plasmalem- Reprint requests to: J.-F. Dubremetz ma of the parasite, decrease in number, and even- tually disappear 10 h after invasion (Entzeroth et al. 1986). It is not known whether their contents are exocytosed into the secondary parasitophorous vacuole or undergo metabolic change within the parasite itself. In the present paper we report on the isolation of S. muris micronemes and the characterization of their contents. We generated polyclonal anti- bodies against the major polypeptides of micro- nemes, using them to localize these molecules in the motile and developing stages of this organism and to analyze their cross-reactivity with the con- tents of micronemes of other coccidia. Materials and methods Parasites S. muris sporocysts were obtained from infected cats and fed to 2- to 3-month-old mice at a dose of 50 sporocysts/animal. From 3 to 6 months later, the skeletal muscles of six mice were minced and digested with 0.25% (w/v) trypsin (Merck, 2000 IU/mg) in phosphate-buffered saline (PBS; 50 mM phos- phate buffer, pH 7.4; 150 mM NaC1) for 10 min at 37 ~ C. The suspension was filtered through three layers of surgical gauze and centrifuged at 500 g for 10 rain. Two additional washes in PBS under the same conditions led to a purified suspension of ca. 20 x 10 9 S. muris cystozoites. Cell cultures, parasite inoculation Cultures of feline embryo cells (FE, Flow Laboratories) were grown in 25-cm 2 culture vessels in minimal essential medium (MEM) with Earle's salts (Gibco), 0.2 mM HEPES buffer, and 10% fetal calf serum. Approximately 106 cystozoites were sus- pended in 1 ml PBS and added to an equal volume of medium containing 3 x 105 cells that had previously been removed from the culture vessel with EDTA-containing buffer. Cystozoites and cells were mixed and centrifuged at 800 g for 5 min, and the pellet was kept at 37 ~ C for 1 10 h to allow zoite-cell inter- actions. The pellet was then fixed for electron microscopy (EM).
Transcript of Characterization and immunolocalization of the protein ... fileof Sarcocystis muris cystozoites...

Parasitol Res (1989) 75:199-205 Parasitnlngy Research �9 Springer-Verlag 1989
Characterization and immunolocalization of the protein contents of micronemes of Sarcocystis muris cystozoites (Protozoa, Apicomplexa)
U. PohP, J.F. Dubremetz 2, and R. Entzeroth 1 1 Zoologisches Insti tut der Universitfit, Poppelsdorfer Schloss, D-5300 Bonn 1, Federal Republic of Germany 2 INSERM, U42, 369, Rue Jules - Guesde, F-59650 Villeneuve-D'Ascq, France
Abstract. Affinity-purified polyclonal antibodies generated against micronemes obtained by the sub- cellular fractionation of Sarcocystis muris contain two major proteins of 16 and 17 kDa. Antibodies cross-react with microneme antigen of S. tenella and S. cuniculi but not with the S. sp. of wild boar (Sus scrofa). Microneme antigens could be traced by immunoelectron microscopy in metro- cytes as early as 47 days postinfection and in ga- monts of S. muris in cell cultures up to 4 h after inoculation.
Sarcocystis muris (Coccidia, Apicomplexa) is an obligate intracellular and two-host parasite that uses cats (gamogony) and mice (merogony) to complete its life cycle. After the ingestion of an infected mouse, the motile merogonic forms of the parasite, called zoites, invade enterocytes of the cat's small intestine, undergo gamogony, and give rise to sporulated oocysts containing two sporo- cysts, each with four sporozoites. The zoites pos- sess an apical complex with characteristic vesicular organelles: rhoptries, dense granules, and micro- nemes. Rhoptries are involved in the early phase of invasion (Entzeroth 1985). At 30-60 min after zoite invasion, the contents of the dense granules are exocytosed into a newly formed secondary par- asitophorous vacuole (Entzeroth etal. 1985), which appears to be involved in adapting the para- site to the host cell. The role of micronemes has not been elucidated. Within a few hours after in- vading the host cell, the parasite rounds and its micronemes organize underneath the plasmalem-
Reprint requests to: J.-F. Dubremetz
ma of the parasite, decrease in number, and even- tually disappear 10 h after invasion (Entzeroth et al. 1986). It is not known whether their contents are exocytosed into the secondary parasitophorous vacuole or undergo metabolic change within the parasite itself.
In the present paper we report on the isolation of S. muris micronemes and the characterization of their contents. We generated polyclonal anti- bodies against the major polypeptides of micro- nemes, using them to localize these molecules in the motile and developing stages of this organism and to analyze their cross-reactivity with the con- tents of micronemes of other coccidia.
Materials and methods
Parasites
S. muris sporocysts were obtained from infected cats and fed to 2- to 3-month-old mice at a dose of 50 sporocysts/animal. From 3 to 6 months later, the skeletal muscles of six mice were minced and digested with 0.25% (w/v) trypsin (Merck, 2000 IU/mg) in phosphate-buffered saline (PBS; 50 m M phos- phate buffer, pH 7.4; 150 m M NaC1) for 10 min at 37 ~ C. The suspension was filtered through three layers of surgical gauze and centrifuged at 500 g for 10 rain. Two additional washes in PBS under the same conditions led to a purified suspension of ca. 20 x 10 9 S. muris cystozoites.
Cell cultures, parasite inoculation
Cultures of feline embryo cells (FE, Flow Laboratories) were grown in 25-cm 2 culture vessels in minimal essential medium (MEM) with Earle's salts (Gibco), 0.2 m M HEPES buffer, and 10% fetal calf serum. Approximately 106 cystozoites were sus- pended in 1 ml PBS and added to an equal volume of medium containing 3 x 105 cells that had previously been removed from the culture vessel with EDTA-containing buffer. Cystozoites and cells were mixed and centrifuged at 800 g for 5 min, and the pellet was kept at 37 ~ C for 1 10 h to allow zoite-cell inter- actions. The pellet was then fixed for electron microscopy (EM).

200 U. Pohl et al. : Protein contents of S. muris cystozoites
Subcellular fractionation
We used a modification of the S. muris and S. tenella subcellular fractionation procedures previously described by Dubremetz and Dissous (1980). Purified cystozoites were suspended into homogenization medium (250 m M sucrose, 1 m M EDTA, 5 m M triethanotamine-HC1, pH 7.5) at a concentration of l0 s cells/ml. They were disrupted in a French pressure cell (Aminco) operated at 50 kg/cm 2. The homogenate was centri- fuged at 500 g for 10 rain to eliminate unbroken cells. The su- pernatant was collected and subjected to 12000 g centrifugation for 10 min in a Beckman L8-55 ultracentrifuge. Pellicles, mito- chondria, dense granules, and amylopectin granules were sedi- mented by this procedure. The supernatant containing the mi- cronelnes was collected and centrifuged for 20 min at 113 000 g in the SW 28 rotor. A pellet highly enriched with micronemes was obtained, which was frozen in liquid nitrogen and immedi- ately processed for EM or electrophoretic analysis. The con- tents of fractions was determined by EM as previously de- scribed elsewhere (Entzeroth et al. 1986).
Sodium dodeeyl sulfate-polyaerylamide gel electrophoresis
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was carried out according to Laemmli (1970) on 15% potyacrylamide gels. Samples to be analyzed were sus- pended into 2% SDS, 0A M dithiothreitol, and 62.5 m M TRIS- HC1 (pH 6.8) and heated for 3 min at 95 ~ C. After electrophore- sis, the gels were stained with 0.25% Coomassie blue R in 25% isopropanol and 10% acetic acid in water. Molecular- weight markers (Pharmacia) were processed simultaneously for calibration.
Isolation of the major microneme polypeptides and antibody pro- duction in rabbits
A pellet of purified micronemes (about 5 mg protein, corre- sponding to 2 • 10 9 cell equivalents) was resuspended in 2 ml electrophoresis sample buffer, heated for 5 min at 95 ~ C, and electrophoresed according to Laemmli (1970) on a 1.5-mm- thick preparative slab gel of 15% polyacrylamide. The gel was then stained with 1% Coomassie blue R in water for 30 rain. The major 16- and 17-kDa bands were identified, excised, and reinserted into other discontinuous acrylamide gels and elec- troeluted in 2 ml glycerol-containing buffer (Mendel-Hartvig 1982). After' electrophoretic control of the eluted fractions, half of the material was emulsified with 1 vol. Freund's complete adjuvant and injected SC into rabbits. The remaining material was emulsified with incomplete adjuvant 6 weeks later and in- jected by the same route. The rabbits were bled 2 weeks later. Immune sera were assayed by indirect fluorescence on air-dried cystozoites and used for immunoelectron microscopic detection.
Western blotting
Purified cystozoites were separated by SDS-PAGE in the pres- ence of 2-mercaptoethanol (106 zoites in a l-cm-wide slot) and electrophoretically transferred to nitrocellulose at 200 mA for 4 h (Towbin et al. 1978). The nitrocellulose sheet was then satu- rated for 30 min in 5% nonfat milk in buffer [140 m M NaCI, 0.5% v/v Tween-20 in 15 m M TRIS-HC1 (pH 8), TNT]. Strips were then cut and incubated with the different antibody solu- tions diluted 1 : 200 in TNT for 1 h at 37 ~ C. After being washed, the sheet was incubated in peroxidase-conjugated anti-mouse IgG serum (Nordic) diluted in TNT and revealed with diamino- benzidine.
Affinity purification of antibodies on nitrocellulose
Two SDS gels (15% acrylamide) were prepared with 50 x 106 cystozoites each of S. tenella. After migration, proteins were transferred to nitrocellulose sheets according to Towbin et al. (1978). The major bands of 20 and 22 kDa (corresponding to S. tenella microneme contents; Dubremetz and Dissous 1980) were located by staining 0.5-cm-wide longitudinal strips taken from the edges and middle of the nitrocellulose sheets with 0.1% amidoschwarz in 50% methanol. The 20- and 22-kDa bands were then excised from the remainder of the nitrocellu- lose sheets and processed separately for affinity purification of rabbit antibodies. Bands were blocked for 30 rain in TNT buffer containing 2% BSA, incubated with 4 ml 1/i0 dilution of anti-16-kDa S. muris rabbit serum for 2 h under agitation, and washed three times for 10 min in TNT, followed by ehition in 2 ml 0.2 M glycine-HC1 (pH 2.8) and 0.5 M NaC1. The eluate was neutralized with 80 ~tl 2 M TRIS base, supplemented with 1% bovine serum albumin (BSA), and dialyzed against PBS.
[mmunoeleetron microscopy
Infected muscles or cultured cells taken 30-60 min after inocu- lation were fixed in 2.5% paraformaldehyde with 0A % glutar- aldehyde in PBS. Specimens were embedded in Lowicryl K4M according to Roth et al. (1981) and sectioned with glass knives on a Reichert Ultracut microtome. Cryoultramicrotomy was carried out according to Tokuyasu (1980). The fixative was removed after 3-6 h and replaced by 10%, 20%, and 30% su- crose in PBS in successive steps. Small pellets were obtained from the fixed cell suspension after centrifugation, placed on specimen holders, and rapidly immersed in melting nitrogen. Semi- and ultrathin sections were cut with glass knives on a Reichert Ultracut microtome with an FC4 attachment. Grids with Lowicryl sections or cryosections were transferred to drop- lets of PBS for 5 min, followed by 10% newborn calf serum (NCS) in PBS for 10 min, then were briefly rinsed in PBS three times for 3 min. Sections were placed on drops of rabbit serum diluted 1:50 in 10% NCS for 15 min, followed by six rinses in PBS for a total of 15 rain. Ultrathin sections were incubated with gold-conjugated goat anti-rabbit IgG (GAR G5, G20; Janssen Life Science Products) diluted 1:50-1:100 for 20-30 min. After a 30-rain rinse in PBS and a final wash in distilled water, sections were poststained in 4% aqueous uranyl acetate and lead citrate. Ultrathin cryosections were addition- ally covered by a thin layer of methylcellulose containing 3% uranyl acetate. Sections were examined under Zeiss EM 10 and Hitachi H U 12 electron microscopes.
Results
The pellet obtained by 113 000-g max., 20-min cen- trifugation of the 12 000 g max., 10-min superna- tant of S. muris zoite homogenate was highly en- riched with micronemes (Fig. 1). The only contam- inating material was a significant amount of elec- tron-lucent vesicular profiles (microsomes) about 0.1 gm in diameter, which pelleted on top of the microneme pellet. High magnification of the iso- lated micronemes showed that they were mem- brane-bound and had retained their elongated �9 shape (Fig. 2). SDS-PAGE analysis of the fraction demonstrated two major bands at 16 and 17 kDa,
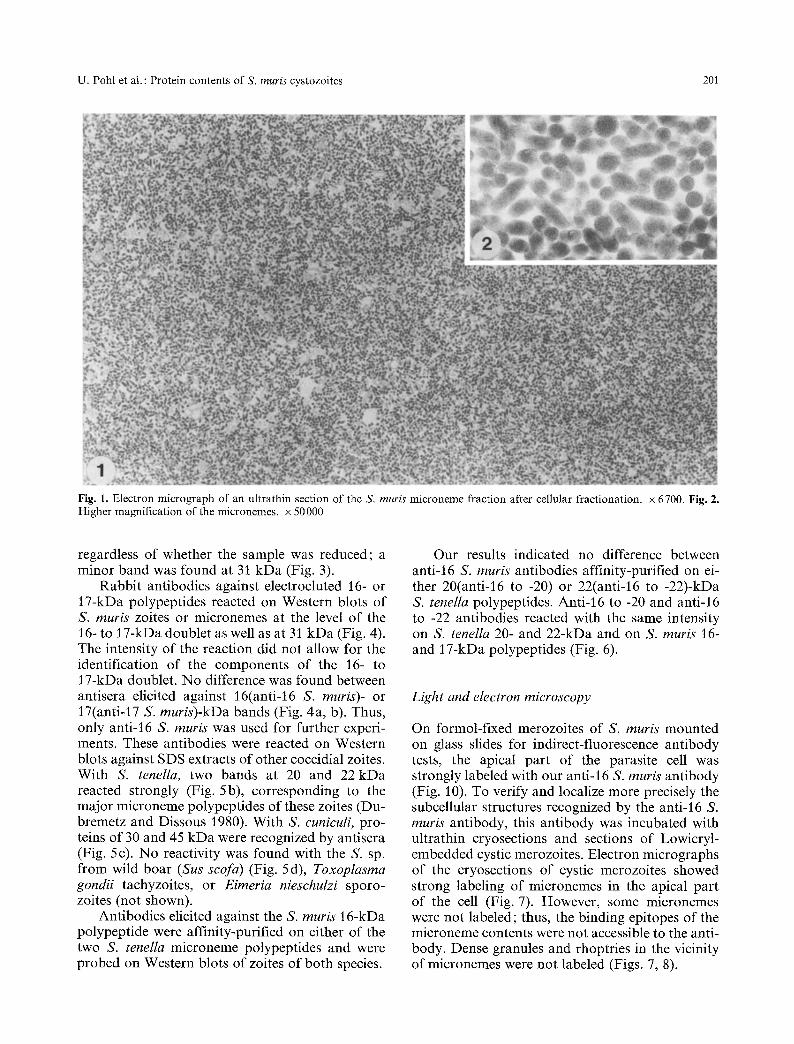
U. Pohl et al. : Protein contents of S. muris cystozoites 201
Fig. 1. Electron micrograph of an ultrathin section of the S. muris microneme fraction after cellular fractionation, x 6700. Fig. 2. Higher magnification of the micronemes, x 50000
regardless of whether the sample was reduced; a minor band was found at 31 kDa (Fig. 3).
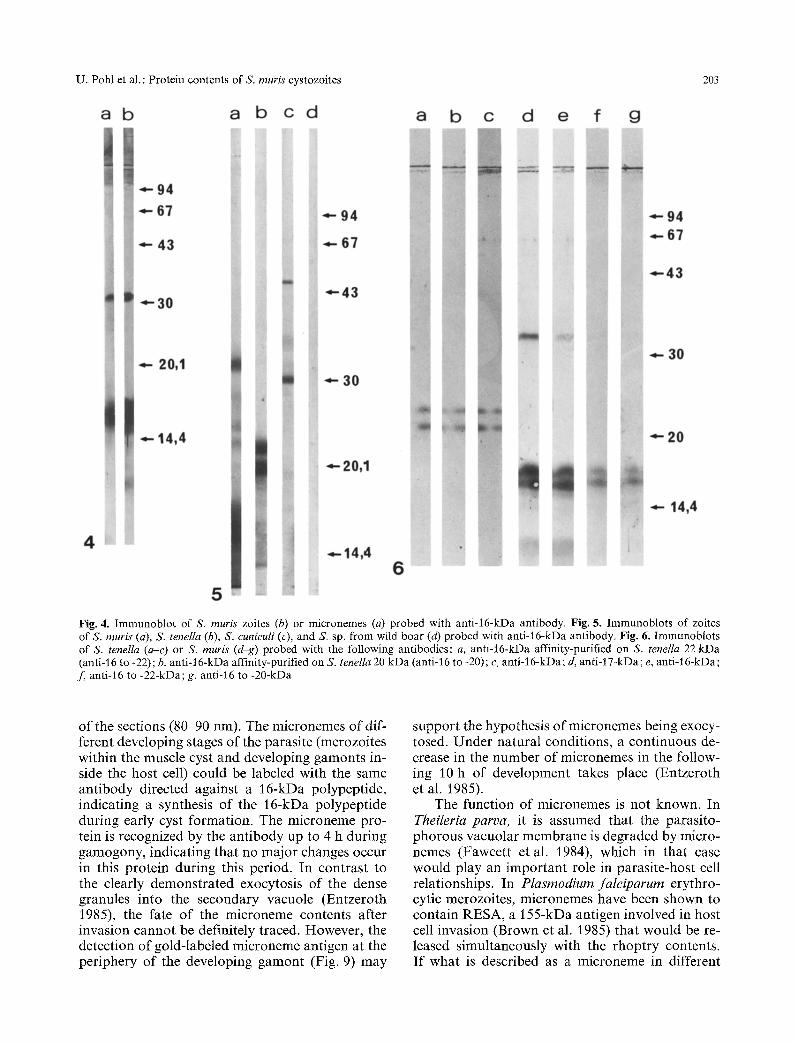
Rabbit antibodies against electroeluted 16- or 17-kDa polypeptides reacted on Western blots of S. muris zoites or micronemes at the level of the 16- to 17-kDa doublet as well as at 31 kDa (Fig. 4). The intensity of the reaction did not allow for the identification of the components of the 16- to 17-kDa doublet. No difference was found between antisera elicited against 16(anti-16 S. muris)- or 17(anti-17 S. muris)-kDa bands (Fig. 4a, b). Thus, only anti-16 S. muris was used for further experi- ments. These antibodies were reacted on Western blots against SDS extracts of other coccidial zoites. With S. tenella, two bands at 20 and 22 kDa reacted strongly (Fig. 5b), corresponding to the major microneme polypeptides of these zoites (Du- bremetz and Dissous 1980). With S. cuniculi, pro- teins of 30 and 45 kDa were recognized by antisera (Fig. 5 c). No reactivity was found with the S. sp. from wild boar (Sus scofa) (Fig. 5d), Toxoplasma gondii tachyzoites, or Eimeria nieschulzi sporo- zoites (not shown).
Antibodies elicited against the S. muris 16-kDa polypeptide were affinity-purified on either of the two S. tenella microneme polypeptides and were probed on Western blots of zoites of both species.
Our results indicated no difference between anti-16 S. muris antibodies affinity-purified on ei- ther 20(anti-16 to -20) or 22(anti-16 to -22)-kDa S. tenella polypeptides. Anti-16 to -20 and anti-16 to -22 antibodies reacted with the same intensity on S. tenella 20- and 22-kDa and on S. muris 16- and 17-kDa polypeptides (Fig. 6).
Light and electron microscopy
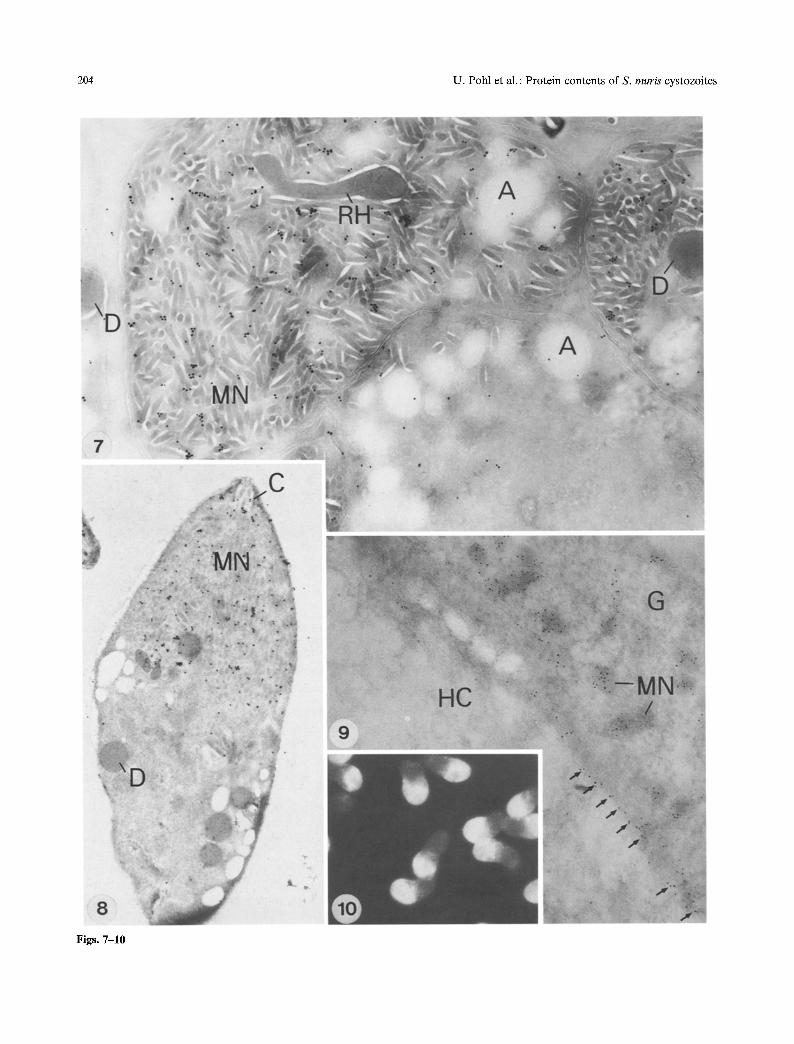
On formol-fixed merozoites of S. muris mounted on glass slides for indirect-fluorescence antibody tests, the apical part of the parasite cell was strongly labeled with our anti-16 S. muris antibody (Fig. 10). To verify and localize more precisely the subcellular structures recognized by the anti-16 S. muris antibody, this antibody was incubated with ultrathin cryosections and sections of Lowicryl- embedded cystic merozoites. Electron micrographs of the cryosections of cystic merozoites showed strong labeling of micronemes in the apical part of the cell (Fig. 7). However, some micronemes were not labeled; thus, the binding epitopes of the microneme contents were not accessible to the anti- body. Dense granules and rhoptries in the vicinity of micronemes were not labeled (Figs. 7, 8).

202 U. Pohl et al. : Protein contents of S. muris cystozoites
Fig. 3. SDS-PAGE of the micronemes of S. muris. Molecular weight markers: phosphorylase B, 94 kDa; albumin, 67 kDa; ovalbumin, 43 kDa; carboanhydrase, 30 kDa; trypsin inhibi- tor, 20.1 kDa; lactalbumin, 14.4 kDa (Calibration Kit, Phar- macia, Uppsala, Sweden)
In developing cystic stages 47 days postinfec- tion, metrocytes were mostly present. After the in- cubation of Lowicryl sections with anti-16 S. muris antibody, specifically labeled micronemes could be detected (Fig. 8) that had their typical shape but were not as abundant as in differentiated mero- zoites.
For the immunodetection of microneme anti- gens in merozoites after invasion and of gamonts during differentiation, cultured feline embryo cells were fixed and embedded in Lowicryl resin 1, 2, 3, and 4 h after parasite-host cell interaction. The micronemes were still present in large numbers in the apical region of the gamonts 45 rain after inva- sion. Dense granules had exocytosed into the sec- ondary parasitophorous vacuole by this time. With the polyclonal anti-16 S. muris antibody, micro- nemes could be labeled up to 4 h after invasion.
During gamont differentiation, they become lo- cated in the peripheral cytoplasm of the parasite and the surface of the gamont (Fig. 9). However, no microneme antigen could be clearly demon- strated outside the parasitic parasitophorous va- cuole or in the host cell cytoplasm.
Discussion
The major components of the S. muris microneme- enriched fraction are two polypeptides of 16 and 17 kDa. This profile is similar to that previously found in S. tenella micronemes, in which two poly- peptides of 20 and 22 kDa were described (Dubre- metz and Dissous 1980). Immunoelectron micro- scopic localization strongly indicates that these proteins are major components of the contents of these organelles. The fate of the minor 31-kDa po- lypeptide is less clear; this molecule might be either a precursor or a complex of the lower-molecular- weight polypeptide (which would explain why anti- 16 S. muris antibodies-cross-react with this mole- cule) or a contaminant of the fraction (proteolytic products that might comigrate at the ] 6- to 17-kDa level and elicit 31-kDa-specific antibodies). Mono- clonal antibodies will be needed to clarify this mat- ter. The major 31-kDa pellicle polypeptide de- scribed in S. muris might correspond with the band observed here (Chbouki and Dubremetz 1985).
The cross-reactivity of anti-S, muris-microneme antibodies with other S. spp. indicates close homo- logies between the organelles in the different spe- cies, especially between S. muris and S. tenella. However, since some S. spp. do not react, this cross-reactivity is limited; it does not extend beyond the generic level.
S. muris and S. tenella micronemes contain a doublet of low-molecular-weight polypeptides. We wanted to know whether the polypeptides within the doublets were distinct from each other. Our experiments showed complete cross-reactivity of antibodies elicited against the 16-kDa S. muris mi- croneme polypeptide and affinity-purified on each component of the 20- to 22-kDa doublet of S. ten- ella, demonstrating that these polypeptides con- tained shared or cross-reacting epitopes.
With our specific polyclonal antibody, the mi- cronemes of S. muris could be clearly labeled and traced during early development. No labeling of rhoptries or dense granules occurred, clearly dem- onstrating that the contents of micronemes was distinct from rhoptries and dense granules. The relatively nonhomogeneous labeling of micro- nemes might be caused by the small size of the micronemes (50-60 nm) relative to the thickness
U. Pohl et al. : Protein contents of S. muris cystozoites 203
Fig. 4. Immunoblot of S. muris zoites (b) or micronemes (a) probed with anti-16-kDa antibody. Fig. ft. Immunoblots of zoites of S. muris (a), S, teneIla (b), S. cuniculi (e), and S. sp. from wild boar (d) probed with antM6-kDa antibody. Fig. 6. Immunoblots of S. tenella (a-c) or S. muris (d-g) probed with the following antibodies: a, anti-16-kDa affinity-purified on S. tenella 22 kDa (anti-16 to -22); b, anti-16-kDa affinity-purified on S. tenella 20 kDa (anti-16 to -20); e, anti-16-kDa; d, anti-17-kDa; e, anti-16-kDa; f , anti-16 to -22-kDa; g, anti-16 to -20-kDa
of the sections (80-90 nm). The micronemes of dif- ferent developing stages of the parasite (merozoites within the muscle cyst and developing gamonts in- side the host cell) could be labeled with the same antibody directed against a 16-kDa polypeptide, indicating a synthesis of the ]6-kDa polypeptide during early cyst formation. The microneme pro- tein is recognized by the antibody up to 4 h during gamogony, indicating that no major changes occur in this protein during this period. In contrast to the clearly demonstrated exocytosis of the dense granules into the secondary vacuole (Entzeroth 1985), the fate of the microneme contents after invasion cannot be definitely traced. However, the detection of gold-labeled microneme antigen at the periphery of the developing gamont (Fig. 9) may
support the hypothesis of micronemes being exocy- tosed. Under natural conditions, a continuous de- crease in the number of micronemes in the follow- ing 10 h of development takes place (Entzeroth et al. 1985).
The function of micronemes is not known. In TheiIeria parva, it is assumed that the parasito- phorous vacuolar membrane is degraded by micro- nemes (Fawcett et al. 1984), which in that case would play an important role in parasite-host cell relationships. In Plasmodium falciparum erythro- cytic merozoites, micronemes have been shown to contain RESA, a 155-kDa antigen involved in host cell invasion (Brown et al. 1985) that would be re- leased simultaneously with the rhoptry contents. If what is described as a microneme in different
204 U. Pohl et al. : Pro te in con ten t s o f S. muris cystozoi tes
Figs. 7-10

U. Pohl et al. : Protein contents of S. muris cystozoites 205
sporozoa cor responds to a funct ional ly identical organelle, one would expect a similar role in each case. Recent da ta on micronemes are still too scarce to clarify this point , and fur ther studies are needed to elucidate their function.
Aknowledgements. The authors are indebted to Mrs. C. Ansel, Mr. Schrauf, and Mrs. Zarbork for their technical assistance. Ultrathin cryosections were kindly prepared by D. Hodick, Bot- any Department, University of Bonn. This study was supported by a Deutsche Forschungsgemeinschaft (DFG) grant to Dr. Entzeroth (En151/1-3) and by INSERM.
References
Brown GV, Culvenor JG, Crewttier PE, Bianco AE, Coppel RL, Saint RB, Stahl MD, Kemp DJ, Anders RF (1985) Localization of the ring-infected erythrocyte surface antigen (RESA) of Plasmodium falciparum in merozoites and ring- infected erythrocytes. J Exp Med/62:774-779
Chbouki N, Dubremetz JF (1985) Structure, isolation and pro- tein composition of the pellicle of Sarcocystis muris cysto- zoites (Protozoa, Coccidia). J Protozool 32:54-58
Dubremetz JF, Dissous C (1980) Characteristic proteins of mi- cronemes and dense granules fromgarcocystis tenella zoites (Protozoa, Coccidia). Mol Biochem Parasitol 1 : 279-289
Entzeroth R (1985) Invasion and early development of Sarco- cystis muris (Apicomplexa, Sarcocystidae) in tissue cultures. J Protozool 32:446-453
Entzeroth R, Chobotar B, Scholtyseck E (1985) Electron micro- scope study of gamogony of Sarcocystis muris (Protozoa,
Apicomplexa) in the small intestine of cats (Felis catus). Protistologica 26: 399408
Entzeroth R, Dubremetz JF, Hodick D, Ferreira E (1986) Im- munoelectron microscopic demonstration of the exocytosis of dense granule contents into the secondary parasitophor- ous vacuole of Sarcocystis muris (Protozoa, Apicomplexa). Eur J Cell Biol 41 : 182-188
Fawcett D, Musake A, Voigt W (1984) Interaction of sporo- zoites of Theileria parva with bovine lymphocytes in vitro : I. Early events after invasion. Tissue Cell 16:873-884
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685
Mendel-Hartvig I (1982) A simple and rapid method for the isolation of peptides from sodium dodecyl sulfate-contain- ing polyacrylamide gels. Anal Biochem 121 : 215-217
Rose ME (1982) Host immune responses. In: Long PL (ed) The biology of coccidia. University Park Press, Baltimore, pp 329-373
Roth JM, Bendeijan M, Carlemalm E, Villiger W, Coravito M (1981) Enhancement of structural preservation and im- munocytochemical staining in low temperature embedded pancreatic tissue. J Histochem Cytochem 29 : 663-671
Ruiz A, Frenkel JK (1976) Recognition of cyclic transmission of Sarcocystis muris by cats. J Infect Dis 133:409-418
Tokuyasu KT (1980) Immunochemistry on ultrathin frozen sec- tions. Histochem J 12:381403
Towbin H, Staehelin T, Gordon J (1978) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellu- lose sheets: procedure and some applications. Proc Natl Acad Sci USA 76:4350-4354
Accepted June 25, 1988
Fig. 7. Ultrathin cryosection of merozoites inside a differentiated muscle cyst 100 days after infection. Note micronemes (MN) after incubation with polyclonal antiserum against a 16-kDa microneme protein and labelling with GAR G 20. A, amylopectin; D dense granules; MN, micronemes; RH, rhoptries, x 31000. Fig. 8. Tangential section through the apical region of a zoite with conoid (C) micronemes (MN) and dense granules (D). Lowicryl K4M, anti-16-kDa rabbit serum, GAR G20. x28000. Fig. 9. Section through the periphery of a gamont (G) 4 h after invasion of a cultured feline embryo cell (HC). Note the labeled micronemes (MN) in the peripheral cytoplasm of the gamont and the labeled antigen at the parasite-host cell interface (arrows). Lowicryl, affinity-purified polyclonal antibody, GAR G 5. x 43200. Fig. 10. Merozoites after incubation with anti-16-kDa rabbit serum and labelling with anti-rabbit FITC (Light micrograph), x 1 100


















