
CHAPTER 5 CHARACTERIZATION AND APPLICATION OF ENZYMES...
24
CHAPTER 5 CHARACTERIZATION AND APPLICATION OF ENZYMES FROM THE CORM
Transcript of CHAPTER 5 CHARACTERIZATION AND APPLICATION OF ENZYMES...

CHAPTER 5
CHARACTERIZATION AND APPLICATION OF ENZYMES FROM THE CORM


63
CHARACTERIZATION AND APPLICATION OF ENZYMES FROM THE CORM
OVERVIEW OF THE CHAPTER
Amorphophallus skinned corms were screened for cellulase and polyphenol oxidase (PPO)
enzyme. The extract showed the presence of cellulase and PPO enzyme activity which was
further confirmed by Congo red plate diffusion and catechol agarose plate assay respectively.
The major useful factors in controlling enzyme activity i.e. temperature, pH, chemicals which
can inhibit enzyme action, substrates alteration were checked. Extracted PPO was purified by
acetone precipitation and ion exchange chromatography and characterized using
spectrophotometric methods. Crude enzyme fractions having cellulase activity could clarify
apple juices where as crude enzyme having polyphenol oxidase activity shows dough raising
application.

64
CHARACTERIZATION AND APPLICATION OF ENZYMES FROM CORM
5.1 INTRODUCTION
Plants usually contain 35–50% cellulose on dry weight basis making it the abundant organic
compound in nature. Cellulose is found in most abundance and represents 1.5x10 12 tons of
total annual biomass production through photosynthesis[149]. It is a highly stable polymer
consisting of β-1, 4-linked glucose units[150-151]. Lignin, hemicelluloses and other back-up
substances provide support against cellulolytic activities. Cellulases refer to cellulose-
degrading enzymes with potential to convert cellulosic material into its subunit-glucose [50].
Increasing demand for renewable energy sources has sparked growing interest in enzymes
capable of degrading cellulose to sugars that can then be used for the production of ethanol
[152] Enzymatic hydrolysis is an economic process in the conversion of cellulose to easily
fermentable low cost sugars. Cellulases have been commercially available for more than 30
years, and these enzymes have represented a target for both academic as well as industrial
research[70], [153], [154]. Higher plant cellulases like Lantana camara and Cuscuta reflexa
are mostly involved in fruit ripening and senescence. Cellulases are used in the textile
industry for cotton softening and denim finishing, in laundry, detergent market for
colour care, cleaning, in the food industry for mashing; Cellulase is used for commercial
food processing in coffee and it also performs hydrolysis of cellulose during drying of
beans[155]–[157]. In pulp and paper industries it has found application in deinking, drainage
improvement, and fibre modifications. High cost and low activity are the major impediments
to the commercial use of cellulases [158].
Unfavourable oxidative browning (mainly due to the oxidation of widely distributed enzyme
polyphenol oxidase) occurs in the damaged tissues during drying processes and other
technological operations. Polyphenol oxidase (PPO, a copper-containing enzyme) is one of
the most common browning agents in nature and enzyme catalyzes the hydroxylation of
monophenols to O-diphenols and oxidation of O-diphenols to O-diquinones. Browning is a
prominent problem for elephant foot yam derived products. This undesirable enzymatic
browning occurs in many plants and vegetables and is of a great concern to food
technologists and processors. In plants, research on PPO is mainly focused on its roles in

65
darkening of the damaged tissue along with browning [159]. Poly-phenolic compounds are
closely related to the sensory, aroma and nutritional qualities of plants. Storage, processing
and handling also leads to browning as demonstrated in the case of potato tubers [160],
artichoke [161][162], yacon roots [163], lettuce [164] and green beans[165] making them
less appealing to consumers and therefore results in the loss of their marketability.
This aroid Amorphophallus paeoniifolius has not been explored much; the identification of
cellulolytic enzymes and PPO enzyme from corm is unique work and can be commercially
harnessed by the fruit juice industry and bakery industry which is among the largest agro-
based industries worldwide. High juice yield is an important goal for juice production. Many
modern processes for fruit and vegetable juice production employ enzymes as important
processing aids to obtain higher yields and clarity[166].
This study will not only provide some fundamental understanding of the PPO and cellulase
enzyme of Amorphophallus but also provide a new enzyme source for different industrial
applications.
5.2 MATERIALS AND METHODS
5.2.1 Plant material and Chemicals
A. paeoniifolius corms were purchased from local vegetable market at Noida, India. Peels
were separated and processed separately for enzyme and phytochemical extraction[40],[167].
In this study, the peeled corm was washed under running water for 30 min to remove surface
contamination and then chopped into small cubes of 4mm, rinsed with distilled water and
stored at 4°C. Pyrocatechol, ascorbic acid, EDTA, D and L Tyrosine was procured from
CDH, New Delhi. PVP (Polyvinylpyrrolidone), potassium mono-hydrogen phosphate and
potassium di-hydrogen phosphate, sodium dodecyl sulphate (SDS), dithiothreitol (DTT) were
procured from Sigma Chemical Co (St. Louis, USA). Macro-prep DEAE support were taken
from BIO RAD. All the other reagents used were of analytical grade.

66
5.2.2 Preparation of corm homogenate as enzyme source Stored peeled corm was cleaned with distilled water, the weighed corm was chopped into
small pieces and homogenized in ratio of 1:3 with 0.1 M Citrate buffer, pH 5.0 for cellulase
enzyme and for PPO A. paeoniifolius peeled corm was homogenized in chilled pH 7.0, 0.1 M
phosphate buffer in a ratio of 1:3 along with 1% of polyvinylpyrrolidone (PVP). All the steps
of enzyme extraction were carried out at 4°C. Crude extract was centrifuged at 10,000 rpm
for 30 min at 4°C in a refrigerated centrifuge to obtain a clear supernatant that was used as
the enzyme source. The pellet was discarded whereas the supernatant was filtered through
Whatman No.1 filter paper and stored at -20°C. Protein and enzyme activity was estimated
for this extract prior to storing at -20°C.
5.2.3 Purification of the PPO extract
The crude protein extract was precipitated out by slowly adding (−20°C) pre chilled acetone.
This solution was stored at 4°C for 2 h. The acetone precipitate was collected and left for 24
h at 4°C to remove acetone. The precipitate was re-suspended in 100 mM phosphate buffer
(pH 7.0), stirred for 10 min and the suspension was centrifuged at 5000 rpm for 10 min. The
supernatant was pooled and loaded on DEAE (Diethylaminoethyl) anion exchanger (Macro-
prep DEAE support) column that had been pre-equilibrated with 100 mM phosphate buffer
(pH 8.0). Unbound of DEAE was collected in fractions of 2 ml each. Elution was done with
buffered solution (pH 8.0) containing 10, 20, 50 mM NaCl. Eluate fractions (2ml) were
collected and dialysed to remove salt. Both unbound and dialysate were assayed for
polyphenol oxidase activity. Active enzyme fraction were pooled and stored at -20°C.
5.2.4 Measurement of the enzyme activity
5.2.4.1 Carboxymethyl-cellulase (CMCase) and Filter-paperase (FPase) enzyme
activity assay
CMCase and FPase activity was assayed using a modified method described by (Wood and
Bhat, 1988),[168] with some modifications. 0.1 ml of supernatant was added to 900 μl of 0.1
M Citrate buffer, pH 5.0 and 1.0 ml of 1% CMC (for CMCase) and Whatman no. 1 filter
paper strip (1 x 3 cm, 25 mg) (for FPase) was added in a test tube and incubated at 50°C for
60 min. The reaction was terminated by adding 3.0 ml of 3, 5-dinitrosalicylic acid (DNS)
reagent and by subsequently placing the reagent tubes in water bath at 100°C for 15 min

67
(Miller, 1959). One ml of Rochelle salt solution 40% was then added to stabilize the colour.
The absorbance was recorded at 540 nm against the blank (without enzyme filtrate). One
international unit of CMCase and FPase activity was expressed as 1 μ mole of glucose
liberated from per ml enzyme per min under assay condition.
5.2.4.2 Polyphenol oxidase enzyme assay
PPO activity was determined spectrophotometrically by measuring the initial rate of quinone
formation, which is indicated by an increase in absorbance at 420 nm. The reaction mixture
contained 1000 µl of 0.1 M phosphate buffer (pH 7), 50 mM catechol solution, and 100 µl of
the enzyme solution and 900 µl of distilled water. The reaction was carried out for 15 min at
35°C. One unit of PPO activity was defined as the amount of enzyme that caused an increase
of 0.001 unit of absorbance per minute under standard assay conditions.
5.2.5 Protein assay Protein concentrations were determined according to the dye binding method of (Bradford,
1976) [169], using bovine serum albumin as standard
5.2.6 Congo red plate diffusion assay The evaluation of cellulase enzyme was identified employing the Congo red plate assay as
described by (Teather and Wood, 1982) [170]. Agar plates were prepared containing 0.5%
carboxy methyl cellulose incorporated into 1% w/v agar in milli Q water. The experimental
steps were conducted in sterile condition in an incubator set at 50°C. The 50 μl enzyme
sample was applied onto each agar well. Following a 24–48 hours incubation period at 50°C,
the wells were washed off with distilled water and were stained with Congo red solution for
30 min. The gels were soaked in 1 M NaCl until clear yellow zones were detected.
5.2.7 Polyacrylamide gel electrophoresis and Activity staining of cellulase
SDS-PAGE was performed on a slab gel containing 10% (w/v) polyacrylamide by the
method of Laemmli (1970) [171]. The gel was stained with Coomassie brilliant blue (0.5%
w/v) for 30 min and de-stained in 10% methanol and 5% acetic acid for a limited period of
time.

68
The activity staining method was performed by modified method as reported by Trudel and
Asselin (1989) [172]. Carboxymethyl cellulose (0.5% and 1%) substrate was boiled and
incorporated into the analytical 10% (w/v) polyacrylamide gel. The isolated protein samples
were run in the substrate incorporated gel at 100V constant voltage for 2-3 hours. After
electrophoresis the gels were soaked in 0.1 M citrate buffer pH 5.0 for two 15 min changes
for 24 hours to renature the enzymes in gel. The polyacrylamide gels were stained for 30 min
in Congo red and de-stained in 1 M NaCl for at least 30 min. Bands with lytic activity
appeared as clear zones against bright red background.
5.2.8 Gel diffusion assay for polyphenol oxidase quantification
Agarose (1.5% (w/v); phosphate buffer pH 7.0) was heated in microwave oven for 5 min, the
solution was gradually cooled to 50°C and 50 mM catechol was added [173]. 25 ml of this
buffered solution was transferred into round petri dishes, cooled at 350 C and stored at 4°C in
dark conditions. Wells of 5 mm diameter were created using a cork borer and gel plugs were
removed. Crude enzyme extract (20-100 ul) was loaded into each 5 mm well and the effect of
altering concentrations on sensitivity and linearity of the gel diffusion assay was studied by
monitoring the dark ring diameters formed after 12 h of incubation. Plates were covered and
left for incubation at 35°C in incubator for 12 h. The diameters of the intensely stained zones
were also recorded.
5.2.9 Detection of PPO activity by SDS PAGE electrophoretic-blot technique
Preparation of Catechol paper
3mm chromatography (15x12 cm) paper was immersed in 50 mM catechol solution for 10
min. The catechol paper was dried at 350C for 15 min and stored in dark condition till further
use.
SDS PAGE and Electrophoretic-blot
20 µg of protein from crude, acetone and unbound fractions were loaded on 10% SDS PAGE
gel. For determining the molecular weight of A. paeoniifolius PPO enzyme, the gel was
checked for activity staining by the electrophoretic blot process. After electrophoresis, one

69
part of gel was stained with Coomassie Blue R-250 to determine the molecular weight
whereas the other side of the glass plate along with the gel was removed and immediately
blotted onto the top of the dried catechol paper. A dark activity brown band developing
within 5- 10 min validates the presence and molecular weight of PPO [174].
5.2.10 Characterization of cellulase 5.2.10.1 Effect of temperature, pH and various metal ions on enzyme activity The cellulolytic activity of the crude enzyme was measured at different temperatures (30 -
90°C), pH values (2 – 10) and with different metal ions (100 mM). The pH was adjusted
using the following buffers (0.1 M) of citrate buffer (pH 2.0- 6.0), Tris (pH 7-10). The
reaction mixture was pre-incubated for 15 min with all the mentioned temperature, pH and
metal ions. After pre-incubation the activity of the enzyme was measured as per standard
assay conditions.
5.2.11 Determination of the PPO enzyme properties
5.2.11.1 Effect of pH and pH stability on PPO activity
The effect of pH on PPO activity was determined under standard assay conditions using 0.1
M hydrochloric acid potassium chloride buffer (HCl-KCl) ( pH 2), 0.1 M citrate buffer (pH
3-6), 0.1 M potassium phosphate buffer (pH 7-8), 0.1 M Tris-HCl buffer (pH 9). The blank
contained only substrate solution and the reaction buffer. The optimum pH and the enzyme
activity were expressed as the percentage of maximum enzyme activity under standard assay
condition.
The pH stability of the enzyme was determined according to a modified method of (Onsa et
al. 2000) [175]. 50µl of enzyme solution was incubated with 1 ml of different pH as
described for optimum activity for 24 h at 4°C, and the residual activity was determined and
calculated as relative activity (%) to the initial activity.
5.2.11.2 Effect of temperature on PPO activity
The buffered enzyme solution was pre-incubated in a water bath at different temperatures
(35°C to 75°C) for 10 minutes prior to addition of substrate 5mM catechol. Relative enzyme

70
activity was calculated as the percentage of the highest activity left after temperature
treatment.
5.2.11.3 Effects of inhibitors
Sodium azide, sodium chloride, ascorbic acid, citric acid CaCl2, ZnSO4 and EDTA were
used as PPO inhibitors. The reaction mixture contained 1 ml of 0.1 M phosphate buffer, pH
(6.9), 950ul of 100 mM inhibitor, 0.50 ml of enzyme extract and 1 ml of 50 mM catechol.
Percentage inhibition was calculated using the following equation:
Inhibition (%) = (Ao - Ai/Ao) 100,
Where Ao: initial PPO activity (without inhibitor) Ai: PPO activity with inhibitor
5.2.12 Application of crude extract
5.2.12.1 Apple juice clarification having cellulase activity
Crude enzyme extract having cellulase activity (30units) was incubated with 5ml of freshly
extracted apple juice, filtered through three layers of cheese cloth for 1 hour at 50°C. The
contents of the tubes were stirred well in order to mix the enzyme with juice. The reaction
was performed in water bath. Clarification of juice was observed after 50 min. Test for starch
and pectin was carried out to test the effectiveness of the enzyme treatment (results not
shown).
5.2.12.2 Dough rising application of crude extract having PPO activity
All the ingredients (salt, sugar, yeast and wheat flour) required to make dough were weighed
and mixed thoroughly. Ingredients were kneaded in water along with 1 ml of crude enzyme
extract (having PPO activity) to form dough whereas control has enzyme blank. The dough
mixture was transferred to marked beaker and kept at room temperature. Rise in dough level
was constantly monitored.

71
5.3 RESULTS AND DISCUSSION
5.3.1 Cellulase from stored Amorphophallus paeoniifolius in clarification of
apple juice
Cellulase enzyme was isolated from corm of Amorphophallus and enzyme activity was
estimated by the method of (Wood and Bhat, 1988)[168]. Optimum pH, temperatures were
determined and investigation was carried out to determine the role of metal ions on enzyme
activity. Optimum temperature for CMCase and FPase both was found to be 60°C as shown
in Figure 5.1(a). Thermal stability studies suggested that cellulase extracted from corm
retained activity even at 90°C. CMCase activity was found to be optimum at pH 10
whereas FPase activity was found to be optimum at pH 5, as shown in Figure 5.1(b).
CMCase retained its activity at both acidic and basic pH but interestingly at pH 7 both
the CMCase and FPase activity decreased.
Figure 5.1: Effect of (a) temperature and (b) pH on enzyme activity CMCase and FPase. Maximum specific
activity obtained for the enzymes was taken as 100% for the calculation of relative activity.
Congo red plate diffusion assay confirmed the presence of cellulase activity in the isolated
enzyme fraction and the clear zone were visualized under white light illumination. Control
with distilled water shows no clear zone. Radial diffusion of enzyme into CMC incorporated
gel produces hydrolysis zones, visualized by staining with congo red dye (see Figure 5.2).

72
Figure 5.2: Congo red plate diffusion assay
1% Agar along with 0.5% carboxymethyl cellulose plates were prepared. 50 microliters of crude extract was
loaded in wells and plates were left for incubation for 24–48-h at 50 °C, the wells were washed with distilled
water and stained with Congo red solution for 30 min., followed by washing in 1 M NaCl. Presence of clear
yellow zones around the wells 1-4 confirmed the presence of CMCase activity. C- control with distilled water
Polyacrylamide gel electrophoresis and zymography studies where substrate CMC was
polymerised in gel yielded bands with lytic activity after staining with Congo red as dark
zones shown in Figure 5.3. The molecular weight of the enzyme is 66 kDa as determined by
the zymographic studies.

73
Figure 5.3: SDS PAGE (a), Native(b) and Zymography(c) of crude enzyme
M: Marker, lane A1 is SDS PAGE analysis of crude extract, lane 1 is the Native gel analysis of the crude
extract, in lane 2 (substrate CMC 1%) and lane 3 (substrate CMC 0.5%) was polymerised in the native gel.
Clear zones in lane 2 and 3 indicate that molecular weight of enzyme is around 66 kDa.
a)
b)
c)

74
Investigation was also carried out to determine the role of metal ions in the stabilization of
cellulase enzymes.
Figure 5.4: Effect of metal ions on cellulase specific enzyme activity
Enzyme was incubated with various metal ions and control without the metal ions was also
taken. CMCase and FPase specific enzyme activity was calculated as shown in Figure 5.4.
Control specific enzyme activity (without the addition of metal ions) was taken as 100%.
Mn2+ has strong inhibitory effect on both CMCase and FPase, whereas Mg2+ activated the
CMCase activity maximum and Fe3+ strongly activated FPase activity.
This crude fraction rich in Cellulase was also checked for apple juice clarification. The
enzyme extract, clarified freshly extracted apple juice. Freshly extracted and filtered apple
juice was incubated with the enzyme extract in the ratio of 5:1 for 1 hour at 50°C.Assay was
done in triplicate and the clear juice was obtained after 1 hour of incubation with the enzyme
as compared to the control tube which did not contain the extract (see Figure 5.5).
0
50
100
150
200
250
Control MgSO4 CuCl 2 FeCl3 MnSO4 NaCl KCl HgCl2
Spe
cifi
c e
nzy
me
act
ivit
y %
Metal ions
CMCase
FPase

75
Figure 5.5: Clarification of apple juice with crude enzyme extract of Amorphophallus corm
Test tube no 1-3 shows clearing of apple juice as compared to the control tube (no 4) which did not contain the
extract.
Carbohydrases level increase on storage and ageing of tuberous crop and we report that these
native enzymes of Amorphophallus corms can degrade polysaccharide material of apple juice
thus facilitating clarification.
5.3.2 Screening, purification and characterization of Polyphenol oxidase
from A. paeoniifolius corm
5.3.2.1 Extraction and partial purification of PPO
Poly phenol oxidase enzyme from corm of A. paeoniifolius was extracted in 0.1 M phosphate
buffer pH 7 and 1% PVP followed by acetone precipitation and ion exchange
chromatography by using DEAE. Undesirable reaction between the natural phenols and PPO
was prevented by adding 1 % polyvinylpolypyrrolidone (PVP). PVP is a phenol adsorbing
agent and added to the extraction buffer to remove phenolic substrates due to its binding
ability to the phenolics in order to prevent phenol-protein interactions. After acetone
precipitation the pellet was suspended and loaded onto a DEAE-cellulose ion-exchange

76
column. Activity rich DEAE unbound fractions were pooled and dialysed prior to studying
its kinetics. DEAE eluates of NaCl 10, 20 and 50 mM did not show any enzyme activity.
We report yield of 18.92 and 1.24 fold purification after acetone precipitation and unbound of
DEAE gave 5.54 fold purification (see Table 5.1).
Table 5.1: Purification of PPO from Amorphophallus paeoniifolius
Purification step Volume
Protein (mg/ml)
Total protein
Activity (U/ml)
Specific Activity(U/mg protein)
Total activity Yield
Purification fold
Crude extract 50 0.074 3.675 833 11333.33 41650 100 1 Acetone precipitation 4 0.14 0.56 1970 14071.43 7880 18.92 1.24 Unbound DEAE Anion Exchange chromatography 2.25 0.010 0.0212 590 62765.96 1327.5 3.19 5.54
Catechol agarose gel diffusion assay for quantification of PPO, Crude enzyme (20, 40, 50,
60, 100 ul) was added in the wells and kept for 12 hours along with control i.e. enzyme blank
well. Diameters of the oxidized zones (area turning black in colour) were measured. Graph
was plotted between the log units of the enzymes and the diameter of the oxidized circles. On
generation of standard curve linear relationship found between the diameters of dark oxidized
circles formed in the gel and the logarithm of the enzyme activity units added to the well,
with correlation coefficient of 0.8921. (see Figure 5.6 a & b).

77
Figure 5.6: Identification and quantitation of PPO activity. (a) catechol agarose plate assay: 5 mm diameter wells was loaded with crude enzyme (20-100 ul) incubated at
35°C in for 12 h in dark.
(b) Diameter (cm) of the dark circles formed was measured and the standard curve between the logarithm of
PPO activity and the diameter of stained zones formed on catechol agarose plates (y = 0.6478x + 0.1475
R² = 0.8921
5.3.2.2 pH optimum and stability
The effect of pH on PPO activity was determined by estimating enzyme activity at different
pH 2.0- 9.0 using catechol as substrate. We report that Amorphophallus PPO shows optimum
activity at pH 6.0 (see Figure. 5.7). The activity was found to increase upto pH 6.0 after
which it dropped considerably giving a bell-shaped curve. Our results were comparable to
PPO activity from other plant sources for example -the pH optimum of 6.5 for apple, pH 7.0
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
0 0.5 1 1.5 2 2.5
Dia
met
er(c
m)
Log enzyme activity (U) b)
a)

78
for artichoke and Barbados cherry respectively.[162], [176], [177] .The enzyme activity
gradually decreased as the pH was shifted either to extreme of alkaline or acidic region. The
enzyme was inactivated at pH 2 but maintained its activity at pH 8; showing relative activity
of 30.57%. Generally, vegetables and fruits show maximum activity at or near neutral pH
values, but these values may vary with the source of enzyme and substrate within a relatively
wide range of pH.
Figure 5.7: Optimal pH requirement as well as effect on stability of enzyme on incubation for 24h at different
pH
Variation in optimum pH of PPO from different sources affected by factors such as cultivar,
nature of phenolic substrate, type of buffer and ionic strength of buffer, purity of enzyme,
extraction method and isoenzyme form [178], [179]. The stability of PPO was measured by
incubating the enzyme in different pH conditions at 4°C for 24 h. The pH stability of an
enzyme is considered as an important parameter for the determination of the conditions that
should be available through isolation, purification, handling and storage of the enzyme.
Protein structure of an enzyme molecule is influenced by the acidity or alkalinity of the
solution due to pH change as its various amino acid residues are in different states of
ionization. As shown in (Figure 5.7) enzyme from A. paeoniifolius when incubated at pH 5.0
retained more than 80% of its original activity. Maximum activity was found at pH 6.
However, the activity reduced to less than 45% of its original activity when incubated at pH
0.00
20.00
40.00
60.00
80.00
100.00
120.00
0 1 2 3 4 5 6 7 8 9 10
Re
lati
ve e
nzy
me
act
ivit
y %
pH
pH optimum
pH stability
79
below 5.0 and retained 20% above 7.0. Therefore, acidic and alkaline conditions can be used
as a potential measure to control browning reaction in Amorphophallus corm.
5.3.2.3 Temperature optimum for PPO
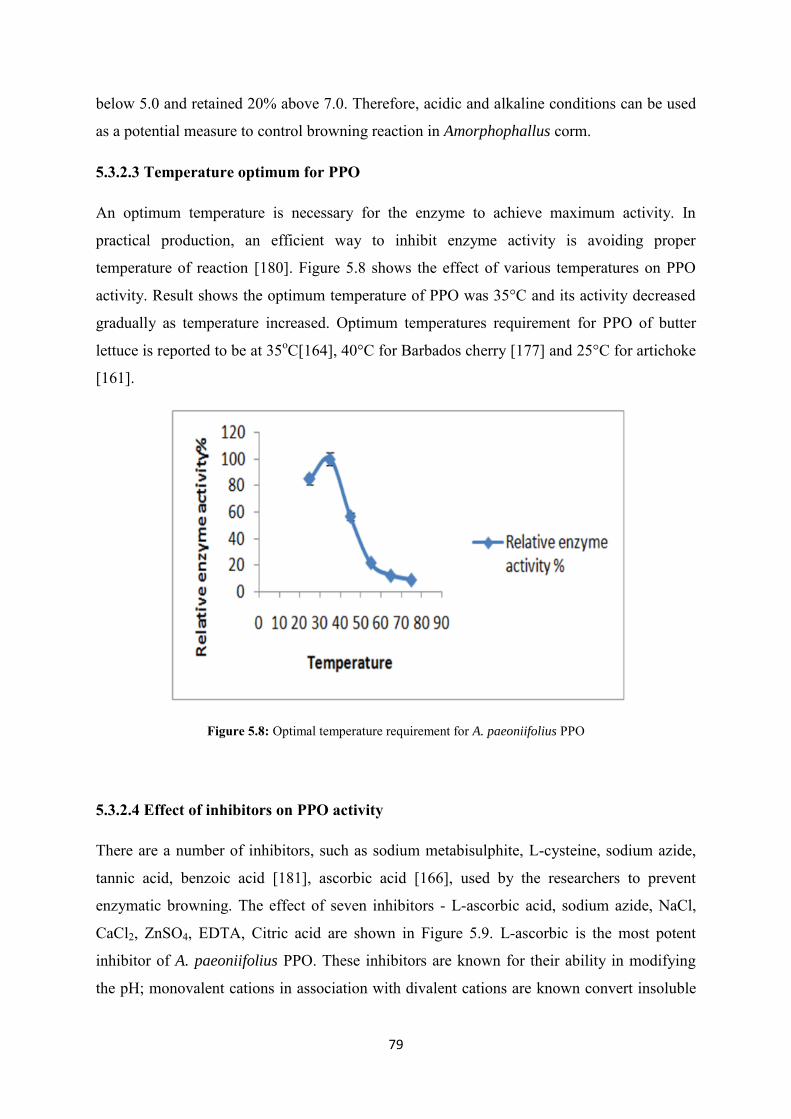
An optimum temperature is necessary for the enzyme to achieve maximum activity. In
practical production, an efficient way to inhibit enzyme activity is avoiding proper
temperature of reaction [180]. Figure 5.8 shows the effect of various temperatures on PPO
activity. Result shows the optimum temperature of PPO was 35°C and its activity decreased
gradually as temperature increased. Optimum temperatures requirement for PPO of butter
lettuce is reported to be at 35oC[164], 40°C for Barbados cherry [177] and 25°C for artichoke
[161].
Figure 5.8: Optimal temperature requirement for A. paeoniifolius PPO
5.3.2.4 Effect of inhibitors on PPO activity
There are a number of inhibitors, such as sodium metabisulphite, L-cysteine, sodium azide,
tannic acid, benzoic acid [181], ascorbic acid [166], used by the researchers to prevent
enzymatic browning. The effect of seven inhibitors - L-ascorbic acid, sodium azide, NaCl,
CaCl2, ZnSO4, EDTA, Citric acid are shown in Figure 5.9. L-ascorbic is the most potent
inhibitor of A. paeoniifolius PPO. These inhibitors are known for their ability in modifying
the pH; monovalent cations in association with divalent cations are known convert insoluble

80
pectins to soluble pectins in the middle lamella of cells[182]. Ascorbate acts as an antioxidant
also by reducing the initial quinine formed by the enzyme to the original diphenol before it
undergoes secondary reactions which lead to browning.
Figure 5.9: Effect of inhibitors (100mM) on enzyme activity respectively
5.3.2.5 Molecular weight determination of Amorphophallus corm PPO
SDS PAGE and the semi native SDS PAGE ( protein sample was diluted with sample buffer
devoid of β-mercaptoethanol and loaded without heat treatment) revealed that the
approximate molecular weight of PPO to be 40 kDa. Figure 5.10(a) depicts the SDS PAGE of
the samples. Figure 5.10(b) shows the electrophorectic blot in which the native gel was kept
on dry catechol paper along with the glassplate for 5 minutes. Single dark brown band was
clearly visible in all the 3 lanes instantly. These bands confirm the presence of PPO in all the
three fractions (C; crude enzyme extract, A; acetone precipitated fraction, U; unbound
fraction of DEAE). Molecular weight of the enzyme can also be estimated very accurately by
this blot zymography studies. Polyphenol oxidase molecular weight ranges between 45 to 67
kDa as reported by many authors [183], [184]

81
a) b)
Figure 5.10: Detection of PPO activity by SDS PAGE(a) and electrophoretic-blot technique.b).
b)Single dark brown band was clearly visible in all the 3 lanes (C, Crude enzyme extract; A, Acetone
precipiated fraction; U, unbound fraction obtained after DEAE ion exchanger) depicts that the molecular weight
of A. paeoniifolius polyphenol oxidase is 40 kDa .The native gel was kept on dry catechol paper along with the
glassplate for 5 minutes.
5.3.2.6 Application in dough rising
Previous studies have reported that polyphenoloxidase affects the dough matrices by
affixing dough enhancement additives to the bread dough, exerting an oxidizing effect to the
dough, strengthening gluten bonds, increasing volume, reducing stickiness which results in
improved freshness of the bread texture, softness of the baked product, improved crumb
structure, flavour and the improved machinability[128]. Role of crosslinking of esterified
ferulic acid on the arabinoxylan portion of the dough also brings about softening
phenomenon as there is breakdown of the cross-linked arabinoxylan network [184].

82
Figure 5.11 Application of crude PPO for dough rising in bakery. ‘A’ indicates control (without addition of
PPO) ‘B’ and ‘C’ shows the PPO treated dough samples with 1ml in duplicate. Bars indicate the dough-raising
in control and PPO treated dough samples. (0.8 cm more rise in height of dough)
Dough raising capability of the crude extract of A. paeoniifolius was monitored at regular
intervals. Maximum dough rising takes about 8 hrs after which there was no increase in
height of the risen dough. Crude enzyme treated dough showed better dough rising property
with a difference of 0.8 cm between control, and enzyme treated dough (Figure 5.11). A.
paeoniifolius PPO may be affecting the different phenolics existing in dough and its
association with biomolecules in flour can be accounted for the observed changes which need
to be investigated further.
5.4 CONCLUSIONS
Researches on cellulases and Polyphenol oxidase have progressed over some time. The
availability of enzymes from plant species for use in industrial processing needs remains a
feasible option as most of the industrial enzyme processes are run at high temperatures.
Crude enzyme fraction from stored corm of Amorphophallus paeoniifolius having cellulase
activity could clarify apple juices and the clear juice obtained indicated the potential for use
of crude extract of Amorphophallus in fruit juice processing. The use of cellulase from
locally produced corms having good shelf life value can be favourable and economical for

83
juice production. It is well known that for clarification of juices by cellulase, xylanases,
pectinases are utilized [185]. The availability of enzymes from plant species remains a best
viable option which needs to be further explored. The stored corm extract showed the
existence of cellulolytic activity with enhanced thermo stability and was stable under both
acidic as well as alkaline conditions. In both substrates (Filter Paper and CMC), the enzyme
extract showed two activity peaks because of dip in activity at pH 7.0. This is due to substrate
heterogeneity, such as the substrate portions degraded easily are hydrolysed first, end product
inhibition, thermal inactivation and irreversible adsorption of part of enzyme by the substrate.
This enzyme has potential to be exploited in the clarification of juices [186].
Enzymatic browning is a biochemical process which occurs in fruits and vegetables by the
enzyme PPO, resulting in the darkening and discoloration of vegetables. PPO activity causes
considerable economic and nutritional loss in the commercial production of fruits and
vegetables. To our knowledge no report is available on the A. paeoniifolius, browning
enzyme PPO. The present study deals with partial purification, characterization, inhibition
and dough rising application of PPO for the first time from A. paeoniifolius. It was shown
that Amorphophallus corm PPO has maximum substrate specificity for catechol. PPO activity
was quite sensitive to some of the common PPO inhibitors, especially to L ascorbic acid,
Citric acid, ZnSO4. This study led to the conclusion that enzymatic browning of A.
paeoniifolius corm might be minimized by addition of these inhibitor solutions. Acidic and
alkaline above pH 7 conditions can be used as a potential measure to control browning
reaction. A. paeoniifolius PPO enzyme will find application in dough raising and baking
industry where stability of enzyme at high temperature would be economical. Also it is to be
noted that further purification steps for the enzyme may be required to make this a good
candidate for some industrial or biotechnical applications.

84


















