
Cellular Components of Nervous Tissue - Semantic Scholar · Cellular Components of Nervous Tissue...
17
CHAPTER 1 Cellular Components of Nervous Tissue Patrick R. Hof, Esther A. Nimchinsky, Grahame Kidd, Luz Claudio, and Bruce D. Trapp Several types of cellular elements are integrated to constitute normally functioning brain tissue. The neuron is the communicating cell, and many neuronal subtypes are connected to one another via complex circuitries, usually involving multiple synaptic con- nections. Neuronal physiology is supported and main- tained by neuroglial cells, which have highly diverse and incompletely understood functions. These include myelination, secretion of trophic factors, maintenance of the extracellular milieu, and scavenging of molec- ular and cellular debris from it. Neuroglial cells also participate in the formation and maintenance of the blood–brain barrier, a multicomponent structure that is interposed between the circulatory system and the brain substance and that serves as the molecular gateway to brain tissue. NEURONS The neuron is a highly specialized cell type and is the essential cellular element in the central nervous system (CNS). All neurological processes are dependent on complex cell–cell interactions between single neurons and/or groups of related neurons. Neurons can be cate- gorized according to their size, shape, neurochemical characteristics, location, and connectivity, which are important determinants of that particular functional role of the neuron in the brain. More importantly, neu- rons form circuits, and these circuits constitute the structural basis for brain function. Macrocircuits involve a population of neurons projecting from one brain region to another region, and microcircuits reflect the local cell–cell interactions within a brain region. The detailed analysis of these macro- and microcircuits is an essential step in understanding the neuronal basis of a given cortical function in the healthy and the dis- eased brain. Thus, these cellular characteristics allow us to appreciate the special structural and biochemical qualities of a neuron in relation to its neighbors and to place it in the context of a specific neuronal subset, circuit, or function. Broadly speaking, therefore, there are five general categories of neurons: inhibitory neurons that make local contacts (e.g., GABAergic interneurons in the cerebral and cerebellar cortex), inhibitory neurons that make distant contacts (e.g., medium spiny neurons of the basal ganglia or Purkinje cells of the cerebellar cor- tex), excitatory neurons that make local contacts (e.g., spiny stellate cells of the cerebral cortex), excitatory neurons that make distant contacts (eg., pyramidal neu- rons in the cerebral cortex), and neuromodulatory neurons that influence neurotransmission, often at large distances. Within these general classes, the struc- tural variation of neurons is systematic, and careful analyses of the anatomic features of neurons have led to various categorizations and to the development of the concept of cell type. The grouping of neurons into descriptive cell types (such as chandelier, double bouquet, or bipolar cells) allows the analysis of popu- lations of neurons and the linking of specified cellular characteristics with certain functional roles. General Features of Neuronal Morphology Neurons are highly polarized cells, meaning that they develop distinct subcellular domains that sub- serve different functions. Morphologically, in a typical 1 From Molecules to Networks, Second Edition Copyright # 2009, Elsevier Inc. All rights reserved.
-
Upload
nguyenminh -
Category
Documents
-
view
220 -
download
0
Transcript of Cellular Components of Nervous Tissue - Semantic Scholar · Cellular Components of Nervous Tissue...

C H A P T E R
1
Cellular Components of Nervous Tissue
Patrick R. Hof, Esther A. Nimchinsky, Grahame Kidd,
Luz Claudio, and Bruce D. Trapp
Several types of cellular elements are integratedto constitute normally functioning brain tissue. Theneuron is the communicating cell, and many neuronalsubtypes are connected to one another via complexcircuitries, usually involving multiple synaptic con-nections. Neuronal physiology is supported andmain-tained by neuroglial cells, which have highly diverseand incompletely understood functions. These includemyelination, secretion of trophic factors, maintenanceof the extracellular milieu, and scavenging of molec-ular and cellular debris from it. Neuroglial cells alsoparticipate in the formation and maintenance of theblood–brain barrier, a multicomponent structure thatis interposed between the circulatory system andthe brain substance and that serves as the moleculargateway to brain tissue.
NEURONS
The neuron is a highly specialized cell type and is theessential cellular element in the central nervous system(CNS). All neurological processes are dependent oncomplex cell–cell interactions between single neuronsand/or groups of related neurons. Neurons can be cate-gorized according to their size, shape, neurochemicalcharacteristics, location, and connectivity, which areimportant determinants of that particular functionalrole of the neuron in the brain. More importantly, neu-rons form circuits, and these circuits constitute thestructural basis for brain function.Macrocircuits involvea population of neurons projecting from one brainregion to another region, and microcircuits reflect the
local cell–cell interactions within a brain region. Thedetailed analysis of these macro- and microcircuits isan essential step in understanding the neuronal basisof a given cortical function in the healthy and the dis-eased brain. Thus, these cellular characteristics allowus to appreciate the special structural and biochemicalqualities of a neuron in relation to its neighbors andto place it in the context of a specific neuronal subset,circuit, or function.
Broadly speaking, therefore, there are five generalcategories of neurons: inhibitory neurons that makelocal contacts (e.g., GABAergic interneurons in thecerebral and cerebellar cortex), inhibitory neurons thatmake distant contacts (e.g., medium spiny neurons ofthe basal ganglia or Purkinje cells of the cerebellar cor-tex), excitatory neurons that make local contacts (e.g.,spiny stellate cells of the cerebral cortex), excitatoryneurons thatmakedistant contacts (eg., pyramidalneu-rons in the cerebral cortex), and neuromodulatoryneurons that influence neurotransmission, often atlarge distances. Within these general classes, the struc-tural variation of neurons is systematic, and carefulanalyses of the anatomic features of neurons haveled to various categorizations and to the developmentof the concept of cell type. The grouping of neuronsinto descriptive cell types (such as chandelier, doublebouquet, or bipolar cells) allows the analysis of popu-lations of neurons and the linking of specified cellularcharacteristics with certain functional roles.
General Features of Neuronal Morphology
Neurons are highly polarized cells, meaning thatthey develop distinct subcellular domains that sub-serve different functions. Morphologically, in a typical
1From Molecules to Networks, Second Edition Copyright # 2009, Elsevier Inc. All rights reserved.
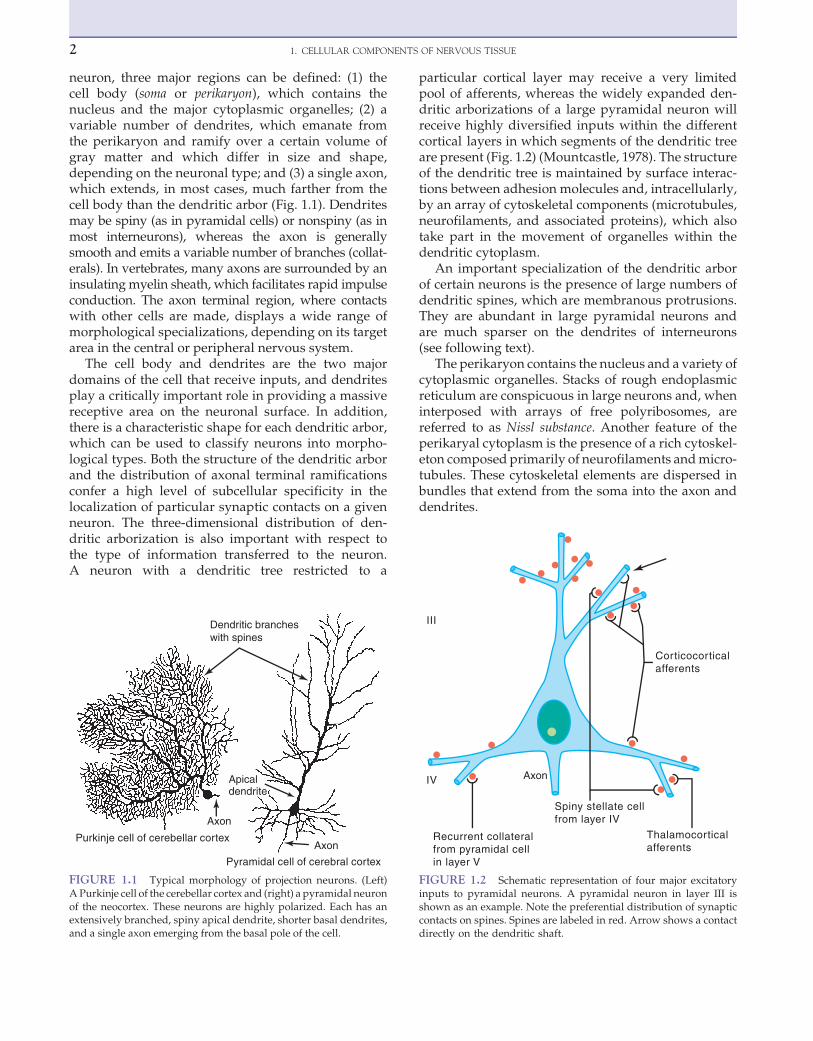
neuron, three major regions can be defined: (1) thecell body (soma or perikaryon), which contains thenucleus and the major cytoplasmic organelles; (2) avariable number of dendrites, which emanate fromthe perikaryon and ramify over a certain volume ofgray matter and which differ in size and shape,depending on the neuronal type; and (3) a single axon,which extends, in most cases, much farther from thecell body than the dendritic arbor (Fig. 1.1). Dendritesmay be spiny (as in pyramidal cells) or nonspiny (as inmost interneurons), whereas the axon is generallysmooth and emits a variable number of branches (collat-erals). In vertebrates, many axons are surrounded by aninsulatingmyelin sheath, which facilitates rapid impulseconduction. The axon terminal region, where contactswith other cells are made, displays a wide range ofmorphological specializations, depending on its targetarea in the central or peripheral nervous system.
The cell body and dendrites are the two majordomains of the cell that receive inputs, and dendritesplay a critically important role in providing a massivereceptive area on the neuronal surface. In addition,there is a characteristic shape for each dendritic arbor,which can be used to classify neurons into morpho-logical types. Both the structure of the dendritic arborand the distribution of axonal terminal ramificationsconfer a high level of subcellular specificity in thelocalization of particular synaptic contacts on a givenneuron. The three-dimensional distribution of den-dritic arborization is also important with respect tothe type of information transferred to the neuron.A neuron with a dendritic tree restricted to a
particular cortical layer may receive a very limitedpool of afferents, whereas the widely expanded den-dritic arborizations of a large pyramidal neuron willreceive highly diversified inputs within the differentcortical layers in which segments of the dendritic treeare present (Fig. 1.2) (Mountcastle, 1978). The structureof the dendritic tree is maintained by surface interac-tions between adhesion molecules and, intracellularly,by an array of cytoskeletal components (microtubules,neurofilaments, and associated proteins), which alsotake part in the movement of organelles within thedendritic cytoplasm.
An important specialization of the dendritic arborof certain neurons is the presence of large numbers ofdendritic spines, which are membranous protrusions.They are abundant in large pyramidal neurons andare much sparser on the dendrites of interneurons(see following text).
The perikaryon contains the nucleus and a variety ofcytoplasmic organelles. Stacks of rough endoplasmicreticulum are conspicuous in large neurons and, wheninterposed with arrays of free polyribosomes, arereferred to as Nissl substance. Another feature of theperikaryal cytoplasm is the presence of a rich cytoskel-eton composed primarily of neurofilaments andmicro-tubules. These cytoskeletal elements are dispersed inbundles that extend from the soma into the axon anddendrites.
Purkinje cell of cerebellar cortex
Pyramidal cell of cerebral cortex
Axon
Axon
Apicaldendrite
Dendritic brancheswith spines
FIGURE 1.1 Typical morphology of projection neurons. (Left)A Purkinje cell of the cerebellar cortex and (right) a pyramidal neuronof the neocortex. These neurons are highly polarized. Each has anextensively branched, spiny apical dendrite, shorter basal dendrites,and a single axon emerging from the basal pole of the cell.
Thalamocorticalafferents
Corticocorticalafferents
Recurrent collateralfrom pyramidal cellin layer V
Spiny stellate cellfrom layer IV
Axon
III
IV
FIGURE 1.2 Schematic representation of four major excitatoryinputs to pyramidal neurons. A pyramidal neuron in layer III isshown as an example. Note the preferential distribution of synapticcontacts on spines. Spines are labeled in red. Arrow shows a contactdirectly on the dendritic shaft.
2 1. CELLULAR COMPONENTS OF NERVOUS TISSUE

Whereas dendrites and the cell body can be charac-terized as domains of the neuron that receive afferents,the axon, at the other pole of the neuron, is responsiblefor transmitting neural information. This informationmaybeprimary, in the case of a sensory receptor, or pro-cessed information that has already been modifiedthrough a series of integrative steps. The morphologyof the axon and its course through the nervous systemare correlated with the type of information processedby the particular neuron and by its connectivity patternswith other neurons. The axon leaves the cell body fromasmall swelling called the axon hillock. This structure isparticularly apparent in large pyramidal neurons; inother cell types, the axon sometimes emerges from oneof themain dendrites. At the axon hillock, microtubulesare packed into bundles that enter the axon as parallelfascicles. The axon hillock is the part of the neuronwhere the action potential is generated. The axon is gen-erally unmyelinated in local circuit neurons (such asinhibitory interneurons), but it is myelinated in neuronsthat furnish connections between different parts of thenervous system. Axons usually have higher numbersof neurofilaments than dendrites, although this distinc-tion can be difficult to make in small elements that con-tain fewer neurofilaments. In addition, the axonmay beextremely ramified, as in certain local circuit neurons; itmay give out a large number of recurrent collaterals, asin neurons connecting different cortical regions, or itmay be relatively straight in the case of projections to
subcortical centers, as in cortical motor neurons thatsend their very long axons to the ventral horn of the spi-nal cord. At the interface of axon terminals with targetcells are the synapses, which represent specializedzones of contact consisting of a presynaptic (axonal) ele-ment, a narrow synaptic cleft, and a postsynaptic ele-ment on a dendrite or perikaryon.
Synapses and Spines
Synapses
Each synapse is a complex of several components:(1) a presynaptic element, (2) a cleft, and (3) a postsynapticelement. The presynaptic element is a specialized part ofthe presynaptic neuron’s axon, the postsynapticelement is a specialized part of the postsynapticsomatodendritic membrane, and the space betweenthese two closely apposed elements is the cleft. Theportion of the axon that participates in the axon isthe bouton, and it is identified by the presence of syn-aptic vesicles and a presynaptic thickening at theactive zone (Fig. 1.3). The postsynaptic element ismarked by a postsynaptic thickening opposite thepresynaptic thickening. When both sides are equallythick, the synapse is referred to as symmetric. Whenthe postsynaptic thickening is greater, the synapse isasymmetric. Edward George Gray noticed this differ-ence, and divided synapses into two types: Gray’stype 1 synapses are symmetric and have variably
FIGURE 1.3 Ultrastructure of dendritic spines (S) and synapses in the human brain. Note the narrow spine necks (asterisks) emanating fromthemain dendritic shaft (D) and the spine head containing filamentousmaterial, and the cisterns of the spine apparatus particularly visible in thelower panel spine. The arrows on the left panels point to postsynaptic densities of asymmetric excitatory synapses (arrows). The apposed axonalboutons (B) are characterized by round synaptic vesicles. A perforated synapse is shown on the lower left panel. The panel at right shows twosymmetric inhibitory synapses (arrowheads) on a large dendritic shaft (D). In this case the axonal boutons (B) contain some ovoid vesicles com-pared to the ones in asymmetric synapses. The dendrites and axons contain numerousmitochondria (m). Scale bar¼ 1 mm. Electronmicrographscourtesy of Drs. S.A. Kirov and M. Witcher (Medical College of Georgia), and K.M. Harris (University of Texas – Austin).
3NEURONS

shaped, or pleomorphic, vesicles. Gray’s type2 synapses are asymmetric and have clear, round vesi-cles. The significance of this distinction is that researchhas shown that, in general, Gray’s type 1 synapses tendto be inhibitory, while Gray’s type 2 synapses tendto be excitatory. This correlation greatly enhanced theusefulness of electron microscopy in neuroscience.
In cross section on electron micrographs, a synapselooks like two parallel lines separated by a very nar-row space (Fig. 1.3). Viewed from the inside of theaxon or dendrite, it looks like a patch of variableshape. Some synapses are a simple patch, or macule.Macular synapses can grow fairly large, reaching dia-meters over 1 mm. The largest synapses have discon-tinuities or holes within the macule and are calledperforated synapses (Fig. 1.3). In cross section, aperforated synapse may resemble a simple macularsynapse or several closely spaced smaller macules.
The portion of the presynaptic element that isapposed to the postsynaptic element is the active zone.This is the region where the synaptic vesicles areconcentrated and where, at any time, a small numberof vesicles are docked and presumably ready forfusion. The active zone is also enriched with voltagegated calcium channels, which are necessary to per-mit activity-dependent fusion and neurotransmitterrelease.
The synaptic cleft is truly a space, but its propertiesare essential. The width of the cleft (�20 mm) is criticalbecause it defines the volume in which each vesiclereleases its contents, and therefore, the peak con-centration of neurotransmitter upon release. On theflanks of the synapse, the cleft is spanned by adhesionmolecules, which are believed to stabilize the cleft.
The postsynaptic elementmay be a portion of a somaor a dendrite, or rarely, part of an axon. In the cerebralcortex, most Gray’s type 1 synapses are located onsomata or dendritic shafts, while most Gray’s type2 synapses are located on dendritic spines, which arespecialized protrusions of the dendrite. A similar segre-gation is seen in cerebellar cortex. In nonspiny neurons,symmetric and asymmetric synapses are often less wellseparated. Irrespective of location, a postsynapticthickening marks the postsynaptic element. In Gray’stype 2 synapses, the postsynaptic thickening (or post-synaptic density, PSD), is greatly enhanced. Amongthe molecules that are associated with the PSD are neu-rotransmitter receptors (e.g., NMDA receptors) andmolecules with less obvious function, such as PSD-95.
Spines
Spines are protrusions on the dendritic shafts ofsome types of neurons and are the sites of synaptic con-tacts, usually excitatory. Use of the silver impregnationtechniques of Golgi or of the methylene blue used by
Ehrlich in the late nineteenth century led to the discov-ery of spiny appendages on dendrites of a variety ofneurons. The best known are those on pyramidal neu-rons and Purkinje cells, although spines occur on neu-ron types at all levels of the central nervous system.In 1896, Berkley observed that terminal axonal boutonswere closely apposed to spines and suggested thatspines may be involved in conducting impulses fromneuron to neuron. In 1904, Santiago Ramon yCajal sug-gested that spines could collect the electrical chargeresulting from neuronal activity. He also noted thatspines substantially increase the receptive surface ofthe dendritic arbor, which may represent an importantfactor in receiving the contacts made by the axonalterminals of other neurons. It has been calculated thatthe approximately 20,000 spines of a pyramidal neuronaccount for more than 40% of its total surface area(Peters et al., 1991).
More recent analyses of spine electrical propertieshave demonstrated that spines are dynamic struc-tures that can regulate many neurochemical eventsrelated to synaptic transmission and modulate synap-tic efficacy. Spines are also known to undergo patho-logic alterations and have a reduced density in anumber of experimental manipulations (such as dep-rivation of a sensory input) and in many develop-mental, neurologic, and psychiatric conditions (suchas dementing illnesses, chronic alcoholism, schizo-phrenia, trisomy 21). Morphologically, spines arecharacterized by a narrower portion emanating fromthe dendritic shaft, the neck, and an ovoid bulb orhead, although spine morphology may vary fromlarge mushroom-shaped bulbs to small bulges barelydiscernable on the surface of the dendrite. Spineshave an average length of �2 mm, but there is consid-erable variability in their dimensions. At the ultra-structural level (Fig. 1.3), spines are characterized bythe presence of asymmetric synapses and contain fineand quite indistinct filaments. These filaments mostlikely consist of actin and a- and b-tubulins. Microtu-bules and neurofilaments present in dendritic shaftsdo not enter spines. Mitochondria and free ribosomesare infrequent, although many spines contain polyri-bosomes in their neck. Interestingly, most polyribo-somes in dendrites are located at the bases of spines,where they are associated with endoplasmic reticu-lum, indicating that spines possess the machinerynecessary for the local synthesis of proteins. Anotherfeature of the spine is the presence of confluent tubu-lar cisterns in the spine head that represent an exten-sion of the dendritic smooth endoplasmic reticulum.Those cisterns are referred to as the spine apparatus.The function of the spine apparatus is not fullyunderstood but may be related to the storage ofcalcium ions during synaptic transmission.
4 1. CELLULAR COMPONENTS OF NERVOUS TISSUE

Specific Examples of DifferentNeuronal Types
Inhibitory Local Circuit Neurons
Inhibitory Interneurons of the Cerebral Cortex Alarge variety of inhibitory interneuron types is presentin the cerebral cortex and in subcortical structures.These neurons contain the inhibitory neurotransmitterg-aminobutyric acid (GABA) and exert strong localinhibitory effects. Their dendritic and axonal arboriza-tions offer important clues as to their role in the regula-tion of pyramidal cell function. In addition, for severalGABAergic interneurons, a subtype of a given mor-phologic class can be defined further by a particularset of neurochemical characteristics. Interneurons havebeen extensively characterized in the neocortex andhippocampus of rodents and primates, but they arepresent throughout the cerebral gray matter andexhibit a rich variety of morphologies, depending onthe brain region as well as on the species studied.
In the neocortex and hippocampus, the targets andmorphologies of interneuron axons are most usefullyclassified into morphological and functional groups. Forexample, basket cells have axonal endings surroundingpyramidal cell somata (Somogyi et al., 1983) and providemost of the inhibitory GABAergic synapses to the somasand proximal dendrites of pyramidal cells. These cellsare also characterized by certain biochemical features inthat the majority of them contain the calcium-bindingprotein parvalbumin, and cholecystokinin appears to bethe most likely neuropeptide in large basket cells.
Chandelier cells have spatially restricted axon ter-minals that look like vertically oriented “cartridges,”each consisting of a series of axonal boutons, or swel-lings, linked together by thin connecting pieces. Theseneurons synapse exclusively on the axon initial seg-ment of pyramidal cells (this cell is also known asaxoaxonic cell), and because the strength of the synapticinput is correlated directly with its proximity to theaxon initial segment, there can be no more powerfulinhibitory input to a pyramidal cell than that of thechandelier cell (Freund et al., 1983; DeFelipe et al., 1989).
The double bouquet cells are characterized by a verti-cal bitufteddendritic tree and a tight bundle of verticallyoriented varicose axon collaterals (Somogyi andCowey,1981). There are several subclasses of double bouquetcells based on the complement of calcium-binding pro-tein and neuropeptide they contain. Their axons contactspines and dendritic shafts of pyramidal cells, as well asdendrites from non-pyramidal neurons.
Inhibitory Projection Neurons
Medium-Sized Spiny Cells These neurons areunique to the striatum, a part of the basal ganglia thatcomprises the caudate nucleus and putamen. Medium-
sized spiny cells are scattered throughout the caudatenucleus, and putamen and are recognized by theirrelatively large size compared with other cellularelements of the basal ganglia, and by the fact that theyare generally isolated neurons. They differ from allothers in the striatum in that they have a highly rami-fied dendritic arborization radiating in all directionsand densely covered with spines. They furnish a majoroutput from the caudate nucleus and putamen andreceive a highly diverse input from, among othersources, the cerebral cortex, thalamus, and certaindopaminergic neurons of the substantia nigra. Theseneurons are neurochemically quite heterogeneous,contain GABA, and may contain several neuropep-tides and the calcium-binding protein calbindin. InHuntington disease, a neurodegenerative disorder ofthe striatum characterized by involuntary movementsand progressive dementia, an early and dramatic lossof medium-sized spiny cells occurs.
Purkinje Cells Purkinje cells are the most salient cel-lular elements of the cerebellar cortex. They arearranged in a single row throughout the entire cerebel-lar cortex between the molecular (outer) layer and thegranular (inner) layer. They are among the largest neu-rons and have a round perikaryon, classically describedas shaped “like a chianti bottle,” with a highly brancheddendritic tree shaped like a candelabrum and extendinginto the molecular layer, where they are contacted byincoming systems of afferent fibers from granule neu-rons and the brainstem. The apical dendrites of Purkinjecells have an enormous number of spines (more than80,000 per cell). A particular feature of the dendritic treeof the Purkinje cell is that it is distributed in one plane,perpendicular to the longitudinal axes of the cerebellarfolds, and each dendritic arbor determines a separatedomain of cerebellar cortex (Fig. 1.1). The axons of Pur-kinje neurons course through the cerebellar white mat-ter and contact deep cerebellar nuclei or vestibularnuclei. These neurons contain the inhibitory neuro-transmitter GABA and the calcium-binding proteincalbindin. Spinocerebellar ataxia, a severedisorder com-bining ataxic gait and impairment of fine hand move-ments, accompanied by dysarthria and tremor, hasbeen documented in some families and is relateddirectly to Purkinje cell degeneration.
Excitatory Local Circuit Neurons
Spiny Stellate Cells Spiny stellate cells are smallmultipolar neurons with local dendritic and axonalarborizations. These neurons resemble pyramidal cellsin that they are the only other cortical neurons withlarge numbers of dendritic spines, but they differ frompyramidal neurons in that they lack an elaborate apicaldendrite. The relatively restricted dendritic arbor of
5NEURONS

these neurons is presumably amanifestation of the factthat they are high-resolution neurons that gather affer-ents to a very restricted region of the cortex. Dendritesrarely leave the layer in which the cell body resides.The spiny stellate cell also resembles the pyramidal cellin that it provides asymmetric synapses that are pre-sumed to be excitatory, and is thought to use glutamateas its neurotransmitter (Peters and Jones, 1984).
The axons of spiny stellate neurons have primarilyintracortical targets and a radial orientation, andappear to play an important role in forming linksamong layer IV, the major thalamorecipient layer,and layers III, V, and VI, the major projection layers.The spiny stellate neuron appears to function as ahigh-fidelity relay of thalamic inputs, maintainingstrict topographic organization and setting up initialvertical links of information transfer within a givencortical area (Peters and Jones, 1984).
Excitatory Projection Neurons
Pyramidal Cells All cortical output is carried bypyramidal neurons, and the intrinsic activity of theneocortex can be viewed simply as a means of finelytuning their output. A pyramidal cell is a highly polar-ized neuron, with a major orientation axis perpendicu-lar (or orthogonal) to the pial surface of the cerebralcortex. In cross section, the cell body is roughly trian-gular (Fig. 1.1), although a large variety ofmorphologictypes exist with elongate, horizontal, or verticalfusiform, or inverted perikaryal shapes. Pyramidalcells are the major excitatory type of neurons and useglutamate as their neurotransmitter. A pyramidalneuron typically has a large number of dendrites thatemanate from the apex and form the base of the cellbody. The span of the dendritic tree depends on thelaminar localization of the cell body, but it may, as ingiant pyramidal neurons, spread over several milli-meters. The cell body and dendritic arborization maybe restricted to a few layers or, in some cases, may spanthe entire cortical thickness (Jones, 1984).
In most cases, the axon of a large pyramidal cellextends from the base of the perikaryon and coursestoward the subcortical white matter, giving off severalcollateral branches that are directed to cortical domainsgenerally located within the vicinity of the cell of origin(as explained later). Typically, a pyramidal cell has alarge nucleus, and a cytoplasmic rim that contains, par-ticularly in large pyramidal cells, a collection of granularmaterial chiefly composed of lipofuscin. Although allpyramidal cells possess these general features, they canalso be subdivided into numerous classes based on theirmorphology, laminar location, and connectivity withcortical and subcortical regions (Fig. 1.4) (Jones, 1975).
Spinal Motor Neurons Motor cells of the ventralhorns of the spinal cord, also called a motoneurons,have their cell bodies within the spinal cord and sendtheir axons outside the central nervous system to inner-vate the muscles. Different types of motor neurons aredistinguished by their targets. The a motoneuronsinnervate skeletal muscles, but smaller motor neurons(the g motoneurons, forming about 30% of the motorneurons) innervate the spindle organs of the muscles.The a motor neurons are some of the largest neuronsin the entire central nervous system and are character-ized by a multipolar perikaryon and a very rich cyto-plasm that renders them very conspicuous onhistological preparations. They have a large numberof spiny dendrites that arborize locally within the ven-tral horn. The a motoneuron axon leaves the centralnervous system through the ventral root of the periph-eral nerves. Their distribution in the ventral horn is notrandom and corresponds to a somatotopic representa-tion of themuscle groups of the limbs and axial muscu-lature (Brodal, 1981). Spinal motor neurons use
A B C D
ThalamusCorticocorticalClaustrum(Callosal)
Spinal cordPonsMedullaTectumThalamusRed nucleusStriatum(Cortical)
CallosalCorticocortical
Corticocortical
I
II
III
IV
V
VI
FIGURE 1.4 Morphology and distribution of neocortical pyra-midal neurons. Note the variability in cell size and dendritic arbor-ization, as well as the presence of axon collaterals, depending onthe laminar localization (I–VI) of the neuron. Also, different typesof pyramidal neurons with a precise laminar distribution projectto different regions of the brain. Adapted from Jones (1984).
6 1. CELLULAR COMPONENTS OF NERVOUS TISSUE

acetylcholine as their neurotransmitter. Large motorneurons are severely affected in lower motor neurondisease, a neurodegenerative disorder characterizedby progressive muscular weakness that affects, at first,one or two limbs but involves more and more of thebody musculature, which shows signs of wasting as aresult of denervation.
Neuromodulatory Neurons
Dopaminergic Neurons of the Substantia NigraDopaminergic neurons are large neurons that residemostly within the pars compacta of the substantianigra and in the ventral tegmental area. A distinctivefeature of these cells is the presence of a pigment,neuromelanin, in compact granules in the cytoplasm.These neurons are medium-sized to large, fusiform,and frequently elongated. They have several largeradiating dendrites. The axon emerges from the cellbody or from one of the dendrites and projects tolarge expanses of cerebral cortex and to the basalganglia. These neurons contain the catecholamine-synthesizing enzyme tyrosine hydroxylase, as well asthe monoamine dopamine as their neurotransmitter.Some of them contain both calbindin and calretinin.These neurons are affected severely and selectivelyin Parkinson disease—a movement disorder differentfrom Huntington disease and characterized by restingtremor and rigidity—and their specific loss is theneuropathologic hallmark of this disorder.
NEUROGLIA
The term neuroglia, or “nerve glue,” was coined in1859 by Rudolph Virchow, who erroneously con-ceived of the neuroglia as an inactive “connective tis-sue” holding neurons together in the central nervoussystem. The metallic staining techniques developedby Ramon y Cajal and del Rio-Hortega allowed thesetwo great pioneers to distinguish, in addition to theependyma lining the ventricles and central canal,three types of supporting cells in the CNS: oligoden-drocytes, astrocytes, and microglia. In the peripheralnervous system (PNS), the Schwann cell is the majorneuroglial component.
Oligodendrocytes and Schwann CellsSynthesize Myelin
Most brain functions depend on rapid communica-tion between circuits of neurons. As shown in depthlater, there is a practical limit to how fast an individualbare axon can conduct an action potential. Organisms
developed two solutions for enhancing rapid commu-nication between neurons and their effector organs.In invertebrates, the diameters of axons are enlarged.In vertebrates, the myelin sheath (Fig. 1.5) evolved topermit rapid nerve conduction.
Axon enlargement accelerates the rate of conductionof the action potential in proportion to the square root ofaxonal diameter. Thus small axons conduct at slowerrates than larger ones. The largest axon in the inverte-brate kingdom is the squid giant axon, which is aboutthe thickness of amechanical pencil lead. This axon con-ducts the action potential at speeds of 10–20m/s. As theaxon mediates an escape reflex, firing must be rapid ifthe animal is to survive. Bare axons and continuous con-duction obviously provide sufficient rates of signalpropagation for even very large invertebrates, such asthe giant squid, and many human axons also remainbare. However, in the human brain with 10 billion neu-rons, axons cannot be as thick as pencil lead, otherwisehuman heads would weigh 100 pounds or more.
Thus, along the invertebrate evolutionary line, theuse of bare axons imposes a natural, insurmountablelimit— a constraint of axonal size—to increasing theprocessing capacity of the nervous system. Verte-brates, however, get around this problem throughevolution of the myelin sheath, which allows 10- to100-fold increases in conduction of the nerve impulsealong axons with fairly minute diameters.
In the central nervous system, myelin sheaths(Fig. 1.6) are elaborated by oligodendrocytes. Duringbrain development, these glial cells send out a fewcytoplasmic processes that engage adjacent axonsand form myelin around them (Bunge, 1968). Myelinconsists of a long sheet of oligodendrocyte plasmamembrane, which is spirally wrapped around an axo-nal segment. At the end of each myelin segment,there is a bare portion of the axon, the node of Ranvier.Myelin segments are thus called “internodes.” Phys-iologically, myelin has insulating properties suchthat the action potential can “leap” from node tonode and therefore does not have to be regeneratedcontinually along the axonal segment that is coveredby the myelin membrane sheath. This leaping of theaction potential from node to node allows axons withfairly small diameters to conduct extremely rapidly(Ritchie, 1984) and is called “saltatory” conduction.
Because the brain and spinal cord are encased inthe bony skull and vertebrae, CNS evolution haspromoted compactness among the supporting cellsof the CNS. Each oligodendrocyte cell body is respon-sible for the construction and maintenance of severalmyelin sheaths (Fig. 1.6), thus reducing the numberof glial cells required. In both PNS and CNS myelin,cytoplasm is removed between each turn of the
7NEUROGLIA

myelin, leaving only the thinnest layer of plasmamembrane. Due to protein composition differences,CNS lamellae are approximately 30% thinner than inPNS myelin. In addition, there is little or no extracel-lular space or extracellular matrix between the mye-linated axons passing through CNS white matter.Brain volume is thus reserved for further expansionof neuronal populations.
Peripheral nerves pass between moving musclesand around major joints and are routinely exposed tophysical trauma. A hard tackle, slipping on an icy side-walk, or even just occupying the same uncomfortableseating posture for too long can painfully compressperipheral nerves and potentially damage them. Thus,evolutionary pressures shaping the PNS favor
robustness and regeneration rather than conservationof space. Myelin in the PNS is generated by Schwanncells (Fig. 1.7), which are different from oligodendro-cytes in severalways. Individualmyelinating Schwanncells form a single internode. The biochemical compo-sition of PNS and CNS myelin differs, as discussed infollowing text. Unlike oligodendrocytes, Schwann cellssecrete copious extracellular matrix components andproduce a basal lamina “sleeve” that runs the entirelength of myelinated axons. Schwann cell and fibro-blast-derived collagens prevent normal wear-and-tearcompression damage. Schwann cells also respond vig-orously to injury, in common with astrocytes butunlike oligodendrocytes. Schwann cell growth factorsecretion, debris removal by Schwann cells after injury,
FIGURE 1.5 An electron micrograph of atransverse-section through part of a myelinatedaxon from the sciatic nerve of a rat. The tightlycompacted multilayer myelin sheath (My) sur-rounds and insulates the axon (Ax). Mit, mito-chondria. Scale bar: 75 nm.
8 1. CELLULAR COMPONENTS OF NERVOUS TISSUE

and the axonal guidance function of the basal laminaare responsible for the exceptional regenerative capac-ity of the PNS compared with the CNS.
The major integral membrane protein of peripheralnerve myelin is protein zero (P0), a member of a verylarge family of proteins termed the immunoglobulingene superfamily. This protein makes up about 80% ofthe protein complement of PNS myelin. Interactionsbetween the extracellular domains of P0 moleculesexpressed on one layer of the myelin sheath withthose of the apposing layer yield a characteristicregular periodicity that can be seen by thin-sectionelectron microscopy (Fig. 1.5). This zone, called theintraperiod line, represents the extracellular appositionof the myelin bilayer as it wraps around itself. On theother side of the bilayer, the cytoplasmic side, thehighly charged P0 cytoplasmic domain probablyfunctions to neutralize the negative charges on thepolar head groups of the phospholipids that makeup the plasma membrane itself, allowing the mem-branes of the myelin sheath to come into close apposi-tion with one another. In electron microscopy, thiscytoplasmic apposition is a bit darker than the intra-period line and is termed the major dense line.In peripheral nerves, although other molecules are
FIGURE 1.6 An oligodendrocyte (OL) in the central nervous system is depicted myelinating several axon segments. A cutaway view ofthe myelin sheath is shown (M). Note that the internode of myelin terminates in paranodal loops that flank the node of Ranvier (N).(Inset) An enlargement of compact myelin with alternating dark and light electron-dense lines that represent intracellular (major dense lines)and extracellular (intraperiod line) plasma membrane appositions, respectively.
FIGURE 1.7 An “unrolled” Schwann cell in the PNS is illus-trated in relation to the single axon segment that it myelinates.The broad stippled region is compact myelin surrounded by cyto-plasmic channels that remain open even after compact myelin hasformed, allowing an exchange of materials among the myelinsheath, the Schwann cell cytoplasm, and perhaps the axon as well.
9NEUROGLIA

present in small quantities in compact myelin andmay have important functions, compaction (i.e., theclose apposition of membrane surfaces without inter-vening cytoplasm) is accomplished solely by P0–P0interactions at both extracellular and intracellular(cytoplasmic) surfaces.
Curiously, P0 is present in the CNS of lower verte-brates such as sharks and bony fish, but in terrestrialvertebrates (reptiles, birds, and mammals), P0 is lim-ited to the PNS. CNS myelin compaction in thesehigher organisms is subserved by proteolipid protein(PLP) and its alternate splice form, DM-20. These twoproteins are generated from the same gene, both spanthe plasma membrane four times, and they differ onlyin that PLP has a small, positively charged segmentexposed on the cytoplasmic surface. Why did PLP–DM-20 replace P0 in CNS myelin? Manipulation ofPLP and P0 content of CNS myelin established an axo-notrophic function for PLP in CNS myelin. Removal ofPLP from rodent CNS myelin altered the periodicity ofcompact myelin and produced a late-onset axonaldegeneration (Griffiths et al., 1998). Replacing PLP withP0 in rodent CNSmyelin stabilized compact myelin butenhanced the axonal degeneration (Yin et al., 2006).These and other observations in primary demyelinationand inherited myelin diseases have established axonaldegeneration as themajor cause of permanent disabilityin diseases such as multiple sclerosis.
Myelin membranes also contain a number of otherproteins such as the myelin basic protein, which is amajor CNS myelin component, and PMP-22, a proteinthat involved in a form of peripheral nerve disease.A large number of naturally occurring gene muta-tions can affect the proteins specific to the myelinsheath and cause neurological disease. In animals,these mutations have been named according to thephenotype that is produced: the shiverer mouse, theshaking pup, the rumpshaker mouse, the jimpymouse, the myelin-deficient rat, the quaking mouse,and so forth. Many of these mutations are well charac-terized and have provided valuable insights into therole of individual proteins in myelin formation andaxonal survival.
Astrocytes Play Important Roles in CNSHomeostasis
As the name suggests, astrocytes are star-shaped,process-bearing cells distributed throughout the centralnervous system. They constitute from 20 to 50% of thevolume of most brain areas. Astrocytes come in manyshapes and forms. The two main forms, protoplasmicand fibrous astrocytes, predominate in gray and whitematter, respectively (Fig. 1.8). Embryonically, astrocytes
develop from radial glial cells, which transversely com-partmentalize the neural tube. Radial glial cells serve asscaffolding for the migration of neurons and play a crit-ical role in defining the cytoarchitecture of the CNS(Fig. 1.9). As the CNS matures, radial glia retract theirprocesses and serve as progenitors of astrocytes. How-ever, some specialized astrocytes of a radial natureare still found in the adult cerebellum and the retinaand are known as Bergmann glial cells and Muller cells,respectively.
Astrocytes “fence in” neurons and oligodendro-cytes. Astrocytes achieve this isolation of the brainparenchyma by extending long processes projectingto the pia mater and the ependyma to form the glialimitans, by covering the surface of capillaries andby making a cuff around the nodes of Ranvier. Theyalso ensheath synapses and dendrites and projectprocesses to cell somas (Fig. 1.10). Astrocytes areconnected to each other by gap junctions, forming asyncytium that allows ions and small molecules todiffuse across the brain parenchyma. Astrocytes havein common unique cytological and immunologicalproperties that make them easy to identify, includingtheir star shape, the glial end feet on capillaries, and a
Molecularlayer
Purkinjecelllayer
Granularlayer
Whitematter
FIGURE 1.8 The arrangement of astrocytes in human cerebellarcortex. Bergmann glial cells are in red, protoplasmic astrocytes arein green, and fibrous astrocytes are in blue.
10 1. CELLULAR COMPONENTS OF NERVOUS TISSUE

unique population of large bundles of intermediatefilaments. These filaments are composed of an astro-glial-specific protein commonly referred to as glialfibrillary acidic protein (GFAP). S-100, a calcium-bindingprotein, and glutamine synthetase are also astrocytemarkers. Ultrastructurally, gap junctions (connexins),desmosomes, glycogengranules, andmembraneortho-gonal arrays are distinct features used by morpholo-gists to identify astrocytic cellular processes in thecomplex cytoarchitecture of the nervous system.
For a long time, astrocytes were thought to physi-cally form the blood–brain barrier (considered laterin this chapter), which prevents the entry of cellsand diffusion of molecules into the CNS. In fact,astrocytes are indeed the blood–brain barrier in lowerspecies. However, in higher species, astrocytes areresponsible for inducing and maintaining the tightjunctions in endothelial cells that effectively formthe barrier. Astrocytes also take part in angiogenesis,which may be important in the development andrepair of the CNS. However, their role in this impor-tant process is still poorly understood.
Astrocytes Have a Wide Range of Functions
There is strong evidence for the role of radial gliaand astrocytes in the migration and guidance of neu-rons in early development. Astrocytes are a majorsource of extracellular matrix proteins and adhesion
molecules in the CNS; examples are nerve cell–nervecell adhesion molecule (N-CAM), laminin, fibronectin,cytotactin, and the J-1 family members janusin andtenascin. These molecules participate not only in themigration of neurons but also in the formation of neu-ronal aggregates, so-called nuclei, as well as networks.
Astrocytes produce, in vivo and in vitro, a verylarge number of growth factors. These factors act sin-gly or in combination to selectively regulate the mor-phology, proliferation, differentiation, survival, or allfour, of distinct neuronal subpopulations. Most ofthe growth factors also act in a specific manner onthe development and functions of astrocytes andoligodendrocytes. The production of growth factorsand cytokines by astrocytes and their responsivenessto these factors is a major mechanism underlying thedevelopmental function and regenerative capacity ofthe CNS. During neurotransmission, neurotransmit-ters and ions are released at high concentration inthe synaptic cleft. The rapid removal of these sub-stances is important so that they do not interfere withfuture synaptic activity. The presence of astrocyteprocesses around synapses positions them well toregulate neurotransmitter uptake and inactivation(Kettenman and Ransom, 1995). These possibilitiesare consistent with the presence in astrocytes of trans-port systems for many neurotransmitters. Forinstance, glutamate reuptake is performed mostly byastrocytes, which convert glutamate into glutamine
MZ
CP
IZ
VZ
Sub VZ
Migrating neuron
Radial processof glial cell
FIGURE 1.9 Radial glia perform support and guidance functions for migrating neurons. In early development, radial glia span the thick-ness of the expanding brain parenchyma. (Inset) Defined layers of the neural tube from the ventricular to the outer surface: VZ, ventricularzone; IZ, intermediate zone; CP, cortical plate; MZ, marginal zone. The radial process of the glial cell is indicated in blue, and a single attachedmigrating neuron is depicted at the right.
11NEUROGLIA

and then release it into the extracellular space. Gluta-mine is taken up by neurons, which use it to generateglutamate and g-aminobutyric acid, potent excit-atory and inhibitory neurotransmitters, respectively(Fig. 1.11). Astrocytes contain ion channels for Kþ,Naþ, Cl–, HCO3, and Ca2þ, as well as displaying awide range of neurotransmitter receptors. Kþ ionsreleased from neurons during neurotransmission aresoaked up by astrocytes and moved away from thearea through astrocyte gap junctions. This is knownas “spatial buffering.” Astrocytes play a major rolein detoxification of the CNS by sequestering metalsand a variety of neuroactive substances of endoge-nous and xenobiotic origin.
In response to stimuli, intracellular calcium wavesare generated in astrocytes. Propagation of the Ca2þ
wave can be visually observed as it moves across thecell soma and from astrocyte to astrocyte. The genera-tion of Ca2þ waves from cell to cell is thought to bemediated by second messengers, diffusing throughgap junctions. Because they develop postnatally inrodents, gap junctions may not play an important rolein development. In the adult brain, gap junctions arepresent in all astrocytes. Some gap junctions have alsobeen detected between astrocytes and neurons. Thus,they may participate, along with astroglial neurotrans-mitter receptors, in the coupling of astrocyte andneuron physiology.
In a variety of CNS disorders—neurotoxicity,viral infections, neurodegenerative disorders, HIV,AIDS, dementia, multiple sclerosis, inflammation,and trauma—astrocytes react by becoming hypertro-phic and, in a few cases, hyperplastic. A rapid andhuge upregulation of GFAP expression and filamentformation is associated with astrogliosis. The forma-tion of reactive astrocytes can spread very far fromthe site of origin. For instance, a localized trauma canrecruit astrocytes from as far as the contralateralside, suggesting the existence of soluble factors in the
FIGURE 1.11 The glutamate–glutamine cycle is an example of acomplex mechanism that involves an active coupling of neurotrans-mitter metabolism between neurons and astrocytes. The systems ofexchange of glutamine, glutamate, GABA, and ammonia betweenneurons and astrocytes are highly integrated. The postulated detox-ification of ammonia and the inactivation of glutamate and GABAby astrocytes are consistent with the exclusive localization of gluta-mine synthetase in the astroglial compartment.
Ependyma
Glia limitans
Pia mater
BV
Myelin
Astrocyte
FIGURE 1.10 Astrocytes (in orange) are depicted in situ in sche-matic relationship with other cell types with which they are knownto interact. Astrocytes send processes that surround neurons andsynapses, blood vessels, and the region of the node of Ranvierand extend to the ependyma, as well as to the pia mater, wherethey form the glial limitans.
12 1. CELLULAR COMPONENTS OF NERVOUS TISSUE

mediation process. Tumor necrosis factor (TNF) andciliary neurotrophic factors (CNTF) have been identi-fied as key factors in astrogliosis.
Microglia Are Mediators of Immune Responsesin Nervous Tissue
The brain has traditionally been considered an“immunologically privileged site,” mainly becausethe blood–brain barrier normally restricts the accessof immune cells from the blood. However, it is nowknown that immunological reactions do take placein the central nervous system, particularly duringcerebral inflammation. Microglial cells have beentermed the tissue macrophages of the CNS, and theyfunction as the resident representatives of the immunesystem in the brain. A rapidly expanding literaturedescribes microglia as major players in CNS develop-ment and in the pathogenesis of CNS disease.
The first description of microglial cells can betraced to Franz Nissl (1899), who used the term rodcell to describe a population of glial cells that reactedto brain pathology. He postulated that rod-cell func-tion was similar to that of leukocytes in other organs.Cajal described microglia as part of his “third ele-ment” of the CNS—cells that he considered to be ofmesodermal origin and distinct from neurons andastrocytes (Ramon y Cajal, 1913).
Del Rio-Hortega (1932) distinguished this third ele-ment into microglia and oligodendrocytes. He usedsilver impregnation methods to visualize the ramifiedappearance of microglia in the adult brain, and heconcluded that ramified microglia could transforminto cells that were migratory, ameboid, and phago-cytic. Indeed, a hallmark of microglial cells is theirability to become reactive and to respond to patholog-ical challenges in a variety of ways. A fundamentalquestion raised by del Rio-Hortega’s studies was theorigin of microglial cells. Some questions about thisremain even today.
Microglia Have Diverse Functions inDeveloping and Mature Nervous Tissue
On the basis of current knowledge, it appears thatmost ramified microglial cells are derived from bonemarrow–derived monocytes, which enter the brainparenchyma during early stages of brain develop-ment. These cells help phagocytose degenerating cellsthat undergo programmed cell death as part of nor-mal development. They retain the ability to divideand have the immunophenotypic properties of mono-cytes and macrophages. In addition to their role inremodeling the CNS during early development,
microglia secrete cytokines and growth factors thatare important in fiber tract development, gliogenesis,and angiogenesis. They are also the major CNS cellsinvolved in presenting antigens to T lymphocytes.After the early stages of development, ameboidmicroglia transform into the ramified microglia thatpersist throughout adulthood (Altman, 1994).
Little is known about microglial function in thehealthy adult vertebrate CNS. Microglia constitute aformidable percentage (5–20%) of the total cells inthe mouse brain. Microglia are found in all regionsof the brain, and there are more in gray than in whitematter. The neocortex and hippocampus have moremicroglia than regions like the brainstem or cerebel-lum. Species variations have also been noted, ashuman white matter has three times more microgliathan rodent white matter.
Microglia usually have small rod-shaped somasfrom which numerous processes extend in a rathersymmetrical fashion. Processes from different micro-glia rarely overlap or touch, and specialized contactsbetween microglia and other cells have not beendescribed in the normal brain. Although each micro-glial cell occupies its own territory, microglia collec-tively form a network that covers much of the CNSparenchyma. Because of the numerous processes,microglia present extensive surface membrane to theCNS environment. Regional variation in the numberand shape of microglia in the adult brain suggeststhat local environmental cues can affect microglial dis-tribution and morphology. On the basis of thesemorphological observations, it is likely that microgliaplay a role in tissue homeostasis. The nature ofthis homeostasis remains to be elucidated. It is clear,however, that microglia can respond quickly and dra-matically to alterations in the CNS microenvironment.
Microglia Become Activated in PathologicalStates
“Reactive”microglia can be distinguished from rest-ingmicroglia by two criteria: (1) change inmorphologyand (2) upregulation of monocyte–macrophage mole-cules (Fig. 1.12). Although the two phenomena gener-ally occur together, reactive responses of microgliacan be diverse and restricted to subpopulations ofcells within a microenvironment. Microglia not onlyrespond to pathological conditions involving immuneactivation but also become activated in neurodegen-erative conditions that are not considered immunemediated. This latter response is indicative of thephagocytic role of microglia. Microglia change theirmorphology and antigen expression in response toalmost any form of CNS injury.
13NEUROGLIA

CEREBRAL VASCULATURE
Blood vessels form an extremely rich network inthe central nervous system, particularly in the cere-bral cortex and subcortical gray masses, whereas thewhite matter is less densely vascularized (Fig. 1.13)(Duvernoy et al., 1981). There are distinct regionalpatterns of microvessel distribution in the brain.These patterns are particularly clear in certain subcor-tical structures that constitute discrete vascular terri-tories and in the cerebral cortex, where regional andlaminar patterns are striking. For example, layer IVof the primary visual cortex possesses an extremelyrich capillary network in comparison with otherlayers and adjacent regions (Fig. 1.13). Interestingly,most of the inputs from the visual thalamus terminatein this particular layer. Capillary densities are higherin regions containing large numbers of neurons andwhere synaptic density is high. Progressive occlusionof a large arterial trunk, as seen in stroke, induces anischemic injury that may eventually lead to necrosisof the brain tissue. The size of the resulting infarc-tion is determined in part by the worsening of theblood circulation through the cerebral microvessels.Occlusion of a large arterial trunk results in rapid
swelling of the capillary endothelium and surround-ing astrocytes, which may reduce the capillary lumento about one-third of its normal diameter, preventingred blood cell circulation and oxygen delivery to thetissue. The severity of these changes subsequentlydetermines the time course of neuronal necrosis,as well as the possible recovery of the surroundingtissue and the neurological outcome of the patient.In addition, the presence of multiple microinfarctscaused by occlusive lesions of small cerebral arte-rioles may lead to a progressively dementing illness,referred to as vascular dementia, affecting elderlyhumans.
The Blood–Brain Barrier Maintainsthe Intracerebral Milieu
Capillaries of the central nervous system form aprotective barrier that restricts the exchange of solutesbetween blood and brain. This distinct function ofbrain capillaries is called the blood–brain barrier(Fig. 1.14) (Bradbury, 1979). Capillaries of the retinahave similar properties and are termed the blood–retina barrier. It is thought that the blood–brain andblood–retina barriers function to maintain a constant
A B C
FIGURE 1.12 Activation of microglial cells in a tissue section from human brain. Resting microglia in normal brain (A). Activated micro-glia in diseased cerebral cortex (B) have thicker processes and larger cell bodies. In regions of frank pathology (C), microglia transform intophagocytic macrophages, which can also develop from circulating monocytes that enter the brain. Arrow in B indicates rod cell. Sectionsstained with antibody to ferritin. Scale bar ¼ 40 mm.
14 1. CELLULAR COMPONENTS OF NERVOUS TISSUE

intracerebral milieu, so that neuronal signaling canoccur without interference from substances leakingin from the bloodstream. This function is importantbecause of the nature of intercellular communicationin the CNS, which includes chemical signals acrossintercellular spaces. Without a blood–brain barrier,circulating factors in the blood, such as certainhormones, which can also act as neurotransmitters,would interfere with synaptic communication. Whenthe blood–brain barrier is disrupted, edema fluidaccumulates in the brain. Increased permeability ofthe blood–brain barrier plays a central role in manyneuropathological conditions, including multiplesclerosis, AIDS, and childhood lead poisoning, and
may also play a role in Alzheimer disease. The cerebralcapillary wall is composed of an endothelial cellsurrounded by a very thin (about 30 nm) basementmembrane or basal lamina. End feet of perivascularastrocytes are apposed against this continuous basallamina. Around the capillary lies a virtual perivascularspace occupied by another cell type, the pericyte, whichsurrounds the capillarywalls. The endothelial cell formsa thin monolayer around the capillary lumen, and asingle endothelial cell can completely surround thelumen of the capillary (Fig. 1.14). A fundamental differ-ence between brain endothelial cells and those of thesystemic circulation is the presence in the brain of inter-endothelial tight junctions, also known as zonula occlu-dens. In the systemic circulation, the interendothelialspace serves as a diffusion pathway that offers littleresistance to most blood solutes entering the surround-ing tissues. In contrast, blood–brain barrier tight junc-tions effectively restrict the intercellular route of solutetransfer. The blood–brain barrier interendothelial junc-tions are not static seals; rather they are a series of activegates that can allow certain small molecules to pene-trate. One such molecule is the lithium ion, used in thecontrol of manic depression.
Another characteristic of endothelial cells of the brainis their low transcytotic activity. Brain endothelium,therefore, is by this index not very permeable. It is ofinterest that certain regions of the brain, such as the areapostremaandperiventricular organs, lack a blood–brainbarrier. In these regions, the perivascular space is indirect contact with the nervous tissue, and endothelialcells are fenestrated and show many pinocytotic vesi-cles. In these brain regions, neurons are known tosecrete hormones and other factors that require rapidand uninhibited access to the systemic circulation.
Because of the high metabolic requirements of thebrain, blood–brain barrier endothelial cells must havetransport mechanisms for the specific nutrientsneeded for proper brain function. One such mecha-nism is the glucose transporter isoform 1 (GLUT1),which is expressed asymmetrically on the surface ofblood–brain barrier endothelial cells. In Alzheimerdisease, the expression of GLUT1 on brain endothelialcells is reduced. This reduction may be due to a lowermetabolic requirement of the brain after extensiveneuronal loss. Other specific transport mechanismson the cerebral endothelium include the large neutralamino acid carrier-mediated system that transports,among other amino acids, L-3,4-dihydroxyphenylala-nine (L-dopa), used as a therapeutic agent in Parkin-son disease. Also on the surface of blood–brainbarrier endothelial cells are transferrin receptors thatallow the transport of iron into specific areas of thebrain. The amount of iron that is transported into the
A
B
FIGURE 1.13 Microvasculature of the human neocortex. (A)The primary visual cortex (area 17). Note the presence of segmentsof deep penetrating arteries that have a larger diameter than themicrovessels and run from the pial surface to the deep corticallayers, as well as the high density of microvessels in the middlelayer (layers IVCa and IVCb). (B) The prefrontal cortex (area 9).Cortical layers are indicated by Roman numerals. Microvesselsare stained using an antibody against heparan sulfate proteoglycancore protein, a component of the extracellular matrix.
15CEREBRAL VASCULATURE

various areas of the brain appears to depend on theconcentration of transferrin receptors on the surfaceof endothelial cells of that region. Thus, the transportof specific nutrients into the brain is regulated duringphysiological and pathological conditions by blood–brain barrier transport proteins distributed accordingto the regional and metabolic requirements of braintissue.
In general, disruption of the blood–brain barriercauses perivascular or vasogenic edema, which isthe accumulation of fluids from the blood aroundthe blood vessels of the brain. This is one of the mainfeatures of multiple sclerosis. In multiple sclerosis,inflammatory cells, primarily T cells andmacrophages,invade the brain bymigrating through the blood–brainbarrier and attack cerebral elements as if these
FIGURE 1.14 Human cerebral capillary obtained at biopsy. Blood–brain barrier (BBB) capillaries are characterized by the paucity of trans-cytotic vesicles in endothelial cells (E), a high mitochondrial content (large arrow), and the formation of tight junctions (small arrows) betweenendothelial cells that restrict the transport of solutes through the interendothelial space. The capillary endothelium is encased within a base-ment membrane (arrowheads), which also houses pericytes (P). Outside the basement membrane are astrocyte foot processes (asterisk), whichmay be responsible for the induction of BBB characteristics on the endothelial cells. L, lumen of the capillary. Scale bar ¼ 1 mm. From Claudioet al. (1995).
16 1. CELLULAR COMPONENTS OF NERVOUS TISSUE

elements were foreign antigens. It has been observedby many investigators that the degree of edema accu-mulation causes the neurological symptoms experi-enced by people suffering from multiple sclerosis.
Studying the regulation of blood–brain barrier per-meability is important for several reasons. Therapeu-tic treatments for neurological disease need to beable to cross the barrier. Attempts to design drugdelivery systems that take therapeutic drugs directlyinto the brain have been made by using chemicallyengineered carrier molecules that take advantage ofreceptors such as that for transferrin, which normallytransports iron into the brain. Development of anin vitro test system of the blood–brain barrier is ofimportance in the creation of new neurotropic drugsthat are targeted to the brain.
References
Altman, J. (1994). Microglia emerge from the fog. Trends Neurosci.17, 47–49.
Bradbury, M. W. B. (1979). “The Concept of a Blood-Brain Barrier,”
pp. 381–407. Wiley, Chichester, UK.Brodal, A. (1981). “Neurological Anatomy in Relation to Clinical
Medicine,” 3rd ed. Oxford University Press, New York.Bunge, R. P. (1968). Glial cells and the central myelin sheath.
Physiol. Rev. 48, 197–251.Claudio, L., Raine, C. S., and Brosnan, C. F. (1995). Evidence of per-
sistent blood-brain barrier abnormalities in chronic-progressivemultiple sclerosis. Acta Neuropathol. 90, 228–238.
DeFelipe, J., Hendry, S. H. C., and Jones, E. G. (1989). Visualizationof chandelier cell axons by parvalbumin immunoreactivity inmonkey cerebral cortex. Proc. Natl. Acad. Sci. USA 86, 2093–2097.
del Rio-Hortega, P. (1932). Microglia. In “Cytology and CellularPathology of the Nervous System” (W. Penfield, ed.), Vol. 2,pp. 481–534. Harper (Hoeber), New York.
Duvernoy, H. M., Delon, S., and Vannson, J. L. (1981). Corticalblood vessels of the human brain. Brain Res. Bull. 7, 519–579.
Freund, T. F., Martin, K. A. C., Smith, A. D., and Somogyi, P. (1983).Glutamate decarboxylase-immunoreactive terminals of Golgi-impregnated axoaxonic cells and of presumed basket cells insynaptic contact with pyramidal neurons of the cat’s visualcortex. J. Comp. Neurol. 221, 263–278.
Griffiths, I., Klugmann, M., Anderson, T., Yool, D., Thomson, C.,Schwab, M. H., Schneider, A., Zimmermann, F., McCulloch,M., Nadon, N., and Nave, K. A. (1998). Axonal swellings anddegeneration in mice lacking the major proteolipid of myelin.Science. 280, 1610–1613.
Jones, E. G. (1984). Laminar distribution of cortical efferent cells. In“Cellular Components of the Cerebral Cortex” (A. Peters and E.G. Jones, eds.), Vol. 1, pp. 521–553. Plenum, New York.
Jones, E. G. (1975). Varieties and distribution of non-pyramidal cellsin the somatic sensory cortex of the squirrel monkey. J. Comp.
Neurol. 160, 205–267.
Kettenman, H., and Ransom, B. R., eds. (1995). “Neuroglia.” OxfordUniversity Press, Oxford.
Mountcastle, V. B. (1978). An organizing principle for cerebral func-tion: The unit module and the distributed system. In “The Mind-ful Brain: Cortical Organization and the Group-Selective Theoryof Higher Brain Function” (V. B. Mountcastle and G. Eddman,eds.), pp. 7–50. MIT Press, Cambridge, MA.
Nissl, F. (1899). Uber einige Beziehungen zwischen Nervenzelle-nerkrankungen und gliosen Erscheinungen bei verschiedenenPsychosen. Arch. Psychol. 32, 1–21.
Peters, A., and Jones, E. G., eds. (1984). “Cellular Components ofthe Cerebral Cortex,” Vol. 1. Plenum, New York.
Peters, A., Palay, S. L., and Webster, H. de F. (1991). “The FineStructure of the Nervous System: Neurons and Their Support-ing Cells,” 3rd ed. Oxford University Press, New York.
Ramon y Cajal, S. (1913). Contribucion al conocimiento de la neuro-glia del cerebro humano. Trab. Lab. Invest. Biol. 11, 255–315.
Ritchie, J. M. (1984). Physiological basis of conduction in mye-linated nerve fibers. In “Myelin” (P. Morell, ed.), pp. 117–146.Plenum, New York.
Somogyi, P., and Cowey, A. (1981). Combined Golgi and electronmicroscopic study on the synapses formed by double bouquetcells in the visual cortex of the cat and monkey. J. Comp. Neurol.
195, 547–566.Somogyi, P., Kisvarday, Z. F., Martin, K. A. C., and Whitteridge, D.
(1983). Synaptic connections of morphologically identified andphysiologically characterized basket cells in the striate cortexof cat. Neuroscience 10, 261–294.
Yin, X., Baek, R. C., Kirschner, D. A., Peterson, A., Fujii, Y., Nave,K. A., Macklin, W. B., and Trapp, B. D. (2006). Evolution of aneuroprotective function of central nervous system myelin.J. Cell Biol. 172, 469–478.
Suggested Readings
Brightman, M. W., and Reese, T. S. (1969). Junctions between inti-mately apposed cell membranes in the vertebrate brain. J. CellBiol. 40, 648–677.
Broadwell, R. D., and Salcman, M. (1981). Expanding the defini-tion of the BBB to protein. Proc. Natl. Acad. Sci. USA 78,7820–7824.
Fernandez-Moran, H. (1950). EM observations on the structure ofthe myelinated nerve sheath. Exp. Cell Res. 1, 143–162.
Gehrmann, J., Matsumoto, Y., and Kreutzberg, G. W. (1995).Microglia: Intrinsic immuneffector cell of the brain. Brain Res.
Rev. 20, 269–287.Kimbelberg, H., and Norenberg, M. D. (1989). Astrocytes. Sci. Am.
26, 66–76.Kirschner, D. A., Ganser, A. L., and Caspar, D. W. (1984). Dif-
fraction studies of molecular organization and membrane inter-actions in myelin. In “Myelin” (P. Morell, ed.), pp. 51–96.Plenum, New York.
Lum, H., and Malik, A. B. (1994). Regulation of vascular endothelialbarrier function. Am. J. Physiol. 267, L223–L241.
Rosenbluth, J. (1980). Central myelin in the mouse mutant shiverer.J. Comp. Neurol. 194, 639–728.
Rosenbluth, J. (1980). Peripheral myelin in the mouse mutant shiv-erer. J. Comp. Neurol. 194, 729–753.
17SUGGESTED READINGS


















