
Changes in Plasma Amino Acids during Development of X-irradiated Chick Embryos
9
Int. J. Rad. Biol., 1961, Vol. 3, No. 5, 449-457 Changes in plasma amino acids during development of X-irradiated chick embryost ELAINE KATZ BERNSTEIN with the technical assistance of Celestia Crawford Division of Biological and Medical Research, Argonne National Laboratory, Argonne, Illinois (Recei*;ed 13 June 1960) 1. Studies were made of the plasma amino acids of chick embryos from clays 5-19 of development after x-irradiation with 600 r on the fourth day of incubation. 2. The levels of amino acids in the 6-8 day embryos decreased : this was a general effect and was not specific for any single amino acid. 3. The levels of amino acids in the 5 day and in the 9-19 day embryos increased : this, again, was a general effect and was not specific for any single amino acid. For days 9-12 the increase in free amino acids was statistically significant. No abnormally occurring amino acids were present. 4. It is believed that the responses of the embryos between 5 and 8 days may be peculiar to developing embryonate tissue, while the responses between 9 and 19 days indicate a basic metabolic disturbance and not one that is peculiar to embryonate tissue. 5. Possible relationships between this increase in free plasma amino acids following x-irradianon and the enhancement of the growth of rickettsial organisms by administration of x-irradiation are discussed. 1. INTRODUCTION Previous work in this laboratory has shown that x-irradiation causes changes in the kind and amount of amino acids excreted in human urine and in the plasma of the newly hatched chick (Bernstein, Steamer and Brues 1956, Katz and Hasterlik 1955, Hempelmann, Lisco and Hoffman 1952). Other investi- gators have shown similar increases in other species (Kay and Entenman 1959, Kay, Harris and Entenman 1956, Hallesy and Doull 1956, Mefferd and Martens 1955, Koszalka, Bowerman and Hempelmann 1955). The increased amino- aciduria and aminoacidemia appear to be part of a general depletion of nitrogen after whole-body x-irradiation (Krise, Williams and Anderson 1957, Williams, Krise, Anderson and Dowlen 1957, Gustafson and Koletsky 1952, Kumta, Gurnami and Sahasrabudhe 1957, Rust, Visek and Roth 1957, Hunter 1957, Weber and Cantero 1957, Jackson and Entenman 1958, Pentz 1958). In both man and the newly-hatched chick the alteration of amino-acid levels occurred immediately after x-irradiation. There was no correlation between (a) the amount of x-irradiation and the degree of aminoaciduria or aminoacidemia or (b) visible evidence of radiation effect and the degree of aminoaciduria or amino- acidemia. t This work was performed under the auspices of the U.S. Atomic Energy Commission. R.B. 2 G Int J Radiat Biol Downloaded from informahealthcare.com by UB Kiel on 11/06/14 For personal use only.
Transcript of Changes in Plasma Amino Acids during Development of X-irradiated Chick Embryos

Int. J. Rad. Biol., 1961, Vol. 3, No. 5, 449-457
C h a n g e s in p l a s m a a m i n o a c i d s d u r i n g d e v e l o p m e n t o f X- i rrad ia ted c h i c k e m b r y o s t
ELAINE KATZ BERNSTEIN
with the technical assistance of Celestia Crawford
Division of Biological and Medical Research, Argonne National Laboratory, Argonne, Illinois
(Recei*;ed 13 June 1960)
1. Studies were made of the plasma amino acids of chick embryos from clays 5-19 of development after x-irradiation with 600 r on the fourth day of incubation.
2. The levels of amino acids in the 6-8 day embryos decreased : this was a general effect and was not specific for any single amino acid.
3. The levels of amino acids in the 5 day and in the 9-19 day embryos increased : this, again, was a general effect and was not specific for any single amino acid. For days 9-12 the increase in free amino acids was statistically significant. No abnormally occurring amino acids were present.
4. It is believed that the responses of the embryos between 5 and 8 days may be peculiar to developing embryonate tissue, while the responses between 9 and 19 days indicate a basic metabolic disturbance and not one that is peculiar to embryonate tissue.
5. Possible relationships between this increase in free plasma amino acids following x-irradianon and the enhancement of the growth of rickettsial organisms by administration of x-irradiation are discussed.
1. INTRODUCTION
Previous work in this laboratory has shown that x-irradiation causes changes in the kind and amount of amino acids excreted in human urine and in the plasma of the newly hatched chick (Bernstein, S teamer and Brues 1956, Katz and Hasterl ik 1955, Hempe lmann , Lisco and Hoffman 1952). Other investi- gators have shown similar increases in other species (Kay and En tenman 1959, Kay, Harris and En tenman 1956, Hallesy and Doull 1956, Mefferd and Mar tens 1955, Koszalka, Bowerman and H e m p e l m a n n 1955). T h e increased amino- aciduria and aminoacidemia appear to be part of a general depletion of ni trogen after whole-body x-irradiation (Krise, Williams and Anderson 1957, Williams, Krise, Anderson and Dowlen 1957, Gustafson and Koletsky 1952, Kumta , G u r n a m i and Sahasrabudhe 1957, Rust, Visek and Roth 1957, Hun te r 1957, Weber and Cantero 1957, Jackson and En tenman 1958, Pentz 1958). In both man and the newly-hatched chick the alteration of amino-acid levels occurred immedia te ly after x-irradiation. The re was no correlation between (a) the amount of x-irradiation and the degree of aminoaciduria or aminoacidemia or (b) visible evidence of radiation effect and the degree of aminoaciduria or amino- acidemia.
t This work was performed under the auspices of the U.S. Atomic Energy Commission.
R.B. 2 G
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.

450 Elaine Katz Bernstein
In other studies it was demonstrated that x-irradiation of chick embryos resulted in more rapid growth of the intracellular organism Rickettsia mooseri (Greiff, Powers and Pinkerton 1957) and inhibition of R. akari (Greiff, Powers, Kisieleski and Pinkerton 1960) cultured in the chorioallantoic membranes of the irradiated embryos. Because of the results described above for chicks and other animals, it was thought that changes in the nitrogen metabolism of the host cells after radiation injury might be responsible for these changes in the rickettsial growth-patterns.
In this series of experiments, studies of the plasma of x-irradiated chick embryos were undertaken in order to expand this work, to extend the chick studies to organisms during their developmental period, and to use an experi- mental subject that would have some extraneous influences eliminated through the use of a closed system. Preliminary reports have been published (Katz and Powers 1955 a, b).
2. MATERIALS AND METHODS
Eggs from White Leghorn hens (Creighton Hatcheries, Warsaw, Indiana), which were received within twenty-four hours of collection at the hatchery, were stored in a cool room (20°c) before incubation. They were white, clean, un- washed, uniform in size, and antibiotic-free; fertility was excellent, averaging about 90 per cent. The eggs were candled immediately before use and infertile ones discarded; almost all were used within 3-5 days of being laid. Handling was kept to a minimum. They were incubated in a large automatic electric incubator-hatcher at 38°c, with forced air circulation, 86 per cent humidity (wet-bulb measurement), and were automatically rotated twice a day.
X-irradiation was carried out in the morning of the fourth day of incubation, as follows: single dose 6C0 r, 66 r/rain, 75 per cent emergent dose, 250kv through 3-0 mm of Cu and 1-0 mm of A1 (3.43 mm Cu HVL). Ten eggs were irradiated at a time. The eggs were out of an incubator only during actual irradiation time (about 10 rain). Irradiation was done at 4 days to duplicate working conditions of other investigators, to permit homogeneous irradiation, and to avoid the peak of natural deaths (Greiff et al. 1957, 1960, Boland 1954).
Since the size, development, and vitality of embryos are dependent upon many factors not under the control of the experimenter, the criteria established by embryologists (Hamilton 1952) were used as standards in order to insure minimum variability of embryos. Embryos that did not resemble the average because of either underdevelopment or overdevelopment were discarded. Selection of x-irradiated embryos was based on the same criteria, but obvious radiation effects had to be considered in the judgment. Since fertile eggs have a lower viability and show greater abnormalities in the summer months, this work was carried out between December and May.
Samples of whole bloed were drawn from embryos on days 5-19 of develop- ment. For sampling the blood of embryos of 5-7 days development, the eggs were placed upright in the incubator for 1-2 hours before use so that the embryos moved to the surface of the egg. The embryo was then floated out into a warm solution of physiological saline and cut away from the yolk sac. A whole mount was made, using half of a small Petri dish as a base. Blood was drawn from the anterior vitelline vein with an extremely fine glass capillary pipette. Capillaries were rinsed with standard heparin solution (1 per cent, Abbott)
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.
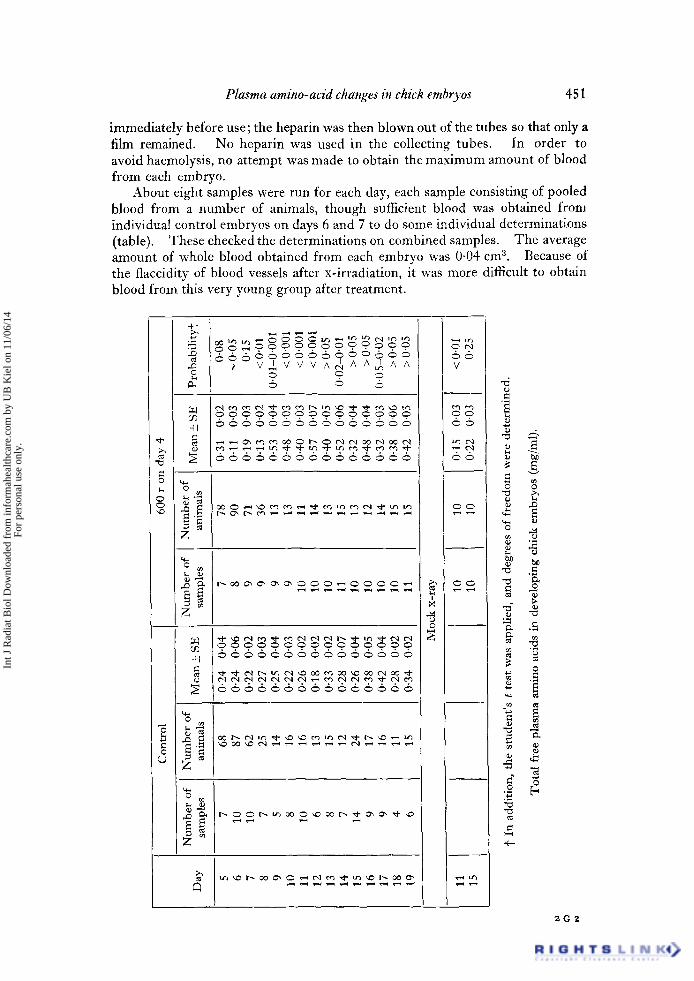
Plasma amino-acid changes in chick embryos 451
immediately before use;the heparin was then blown out of the tubes so that only a film remained. No heparin was used in the collecting tubes. In order to avoid haemolysis, no attempt was made to obtain the maximum amount of blood from each embryo.
About eight samples were run for each day, each sample consisting of pooled blood from a number of animals, though sufficient blood was obtained from individual control embryos on days 6 and 7 to do some individual determinations (table)• These checked the determinations on combined samples. The average amount of whole blood obtained from each embryo was 0.04 cm 3. Because of the flaccidity of blood vessels after x-irradiation, it was more difficult to obtain blood from this very young group after treatment.
§
(,.)
+ -
,,.D V ~ V V V A ~ A A ~ A A
+1
X
'2
Z
Z
+ I
C,
V
e ~
e q .,..~ ¢.q
S S
x..--¢
©
.-&
a
+ -
,.-.,
o
o
0J
o
. 2 0 2
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.

452 Elaine Katz Bernstein
Samples were obtained from the embryos of 8-19 days development by opening the eggs at the air space and making the shell membrane translucent with mineral oil (using a minimum amount). Glass capillary pipettes, rinsed with heparin immediately before use and treated as described above, were used for all samples; dried heparin was added to the collecting tubes (micro-haematocrit tubes) for samples from days 12-19 in amounts as follows:
0.013 cm 3 heparin for days 12-14 (1 drop), 0.026 cm a heparin for days 15-17 (2 drops), 0.04 cm a heparin for days 18-19 (3 drops).
All blood samples in this age group were taken from the distal portion of the allantoic vein directly under the air sac. An egg stand made with rubber and lucite to specifications already described (Beveridge and Burnet 1946) was invaluable.
There is no difficulty in obtaining blood from embryos after the eighth day of development. For all days, the quality of the capillary pipettes is crucial in obtaining samples that are unhaemolyzed and uncontaminated with allantoic fluid.
Amino-acid determinations were made on plasma from unhaemolyzed blood. Samples were centrifuged immediately; the plasma was separated and the pro- tein precipitated with 80 per cent ethanol in a 1 : 10 dilution, and filtrates eva- porated over a water bath to the original plasma volume. Amino-acid deter- minations were done immediately in triplicate with aliquots of 25, 50 and 100/xl and using two-dimensional paper chromatography in the conventional manner- - phenol: water: ammonia for the first solvent, and collidine: lutidine: water for the second. Reading the sheets for identification and quantitation of amino acids was done by visual comparison of the unknown amino acids with the amino-acid standards, using a commercial 14in. x 17 in. x-ray viewing box. In all cases the amino-acid standard solutions were run on the same sheet as the sample. Standard colour tests, position matching of unknown amino acids, etc., were employed for the identification of questionable substances. A full descrip- tion of the chromatographic techniques used and the method of quantitation of individual amino acids has been previously published (Bernstein et al. 1956, Katz and Hasterlik 1955).
3. RESULTS
Radiation-induced changes noted in the embryos included inconspicuous haemorrhages, flaccidity of blood vessels, change in the consistency of the shell membrane, enlarged and peculiarly-shaped air sacs, and the presence of fluid between the chorioallantoic sac and shell membrane. This latter has been described as an abnormality sometimes found in untreated embryos and often found in infected embryos (Beveridge and Burnet 1946), but has never been seen in this laboratory in an unirradiated embryo. In addition to these general effects, radiation-induced changes related to specific days of development were as follows :
(a) Days 5 and 6: evidence of' clotting ', or a seemingly similar phenomenon ;
(b) Days 6-I0: poor and seemingly retarded development of blood vessels in both size and distribution;
(c) Days 12-19: prolonged clotting time.
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.
Plasma amino-acid changes in chick embryos 453
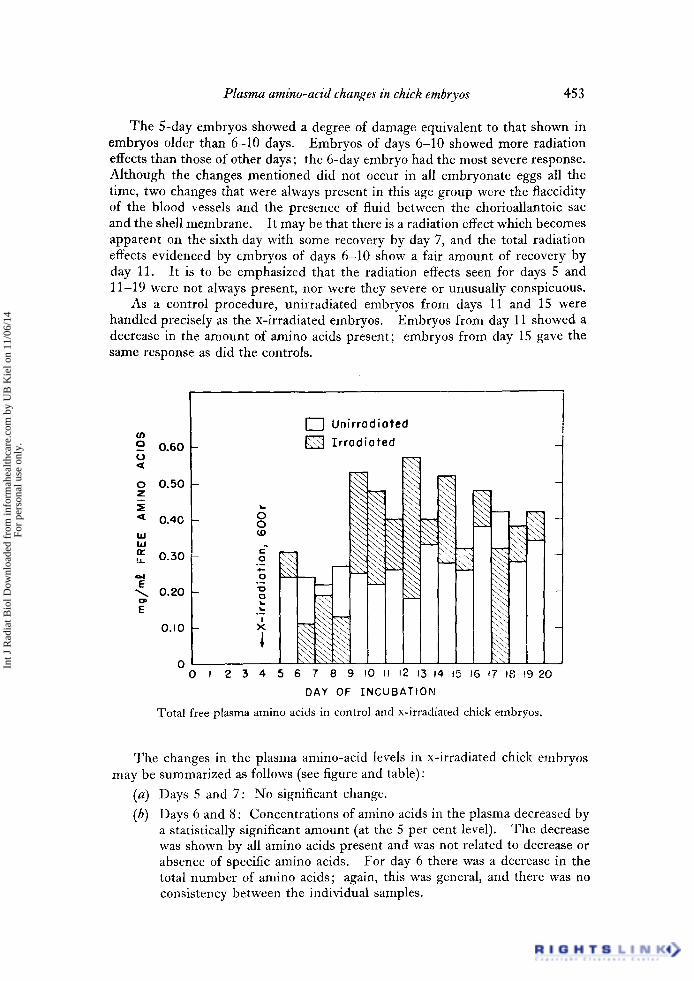
The 5-day embryos showed a degree of damage equivalent to that shown in embryos older than 6-10 days. Embryos of days 6-10 showed more radiation effects than those of other days; the 6-day embryo had the most severe response. Although the changes mentioned did not occur in all embryonate eggs all the time, two changes that were always present in this age group were the flaccidity of the blood vessels and the presence of fluid between the chorioallantoic sac and the shell membrane. It may be that there is a radiation effect which becomes apparent on the sixth day with some recovery by day 7, and the total radiation effects evidenced by embryos of days 6-10 show a fair amount of recovery by day 11. It is to be emphasized that the radiation effects seen for days 5 and 11-19 were not always present, nor were they severe or unusually conspicuous.
As a control procedure, unirradiated embryos from days 11 and 15 were handled precisely as the x-irradiated embryos. Embryos from day 11 showed a decrease in the amount of amino acids present; embryos from day 15 gave the same response as did the controls.
9 o.6o r o
o 0.50 Z
'¢ 0 . 4 0 I.IJ W
n: 0 . 3 0
E 0.20
E
OAO
0
¢-
0
0
1 0
° - - I
X
[ ~ ] Unir rodi0ted
I r r ad ia ted
,xg
1
0 0 I 2 3 4 5 6 7 8 9 l0 ii 12 r3 14 ~5 t6 e7 ~8 ~9 20
DAY OF I N C U B A T I O N
Total free plasma amino acids in control and x-irradiated chick embryos.
The changes in the plasma amino-acid levels in x-irradiated chick embryos may be summarized as follows (see figure and table):
(a) Days 5 and 7: No significant change.
(b) Days 6 and 8 : Concentrations of amino acids in the plasma decreased by a statistically significant amount (at the 5 per cent level). The decrease was shown by all amino acids present and was not related to decrease or absence of specific amino acids. For day 6 there was a decrease in the total number of amino acids; again, this was general, and there was no consistency between the individual samples.
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.

454 Elaine Katz Bernstein
(c) Days 9-19: A general increase was found in the total amount of amino acids present, and some values from individual embryos were three or four times higher than control values. For days 9-12 the increase in free amino acids was statistically significant. The spread of values for days 9-19 was large, and for days 13-19 individual values were about two times as high as the highest control levels. The increase in total amount of free amino acids was general and was not related to a large increase in specific amino acids. In the 19-day embryos there was a slight increase in the number of amino acids found.
By using the chromatographic technique described, we observed a number of amino acids. Other amino acids are probably normally present, and could be demonstrated either by modifications of this technique or by use of other chromatographic or microbiological methods. Some combination of the following amino acids was found in the plasma of the normal chick embryo; each amino acid was not observed in each sample.
Aspartic acid, cystine, glutamic acid, serine, glycine, taurine, threonine, alanine, glutamine, tyrosine, /?-alanine, 7,-amino-n-butyric acid, histidine, tryptophane, hydroxyproline, valine, leucine, methionine, lysine and/or ornithine, proline, arginine, 7-aminobutyric acid, methylhistidine, ethanol- amine, and phosphoethanolamine.
After x-irradiation no abnormally-occurring amino acids were present. There was one instance, in a 12-day embryo, of the appearance of fi-amino-isobutyric acid, but this should not be classified as an abnormally-appearing amino acid on the basis of one occurrence.
In almost all samples from irradiated animals, however, certain amino acids, which appeared only sporadically in the controls, appeared and persisted during days 9-19. These are taurine, fi-alanine, 7-aminobutyric acid, methylhistidine, cystine (as cysteic acid), hydroxyproline, ethanolamine, and phosphoethanol- amine.
4. DIscussiON
The results show that x-irradiation causes a generalized disturbance in amino-acid and protein metabolism in the embryonate egg. This disturbance is not closely correlated with obvious gross morphological changes induced by radiation. Although physiological evidences of radiation injury were seen, they were not conspicuous even in embryos showing marked changes in amino acids. There was, moreover, no obvious difference in appearance between embryos showing definite amino-acid changes and those giving a non-significant response.
Several physiological mechanisms which might be responsible for the increase in free plasma amino acids may be noted. Decreased oxidation and utilization of amino acids by the liver and other tissues of the embryo may be a cause. The embryo catabolizes protein by at least the fourth day (Patten 1951); by the 8th day of incubation the liver is synthesizing glycogen, bile, cholesterol, and creatine, so that under normal circumstancs the 8-day embryo actively utilizes amino acids and synthesizes protein (Needham 1950). Under normal circumstances, moreover, the yolk is in its greatest catabolic period at about the 10th day (Needham 1950). The fact that the most significant increases in plasma amino acids occur during days 9 to 12 is consistent with inhibition of utilization or protein synthesis by the liver and tissues at that time. In studies of normal
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.

Plasma amino-acid changes in chick embryos 455
embryos (Rupe and Farmer 1955), amino-acid concentrations and appearance have been shown not to be a result of the appearance of specific amino acids at a particular time. All amino acids accumulate in the embryo at the same rate, and the amounts of the amino acids, with the exception of histidine and valine, which show a slight decrease, remain constant throughout incubation. Free amino acids in the yolk, however, show progressive increase from the fourth to tenth day, and decrease after this until the twelfth day (Williams, DaCosta, Newman and Marshall 1954). This means that the embryo is mobilizing these compounds into protein during this period, and the occurrence of an increase in concentration after irradiation can reasonably be interpreted as the consequence of a disturbance in the mobilizing mechanism or mechanisms.
Accelerated proteolysis in the yolk sac or the white of the embryonate egg might be another explanation for the increased plasma amino-acid levels. It has been shown that some protease (cathepsin) inhibitors are inactivated by x-irradiation (Feinstein and Ballin 1953). If such mechanisms were operative in the chick embryo, more amino acids would be released than the growing embryo or tissues could utilize. Increased protein breakdown of embryonic tissue as a possible cause of these changes should also be considered.
Since changes in amino-acid levels are a function of two processes, proteolysis and protein synthesis, it is difficult to reach any conclusion as to where the abnormality may lie.
The adrenal is also functioning by the eighth dayof incubation, and the adrenal response may be a partial explanation of the changes seen. Administration of cortisone has been shown to cause a marked increase of hydroxyproline in the tissues of chick embryos (Roberts, Karnofsky and Frankel 1951). It has been postulated that the extensive protein breakdown found after administration of A C T H and cortisone is caused partly by inhibition of protein synthesis and partly by accelerated mobilization of protein (Katz and Hasterlik 1955).
Recent work (Kay and Entenman 1959, Jackson and Entenman 1958, Pentz 1958) has shown that adrenalectomy or splenectomy prior to irradiation does not prevent excessive taurine excretion in the rat after irradiation, though adrenalectomy does prevent post-irradiation increase in urea. In a recent review of the literature, one of these investigators (Pentz 1958) points out that " it is difficult to interpret the high taurine values reported for adrenalectomized animals after exposure to radiation "
Hydroxyproline and taurine have never been detected in the yolk of the developing embryo (Lowe 1954, Newman 1950). These, then, must result from the synthetic activity of embryonate tissues, and the increase of these two amino acids after x-irradiation would indicate either of the mechanisms discussed, inhibition of protein-synthesis or increase in proteolysis, as well as being a result of the adrenal response.
It is believed that the response of the 5-8 day embryos may be peculiar to developing embryonate tissue, while the response of the 9-19 day embryos indicates a basic metabolic disturbance. A possibility not to be excluded is that x-irradiation causes such stimulus to protein synthesis at this point in embryonic development that more amino acids are being used than under normal circum- stances.
The studies of the newly-hatched chick after lethal x-irradiation showed a plasma amino-acid pattern for normal animals similar to that found in the
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.

456 Elaine Katz Bernstein
embryo. X-irradiation did not affect the patterns in either case, although in newly-hatched chicks there was unusual and early excretion of some amino acids (taurine, valine, leucine, hydroxyproline and fi-alanine). This would seem to indicate a basic metabolic disturbance and not one that is peculiar to embryon- ate tissue. The amino-acid response to a smaller dose-level in the chick embryo is similar to the response obtained in man.
If the post-irradiation increase in free amino acids in the serum indicates a similar increase in the cells of the chorioallantoic membrane or the yolk sac, then the post-irradiation increase in rate of growth of Rickettsia mooseri (Greiff et al. 1957) is understandable. The stimulation of growth of the rickettsial organism may be observed days after irradiation, and it is clearly an effect on the embryo and not the rickettsiae. The increase in free amino acids in the chick embryo after irradiation also lasts for days. Furthermore, chick cells with larger amounts of available amino acids can be considered as better culture media for the rickettsial organism. As far as this evidence goes, then, the biochemical changes recorded here constitute a reasonable explanation for the increases in growth of R. mooseri observed after irradiation of chick embryos.
The contrary result observed with respect to the growth of R. akari after irradiation, namely the inhibition of its growth (Greiff et al. 1960), may be associated with the fact that R. ahari grows mainly in the nucleus and may not be in the region of increased available amino acids. In both instances, and especially in the first, experiments to test the role of amino acids for rickettsial growth suggest themselves.
ACKNOWLEDGMENTS
The author is grateful to Dr. E. L. Powers for many helpful discussions, and to Dr. John H. Pomeroy for his critical reading of the manuscript. She also wishes to thank Dr. Sylvanus Tyler, who performed the statistical analysis.
1. Les acides aminds plasmiques des embryons de poulet soumis h 600 r rayons x h 4 jours d'incubation ont dtd etudids du 5 e au 19 e jour.
2. La teneur en acides aminds ddcroit du 6 e au 8 e jour: cet effet est gdn4ral et non pas spdcifique pour aucun acide amin6 particulier.
3. La teneur en acides amin6s augmente au cours du 5 e jour et du 9 e au 19 e jour: ici encore l'effet est gdndral et non pas spdcifique pour aucun acide amind particulier. L'616vation du taux d'acides aminds libres entre 9 et 12 jours est hautement signifieative. Aucun acide amind anormal n'est prdsent.
4. Nous pensons que la rdaction des embryons entre 5 et 8 jours peut-etre propre au tissu embryonnaire en cours de d~veloppement, tandis que la rdaction entre 9 et 19 jours indique un ddrangement mdtabolique fondarnental et non spdcifique au tissu embryonnaire.
5. Les observations sugg6rent la possibilitd d'une relation entre l'augnlentation de la teneur en acides aminds suivant l'irradiation par les rayons x et l'aceroissement d'organismes de Rickettsia mooseri apr~s irradiation.
1. Die Arbeit bringt eine Untersuchung der Aminosaeuren im Plasma yon Huehner- embryonen vom 5. bis sum 9. Tag der Entwicklung nach Roentgenbestrahlung yon 600 r am vierten Tag der Bebruetung.
2. Eine Abnahme der Mengen der Arninosaeuren wurde bei den 6-8 Tage alten Embryonen gefunden. Dies war ein aUgemeiner Effekt ohne Spezifitaet fuer eine bestimmte Aminosaeure.
3. Eine Zunahme der Aminosaeuren wurde in den Embryonen vom 5. und vom 9. bis 12. Tag festgestellt, wieder als allgemeiner Effekt. Die Zunahme der freien Arninosaeuren in
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.

Plasma amino-acid changes in chick embryos 457
der Zeit vom 9. bis 12. Tag war statistisch signifikant. Keine nicht normaler Weiser vorkommenden Aminosaeuren waren vorhanden.
4. Es wird fuer moeglich gehalten, dass die embryonale Reaktion zwischen dem 5. und 8. Tag eine Besonderheit des embryonalen Gewebes sein koennte, waehrend die Reaktion zwisehen dem 9. und 19. Tag aug eine allgemeine Stoffwechselstoerung hinweist und nicht auf einen fuer das embryonale Gewebe eigentuemlichen Vorgang.
5. Line moegliche Beziehung zwischen der Zunahme der freien Aminiosaueren im Plasma nach der Bestrahlung und der Erhoehung des Wachstums yon Riekettsienbei Roentgenbestrahlung wird diskutiert.
REFERENCES
BERNSTEIN, E. K., STEARNER, S. P., and BRUES, A. M., 1956, Amer. J. Physiol., 186, 543. BEVERIDCE, W., and BURNET, F. M., 1946, The Cultivation of Viruses and Rickettsiae in the
Chick Embryo (London: H.M.S.O.), Medical Research Council, Special Report Series No. 256.
BOLAND, J., 1954, Brit. J. Radiol., 27, 680. FEINSTEIN, R. N., and BALLIN, J. C., 1953, Proc. Soc. exp. Biol., N.Y. , 83, 6. GREIFF, D., POWERS, E. L., and PINKERTON, H., 1957, J. exp. Med., 105, 217. GREIFF, D., POWERS, E. L., KISIELESKI, W., and PINKERTON, H., 1960, J. exp. Med., 111,
841. GUSTAESON, G. E., and KOLETSKY, S., 1952, Amer. J. Physiol., 171, 319. HALLESY, D. W., and DOULL, J., 1956, USAF Radiation Lab. Quart. Pr@ Rep. No. 20, 5l. HAMILTON, H. L., 1952, Lillie's Development of the Chick, 3rd edition (New York: Holt &
Co.). HEMPELMANN, L. H., LIsco, H., and HOFFMAN, J. G., 1952, Ann. Int. Med., 36, 279. HUNTER, C. G., 1957, Proc. Soc. exp. Biol., N.Y. , 97, 794. JACKSON, K. L., and ENTENMAN, C., 1958, Proc. Soc. exp. Biol., N.Y. , 97, 184. KATZ, E. J., and HASTERLIK, R. J., 1955, J. nat. Cancer Inst., 15, 1085. KATZ, E. J., and POWERS, E. L., 1955 a, Argonne National Laboratory Report, 5456, 7;
1955 b, Radiat. Res., 3, 331. KaY, R. E., and ENTENMAN, C., 1959, Radiat. Res., 11, 357. KAY, R. E., HARRIS, D. C., and ENTENMAN, C., 1956, Amer. J. Physiol., 186, 175. KOSZALm% T. R., BOWERMAN, C. I., and HEMPELMANN, L. H., 1955, Proc. Soc. exp. Biol.,
N.Y., 89, 662. KRISE, G. M., WILLIAMS, C. M., and ANDERSON, D. R., 1957, Proc. Soc. exp. Biol., N.Y. ,
95, 764. KUMTA, U. S., GUm'~ANI, S. U., and SAHASRABUDHE, M. B., 1957, J. Sci. industr. Res. C,
16, 179. LOWE, I. P., 1954, Patterns of Free Amino Acids and Related Compounds in Tissues of
Several Species. Doctoral Dissertation, Washington University (St. Louis, Mo.). MEFFERD, R. B., and MARTENS, H. H., 1955, Science, 122, 829. NEEDHAM, J., 1950, Biochemistry and iVIorphogenesis (Cambridge: University Press). NEW~aAN, R. E., 1950, Proc. Soc. exp. Biol., N.Y. , 75, 37. PATTEN, B. M., 1951, Early Embryology of the Chick, 4th edition (New York: Blakiston). PENTZ, E. I., 1958, J. Biol. Chem., 231, 165. ROBERTS, E., KARNOFSKY, D. A., and FRANKEL, A. S., 1951, Proc. Soc. exp. Biol., N.Y. , 76,
289. RVPE, C. O., and FARMER, C. J., 1955, J. biol. Chem., 213, 899. RUST, J. H., VISEK, W. J., and ROTH, L. J., 1957, Arch. Biochem. Biophys., 72, 189. WEBER, G., and CANTERO, A., 1957, Radiat. Res., 4, 459. WILLIAMS, C. M., KRISE, G. M., ANDERSON, D. R., and DOWLEN, R. M., 1957, Radiat. Res.,
7, 176. WILLIAMS, M. A., DACOSTA, W. A., NEWMAN, L. H., and MARSHALL, L. M., 1954, Nature,
Lond., 173, 490.
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B K
iel o
n 11
/06/
14Fo
r pe
rson
al u
se o
nly.


















