
Chapter-2 Review of...
36
Chapter-2 Review of Literature
Transcript of Chapter-2 Review of...

Chapter-2 Review of Literature

Chapter-2 Review of Literature
6
2.1. Plastic waste: An environmental issue
With increasing rate of urbanization in India, there is an increasing demand of plastic.
The growth of plastic industry in India is quite phenomenal having a growth rate of
17% which is higher than elsewhere in the world. This increasing demand and
abilities of plastic to fulfill these demands at cheaper price has caused an increase in
consumption of plastic in last few years. Today almost all the available plastics are
manufactured synthetically. They have much versatile qualities than naturally
occurring polymers i.e. strength, lightness, durability, resistance to degradation,
processability into wide range of shapes and strength. Increasing consumption of
plastic is having significant impact both visible and invisible on environment which
becomes more severe with no proper disposal strategies available. More than 100
million tones of plastic are produced annually worldwide and the major portion is
discarded into landfills annually. These landfills do not present a proper solution to
their disposal problem and themselves are becoming problem to municipality
worldwide as they lose capacity because of the accumulation of synthetic plastics.
Disposal of synthetic plastic is also threatening the natural terrestrial, marine
environment, soil fertility, depletion of underground water. The situation is more
acute in countries such as India where economic growth as well as urbanization is
quite rapid. In India 36.5 million tones/year of municipal solid waste is generated and
plastic waste amounting to the total 2-4 million tones/years. Mounds of plastic
polyethene bags strewn across roadsides are a familiar sight in urban India. It is
worrisome to think it would take one million years for its degradation. Recycling of
the plastic can be done as a solution but it is very tedious and expensive. The major
task is the sorting of wide variety of discarded plastic which is very time consuming.
Also, the presence of a wide variety of additives such as pigments, coatings, fillers
limits the use of the recycled material. Moreover, there are certain issues related with
the plastic industry and recycling of the plastic which need immediate attention.
These issues are related to the health and hygiene of the workers involved in the
processing trade, upgrading of processing equipment used in recycling, quality of the
effluent from the recycling plant and finally the quality of the product from recycled
plastic wastes. Given this scenario of intensive use of synthetic plastics, it is crucial

Chapter-2 Review of Literature
7
for India to check the use of plastic. The existing policies have not been able to
provide any respite against littering and its associated problem. Therefore, there is an
urgent need to adopt policies that can help in establishing an efficient waste
management process and ensure efficient resources use in country.
In search of sustainable product development there has been a lot of research to find
different ways of using energy efficient, non-toxic, renewable sources rather than
finite sources. It has been realized that the solution to the problem of ‘polymer
garbage’ is the production and replacement of non-biodegradable by a wide range of
polymers that are degradable under varying environmental conditions.
2.2. Bioplastic
Bioplastics are the most promising alternative that can change the scenario of plastic
waste management. These are an important class of biomaterial consisting of
polyesters that are widely distributed in nature. These have physio-chemical
properties resembling that of synthetic plastic but have an advantage of being
biodegradable under different environmental conditions. There are three types of
biodegradable plastic reported:
1. Photodegradable: Photodegradable plastic, having light sensitive groups
incorporated into the backbone of the polymer as additives, can be disintegrated
by using extensive UV radiations. This makes them susceptible for further
bacterial degradation (Kalia et al., 2000a).
2. Semi-biodegradable: These are starch linked plastics where starch is
incorporated to hold together short fragments of PE. Starch in the plastic is
degraded by bacteria but bacteria are turned off by PE fragments which thereby
remain non-biodegradable (Tang and Alavi, 2011).
3. Completely biodegradable: These plastics are the most promising because of its
actual and complete degradation by bacteria e.g. PHA, PLA, aliphatic polyesters. Biodegradable polymers, such as cellophane, polylactic acid and PHAs (PHAs) have
assumed increasing importance in last few years. Out of these, PHA polymers are of
prime interest as these can be produced by bacterial fermentation using renewable

Chapter-2 Review of Literature
8
sources. In contrast, in others either the monomer composing the polymers are derived
from the fossil sources or if produced naturally like cellulose and lactic acid further
polymerization is needed under biotechnological process. PHAs are synthesized by
bacteria as energy and food reserve material in the form of granules up to the level of
90% of total Cell Dry Weight (cdw) and can be recovered directly using various
methods.
Product applications for bioplastics largely will depend on its material properties like
its strength, life span, resistance to heat and water, the ability to be used in packaging
and its cost. However, there are certain issues which are limiting the large scale
production and commercialization of bioplastic. The improper implementation of
legislative measurements and high production cost are major issues. Policies are not
effectively implemented to enhance the use of biodegradable polymers in India e.g.
for export of products in certain countries the mandatory condition of using
Biodegradable Plastics for packaging is enforced.
Expensive production process in mainly attributed to the substrate cost and efficiency
of downstream processes depending on polymer content, productivity (PHA/unit
volume/unit time), cost of maintaining an axenic, sterile conditions and recovery
processes (Chen, 2009). The cost of PHA production is around 20 times higher than
polypropylene. Using renewable cheap sources and recombinant E. coli as PHA
producer can reduce the price comparable to other biodegradable plastics such as PLA
and aliphatic polyesters but the price still remains higher than that of petroleum based
plastic. Another major factor limiting PHAs extensive use is the brittle nature of
homopolymers. Copolymers are known to attribute more strength and flexibility of
polymers and can be produced by feeding co-monomer precursor substrates.
Exploitation of bacterial strains for their ability to use renewable sources like
biowaste as substrate to accumulate PHA copolymers is the most promising strategy.
2.3. Polyhydroxyalkanoates (PHAs)
PHAs are the polyesters of hydroxyacids hydroxylated at positions 3, 4, 5 and 6, all of
which are (R)-form chiral molecules. These are accumulated by various microbes
under nutrient (N, P, S, O and Mg) limiting and excess carbon source conditions, in

Chapter-2 Review of Literature
9
the form of storage granules as sink for carbon and reducing equivalent (Reddy et al.,
2003; Singh et al., 2009; Koller et al., 2010; Rehm, 2010). Figure 2.1a represents the
general structural formula (Keshavarz and Roy, 2010). Depending on the number of
carbon atoms in the monomer unit, PHAs can be divided into three groups: 1) Short
chain length PHAs (SCL-PHAs), which consist of 3-5 carbon atoms (Figure 2.1 b, c).
PHB, the first PHA to be discovered in B. megaterium by Lemoigne in 1923, is the
most widely studied and best characterized PHA. It has a perfectly isotactic structure
with only the (R)-configuration; 2) Medium chain length PHAs (MCL-PHAs), which
consist of 6-14 carbon atoms, and 3) Long chain length PHAs (LCL-PHAs), which
consist of 17 and 18 carbon atoms. This division of polymers into groups is based on
the substrate specificity of PHA synthases that can only accept certain
hydroxyalkanoic acids in course of polymerization that depends on strain and culture
conditions (Keshavarz and Roy, 2010).
a.
b.
c.
Figure 2.1: Structure of PHAs: a) General formula; b) P(3HB-co-3HV) and c) P(3HB-co-4HB)
PHAs have physical properties similar to petroleum based synthetic plastics and are
degradable under both aerobic and anaerobic environment. The physical properties of
PHA granules are quite distinguished between two physical states in which PHA
occurs. In the prokaryotes, PHA occurs either as inclusion bodies or as complexes of
Ca+2 and polyphosphates in the cytoplasmic membranes. In the cytoplasm of bacteria,

Chapter-2 Review of Literature
10
the polymer aggregates to form a spherical inclusion or granule of usually 50-500 nm
in diameter. It has a amorphous hydrophobic PHA polyester at the core forming
rubbery state and is surrounded with a layer consisting of phospholipids with attached
or embedded granule-associated proteins at the surface including the PHA synthase,
PHA depolymerases, structural, and regulatory proteins (Potter and Steinbuchel,
2005; Potter et al., 2005; Grage et al., 2009). The hydrophobic polyester core is with
water as a component that prevents crystallization by acting as a plasticizer. This is
the mobile state of PHA, that is, the form that is subject to the action of synthesizing
and degrading enzymes. During extraction of granules from the cell, the phospholipid
and protein layer is damaged or lost. After isolation, extracellular PHA granule is
often crystalline having about 50-80% of crystallinity (Tsuge, 2002; Grage et al.,
2009). The molecular weight of these compounds range from 2×105 to 3×106 Da. The
densities of crystalline and amorphous PHB are 1.26 and 1.18 g/cm3, respectively.
The Mw of PHB produced from wild-type bacteria is usually in the range of 1×103 to
3×106 g/mol. Thus, PHAs have sufficiently high molecular mass to have polymer
characteristics that are similar to conventional plastics such as polypropylene
(Madison and Huisman, 1999; Tsuge, 2002).
A few PHAs, such as PHB and copolymers of 3HB, 3HV and/or 4HB are produced
by various industries (Biocycle, Biomer, Biopol, Enmat, Mirel, and Nodax) (Chen,
2009). The history of commercialized PHAs goes back to 1959. W. R. Grace and
Company produced PHB in the U.S. for possible commercial applications. However,
the company shut down the process due to low production efficiency and a lack of
suitable purification methods. In 1970, P(3HB-co-3HV) was commercialized by
Imperial Chemical Industries Ltd. (ICI/Zeneca BioProducts, Bellingham, UK) under
the trade name of Biopol™ (Chen, 2009; Chanprateep, 2010; Meyer, 2011). In 1996,
the technology was sold to Monsanto and then to Metabolix, Inc. Now a days, a
number of different companies are developing PHAs for use in plastics worldwide,
e.g. Kaneka in Japan, and P&G Chemical, BP and Metabolix (DeMacro, 2005; Noda
et al., 2005) in the US and Imperial Chemical Industries in the UK (Khanna and
Srivastava, 2005). Kaneka and P&G Chemical have teamed up to commercialize a
product called Nodax (also known as Nodak™) which is a specialized PHA. Nodak

Chapter-2 Review of Literature
11
™ has already been made into a variety of different prototype objects such as plastic
fiber or twine and molded plasticware such as plates and cups (Noda et al., 2005).
Metabolix is already producing preliminary PHA materials, but is teaming up with BP
for two years to produce bioplastics.
2.4. PHA biosynthesis
PHAs are synthesized by diverting intermediates of carbon metabolism to hydroxyacyl-
CoA thioesters. Pathways for PHA synthesis consist of three enzymatic reactions
catalyzed by successive action of β-Ketoacyl-CoA thiolase (phbA), acetoacetyl CoA
(phbB) reductase, PHA polymerase (phbC) (Madison and Huisman, 1999). PHA
synthase or polymerase is the key enzyme that in various pathways catalyses the
committed step of polymerizing (R)-3-hydroxyacyl-CoA thioester monomers into
polyester with the release of CoA. The nucleotide sequences of 59 PHA synthase genes
have been obtained from 45 different bacteria (Rehm 2003). The multiple alignments of
the primary structures of PHA synthases show an overall identity of 8-96% with only
eight strictly conserved amino acid residues (Rehm, 2003). Depending upon the
primary structures deduced from these sequences, the substrate specificities of the
enzymes and the subunit composition, PHA synthases differ in different organisms
(Rehm, 2006; Valappil et al., 2007a). Four major classes of PHA synthases can be
distinguished (Table 2.1). Class I and class II PHA synthases comprise enzymes
consisting of only one type of subunit (PhaC) with molecular masses between 61 kDa
and 73 kDa. Class I PHA synthases preferably utilize CoA thioesters of various (R)-3-
hydroxy fatty acids having 3 to 5 carbon atoms, whereas class II PHA synthases
preferentially utilize CoA thioester of various (R)-3-hydroxy fatty acids containing 6 to
14 carbon atoms. In class III PHA synthases two different types of subunits are present:
(i) the PhaC subunit (molecular mass of approx. 40 kDa) showing amino acid sequence
similarity of 21-8% to class I and II PHA synthases and (ii) the PhaE subunit (molecular
mass of approx 40 kDa) with no similarity to PHA synthases. These PHA synthases
prefer CoA thioesters of (R)-3-hydroxy fatty acids comprising 3 to 5 carbon atoms.
Class IV PHA synthases resemble the class III PHA synthases, but PhaE is replaced by
PhaR (molecular mass of approx. 20 kDa) which is different from the transcriptional

Chapter-2 Review of Literature
12
regulator PhaR in R. eutropha. The PHA synthases of R. eutropha, Pseudomonas
aeruginosa, Allochromatium vinosum and B. megaterium represent Classes I, II, III, and
IV respectively (Rehm 2003; Naik et al., 2008).
Table 2.1: Four classes of PHA synthases (PhaC) (Rehm, 2006)
All PHA synthases share a conserved cysteine (Cys319 in R. eutropha PHA synthase)
as a catalytic active site to which the growing PHA chain is covalently attached. The
active-site cysteine, along with other conserved amino acids (histidine and aspartate),
constitutes a catalytic triad similar to esterases. Comparison of PHA synthase amino
acids sequences with sequences of esterases with known structures strongly suggests
that PHA synthases have an α/β-hydrolase fold.
Pathways
There are different pathways for PHA synthesis varying among different type of
organism (Suriyamongkol et al., 2007).
2.4.1. Biosynthetic pathway in R. eutropha
Most of the organisms synthesize PHA using this pathway, well established in R.
eutropha, Zoogloea ramigera, and Azotobacter beijerinckii. In R. eutropha,
metabolism of carbohydrates leads to the biosynthesis of PHA (Figure 2.2). β-
Ketothiolase, encoded by phaA, condenses two molecules of acetyl-CoA to
acetoacetyl-CoA which is subsequently reduced to (R)-3-hydroxybutyryl-CoA by the
NADPH-dependent acetoacetyl-CoA reductase, encoded by phaB. The PHB synthase,
encoded by phaC then polymerises the (R)-3-hydroxybutyryl CoA enantiomer into
the polyoxoester PHB (Tsuge, 2002; Naik et al., 2008). When propionic acid is used

Chapter-2 Review of Literature
13
as a substrate acetyl-CoA is formed by the elimination of carbonyl carbon from
propionyl-CoA. Two moles of acetyl-CoA are used to form a HB unit of the
copolymer, while a HV unit is formed by the reaction of acetyl CoA and propionyl-
CoA resulting in final production of copolymer P(HB-co-HV) (Naik et al., 2008).
Figure 2.2: PHA biosynthetic pathway in Ralstonia eutropha (Tsuge 2002; Naik et al., 2008)
2.4.2. Biosynthetic pathway in Rhodopseudomonas rubrum
This pathway is similar to the R. eutropha pathway and found in R. rubrum where β-
oxidation of fatty acid leads to the biosynthesis of PHA (Figure 2.3).
Figure 2.3: PHA biosynthetic pathway from fatty acids oxidation in Rhodopseudomonas
rubrum (Naik et al., 2008)

Chapter-2 Review of Literature
14
The first reaction remains the same as that for R. eutropha leading to the formation of acetoacetyl CoA, which is then reduced into L-(+)- 3-hydroxybutyryl CoA by a NADH dependent reductase. Two enoyl-CoA hydratases are also involved in the second step of catalyzing the conversion of L-3-hydroxybutyryl-CoA to D-3- hydroxybutyryl-CoA via crotonyl-CoA (Tsuge, 2002; Khanna and Srivastava, 2005; Naik et al., 2008). 2.4.3. Biosynthetic Pathway in Pseudomonas group I
This type of PHA biosynthetic pathway is found in most Pseudomonas species, belonging to rDNA homology group-I e.g. P. oleovorans. These organisms produce MCL-PHAs (from C6-C9) from MCL-alkanes, alcohols, or alkanoates using intermediates from fatty acid β-oxidation pathway (Khanna and Srivastava, 2005). SCL-PHAs, i.e., PHB homopolymer and PHB-PHV copolymer, cab be produced by these organisms in small amount i.e. less than 1.5%.
2.4.4. Biosynthetic pathway in Pseudomonas group II
This is the fourth type of PHA biosynthetic pathway found in Pseudomonas belonging to rDNA homology group-II e.g. P. aeruginosa (Figure 2.4). Most Pseudomonads from the rDNA homology group I except P. oleororans also produce MCL PHAs using this pathway. Synthesis of PHA results from de novo fatty acid synthesis pathway which involves the synthesis of copolymers of MCL 3-hydroxyalkanotae (3HA) from acetyl-CoA utilizing unrelated substrates, e.g., gluconate or acetate (Tsuge, 2002; Naik et al., 2008).
Figure 2.4: PHA biosynthetic pathway from “de novo fatty acid synthesis” in Pseudomonas
group II (Naik et al., 2008)

Chapter-2 Review of Literature
15
2.5. Homopolymer vs Copolymer
Approximately 150 different PHA monomers have been characterized. Besides 3-, 4-,
5-, and 6-hydroxyalkanoates, different functionalized hydroxyalkanoates such as
those with halogenated and aromatic side chains have been described as constituents
of PHAs (Suriyamongkol et al., 2007). The crystalline homopolymer PHB melts in
the range of 170-180 °C and have a glass transition temperature around 4 °C. PHB in
the amorphous fraction is characterized by the same glass transition temperature as
PHB in native granules (Tg ≈ 0 °C). The crystalline behavior of the polymer (55-80%
crystallanity) results in fairly stiff and brittle materials, somewhat limiting its
applications and processability (e.g. elongation to break is about 2-10% compared to
up to 400% for some polyolefins). The tensile strength of PHB (43 MPa) is close to
that of polypropylene (38 MPa) (Potter and Steinbuchel, 2005; Albuquerque et al.,
2011). It is well established that PHA’s thermal and mechanical properties depend
directly on the polymer composition and structure. In contrast to homopolymer PHB,
copolymers are more ductile, tensile, easier to mold with increased flexibility (higher
elongation to break), having more favorable thermoplastic properties, better film
forming capacity and mechanical properties similar to low-density polyetylene. These
features improve their strength and processability (Tsuge, 2002; Valappil et al.,
2007b; Lee et al., 2008; Albuquerque et al., 2011). The incorporation of different
monomer types reduces polymer crystallinity by disturbing the crystal lattice. There
are studies reporting that melting and glass transition temperature of P(3HB-co-3HV)
steadily decrease from 0 to 30 mol% fraction of 3HV (Lee et al., 2008; Allen et al.,
2010; Bengtsson et al., 2010a). The lower melting temperatures of these copolymers
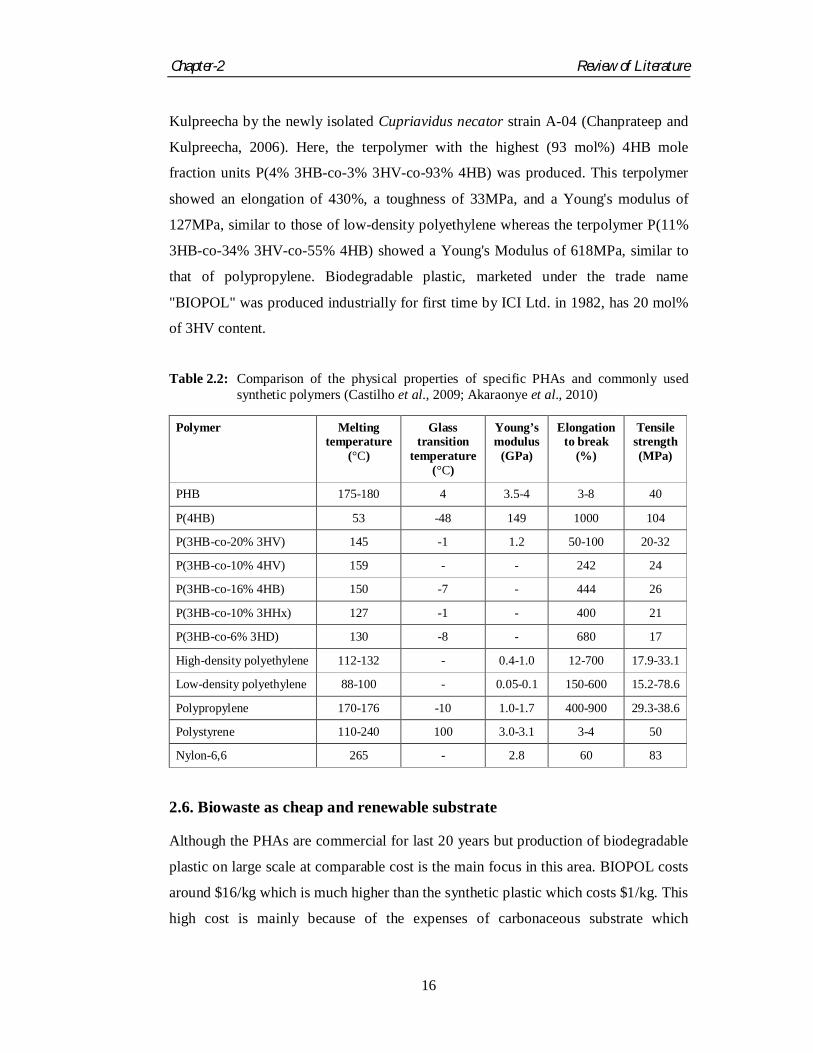
allow processability for a wider temperature range. Table 2.2 summarizes the physical
properties of commonly used PHAs and synthetic polymers.
Copolymer production can be achieved by feeding copolymer precursors substrates
where their composition can be varied with different feeding strategies (Valappil et
al., 2007b; Singh and Mallick, 2008). The use of continuous feeding was observed to
increase the HV content relatively in comparison to pulse wise feeding (Albuquerque
et al., 2011; Hafuka et al., 2011). There have been reports on the production of the
terpolymer P(3HB-co-3HV-co-4HB) reported for the first time by Chanprateep and
Chapter-2 Review of Literature
16
Kulpreecha by the newly isolated Cupriavidus necator strain A-04 (Chanprateep and
Kulpreecha, 2006). Here, the terpolymer with the highest (93 mol%) 4HB mole
fraction units P(4% 3HB-co-3% 3HV-co-93% 4HB) was produced. This terpolymer
showed an elongation of 430%, a toughness of 33MPa, and a Young's modulus of
127MPa, similar to those of low-density polyethylene whereas the terpolymer P(11%
3HB-co-34% 3HV-co-55% 4HB) showed a Young's Modulus of 618MPa, similar to
that of polypropylene. Biodegradable plastic, marketed under the trade name
"BIOPOL" was produced industrially for first time by ICI Ltd. in 1982, has 20 mol%
of 3HV content.
Table 2.2: Comparison of the physical properties of specific PHAs and commonly used
synthetic polymers (Castilho et al., 2009; Akaraonye et al., 2010)
Polymer Melting temperature
(°C)
Glass transition
temperature (°C)
Young’s modulus
(GPa)
Elongation to break
(%)
Tensile strength (MPa)
PHB 175-180 4 3.5-4 3-8 40
P(4HB) 53 -48 149 1000 104
P(3HB-co-20% 3HV) 145 -1 1.2 50-100 20-32
P(3HB-co-10% 4HV) 159 - - 242 24
P(3HB-co-16% 4HB) 150 -7 - 444 26
P(3HB-co-10% 3HHx) 127 -1 - 400 21
P(3HB-co-6% 3HD) 130 -8 - 680 17
High-density polyethylene 112-132 - 0.4-1.0 12-700 17.9-33.1
Low-density polyethylene 88-100 - 0.05-0.1 150-600 15.2-78.6
Polypropylene 170-176 -10 1.0-1.7 400-900 29.3-38.6
Polystyrene 110-240 100 3.0-3.1 3-4 50
Nylon-6,6 265 - 2.8 60 83
2.6. Biowaste as cheap and renewable substrate
Although the PHAs are commercial for last 20 years but production of biodegradable
plastic on large scale at comparable cost is the main focus in this area. BIOPOL costs
around $16/kg which is much higher than the synthetic plastic which costs $1/kg. This
high cost is mainly because of the expenses of carbonaceous substrate which

Chapter-2 Review of Literature
17
contribute around 45% of the total production cost (Reddy et al., 2003; Kumar et al.,
2009). Use of renewable sources like biowaste as cheap carbon source can reduce the
cost attributed to substrate as well as can also solve another problem related to the
handling and treatment of large amount of waste.
2.6.1. Waste management
Waste management is becoming political priority in many developed and developing
countries. Waste management methods used today commercially include: i)
transportation of waste to low lying areas/landfills, ii) burning of waste on site or in
the incinerators, iii) composting, iv) briquetting, (v) recycling of waste matter and vi)
microbial treatment (aerobic and anaerobic) etc. As each of these methods has its own
advantages and can be employed to certain types of waste an integrated approach is
required for complete disposal of waste (Talyan et al., 2007). Disposal of wastes in
unorganized landfills causes environmental pollution due to slow and uncontrolled
fermentation, which generates gases like CO2, CH4 and H2S. Natural degradation in
landfills is very slow continued over scores of years and only 10-15% of the available
energy content can be recovered. Burning of wastes on site is a conventional method
of disposing wastes. Out of the 4000-5000 tons of waste generated per day in
metropolitan cities about 60-70% is collected and discarded into landfills. Burning is
possible only for dry wastes and these wastes have up to 65% of materials, which can
be combusted. However, it results in generation heat at the rate of around 2700
BTU/kg waste (1 BTU, British thermal unit = 2.9x 10-4 KWH) (Kalia, 1991) and
obnoxious gases affects the environment and human health. Composting is the highest
form of recycling. Composting is inexpensive, rapidly implemented and a publicly
acceptable treatment process that recycle the organic matter to be reused in beneficial
manner. Briquetting process consists of heating the feedstock’s under controlled
conditions, without any contact with air and converts highly voluminous and
troublesome waste materials in to clean, non-polluting fuel. Being compact,
transportation and storage costs are reduced. It is environmentally superior technology
for handling different ligno-cellulosic materials (Kalia, 1992, 2007, Kalia et al.,
1992a, b; 1995; McDowall and Eames, 2007).

Chapter-2 Review of Literature
18
2.6.2. Anaerobic digestion
The fermentation technology is the best technology for treatment and recycling of
organic wastes being more hygienic and convenient process than other means like
briquetting, burning, composting and landfilling. Anaerobic digestion (AD) process is
one of the oldest processes used for the treatment stability of waste materials. AD is a
series of processes in which microorganisms break down biodegradable material in
the absence of O2. Unlike uncontrolled and slow fermentation of wastes in a landfill,
biodegradation is using AD system results in energy generation and production of
nutrient rich biomanure providing a better approach for waste recycling and
stabilisation. The intermediate of this process are primarily converted to H2 and
finally in to CH4 and CO2 during methanogenesis (Kalia and Luthra, 1994; Kalia,
1995; Kumar et al., 1995; Kalia, 2007; Chen et al., 2008; Holm-Nielsen et al., 2009).
2.6.3. Steps involved in anaerobic digestion
There are four key stages of AD process:
(i) Hydrolysis
Through this first step of AD the complex organic molecules are broken down into
simple sugars, amino acids and fatty acids. Extracellular enzymes such as such as
amylase, cellulase, lipase and protease, which are excreted by hydrolytic fermentative
bacteria catalyse this hydrolytic process which provide substrates to next group of
bacteria (Kalia et al., 2000a, b).
(ii) Acidogenesis
The biological process of acidogenesis leads to further breakdown of the remaining
components by acidogenic (fermentative) bacteria. This step is fastest in the AD
processes with acidogenic bacteria having with minimum doubling time of around 30
min. Here, low molecular weight compounds such VFAs are created along with NH3,
CO2 and H2 well as other byproducts.

Chapter-2 Review of Literature
19
(iii) Acetogenesis
In the third stage AD simple molecules created through the acidogenesis phase are
further digested by acetogens to produce largely acetic acid as well as CO2 and H2.
The acetogens are slow growing, doubling time of around 1.4 days and among them
more co-operations are needed for efficiency.
(iv) Methanogenesis
Biological process of methanogenesis constitutes the last step where methanogens
catabolize acetate and a mixture of H2 + CO2 to CH4. Methanogens are unique among
prokaryotes and are classified as members of archaebacteria, a group of
phylogenetically different organisms.
All these processes occur simultaneously and synergistically, the first group has to
perform its metabolic action before the next stage bacterial group take over and so
forth. AD of animal manure offers several environmental, agricultural and socio-
economic benefits throughout production of biogas as clean and renewable fuels
(Holm-Nielsen et al., 2009; Lew et al., 2009). Biological oxygen demand (BOD) and
chemical oxygen demand (COD) are reduced by these pathways.
Figure 2.5: Strategy for efficient degradation of biological wastes (Kalia and Purohit, 2008)

Chapter-2 Review of Literature
20
2.6.4. Anaerobic Digestion and PHA production
AD is generally accepted to be effective and economical method for treatment of
waste treatment in which pollution control and energy recovery can be achieved. At
present anaerobic treatment is successfully implemented for various types of
industrial as well as domestic wastes. Hydrolyzing bacteria solubilize and convert
complex organic waste into simpler and utilizable products (VFAs and other soluble
organic compounds) in acidogenic stage og anaerobic digestion process (Figure 2.5).
These hydrolysis products can be utilized by microbial culture as substrate for
bioproduct formation such as PHA (Sonakya et al., 2001; Sun et al., 2007). Various
agricultural, industrial and food waste are being exploited for this purpose e.g.
extruded rice bran, municipal sludge, palm oil, paper mill waste water, soy molasses,
sugar cane molasses, waste cooking oil, and whey, etc. (Khardenavis et al., 2007;
Bengtsson et al., 2008; Castilho et al., 2009; Ntaikou et al., 2009; Chanprateep,
2010). So, we need to search and exploit microbial diversity for bacteria having
ability to produce PHA in high concentration as well as utilize inexpensive sources as
substrate (Kalia et al., 2003; Kalia and Purohit, 2008). The best PHAs-producing
species should satisfy several demands such as the fast growing population, being
able to utilize cheap carbon and having a high production rate.
2.6.5. Potential substrates for PHA production
To make PHAs production more economical, some researchers have tried to produce
PHAs from inexpensive carbon sources. Among various substrates used for PHA
production starch, methanol, dairy byproducts, agro-industrial byproducts, oily waste
are generally used (Tian et al., 2009; Akaraonye et al., 2010). These are readily
utilized by various bacterial strains belonging to gram negative and gram positive
(Table 2.3 & 2.4) group as potential substrates for economic PHA production.
(i) Starch
Starch is a potential carbon feedstock that can be used for PHA production as it is
quite cheap and is readily available in large amount from plant sources (Chen et al.,
2006; Huang et al., 2006). However, to utilize starch as carbon source in PHA
production it needs to be hydrolyzed first because it is complex in nature (Huang et

Chapter-2 Review of Literature
21
al., 2006; Halami, 2008). P(3HB-co-3HV) production has been reported from
Haloferax mediterranei with enzymatically extruded starch in pH stat fed batch
fermentation process achieving total 1.14 g/L biomass with PHA content of 0.84 g
amounting to 43% of total cdw. Here, the solution feed contained extruded starch/
yeast extract in ratio 1/1.7 g/g (Chen et al., 2006). Halomonas boliviensis produced
1.2 g/L biomass containing 56% of cdw PHB in shake flask cultures when starch
hydrolysate was used. However, under batch fermentation, the PHB content was 35%
of cdw under optimum O2 supply and increased to 41% of cdw under O2 limitation
during active growth but biomass was adversely affected (Quillaguaman et al., 2005).
PHA production has also been reported using B. cereus CFR06 which under batch
culture produced 48% of cdw PHA and 1 g/L biomass (Halami, 2008).
Saccharification hydrolyse the starch into more utilizable sugars that can further be
utilized for PHA production. Saccharified waste potato starch was used as carbon
source where 94 g/L PHB was produced. It was equivalent to 55% of total cdw in fed
batch fermentation with R. eutropha NCIM 5149 (Haas et al., 2008).
(ii) Methanol
Methanol produced by anaerobic digestion of woody material and organic substrates
can be used as cheap carbon source for PHA production. Species belonging to
Methylobacterium are shown to utilize methanol for PHA production.
Methylobacterium sp. GW2 has been reported to produce maximum PHB content (out
of cdw) of 40% utilizing methanol as substrate. The PHB accumulation in this
organism is growth associated thereby removing the need for two-stage fermentation
(Yezza et al., 2006). Homopolymer PHB production was reported from methanol
using a 2 L fed-batch fermentation using Methylobacterium extorquens. Here, a total
biomass of 9 g/L and PHB content (out of cdw) of 30-33% were achieved. However,
the organism was able to accumulate copolymer of P(3HB-co-20-mol% 3HV) when
valerate was added with methanol (Bourque et al., 1992). Mokhtari-Hosseini et al.
carried out PHB production in 5L fermentor achieving total biomass of 15.4 g/L with
PHB content of 62.3% on methanol as substrate using M. extorquens DSMZ 1340
(Mokhtari-Hosseini et al., 2009). Recently, mixed culture has been shown to utilize
methanol as substrate under a non-sterile process and producing PHB to the level of

Chapter-2 Review of Literature
22
0.424 g/L while the biomass was achieved to level 1.54 g/L. Here, optimum PHB was
produced with methanol concentration of 0.15% (v/v) (Dong et al., 2011).
(iii) Agro-industrial waste
A number of agricultural wastes like wheat bran, rice bran, corn starch, molasses, PS and
other agricultural byproducts are shown to be potential substrates for PHA production.
H. boliviensis LC1 was shown to produce 1.08 g/L of PHA equivalent to 33.8% of cdw
on wheat bran hydrolysate in a shaken flask study (Van-Thuoc et al., 2008). Huang et al.
have successfully used inexpensive extruded rice bran and corn starch in PHA
production from H. mediterranei (Huang et al., 2006). Here, PHA concentration and
PHA content of 77.8 g/L and 55.6% of cdw, respectively, were achieved using extruded
rice and extruded corn starch in the ratio of 1: 8 in a pH-stat fed-batch fermentation.
Molasses contain large amount of sugars, sucrose being predominant (appox. 50%) and
therefore can be considered as a potential source for PHA production (Solaiman et al.,
2006; Akaraonye et al., 2011). Mixed bacterial culture have used for PHA production on
sugar cane molasses in a sequencing batch reactor (SBR) operated under aerobic
dynamic feeding (Albuquerque et al., 2010a, b; Bengtsson et al., 2010a, b). Albuquerque
et al. achieved the production of 30% of cdw P(3HB-co-3HV) with a biomass of 3.5 g/L.
Bacterial species belonging to Bacillus were also shown to utilize molasses for PHA
production (Santimano et al., 2009). Further, Solaiman et al. successfully utilized soy
molasses for the production of MCL-PHA P(3HDD-co-3HO-co-3HTD) using P.
corrugate. A cell density of 3.4 g/L and a total PHA content of 5-17% cdw were
achieved when 5% (w/v) soy molasses was added to the E-medium (Solaiman et al.,
2006). Recently, PS were also shown in our laboratory as potential substrate for PHB
production where biowaste slurry was successfully supplemented upto 50% of total feed
(Kumar et al., 2009). Further improvement in using only hydrolyzed PSS as medium
was achieved for successful PHA production (Patel et al., 2011b). Conversion of agro-
industrial waste water into PHB was successfully employed by various researchers
(Khardenavis et al., 2007). Rice grain based spent wash has produced PHB yield upto
67% of cdw while the yield was upto 42.3% with jowar grain based distillery spentwash
(Khardenavis et al., 2007). Mixed cultures enriched under feast and famine conditions
have been used produce PHA from VFAs obtained in fermented paper mill wastewater.

Chapter-2 Review of Literature
23
Here the yield achieved was 0.11 kg PHA/kg COD treated containing 53.69 mol% of
3HV (Bengtsson et al., 2008).
(iv) Dairy byproducts
Whey has been the most extensively for PHA production by various micro-organism.
It represents about 80-90% of the volume of processed milk. Only about half of the
whey produced is converted into useful products such as human and animal feed
while the rest is disposed of as waste causing environmental problems due to its high
oxygen demand. Koller and co-workers have found that hydrolyzed whey can be used
by P. hydrogenovora to accumulate up to 5 g/L biomass containing 1.27 g/L and
12% of cdw PHB monomers (Koller et al., 2008). Recombinant E. coli has been used
extensively for the purpose and a PHB yield of 168 g/L contributing to 87% of cdw
was achieved on whey in feed containing 280 g lactose/L under fed batch culture
(Ahn et al., 2001). Using E. coli K24K, Nikel and collegues achieved 72.9% of cdw
PHB amounting to 51.1 g/L under pH controlled fed-batch fermentation with whey
and corn steep liquor as carbon and nitrogen sources was used PHA production
achieving (Nikel et al., 2006 ). Thermus thermophilus utilized whey as substrate to
produce PHA upto the content of 35% containing 3HV), 3HHp, 3HN and 3HU on
media containing 24% (v/v) whey under nutrient limitation (Pantazaki et al., 2009).
Koller et al., have reported successful PHA production using Hydrogenophaga
pseudoflava on whey using an antibiotic (vancomycin) based strategy against the B.
cereus contamination (Koller et al., 2011b). Thus, the successful use of whey as a
carbon source for PHA production is certainly a step forward towards the reduction of
the cost of PHA production.
(v) Oily waste
Improper disposal of the waste vegetable oil leads to an increase in the biological
oxygen demand (BOD) and chemical oxygen demand (COD), which in turn leads to the
deoxygenation of water, an infiltration into soil sediments and aquifer contamination.
These waste vegetable oils can be utilized as inexpensive carbon sources for PHA
production (Mumtaz et al., 2010). This will not only help in converting waste to a
useful material but also aid in the disposal management of the waste vegetable oil.

Chapter-2 Review of Literature
24
Various researchers have shown PHA production using oily wastes like corn oil, olive
oil, olive oil waste water (alpechin), palm kernel oil and soybean oil. There are various
reports of PHA production from C. necator on PKO (Sudesh et al., 2011). Production
of P(3HB-co-3HV) upto 79% of cdw by C. necator was reported when grown on palm
kernel oil with sodium valerate and propionate as 3HV precursors which ranged from 0-
60 mol% (Bhubalan et al., 2008). Copolymer, P(3HO-co-3HD-co-3HDD) upto 63% of
cdw was achieved using P. guezenneibiovar tikehau when coprah oil was used as the
main carbon substrate (Simon-Coin et al., 2008). Different copolymer (P(3HB-co-
3HHx) with different %mol ratios of 3HHx production has been reported using
recombinant strains harbouring the genes mostly belonging to Aeromonas and
Pseudomonas. P. putida KT2442 strain KTT2 on olive oil waste water (alpechin)
produces a maximum PHA concentration and PHA content of 0.1 g/L and 3.59% of
cdw respectively with a corresponding biomass accumulation of 4.2 g/L (Ribera et al.,
2001). PHA production by Pseudomonas sp. on rapeseed oil (Mozejko et al., 2011) and
P. oleovorans on jutropha carcus oil (Allen et al., 2010) have been reported recently. In
a recent report, C. necator produced PHB on derivatives of rapeseed oil upto 1.2 g/L
with waste frying oil which was higher than that observed with pure oil (0.62 g/L) and
heated oil (0.9 g/L) (Verlinden et al., 2011).
2.7. PHA producing microorganisms
PHAs play an important function in ecosphere. PHA producing microorganisms
represents a widespread phylogenetic diversity and are present in ecosystems where
fluctuating availability of C source and nutriens prevails (Koller et al., 2011a).
Although more than 300 different microorganisms belonging to 90 genera are known
so far to synthesize PHAs (Hazer and Steinbuchel, 2007), a perusal of the capacities
of different microbes to produce homopolymers and copolymers of PHAs reveals that
Alcaligenes, Pseudomonas and Ralstonia belonging to Gram-negative lead this group.
These have abilities to utilize pure substrates, agricultural wastes, oily wastes, dairy
products and CO2 for PHA production at large scale (Singh et al., 2009) (Table 2.3 &
2.4). Recombinant strain of E. coli harboring genes for PHA synthesis have also been
used extensively (Suriyamongkol et al., 2007). Among gram positive group Bacillus
sp. are most commonly used having the potential to utilize wide range of substrates
for PHA production (Table 2.3 & 2.4).

Chapter-2 Review of Literature
25
Table 2.3: Microbial biodiversity of PHA homopolymer producers
Substrate Organism Polyhydroxybutyrate Reference g/L % of cdw
Gram negative group
Glucose
Aeromonasa 0.49 36.8 Chien and Ho, 2008 Azotobacter 3.7 77.3 Myshkina et al., 2008 Comamonas 1.27 53 Lee et al., 2004 Escherichiaa 51.1 72.9 Nikel et al., 2006 Pseudomonas 0.06 5.7 Bertrand et al., 1990 Ralstonia 0.97 84 Nurbas and Kutsal, 2004 Vibrio 0.02 14.2 Chien et al., 2007
Fructose Comamonas 0.05 3.0 Lee et al., 2004 Ralstonia 5.5 19.9 Khanna and Srivastava, 2008
Sucrose Alcaligenes 5.48 67.4 El-sayed et al., 2009 Comamonas 0.04 2.0 Lee et al., 2004 Vibrio 0.17 45.5 Chien et al., 2007
Lactose
Comamonas 0.07 4.0 Lee et al., 2004 Hydrogenophaga 0.05 - Povolo and Casella, 2003 Methylobacterium 1.16 - Nath et al., 2008 Paracoccus 0.03 - Povolo and Casella, 2003 Pseudomonas 0.15b 56.0 Young et al., 1994 Sinorhizobium 0.01 14.0 Povolo and Casella, 2003
Fatty acids (& derivatives)
Brachymonas - 64.0 Shi et al., 2007 Comamonas 0.08 4.0 Lee et al., 2004 Escherichia 1.20 54.3 Chien et al., 2010 Pseudomonas 70.0-80.0 Lopez et al., 2009 Spirulina - 10.0 Jau et al., 2005 Vibrio 0.07 12.1 Chien et al., 2007
Maltose Comamonas 0.02 1.0 Lee et al., 2004 Methanol Methylobacterium 15.4 62.3 Mokhtari-Hosseini et al., 2009
Starch
Aeromonas a 0.60 32.7 Chien and Ho, 2008 Azotobacter 3.2 63.0 Myshkina et al., 2008
Haloferax 6.48 32.4 Lillo and Rodriguez-Valera, 1990
Protomonas - - Suzuki et al., 1986
Glycerol
Escherichia a (batch) 3.52 42.0 Nikel et al., 2008 (fed batch) 10.81 51.0 Methylobacterium 10.5 50.0 Bormann and Roth, 1999 Ralstonia - 70.0 Mothes et al., 2007 Vibrio 0.19 42.8 Chien et al., 2007
Xylose Burkholderia 36.0 60.0 Silva et al., 2004 Pseudomonas 0.41 19.7 Bertrand et al., 1990

Chapter-2 Review of Literature
26
Substrate Organism Polyhydroxybutyrate Reference g/L % of cdw
Agricultural Waste
Alcaligenes - - Wang et al., 2007 Azotobacter 22.0 66.0 Page and Cornish, 1993 Burkholderia 2.73 62.0 Silva et al., 2004 Escherichia a 31.6 80.0 Liu et al., 1998 Haloferax 23.0 27.0 Huang et al., 2006 Klebsiella a - 70.0 Zhang et al., 1994 Pseudomonas 0.94 57.7 Aremu et al., 2010 Ralstonia 94.0 52.5 Haas et al., 2008
Dairy Products
Escherichia a 51.1 72.9 Nikel et al., 2006 Hydrogenophaga 0.02 4.4 Povolo and Casella, 2003 Methylobacterium 3.91 66.0 Nath et al., 2008 Pseudomonas 1.27 12.0 Koller et al., 2008 Sinorhizobium 0.02 3.5 Povolo and Casella, 2003
Oily Waste Ralstonia 0.84b 79.0 Budde et al., 2011
Industrial Waste Azotobacter 2.4 48.2 Sathesh Prabu and
Murugesan, 2010 Burkholderia 2.47 57.4 Alias and Tan, 2005 Pseudomonas 22.0 70.0 Jiang et al., 2008
Gram positive group
Glucose Bacillus
0.43 50.0 Kumar et al., 2009; 0.81 38.0 Valappil et al., 2007b
Streptococcus - - Yuksekdag et al., 2008 Streptomyces 1.5-11.8 Verma et al., 2002
Fructose Bacillus 0.39 0.75 Kumar et al., 2009
Sucrose Bacillus
1.2 48.0 Anil-Kumar et al., 2007; 0.17 35.0 Kumar et al., 2009
Streptococcus - - Yuksekdag et al., 2008
Lactose Lactobacillus - - Yuksekdag et al., 2008 Lactococcus - - Yuksekdag et al., 2008 Streptococcus - - Yuksekdag et al., 2008
Fatty acids (& derivatives) Bacillus 0.45 80.1 Valappil et al., 2007b
Agricultural waste
Bacillus - - Akaraonye et al., 2011
3.3 41.0 Kumar et al., 2009 Staphylococcus - - Wang et al., 2007
Industrial waste
Actinobacillus 2.25 46.5 Son et al., 1996 Bacillus 8.2 51.2 Vijayendra et al., 2007
a: Recombinant strains b: g/g substrate consumed

Chapter-2 Review of Literature
27
Table 2.4: Microbial biodiversity of PHA copolymer producers
Substrate Organism Polyhydroxyalkanoates Reference
g/L % of cdw
Composition
Gram negative group
Glucose Pseudomonas 0.09 25.5 3HB:3HV:3HHD:3HOD Singh and Mallick, 2009
Sucrose Rhizobium 1.62 58 3HB:3HV Anil-Kumar et al., 2007 Sphingomonas 0.99 45 3HB:3HV Anil-Kumar et al., 2007
Fatty acids (& derivatives)
Aeromonasa 0.92 59.4 3HB:3HHx Chien and Ho, 2008 Azotobacter 3.41 75.8 3HB:3HV Myshkina et al., 2010 Comamonas 0.63 25.0 3HB:4HB Lee et al., 2004
Escherichiaa 0.39 44.0 3HB:3HV:3HHx: 3HO:3HD:3HDD Davis et al., 2008
Pseudomonas 0.57 38.0 3HHx:3HO:3HUde :3HNe:3HHpe Hartmann et al., 2006
Ralstonia 0.13-0.56 <84 3HB:3HV Nurbas and Kutsal, 2004
Methanol Methylobacterium 2.98 33.0 3HB:3HV Bourque et al., 1992
Agricultural waste
Haloferax 77.8 55.6 3HB:3HV Huang et al., 2006 Klebsiella - - 3HB:3HV Wang et al., 2007
Pseudomonas 0.22-0.25
7.0-17.0
3HHx:3HO:3HD: 3HDD:3HDde: 3HTD:3HTde
Solaiman et al., 2006
Rhizobium 0.71 31.0 3HB:3HV Anil-Kumar et al., 2007 Sphingomonas 0.44 22.0 3HB:3HV Anil-Kumar et al., 2007
Dairy products
Haloferax 14.7 87.5 3HB:3HV:4HB Koller et al., 2007 Pseudomonas 1.44 12.0 3HB:3HV Koller et al., 2008 Ralstonia 0.98 20.6 3HB:3HV Marangoni et al., 2002
Oily waste
Comamonas - 87.5 3HHx:3HO:3HD: 3HDD:3HTDa
Thakor et al., 2005
Pseudomonas - 26.06 3HB:3HV Allen et al., 2010
Ralstoniaa 20.0 95.0 3HB:4HB Park and Kim, 2011
Industrial waste Azotobacter 5.48 58.0 3HB:3HV Ryu et al., 2008
Gram positive group
Glucose Bacillus 0.28 32.9 3HB:3HHx Tajima et al., 2003 Microlunatus 1.41 26.0 3HB:3HV Akar et al., 2006
Fructose Bacillus 0.50 40.3 3HB:4HB Valappil et al., 2007b Sucrose Bacillus 0.64 38.4 3HB:4HB Valappil et al., 2007b
Fatty acids Bacillus 0.23 47.4 3HB:3HV Valappil et al., 2007b 0.33 64.5 3HB:3HV:4HB:3HHx Tajima et al., 2003
Agricultural waste Bacillus 2.0 54.0 3HB:3HV Anil-Kumar et al., 2007
Industrial waste Rhodococcus 0.03 2.4 3HB:3HV Fuchtenbusch and
Steinbuchel, 1999
a: Recombinant strains

Chapter-2 Review of Literature
28
2.7.1. Alcaligens and Ralstonia
Ralstonia and Alcaligenes are versatile organisms with well established abilities to
utilize pure substrates, agricultural wastes, oily wastes, and dairy products for PHA
production. R. eutropha which is most commonly and extensively studied for PHA
production accumulates homopolymers (PHB) to tercoploymers (P(3HB-co-3HV-co-
3HHx) from carbon sources like sugars, organic acids, vegetable oils and CO2 (Tsuge,
2002). There are several other taxonomic names of R. eutropha are found in literature
including Hydrogenomonas eutropha, A. eutrophus, Wautersia eutropha, and C.
necator (Vandamme and Coenye, 2004). Ralstonia has been shown to produce 232
g/L of PHA (82% of cdw) from glucose (Ryu et al., 1997) and up to 96 g/L of PHA
(76% of cdw) from oily wastes (Kahar et al., 2004). Terpolymers P(3HB-co-3HB-co-
3HHx) was reported from mixtures of palm kernel oil and 3HV precursors (Bhubalan
et al., 2008; Bhubalan et al., 2010). Nutritional supplementation with oleic acid
resulted in the improving the physical characteristics of PHB (Grigull et al., 2008).
PHA production by R. eutropha has been exploited in high cell density cultivation
reactor for PHB production on fructose achieving 27.7 g/L biomass with PHB
concentration of 5.5 g/L and productivity of 0.55 g/L/h (Khanna and Srivastava,
2008). Production of P(3HB-co-4HB) was reported by on sugarcane, brown
sugarcane, rock sugar, toddy palm sugar and coconut palm sugar in fed batch cultures.
Here, the mole fraction of 4HB units could be varied from 0 to 94mol% by switching
of the ratios of carbon to nitrogen, together with the ratios of fructose to c-
Hydroxybutyric acid or 1,4-butanediol. The total P(3HB-co-4HB) content was 71% of
cdw with a 4HB unit mole fraction of 30 mol% in the copolymer. The study of
kinetics of P(3HB-co-4HB) production by C. necator strain A-04 has shown that
synthesis of 4HB units was growth-associated under nitrogen-sufficient conditions
and the synthesis of 3HB units was enhanced under nitrogen deficient conditions
(Chanprateep et al., 2008). In further study, upto 112 g and 73 g P(3HB-co-4HB)
were achieved containing 38% mole fraction of 4HB content under optimum
condition (Chanprateep et al., 2010). C. necator was recently reported to produce
PHB on various derivatives of rapeseed oil (Verlinden et al., 2011). By co-feeding
soybean oil and γ-butyrolactone as carbon sources, P(3HB-co-4HB) could be

Chapter-2 Review of Literature
29
produced by R. eutropha KCTC 2662 with cdw of 10-21 g/L, yields of 0.45-0.56 g-
PHA/g-soybean oil used and 4HB fractions of 6-10 mol% (Park and Kim, 2011).
2.7.2. Pseudomonas
Pseudomonas has been widely observed to synthesize MCL-PHA on various aliphatic
alkenes or fatty acids, agricultural and oily wastes (Khardenavis et al., 2007; Jiang et
al., 2008). P. aeruginosa, P. oleovorans, P. resinovorans and other Pseudomonas sp.
have been reported to simultaneously produce C4 to C12 3HA units i.e. SCL-MCL-
PHAs (Singh and Mallick, 2009). Pseudomonas corrugata 388 accumulated MCL-
PHA 5 to 17% of cdw, constituted by 3HO, 3HDD, 3HDde and 3HTde on 2 to 5%
w/v soy molasses (Solaiman et al., 2006). Attempts to utilize agro-industrial oily
wastes like waste frying oil and waste free fatty acid from soybean oil as feed resulted
in 29 to 66% PHA by P. aeruginosa (Fernandez et al., 2005). P. oleovorans can
produce a natural-synthetic hybrid block copolymer polyhydroxyoctanoate-diethylene
glycol which has lesser molecular mass (50%) and reduced polydispersity. It had
similar thermal and crystalline properties as homopolymers (Foster et al., 2005).
Here, polyethylene glycol (PEG106) in 1000:1 ratio affected the polymer yield and
processing.
Recently, homopolymer poly-3-hydroxydecanoate (PHD) and P(3HD-co-84mol%
3HDD) has been reported on decanoic acid or dodecanoic acid by mutant P. puitda
KTQQ20 (Liu et al., 2011). This mutant strain was constructed by knocking out the
genes fatty acid degradation enzymes fadB, fadA, fadB2x, fadAx, 3-hydroxyacyl-
CoA dehydrogenase and acyl-CoA dehydrogenase encoded by the genes PP2136,
PP2137, PP2214, PP2215, PP2047 and PP2048 and also 3-hydroxyacyl-CoA-
acylcarrier protein transferase encoded by PhaG leading to a significant reduction of
fatty acid β-oxidation activity.
2.7.3. Haloferax
Presence of PHA in members of the family Halobacteriaceae of the archaea has also
been shown (Quillaguaman et al., 2010). Although not widely studied, certain
archaeal strains of Haloferax, Halobacterium, Haloarcula and Haloquadratum have

Chapter-2 Review of Literature
30
been reported for their abilities to synthesize PHA from inexpensive carbon sources as
feed material (Han et al., 2007; Huang et al., 2006). phaEC encoding PHA synthase
in H. mediterranei has been reported in various studies to produce P(3HB-co-3HV)
copolymer upto 60% of cdw from starch, glucose, or other cheaper carbon sources,
including industrial by-products (Lillo and Rodriguez-Valera, 1990; Koller et al.,
2007; Lu et al., 2008) and P(3HB-co-11% 3HV) on glucose with PHA content of
48.6% and productivity 0.36 g/L/h (Don et al., 2006). H. mediterranei has high
growth rate, metabolic versatility and genetic stability (Lu et al., 2008). Since H.
mediterranei survives in extreme salinity, it circumvents contamination problem and
consequently greatly reduces the sterility requirement. A study with the same
microorganism on hydrolyzed whey as substrate resulted in P(3HB-co-6% 3HV)
production with yield of 72% and productivity of 0.09g/L/h but P(3HB-co-21.8%
3HV-co-5.1% 4HB) was obtained with yield increased to 87.5% when sodium
valerate and γ-butyrolactone was supplemented into feed (Koller et al., 2007).
2.7.4. Recombinant E. coli
The PHA production by recombinant E. coli on different substrates has been widely
reported (Li et al., 2007b). PHA composition can be altered by engineering desirable
genes into E. coli (Li et al., 2010; Tomizawa et al., 2011). In a cell-recycle fed batch
fermentation strategy, recombinant E. coli CGSC4401 (harboring Alcaligenes latus
PHA genes) yielded 4.6 g PHB/L/h from whey (Ahn et al., 2001). E. coli
phosphotransferase system mutant strain LR1010 harboring phaCRe and phaABRe
genes from R. eutropha could simultaneously consume glucose and xylose to
accumulate SCL-PHA. Here total cdw was achieved 2.3 g/L, with PHA equivalent to
11.5% of cdw. However, E. coli LR1110, harboring phaC1 gene from P. aeruginosa
could accumulate MCL-PHA (2.3 g/L of cdw) by using a mixture of glucose and fatty
acids (Li et al., 2007a). More recent attempts of co-expression of phaA and phaB
genes from Bacillus sp. and phaC1 and phaJ1 from Pseudomonas sp. in E. coli
JC7623 resulted in production of PHA with SCL and MCL in the range of 78:22 to
18:82. The PHA yields by E. coli JC7623ABC1J1 were further improved from 28 to
34% of cdw by an additional 3 to 11% as a result of the action of acrylic acid, an
inhibitor of β-oxidation (Davis et al., 2008). Ultra high molecular weight PHB was

Chapter-2 Review of Literature
31
produced by recombinant E. coli (phaCABRe of R. eutropha) (Park et al., 2005)
resulting in successful preparation of PHB stretched films and improvement in
mechanical properties (Tsuge, 2002). Recently, a Novel lactic acid (LA)-based
terpolymer, P(LA-co-3HB-co-3HV), was produced in LA-overproducing mutant E.
coli JW0885 on glucose and propionate. Here, the mutant strain harbored the genes
encoding LA-polymerizing enzyme (mutant of phaC1 from Pseudomonas sp. 61-3),
phaA and phaB from R. eutropha and propionyl-CoA transferase gene from
Megasphaera elsdenii (Shozui et al., 2010). Poly (3-hydroxypropionate) (3HHp) was
recently reported with recombinant E. coli harbouring the genes for glycerol
dehydratase of Clostridium butyricum, propionaldehyde dehydrogenase of Salmonella
enterica serovar Typhimurium LT2 and phaC1 gene of R. eutropha. A yield of
11.98% of cdw was achieved in two step fed batch fermentation using glycerol as
substrate (Andreessen et al., 2010).
2.7.5. Bacillus: Future PHA producer
A wide range of Bacillus spp. has been reported to accumulate PHB: B.
amyloliquefaciens DSM7 (11% of cdw), B. laterosporus (14%), B. licheniformis
(21%), B. macerans (28%), B. cereus (32%) and B. circulans (34%), B. firmus G2
(15%), B. subtilis K8 (32%), B. sphaericus X3 (36%), B. megaterium Y6 (48%), B.
coagulans (24%), B. brevis (41%), B. sphaericus ATCC 14577 (6%), B.
thuringiensis (52%), B. mycoides RLJ B-017 (69%) (Yilmaz et al., 2005; Porwal et
al., 2008; Singh et al., 2009). Bacillus sp. INT005 and B. cereus UW85 could
produce PHA with a wide range of compositions varying from PHB, P(3HB-co-
3HV), P(3HB-co-3HHx), P(3HB-co-4HB-co-3HHx) to P(3HB-co-6HHx-co-3HHx)
depending up on the substrate (Labuzek and Radecka, 2001; Tajima et al., 2003;
Valappil et al., 2008). Various Bacillus spp. have been shown by different
researchers to synthesize copolymers when co-fed with various substrates. Using B.
cereus UW85, the production of terpolymer of 3HB, 3HV and 6HHx was recorded
with ε- caprolactone as sole C source in mineral salts medium without any glucose.
However, addition of glucose along with ε-caprolactone seemed to suppress
copolymer synthesis and the result was the production of PHB (Labuzek and
Radecka, 2001). Bacillus sp. INT005 could accumulate PHB when glucose was used

Chapter-2 Review of Literature
32
alone as C substrate in the medium. However, addition of various C sources along
with very low glucose concentration resulted in copolymers of 3HB and 3HHx on
octanoate and decanoate, copolymers of 3HB-4HB-4HHx on 4-hydroxybutanoate
and 3HB-3HHx-6HHx on supplementation with ε-caprolactone (Tajima et al.,
2003). Recent studies have produced still more interesting information. B. cereus
SPV when grown on structurally unrelated C sources such as fructose, sucrose and
gluconate resulted in the incorporation of 4HB with the first two substrates and 4HB
and 3HV with gluconate in the medium (Valappil et al., 2007b). Although limitation
of nitrogen (N), phosphorous (P) and oxygen in the culture conditions are known to
influence PHB production, however, potassium limiting media led to the production
of a copolymer containing 3HB and 3HV monomers in contrast to only PHB under
sulphur, P or N limitation (Valappil et al., 2008). B. cereus CFR06 yielded PHB on
starch (Halami, 2008) and other Bacillus spp. could also produce PHB from
industrial food waste water, soya waste and malt waste from beer brewery plant and
PSS (Kumar et al., 2009). Bacillus sp. 256 utilize an unrefined natural substrate –
mahua (Madhuca sp.) flower as C source (containing 57% w/w as sugars) to
produce copolymers P(3HB-co-10% 3HV) (Anil-Kumar et al., 2007). Recently a
highy polymer yield of 61.07 % dcw, in 1L shaken flask study and 51.37 % dcw in
2L fermenter study has been reported, using B. cereus SPV with sugarcane molasses
as the main carbon source (Akaraonye et al., 2011). B. subtilis has been recently
used as a host for over expression of phaCAB genes from P. aeruginosa and R.
eutropha. Expression of phaC1 from P. aeruginosa and phaAB genes from R.
eutropha in B. subtilis resulted in the production of copolymers P(HD-co-HDD and
P(HB-co-HD-co-HDD) from malt waste (Wang et al., 2006). Since B. subtilis is not
a human pathogen, further supporting its usage as a host for expression of foreign
genes (Law et al., 2003). Here, B. megaterium phaPQRBC genes were cloned into
B. subtilis 1A304 (Φ105MU331). Recombinant B. subtilis could utilize malt waste
as a C source, further raising the hopes for producing PHA at cheaper rates. This
study showed that phaPQ of B. megaterium was essential for PHA production along
with phaRBC, although they could not observe any sequence homology in NCBI
database.

Chapter-2 Review of Literature
33
Several species of Bacillus, a Gram-positive bacterium have certain characteristics on
the basis of which they can be considered as potential candidate(s) for PHA production
on industrial scale. A brief description of their unique abilities is presented here.
(i) Lacks Lipopolysachharides (LPS)
Gram-negative bacteria such as C. necator (Vandamme and Coenye, 2004), A. latus
and recombinant E. coli are among those which have been exploited even for
industrial scale PHB production. However, in these organisms, the outer membrane
LPS are endotoxins (Stewart et al., 2006), which co-purify with the PHAs. Since LPS
induces a strong immunogenic reaction, this feature is undesirable for biomedical
applications of the PHAs (Chen and Wu, 2005; Valappil et al., 2007a). A review on
the toxic nature of LPS reveals that cyanobacterial (Gram- positive bacteria) LPS are
less toxic compared to those produced by members of Enterobacteraceae (Stewart et
al., 2006). Gram positive bacteria such as B. subtilis offers the advantage of lacking
LPS and excreting proteins at a high rate into the medium (Morimoto et al., 2008). It
thus stands a better choice as PHA producers for biomedical applications.
(ii) Possible candidate to replace E. coli as expression host
E. coli has been extensively used as a host for over expression of foreign genes,
including those involved in PHA biosynthesis. However, a major limitation of this
system is the secretion of proteins which result in the formation of inactive inclusion
bodies (Leonbartsberger, 2006; Chen et al., 2007). B. subtilis has been recently used
as a host for over expression of phaCAB genes from P. aeruginosa and R. eutropha. It
was chosen as host over E. coli, because of its short generation time, absence of
endotoxins and secretion of hydrolytic enzymes (amylases, proteinases, etc) for
utilizing biowastes as feed (Porwal et al., 2008), which may help in reducing the cost
of PHA production (Wang et al., 2006). Expression of phaC1 from P. aeruginosa and
phaAB genes from R. eutropha in B. subtilis resulted in the production of copolymers
P(HD-co-HDD) and P(HB-co-HD-co-HDD) from malt waste (Wang et al., 2006).
Since B. subtilis is not a human pathogen, it thus further supports its usage as a host
for expression of foreign genes (Law et al., 2003).

Chapter-2 Review of Literature
34
(iii) Potential as hydrogen producer
The use of CO2 as a potential inexpensive renewable C source can help in reducing
PHA production cost (Tsuge, 2002). Synechococcus sp. MA19 was observed to
produce PHA upto 55% of cdw from CO2 (Nishioka et al., 2001). A. eutrophus was
shown to accumulate PHA at the rate of 1.55g/L/h, which was higher than that
recorded with cyanobacteria or photosynthetic bacteria (Tanaka et al., 1995). The
strategy being proposed is that if hydrogen (H2) production becomes cheap then R.
eutropha can produce PHB from CO2 and oxidation of H2, under dark fermentative
conditions (Ishizaki et al., 2001). Bacillus seems to meet the requirements of this
proposal. Bacillus spp. such as B. coagulans, B. licheniformis and B. subtilis have
been shown to evolve 1.5 to 2.36 mol H2/mol glucose (Kalia and Purohit, 2008).
Biowastes rich in starch such as damaged wheat grains have been employed as feed
for generating H2 (45 to 64 L/kg Total solids) by B. licheniformis and B. subtilis
(Sonakya et al., 2001). B. cereus strain EGU43 and B. thuringiensis strain EGU45
have been reported to generate 0.63 mol of H2/mol of glucose and up to 500 mg
PHB/L in different conditions (Porwal et al., 2008). More re cently, these strain have
been reported to produce both H2 followed by PHB on same media from glucose in
two stage system which further enhances the efficiency of the system (Patel et al.,
2011a).
2.7.6. PHA production by mixed microbial cultures (MMCs)
Among other factors causing high production cost is the sterilisation process. As an
alternative, using the combination of the MMCs on biowaste allows decreasing
operating costs by reducing the cost of substrate and that of the energy used (since no
sterilization is required) (Albuquerque et al., 2011). The use of MMC to produce
added value products (such as biochemicals and biomaterials) using ecological
selection principles to engineer the microbial consortium has been designated as
ecobiotechnology (Kleerebezem and van Loosdrecht, 2007). Enrichment of PHA
accumulating organisms is generally carried out by subjecting mixed cultures to
transient conditions of carbon supply, designated as Aerobic Dynamic Feeding or
Feast and Famine (Albuquerque et al., 2011).

Chapter-2 Review of Literature
35
At early stages research was focused on the use of chemically defined media
supplemented with synthetic volatile fatty acids. Mixed microbial cultures collected
from activated sludge under feast and famine condition have been shown to
potentially producing PHA (Serafim et al., 2007; Jiang et al., 2008; Albuquerque et
al., 2010a, b). A maximum PHA content of 90% (PHA concentration; 4.3 g/L) from
lactate (Jiang et al., 2008) followed by 89% (PHA concentration; 1.2g PHA/g
substrate/h) from acetate (Johnson et al., 2009) and 78.5% from acetate pulse feeding
(Serafim et al., 2007) have been reported by mixed microbial cultures. However, the
use of low cost agro-industrial surplus feedstocks wastes and by-products is gaining
interest recently as reported by several authors e.g. fermented molasses (Albuquerque
et al., 2007; 2010a, b; Bengtsson et al., 2010a, b), fermented paper mill effluents
(Bengtsson et al., 2008), industrial wastewaters (Dionisi et al., 2006), fermented olive
oil mill effluents (Dionisi et al., 2005; Beccari et al., 2009), municipal wastewaters
(Coats et al., 2007), olive oil mill effluents (Dionisi et al., 2005; Beccari et al., 2009),
paper mill effluent (Bengtsson et al., 2008). Use of VFAs, which are the major
products of fermentation of biowaste, as feed for PHA production is the most
promising and efficient strategy employed by various researchers. Albuquerque et al.
developed a three-stage PHA production process from sugar cane molasses and have
reported a PHA content of 74.5% of cdw from VFA produced in fermented sugar
molasses attaining PHA concentration to the level of 0.49 g PHA/g substrate/h
(Albuquerque et al., 2007; Albuquerque et al. 2010a, b). An advantage with the use of
mixed culture is the production of co-polymers with a broad range of PHA
compositions. PHA containing different HA monomers can be produced on fermented
feedstocks (containing mixtures of organic acids such as acetate, propionate, butyrate
and valerate) without the need for added co-substrates as is the case for pure culture
fermentations and thus offers the possibility of producing PHA with a wide range of
thermal and mechanical properties. Lee et al., 2008 reported that pure culture
produced solely PHB and required large amounts of co-substrates (such as alcohols or
organic acids) to produce polymers with relatively small fractions of monomers (2-
8% HV) with same amount of feedstock. Co-polymers of P(HB-co-HV) with HV
ranging from 17 to 85% were reported (Lemos et al., 2006) with acetate and
propionate as substrate. 3HV content of upto 69 mol% is reported using mixed

Chapter-2 Review of Literature
36
microbial cultures (Bengtsson et al., 2008; Arcos-Hernandez et al., 2010). Other
monomers reported include 3H2MV or 3HHx (Lemos et al., 2006; Bengtsson et al.,
2010b). A termopolymer of 3HB:3HV:3H2MV (molar ratio-6:58:24) was reported by
Lemos et al. (2006) from acetate/propionate mixtures.
Thus, mixed bacterial culture production has the potential to produce large amounts of
PHAs with seemingly lower costs. With the use of wastes and open cultures, a more
robust process could be obtained since, compared to pure cultures, mixed cultures are
more amenable to complex feedstocks. Although potential of mixed microbial culture
to produce PHA have been shown in early reports also, but potential of defined mixed
microbial cultures (consortia) to produce PHA have not been reported yet either on
synthetic media or biowaste.
2.8. Genomic status of PHA biosynthesis
The genes and enzymes involved in the biosynthesis of PHA vary in relation to their
organization among different organisms. The diversity of the PHB biosynthetic
pathways implies how far the pha loci have diverged. The phb (genes encoding
enzymes for SCL-PHA) and pha (genes encoding enzymes for MCL-PHA) are not
necessarily clustered and the gene organization varies from species to species (Reddy
et al., 2003). According to the need of organisms, with evolution phaC gene got
arranged with genes supplying monomers like phaAB and phaJ and while in some
organisms with genes involved in PHA regulation like phaZ and phaP (Naik et al.,
2008).
Among β-proteobacteria such as R. eutropha, Burkhelderia sp. A. latus, Delftia
acidovorans operon arrangement of PHA biosyhthesis genes is related to SCL-PHA
biosynthesis (class I PHA synthase gene) (Rehm, 2003). Here the genes are arranged
tandemly on the chromosome (Figure 2.6 & 2.7). P. acidophila, R. eutropha contains
the similar arrangement whereas Acinetobacter spp. contain the complete phaCAB
operon but genes are not in the same order (Rehm and Steinbuchel, 1999; Rehm,
2003; Suriyamongkol et al., 2007; Naik et al., 2008).

Chapter-2 Review of Literature
37
Figure 2.6: Biosynthetic pathway and gene arrangement in Ralstonia eutropha
In bacteria belonging to α-proteobacteria such as Azorhizobium caulinodans,
Caulobacter crescentus, M. extorquens, Paracoccus denitrificans , Rhizobium metli
and Sinorhizobium meliloti containing class I PHA synthase genes phaC and phaAB
loci are unlinked or uninterrupted i.e. phaC is found elsewhere in the chromosome
(Rehm and Steinbuchel, 1999; Rehm, 2003). Other than α-proteobacteria, bacteria
such as such as Z. ramigera (β-proteobacterium), Aeromonas punctata (γ -
proteobacterium) and Gordonia rubripertinctus (a firmicute) contain similar
biosynthetic genes arrangement that are not co-localized (Naik et al., 2008).
Pseudomonas which produces MCL-PHA posses class II synthase e.g. P. oleovorans
and P. aeruginosa containing two different phaC genes i.e. phaC1 and phaC2
seperated by an additional phaZ gene encoding intracellular PHA depolymerase
enzyme (Figure 2.7). Enzymes encoded by phaC1 and phaC2 have similar primary
structures and substrate specificity. Downstream to the synthase gene arrangement,
phaD is located followed by phaI and phaF encoding structural and regulatory
proteins (Rehm, 2003). In bacteria having class III PHA syhthase, phaC and phaE
encoding the two subunits of polymerase enzyme are directly linked constituting a
single operon. phaEC and phaAB are present in one loci but are having opposite
orientation e.g. A. vinosum, Thiocystis violacea and T. pfennigii, Synechocystis sp.. In
Synechocystis sp. phaA and phaB do not map close to phaEC locus (Rehm, 2003). In
M. extorquens, Nocardia corallina, R. metli and Rhodococcus ruber an additional
gene phbP is present with an unknown function in the PHB regulation. Here also
phbCP and phbAB are present in the same locus but with different orientations

Chapter-2 Review of Literature
38
therefore only the PHB polymerase encoding gene phbC has been identified so far
(Hustede and Steinbuchel, 1993). In A. caviae the PHB polymerase encoding gene
phbC has a flanking gene phbJ, which provides monomer for the enzyme. In C.
vinosum, P. acidophila, R. eutropha, R. meliloti and T. violacea an additional gene
phbF is present with an unknown function in PHA regulation (Suriyamongkol et al.,
2007). In P. acidophila, R. eutropha and Z. ramigera have a gene present upstream of
phbCAB operon and which is identical to the hypothetical E. coli protein YfiH (Naik
et al., 2008).
Figure 2.7: Genetic organization of representative PHA biosynthesis genes encoding the
various classes of enzymes (Rehm, 2003; Naik et al., 2008)
In Bacillus that contain class IV synthase phaR and phaC encoding PhaR and PhaC
subunits are present separated by phaB (McCool and Cannon, 2001; Rehm, 2003;
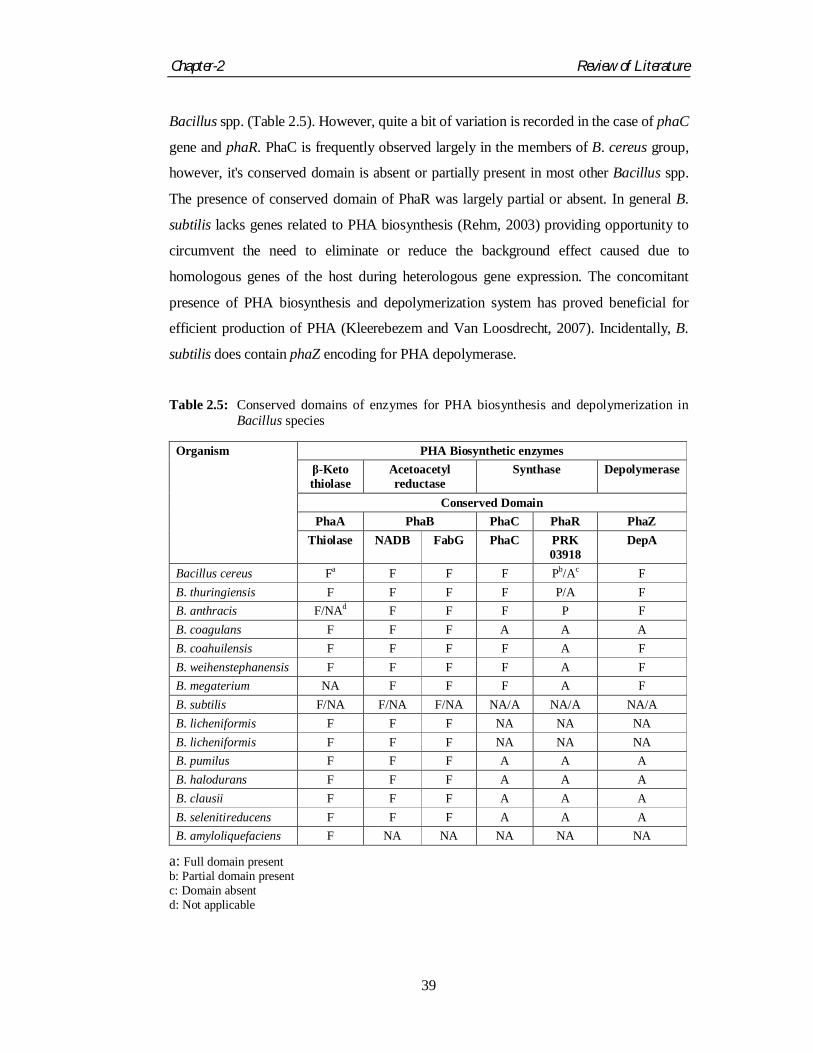
Valappil et al., 2007a). Screening of metabolic (KEGG) and genomic (NCBI) databases
for the presence of enzymes (Kalia et al., 2003) involved in PHA biosynthesis reveals that
genes for PhaA and PhaB are observed to be present in almost all the sequenced genomes
of Bacillus except certain strains of B. megaterium, B. thuringiensis, B. subtilis and
Chapter-2 Review of Literature
39
Bacillus spp. (Table 2.5). However, quite a bit of variation is recorded in the case of phaC
gene and phaR. PhaC is frequently observed largely in the members of B. cereus group,
however, it's conserved domain is absent or partially present in most other Bacillus spp.
The presence of conserved domain of PhaR was largely partial or absent. In general B.
subtilis lacks genes related to PHA biosynthesis (Rehm, 2003) providing opportunity to
circumvent the need to eliminate or reduce the background effect caused due to
homologous genes of the host during heterologous gene expression. The concomitant
presence of PHA biosynthesis and depolymerization system has proved beneficial for
efficient production of PHA (Kleerebezem and Van Loosdrecht, 2007). Incidentally, B.
subtilis does contain phaZ encoding for PHA depolymerase.
Table 2.5: Conserved domains of enzymes for PHA biosynthesis and depolymerization in
Bacillus species
Organism PHA Biosynthetic enzymes β-Keto thiolase
Acetoacetyl reductase
Synthase Depolymerase
Conserved Domain PhaA PhaB PhaC PhaR PhaZ
Thiolase NADB FabG PhaC PRK 03918
DepA
Bacillus cereus Fa F F F Pb/Ac F B. thuringiensis F F F F P/A F B. anthracis F/NAd F F F P F B. coagulans F F F A A A B. coahuilensis F F F F A F B. weihenstephanensis F F F F A F B. megaterium NA F F F A F B. subtilis F/NA F/NA F/NA NA/A NA/A NA/A B. licheniformis F F F NA NA NA B. licheniformis F F F NA NA NA B. pumilus F F F A A A B. halodurans F F F A A A B. clausii F F F A A A B. selenitireducens F F F A A A B. amyloliquefaciens F NA NA NA NA NA
a: Full domain present b: Partial domain present c: Domain absent d: Not applicable

Chapter-2 Review of Literature
40
As observed through this diverse arrangement of genes it can be concluded that
evolutionary forces has caused the clustering of phb genes in an operon with same
transcriptional units as in P. acidophila, R. eutropha, Acinetobacter sp., A. latus, A.
caviae and at times in separate transcriptional units as in Z. ramigera, P. denitrificans,
R. meliloli, C. vinosum, T. violaecae, P. oleovorans, P. putida etc. In some cases it
resulted in diverse orientation and varying flanking regions ultimately resulting in
diverged rearrangements in the phb loci.


















