
CD26: A Negative Selection Marker for Human Treg Cells · Department of Biochemistry and Molecular...
13
CD26: A Negative Selection Marker for Human Treg Cells Francisco J. Salgado, Amparo Pe ´rez-Dı´az, Nora M. Villanueva, Olaya Lamas, Pilar Arias, Montserrat Nogueira * Abstract A major obstacle hampering the therapeutic application of regulatory T (Treg) cells is the lack of suitable extracellular markers, which complicates their identification/isola- tion. Treg cells are normally isolated via CD25 (IL-2Ra) targeting, but this protein is also expressed by activated CD4 1 effector T (Teff) lymphocytes. Other extracellular (positive or negative) Treg selection markers (e.g., HLA-DR, CD127) are also nonspeci- fic. CD26 is an extracellular peptidase whose high expression has been traditionally used as an indicator of immune activation and effector functions in T cells. Now, we provide flow cytometry data showing high levels of CD26 within CD4 1 CD25 2 or CD4 1 FoxP3 2/low effector T (Teff) lymphocytes, but negative or low levels (CD26 2/low ) in Treg cells selected according to the CD4 1 CD25 high or the CD4 1 FoxP3 high phenotype. Unlike the negative marker CD127 (IL-7Ra), which is down modulated in CD4 1 Teff lymphocytes after TCR triggering, most of these cells upregulate CD26 and take a CD4 1 CD25 1/high CD26 1 phenotype upon activation. In contrast, there is only a slight upregulation within Treg cells (CD4 1 CD25 high CD26 2/low ). Thus, differences in CD26 levels between Treg and Teff subsets are stable, and assessment of this marker, in combination with others like CD25, FoxP3, or CD127, may be useful during the quantitative evaluation or the isolation of Treg cells in samples containing activated Teff lymphocytes (e.g., from patients with autoimmune/inflammatory diseases). ' 2012 International Society for Advancement of Cytometry Key terms CD26; regulatory T cells; effector T cells; flow cytometry; magnetic sorting REGULATORY cells are included in different leukocyte populations, like CD8 1 (1) or CD4 1 (2,3) T subsets. CD4 1 CD25 high natural Treg (nTreg) cells constitute a small CD4 1 subpopulation ( \5% of CD4 1 T lymphocytes) with a thymic origin. Numeri- cal or functional deficit of nTreg cells is linked to autoimmune diseases such as multi- ple sclerosis (MS), Type 1 diabetes or rheumatoid arthritis (RA) (4). In addition, they are also important to prevent (or delay) grafts rejection or disproportionate responses to bacterial/viral antigens (5,6). Alternative subpopulations of regulatory T cells (e.g., T H 3 and T r 1), called ‘‘adaptive’’ or ‘‘induced’’ regulatory CD4 1 T cells (iTreg), are generated in the periphery (7). Both nTreg and iTreg lymphocytes (altogether, Tregs) limit the biological activities (e.g., proliferation, cytokine production) of adaptive ‘‘effector’’ cells, a miscellaneous group consisting of CD4 1 helper T cells (T H 1, T H 2, T H 17, T FH ,T H 9, T H 22), CD8 1 cytotoxic T lymphocytes and B cells. To finely tune the strength of effector responses, Treg cells employ various suppressor mechanisms, like inhibitory soluble molecules (adenosine, TGFb , IL-10) or cell contact-mediated path- ways (e.g., membrane cytokines like TGFb1 or surface molecules like CTLA-4) (8,9). Any research aimed at controlling Treg function, either enhancing (e.g., in auto- immune diseases) or blocking it (e.g., in cancer), will hold a great interest (5,6,10,11). However, this kind of research faces several challenges; for example, how to distinguish regulatory (Treg) from effector (Teff) CD4 1 T populations. Human Department of Biochemistry and Molecular Biology, Faculty of Biology/ CIBUS, University of Santiago de Compostela, Coru~ na, Espa~ na Received 19 September 2011; Revision Received 18 May 2012; Accepted 24 July 2012 Grant sponsor: Xunta de Galicia; Grant number: INCITE08PXIB200062PR (to A.P.-D.). Additional Supporting Information may be found in the online version of this article. Francisco J. Salgado, Nora Mart ınez- Villanueva, Amparo Pe ´rez-D ıaz, and Olaya Lamas did the experimental work and data analysis. Francisco J. Salgado, Pilar Arias, and Montserrat Nogueira designed the research, supervised the work, and wrote the article. *Correspondence to: Montserrat Nogueira. Laboratorio BQ1 (2 o piso) CIBUS. Departamento de Bioqu ımica y Biolog ıa Molecular, Universidad de Santiago de Compostela. C/Lope Go ´mez de Marzoa s/n (Campus vida), Santiago de Compostela, CP. 15782, Coru~ na, Espa~ na Email: [email protected] Published online 4 September 2012 in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/cyto.a.22117 © 2012 International Society for Advancement of Cytometry Original Article Cytometry Part A 81A: 843855, 2012
Transcript of CD26: A Negative Selection Marker for Human Treg Cells · Department of Biochemistry and Molecular...

CD26: A Negative Selection Marker for
Human Treg Cells
Francisco J. Salgado, Amparo Perez-Dıaz, Nora M. Villanueva, Olaya Lamas,Pilar Arias, Montserrat Nogueira*
� AbstractA major obstacle hampering the therapeutic application of regulatory T (Treg) cells isthe lack of suitable extracellular markers, which complicates their identification/isola-tion. Treg cells are normally isolated via CD25 (IL-2Ra) targeting, but this protein isalso expressed by activated CD41 effector T (Teff) lymphocytes. Other extracellular(positive or negative) Treg selection markers (e.g., HLA-DR, CD127) are also nonspeci-fic. CD26 is an extracellular peptidase whose high expression has been traditionallyused as an indicator of immune activation and effector functions in T cells. Now, weprovide flow cytometry data showing high levels of CD26 within CD41CD252 orCD41FoxP32/low effector T (Teff) lymphocytes, but negative or low levels (CD262/low)in Treg cells selected according to the CD41CD25high or the CD41FoxP3high phenotype.Unlike the negative marker CD127 (IL-7Ra), which is down modulated in CD41 Tefflymphocytes after TCR triggering, most of these cells upregulate CD26 and take aCD41CD251/highCD261 phenotype upon activation. In contrast, there is only aslight upregulation within Treg cells (CD41CD25highCD262/low). Thus, differencesin CD26 levels between Treg and Teff subsets are stable, and assessment of thismarker, in combination with others like CD25, FoxP3, or CD127, may be usefulduring the quantitative evaluation or the isolation of Treg cells in samples containingactivated Teff lymphocytes (e.g., from patients with autoimmune/inflammatorydiseases). ' 2012 International Society for Advancement of Cytometry
� Key termsCD26; regulatory T cells; effector T cells; flow cytometry; magnetic sorting
REGULATORY cells are included in different leukocyte populations, like CD81 (1) or
CD41 (2,3) T subsets. CD41CD25high natural Treg (nTreg) cells constitute a small
CD41 subpopulation (\5% of CD41 T lymphocytes) with a thymic origin. Numeri-
cal or functional deficit of nTreg cells is linked to autoimmune diseases such as multi-
ple sclerosis (MS), Type 1 diabetes or rheumatoid arthritis (RA) (4). In addition, they
are also important to prevent (or delay) grafts rejection or disproportionate responses
to bacterial/viral antigens (5,6). Alternative subpopulations of regulatory T cells (e.g.,
TH3 and Tr1), called ‘‘adaptive’’ or ‘‘induced’’ regulatory CD41 T cells (iTreg), are
generated in the periphery (7). Both nTreg and iTreg lymphocytes (altogether, Tregs)
limit the biological activities (e.g., proliferation, cytokine production) of adaptive
‘‘effector’’ cells, a miscellaneous group consisting of CD41 helper T cells (TH1, TH2,
TH17, TFH, TH9, TH22), CD81 cytotoxic T lymphocytes and B cells. To finely tune the
strength of effector responses, Treg cells employ various suppressor mechanisms, like
inhibitory soluble molecules (adenosine, TGFb, IL-10) or cell contact-mediated path-
ways (e.g., membrane cytokines like TGFb1 or surface molecules like CTLA-4) (8,9).
Any research aimed at controlling Treg function, either enhancing (e.g., in auto-
immune diseases) or blocking it (e.g., in cancer), will hold a great interest
(5,6,10,11). However, this kind of research faces several challenges; for example, how
to distinguish regulatory (Treg) from effector (Teff) CD41 T populations. Human
Department of Biochemistry andMolecular Biology, Faculty of Biology/CIBUS, University of Santiago deCompostela, Coru~na, Espa~na
Received 19 September 2011; RevisionReceived 18 May 2012; Accepted 24 July2012
Grant sponsor: Xunta de Galicia; Grantnumber: INCITE08PXIB200062PR (to A.P.-D.).
Additional Supporting Information may befound in the online version of this article.
Francisco J. Salgado, Nora Mart�ınez-Villanueva, Amparo Perez-D�ıaz, andOlaya Lamas did the experimental workand data analysis. Francisco J. Salgado,Pilar Arias, and Montserrat Nogueiradesigned the research, supervised thework, and wrote the article.
*Correspondence to: MontserratNogueira. Laboratorio BQ1 (2o piso)CIBUS. Departamento de Bioqu�ımica yBiolog�ıa Molecular, Universidad deSantiago de Compostela. C/Lope Gomezde Marzoa s/n (Campus vida), Santiagode Compostela, CP. 15782, Coru~na,Espa~na
Email: [email protected]
Published online 4 September 2012 inWiley Online Library(wileyonlinelibrary.com)
DOI: 10.1002/cyto.a.22117
© 2012 International Society forAdvancement of Cytometry
Original Article
Cytometry Part A � 81A: 843�855, 2012

Treg cells constitutively express surface proteins like CD25,
CD45RO, CTLA-4, HLA-DR, or GITR (see Table 1 in Sup-
porting Information) (2,12), but these markers are neither
present in 100% of Tregs or exclusive of this cell lineage. Three
examples can illustrate this point: CD25 (IL-2Ra), FoxP3, and
CD127 (IL-7Ra).
The majority of human Treg cells strongly and constitu-
tively express CD25 (CD25high). However, conventional/effec-
tor T cells (2,12,13) and a portion of CCR71 central memory
T lymphocytes start expressing CD25 upon TCR-mediated
activation (14). Therefore, even highly pure CD41CD25high
Treg populations may contain a significant fraction of proin-
flammatory Teff cells (15). On the other hand, FoxP3 is the
most specific Treg marker (2). Despite this fact, FoxP3 still
shows a transitory and activation-dependent expression in
CD41CD252 Teff cells, which together with its intracellular
nature disqualifies this marker for Treg identification and,
especially, isolation purposes (16). CD127 is another protein
whose levels inversely correlate with FoxP3 expression in Treg
lymphocytes (17,18). Nevertheless, the mere existence of an
underlying disease (e.g., HIV infection) (19) or the in vitro
activation (20) cause an intense CD127 down modulation on
formerly CD1271 Teff cells. Other alternative markers have
arisen more recently. Markus Kleinewietfeld et al. (21) have
reported CD49d (a chain of VLA-4 integrin) as expressed in
most of IFN-c or IL-17-producing proinflammatory T cells
but reduced on Tregs, even though data from the same group
(22) as well as other researchers (23) still reflect some degree of
CD49d expression in some subsets of Treg cells. Mandapathil
et al. (24) have focused on CD39 as a positive selection marker.
This ectoenzyme catalyzes the generation of AMP from ATP,
which is necessary to produce adenosine, an important media-
tor of active suppression (22). Fifty to 90% of CD41CD391 T
lymphocytes are FoxP31 and express low levels of CD127 (24).
However, activated T cells upregulate CD39 (25), and a novel
population of human CD41CD391FoxP32 T cells that produce
IFN-c and IL-17 has been found (26). Thus, it seems that more
phenotypic studies on Treg cells are still necessary (27).
CD26 is a serine protease with dipeptidyl peptidase IV
(DPPIV) activity (28). Activated/memory T cells display a
CD26high phenotype, and TH1 cytokines like IL-12 raise the
number of CD26 molecules on T lymphocytes (28,29). Thus,
high surface levels of this protease are an indication of, at least,
TH1 effector responses (28,29). Ligation of CD26 leads to acti-
vation of signal transduction proteins (e.g., ERK, p56lck,
CD3f, ZAP-70, CARMA-1/NF-jB), cell proliferation, and
cytokine (IL-2, IFN-c) production (28,30). Both cis-interac-
tion of CD26 with CD45 (31,32) and trans-association of
CD26 (T cell) with caveolin-1 (APCs) (33) seem important
for CD26 functions. However, despite the large amount of
data supporting the costimulatory role of CD26 in T cells, an
extensive research on CD26 levels in Treg cells has not been
undertaken. The present work shows that CD26 is present on
FoxP3-expressing activated CD41 Teff cells, but reduced or
absent from Tregs. This CD262/low phenotype is stable, and
therefore useful to differentiate these two antagonistic CD41
T cell subsets.
MATERIALS AND METHODS
Materials
Phytohemagglutinin (PHA; catalog no. L1668), parafor-
maldehyde (PFA; catalog no. P-6148), penicillin-streptomycin
solution (catalog no. P0781), 5(6)-Carboxyfluorescein diace-
tate N-succinimidyl ester (CFSE; catalog no. 21888-25MG-F),
and RPMI-1640 culture medium (catalog no. R4130) were
obtained from Sigma-Aldrich (Spain), fetal bovine serum
(FBS; catalog no. DE14-801F) from Biowhittaker (Lonza Iber-
ica, Spain), and Ficoll-Paque PLUS (catalog no. 17-1440-03)
from GE Healthcare (Spain). Mouse antibodies against human
molecules CD127 (CD127-PE, catalog no. 557938, and CD127
Alexa Fluor 647, catalog no. 558598; both IgG1 clone hIL-7R-
M21), CD25 (CD25-FITC, catalog no. 555431, and CD25-
APC, catalog no. 555434; both IgG1, clone M-A251), CD39
(CD39-FITC, clone TU66, catalog no. 561444), CD49d
(CD49d-APC, IgG1, clone 9F10, catalog no. 559881), FoxP3
(anti-FoxP3-PE, IgG1, clone 259D/C7, catalog no. 560046),
and IFN-c (anti-IFN-c-PE, IgG1, clone B27, catalog no.
554701), as well as the mouse isotypic control IgG1-APC
(clone MOPC-21, catalog no. 555751) and the Alexa Fluor
647 mouse IgG1 isotype control (clone MOPC-21, catalog no.
557714), were from BD Pharmingen (BD Biosciences, Spain).
Mouse antibodies against human CD4 (CD4-PerCP, IgG1,
clone SK3, catalog no. 345770) and CD26 (CD26-FITC,
IgG2a, clone L272, catalog no. 340426) were purchased from
BD Immunocytometry Systems (BD Biosciences). Mouse
mAb against human CD26 (TP1/16) was purified from a
hybridome supernatant and used pure (in combination or not
with APC goat anti-mouse Ig; catalog no. 550826, BD Phar-
mingen) or FITC-labeled (Fluorotag FITC Conjugation Kit,
catalog no. FITC1-1KT; Sigma). Murine mAb against human
CD26 clone TP1/19 (CD26-FITC or APC, IgG2b, catalog no.
26F-100T or 26A-100T) was provided by Immunostep (Sala-
manca, Spain). Isotypic controls IgG1-FITC (clone MOPC-21;
catalog no. F6397) and IgG1-PE (clone MOPC-21; catalog no.
P4685) were purchased to Sigma. Buffers used for intracellular
staining of FoxP3 and IFN-c were BD Pharmingen stain buffer
(catalog no. 554656), BD FACSTM lysing buffer (catalog no.
349202), Human FoxP3 buffer set (catalog no. 560098), BD
Cytofix/Cytoperm bufferTM Plus (catalog no. 555028), and BD
Perm/WashTM (catalog no. 554723), all from BD Biosciences.
Methods
PBMCs purification and culture conditions. Human buffy
coats from healthy donors were kindly provided by Centro de
Transfusiones de Galicia (Santiago de Compostela, Spain).
Once informed consent for the donation was obtained, blood
samples (healthy subjects) were drawn into EDTA/K2E or LH/
170 IU tubes (BD vacutainer; BD Biosciences; catalog no.
367525 and 367526, respectively) via sterile venipuncture
(Medical Service at the University of Santiago) according to
the ethics committee guidelines. Peripheral blood mononu-
clear cells (PBMCs) were isolated from either buffy coats or
anticoagulant treated blood samples using Ficoll1 density gra-
dient centrifugation, as previously described (29,32). PBMCs
ORIGINAL ARTICLE
844 CD26 as a Negative Treg Selection Marker

were either used directly or in vitro cultured (378C, 5% CO2
humidified atmosphere). RPMI-1640 growth medium was
supplemented with 10% FBS, 100 lg/ml streptomycin and 100
UI/ml penicillin (complete medium). Stimulation of PBMCs
was performed by using 1 to 2 lg/ml PHA (phytohemaggluti-
nin from Phaseolus vulgaris) for 5 days in either 6/24-well
plates or 75/150 cm2 flasks. Alternatively, PBMCs were acti-
vated with beads coated with monoclonal antibodies against
CD2/CD3/CD28 (Treg suppression inspector human, Miltenyi
Biotec, Auburn, CA; 0.5–1 bead per lymphocyte; catalog no.
130-092-909). In IFN-c assays, culture medium was supple-
mented with GolgiPlugTM (BD Biosciences; 1/1,000 dilution)
during the last 4 h of incubation before immunostaining.
Human Treg purification. Viable (>90%; trypan blue exclu-
sion assay) Treg cells were isolated from either resting or acti-
vated PBMCs by two different magnetic methods. The first
one (Dynabeads1 Regulatory CD41CD251 T cell Kit; Life-
Technologies, Spain; catalog no. 113.63D) was a ‘‘classical’’
procedure. During the first step, CD41 T cells were enriched
by negative selection by means of a cocktail of monoclonal
antibodies (Antibody Mix Human CD4; mouse IgG antibodies
against CD14, CD16a, CD16b, CD56, CDw123, CD36, CD8,
CD19 and glycophorin A) and anti-mouse IgG Abs linked to
super paramagnetic polystyrene beads (Depletion MyOneTM
Dynabeads). Afterwards, both CD41CD252 Teff and CD25high
Treg cells were purified from CD41 T cells by means of beads
coated with antibodies against CD25 (Dynabeads CD25).
Finally, magnetic beads were removed from Treg cells (DETA-
CHaBEAD1 buffer). Cell recovery was calculated as follows:
(number of cells within the Treg enriched fraction 3 Treg pu-
rity)/(starting cell number 3 starting Treg percentage).
The second approach was the Human CD41
CD127lowCD49d2 Regulatory T Cell Enrichment Kit (Stem-
Cell Technologies, Grenoble, France; catalog no. 19232). This
fully negative selection protocol uses CD127 and CD49d to
isolate the Treg population from a PBMCs suspension. We
have followed strictly the manufacturer’s manual (EasySepTM
protocol) in combination with the EasySep magnet (for stand-
ard 5 ml polystyrene tubes; catalog no. 18000). Despite this
fact, a low Treg viability was always observed.
Flow cytometry assays. All the incubations during immuno-
fluorescence protocols were performed in the dark, and
unstained cells, isotype controls, and single fluorochrome
stained cells were used to set-up the flow cytometer. In all
cases, recommended concentrations were employed for all
monoclonal antibodies. In both proliferation and immunoflu-
orescence experiments, sample acquisition (typically 20,000 to
200,000 events) was performed on a FACScalibur Flow Cyto-
metry System (BD Biosciences), a 2-laser, 4-color flow cytom-
eter used for clinical samples at the USC University Hospital
Complex (CHUS). This instrument has the following laser-
fluorochrome combinations: (1) 488 nm blue laser—fluores-
cein isothiocyanate (FITC), 5(6)-carboxyfluorescein diacetate
N-succinimidyl ester (CFSE), phycoerythrin (PE), and peridi-
nin-chlorophyll-protein complex (PerCP); (2) 635-nm red-
diode laser—allophycocyanin (APC) and AlexaFluor 647. We
used WinMDI software (Dr. Joe Trotter, The Scripps Institute,
Flow Cytometry Core Facility) for data analysis.
For CFSE-based proliferation studies, PBMCs (RPMI me-
dium, 10 3 106 cells/ml) were firstly incubated with 5 mM
CFSE for 8 min (RT) in the dark. To block the reaction, an
equal volume of FBS was added and cells were thoroughly
washed before counting. Initial cell density was 50,000 cells/
well (0.25 3 106 PBMC/ml; 96-round bottom well micro-
plates), and 2 lg/ml PHA was used as mitogen (34). Each ex-
perimental condition was tested nine times. At the end of the
incubation period (5 days), wells were pooled to generate sin-
gle triplicates. Cells were washed with PBS pH 7.4, and indir-
ectly labeled with either isotype antibody or a purified mAb
against CD26 (TP1/16) plus GAM-APC. Unlabeled (CFSE2)
cells served as negative controls, while unstimulated CFSE-la-
beled PBMCs allowed us to identify those lymphoblasts that
did not divide.
For the multicolor extracellular staining of unfractionated
leukocytes, either isotype (isotype-FITC, isotype-PE, isotype-
APC) or specific (CD26-TP1/19-FITC, CD127-PE, CD4-
PerCP, or CD25-APC) monoclonal antibodies were added to
test tubes. Hundred microliters of EDTA anticoagulated whole
blood was mixed with antibodies by gentle vortexing and left
for 30 min (RT). Afterwards, erythrocytes were lysated (2 ml
13 BD FACS lysing solution/test tube; 15 min, RT), and sam-
ples centrifuged (200g, 5 min, RT), washed with 2 ml of PBS
pH 7.4, 1% FBS, 0.1% sodium azide (washing solution), and
fixed with 2 ml ice-cold 2% PFA in PBS pH 7.4 (30 min, RT).
Finally, cells were washed again and resuspended in 1 ml of
washing solution.
During the simultaneous detection of extracellular (CD4,
CD25, CD26, CD49d, CD39) and intracellular (FoxP3) pro-
teins in either resting or PHA-activated PBMCs, samples were
washed with BD Pharmingen Stain Buffer, counted (hemocyt-
ometer) and adjusted to 10 3 106 cells/ml. For extracellular
labeling, adequate volumes of isotype (isotype-FITC and iso-
type-APC) or specific antibodies (CD26-FITC TP1/16, CD4-
PerCP, and either CD25-APC or CD49d-APC) were placed
inside flow cytometry tubes and 1 3 106 cells (100 ll) added.
In some experiments (Fig. 5), samples were simultaneously
stained with CD49d-APC and CD127 Alexa Fluor 647 (same
fluorescence channel). After incubation (20 min, RT), cells
were washed (2 ml of BD Pharmingen Stain Buffer per tube)
and centrifuged at 250g 10 min RT. Then, cell pellet was fixed
with 13 Human FoxP3 buffer A (2 ml/tube; 10 min at RT),
centrifuged again (500g, 5 min, RT) and the wash step
repeated. Fixed and extracellularly labeled cells were permeab-
ilized with 13 Human FoxP3 buffer C (0.5 ml/tube) for 30
min at RT. After washing, permeabilized cells were incubated
(30 min, RT) with isotype-PE or anti-FoxP3-PE. Cells were
washed again, centrifuged (500g, 5 min, RT) and fixed (1 ml,
2% PFA) during 30 min at RT. Finally, leukocytes were resus-
pended in 1 ml of BD Pharmingen Stain Buffer per tube.
We used a similar protocol to evaluate the percentage
of IFN-c-producing T lymphoblasts. In brief, GolgiPlugTM
ORIGINAL ARTICLE
Cytometry Part A � 81A: 843�855, 2012 845

treated lymphoblasts were also extracellularly labeled with
diverse combinations of isotypic (FITC- and APC-labeled) or
specific (CD26-FITC TP1/16, CD25-FITC, CD4-PerCP, and
CD49d-APC or CD25-APC) mAbs. Once washed, cells were
fixed and permeabilized (20 min, 48C) with 250 ll of BD
Cytofix/Cytoperm buffer, washed two times with 1 ml of 13
BD Perm/WashTM and incubated with anti-IFN-c-PE or iso-
type-PE in 50 ll of 1X BD Perm/Wash (30 min, 48C). Finally,
leukocytes were washed twice (1 ml/each, 1X BD Perm/Wash)
and resuspended in BD Pharmingen Stain Buffer (1 ml).
In Vitro Suppression Assays
Human PBMCs were activated with 2 lg/ml PHA for 5
days to generate T lymphoblasts. To favor Treg cell prolifera-
tion (2- to 10-fold), culture medium was also supplemented
(day 3) with 200 IU/ml IL-2 (Peprotech). CD41CD25high
and CD41CD25highCD262/low Treg subsets were purified
from T lymphoblasts (Dynabeads1 Regulatory CD41CD251
T cell Kit; Life-Technologies, Spain) and tested for suppres-
sion in co-cultures with autologous CFSE-labeled PBMCs as
responder cells (1 3 105/well; 250 ll/well). The suppression
assay was performed in round-bottom 96-well plates with a
dilution series ranging from 1:1 to 4:1 of responder cells:Treg
cells. To induce proliferation, responder cells were stimulated
with beads coated with mAbs against CD2/CD3/CD28 (Treg
suppression inspector human, Miltenyi Biotec, Auburn, CA;
1 bead per cell) in the presence of 200 IU/mL IL-2 for 4 to 5
days. Additionally, responder cells and Treg cells were cul-
tured separately, either with or without beads/IL-2. Measure-
ments were always carried out in triplicate. Potency of
suppression was calculated at 1:1 ratio as [1-(proliferation
of Treg:responders coculture/proliferation of responders
alone)].
Statistics
P values were calculated by the nonparametric Mann–
Whitney U-test using IBM SPSS statistics 19 software. P values
=0.05 were considered to be significant.
RESULTS
As previously reported (29), resting lymphocytes express
low levels of CD26. However, CD26 is upregulated during cell
activation (plateau phase at 4–6 days; Fig. 1A), and CD26
levels are positively correlated with cell size (forward scatter/
FSC measurements) (Fig. 1A). CD26 up regulation takes place
especially amongst CD4 T-cells (Fig. 1B), and actively dividing
T lymphocytes display higher numbers of CD26 molecules as
they progress through new division rounds (Fig. 1C, 5-days
PHA blasts, [90% enriched in CD31 T cells; see CD26 geo-
mean values plotted against the number of cell divisions in
line graph). Moreover, as the percentage of IFN-c1 secreting
CD41 T cells that also stained for CD26 after 5 days of stimu-
lation with PHA is very high (81.5 � 15.7%; n 5 5 donors),
this extracellular peptidase allows tracing IFN-c-producing
TH1 cells. Therefore, CD26 can be used, at least, as an activa-
tion/TH1 marker.
Treg cells display an ‘‘activated-like’’ (CD41CD25high
CD45RO1) phenotype. Consequently, we set out to determine
whether Treg cells at the steady-state expressed CD26. About
5.3 � 2.2% (n 5 10; data not shown) of resting CD41 T lym-
phocytes are CD251 (2,12), but only those displaying the
highest expression of CD25 (CD25high, �1–2% CD41 cells)
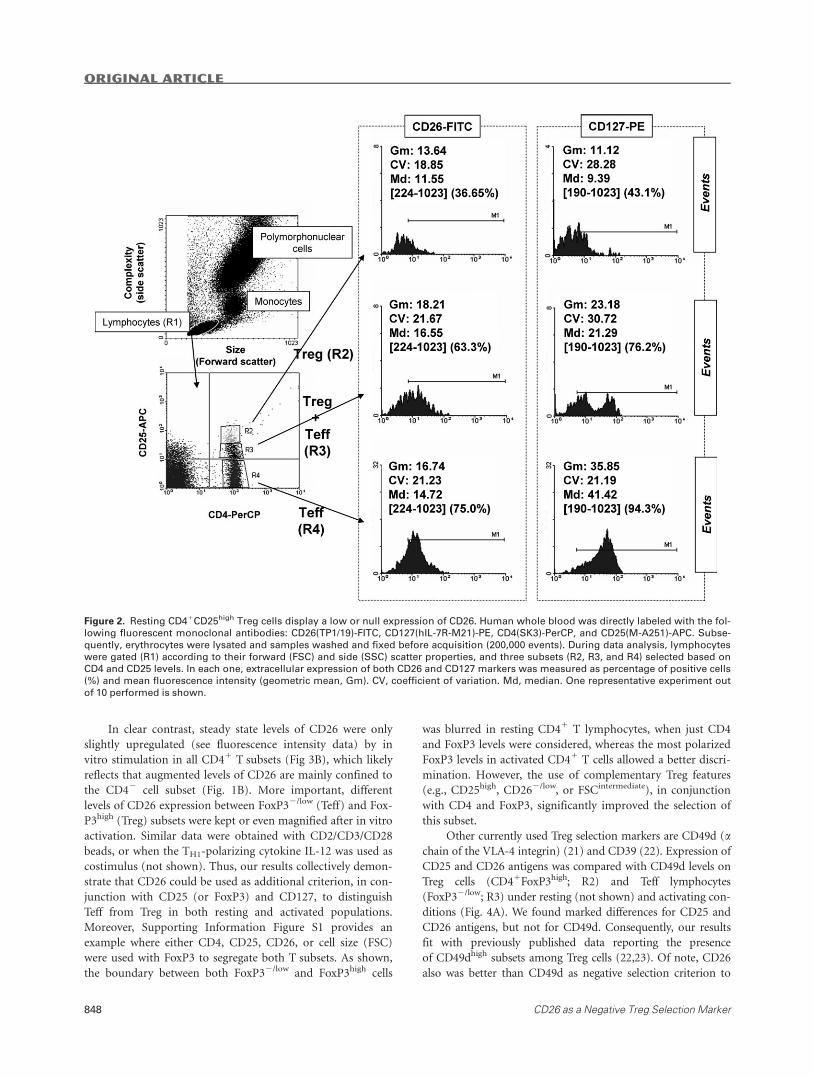
are actual Treg cells (Fig. 2) (2). In contrast, CD41 popula-
tions with intermediate levels of CD25 (CD25low) contain a
mixture of Teff and immature Treg lymphocytes (2). This fact,
together with the continuous expression of CD25, makes the
boundary for CD25high Treg populations not exempt of arbi-
trariness (10). However, we observed that CD41CD25high Treg
lymphocytes (R2 gate) were the subset with the lowest per-
centage of positive cells and the most reduced molecular den-
sity for both CD26 and CD127 antigens (Fig. 2). In contrast,
high levels of CD127 and CD26 were found amongst
CD41CD252 Teff cells (R4 gate), whereas CD41 lymphocytes
expressing intermediate densities of CD25 (R3 gate) consisted
of regulatory (CD262/lowCD1272/low) and effector
(CD261CD1271) populations (more clearly observed for
CD127). We confirmed these results with three different CD26
mAbs clones (TP1/19, TP1/16, and L272), pointing out that
this result does not depend on a particular epitope. Thus,
when CD26 levels were measured with the TP1/16 mAb
(�20% stronger staining than the TP1/19 mAb in Fig. 2), the
following results were obtained: 68.3 � 12.2% of CD41CD25high
T cells, 76.9 � 11% of CD41CD251, and 91.4 � 5.3% of
CD41CD252 T cells were CD261 (n 5 10). In addition, intra-
cellular staining of CD41CD25high Treg cells also revealed low
levels of CD26; i.e., there is not internal pool (not shown).
FoxP3 is still the preferable marker to accurately identify
the Treg subset, even though the existence of nonregulatory
FoxP3low T cells in normal individuals also precludes the use
of FoxP3 as a sole marker for Treg cells (2). For that reason,
cell size (FSC), CD25, CD127, and especially, CD26 levels,
were evaluated in resting CD41FoxP3high (R2), CD41FoxP3low
(R3), and CD41FoxP32 (R4) T cells from healthy individuals
(Fig. 3A). Our results clearly showed that, compared with
CD41FoxP32/low lymphocytes (R3 and R4), CD41FoxP3high
Treg cells (R2) were mainly CD25high leukocytes with an interme-
diate size and a dim expression of CD4, CD127, and CD26
(82.2–85.2% vs. 46.5% for CD127; 82.9–85.2% vs. 39.4% for
CD26). Thus, our findings suggest that CD262/low levels are
linked to Treg cells in resting lymphocyte populations, while high
CD26 levels can be found in CD41 subsets with effector func-
tions, which is in line with data from Mandapathil et al. (24).
However, both CD25 and CD26 antigens also reflect the
cell activation status. Consequently, we wanted to exclude the
possibility of blurry limits between different T helper subsets
for CD26 levels because of cell activation. Thus, data in resting
PBMCs were compared with those obtained after subjecting
the same cells to 5 days of in vitro polyclonal activation (Fig
3B). As expected, the nonspecific mitogenic stimuli (PHA)
caused an intense FoxP3 up regulation in both CD41 and
CD81 T cells (16), as well as augmented levels of CD25 within
the CD41FoxP32/low (R3 and R4) and, especially, the
CD41FoxP3high Treg subset (R2 gate) (14). On the contrary,
ORIGINAL ARTICLE
846 CD26 as a Negative Treg Selection Marker

the percentage of CD1271 cells (but not the fluorescence in-
tensity value) was diminished (Fig. 3B) in all CD41 T subsets
(R2-R4) (19,20). Therefore, our data support the notion that
there is not a clear boundary between Treg and Teff lympho-
cytes under activating conditions when selection is exclusively
based on extracellular markers such as CD25 or CD127.
Figure 1. CD26 is an activation marker. (A) Human PBMCs, cultured and activated with 2 lg/ml PHA for 5 days in 24-well plates (0.25 3 106
cells/ml), were harvested and immunostained with CD4(SK3)-PerCP and CD26(TP1/16)-FITC. During flow cytometry data acquisition
(20,000 events) and analysis, lymphocytes were gated (R1; not shown) according to their size (FSC) and complexity (SSC), three subsets
differentiated based on cell size (small, medium, big), and CD26 levels detected in each one of these subpopulations. The discontinuous
line indicates the CD26 expression in resting lymphocytes (day 0). (B) CD26 levels in CD41 and CD42 populations from resting (day 0) and
PHA-activated lymphocytes (day 5). (C) CFSE-labeled PBMCs, seeded at 0.25 3 106 cells/ml in 96-round well microplates, were either kept
unstimulated (upper left dot plot) or activated during 5 days with 2 lg/ml PHA (upper right dot plot). Then, cells were indirectly labeledwith isotype control antibody or mAb against CD26 (TP1/16) plus GAM-APC, and CD26 expression (percentage and geomean) plotted
against the number of cell divisions of gated lymphocytes (FSC vs. SSC plot; R1 gate, not shown). Results in (A), (B) and (C) belong to rep-
resentative experiments. Data in (A) and (C) are shown as mean � standard deviation (SD) of triplicate measurements.
ORIGINAL ARTICLE
Cytometry Part A � 81A: 843�855, 2012 847
In clear contrast, steady state levels of CD26 were only
slightly upregulated (see fluorescence intensity data) by in
vitro stimulation in all CD41 T subsets (Fig 3B), which likely
reflects that augmented levels of CD26 are mainly confined to
the CD42 cell subset (Fig. 1B). More important, different
levels of CD26 expression between FoxP32/low (Teff) and Fox-
P3high (Treg) subsets were kept or even magnified after in vitro
activation. Similar data were obtained with CD2/CD3/CD28
beads, or when the TH1-polarizing cytokine IL-12 was used as
costimulus (not shown). Thus, our results collectively demon-
strate that CD26 could be used as additional criterion, in con-
junction with CD25 (or FoxP3) and CD127, to distinguish
Teff from Treg in both resting and activated populations.
Moreover, Supporting Information Figure S1 provides an
example where either CD4, CD25, CD26, or cell size (FSC)
were used with FoxP3 to segregate both T subsets. As shown,
the boundary between both FoxP32/low and FoxP3high cells
was blurred in resting CD41 T lymphocytes, when just CD4
and FoxP3 levels were considered, whereas the most polarized
FoxP3 levels in activated CD41 T cells allowed a better discri-
mination. However, the use of complementary Treg features
(e.g., CD25high, CD262/low, or FSCintermediate), in conjunction
with CD4 and FoxP3, significantly improved the selection of
this subset.
Other currently used Treg selection markers are CD49d (achain of the VLA-4 integrin) (21) and CD39 (22). Expression of
CD25 and CD26 antigens was compared with CD49d levels on
Treg cells (CD41FoxP3high; R2) and Teff lymphocytes
(FoxP32/low; R3) under resting (not shown) and activating con-
ditions (Fig. 4A). We found marked differences for CD25 and
CD26 antigens, but not for CD49d. Consequently, our results
fit with previously published data reporting the presence
of CD49dhigh subsets among Treg cells (22,23). Of note, CD26
also was better than CD49d as negative selection criterion to
Figure 2. Resting CD41CD25high Treg cells display a low or null expression of CD26. Human whole blood was directly labeled with the fol-
lowing fluorescent monoclonal antibodies: CD26(TP1/19)-FITC, CD127(hIL-7R-M21)-PE, CD4(SK3)-PerCP, and CD25(M-A251)-APC. Subse-
quently, erythrocytes were lysated and samples washed and fixed before acquisition (200,000 events). During data analysis, lymphocytes
were gated (R1) according to their forward (FSC) and side (SSC) scatter properties, and three subsets (R2, R3, and R4) selected based on
CD4 and CD25 levels. In each one, extracellular expression of both CD26 and CD127 markers was measured as percentage of positive cells
(%) and mean fluorescence intensity (geometric mean, Gm). CV, coefficient of variation. Md, median. One representative experiment out
of 10 performed is shown.
ORIGINAL ARTICLE
848 CD26 as a Negative Treg Selection Marker

discriminate IFN-c-producing cells within the CD41CD25high
compartment in PHA-activated PBMCs (Supporting Information
Fig. S2).
Unlike CD49d, CD39 is a marker with a clearly divergent
expression pattern for Treg (R2; �72% CD391) and Teff (R3;
�3% CD391) cells cultured under activating conditions
(Fig. 4B), but not at the steady state (data not shown; Treg,
�3% CD391; Teff �0.1% CD391). Moreover, CD39 is linked
to adenosine production (22), an immunosuppressive molecule
whose accumulation is also favored by the CD262/low phenotype
of activated CD391 Treg cells (Fig 4B), since the CD26 protease
is the main adenosine deaminase (ADA) binding protein (28).
Figure 3. Phenotypic stability of CD41CD25highCD262/low Treg cells upon activation. PBMCs were isolated by Ficoll density gradient
separation, and either directly labeled (resting, day 0) (A) or activated in complete medium supplemented with 1 lg/ml PHA before immu-nofluorescent labeling with antibodies (activated, 5 days) (B). In both cases, cells were initially incubated with the following mAbs: CD26-
FITC (clone TP1/16), CD4-PerCP (clone SK3), and either CD25-APC (clone M-A251) or CD127-Alexa Fluor 647 (clone hIL-7R-M21). Then, cells
were fixed/permeabilized and stained with anti-FoxP3-PE (clone 259D/C7). Background values were set with the corresponding isotype
mAbs. During data analysis (WinMDI), the lymphocyte population was selected according to its cell size (FSC) and complexity (SSC) (R1).
Three subpopulations of CD41 T cells were identified as a function of its FoxP3 expression (R2, CD41FoxP3high Treg; R3, CD41FoxP3low;
R4, CD41FoxP32), and the level of CD25, CD127, and CD26 as well as the cell size (FSC) quantified afterwards. Numbers on top of histo-
grams represent the percentage of positive cells for each marker (%) as well as the corresponding intensity of fluorescence (geomean; in
brackets). Data belong to one representative experiment out of four performed.
ORIGINAL ARTICLE
Cytometry Part A � 81A: 843�855, 2012 849

According to Kleinewietfeld et al. (21), antibodies against
CD49d and CD127 alone are sufficient to isolate ‘‘untouched’’
FoxP31 Treg cells free of contaminating CD251 Teff cells. To
determine whether the combined use of these two markers is
enough to accurately discriminate Treg cells from activated
Teff cells, ‘‘untouched’’ CD41CD127lowCD49d2 Treg lympho-
cytes were purified from resting or in vitro expanded PBMCs
using the EasySepTM Human CD41CD127lowCD49d2 Regula-
tory T Cell Enrichment Kit (StemCell Technologies). Treg cells
isolated from resting PBMCs were a homogeneous population
(see CD4 and FoxP3 levels), mostly negative/low for CD127
and CD49d (as expected) and with a homogeneous CD262/low
phenotype (see R2 and R3 gates in Fig. 5A). On the contrary,
the CD41CD127lowCD49d2 cell fraction purified from acti-
vated PBMCs was heterogeneous, whether regarding the level
of CD4 and FoxP3 molecules or according to CD26 levels (see
Figure 4. Levels of CD26, CD49d and CD39 antigens in Treg (FoxP3high) and Teff (FoxP3low) lymphocytes. PBMCs were purified and
expanded with PHA during 5 days before inmunofluorescent labeling. Cells were extracellularly stained with CD4-PerCP (cloneSK3) and
different combinations of CD26-FITC (clone TP1/16), CD39-FITC (clone TU66), CD25-APC (clone M-A251), CD26-APC (clone TP1/19), and
CD49d-APC (clone 9F10) mAbs. After that, cells were fixed/permeabilized and stained with anti-FoxP3-PE (clone 259D/C7). During analysis,
lymphocytes were gated (size vs. complexity plots; R1) and two subpopulations (R2 and R3) distinguished based on CD4 and FoxP3
expression: CD41FoxP31/high Treg cells (R2; depicted as grey color in A) and CD41FoxP3low/2 Teff lymphocytes (R3; depicted as black color
in A). Finally, either plasma membrane levels of CD25, CD26, and CD49d, each one in combination with FoxP3 (A), or both CD39 and CD26
antigens (B), were evaluated. The figure shows one representative experiment out of three performed.
ORIGINAL ARTICLE
850 CD26 as a Negative Treg Selection Marker

R2 and R3 gates in Fig. 5B). This heterogeneity was caused by
a preferential isolation of a FoxP3low subset (R3), which dis-
plays a higher CD26 expression than the FoxP3high subset (R2)
(Fig. 5B). Due to the activation-dependent reduction of
CD1271 cells within the Teff subset (Fig. 3), we speculate that
this FoxP3lowCD261 phenotype (R3) may be the signature of
T cells with unstable FoxP3 expression, activated-memory
phenotype, and the capability of producing IL-2, IFN-c or
IL-17 (35,36).
According to our findings, inclusion of CD26-specific
mAbs in Treg purification protocols may help to deplete con-
taminating FoxP3low T cells and allow the isolation of highly
enriched FoxP3high cells, especially from previously expanded
‘‘bulk’’ T-cell populations or samples from inflammatory/
autoimmune diseases. Most of commercially available proto-
cols are based on the CD41CD25high phenotype. According to
our data in resting T lymphocytes, 81.9 � 11.2% (n 5 10) of
CD41CD251 T cells were CD262. Thus, unlike the use of
CD45RA to isolate ‘‘naive’’ CD41CD25high Treg cells (24–61%
of resting Treg cells) (37), the CD26 strategy should yield
higher Treg numbers (38). As a first approach, we determined
by means of a pilot flow cytometry experiment whether the
strategy of selecting the CD41CD25highCD262/low yielded
more homogeneous Treg populations (according to cell size
and FoxP3; Supporting Information Fig. S3) than the usual
CD41CD25high phenotype. We used a broader gating strategy
and a lower than usual CD25 boundary. CD41CD252 T cells
were also gated for comparison, revealing that �95% of them
(R4 gate) were FoxP32/low cells. However, FoxP3low cells
(40–60%) were also detected amongst the CD41CD25high
Treg lymphocytes (R2 1 R3 gates). In contrast, the use of a
CD41CD25highCD262 phenotype (R2 gate) to select the Treg
subset yielded a more homogeneous population, as the per-
centage of FoxP3high cells reveals (in Supporting Information
Fig. S3, 43%?61% for resting PBMCs, and 59?75% for acti-
vated PBMCs).
Given these preliminary data, both CD41CD25high Treg
and CD41CD252 Teff lymphocytes were isolated from either
freshly isolated or PHA-expanded PBMCs by means of the
Dynabeads1 Regulatory CD41CD251 T cell Kit (Life Tech-
nologies/Invitrogen). As expected, the starting percentage of
Treg cells amongst resting PBMCs was low (0.84 � 0.37%; n
5 6), recovering �86% (n 5 3) of these Treg cells as
CD41CD25high fractions with a cell purity of 87% (assessed by
CD25 expression; n 5 3). In contrast, we have detected a
slightly higher starting percentage of CD41CD25high cells
amongst PHA-activated PBMCs (3.34 � 2.08%, n 5 6). Only
68% (n 5 7) of these lymphocytes was collected within
CD41CD25high fractions. Moreover, the percentage of positive
cells for CD25 and FoxP3 was, respectively, 83.6 � 5.14% and
63.63 � 9.12% (n 5 3), which can be explained because of the
presence of activated CD251 Teff cells.
To improve the purity of Treg cells obtained from PHA-
expanded PBMCs, we partially removed CD261 cells from
CD41 T lymphocytes by means of the addition of TP1/16
mAb to the cocktail of mouse IgG mAbs during the first step.
Consequently, a significant decrease in the number of CD41
Figure 5. The combined use of CD127 and CD49d does not avoid
the presence of FoxP3lowCD26high T cells within Treg populations
isolated from activated PBMCs. Regulatory T cells were purified
from resting (A) or activated PBMCs (1 lg/ml PHA, 5 days) usingthe EasySepTM Human CD41CD127lowCD49d2 Regulatory T Cell
Enrichment Kit (StemCell Technologies). Treg cells were stained
with CD26-FITC (clone TP1/16), anti-FoxP3-PE (clone 259D/C7),
CD4-PerCP (clone SK3), and CD49d-APC/CD127-AlexaFluor 647
(Clone 9F10/hIL-7R-M21) mAbs. During analysis (A and B), Treg
lymphocytes were gated (forward vs. side scatter; R1) and two
subpopulations (R2 and R3) distinguished based on the expres-
sion of CD4 and FoxP3: CD41FoxP3high (R2) and CD41FoxP3low
(R3). Finally, both CD26 and CD127/CD49d levels were evaluated
for R2 and R3 in new density plots, reflecting that Treg lympho-
cytes purified from PBMCs (A) are a more homogeneous popula-
tion as compared with the Treg population isolated from activated
PBMCs (B). A high mortality rate has been constantly observed
for Treg cells purified with this protocol (see forward vs. side scat-
ter plot is in A and B). One representative experiment out of three
performed is shown.
ORIGINAL ARTICLE
Cytometry Part A � 81A: 843�855, 2012 851

T cells was noted: 27.39 � 8.93 ?18.67 � 5.53 (3 106) CD41
T cells, 28.84 � 5.47% reduction (n 5 7). Once purification
was completed, flow cytometric analysis revealed a low expres-
sion of CD26 in CD41CD252 Teff cells and a bimodal pattern
for the CD41CD25high Treg subset, which comprises CD262/
low and CD26high subtypes (Fig. 6A). The use of mAbs against
CD26 caused an almost full depletion of CD261 cells in
CD41CD252 Teffs, but only a partial decline amongst
CD41CD25high Tregs (�40%; Figs. 6B and 6C). However, de-
spite this incomplete CD26 depletion, the percentage of positive
cells for CD25 (83.6 � 5.14% ? 91.2 � 5.05%; n 5 3) and
FoxP3 (63.63 � 9.12% ? 75.33 � 10.91%; n 5 3) (Fig. 6C)
was raised, as well as the percentage of CD25highFoxP3high cells
(e.g., from 66.8 to 84.5% in Figs. 6A and 6B). Moreover, there
was a significant rise in the number of CD1271 Treg cells after
CD26-depletion (8.9 � 9.2% CD1271 Treg ? 29.7 � 0.8%
CD1271 Treg, n 5 2 donors), which fits with a higher
suppressive activity (Fig. 6D; 8.29 � 2.17% ? 17.98 � 1.93%).
DISCUSSION
CD26 is an activation/memory marker capable of trans-
mitting costimulatory signals (28,30). Thus, interaction
between CD26 (T cell) and caveolin-1 (APCs) leads to NF-jB
activation and recruitment of CARMA1/Bcl10/IjB (28,33), a
complex implied in thymic development of Treg cells (39),
while blockade of this costimulatory interaction causes T cell
cycle arrest (40,41). Nevertheless, CD26 may also have inhibi-
tory functions. For example, disease severity is increased in
CD262/2 mice in certain autoimmune models (42,43), with
augmented T-cell proliferation rates (43) and production of
TH1 cytokines (IFN-c, TNF-a) (42,43) but diminished TGF-b1
synthesis (43). Indeed, several inhibitory peptides or naturally
occurring ligands of CD26/DPPIV promote the secretion of
TGF-b1 (43-45). Therefore, all these findings point to some
kind of connection between CD26 and the Treg phenotype
that deserves to be ascertained, especially because Treg lym-
phocytes, with an ‘‘activated-like’’ (e.g., CD25high, HLA-DR1)
and ‘‘memory’’ (CD45RO1) phenotype, were expected to be
CD261 (46,47) but not CD262/low cells. However, our results
are in agreement with the CD262/low profile of Treg-like leu-
kocytes in classical Hodgkin’s lymphoma (48). Moreover, they
fit with recent studies in human Treg cells (24), the latest in
April this year (49), where authors describe variable CD26
levels in different TH subsets: TH17[TH1[TH2[Treg cells.
Nevertheless, our data go beyond, as they show that this
CD262/low phenotype is stable upon in vitro T-cell expansion
and can be used during the isolation/identification of Treg
cells. In addition, this CD262/low phenotype may explain why
the TH1 cytokine IL-12 causes a strong upregulation of CD26
on ‘‘bulk’’ T-cell cultures, whereas IL-2 (CD25-ligand) induces
only a slight increase (29). Still, several questions remain
unanswered: for example, whether this CD262/low phenotype
is based on differential transcription/translation, shedding, or
exosomes/microvesicles releasing processes, whether CD26
level on Tregs depends on their tissue localization (50), or how
the various Treg/Teff cells ratios found in different diseases
correlate with levels of circulating sCD26 (51).
Researchers can only get high numbers of Treg cells by
means of tedious two-step magnetic procedures based on
CD25 (35-38). However, CD25 is also expressed by FoxP3low T
cells, a group of lymphocytes that might fit with naıve or
uncommitted Tregs but also activated Teff cells (2,37,52,53).
For that reason, more efficient technologies have been placed
on the research market. Nevertheless, these new protocols may
not be entirely effective yet, as our article points out. Thus,
negative selection antigens, like CD127 (17,18) or CD49d
(21), certainly provide an additional way of ascertaining the
right phenotype. Nevertheless, this does not seem to be suffi-
cient because of the intense reduction in CD1271 cells upon
TH-cell activation (19,20,54, our data), the presence of
CD127high subsets within activated Treg cells (50, our data), or
the small differences for CD49d between FoxP3high and
FoxP32/low cells (our data).
The limited amount of biological samples or the low
abundance of Treg cells in peripheral blood are additional pro-
blems. On this regard, several protocols have been developed
for the in vitro expansion of Treg cells (35-38), but many of
them are associated with potential risks: e.g., the in vitro out-
growth of contaminating Teff cells or the possibility of
‘‘plastic’’ Treg cells reverting to FoxP3low Teff cells (2,35,37).
To circumvent these problems, CD45RA have been promoted
as a discriminating marker to identify a subset of naive
CD41CD25high Treg cells that maintain FoxP3 expression dur-
ing in vitro culture (37). However, it remains to be determined
if there is a sufficient number of such ‘‘naive’’ Treg cells in
blood samples of patients for in vitro expansion and autolo-
gous adoptive transfer (37), especially considering the age-de-
pendent differentiation from naive to effector memory Treg
cells (55). Moreover, it is known that even CD45RAhigh Treg
cells can give rise to nonregulatory T cells when cultured in a
milieu containing high-dose IL-2 (2,35,37). In clear contrast,
our study suggests using a different strategy. This new
approach consist in the in vitro expansion of bulk lympho-
cytes before isolating the Treg subset on the basis of a more
reliable Treg/Teff differentiation marker: CD26. Thus, antibo-
dies against this peptidase could be included during the first
negative selection step of any magnetic protocol to get rid of
unwanted CD261/high Teff (TH1, TH2, TH17) lymphocytes and
achieve higher numbers of either CD41CD25high or
CD41CD39high Treg populations (our data, 24,49). Moreover,
the phenotype CD41CD127lowCD49d2CD262 may be also
enough to select Treg cells through a fast and activation-degree
independent ‘‘single-step’’ purification protocol.
Which functional explanation there might be for this
CD262/low phenotype in human Treg cells? Several ligands
have been found for CD26, as the tyrosine phosphatase
CD45RO (31,32) or adenosine deaminase (ADA). The last one
is an ectoenzyme involved in degradation of adenosine, which
is an immunosuppressive molecule (22,56). Consequently, a
low level of CD26 could alter the membrane location of
CD45RO (32) or reduce the amount of surface ADA on Treg
cells (57), which may explain either their anergic phenotype in
vitro or their net production of adenosine (22). Alternatively,
interaction of Caveolin-1 (APC) with CD26 (T-cell) leads to
ORIGINAL ARTICLE
852 CD26 as a Negative Treg Selection Marker

Figure 6. The use of CD26 as a negative selection marker enhances the levels of FoxP3 and CD25 in CD41CD25high Treg populations iso-
lated from activated PBMCs. CD41CD25high Treg cells were purified from activated PBMCs (1 lg/ml PHA, 5 days) using the Dynabeads1
Regulatory CD41CD251 T Cell Kit (Life Technologies/Invitrogen). During the depletion of non-CD41 cells, the cocktail of monoclonal antibo-
dies was supplemented (B) or not (A) with CD26 (TP1/16) mAb in order to deplete CD261 cells. Isolated CD41CD25high Treg and CD41CD252
Teff cells were stained with CD26-FITC (TP1/16), anti-FoxP3-PE (259D/C7), CD4-PerCP (SK3), and CD25-APC (M-A251) mAbs. During analy-
sis, viable CD41 lymphocytes were selected (R1 and R2) and FoxP3/CD25 levels and CD26 expression evaluated. According to CD26, two
different subsets (CD262/low and CD26high) were observed within CD41CD25high Treg cells (A), which points out to the presence of contami-
nating activated Teff cells (CD26high). Reduction of this CD26high subset within the CD41CD25high Treg lymphocytes raised the percentage
of CD25/FoxP3 double positive cells (B). (C) Columns plot summarizes the percentage of reduction/increase in the fluorescence intensity of
CD26, FoxP3, and CD25 in Tregs from three different healthy donors after the partial depletion of CD261 cells (mean � standard deviation).(D) Suppression of autologous (CFSE-labeled) responder PBMCs proliferation (y-axis; % of cells that divided at least once) by coculture
with increasing numbers of either CD41CD25high (Treg) or CD41CD25highCD262/low (CD26-depleted Treg) lymphocytes isolated from
preactivated (PHA 1 IL-2; 5 days) PBMCs (x-axis). Measurements were carried out in triplicate. Significant results are indicated by single
asterisks (P value is =0.05; Mann-Whitney U-test).
ORIGINAL ARTICLE
Cytometry Part A � 81A: 843�855, 2012 853

the up regulation of CD86 on APCs (58). Interestingly, Treg
lymphocytes down-regulate CD80 and CD86 on APCs
through a CTLA-4-dependent (9) mechanism. CD80/CD86
molecules are differentially regulated (59) and seem to have
divergent functions (60). For example, CD86 is highly respon-
sive to proinflammatory stimuli (LPS, TNFa, IFN-c, CD40-
CD40L ligation) and shows a preference for CD28, whereas
CD80 seems rather specific for the inhibitory receptor CTLA-
4 (60,61). Therefore, if we assume a co-stimulatory role for
the CD26/caveolin-1 interaction and a subsequent CD86 (but
not CD80) upregulation on APCs (58), a CD262/low pheno-
type in Treg cells may contribute to keep low levels of proin-
flammatory CD86 molecules on APCs, leading indirectly to a
lack of response (anergy) or apoptosis in antigen-primed naıve
T cells.
In conclusion, the present data substantiate a link between
null/low levels of CD26 and immunosuppressive functions
within either CD41CD25high, CD41CD39high, or
CD41CD1272/lowCD49d2 subsets. In addition, they support
further investigation to determine whether the CD262/low phe-
notype is a reliable alternative for the delineation or purification
of human Treg cells by means of extracellular markers, which
may stimulate the arising of new protocols and the progress in
Treg field. Finally, a patent application for the data described in
this paper was done on January 26, 2010, which has recently
entered the Patent Cooperative Treaty (PCT) phase.
ACKNOWLEDGMENTS
The authors are grateful to the Medical Service at the
University of Santiago, and to Dr. Juan E. Viuela, Immunology
Service, Clinic University Hospital of Santiago (CHUS), for
technical assistance. In addition, the authors want to thank
Ana M. Carballido and Alejandro Gonzalez for their colla-
boration.
LITERATURE CITED
1. Lu L, Cantor H. Generation and regulation of CD81 regulatory T cells. Cell MolImmunol 2008;5:401–406.
2. Sakaguchi S, Miyara M, Costantino CM, Hafler DA. FOXP31 regulatory T cells inthe human immune system. Nat Rev Immunol 2010;10:490–500.
3. Feuerer M, Hill JA, Mathis D, Benoist C. Foxp31 regulatory T cells: Differentiation,specification, subphenotypes. Nat Immunol 2009;10:689–695.
4. Sakaguchi S, Ono M, Setoguchi R, Yagi H, Hori S, Fehervari Z, Shimizu J, TakahashiT, Nomura T. FoxP31CD251CD41 natural regulatory T cells in dominant self-toler-ance and autoimmune disease. Immunol Rev 2006;212:8–27.
5. Boros P, Bromberg JS. Human FOXP31 regulatory T cells in transplantation. Am JTransplant 2009;9:1719–1724.
6. Joosten SA, Ottenhoff TH. Human CD4 and CD8 regulatory T cells in infectious dis-eases and vaccination. Hum Immunol 2008;69:760–770.
7. Curotto de Lafaille MA, Lafaille JJ. Natural and adaptive FoxP31 regulatory T cells:More of the same or a division of labor? Immunity 2009;30:626–635.
8. Corthay A. How do regulatory cells work? Scand J Immunol 2009;70:326–336.
9. Oderup C, Cederbom L, Makowska A, Cilio CM, Ivars F. Cytotoxic T lymphocyte anti-gen-4-dependent down-modulation of costimulatory molecules on dendritic cells inCD41CD251 regulatory T-cell-mediated suppression. Immunology 2006;118:240–249.
10. Tang Q, Bluestone JA. Regulatory T-cell physiology and application to treat autoim-munity. Immunol Rev 2006;212:217–237.
11. Nishikawa H, Sakaguchi S. Regulatory T cells in tumor immunity. Int J Cancer2010;127:759–767.
12. Baecher-Allan C, Brown JA, Freeman GJ, Hafler DA. CD41CD25high regulatory cellsin human peripheral blood. J Immunol 2001;167:1245–1253.
13. Aerts NE, Dombrecht EJ, Ebo DG, Bridts CH, Stevens WJ, De Clerck LS. Activated Tcells complicate the identification of regulatory T cells in rheumatoid arthritis. CellImmunol 2008;251:109–115.
14. Jourdan P, Vendrell JP, Huguet MF, Segondy M, Bousquet J, Pene J, Yssel H. Cyto-kines and cell surface molecules independently induce CXCR4 expression on CD41CCR71 human memory T cells. J Immunol 2000;165;716–724.
15. Dieckmann D, Plottner H, Berchtold S, Berger T, Schuler G. Ex vivo isolation andcharacterization of CD4(1)CD25(1) T cells with regulatory properties from humanblood. J Exp Med 2001;193:1303–1310.
16. Tran DQ, Ramsey H, Shevach EM. Induction of FOXP3 expression in naive humanCD41FOXP32 T cells by T cell receptor stimulation is TGFb-dependent but does notconfer a regulatory phenotype. Blood 2007;110:2983–2890.
17. Liu W, Putnam AL, Xu-Yu Z, Szot GL, Lee MR, Zhu S, Gottlieb PA, Kapranov P, Gin-geras TR, Fazekas de St Groth B, et al. CD127 expression inversely correlates withFoxP3 and suppressive function of human CD41 T reg cells. J Exp Med2006;203:1701–1711.
18. Seddiki N, Santner-Nanan B, Martinson J, Zaunders J, Sasson S, Landay A, SolomonM, Selby W, Alexander SI, Nanan R, et al. Expression of interleukin (IL)-2 and IL-7receptors discriminates between regulatory and activated T cells. J Exp Med2006;203:1693–1700.
19. Del Pozo-Balado M del M, Leal M, Mendez-Lagares G, Pacheco YM.CD4(1)CD25(1/hi)CD127(lo) phenotype does not accurately identify regula-tory T cells in all populations of HIV-infected persons. J Infect Dis 2010;201:331–335.
20. Alves NL, van Leeuwen EM, Derks IA, van Lier RA. Differential regulation of humanIL-7 receptor alpha expression by IL-7 and TCR signaling. J Immunol2008;180:5201–5210.
21. Kleinewietfeld M, Starke M, Di Mitri D, Borsellino G, Battistini L, Rotzschke O, FalkK. CD49d provides access to ‘‘untouched’’ human FoxP31 Treg free of contaminatingeffector cells. Blood 2009;113:827–836.
22. Borsellino G, Kleinewietfeld M, Di Mitri D, Sternjak A, Diamantini A, Giometto R,Hopner S, Centonze D, Bernardi G, Dell’Acqua ML, et al. Expression of ectonucleo-tidase CD39 by Foxp31 Treg cells: Hydrolysis of extracellular ATP and immune sup-pression. Blood 2007;110:1225–1232.
23. Venken K, Hellings N, Thewissen M, Somers V, Hensen K, Rummens JL, Medaer R,Hupperts R, Stinissen P. Compromised CD41 CD25(high) regulatory T-cell functionin patients with relapsing-remitting multiple sclerosis is correlated with a reducedfrequency of FOXP3-positive cells and reduced FOXP3 expression at the single-celllevel. Immunology 2008;123:79–89.
24. Mandapathil M, Lang S, Gorelik E, Whiteside TL. Isolation of functional human reg-ulatory T cells (Treg) from the peripheral blood based on the CD39 expression. JImmunol Methods 2009;346:55–63.
25. Alam MS, Kurtz CC, Rowlett RM, Reuter BK, Wiznerowicz E, Das S, Linden J, CroweSE, Ernst PB. CD73 is expressed by human regulatory T helper cells and suppressesproinflammatory cytokine production and Helicobacter felis-induced gastritis inmice. J Infect Dis 2009;199:494–504.
26. Moncrieffe H, Nistala K, Kamhieh Y, Evans J, Eddaoudi A, Eaton S, Wedderburn LR.High expression of the ectonucleotidase CD39 on T cells from the inflamed site iden-tifies two distinct populations, one regulatory and one memory T cell population. JImmunol 2010;185:134–143.
27. Bluestone JA, Thomson AW, Shevach EM, Weiner HL. What does the future hold forcell based tolerogenic therapy? Nat Rev Immunol 2007;7:650–654.
28. Ohnuma K, Dang NH, Morimoto C. Revisiting an old acquaintance: CD26 and itsmolecular mechanisms in T cell function. Trends Immunol 2008;29:295–301.
29. Cordero OJ, Salgado FJ, Viuela JE, Nogueira M. Interleukin-12 enhances CD26expression and Dipeptidyl peptidase IV function on human activated lymphocytes.Immunobiology 1997;197:522–533.
30. Huhn J, Ehrlich S, Fleischer B, von Bonin A. Molecular analysis of CD26-mediatedsignal transduction in T cells. Immunol Lett 2000;72:127–132.
31. Torimoto Y, Dang NH, Vivier E, Tanaka T, Schlossman SF, Morimoto C. Coassocia-tion of CD26 (dipeptidyl peptidase IV) with CD45 on the surface of human T lym-phocytes. J Immunol 1991;147:2514–2517.
32. Salgado FJ, Lojo J, Fernandez-Alonso CM, Alonso-Lebrero JL, Lluis C, Franco R,Viuela JE, Cordero OJ, Nogueira M. A role for IL-12 in the regulation of T cell plasmamembrane compartmentation. J Biol Chem 2003;278:24849–24857.
33. Ohnuma K, Uchiyama M, Yamochi T, Nishibashi K, Hosono O, Takahashi N, Kina S,Tanaka H, Lin X, Dang NH, et al. Caveolin-1 triggers T-cell activation via CD26 inassociation with CARMA1. J Biol Chem 2007;282:10117–10131.
34. Canda-Sanchez A, Salgado FJ, Perez-Dıaz A, Varela-Gonzalez C, Arias P, Nogueira M.IL-12-dependent activation of ERK1/2 in human T lymphoblasts. Immunobiology2009;214:187–196.
35. Hoffmann P, Boeld TJ, Eder R, Huehn J, Floess S, Wieczorek G, Olek S, Dietmaier W,Andreesen R, Edinger M. Loss of FOXP3 expression in natural human CD41CD251regulatory T cells upon repetitive in vitro stimulation. Eur J Immunol 2009;39:1088–1097.
36. Earle KE, Tang Q, Zhou X, Liu W, Zhu S, Bonyhadi ML, Bluestone JA. In vitroexpanded human CD41CD251 regulatory T cells suppress effector T cell prolifera-tion. Clin Immunol 2005;115:3–9
37. Hoffmann P, Eder R, Boeld TJ, Doser K, Piseshka B, Andreesen R, Edinger M. Onlythe CD45RA1 subpopulation of CD41CD25high T cells gives rise to homogeneousregulatory T-cell lines upon in vitro expansion. Blood 2006;108:4260–4267.
38. Riley JL, June CH, Blazar BR. Human T regulatory cell therapy: Take a billion or soand call me in the morning. Immunity 2009;30:656–665.
39. Molinero LL, Yang J, Gajewski T, Abraham C, Farrar MA, Alegre ML. CARMA1 con-trols an early checkpoint in the thymic development of FoxP31 regulatory T cells. JImmunol 2009;182:6736–6743.
40. Ohnuma K, Ishii T, Iwata S, Hosono O, Kawasaki H, Uchiyama M, Tanaka H, Yamo-chi T, Dang NH, Morimoto C. G1/S cell cycle arrest provoked in human T cells byantibody to CD26. Immunology 2002;107:325–333.
41. Ohnuma K, Uchiyama M, Hatano R, Takasawa W, Endo Y, Dang NH, Morimoto C.Blockade of CD26-mediated T cell costimulation with soluble caveolin-1-Ig fusion proteininduces anergy in CD41 T cells. Biochem Biophys Res Commun 2009;386:327–332.
ORIGINAL ARTICLE
854 CD26 as a Negative Treg Selection Marker

42. Busso N, Wagtmann N, Herling C, Chobaz-Peclat V, Bischof-Delaloye A, So A,Grouzmann E. Circulating CD26 is negatively associated with inflammation inhuman and experimental arthritis. Am J Pathol 2005;166:433–442.
43. Preller V, Gerber A, Wrenger S, Togni M, Marguet D, Tadje J, Lendeckel U, Rocken C,Faust J, Neubert K, et al. TGF-beta1-mediated control of central nervous systeminflammation and autoimmunity through the inhibitory receptor CD26. J Immunol2007;178:4632–4640.
44. Steinbrecher A, Reinhold D, Quigley L, Gado A, Tresser N, Izikson L, Born I, Faust J,Neubert K, Martin R, et al. Targeting dipeptidyl peptidase IV (CD26) suppressesautoimmune encephalomyelitis and up-regulates TGF-beta 1 secretion in vivo. JImmunol 2001;166:2041–2048.
45. Matteucci E, Giampietro O. Dipeptidyl peptidase-4 (CD26): knowing the functionbefore inhibiting the enzyme. Curr Med Chem 2009;16:2943–2951.
46. Skripuletz T, Schmiedl A, Schade J, Bedoui S, Glaab T, Pabst R, von Horsten S, Ste-phan M. Dose-dependent recruitment of CD251 and CD261 T cells in a novelF344 rat model of asthma. Am J Physiol Lung Cell Mol Physiol 2007;292:L1564–1571.
47. Stephens LA, Barclay AN, Mason D. Phenotypic characterization of regulatoryCD41CD251 T cells in rats. Int Immunol 2004;16,365–375.
48. Ma Y, Visser L, Blokzijl T, Harms G, Atayar C, Poppema S, van den Berg A. TheCD41CD262 T-cell population in classical Hodgkin’s lymphoma displays a distinc-tive regulatory T-cell profile. Lab Invest 2008;88:482–490.
49. Bengsch B, Seigel B, Flecken T, Wolanski J, Blum HE, Thimme R. Human Th17 cellsexpress high levels of enzymatically active dipeptidylpeptidase IV (CD26). J Immunol2012;188:5438–5447.
50. Simonetta F, Chiali A, Cordier C, Urrutia A, Girault I, Bloquet S, Tanchot C, Bour-geois C. Increased CD127 expression on activated FOXP31CD41 regulatory T cells.Eur J Immunol 2010;40:2528–2538.
51. Cordero OJ, Salgado FJ, Nogueira M. On the origin of serum CD26 and its alteredconcentration in cancer patients. Cancer Immunol Immunother 2009;58:1723–1747.
52. Komatsu N, Mariotti-Ferrandiz ME, Wang Y, Malissen B, Waldmann H, Hori S. Het-erogeneity of natural FoxP31 T cells: A committed regulatory T-cell lineage and an
uncommitted minor population retaining plasticity. Proc Natl Acad Sci USA2009;106:1903–1908.
53. Kmieciak M, Gowda M, Graham L, Godder K, Bear HD, Marincola FM, Manjili MH.Human T cells express CD25 and Foxp3 upon activation and exhibit effector/mem-ory phenotypes without any regulatory/suppressor function. J Trans Med 2009;7:89.
54. Potter SJ, Lacabaratz C, Lambotte O, Perez-Patrigeon S, Vingert B, Sinet M, Colle JH,Urrutia A, Scott-Algara D, Boufassa F, et al. Preserved central memory and activatedeffector memory CD41 T-cell subsets in human immunodeficiency virus controllers:An ANRS EP36 study. J Virol 2007;81:13904–13915.
55. Santner-Nanan B, Seddiki N, Zhu E, Quent V, Kelleher A, Fazekas de St Groth B,Nanan R. Accelerated age-dependent transition of human regulatory T cells to effec-tor memory phenotype. Int Immunol 2008;20:375–383.
56. Pacheco R, Martinez-Navio JM, Lejeune M, Climent N, Oliva H, Gatell JM, Gallart T,Mallol J, Lluis C, Franco R. CD26, adenosine deaminase, and adenosine receptorsmediate costimulatory signals in the immunological synapse. Proc Natl Acad Sci USA2005;102:9583–9588.
57. Cordero OJ, Salgado FJ, Fernandez-Alonso CM, Herrera C, Lluis C, Franco R,Nogueira M. Cytokines regulate membrane adenosine deaminase on human activatedlymphocytes. J Leukoc Biol 2001;70:920–930.
58. Ohnuma K, Yamochi T, Uchiyama M, Nishibashi K, Yoshikawa N, Shimizu N, IwataS, Tanaka H, Dang NH, Morimoto C. CD26 up-regulates expression of CD86 onantigen-presenting cells by means of caveolin-1. Proc Natl Acad Sci USA2004;101:14186–14191.
59. Larsen CP, Ritchie, SC, Hendrix R, Linsley PS, Hathcock KS, Hodes RJ, Lowry RP,Pearson TC. Regulation of immunostimulatory function and costimulatory molecule(B7-1 and B7-2) expression on murine dendritic cells. J Immunol 1994;152:5208–5219.
60. Rudy W, Guckel B, Siebels M, Lindauer M, Meuer SC, Moebious U. Differential func-tion of CD80- and CD86-transfected human melanoma cells in the presence of IL-12and IFN-c. Int Immunol 1997;9:853–860.
61. Zheng Y, Manzotti CN, Liu M, Burke F, Mead KI, Sansom DM. CD86 and CD80 dif-ferentially modulate the suppressive function of human regulatory T cells. J Immunol2004;172:2778–2784.
ORIGINAL ARTICLE
Cytometry Part A � 81A: 843�855, 2012 855


















