
CHARACTERISATION OF THE DEVELOPMENT … OF THE DEVELOPMENT AND HORMONAL REGULATION OF THE OVARIAN...
34
CHARACTERISATION OF THE DEVELOPMENT AND HORMONAL REGULATION OF THE OVARIAN LYMPHATIC VASCULATURE Hannah Mary Brown Robinson Institute School of Paediatrics and Reproductive Health Research Centre for Reproductive Health Discipline of Obstetrics and Gynaecology University of Adelaide, Adelaide Australia Thesis submitted to the University of Adelaide in fulfillment of the requirements for admission to the degree Doctor of Philosophy April 2009
Transcript of CHARACTERISATION OF THE DEVELOPMENT … OF THE DEVELOPMENT AND HORMONAL REGULATION OF THE OVARIAN...

CHARACTERISATION OF THE DEVELOPMENT AND HORMONAL REGULATION OF THE OVARIAN
LYMPHATIC VASCULATURE
Hannah Mary Brown
Robinson Institute School of Paediatrics and Reproductive Health
Research Centre for Reproductive Health Discipline of Obstetrics and Gynaecology
University of Adelaide, Adelaide Australia
Thesis submitted to the University of Adelaide in fulfillment of the requirements for admission to the degree Doctor of Philosophy
April 2009

CHAPTER 4 OVARIAN LYMPHANGIOGENESIS IS ASSOCIATED WITH FSH-
MEDIATED FOLLICLE GROWTH

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
54 BROWN HM
4.1 INTRODUCTION The process of primary lymphangiogenesis (or embryonic lymphatic development) has been well described and involves a number of differentiation and migration events and the tight regulation of a number of genes including Prox1, Vegfc and Vegfr3. In the mouse, at around E 10.5, the lymphatic vasculature is developing centrifugally by budding from central veins. These lymph sacs subsequently give rise to lymphatic vessel plexuses and networks. Between E 11.5 and 14.5, the lymphatic vasculature becomes completely independent of the blood circulation [102]. By E 15, the lymphatic vasculature is present within distal organs, including the heart, diaphragm, lungs, gut and skin and expansion continues throughout embryonic development and early postnatal life [109, 110]. The mechanisms controlling the generation and expansion of the lymphatic vasculature within organs during early development however remain elusive, and virtually unexplored.
Secondary lymphangiogenesis or adult lymphangiogenesis is the process by which lymphatic vessels are remodelled, and is thought to be associated with pathophysiological events, much like adult angiogenesis [119]. Lymphatic vascular remodelling and growth is thought to be associated with wound healing and tissue regeneration, tumour growth and inflammation. Most recently, the lymphatic vasculature has been described in the cyclically remodelling endometrium; however whether the vessels are remodelled in the uterus is not determined. New lymphatic vessels are believed to sprout from existing vessels, much like the angiogenic process [121], which is well described in most systems. There are however, suggestions that bone-marrow derived progenitor cells may also contribute to adult lymphangiogenesis. Recent evidence also suggests that mouse corneal macrophages can trans-differentiate into lymphatic endothelial cells (LECs) [183], and host LEC progenitor cells have been reported to be present in inflamed kidney transplants [123], however this area remains controversial and is limited by the ability to identify LECs from other cells expressing LEC related genes. A number of animal post-surgical models of adult lymphangiogenesis have been described, however the mechanism by which lymphangiogenesis is achieved is poorly understood, and is likely to include multiple factors including inflammation, physical changes to interstitial fluid flow, MMP activity (ECM remodelling) and Vegfc/Vegfr3 signalling [115-119]. Vegfc, Vegfd, hepatocyte growth factor (Hgf) and platelet derived

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
55 BROWN HM
growth factor (Pdgf)-BB have been implicated in the promotion of tumour lymphangiogenesis and lymph node/systemic metastasis [119]. It also remains unclear whether the regulators of developmental lymphangiogenesis will have the same relevance in adult lymphangiogenesis or lymphatic regeneration, particularly considering that many of these factors play multiple roles in early development of both blood and lymphatic vessels before becoming specific to one type of endothelial cell in adulthood [104, 119].
Uniquely, the female reproductive organs undergo adult blood vascular remodelling as part of the necessary remodelling events associated with the reproductive cycle. Follicular angiogenesis coincides with folliculogenesis and is minimal until formation of the follicular antrum, which occurs under the tight control of FSH [7, 21]. FSH is a pituitary produced and secreted hormone which acts via the gonad specific, G-protein coupled, FSH-receptor found on granulosa cells. FSH promotes follicle maturation by stimulation of granulosa cell proliferation and synthesis of the androgen aromatising enzyme Cyp19a1 (P450 Aromatase), and protects follicles from death via atresia [7, 21]. During this same period of development, a wreath-like structure of blood vessels forms around the follicle, consisting of two concentric networks of vessels within the theca interna and externa, and is closely linked with growth and development of the follicle. The blood vasculature is not directly responsive to FSH; blood vascular endothelial cells do not express the FSH-receptor, instead FSH induces angiogenic promoting molecules (Vegfa) within the granulosa cells, which acts in a paracrine manner to promote vascular remodelling within the thecal layers [29, 184, 185]. The blood vasculature around the rapidly growing follicle sequesters an increasing supply of gonadotrophins, growth factors, oxygen, lipids and steroid precursors and other requirements, as well as removing waste products. The vessels increase in size and number, but never penetrate the basement membrane, keeping the granulosa layers and oocyte completely avascular until just prior to ovulation. The necessity of blood vascular remodelling and expansion process has been well characterised in vivo by the inhibition of both Vegfa and Vegfr2, both of which prevent follicle maturation, demonstrating that vascular remodelling is necessary for gonadotrophin supported follicle growth [29, 73]. It is currently unclear whether the lymphatic vasculature undergoes remodelling (lymphangiogenesis) in association with FSH-dependent follicle growth.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
56 BROWN HM
While the ovarian lymphatic vasculature remains virtually unexplored, we recently identified an ovarian lymphatic defect in the Adamts1-/- mouse. Adamts1 (a disintegrin and metalloproteinase with thrombospondin type motifs 1) is an extracellular matrix (ECM) protease responsible for the degradation (cleavage) of proteoglycans in a wide range of tissues under a wide range of normal and pathophysiological circumstances including ovarian remodelling [145, 186]. The gene for Adamts1 is FSH, LH and progesterone responsive [143, 144]. Female mice with a null mutation in Adamts1 have severe fertility defects which have been well characterised. Ovarian defects include dysgenic follicle growth and abnormal follicle loss, aberrant cumulus matrix formation and ovulatory disruption, all contributing to a severely sub-fertile phenotype [142]. Most interestingly, these female mice also have an ovarian lymphatic defect, whereby at PND 10 and d 21, prior to puberty, they had developed no ovarian lymphatic network. While some lymphatic vessels are present in the adult Adamts1-/- ovary, it is currently unclear when and if the Adamts1 nulls develop a normal ovarian lymphatic network, and if it is ever comparable to that of their Adamts1+/- counterparts [142].
In Chapter 3, I demonstrated that the ovarian lymphatic vasculature developed postnatally, in association with the induction of ovarian Aromatase expression and the first wave of estrogenic-follicle growth. Given the propensity and necessity for blood vascular remodelling during FSH-dependent follicle growth, in this chapter I hypothesised that much like the blood vasculature, the lymphatic vasculature is remodelled in association with follicle growth, in response to hormones that regulate folliculogenesis. To examine this hypothesis, I used FSH-analogue, eCG and examined the associated temporal and spatial changes to the Lyve1-positive lymphatic vasculature. Additionally, I used the same gonadotrophin manipulation to examine whether it was possible to influence the ovarian lymphatic vasculature during its establishment window (from PND 8.5) in postnatal development. Finally, given our previous data reporting the lymphatic vessel phenotype in the Adamts1-/- ovary, I exploited this model to examine Adamts1-dependent ovarian lymphangiogenesis and whether gonadotrophic promotion of follicle growth could restore the Adamts1-/- Lyve1-positive ovarian lymphatic vasculature.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
57 BROWN HM
4.2 MATERIALS AND METHODS
4.2.1 Animal treatment and tissue collection
eCG treatment in prepubertal mice: Adamts1+/- and Adamts1-/- female mice were i.p. injected at 1400 h on d 21 with 5IU eCG. 44 h later, mice were sacrificed via cervical dislocation and dissected ovaries were either snap frozen whole in liquid nitrogen for mRNA analysis or collected into 4% paraformaldehyde fixative and paraffin embedded for immunohistochemistry. Ovaries were also collected from control mice which had received no treatment at d 23 of age. A total of five mice were used for each treatment and genotype (Figure 4.1 A).
eCG treatment in postnatal mice: Mice were i.p. injected at 1400 h on PND 8.5 of age with eCG (500IU/kg). 44 h later, mice were sacrificed via decapitation and ovaries were either snap frozen whole in liquid nitrogen for mRNA analysis or collected and fixed in 4% paraformaldehyde and paraffin embedded for immunohistochemistry. Uteri were weighed as a bioassay for FSH-mediated estradiol production. A total of 6-7 mice were collected for each treatment (Figure 4.1 B).
Additionally, C57BL/6 mouse ovaries were collected unstimulated (d 21), following 44 h eCG stimulation (d 23), following eCG + 12 h hCG stimulation (d 23), or naturally cycling at two months of age (d 60). Ovaries were collected and fixed in 4% paraformaldehyde and paraffin embedded for immunohistochemical localisation of blood and lymphatic vessels (Figure 4.1 C).
All mice were maintained in 12 h/ 12 h light dark and given water and rodent chow ad libitum. All experiments were approved by the University of Adelaide Animal Ethics Committee and were conducted in accordance with the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes.
4.2.2 Immunohistochemistry
Immunohistochemical localisation of Lyve1 and Von Willebrand Factor were undertaken on ovarian tissue sections as per Section 2.2.5 and 3.2.2. Both the Lyve1 and Von Willebrand Factor antibodies (Millipore) required Proteinase K antigen retrieval.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
58 BROWN HM
Figure 4.1 Schematic representation of sample collection schedule/experimental design
A). Adamts1+/- and Adamts1-/- mouse ovaries were collected on d 23 of age following either no treatment or 44 h eCG treatment. Ovaries were collected into paraformaldehyde and paraffin embedded for histological analysis, or snap frozen in liquid nitrogen for subsequent mRNA analysis (n=five mice/treatment/genotype). B). C57BL/6 mouse ovaries were collected on PND 10.5 following either no treatment, or 44 h eCG stimulation. Ovaries were either snap frozen whole (independently) in liquid nitrogen for mRNA analysis or fixed in paraformaldehyde and paraffin embedded for immunolocalisation of lymphatic vessels (n=6-7 mice/treatment). C). C57BL/6 female mice were collected unstimulated (d 23), following 44 h eCG stimulation (d 23), following eCG + 12 h hCG stimulation (d 23), or at two months of age (d 60) and fixed in 4% paraformaldehyde and paraffin-embedded for immunohistochemical localisation of blood and lymphatic vessels.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
59 BROWN HM

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
60 BROWN HM
4.2.3 Image capture and morphometric analysis
Following Lyve1 immunohistochemistry on serial ovary sections from every ovary, the microscope slides were scanned and images of whole ovary sections were captured at high resolution using NanoZoomer Digital Pathology slide scanner (Hamamatsu Photonics K.K., Japan). Images of ovarian sections, taken
at 35�m increments through the entire ovary for each experiment and treatment, were then captured at
10x magnification in preparation for image analysis using NDP View (Digital Slide Viewer, Hamamatsu Photonics K.K., Japan). Images of each entire section were imported into Analysis™ software (Olympus, NSW Australia) where morphometric analysis including ovarian area, follicle area (growing follicles of stage 4 and greater) [187] and lymphatic vessel number and cross sectional area (size) were measured.
From this analysis, lymphatic vessels were grouped into four size categories based on vessel cross-
sectional area to discriminate between the microvasculature (small; <280 �m2), medium (281-1870
�m2), large (1871-9350 �m2) and extra large collecting (>9350 �m2) vessel populations (Table 4.1). To
classify the spatial distribution of the lymphatic vessels within the ovary, vessels were assessed in three
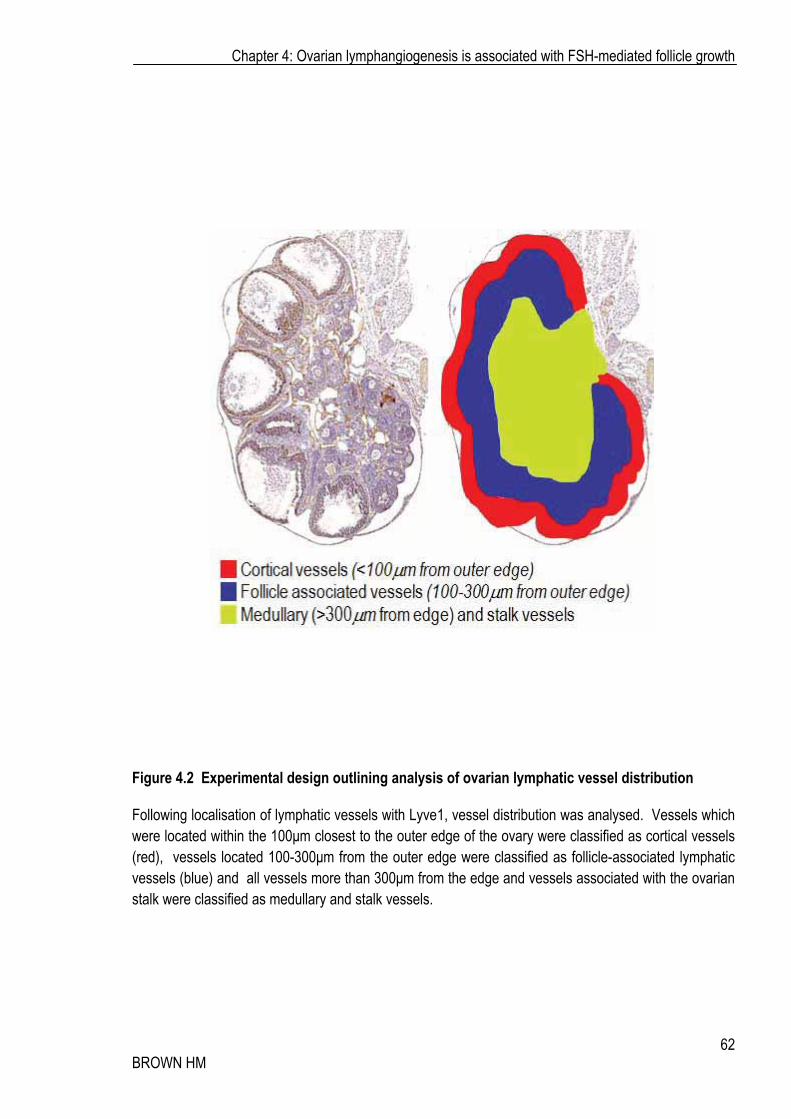
regions. Cortical vessels were those within 100�m of the outer surface epithelium of the ovary, follicle
associated vessels were those found between 100-300�m from the outer edge of the ovary where most
growing follicles reside, and the medullary and/or stalk vessels which were classified as greater than
300�m from the edge, and or associated with the ovarian stalk (Figure 4.2).
4.2.4 Real Time RT-PCR
Real Time RT PCR was performed as per Chapter 2.2.7 with primers listed in Table 2.1. Genes measured were Lyve1, Vegfc, Vegfd, Vegfr3 and Aromatase. Data are presented as 2-��CT (mean ± SEM) normalised to the same calibrator sample to facilitate comparison of fold change with treatment as well as relative expression between Adamts1 genotype and treatment.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
61 BROWN HM
Table 4.1 Lymphatic vessel size classifications
Vessel size characterisation Size in microns (�m2)
Small <280 �m2
Medium 281-1870 �m2
Large 1871-9350 �m2
Extra large >9350 �m2
Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
62 BROWN HM
Figure 4.2 Experimental design outlining analysis of ovarian lymphatic vessel distribution
Following localisation of lymphatic vessels with Lyve1, vessel distribution was analysed. Vessels which were located within the 100�m closest to the outer edge of the ovary were classified as cortical vessels (red), vessels located 100-300�m from the outer edge were classified as follicle-associated lymphatic vessels (blue) and all vessels more than 300�m from the edge and vessels associated with the ovarian stalk were classified as medullary and stalk vessels.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
63 BROWN HM
4.2.5 Statistical analysis
Data was analysed using One Way ANOVA with Tukey Post-Hoc and statistical significance considered if P�0.05 (SPSS 11.5; SPSS Inc., Chicago, IL). Alternatively, when comparing the difference between only two groups, Student’s t-test was used and statistical significance considered if P�0.05 (SPSS 11.5; SPSS Inc., Chicago, IL).
4.3 RESULTS To assess whether FSH-mediated folliculogenesis was associated with changes to the ovarian lymphatic vasculature, pre-pubertal and PND 8.5 mice were stimulated with eCG and lymphatic vessels analysed. The prepubertal paradigm provided a model to examine secondary lymphangiogenesis, or remodelling of an existing lymphatic vascular network, whilst the postnatal model allowed examination of the effects of modulating the lymphatic vasculature during its establishment window. Furthermore, the Adamts1-/- model, lacking ovarian lymphatic vasculature, afforded the opportunity to examine whether ovarian lymphatic establishment could be induced upon stimulation of follicle growth in response to eCG.
4.3.1 Ovarian growing follicle area is greater in the presence of Adamts1
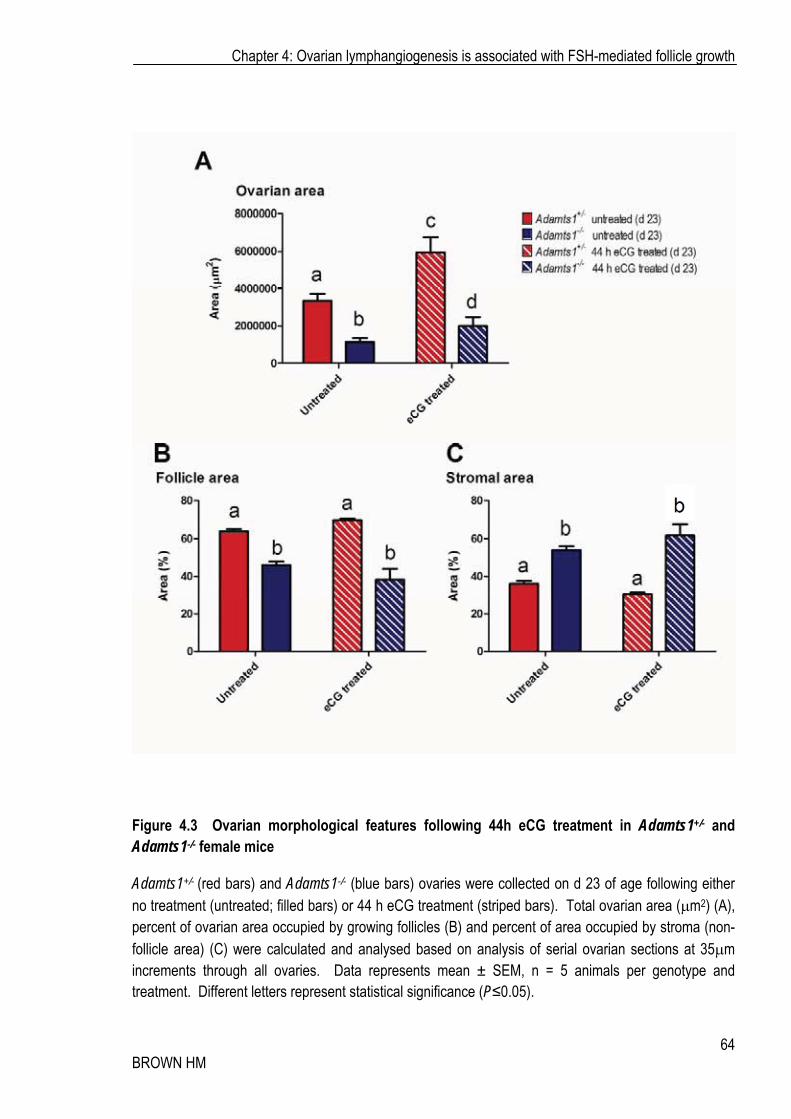
Ovarian area significantly increased following 44 h eCG administration, as expected, and was significantly lower in the absence of Adamts1 (Adamts1-/-) (Figure 4.3A). Whilst eCG significantly increased total ovarian area in Adamts1-/- mice, it remained significantly lower than that of their Adamts1 heterozygous counterparts. In Adamts1+/- ovaries, the percentage of growing follicle area (of total ovarian area) was significantly higher and inversely, stromal/thecal area was lower, consistent with previous findings regarding follicle growth kinetics in the Adamts1 nulls [142] (Figure 4.3 B, C). This increase in cross-sectional area and its potential influence on number of lymphatic vessels observed was accommodated in subsequent analysis by normalising lymphatic vessel area to total ovarian area or that of the growing follicles or stroma.
Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
64 BROWN HM
Figure 4.3 Ovarian morphological features following 44h eCG treatment in Adamts1+/- and Adamts1-/- female mice
Adamts1+/- (red bars) and Adamts1-/- (blue bars) ovaries were collected on d 23 of age following either no treatment (untreated; filled bars) or 44 h eCG treatment (striped bars). Total ovarian area (�m2) (A), percent of ovarian area occupied by growing follicles (B) and percent of area occupied by stroma (non-follicle area) (C) were calculated and analysed based on analysis of serial ovarian sections at 35�m increments through all ovaries. Data represents mean ± SEM, n = 5 animals per genotype and treatment. Different letters represent statistical significance (P�0.05).
Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
65 BROWN HM
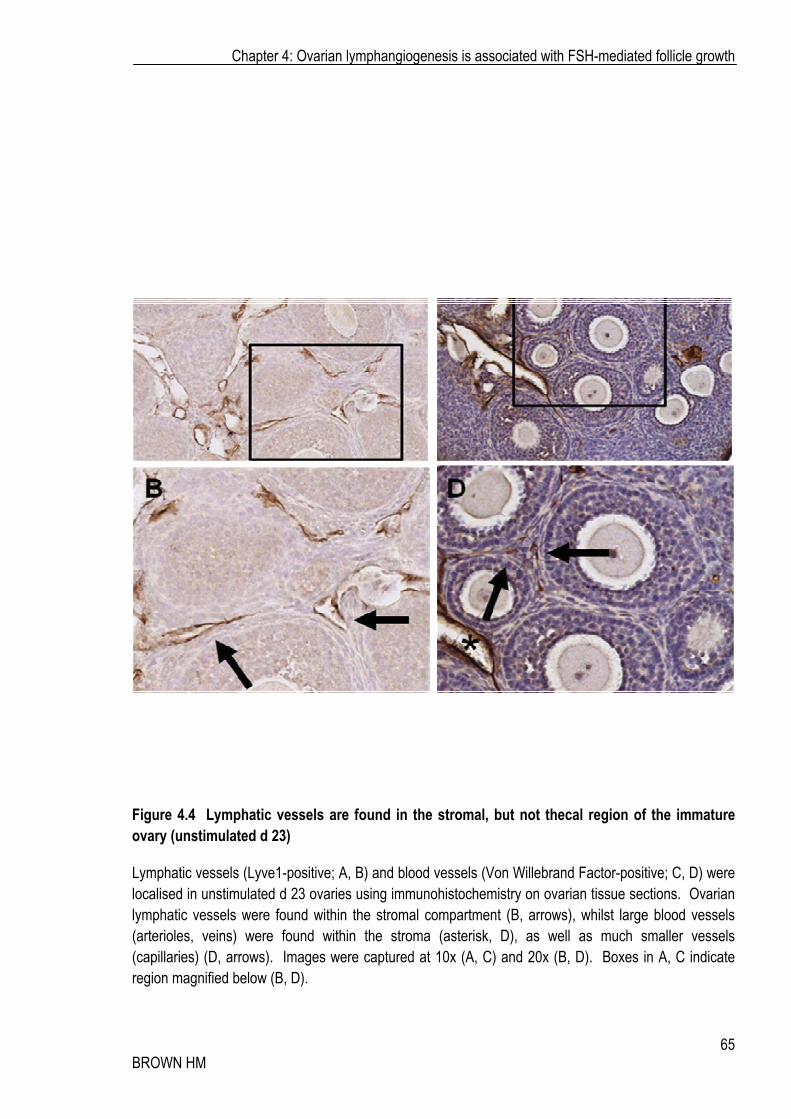
Figure 4.4 Lymphatic vessels are found in the stromal, but not thecal region of the immature ovary (unstimulated d 23)
Lymphatic vessels (Lyve1-positive; A, B) and blood vessels (Von Willebrand Factor-positive; C, D) were localised in unstimulated d 23 ovaries using immunohistochemistry on ovarian tissue sections. Ovarian lymphatic vessels were found within the stromal compartment (B, arrows), whilst large blood vessels (arterioles, veins) were found within the stroma (asterisk, D), as well as much smaller vessels (capillaries) (D, arrows). Images were captured at 10x (A, C) and 20x (B, D). Boxes in A, C indicate region magnified below (B, D).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
66 BROWN HM
4.3.2 The lymphatic vasculature is found within the stroma, but not theca of immature and eCG-stimulated ovaries
In immature ovaries (d 21), the ovarian medulla contained both large blood and lymphatic vessels. Small follicles undergoing the pre-antral to antral transition contained blood vascular capillaries within the theca identified by Von Willebrand Factor staining (Figure 4.4 C-D, arrows) were also associated with larger arteriole-like vessels (asterisk). Lymphatic capillaries were not found in the developing theca. Large Lyve1-positive lymphatic vessels (much larger than blood capillaries) were found in the stroma associated with growing follicles (Figure 4.4 A-B; arrows).
The distribution of lymphatic vessels following eCG stimulation was very similar to that seen in immature d 21 ovaries. Following eCG stimulation, the ovarian medulla contained both large blood and lymphatic vessels. Growing follicles had developed a rich supply of blood capillaries within the theca (Figure 4.5 C-D, arrows) and were also associated with larger arteriole-like vessels (asterisk). Again, lymphatic capillaries were not found in the expanding theca; however large antral follicles were closely associated with lymphatic vessels, much larger in size than the blood capillaries (Figure 4.5 A-C; arrows).
4.3.3 eCG promotes ovarian lymphatic vessel growth and expansion
Mice lacking Adamts1 had an average of 8.5 ovarian lymphatic vessels per ovary, compared to their Adamts1 heterozygous counterparts, who averaged 140 vessels per ovary, consistent with our previous qualitative reports [142]. Following eCG stimulation, in Adamts1+/- ovaries, vessel number was unchanged (Figure 4.6 A), however the vessels increased in size demonstrated by a significant reduction in the number of small vessels and a significant increase in the number of large vessels (Figure 4.6 B), as well as an increase in the presence of extra-large vessels, and a resulting increase in mean ovarian lymphatic vessel size (Figure 4.7).
In Adamts1-/- ovaries, eCG treatment stimulated primary establishment of the lymphatic vascular network within the ovary. The number of lymphatic vessels per ovary increased 6.6-fold in eCG treated Adamts1-/- mice (Figure 4.6 A). Vessel size also increased, reflected in this case, by significant increases in the number of small and medium vessels within the ovary (Figure 4.6 C). This resulted in a significant increase in the average vessel size following eCG treatment in the absence of Adamts1 (Figure 4.7). Importantly, the lymphatic vessels in Adamts1-/- ovaries remained fewer in number (Figure

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
67 BROWN HM
Figure 4.5 Lymphatic vessels are found in the stroma close to the theca, but not within the thecal region of pre-ovulatory follicles
Lymphatic vessels (Lyve1-positive; A, B) and blood vessels (Von Willebrand Factor-positive; C, D) were localised in ovaries following 44 h eCG stimulation using immunohistochemistry on ovarian tissue sections. Ovarian lymphatic vessels were found associated with growing follicles (B, arrow) within the stromal compartment, whilst large blood vessels (arterioles, veins) were found within the stroma (asterisk, D), and smaller vessels (capillaries) were in the theca layer (D, arrows). Images were captured at 10x (A, C) and 20x (B, D). Boxes in A, C indicate region magnified below (B, D).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
68 BROWN HM
Figure 4.6 eCG increases ovarian lymphatic vessel size and number
Ovarian lymphatic vessels were counted and area measured in Adamts1+/- (red bars) and Adamts1-/- (blue bars) ovaries following either no treatment (untreated; filled bars) or 44 h eCG treatment (striped bars). Total ovarian lymphatic vessel number (A) and vessel size (cross sectional area) was analysed (B, C). Lymphatic vessel size was classified based on cross sectional areas to discriminate the microvasculature (small; <280 �m2), medium (281-1870 �m2), large (1871-9350 �m2) and extra large collecting (>9350 �m2) vessel populations (Table 4.1). Morphometric analysis was based on analysis of serial ovarian sections at 35�m increments through all ovaries. Data represents mean ± SEM, n = 5 animals per genotype and treatment. Different letters (or *) represent statistical significance (P�0.05).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
69 BROWN HM
Figure 4.7 eCG increases mean ovarian lymphatic vessel size
Mean ovarian lymphatic vessel size (or cross sectional area) was calculated in Adamts1+/- (red bars) and Adamts1-/- (blue bars) ovaries following either no treatment (untreated; filled bars) or 44 h eCG treatment (striped bars). Morphometric analysis was based on analysis of serial ovarian sections at 35�m increments through all ovaries. Data represents mean ± SEM, n = 5 animals per genotype and treatment. Different letters represent statistical significance (P�0.05).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
70 BROWN HM
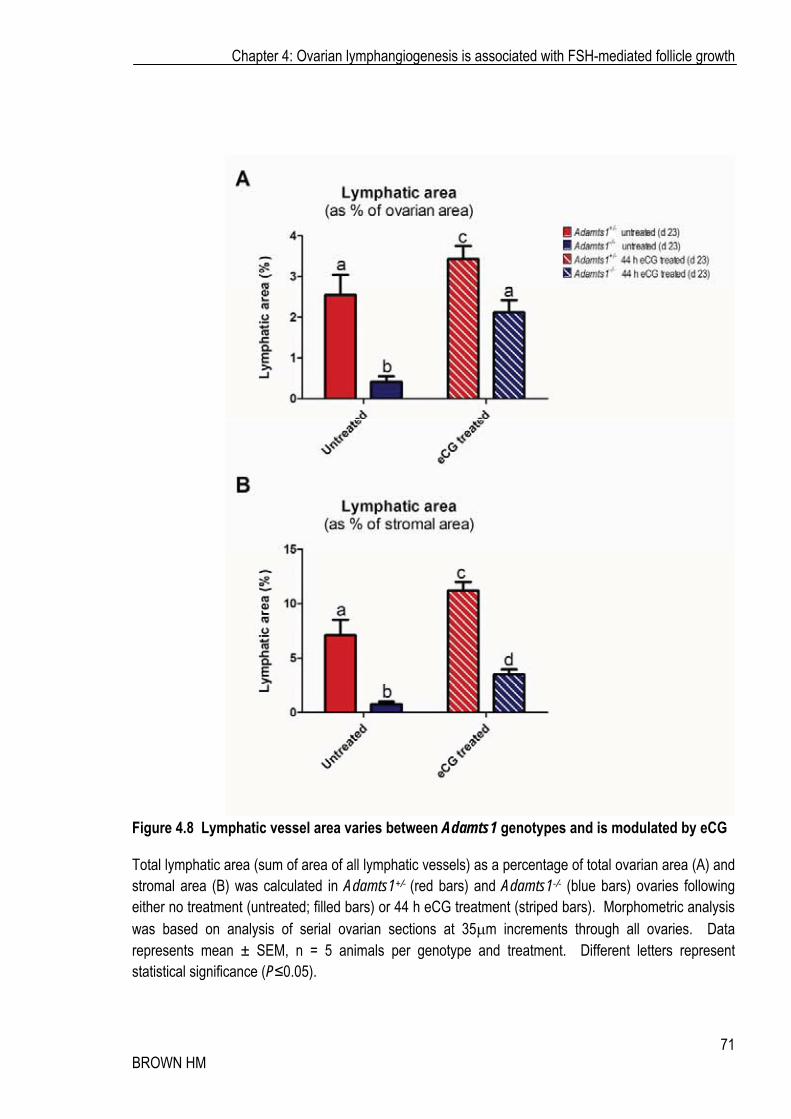
4.6 A) and smaller (Figure 4.6 C, 4.7), than their heterozygous counterparts, even after the effects of exogenous eCG treatment. Through increasing either vessel size in Adamts1+/- mice or number and size in Adamts1-/- ovaries, eCG treatment significantly increased the lymphatic vessel area: total ovarian area and lymphatic vessel area: stromal area ratios (Figure 4.8 A, B). This demonstrates that eCG was able to increase Adamts1-independent lymphangiogenesis (in the Adamts1-/-), but Adamts1-dependent mechanisms also remain necessary, as indicated by the reduced number of vessels in the Adamts1-/-, even following eCG treatment.
4.3.4 eCG mediates changes in lymphatic vessel spatial distribution
Lymphatic vessel location was categorised as either cortical, follicle associated or medullary and/or stalk region (see Figure 4.2). Lymphatic vessels were present in all three regions independent of treatment or genotype, albeit sparsely in Adamts1 nulls (Figure 4.9 A-B). In the presence of Adamts1, eCG did not alter the distribution of lymphatic vessels in any of the regions, consistent with the lack of change in total vessel number. Examining the spatial vessel distribution revealed that the increase in lymphatic vessel size in the Adamts1+/- ovaries was due to a significant two-fold increase in the medullary/stalk vessel size and a trend towards an increase in cortical vessel size (Figure 4.9 C). In Adamts1-/- ovaries eCG stimulation induced an increase in vessel number in each of the three zones (Figure 4.9 B) and vessel size was also increased in each region, with a significant three-fold increase in vessels of the cortical region (Figure 4.9 D). In both Adamts1 genotypes, the lymphatic vessels found within the medullary/stalk region were the largest (Figure 4.9 C, D), however, even after this exogenous eCG treatment the largest population of vessels remained approximately 50% smaller in Adamts1-/- than Adamts1+/- ovaries (Figure 4.9 C, D).
4.3.5 eCG mediated ovarian expression of lymphangiogenic genes in the presence and absence of Adamts1
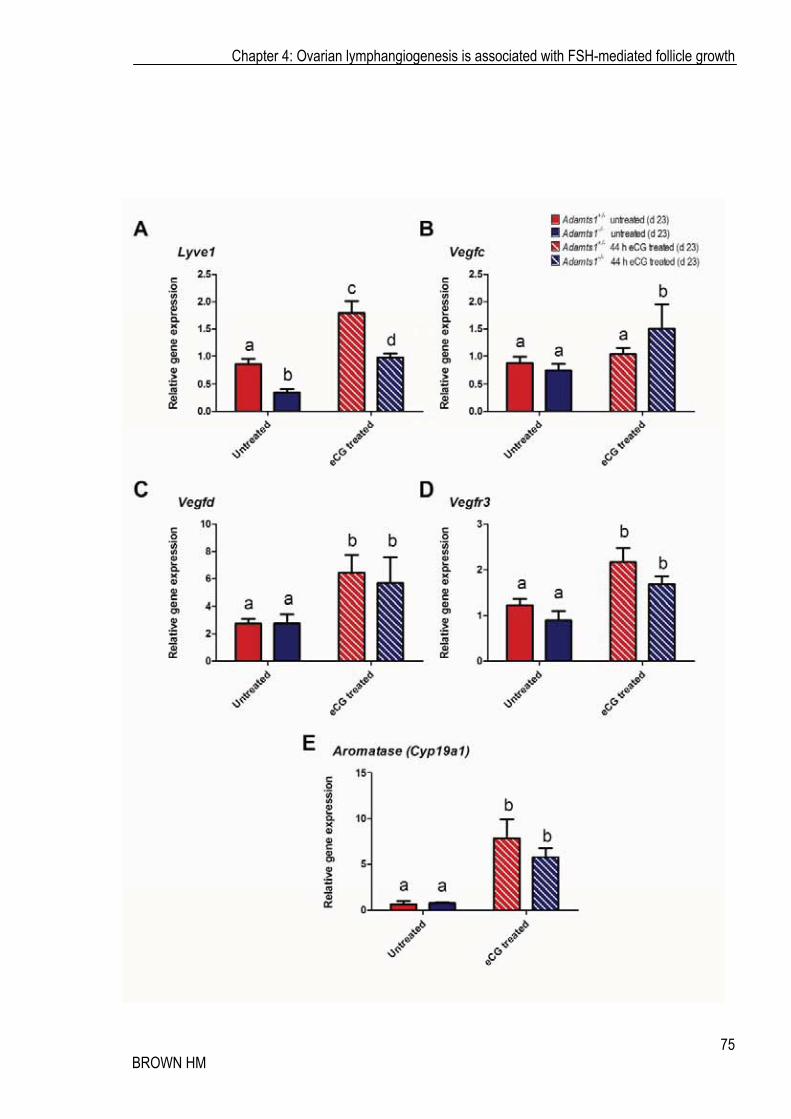
Gene expression in the whole ovary analysed by real time RT-PCR revealed that Lyve1 gene expression closely resembled the pattern of lymphatic vessel number/size. Lyve1 expression significantly increased in whole ovary, in both heterozygous and Adamts1-/- ovaries following eCG treatment. Likewise, Lyve1 expression remained significantly lower in the absence of Adamts1 (Figure
Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
71 BROWN HM
Figure 4.8 Lymphatic vessel area varies between Adamts1 genotypes and is modulated by eCG
Total lymphatic area (sum of area of all lymphatic vessels) as a percentage of total ovarian area (A) and stromal area (B) was calculated in Adamts1+/- (red bars) and Adamts1-/- (blue bars) ovaries following either no treatment (untreated; filled bars) or 44 h eCG treatment (striped bars). Morphometric analysis was based on analysis of serial ovarian sections at 35�m increments through all ovaries. Data represents mean ± SEM, n = 5 animals per genotype and treatment. Different letters represent statistical significance (P�0.05).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
72 BROWN HM
Figure 4.9 eCG increases lymphatic vessel size in specific regions of the ovary
Ovarian lymphatic vessels were characterised by number, size and location (distribution) Adamts1+/-
(red bars) and Adamts1-/- (blue bars) ovaries following either no treatment (untreated; filled bars) or 44 h eCG treatment (striped bars). Lymphatic vessels number and distribution were analysed (A, B) as well as size and distribution (C, D). Morphometric analysis was based on analysis of serial ovarian sections at 35�m increments through all ovaries. Data represents mean ± SEM, n = 5 animals per genotype and treatment. * represents statistical significance (P�0.05). # represents P�0.07.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
73 BROWN HM
4.10 A). Importantly, the genes known to be necessary for lymphangiogenesis, Vegfc, Vegfd and Vegfr3 were also significantly increased in the whole ovary in response to eCG treatment. The Vegfc mRNA expression increase following eCG stimulation was significant only in the Adamts1-/- ovaries (Figure 4.10 B), whilst Vegfd was significantly induced in both genotypes (Figure 4.10 C). Vegfr3, the receptor responsible for mediating the lymphangiogenic effects of Vegfc and Vegfd was also significantly increased in both genotypes in response to treatment (Figure 4.10 D). As expected, Aromatase expression was significantly increased after eCG treatment and was not significantly different between genotypes (Figure 4.10 E). These results indicate that FSH action has a specific effect on an ovarian lymphangiogenic regulatory mechanism, which, as shown above, promotes significant lymphatic vascular changes.
4.3.6 Primary ovarian lymphangiogenic establishment is exaggerated by eCG treatment
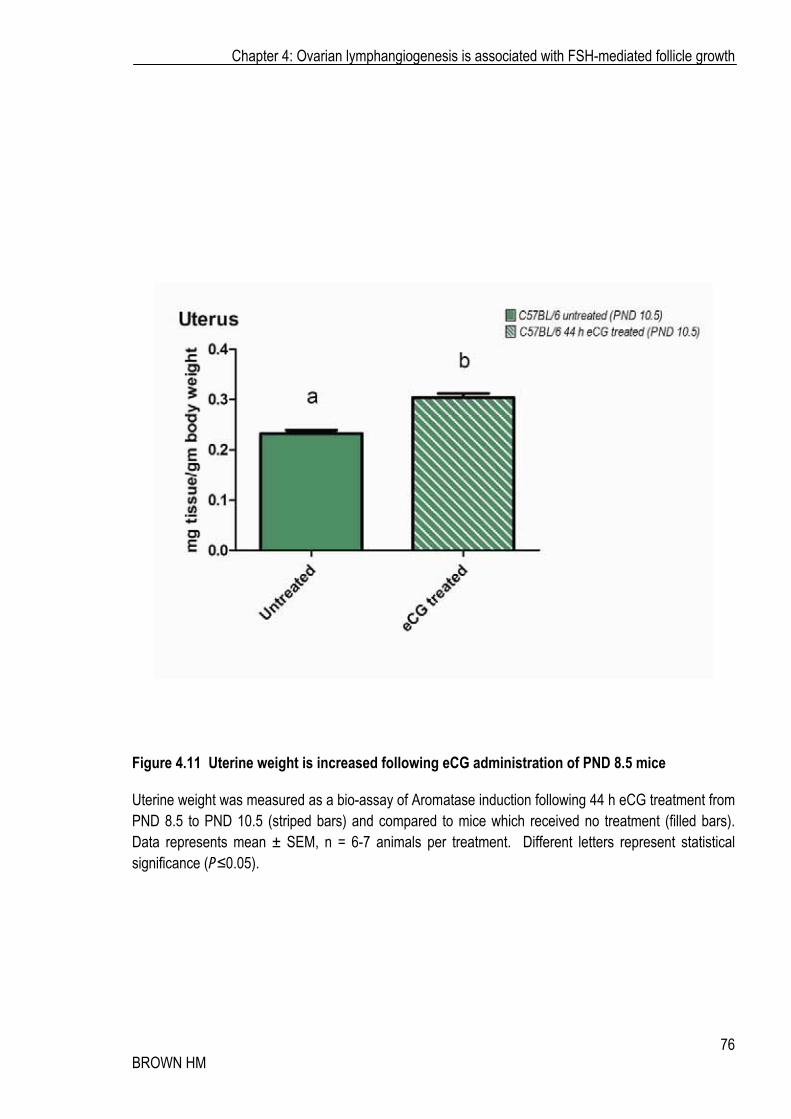
After demonstrating the ability of FSH-analog eCG to mediate changes to the ovarian lymphatic vasculature of the prepubertal ovary, I examined if eCG had the capacity to modulate the initial postnatal ovarian establishment (as described in Chapter 3). Mice were administered with eCG (equivalent dose for weight; 500IU/kg) at PND 8.5 (Figure 4.1 B); the time when stalk vessels are first present, but no vessels are associated with follicles or cortex. eCG stimulation induced a small but significant increase in uterine weight (Figure 4.11 A); indicative of enhanced estrogen secretion and indicates the induction of biologically relevant induction FSH-associated pathways in the ovary.
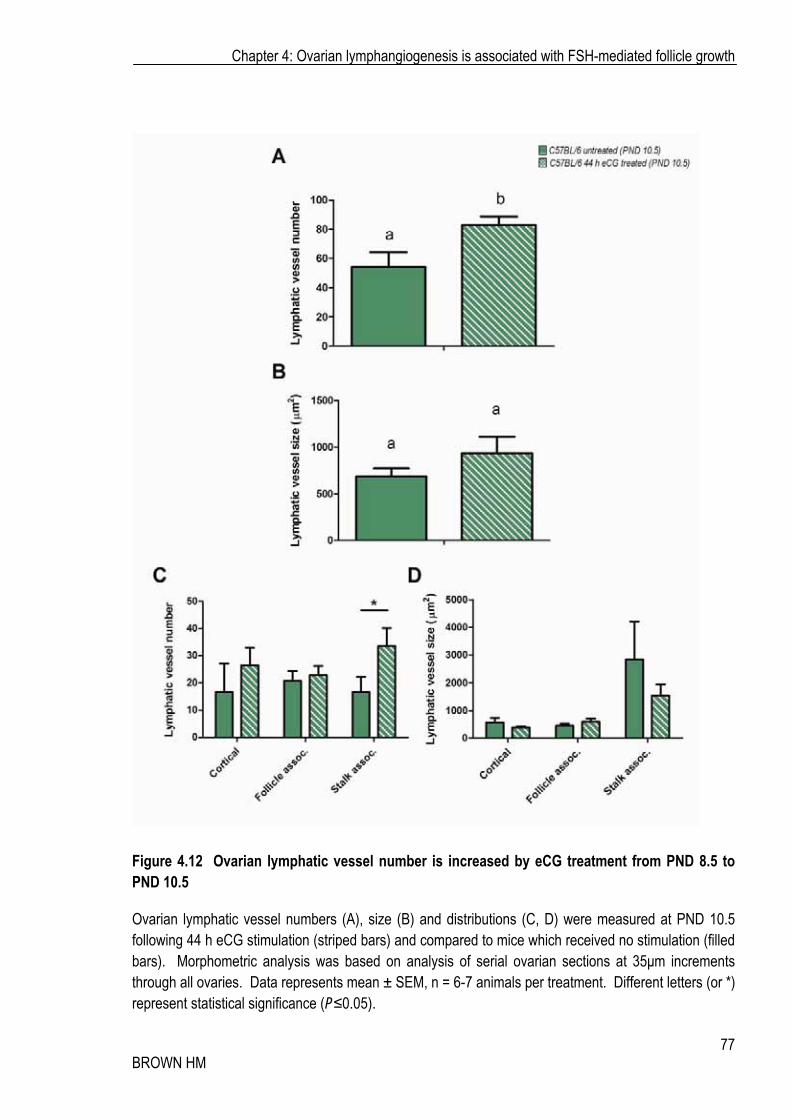
Stimulation with eCG significantly increased the number of lymphatic vessels per ovary by approximately 50%, from 54 to 82 (Figure 4.12 A). Lymphatic vessel number significantly increased in the medullar/stalk region but was unchanged in the cortical or follicle associated regions (Figure 4.12 C), while the vessel size was unaffected (Figure 4.12 B, D). eCG stimulated a moderate, but non-significant increase in the mRNA expression of Vegfd, and had no effect on expression of Vegfr3 (Figure 4.13 A-B).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
74 BROWN HM
Figure 4.10 eCG regulated mRNA expression of lymphangiogenic-related genes in the ovary
Using real time RT-PCR analysis, Lyve1 (A), Vegfc (B), Vegfd (C), Vegfr3 (D) and Aromatase (Cyp19a1) (E) expression was analysed Adamts1+/- (red bars) and Adamts1-/- (blue bars) whole ovaries following either no treatment (untreated; filled bars) or 44 h eCG treatment (striped bars). Data represents mean (2-��CT) ± SEM, n = 5 animals per genotype and treatment. Different letters represent statistical significance (P�0.05).
Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
75 BROWN HM
Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
76 BROWN HM
Figure 4.11 Uterine weight is increased following eCG administration of PND 8.5 mice
Uterine weight was measured as a bio-assay of Aromatase induction following 44 h eCG treatment from PND 8.5 to PND 10.5 (striped bars) and compared to mice which received no treatment (filled bars). Data represents mean ± SEM, n = 6-7 animals per treatment. Different letters represent statistical significance (P�0.05).
Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
77 BROWN HM
Figure 4.12 Ovarian lymphatic vessel number is increased by eCG treatment from PND 8.5 to PND 10.5
Ovarian lymphatic vessel numbers (A), size (B) and distributions (C, D) were measured at PND 10.5 following 44 h eCG stimulation (striped bars) and compared to mice which received no stimulation (filled bars). Morphometric analysis was based on analysis of serial ovarian sections at 35�m increments through all ovaries. Data represents mean ± SEM, n = 6-7 animals per treatment. Different letters (or *) represent statistical significance (P�0.05).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
78 BROWN HM
Figure 4.13 eCG administration from PND 8.5 to PND 10.5 modulates ovarian mRNA expression of Vegfd, but not Vegfr3
Vegfd (A) and Vegfr3 (B) mRNA expression was analysed using real time RT-PCR in whole ovary samples following 44 h eCG treatment (striped bars) or in mice which received no treatment (filled bars). Data represents mean (2-��CT) ± SEM, n = 6-7 animals per treatment. # represents P�0.07, NS represents no significant difference.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
79 BROWN HM
4.3.7 Ovarian lymphatic vessel distribution in the ovulating follicle and corpus luteum differs from the blood vasculature
To examine the distribution of ovarian lymphatic vessels as the follicle transitions into a corpus luteum, ovaries were isolated from female mice that were either cycling, or which has been treated with an ovulatory dose of hCG to mimic the LH surge. The theca, at this time undergoes the migratory/invagination process by which the thecal cells invade the follicle structure to vascularise the developing CL. While known to be rich in blood vascular endothelial cells preparing to invade and colonise the newly forming CL, the invading theca were void of Lyve1-positive lymphatic vessels. (Figure 4.14 A-B; boxed area). Interestingly, it appeared that Lyve1 staining was present within the granulosa cells of the peri-ovulatory follicle, which was further explored in Chapter 5. Corpora lutea were rich in blood vascular capillaries as has been well described [188] (Figure 4.15 C-D, arrows), while in contrast, CLs were completely void of lymphatic vessels (Figure 4.15 A-B). Large lymphatic vessels were present in the stroma closely associated with the edge of the CL.
4.4 DISCUSSION Whilst the changes to the pattern of blood vessels around growing follicles and within the CL have been widely explored, the dynamics of the ovarian lymphatic vasculature has not been examined, even considering that lymphatic vessels have been reported in mouse [142], human [189], Caiman crocodile [190], dog [191, 192], cat [193], rabbit [125, 194] and rat [124]. I took advantage of two novel models to better characterise the lymphatic vessel deficient Adamts1-/- mouse and the PND 8.5 to 10.5 mouse in which ovarian lymphangiogenesis first occurs, to interrogate the lymphatic response in association with gonadotrophin-dependent follicle growth and investigate the role for Adamts1.
Taking into consideration the extensive blood vascular remodelling concomitant with rapid follicle growth, I hypothesised that the lymphatic vasculature would undergo similar remodelling in response to FSH. Indeed, eCG approximately doubled the mean size of the ovarian lymphatic vessels in Adamts1+/-

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
80 BROWN HM
Figure 4.14 Lymphatic vessels are found in the stroma, but not within the invaginating thecal region of peri-ovulatory follicles
Lymphatic vessels (Lyve1-positive; A, B) were localised in ovaries following 44 h eCG + 10 h hCG stimulation using immunohistochemistry on tissue sections. Ovarian lymphatic vessels were found associated with growing follicles (B) within the stromal compartment, however no lymphatic vessels or capillaries were detectable within the invaginating theca (B, boxed). Images were captured at 10x (A) and 20x (B).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
81 BROWN HM
Figure 4.15 Lymphatic vessels are found in alongside, but not within the corpus luteum
Lymphatic vessels (Lyve1-positive; A, B) and blood vessels (Von Willebrand Factor; C, D) were localised in adult cycling ovaries using immunohistochemistry on ovarian tissue sections. Ovarian lymphatic vessels were found within the stromal compartment of the ovary (A, B), whilst blood vessels were found in and around the CL. No lymphatic vessels or capillaries are detectable within the CL (B) compared with the extensive blood vascular network within the CL (D, arrows). Images were captured at 10x (A, C) and 20x (B, D).

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
82 BROWN HM
control mice, while it had no effect on the vessel number. This suggests that once the lymphatic vasculature is established within the ovary, vessel number remains static, and perhaps a limitation exists on the number of vessels within the organ; but that vessel size may be modulated to provide the necessary functional adaptations needed to accommodate a large growing follicle pool. Conversely, in the Adamts1-/- ovaries, where normal lymphatic vascular establishment fails, eCG promoted lymphangiogenesis and the establishment of an ovarian lymphatic network; increasing vessel number and size. Mice lacking Adamts1 fail to develop a normal ovarian lymphatic network postnatally and have a very minimal number of vessels at d 21 (less than 6% of their heterozygous counterparts). Following stimulation with eCG, the ovarian lymphatic vessel population expanded more than five-fold in number and increased in size; however both size and number remained significantly lower than their Adamts1+/- counterparts, further confirming a non-redundant, essential role for Adamts1 in normal ovarian lymphangiogenesis.
These vascular remodelling events are novel and exciting as they are the first evidence that the lymphatic vasculature is capable of non-pathological remodelling in response to hormonal cues. Secondary lymphangiogenesis is poorly understood, and until now, thought only to occur during the pathophysiological circumstances of wound healing and tumourigenesis, much like angiogenesis. Small lymphatic vessels and capillaries are anchored to the ECM by filaments, and as such, can expand in situations of high flow to allow the passage of large protein moieties. As a result, it the increase in vessel size described here in response to eCG stimulation may in fact be as a result of increased flow and sheer stress, rather than active vessel remodelling. The large (two-fold) increase in vessel size and Lyve1 expression along with the significant induction of the lymphangiogenic mechanism (Vegfc and Vegfr3 expression) suggests that there is, in fact, active vascular growth and remodelling, not just as a consequence of flow, within the ovary in response to FSH-analog eCG. These results suggest that there is significant vascular remodelling associated with FSH-mediated follicle growth within the ovary.
Excitingly, when treated at PND 8.5, prior to (and during) the establishment of the ovarian lymphatic vasculature, eCG was able to significantly modulate lymphangiogenesis. During this period of vascular establishment, the lymphatic vessels are expected to be actively dividing and colonising the tissue. Hormonal manipulation during this establishment window resulted in a significant increase in the

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
83 BROWN HM
number of lymphatic vessels within the normal mouse ovary and an increase in the expression of lymphangiogenic growth factor, Vegfd. In most tissues, including gut, lung, heart and skin the lymphatic vasculature is present well before birth (prior to or at E 15) [109], however the developmental aetiology of their lymphatic vascularisation is unknown. Expansion of these vessels occurs following, or as part of the initial colonisation of the organ, and is mediated in part by neuropilin 2 (Nrp2), a non-kinase trans-membrane receptor involved in axon guidance and known to bind members of the Vegf family including Vegfc [109]. It is likely that Vegfc and Vegfr3 are also highly implicated in this process; however the extent of their involvement has likely been hindered by the embryonic lethality and failure to establish any lymphatic vasculature in their absence. The present data suggests that, within the ovary, hormonal manipulation during lymphatic establishment of the organ promotes development and expansion of the lymphatic vessel network.
Two previous reports have suggested a hormonal influence on lymphangiogenesis. The lymphatic vasculature has been described in the oviduct and/or ovarian bursa for species including mouse [195], guinea pig [196, 197], pig, [198] and European beaver [199]. Estradiol treatment increased mouse ovarian bursal lymphatic stomata (vessel-opening into the bursa) size and intrabursal absorption (vessel function) [195], whilst pregnancy or PMSG+hCG treatment also increased size and function (tracer absorbance) in the guinea-pig ovarian bursa [196]. This supports the hormonal changes to the lymphatic vasculature I have described within the ovary, and also suggests that the change in lymphatic vessel size may be of functional importance.
Interestingly, the spatial distribution of the lymphatic vasculature does not always closely match the ovarian blood vasculature. The lymphatic vasculature does not appear to form a capillary network around growing follicles during pre-antral, antral or pre-ovulatory growth; unlike the blood vasculature, which develops a rich network within the thecal layers during gonadotrophin-dependent follicle growth [62]. The lymphatic vessels that were associated with growing follicles were much larger than capillaries, found within the theca or stroma, and often in close association with the follicle edge. The lymphatic vasculature was also present in the medullary region of the ovary, which is also rich in larger blood vessels, and was more often than not, very closely associated with the blood vasculature, often directly alongside them. In stark contrast to the intense blood angiogenesis seen within the CL, it was

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
84 BROWN HM
devoid of lymphatic vessels. I was able to localise lymphatic vessels to the outer periphery of CLs, in the adjacent stroma, but not within the CL itself. Other reports suggest that lymphatic vessels are found in the peripheral zone of the CL in later stages of development in rats [124] and lymphatic capillaries have also been observed among the theca lutein, but not granulosa lutein cells three days following hCG stimulation in the rabbit [125]. Both of these observations were made prior to the identification of a lymphatic specific marker. It is possible that species differences exist with regard to the lymphatic vasculature within the CL, and detailed multi-species analysis using known lymphatic markers could further elucidate these details. It is interesting that the distribution of the microvasculature of both vessel types is different within the ovary. The absence of Lyve1-positive lymphatic capillaries within both the theca of growing follicles and also the CL suggest that, at least within these compartments of the ovary, tight interactions between the two vasculatures may not exist. Whilst purely speculative, the lack of lymphatic vasculature within the theca may be necessary for the accumulation of extravascular fluid during antrum formation, which would normally be removed by the lymphatic vasculature if associated with an edematous pathology.
Whilst the functions of the lymphatic vasculature within the ovary remain speculative, the possibilities of increased functional requirements following eCG (or FSH) stimulation are considerable. Antrum formation requires rapid changes in the ovarian fluid dynamics and the lymphatic vasculature may play an important role. The association of lymphangiogenesis with hormone-mediated folliculogenesis and the association of failed lymphangiogenesis and folliculogenesis in Adamts1-/- mice implicates a role for lymphatics in supporting normal follicle growth. The removal of cellular debris from regressing follicles and the CL by macrophages may require a robust lymphatic network for trafficking macrophages out of the ovary. Furthermore, lymphatic vasculature may participate in normal immune cell trafficking and surveillance of the ovary. Localisation of antigen presenting cells (APCs) including macrophages and dendritic cells using markers including F-4/80, CD11b and CD11c or the pan-immune cell marker CD45 (Common Leukocyte Antigen) both prior to, and following establishment of the lymphatic vasculature in the Adamts1-/- may provide valuable information regarding both the immune trafficking functions of the lymphatic vasculature, as well as insight into the other reproductive phenotypes.

Chapter 4: Ovarian lymphangiogenesis is associated with FSH-mediated follicle growth
85 BROWN HM
The experiments in this chapter clearly demonstrate that FSH-analog eCG is able to promote ovarian lymphangiogenesis. This work provides novel insight into both the dynamics of the ovarian lymphatic vasculature in association with FSH-mediated follicle growth and its hormonal regulation; however what remains unclear is the how eCG mechanistically promotes lymphangiogenesis. In response to FSH, the growing follicle is capable of the production of blood angiogenic mediators and the paracrine regulation of the blood vasculature [29, 184, 185]. Increased Vegfc/d reported here indicates that similar mechanisms may be active in the promotion of ovarian lymphangiogenesis in response to FSH, whereby the follicle can produce lymphangiogenic mediators. The regulation of the blood and lymphangiogenic factors in the ovary is surprisingly poorly understood and will be further explored in the subsequent chapter.


















