
CARACTERIZACION MOLECULAR Y SEROLOGICA DE VIBRIOSIS EN...
84
UNIVERSIDAD DE CHILE Facultad de Ciencias Químicas y Farmacéuticas CARACTERIZACIÓN MOLECULAR Y SEROLÓGICA DE AGENTES CAUSALES DE VIBRIOSIS EN SALMÓNIDOS EN EL SUR DE CHILE MEMORIA PARA OPTAR AL TÍTULO PROFESIONAL DE BIOQUÍMICO Andrés Alejandro Silva Rubio Santiago - Chile 2007
-
Upload
nguyenkhue -
Category
Documents
-
view
217 -
download
0
Transcript of CARACTERIZACION MOLECULAR Y SEROLOGICA DE VIBRIOSIS EN...

UNIVERSIDAD DE CHILE Facultad de Ciencias Químicas y Farmacéuticas
CARACTERIZACIÓN MOLECULAR Y SEROLÓGICA DE
AGENTES CAUSALES DE VIBRIOSIS EN SALMÓNIDOS
EN EL SUR DE CHILE
MEMORIA PARA OPTAR AL TÍTULO PROFESIONAL DE BIOQUÍMICO
Andrés Alejandro Silva Rubio
Santiago - Chile
2007

Directores de Memoria Dr. Beltrán Jaureguiberry
Claudia Acevedo, Bq., Med. Vet.
Laboratorio de Investigación y Desarrollo -Veterquímica Ltda.
Profesor Patrocinante
Dra. Daniela Seelenfreund
Dpto. de Bioquímica y Biología Molecular Facultad de Cs. Químicas y Farmacéuticas
Universidad de Chile

PUBLICACIONES EN ESTA TESIS
Este trabajo ha dado lugar a las siguientes publicaciones:
1. Antigenic and molecular characterization of Vibrio ordalii strains isolated from Atlantic
salmon (Salmo salar) in Chile” enviado a revision a la revista Diseases of Aquatic
Organisms.
2. “First report of Listonella (V) anguillarum isolated from salmonids farming in brackish
water and seawater in the south of Chile” enviado a revision a la revista Bulletin of the
European Association of Fish Pathologists.
Esta tesis fue realizada gracias a un proyecto FONTEC Código 204-4044 “Vacunas Autógenas
Especificas para la Salmonicultura Chilena Usando Herramientas Biotecnológicas” otorgado a la empresa Veterquímica Ltda.

AGRADECIMIENTOS
Gracias a la gente que me a apoyado, que ha creído en mi y he sido parte de sus
vidas durante estos últimos años, a ellos expreso mi gratitud por sus enseñanzas por
contribuir con su experiencia y forma de ver la vida.
En primer lugar agradezco al Dr. Hugo Zunino como parte de la empresa
Veterquímica Ltda, la que me ha acogido y apoyado, a mis directores de memoria el Dr.
Beltrán Jaureguiberry y Claudia Acevedo, los que me han dado la oportunidad de
desarrollar el tema con gran libertad confiando en mi desempeño profesional, pero
estando siempre presentes y dispuestos para ayudar en caso de problemas
experimentales y también han sido un gran apoyo en el desarrollo del trabajo escrito de
esta tesis.
Gracias a mis compañeros de laboratorio, los cuales además de una relación
laboral, ahora son parte de mi vida: Gloria Adasme (Glory), Susana Velásquez, Verónica
Soto, Iván Valdés, Carolina Pérez, Dra. Patricia Olivares y su equipo de trabajo
(Maggy, Alejandra, Sra. Isabel, Sra. María, Naty, Mauricio Álvarez).
A quienes me ayudaron en el desarrollo de mi tesis y son un aporte en mi
desarrollo personal y profesional, personas como Loreto Egaña tiempo a tras y ahora
Rubén Avendaño.
Gracias a mi profesor patrocinante la Dra. Daniela. Seelenfreund, por su apoyo y
su constante guía, por su excelencia académica, representante de la docencia impartida
en nuestra Universidad y en ella expreso mi agradecimientos a todos los docentes que
son parte de mi formación profesional.
Gracias a mis amigos de Universidad, aquellos con los que compartimos
intereses comunes, la ciencia y buenos momentos en la facultad: Jocelyn Brito, Romina
Barría, Judith Tello, Patricia Santis, Fernando Sepúlveda, Alexis Peralta, Bendelin
Hernández. Gracias a mis amigos de la vida, aquellos especiales únicos Miguel A.
Muños M (gran y mejor amigo), Alejandro Collao y Cristian Martínez.
Gracias a mi Familia por su incondicional apoyo, por creer en mi y alentarme a
seguir mis sueños, entregando lo máximo de ellos en mi beneficio, no importando las
veces que he tropezado, sino destacando las veces que he obtenido pequeños logros y
quiero a través de esta tesis reconocer sus esfuerzo y entrega hacia a mi, gracias a mi
papá Rubén Silva, a mi mamá Cecilia Rubio y mis hermanos Pato y Emilio.
Y por ultimo, mi pilar, mi fe, quien es mi modelo a seguir…., a Cristo.

RESUMEN
La aparición de vibriosis en los cultivos de peces en Chile ha causado masivas mortalidades de
salmónidos cultivados en el sur de nuestro país. Debido a la gran trascendencia económica de esta
patología se han estudiado 77 aislados, los cuales han sido identificados a nivel de especie y caracterizados
genética, molecular y antigénicamente con el fin de desarrollar futuras medidas de prevención.
El análisis de la secuencia del gen rRNA 16S de todos los aislados, permitió identificarlos dentro de las
especies Listonella anguillarum (antes Vibrio anguillarum) y Vibrio ordalii. Trece aislados se
relacionaron genéticamente a la especie L. anguillarum y un grupo mayoritario de 61 aislados presentó
homología con la secuencia tipo de V. ordalii.
Desde el punto de vista genético, molecular y antigénico, las cepas chilenas de V. ordalii constituyen
un grupo homogéneo. Por el contrario, las cepas de L. anguillarum son heterogéneas en cada una de las
propiedades señaladas anteriormente.
De esta forma, mediante la técnica de electroforésis de campo pulsado (PFGE) fue posible observar al
menos 3 grupos genéticos dentro de las cepas chilenas de L. anguillarum. Sin embargo, al emplear
métodos basados en la Reacción en Cadena de la Polimerasa de elementos repetitivos (PCR-ERIC y PCR-
REP) fue posible obtener mayores diferencias, las que podrían ser empleadas para el seguimiento de un
aislado en particular con fines de llevar a cabo estudios epidemiológicos.
Además, se determinó la existencia de dos grupos serológicos mayoritarios dentro de los aislados
chilenos de L. anguillarun, los cuales no pertenecen a los serotipos O1 y O2 que comunmente están
asociados a la vibriosis clásica que afecta a especies salmónideas.
Los estudios de virulencia demostraron que las cepas de L. anguillarum y V. ordalii son capaces de
causar mortalidad en peces de la especie salmón del Atlántico (Salmo salar), reproduciendo fielmente los
signos característicos de ambas patologías.
Esta tesis constituye el primer reporte de L. anguillarum como agente causal de vibriosis en
salmónidos en Chile.

SUMMARY
Molecular and serological characterization of vibriosis causing agents in salmonids in the south of
Chile
The outbreaks of vibriosis in Chilean fish farms have caused massive mortalities of reared salmonids
in our country. Due to the great economical importance of this pathology we have studied 77 isolates,
which have been identified to the level of species and also have been characterized genetically,
molecularly and antigenically with the purpose of developing future control measures.
The analysis of the 16S rRNA gene of the total number of isolates, allowed us to identify them as
belonging to the species Listonella anguillarum (formerly Vibrio anguillarum) and Vibrio ordalii. In this
way, 13 isolates were genetically classified as L. anguillarum while a major group of 61 isolates showed
homology with V. ordalii.
The Chilean strains of V. ordalii constitute a homogeneous group according to genetic, antigenic and
molecular criteria. On the contrary, the isolates of L. anguillarum are heterogeneous in each of the
properties indicated above.
When we analysed the Chilean isolates of L. anguillarum with the technique of pulse field gel
electrophoresis (PFGE), three genetic groups could be identified. Furthermore, more differences were
observed when PCR-based techniques such as PCR-ERIC and PCR-REP were used. These could be used
for the follow-up of isolates in epidemiological studies.
Also, we have described the existence of two major serological groups among Chilean isolates of L.
anguillarum, which are not related to any of those commonly found that causing infection in salmonid fish
elsewhere (serotypes O1 and O2).
On the other hand, virulence studies have shown that isolates of L. anguillarum and V. ordalii are
capable of causing mortality in fish of Atlantic salmon (Salmo salar). The characteristic signs of both
pathogens were observed.
This thesis constitutes the first report of L. anguillarum as being responsible of vibriosis of salmonids
in Chile.

i
INDICE GENERAL
AGRADECIMIENTOS
RESUMEN
SUMMARY
INDICE GENERAL ……………………………………………… i
INDICE DE FIGURAS ........................................................................ v
INDICE DE TABLAS ........................................................................ vi
ANEXOS ......................................................................... vi
ABREVIATURAS .......................................................................... vii
1. INTRODUCCIÓN ………………………………………………. 1
1.1. Generalidades ……………………………………………..... 1
1.2. Vibriosis en Chile ……………………………………………..... 2
1.3. La vibriosis ……………………………………………..... 3
1.3.1. Epizooetiología …………………………………………...….. 3
1.3.2. Signos clínicos ………………………………………………. 3
1.3.3. Histopatología ………………………………………………. 4
1.4. Reservorio y transmisión …………………………………….... 4
1.5. Listonella anguillarum y Vibrio ordalii: ……………………….......... 5
1.5.1. Posición taxonómica …………………………………….... 5
5.1.5.2. Caracterización bioquímica ……………………………………… 6
1.5.3. Caracterización serológica …………………………….………... 7

ii
1.5.4. Caracterización genética ……………………………………… 9
1.6. Factores de virulencia ……………………………………… 10
1.7 Tratamiento y prevención ………………………………...……. 11
2. HIPÓTESIS ………………………………….....…..…….. 12
3. OBJETIVO GENERAL ……………………...…………....…. 12
3.1 OBJETIVOS ESPECIFICOS ………………………………............ 12
4. MATERIALES ………………………………………..…….. 13
4.1. Aislados incluidos en este estudio. ……………………………. 13
4.2. Animales experimentales: …………………………………..…. 13
4.3. Enzimas ……………………………………………………… 13
4.4. Anticuerpos. ……………………………………………………… 14
4.5. Material fungible …………………………………………….... 14
4.5.1. Reactivos. ………………………………………………………. 14
5. METODOLOGÍA ……………………………………………… 15
5.1. Procesamiento de los aislados ……………………………………... 15
5.1.1. Muestreo ……………………………………………………..... 15
5.1.2. Medios y condiciones de cultivos ………………………….…. 15
5.1.3. Ensayos bioquímicos ……………………………………………... 15
5.1.4. Extracciones ……………………………………………………… 16
5.1.4.1. Extracción de ADN ……………………………………………… 16
5.1.4.2. Extracción de proteínas totales y membrana externa …………… 16
5.1.4.3. Extracción de lipopolisacáridos (LPS) ………………………... 17
5.1.4.4. Extracción de antígeno “O” de bacterias …..…………………. 18
5.2. Identificación Molecular …………………………………… 18
5.2.1. Análisis filogenético basado en el gen ARNr 16S …………. 18

iii
5.2.1.1 Amplificación del gen ARNr 16S mediante partidores universales .. 18
5.2.1.2. Análisis de las secuencias …………………………………….... 19
5.2.2. Amplificación de la región intergénica (ITS) entre el 16S – 23S ARNr. 19
5.3. Caracterización Molecular ……………………………………… 20
5.3.1. Análisis de los aislados mediante electroforesis de campo pulsado…... 20
5.3.2. Análisis de restricción de los espaciadores intergénicos …………….. 21
5.3.3. Análisis de secuencias intergénicas repetitivas (PCR–ERIC)................ 22
5.3.4. Análisis de elementos extragénicos repetitivos (PCR-REP)…………. 22
5.3.5. Electroforesis en geles de agarosa …………………………….... 22
5.4. Caracterización serológica ………………………………………. 24
5.4.1.1. Obtención de sueros ……………………………………………….. 24
5.4.1.1. Formulación del antígeno ……………………………………… 24
5.4.1.2. Inmunización de conejos ……………………………………… 24
5.4.1.3. Inmunización de peces ……………………………………… 24
5.4.1.4. Absorción de sueros ……………………………………… 25
5.4.2. Titulación de los sueros por ELISA ……………………………… 25
5.4.3. Ensayos de Aglutinación ……………………………………… 26
5.4.4. Ensayos de dot-blot ………………………………………………. 26
5.4.5. Revelado para dot-blot y western-blot ……………………………... 27
5.4.6. Ensayo de Western-blot ………………………………………. 27
5.5. Estudios de virulencia ………………………………………. 28
6. RESULTADOS ……………………………………………….. 29
6.1. Identificación de los aislados de vibriosis ……………………… 29
6.1.1. Análisis filogenético del gen ARNr 16S ……………………. 29

iv
6.2. Caracterización genética ……………………………………... 32
6.2.1. Amplificación de ITS entre el gen ARNr 16S y el 23S …………….. 32
6.2.2. RFLP-ITS ………………………………………………………. 33
6.2.3. Electroforesis de campo pulsado (PFGE) …………………….. 33
6.2.4. Análisis de elementos extragénicos repetitivos (PCR-REP)................. 36
6.2.5. Análisis de secuencias intergénicas repetitivas (PCR-ERIC)………… 37
6.3. Caracterización molecular de componentes estructurales …………… 38
6.3.1. Proteínas totales y externas ………………………………………. 38
6.3.2. Lipopolisacáridos (LPS) …………………………………………… 40
6.3. Estudios serológicos de agentes causales de vibriosis ……………. 40
6.3.1. Obtención de sueros policlonales en conejos y su titulación por ELISA 41
6.3.2. Ensayo de aglutinación en porta-objeto …………………….. 42
6.3.3. Ensayos de Dot-blot ……………………………………… 43
6.3.4. Ensayos de Western-blot ……………………………………… 46
6.4. Estudios de virulencia ……………………………………… 51
7. DISCUSIÓN ……………………………………………… 52
7.1. Identificación …………………………………………..….. 52
7.2 Caracterización genética ……………………………………... 53
7.3. Caracterización de los componentes de la envoltura celular …… 56
7.4. Caracterización antigénica ……………………………………... 56
8. CONCLUSIÓN ……………………………………………… 59
9. REFERENCIAS ……………………………………………… 601

v
INDICE DE FIGURAS
Figura 1. Colonias típicas de los aislados chilenos de vibriosis …………….. 7
Figura 2. Comparación de las secuencias del gen ARNr 16S de aislados chile-
nos y aquellos disponibles para especies del género Vibrio..……..….. 30
Figura 3. Árbol filogenético a partir del gen ARNr 16S de aislados chilenos
causales de vibriosis en el sur de Chile .……………………...…..…. 31
Figura 4. Análisis de los aislados mediante la técnica de PCR-ITS ……..….. 32
Figura 5. Patrón de PFGE y dendograma de aislados de vibriosis en Chile …. 34
Figura 6. Gel comparativo entre los patrones electroforéticos de PFGE de los
aislados de V. ordalii y L. anguillarum nacionales………………….. 35
Figura 7. Electroforesis de PCR-REP de los aislados de vibriosis estudiados.… 36
Figura 8. Perfil electroforético del producto de amplificación PCR-ERIC de
aislados de V. ordalii y L. anguillarum ……………………………..... 37
Figura 9 Electroforesis en SDS-PAGE de proteínas y dendograma para V.
ordalii……………………………………………………………………… 38
Figura 10. Electroforesis en SDS-PAGE de proteínas y dendograma para L.
anguillarum…………………………………………………………..…… 39
Figura 11. Perfiles electroforéticos de LPS de los aislados de vibiosis en Chile. 40
Figura 12. Gráfica de titulación por ELISA de sueros anti-L. anguillarum…… 42
Figura 13. Ensayo de dot-blot de los aislados chilenos de V. ordalii…………... 45
Figura 14. Ensayo de dot-blot de los aislados chilenos de L. anguillarum………. 45
Figura 15. Inmunoblot de proteínas de aislados chilenos de V. ordalii……. …... 47

vi
Figura 16. Inmunoblot de proteínas de aislados chilenos de L. anguillarum ……. 48
Figura 17. Inmunoblot de lipopolisacáridos de aislados chilenos de V. ordalii…. 49
Figura 18. Inmunoblot de lipopolisacáridos de aislados chilenos de L. anguillarum 50
Figura 19. Mortalidad en los ensayos de virulencia ……………………………… 51
INDICE DE TABLAS
Tabla 1. Partidores utilizados en los estudios de caracterización genética……... 23
Tabla 2. Aislados representativos de L. anguillarum y V. ordalii seleccionados
para su caracterización serológica. ……………..……………….. 41
Tabla 3. Títulos obtenidos a partir de conejos inmunizados con los aislados
seleccionados de V. ordalii y L. anguillarum …………………….. 41
Tabla 4. Ensayo de aglutinación en portaobjeto usando los antisueros de L.
anguillarum y V. ordalii para los aislados chilenos ..………………… 43
ANEXOS
Anexo 1: Principales características fenotípicas, fisiológicas y bioquímicas de las
especies L. anguillarum y V. ordalii ……………………………..… 68
Anexo 2: Aislados bacterianos estudiados en la presente tesis, los cuales fueron asociados
a mortalidades de salmónidos entre octubre 2004 y agosto2005..…. 69

vii
ABREVIATURAS
-ME -mercaptoetanol
ADN ácido desoxirribonucléico
Ag O Antígeno O
ARNr: ARN (ácido ribonucleico) ribosomal
BCIP 5-bromo-4-cloro-3-indol fosfato p-toloudin
BrEt bromuro de etidio
BSA seroalbúmina de bovino
CBB Coomassie Brilliant Blue
DMSO dimetilsulfóxido
DTT ditiotreitol
EDTA ácido etilen-diamino-tetra-acético
ELISA (Enzyme Linked Inmunoabsorbent Assay ) ensayo inmunoenzimático
ERIC secuencias consenso repetitivas intragénicas de enterobacterias.
h hora / horas
HRP peroxidasa de rábano
IgG inmunoglobulina tipo G
ITS (Internal Transcribed Spacer) espaciador entre genes ARNr 16S y 23S
kDa kilo Dalton
LPS lipopolisacárido
mA miliamper
NBT azul de nitrotetrazolium
NC nitrocelulosa
nM nanomolar
OPD o-fenilendiamina
PAGE electroforesis en Geles de Poliacrilamida
pb pares de bases
PBS tampón fosfato salino

viii
PCR: (Polymerase Chain Reaction) Reacción en cadena de la polimerasa.
PFGE electroforesis de campo pulsado
PM peso molecular
PMSF fenil-metil-sulfonil-fluoruro
PSA persulfato de amonio
REP secuencias palindrómicas repetitivas extragénicas
RFLP polimorfismo del largo de los fragmentos de restricción
RNA ácido ribonucleico
rpm revoluciones por minuto
SDS dodecil sulfato de sodio
SRS síndrome rickettsial del salmón.
TA temperatura ambiente
TAE tampón tris, acetato, EDTA
TEMED N, N, N’, N’-tetrametil etilen diamina.
TCBS agar tiosulfato-citrato-sacarosa con sales biliares
Tris-HCl clorhidrato de Tris (hidroximetil) Aminometano
TSA medio de cultivo Agar tripticasa de soya
U unidades
V.p. Vibrio parahemolítico
V volts

1
1. INTRODUCCIÓN
1.1. Generalidades
En la última década, en Chile se ha expandido rápidamente el cultivo de diversas
especies de salmónidos, particularmente salmón del Atlántico (Salmo salar), salmón del Pacífico
(Oncorhynchus kisutch) y trucha arco iris (Oncorhynchus mykiss), siendo actualmente el
segundo productor del mundo después de Noruega (Instituto Tecnológico del Salmón, 2000).
Así, la producción mundial de salmónidos y trucha en el año 2005 fue de 1.586.000 toneladas,
de la cuales Noruega aportó 602.000 toneladas, correspondiendo al 38,0% seguido por Chile
con volumenes sercanos de producción que aportaron al país alredeor de US$ 1.721 millones
anuales, esto corresponde al 37,9 % de la producción mundial (Salmón Chile, 2006).
El explosivo crecimiento de la producción de salmón, debido principalmente a la
intensificación del cultivo de salmónidos, ha originado un aumento en la aparición de patologías,
ya sea causadas por microorganismos autóctonos de los sistemas de cultivo o emergentes (Hill,
2005; Instituto Tecnológico del Salmón, 2000) que pueden ser consideradas estrictamente como
patógenos obligados. La gran mayoría de los microorganismos son patógenos oportunistas que
constituyen parte de la microbiota del agua. Sin embargo, son los sistemas de cultivo de peces
donde se dan las condiciones más adecuadas para que estas bacterias oportunistas puedan
colonizar al pez y causar efectos adversos.
De esta forma, la actividad acuícola somete al medio ambiente a drásticos cambios entre
la microbiota bacteriana y sus relaciones con los peces en cultivo. Así por ejemplo, los sistemas
intensivos con alta biomasa, suelen ser una de las causas más habituales de infección bacteriana
debido principalmente a las variaciones en la concentración de gases disueltos, en el incremento
de la cantidad de amonio y en el aumento de desechos orgánicos (alimento y fisiológico). Por
ende, cada uno de estos factores o su conjunto pueden causar un incremento en la susceptibilidad
de los peces frente a estos patógenos (Schiewe y Harrell, 1983).
Los microorganismos patógenos más comunes en la industria acuícola mundial, tanto de
moluscos, crustáceos y peces son aquellos pertenecientes al género Vibrio, particularmente
Listonella anguillarum (antes V. anguillarum), V. ordalii, V. salmonicida, V. vulnificus biotipo 2

2
y V. harveyi. Estas bacterias generalmente son constituyentes naturales del hábitat acuático y
causan infecciones denominadas vibriosis. Así, un estudio de 25 brotes de enfermedades que
afectaron a peces en cultivo, identificó que el 69% de los patógenos bacterianos correspondieron
al género Vibrio (Zorrilla y cols., 2003), demostrando que especies de este género están
comúnmente asociadas a patologías presentes en la industria salmonera.
La trascendencia económica de la aparición de este grupo bacteriano en los cultivos, ha
incrementado el interés por estudiar estos microorganismos centrándose fundamentalmente en su
caracterización taxonómica, bioquímica, fisiológica y molecular, así como en el estudio de los
factores de virulencia. El conocimiento de estos aspectos ayuda a desarrollar medidas que
permitan diagnosticar rápidamente y contrarrestar la proliferación de estas cepas, y por lo tanto a
la prevención y control de la vibriosis.
1.2. Vibriosis en Chile
La primera descripción de vibriosis en Chile fue hecha en 1995 por Riquelme y cols.,
quienes aislaron una variante de L. anguillarum a partir de mortalidades masivas ocurridas en
criaderos de ostión del norte (Argopecten purpuratus), un molusco bivalvo cultivado en las
regiones II, III y IV de nuestro país.
Sin embargo, a partir del año 2003 se informan los primeros casos de vibriosis ocurridos
en peces cultivados, particularmente salmón del Atlántico y posteriormente en salmón del
Pacífico. Precisamente, a partir de algunos de estos casos Colquhoun y cols., (2004) lograron
aislar el agente causal de estas enfermedades, identificándolo como Vibrio ordalii. Hasta
entonces, esta especie sólo había sido aislada en las costas de Norte América, Japón, Australia y
Nueva Zelanda, por lo que ahora se sabe que V. ordalii ha extendido su distribución a áreas del
suroeste del Pacífico. Además, estos mismos investigadores confirmaron la patogenicidad de
algunos de los aislados chilenos mediante ensayos de virulencia en salmón del Atlántico
(Colquhoun y cols., 2004).
Es importante señalar que si bien los aislados chilenos mostraron una homología
superior al 99% con la secuencia del gen ARNr 16S de la cepa tipo de V. ordalii (ATCC
33509T), bioquímicamente las cepas aisladas a partir de salmónidos se comportan de modo muy
diferente. Además, los aislados chilenos no fueron serotipificados, aún cuando emplearon los

3
sueros anti -serotipo O2 de L. anguillarum debido a que todas las cepas de V. ordalii presentaron
autoaglutinación (Colquhoun y cols., 2004).
Por otro lado, durante el último año, L. anguillarum también ha sido identificada como
agente causal de vibriosis en salmónidos cultivados en nuestro país (Bohle y cols., 2006).
1.3. La vibriosis
1.3.1. Epizooetiología
La vibriosis es caracterizada por una compleja interacción entre el patógeno, el
hospedero, los factores ambientales y el manejo de las condiciones de cultivo. Entre los factores
ambientales que influyen en el desarrollo de la patología se encuentran: a) el incremento de la
temperatura del agua; b) exceso de material orgánico y c) cambios en la calidad del agua de
cultivo. En cuanto al manejo del sistema, una inadecuada alimentación (calidad y cantidad de
comida) y/o manipulación de los peces y particularmente sobrepoblación en los sistemas
acuícolas, aumentan considerablemente la incidencia de la vibriosis. Es importante denotar que
se requiere sólo un pequeño número de peces susceptibles a L. anguillarum o V. ordalii, para la
aparición de un brote de vibriosis, ya que un pez enfermo fácilmente contagia a uno sano,
iniciándose así la propagación de la enfermedad. Además, la erradicación total de la vibriosis en
un centro de cultivo es compleja debido a que una cepa puede persistir al menos dos años en un
lugar geográfico determinado (Pedersen y Larsen, 1998).
1.3.2. Signos clínicos
La vibriosis en peces está descrita como una septicemia hemorrágica y posee diferentes
manifestaciones dependiendo el hospedero. Esta enfermedad corresponde a un cuadro
septicémico, debido a la gran amplitud que abarca la infección y las lesiones que están presentes
en todos los órganos como piel, aletas, ojos, ano, músculo, intestino, bazo e hígado, entre otros.
Los síntomas característicos de vibriosis causados por L. anguillarum son lesiones necróticas
sobre las áreas ventral y lateral del pez, lesiones oscuras e inflamadas en la piel que ulceran y
liberan exudados sanguinolentos, lesiones corneales opacas seguido por una ulceración del
contenido orbital, intestino distendido y recto tapado por un líquido viscoso (Actis y cols., 1999).
Internamente, se puede observar hemorragia y necrosis del tejido muscular y órganos como

4
bazo, hígado, riñón y sistema digestivo, los cuales en algunos casos sólo pueden mostrar
procesos inflamatorios (Actis y cols., 1999; Bassler y cols., 1994). En epizootias agudas, el curso
de la vibriosis es rápido, sin signos clínicos y se detecta sólo cuando el pez infectado muestra
zonas de inflamación en la piel y liberación de fluidos sanguinolentos antes de su muerte
(Valerie y cols., 1993).
En el caso de Vibrio ordalii, los signos de vibriosis son diferentes y particularmente
asociados a lesiones hemorrágicas en la aleta ventral y el área que rodea el ano, incluyendo el
poro anal. En algunos casos es posible detectar la presencia de abscesos con contenidos
hemorrágicos en la piel (Ransom y cols., 1984; Vera, 2005)
A escala macroscópica, internamente V. ordalii provoca palidez del hígado,
hepatomegalia, esplenomegalia, palidez de bazo o estado de mórula, renomegalia, enteritis,
hemorragia de la vejiga natatoria, hígado, grasa visceral y en ocasiones es factible observar
cavernas musculares y pericarditis (Ransom y cols., 1984).
1.3.3. Histopatología
La histopatología de peces enfermos con vibriosis es principalmente caracterizada por
septicemia hemorrágica. El número de leucocitos disminuye drásticamente, se encuentra una alta
concentración de bacterias en la sangre y existe una severa destrucción del tejido
hematopoyético, generando una anemia aguda. Al realizar frotis histológicos, se pueden observar
agregados de colonias bacterianas que pueden ocupar extensas áreas en los órganos afectados,
tales como piel, músculo y riñón, incluso en el corazón (Ransom y cols., 1984).
1.4. Reservorio y transmisión
El reservorio natural de L. anguillarum y V. ordalii es el agua de cultivo, aunque ambas
bacterias han sido aisladas de sedimentos y organismos marinos como crustáceos, moluscos,
Cnidarios (Cnidaria) que son especies de animales marinos relativamente simples, entre otros.
En cuanto al modo de transmisión y ruta de infección de L. anguillarum, el agua de
cultivo es fundamental. Se han descrito casos en que el patógeno puede migrar y ser adquirido
lejos de la fuente de infección (Toranzo y Barja, 1990). En el caso de V. ordalii, la ruta de

5
infección ocurre por cohabitación de peces enfermos con ejemplares sanos, existiendo dos vías
principales de ingreso, a través del recto e intestino posterior y a través de la piel cuando
presenta pérdida de su integridad, (Ransom y cols., 1984). Además, es importante señalar que la
vibriosis a veces se encuentra asociada a otras patologías, particularmente a las causadas por las
bacterias Pisciricketsia salmonis, Renibacterium salmoninarum y Streptococcus phocae.
1.5. Listonella anguillarum y Vibrio ordalii.
1.5.1. Posición taxonómica
Los vibrios son microorganismos acuáticos comunes que viven libremente, Gram-
negativos no esporulados rectos o curvos, entre 0,5 - 0,8 μm de longitud y son móviles en medio
líquido, ya que poseen un flagelo polar, aunque existen especies con más de un flagelo. Estas
bacterias son anaerobios facultativos (con metabolismo respiratorio y fermentativo), la mayoría
de ellos son oxidasa y catalasa positivos, fermentan glucosa sin producción de gas y son
sensibles a agentes vibrioestáticos O/129 (2,4 diamino 6,7-diisopropil pteridina fosfato). Las
especies del género Vibrio son quimioorganotróficos, por ello los iones de sodio estimulan su
crecimiento y en algunos casos son esenciales para su desarrollo. Estas bacterias se pueden
encontrar libres o formando biopelículas en asociación con corales, peces, moluscos, algas,
camarones y zooplancton, incluso dentro del contenido intestinal de estos organismos (Watnick
y Kolter, 2000; Donlan, 2002; Schembri y cols., 2002).
Listonella anguillarum más conocida por su sinónimo taxonómico Vibrio anguillarum,
es el agente causal de la vibriosis clásica, siendo considerada una de las infecciones de mayor
importancia dentro de las patologías presentes en animales de agua de mar. Esta bacteria afecta
alrededor de 50 especies de peces, y es más frecuente aislarla a partir de especies en cultivo que
de especies en estado salvaje (Toranzo y Barja, 1990). Listonella anguillarum se aisló por
primera vez de anguilas y denominada como Bacterium anguillarum por Canestrini en 1893. En
1909 Bergeman la renombra como Vibrio anguillarum (Powell y Loutit, 1990), permaneciendo
dentro de este género bacteriano hasta que MacDonell y Colwell (1985) la proponen como
Listonella anguillarum (Género IV dentro de la familia de las Vibrionaceae, corrección de
Bergeman 1909). Sin embargo, a la fecha, la mayoría de los patólogos y microbiólogos se

6
refieren a esta bacteria como un miembro del género Vibrio.
Vibrio ordalii es otro agente causal de vibriosis, que durante 10 años fue considerado
una bacteria relacionada a L. anguillarum, y descrito como V. anguillarum biotipo 2 (Schiewe y
cols., 1977). Sin embargo, el mismo grupo de investigadores en el año 1981, demostró la
existencia de diferencias entre las dos especies, basándose principalmente en la morfología de la
colonia, características bioquímicas y en la composición de la secuencia de ácido
desoxirribonucleico (ADN) mediante la técnica de hibridación ADN-ADN (Schiewe y cols.,
1981). Así, el nombre propuesto para la clasificación de esta especie fue Vibrio ordalii en honor
del investigador Erling Ordal por su contribución al estudio de bacterias patógenas de peces. El
primer aislado de V. ordalii depositado en la Colección Americana de Cultivos Tipos (ATCC) se
realizó a partir de salmón del Pacífico, depositado como ATCC 33509T (Schiewe y cols., 1981).
1.5.2. Caracterización bioquímica
Si bien V. ordalii y L. anguillarum son agentes causales de la misma patología, presentan
diferencias morfológicas y bioquímicas que son útiles para poder distinguirlos. Las principales
características bioquímicas que diferencian los aislados de L. anguillarum de cepas de V. ordalii
son la presencia de actividad β-galactosidasa, arginina dihidrolasa, utilización de citrato,
triptófano, indol, reducción de nitrato, Voges Proskauer positivo, degradación de gelatina y
fermentación de manitol, sorbitol, amigdalina y arabinosa. Además, L. anguillarum es un
microorganismo halófilo capaz de tolerar hasta un 5% de cloruro de sodio y temperaturas de
incubación entre 4 a 37ºC. En el anexo 1 se muestran las principales propiedades bioquímicas de
L. anguillarum y V. ordalii.
Por otro lado, ambos microorganismos pueden ser cultivados en medios generales
líquidos y sólidos como Zobell 2216, Luria-Bertani y tripticasa de soya suplementado con
cloruro de sodio en concentraciones no superiores a 2%. Es interesante destacar que sólo L.
anguillarum es capaz de crecer en el medio selectivo para especies del género Vibrio
thiosulphate-citrate-bile salts-sucrose (TCBS), lo cual es una propiedad que puede ser empleada
con fines de diagnóstico, ya que Vibrio ordalii no crece en este medio. Todos los medios
anteriormente descritos son adecuados para el aislamiento de ambos patógenos, tanto a partir de
muestras de agua como de organismos marinos. Por otro lado, para la realización de
antibiogramas, The Clinical and Laboratory Standards Institute (CLSI, antes NCCLS)

7
A BA B
recomienda para los dos microorganismos el medio de cultivo Müeller-Hinton suplementado con
cloruro de sodio.
En general, L. anguillarum, independiente del medio de cultivo empleado, crece en
forma óptima después de 24 a 48 h de incubación y a una temperatura de 20ºC. Las colonias son
circulares elevadas de color café-amarillo de 3 a 5 mm. Por el contrario, el crecimiento de V.
ordalii es mas tardío (entre 4 a 6 días) y las colonias típicas son blancas translúcidas, circulares,
convexas, alrededor de 1-2 mm (Figura 1).
Figura 1. Colonias típicas de los aislados chilenos de vibriosis
Se observa las colonias de Listonella anguillarum (A) y Vibrio ordalii (B) cultivadas en
placas de tripticasa de soya.
1.5.3. Caracterización serológica
Numerosos autores han demostrado que L. anguillarum posee heterogeneidad
serológica. Los primeros estudios se remontan a 1969, cuando Pacha y Kiehn describieron la
existencia de 3 serotipos dentro de la especie L. anguillarum. El serotipo 1, dado por los aislados

8
obtenidos a partir de salmónidos del Noreste; el serotipo 2 conformado por las cepas de L.
anguillarum procedentes de Europa y el serotipo O3 que agrupa a los aislados del Océano
Pacífico (Toranzo y Barja,1990). Sin embargo, con el desarrollo de las herramientas serológicas,
diversos investigadores demostraron la existencia de una mayor heterogeneidad dentro esta
especie, siendo descritos 3 serotipos mayoritarios en Noruega, 2 a 3 serotipos en los Estados
Unidos (citado por Grizes y Ollevier, 1995) y seis serotipos en Japón (Kitao y cols., 1983).
Ezura y cols., (1980) compararon algunos de estos serotipos, y propusieron que era
pertinente desarrollar un sistema inequívoco de serotipificación para este patógeno. Así,
Sørensen y Larsen (1986), estandarizaron la nomenclatura y presentaron un nuevo esquema de
tipificación, utilizando la técnica de aglutinación en portaobjeto, designando el prefijo O y luego
un número al serotipo. Esta nomenclatura se coordinó con los sistemas americano y japonés. En
resumen, el sistema propuesto por Sørensen y Larsen (1986) reconoce para L. anguillaum la
existencia de 10 serotipos mayoritarios (O1 a O10), siendo este sistema de serotipado el más
empleado a nivel mundial. Pedersen y cols., (1999) han descrito la existencia de 23 serotipos
para esta especie.
Dentro de la heterogeneidad antigénica presentada por L. anguillarum, existe
uniformidad en señalar que los serotipos O1, O2 y en menor grado el grupo O3 son los
responsables de gran parte de los brotes ocurridos en la industria acuícola (Toranzo y Barja,
1990). Los aislados pertenecientes a los restantes serotipos (O4 a O10) son considerados
ambientales y no patogénico. Cabe destacar que el serotipo O1 es antigénicamente homogéneo
(Bolinches y cols., 1990), mientras que los serotipos O2 y O3 presentan heterogeneidad
antigénica, siendo divididos en dos sub-grupos denominados O2α, O2β y O3A, O3B
respectivamente. El subgrupo serológico O2 está asociado a la infección de salmónidos y no
salmónidos, mientras que O2 ha sido aislado exclusivamente de especies no salmónidos como
rodaballo, bacalao, dorada, entre otros (Toranzo y cols., 1990). En cuanto al serotipo O3, sólo el
sub-grupo O3A ha sido causal de mortalidad en peces no salmónidos, mientras que el subgrupo
O3B como los restantes serotipos (O4 a O10), son considerados propios del agua y sedimento
marino (Austin y cols., 1993; Toranzo y cols., 2005). Por su parte, Vibrio ordalii es una especie
bacteriana homogénea y comparte componentes antigénicos con el serotipo O2 de L.
anguillarum (Schiewe y cols., 1981; Toranzo y cols., 1990).

9
1.5.4. Caracterización genética
Desde el punto de vista genético, las cepas de V. ordalii presentan una gran homología
entre sí, mostrando porcentajes de similitud que fluctúan entre los 83 a 100%, mientras que las
cepas de L. anguillarum sólo muestran una homología mayor al 70%. En cuanto al porcentaje de
similitud entre las dos especies bacterianas, éstos se encuentran entre el 53 a 69% (Schiewe y
cols., 1981).
Más recientemente, Skov y cols., (1995) y Tiainen y cols., (1995) usando técnicas de
ribotipado y electroforesis de campo pulsado (PFGE) han demostrado que existe una estrecha
relación entre las líneas clonales y los serotipos a los que pertenecen las cepas de L. anguillarum.
Paradójicamente, los estudios realizados con cepas de V. ordalii han mostrado la existencia de
clonalidad genética, independiente del huésped de aislamiento y origen geográfico del aislado
(Wards y Patel, 1991).
Las técnicas de PCR son útiles para diferenciar grupos de cepas no relacionadas
clonalmente, pero son menos discriminativas que la PFGE para diferenciar subtipos entre cepas
relacionadas. Existen diversas técnicas moleculares basadas en la PCR que son muy útiles como
método inicial de tipificación para estudiar la relación clonal entre aislados de una misma
especie. El análisis de secuencias palindrómicas repetitivas extragénicas (REP), que son
segmentos extragénicos de largos variables que se transcriben pero que no se traducen y las
secuencias consenso repetitivas intragénicas de enterobacterias (ERIC), que son secuencias
altamente conservadas dentro del genoma bacteriano y su distribución varía según la especie.
Estas secuencias son algunas de las estrategias que más se han utilizado en estudios
epidemiológicos de brotes infecciosos. Estas técnicas de análisis se caracterizan por su
simplicidad pues no requieren del uso de enzimas de restricción, ni técnicas electroforéticas
especiales. Además, son rápidas (menos de 24 h), de bajo costo y la interpretación de las bandas
suele ser sencilla. Los patrones de ADN que se obtienen con ERIC-PCR suelen ser menos
complejos que los generados mediante REP-PCR (Fernández-Cuenca, 2004).
La PCR-ITS se basa en la amplificación de las regiones espaciadoras que separan los
genes ribosomales 16S y 23S. Esta técnica es reproducible, aunque posee un poder de
discriminación bajo para algunos microorganismos. Para una mejor caracterización, anexo a la

10
amplificación, se puede incrementar el poder discriminativo con la utilización de enzimas de
restricción sobre el producto PCR (RFLP-ITS).
1.6. Factores de virulencia
Los lipopolisacáridos son biomoléculas que están presente en todas las bacterias Gram-
negativas, no sólo cumpliendo un rol estructural, sino que determinan muchas interacciones
bacteria-hospedero, juegan un rol importante en la patogénesis y en el desarrollo del cuadro
infeccioso por diversos microorganismos, incluyendo L. anguillarum. Asimismo, el LPS
bacteriano otorga a los miembros del género Vibrio, particularmente a los patogénicos, la
capacidad de resistir la actividad bactericida del suero normal o no inmune de los peces (Trust
y cols., 1981).
Existen proteasas en L. anguillarum que producen una mio-necrosis focal severa en los
peces, la cual lleva a una licuefacción muscular (Valerie y cols., 1993). Por otro lado, la
presencia de células de V. ordalii y la de un factor leucocítico lleva a una disminución de las
células del sistema inmune en la sangre de peces moribundos (Ransom y cols., 1984).
El sistema captador de hierro es el factor de virulencia más estudiado en L. anguillarum,
siendo empleado por la bacteria como mecanismo de defensa frente al hospedero y en el proceso
de infección de la misma. Esta molécula es liberada por la bacteria al medio externo y tiene
como función capturar el hierro desde las proteínas transportadoras (transferrinas y ferritinas) del
huésped (Valerie y cols., 1993), produciendo anemia hemolítica en los peces. Este sistema está
asociado a un receptor de membrana compuesto de cuatro proteínas (Actis y cols., 1986, Koster
y cols., 1991), las cuales participan en forma directa e indirecta en la captación del mineral. El
sistema de captación de hierro está codificado en un plasmidio de 65 Kb conocido como pJM1,
que permite a L. anguillarum crecer y sobrevivir en ambientes adversos (Crosa, 1980).
En L. anguillarum y V. ordalii, el flagelo bacteriano participa en la unión de la bacteria a
la superficie del pez. Además, esta estructura presenta motilidad quimiotáctica, la cual es capaz
de evitar la acción defensiva del mucus y romper los integumentos del pez. (Milton y cols.,
1996; Ormonde y cols., 2000).

11
1.7. Tratamiento y prevención
Aunque en la actualidad la vibriosis aún es controlada mediante la entrega de
quimioterápicos, el cuestionamiento de la comunidad mundial ha permitido disminuir
considerablemente las miles de toneladas usadas hasta hace unos años atrás. Esta situación es
debida a que los quimioterápicos son administrados como aditivos en el alimento o directamente
agregados al agua, por lo que se requieren grandes cantidades de fármaco para controlar la
enfermedad. Este tipo de tratamiento puede desencadenar la aparición de cepas resistentes y una
propagación de esta resistencia a los microorganismos del ambiente natural.
Una alternativa de prevención contra la vibriosis es la vacunación, existiendo en el
mercado mundial diversas vacunas comerciales tanto para L. anguillarum como V. ordalii. Estas
formulaciones están basadas sobre bacteria muerta o inactivada como también en componentes
de la membrana celular (proteínas o LPS). En nuestro país, las vacunas empleadas en la industria
acuícola son preparadas a partir de cepas madres no nativas, particularmente microorganismos
aislados en Noruega, por lo que se podría explicar que en algunos casos no se ha logrado los
efectos esperados.
Por ello, el uso de vacunas preparadas a partir de cepas autóctonas chilenas, bacterias
completas y/o selección de estructuras antigénicas, sigue siendo la mejor opción. Los estudios de
identificación, caracterización molecular y serológica son necesarios para la elección de la cepa
bacteriana y del serotipo a emplear, ya que la composición antigénica de la vacuna está adaptada
a la especie de pez y al área geográfica de uso.
Este trabajo se enmarca en un proyecto para el estudio de agentes causales de vibriosis
en salmónidos del sur de Chile a través de herramientas moleculares y serológicas, con el fin de
desarrollar una medida de prevención eficiente como el uso de autovacunas o vacunas para este
tipo de patógenos emergentes.
En la presente tesis se muestran los resultados obtenidos con cepas chilenas de V. ordalii
y L. anguillarum, correspondiendo estas últimas a la primera descripción de este patógeno en
cultivos de salmónidos de nuestro país.

12
2. HIPÓTESIS
La vibriosis en salmónidos cultivados en el sur de Chile es causada por cepas autóctonas
de microorganismos, con potencial patogénico, presentando características particulares.
3. OBJETIVO GENERAL
Identificar a nivel de especie el o los agente(s) causante(s) de vibriosis en la
salmonicultura chilena, con el fin de caracterizar genética, molecular y antigénicamente las
cepas aisladas a partir de mortalidades de peces cultivados, siendo el conocimiento obtenido la
base para el desarrollo de una futura vacuna contra esta enfermedad.
3.1 OBJETIVOS ESPECIFICOS
1. Identificar las especies bacterianas causantes de las mortalidades de salmónidos
mediante el estudio del gen ARNr 16S.
2. Caracterizar genéticamente estos aislados provenientes de casos clínicos usando las
técnicas de ITS, RFLP-ITS, PFGE, RAPD, ERIC-PCR y REP-PCR, con el fin de
evaluar su utilidad en estudios epidemiológicos futuros.
3. Caracterizar los componentes de la envoltura celular de estos microorganismos, tanto a
nivel proteico como de lipopolisacárido.
4. Caracterizar antigénicamente los aislados estudiados, con el fin de establecer relaciones
serológicas usando técnicas de ELISA, aglutinación en portaobjeto, dot-blot y western-
blot.
5. Evaluar la virulencia de algunas cepas en peces de la especie salmón del Atlántico
(Salmo salar) y así lograr establecer protocolos de desafíos para el futuro desarrollo de
formulación de vacunas.

13
4. MATERIALES
4.1. Aislados incluidos en este estudio.
Las muestras empleadas en este estudio proceden de casos clínicos de vibriosis ocurridos
en salmónidos al sur de Chile. Esta colección bacteriana incluye un total de 77 cepas aisladas por
médicos veterinarios especialistas en enfermedades en peces de las empresas Aquagestión de
Fundación Chile y Advance Diagnostic Laboratory (ADL), quienes efectuaron muestreos
dirigidos a ejemplares muertos o moribundos de salmón del Atlántico (Salmo salar), salmón del
Pacífico (Oncorhynchus kisutch) y trucha arco iris (Oncorhynchus mykiss) procedentes de
diversas áreas geográficas, principalmente la X Región del país (anexo 2).
Además, con fines comparativos se incluyeron las cepas de referencia de L. anguillarum
ATCC 43305 y ATCC 43306 (representantes del serotipo O1 y O2, respectivamente de la
especie) y Vibrio ordalii ATCC 33509, procedentes de la Colección Americana de Cultivos
Tipos (ATCC).
4.2. Animales experimentales:
a) Conejos: Con el fin de obtener sueros que permitieran realizar la caracterización serológica
de los aislados incluidos en esta tesis, se utilizaron 30 conejos hembras de raza Neozelandesa
(1,5 – 2 Kg).
b) Peces: Para la evaluación de la virulencia de los aislados y confirmación de los postulados de
Koch, los ensayos se realizaron en peces de la especie salmón del Atlántico (Salmo salar) con
pesos entre 6 a 8 grs.
4.3. Enzimas
Las endonucleasas empleadas en los estudios fueron las siguientes: BamHI, BstYI, DnaI,
EcoO109, EcoRI, HaeIII, HindIII, HinfI, HpaI, HapII, MluI, MnlI, PvuI, PvuII, RsaI, SalI,
Tth111I, XbaI y NotI, de New England Biolabs (Beverly, EEUU).

14
4.4. Anticuerpos.
Para la caracterización serológica se emplearon sueros anti-IgG de conejo preparados en
cabra y conjugados con peroxidasa de rábano (HRP) Bio-Rad ((Madrid,, España), así como un
anticuerpo monoclonal C11-HRP anti-IgM de salmón del Atlántico (salmo salar) de Aquatic
Diagnostic Ltda. (Stirling, Escocia). Además se utilizó sueros hechos en conejos antiserotipo O1
y O2 de L. anguillarum, donados gentilmente por la Dra. Alicia Toranzo, Universidad de
Santiago de Compostela, España.
4.5. Material fungible
4.5.1. Reactivos.
Los reactivos empleados en esta tesis fueron: Tween 20, acrilamida y bis-acrilamida
grado BM, antipapaína, azul brillante de Coomassie R-250, azul de bromofenol, BCIP, b-
mercaptoetanol, bromuro de etidio, CaCl2, DMSO, glicina, KCl, leupeptina, MgCl2, NaCl, NBT,
OPD, pepstatina, persulfato de amonio, PMSF, SDS, TEMED, los cuales fueron adquiridos a
Sigma Chemical Co. (St. Louis, MO, EEUU).
Acetona, ácido acético glacial, ácido cítrico, BSA, etanol, formaldehído, fosfato
disódico, glicerol, HCl, isopropanol, mertiolato, metanol, Tris-Base se adquirieron en Merck,
(Darmstadt, Alemania).
Los componentes empleados en la reacción de PCR se adquirieron a Bioron
(Ludwigshafen, Alemania), particularmente los desoxinucleótidotrifosfato (dNTPs) 10 mM y
MgCl2 50 mM, así como la Proteinasa K. Los partidores empleados en la reacción de PCR de los
ITS, es decir, G1 y L1, también la DNA polimerasa Taq 5 U/mL proceden de GIBCO Invitrogen
Corporation (Grand Island, NY, EEUU). El estándar de peso molecular de 100 pb es de Winkler
(Santiago de Chile).
Se utilizaron los siguientes sistemas comerciales API 20E de Biomérieux S.A. (Madrid,
España), y Micro BSA de Pierce Biotechnology (Francia) importado por Biosonda S.A
(Santiago, Chile), para el análisis filogenético Chelex®. 100 Resin, y el sistema comercial de
extracción InstGeneTM
Matrix de BioRad (Madrid, España) y Ready-To-GoTM
PCR beads de
Amersham Pharmacia Biotech ( New York, EEUU). Los aislados se conservaron en viales
comerciales Cryo-billes de AES Laboratoire (Rennes, Francia).

15
5. METODOLOGÍA
5.1. Procesamiento de los aislados
5.1.1. Muestreo
Se realizó un muestreo dirigido a cuadros clínicos de vibriosis o patologías asociadas a
esta enfermedad en diferentes planteles acuícolas, bajo la supervisión de médicos veterinarios en
terreno, pertenecientes a laboratorios especializados en salud de peces. Las muestras fueron
recolectadas en la IX, X, y XI regiones del país durante octubre 2004 y agosto del año 2005.
Los laboratorios responsables de los aislamientos fueron Aquagestión (Fundación Chile)
y Advance Diagnostic Laboratory (ADL). Con el fin de identificar y caracterizar las cepas
bacterianas, éstas fueron enviadas en agar de tripticasa de soya (TSA) al Laboratorio de
Microbiología de Veterquímica, en donde, una vez que se confirmó la pureza de los aislados, se
procedió a realizar estos estudios.
5.1.2. Medios y condiciones de cultivos
Durante el desarrollo de esta tesis, todos los microorganismos fueron rutinariamente
crecidos en agar o caldo Luria-Bertani e incubados a 20 1ºC durante 24 a 48 horas. La
conservación de los aislados se llevó a cabo empleando viales comerciales Cryo-billes a -80ºC.
5.1.3. Ensayos bioquímicos
La identificación bioquímica se realizó mediante el sistema comercial Api20E, un
sistema estandarizado que permite la identificación de Enterobacteriaceae y otros bacilos Gram
negativos no exigentes, como Vibrionaceae. El sistema estandarizado en microtubos que
contiene sustratos deshidratados, está compuesto de 21 pruebas bioquímicas. Se reconstruyó la
matriz por inoculación de una suspensión bacteriana en solución salina 0,85 % de NaCl. La
utilización del sustrato por la bacteria se traduce en un cambio de color de la matriz (positivo).
La interpretación de los resultados se llevó a cabo mediante la comparación con una tabla de
referencia, la cual entrega un código de 7 cifras correspondiente a una serie de identificación,
que es comparado contra una base de datos.

16
5.1.4. Extracciones
5.1.4.1. Extracción de ADN
Método 1: La extracción del ADN se realizó utilizando el método clásico fenol-cloroformo, a
partir de 2 mL de un cultivo fresco (5.1.2.), el cual se lavó 2 veces con PBS estéril y se
centrifugó a 13000 rpm durante 5 min, en microcentrifuga. El precipitado se resuspendió en 400
L de solución de lisis (SDS 1%, NaCl 0,15 M, EDTA 1,25 mM pH 8,0, Tris-HCl 0,1 M pH
7,5) en un tubo de centrífuga de 2 mL y la muestra se calentó a baño María (100ºC durante 10
min.). Una vez transcurrido este período, se agregó 20 L de proteinasa K (10 mg/mL) y se
incubó 30 min a 55ºC. Se agregó un volumen de solución fenol-cloroformo básico y se extrajo
agitando con vortex (máxima velocidad) durante 30 s. Se centrifugó a 10.000 x g durante 15 min
y se transfirió la fase acuosa (superior) a un tubo nuevo. La precipitación del ADN se realizó con
dos volúmenes de isopropanol y se mezcló suavemente, luego se centrifugó a 13.000 x g durante
15 min y se eliminó el sobrenadante, cuidando de no arrastrar el ADN. El precipitado se lavó 2
veces con etanol al 75%, se dejó secar a Tº ambiente y se resuspendió en agua calidad PCR.
Método 2, La extracción del ADN se realizó usando dos productos comerciales distintos. Una
resina quelante (Chelex®. 100) para el análisis filogenético del gen ARNr 16S y el sistema
comercial de extracción InstGeneTM
Matrix, siguiendo las recomendaciones del fabricante.
5.1.4.2. Extracción de proteínas totales y membrana externa
Las proteínas totales de los aislados fueron preparadas a partir de cultivos frescos (según
5.1.2.). Se centrifugó un volumen aproximado de 30 mL de cultivo a 9.600 rpm durante 5 min a
4°C (Centrifuga IEC (Internacional Equipment Company)) B-22M, rotor cat. 875 2-95). El
precipitado se lavó 2 veces mediante centrifugación con PBS estéril a 9.000 rpm durante 5 min a
4ºC. El precipitado se resuspendió en 3 mL de tampón Tris 10 mM pH 8,0 con 0,3% de NaCl y
la disgregacion celular se efectuó en dos etapas de 30 pulsos con 90% de amplitud y 0,6 ciclos
de sonicación, manteniendo las muestras en hielo para evitar el sobrecalentamiento (sonicador
Dr. Hielscher GMBH modelo 350601624). Si no se observó ruptura de las bacterias, se repitió el
ciclo de sonicación. Posteriormente, se centrifugó a 10.000 x g por 2 min en microcentrífuga
para separar los restos celulares.

17
El sobrenadante se separó en dos viales (uno para proteínas totales y otras proteínas de
membranas externas) y se centrifugó a 15.000 x g durante 60 min a 4°C.
El precipitado de un tubo, correspondiente a las proteínas totales, se resuspendió en agua
calidad PCR agregando una mezcla de inhibidores de proteasas constituida por leupeptina (2
mg/mL), antipaina (2 mg/mL), pepstatina (2 mg/mL) y PMSF (200 mM).
Uno de los 2 viales obtenidos con el procedimiento anterior (5.1.4.2), se utilizó para
extraer las proteínas de membranas externas. La fracción de membranas externas se obtuvo por
el método de Filip y cols. (1973), la cual utiliza el detergente no iónico, lauril sarcosinato de
sodio (sarkosyl) (N-lauril sarcosinato de sodio 3% en Tris-HCl 10 mM pH 8,0). Se resuspendió
el sedimento proteico en 200 µL de sarkosyl 3%, se dejó reaccionar a temperatura ambiente 30
min y posteriormente se centrifugó a 15.000 rpm durante 60 min en microcentrífuga.
Se resuspendió en agua calidad PCR agregando una mezcla de inhibidores de proteasas
constituida de igual manera que para la extracción de proteinas totales.
La cuantificación de proteínas se realizó con una alícuota del producto anteriormente
procesado, mediante el sistema comercial “Micro BCA Protein Assay” (PIERCE), siguiendo las
recomendaciones del fabricante. El resto de las muestras se almacenaron a –20 ºC para su uso
posterior.
Las proteínas fueron analizadas mediante electroforesis en condiciones desnaturantes o
SDS-PAGE (Laemmli, 1970) en mini-geles de poliacrilamida, los polipéptidos se visualizaron
mediante tinción con azul de Coomassie brillante R-250, y se estimó su tamaño por comparación
con el estándar de peso de amplio rango de 116 a 14,4 kDa.
5.1.4.3. Extracción de lipopolisacáridos (LPS)
Los LPS se purificaron por el método de Hitchcock y Brown (1983), a partir de un
cultivo fresco en caldo LB-NaCl 1,5% crecido durante la noche. El cultivo se precipitó y lavó 2
veces con PBS estéril y centrifugación 9.600 x g durante 10 min, obteniendo una suspensión
libre de medio. Se ajustó la suspensión a 0,2 de absorbancia, a una longitud de 600 nm ( A600 =
0.2) en PBS. Un volumen de 1,5 mL de la suspensión bacteriana ajustada, se centrifugó a 13.000
rpm durante 5 min (Micro High Speed refrigerated centrifuge VS-15000 cfn II, Vision). El

18
sedimento se resuspendió en 90 L de solución de lisis para LPS (SDS 2%, -mercaptoetanol
4%, glicerol 10% y azul de bromofenol 0,002% en Tris – HCl 1 M pH 6,8) y se calentó durante
10 min a 100ºC. Los restos celulares fueron separados por microcentrifugación (13.000 rpm
durante 5 min) y el sobrenadante se paso a un tubo nuevo, donde se agregó 10 L de una
solución de proteinasa K (20 mg/mL), se incubó durante 30 min a 60ºC y se almacenó a -20ºC
hasta su uso.
Los LPS fueron analizados a través de electroforesis en condiciones desnaturantes o
SDS-PAGE (Laemmli, 1970) en mini-geles de poliacrilamida y se visualizaron mediante tinción
con plata (Tsai y Franch, 1982) con el sistema comercial de tinción de plata Bio-Rad siguiendo
las instrucciones de los fabricantes.
5.1.4.4. Extracción de antígeno “O” de bacterias
El antígeno “O” (AgO) es una estructura termoestable, por lo tanto, este segmento del
LPS se obtuvo por calentamiento de 3 mL de una suspensión bacteriana de 109 ( A600 = 0,2) en
PBS estéril a baño María (100ºC x 60 min) y se centrifugó a 5.000 x g durante 5 min para
precipitar los restos celulares.
5.2. Identificación Molecular
5.2.1. Análisis filogenético basado en el gen ARNr 16S
5.2.1.1 Amplificación del gen ARNr 16S mediante partidores universales
Para este estudio el ADN fue extraído según 5.1.4.1. (Método 2). Se amplificó un
fragmento de aproximadamente 1416 pb, usando los partidores universales 1500R (Toyama y
cols., 1996) y SalI-fD1 (Weisburg y cols., 1991).
La reacción de PCR se realizó en un volumen final de 50 L, que contenía 5 L de ADN
purificado, 5 L de tampón 10X, 100 M de cada uno de los desoxinucleótidos trifosfato
(dNTPs), 25 M de cada partidor, 1,5 mM de MgCl2 y 2 U de ADN Polimerasa Taq. La
amplificación fue llevada a cabo en un termociclador 2700 de Applied Biosystems S, (Applied
Biosystems Foster, EEUU) y consistió en 35 ciclos de 1 minuto de desnaturación a 94ºC, 1

19
minuto de hibridación a 55ºC y 1 minuto de extensión a 72ºC. El producto amplificado fue
purificado en microcolumnas Omega Bio-tek de acuerdo a las recomendaciones del fabricante.
La secuenciación del producto amplificado y purificado, fue realizada por el
Departamento de Secuenciación de la Pontificia Universidad Católica de Chile.
5.2.1.2. Análisis de las secuencias
Todas las secuencias del gen ARNr 16S de los aislados causantes de vibriosis, fueron
analizados y comparados con secuencias disponibles en la base de datos RDP-II (Ribosomal
Database Project-II) y GenBank del Centro Nacional de Información de Biotecnología (NCBI).
Los alineamientos fueron realizados a través del programa ClustalX y el análisis filogenético
fue realizado con el programa PHYLIP (Saitou y Nei, 1987) disponible en http://
evolution.genetics.washington.edu/pub/phylip). La distancia evolutiva fue estimada por el uso de
DNADIST (método de 2 parámetros de Kimura) y las relaciones filogenéticas fueron
determinadas usando NEIGHBOR (método neighbour.joining). La reproducibilidad de los
patrones fue realizada con el SEQBOOT (método bootstrap; 100 réplicas) y el dendograma de
consenso fue generado con el software CONSENCE del programa PHYLIP (trabajo realizado
por el Dr. Jorge Fernández del Instituto de Salud Pública – ISP).
5.2.2. Amplificación de la región intergénica (ITS) entre el 16S – 23S ARNr
El ADN cromosomal de cada aislado bacteriano se extrajo según se describe en 5.1.4.1.
(Método 1 ó 2), la reacción de PCR se llevó a cabo en un volumen total de 30 L, con los
partidores universales G1 y L1 descritos por Jensen y cols. (1993) a una concentración de 0,25
M, en una mezcla de reacción conteniendo MgCl2 2 mM, nucleótidos 0,2 mM cada uno y 1,5 U
de DNA polimerasa Taq por reacción.
Se utilizó 1 L de muestra de ADN directo o diluciones por reacción. La amplificación
se realizó en un termociclador Applied Biosystems utilizando el siguiente programa: 94°C
durante 5 min; 30 ciclos de 94°C 1 min, 58°C 1 min, 72°C 1 min; 5 min de extensión a 72°C y
mantención de los productos a 4°C. La visualización del producto PCR se realizó en mini geles
de poliacrilamida teñidos al 8% con plata.

20
La tinción consistió en la fijación del gel en una mezcla de etanol-ácido acético, luego se
agregó una solución de plata al 0,18% p/v, y se reveló con una solución de NaOH al 3% y
formaldehído (0,5 mL/50 mL de solución). Una vez revelado se fijó el gel en una solución
etanol-ácido acético (método de Tsai y Franch, 1982) o bien por un método rápido en
microondas:
1. Se fijó el gel en una solución de etanol al 9,5 % y ácido acético al 0,5 %, durante 10
segundos, en microondas a máxima potencia (800 Watts).
2. Se cambió la solución por 50 mL de AgNO3 al 9 % durante 10 segundos en microondas a
máxima potencia.
3. El gel se lavó suavemente dos veces durante 10 segundos con agua destilada a temperatura
ambiente.
4. Se reveló con solución de NaOH al 3 % en agua destilada y 1 % de formaldehído durante 15
segundos, en microondas a máxima potencia, y luego con agitación a temperatura ambiente
hasta visualizar las bandas.
5. Se lavó una vez con agitación suave en agua destilada.
6. La tinción se fijó con la mezcla de etanol al 9,5 % y ácido acético al 0,5 %, preparada como
se describe en el punto 1.
5.3. Caracterización Molecular
5.3.1. Análisis de los aislados mediante electroforesis de campo pulsado (PFGE)
Se trabajó con 1 mL de bacterias en caldo de cultivo LB, con una cantidad equivalente a
1x108 células/mL por aislado, se centrifugó a 8.000 x g durante 4 min, el sedimento se
resuspendió en 150 L de solución 1 (Tris – HCl 100 mM, NaCl 1 M, EDTA 50 mM pH 8), se
agregó 150 L de agarosa de bajo punto de fusión (LMP) 1,6% a 45°C, sin generar burbujas,
dejando gelificar a temperatura ambiente (TA) por 20 min y luego se dejó a 4°C por un tiempo
similar. Se extrajo los bloques y se agregaron a un tubo de 1,5 mL estéril, incubando los bloques
durante 3 h a 37°C con 1 mL de solución de lisozima por tubo (lisozima 1 mg/mL en Tris – HCl
10 mM, NaCl 1 M, EDTA 100 mM pH 8, desoxicolato de sodio 0,2%, SDS (N-lauril-sarcosin de
sodio) 0,5% y RNAsa 2 g/mL) durante 4 horas a 37ºC. Posteriormente, se incubó con 1 asmL

21
de solución de proteinasa K (1 mg/mL proteinasa K en 50 mM EDTA pH 8, SDS 1%) por 4 h a
56°C, se lavó los bloques con solución de lavado por 2 h a TA con agitación suave y se lavó con
1mL por bloque de solución de PMSF (0,1 M en etanol, diluído 1/100 en solución 1) durante 1 h
a TA y luego toda la noche a 4°C. Se lavó 2 veces con solución 1 para eliminar los restos de
PMSF y se colocó los bloques en TE 1X (Tris – HCl 20 mM, EDTA 50 mM pH 8) a 4°C.
Posterior a esto, se ambientaron los bloques en 100 µL de tampón de digestión de NotI en tubos
de vidrio de fondo plano por 1 h a TA. Se digirió con 50 µL de tampón fresco que contenían 50
U de enzima y se incubó toda la noche a 37°C (o como lo indica el fabricante). Se eliminó la
solución anterior y se agregó 20 volúmenes de TE 1X por más de 2 h, posterior a ésto se agregó
100 µL de solución de detención (0,25% de azul de bromofenol en TE) por 15 min.
Se realizó el PFGE en un equipo Cheff (Bio-Rad) empleando 150 mL al 1 % de agarosa
para campo pulsado en TBE 0,5 X a 14°C y se equilibró a 50ºC antes de vaciar. La electroforesis
se realizó a 6 V/cm (200 V) con una gradiente de pulsos de i) 1-18 seg. durante 12 h., ii) 3-80
seg. durante 12 horas. Se tiñó con bromuro de etidio 0,5 g/mL en 150 mL de agua destilada
durante 30 min. La intensidad de la tinción se reguló por desteñido en 250 mL de agua destilada
con agitación suave.
5.3.2. Análisis de restricción de los espaciadores intergénicos 16S-23S ARNr (RFLP –ITS)
Con el fin de diferenciar los aislados con igual patrón de ITS, el producto de
amplificación por PCR se digirió con diferentes enzimas de restricción. Las enzimas utilizadas
fueron las siguientes:
Bam HI, Bst YI, Dna I, Eco O109, Eco RI, Hae III, Hind III, Hinf I, Hpa I, Hap II, Mlu I, Mnl I,
Pvu I, Pvu II, Rsa I, Sal I, Tth 111I, Xba I.
A 5 L de producto PCR-ITS, se agregó un volumen de 10 L de reacción enzimática
según lo recomendado por el fabricante. En la sección 5.2.2., se describe el protocolo de
amplificación y visualización de estos productos.

22
5.3.3. Análisis de secuencias intergénicas repetitivas de enterobacterias (PCR–ERIC)
La amplificación de los fragmentos ERIC-PCR se realizaron con el sistema comercial
Ready-To-GoTM
PCR beads. Los partidores utilizados fueron ERIC 1 y ERIC 2 (Tabla 1). En
tubos de microcentrífuga de 0,2 mL se agregaron las micro esferas, 22 µL de agua calidad PCR,
1 µl de cada partidor y 1 µL de ADN y se incubó en el siguiente programa: Desnaturación inicial
(95ºC durante 5 min); 35 ciclos (desnaturación a 92ºC durante 45 seg; alineamiento a 52ºC
durante 1 min y extensión a 70ºC durante 10 min) y una extensión final a 70ºC durante 20 min.
El control negativo consistió en la mezcla de reacción sin ADN. Los productos se visualizaron
en geles de agarosa.
5.3.4. Análisis de elementos extragénicos palindrómicos repetitivos (PCR-REP)
El mismo sistema comercial Ready-To-GoTM
PCR beads se utilizó para las
amplificaciones REP, utilizando los siguientes partidores REP 1D y REP 2D. La reacción de
PCR se llevó a cabo en 25 µL, en tubos de microcentrífuga de 0,2 mL se agregó las micro
esferas, 22 µL de agua calidad PCR, 1 µL de cada partidor y 1 µL de ADN y se incubó en el
siguiente programa: Desnaturación inicial (95ºC durante 7 min); 35 ciclos (desnaturación a 92ºC
durante 45 seg; alineamiento a 40ºC durante 1 min y extensión a 70ºC durante 8 min) y una
extensión final a 72ºC durante 15 min. El control negativo consistió en la mezcla de reacción sin
ADN. Los productos se visualizaron en geles de agarosa.
5.3.5. Electroforesis en geles de agarosa
Los productos obtenidos se separaron utilizando electroforesis horizontal (100 V) en geles de
agarosa al 1,5% en tampón TAE 1X (Tris 0,04 M, EDTA 0,1 mM, pH 8,0) y 0,06 µg/mL de
bromuro de etidio y se fotografiaron bajo luz UV.

23
TABLA1: Partidores utilizados en los estudios de caracterización genética
Análisis de secuencia del gen ARNr 16S
Partidor Secuencia 5’ – 3’ Tm Sitio de unión Tamaño del
amplificado Fuente
1500R GGTTACCTTGTTACGACTT 55ºC gen ARNr 16S 1500pb Toyama y
cols., (1996)
Belt4 CGGTCGACAGAGGTTTGATCCTGGCTCAG
55ºC gen ARNr 16S 1500pb Weisburg y cols., (1991)
Análisis de Región Intergénica 16S-23S
Partidor Secuencia 5’ – 3’ Tm Sitio de unión Tamaño del
amplificado Fuente
G1 Forward GAAGTCGTAACAAGG 58ºC Región intergénica
16S – 23S (ITS)
Entre
600-700 pb Jensen y cols., (1993)
L1 Reverse CAAGGCATCCACCGT 58ºC Región intergénica 16S – 23S (ITS)
Entre 600-700 pb
Jensen y cols., (1993)
Análisis de Secuencias Consenso Intergénicas Repetitivas de Enterobacterias (ERIC)
Partidor Secuencia 5’ – 3’ Tm Sitio de unión Tamaño del
amplificado Fuente
ERIC 1 Forward
ATGTAAGCTCCTGGGGATTCAC
52ºC Secuencia IRUs Vesalovic y cols., (1991)
ERIC 1 Reverse
AAGTAAGTGACTGGGGTGAGCG
52ºC Secuencia IRUs Vesalovic y cols., (1991)
Análisis de Elementos ExtragénicosPalindrómicos Repetitivos (REP)
Partidor Secuencia 5’ – 3’ Tm Sitio de unión Tamaño del
amplificado Fuente
REP 1D Forward NNNRCGYCGNCATCMGGC 40ºC
Regiones palindrómicas externas
Vesalovic y cols., (1991)
REP 2D Reverse RCGYCTTATCMGGCCTAC 40ºC
Regiones
palindrómicas externas
Vesalovic y cols., (1991)
Donde: M es A o C; R es A o G; Y es C o T y N es cualquier nucleótido.

24
5.4. Caracterización Serológica
5.4.2. Obtención de Sueros
5.4.1.1. Formulación del antígeno
El cultivo bacteriano fresco se lavó tres veces por centrifugación (10.000 x g durante 5
min) en tampón fosfato (PBS) pH 7,4 estéril. Se resuspendió en una solución al 0,7% de
formalina en PBS, luego se incubó durante 3 h a TA y a 4ºC durante toda la noche.
Posteriormente, se lavó 3 veces con PBS y se centrifugó a 10.000 x g por 15 min. El precipitado
bacteriano se resuspendió en una solución al 0,2% de formalina y se ajustó la concentración
bacteriana a 109 células por mL. Esta solución se utilizó para la inmunización.
Para controlar la inactivación de los microorganismos se evaluó la viabilidad de la
suspensión bacteriana. Una alícuota de la solución se sembró en placas de LB o TSA
suplementado con NaCl al 1,5% de y se observó la presencia o ausencia de crecimiento
bacteriano.
5.4.1.2. Inmunización de conejos
Los sueros fueron obtenidos según el protocolo descrito por Sorensen y Larsen (1986).
Se inyectaron conejos neozelandeses hembras de aproximadamente 1,5 kg, por vía intravenosa
(i.v.) en la vena del pabellón externo de la oreja cada 4 días. Las dosis fueron incrementadas al
doble en cada inoculación (0,2; 0,4; 0,8; 1,6; y 3,2 mL), una semana después de la última
inoculación se extrajo el suero. La sangre se dejó durante 3h a 37ºC para la formación del
coágulo y se dejó durante toda la noche a 4ºC. Posteriormente, se separó el suero y se almacenó
a –20 ºC hasta su uso. Los aislados utilizados como antígenos fueron L. anguillarum (PF2, PF4,
PF7, PF8 y PF10 ) y V. ordalii (PF181, PF186, Au2, Au3 y Au10). Los antígenos fueron
preparados según el protocolo 5.4.1.1.
5.4.1.3. Inmunización de Peces
140 peces formando 4 grupos de 35 (cada pez de aproximadamente 5 grs), se inocularon
con 0,2 mL vía intraperitonial (i.p.). Un grupo se inoculó con el aislado Au2 de V. ordalii y dos
grupos con los aislados PF2 y PF10 de L. anguillarum .El cuarto grupo correspondió al control
inoculado sólo con solución salina (PBS).

25
Se realizó un muestreo de 4 peces por grupo a distintos intervalos de tiempo (0, 13, 26,
42, 60, 84 y 120 días) siendo el día 0 antes de la inoculación (control preinmune). A estos peces
se les extrajo la sangre, la que se mantuvo a 16ºC por 2h y luego durante toda la noche a 4ºC
para la formación de coágulo. Posteriormente se separó los sueros y se almacenaron a -20ºC
hasta su uso.
5.4.1.4. Absorción de sueros
Con el fin de establecer la existencia o no de epítopes comunes entre los aislados, los
sueros obtenidos en conejos o peces, se absorbieron con el aislado homólogo (el mismo de la
inmunización) o con un aislado heterólogo (comparativo). Para las absorciones se mezcló un
volumen de suero (entre 70 a 100 L) con un volumen de antígeno “O”. Se incubó 1h a 37°C
(conejo) o 16ºC (peces), y se almacenó durante toda la noche a 4°C. Posteriormente, se
centrifugó a 4.000 x g durante 5 min, para precipitar el complejo antígeno – anticuerpo (Ag-Ac),
y se repitió el protocolo de absorción 2 veces más.
5.4.2. Titulación de los sueros por ELISA
Para estimar el título de los anticuerpos obtenidos por las inmunizaciones en peces o
conejos, se realizó los ensayos de ELISA en placas de 96 pocillos. Se activaron con 100 µl de
una suspensión bacteriana, con una DO600 0,1 a 0,2, durante toda la noche. Se lavó 3 veces con
solución Low-salt (Tris 20 mM, NaCl 0,38 M, mertiolato 0,01%, Tween20 0,05%) y
posteriormente, se bloqueó con una solución de PBS-BSA al 1% durante 3 h a 37ºC lavandose
posteriormente los pocillos 3 veces con solución Low-salt.
Se agregó 100 µL de PBS-BSA 1% a toda la placa y se agregaron 100 µL de suero a
titular en uno de los pocillos y se procedió a hacer diluciones seriadas en base 2 en los pocillos
siguientes. La placa se incubó por 1 hora a 37ºC, se lavó 3 veces con solución High-salt (Tris 20
mM, NaCl 0,5 M, mertiolato 0.01%, Tween 20 0,1%) y se agregó el segundo anticuerpo
conjugado a fosfatasa alcalina, que reconoce a las IgG de conejo o IgM de salmón, diluido
1:3000 en solución tampón de conjugado (Low salt-BSA 1%). Se incubó por 1h a 37ºC y se
lavó 3 veces con solución High-salt.

26
Las placas se revelaron agregando 100 µL/pocillo de solución de sustrato (OPD 0,4
mg/mL, ácido cítrico 0,024 M, fosfato disódico 0,051 M), por 10 min., y la reacción se detuvo
con 50 µl/pocillo de H2SO4 2M. Finalmente, las placas fueron leídas a 492 nm en lector de
ELISA Multiskan MS (Labsystems).
El criterio utilizado para determinar el título de los sueros por ELISA se definió como:
tres veces el valor promedio de la absorbancia de la lectura de fondo del suero control o no
inmune. El título del suero se obtuvo por interpolacion de este valor (absorbancia) en la abscisa
de la gráfica.
5.4.3. Ensayos de Aglutinación
Sobre un portaobjeto se agregó un volumen de suero policlonale no absorbidos en una
dilución 1/10. Luego, se agregó mezclando con igual volumen de antígeno “O” termoestable
(AgO). La visualización del ensayo se realizó a contraluz sobre una superficie obscura. La
aparición de una aglutinación rápida y clara fue considerada positiva (respecto a los controles).
Una reacción débil y tardía se consideró negativa.
Para descartar la unión inespecífica o auto-aglutinación se mezclaron los sueros y
antígenos por separado con igual volumen de PBS, en donde no se debía observar reación. Como
control negativo se utilizó suspensiones de antígeno “O” y sueros pre-inmunes, descartando de
esta forma la existencia de reacciones inespecíficas o de autoaglutinación. En este estudio se
utilizaron sueros policlonales anti-O1 y anti-O2 de L. anguillarum facilitados por la Dra.
Toranzo de la Universidad Santiago de Compostela, España.
5.4.4. Ensayos de dot-blot
Se utilizó el protocolo de Cipriano y cols. (1985), con algunas modificaciones. Se
humedecieron las membranas de nitrocelulosa (0,45 m) en TBS (Tris 20 mM, NaCl 500 mM,
pH 7,5) por 5 min. Se eliminó el tampón y se dejó secar a temperatura ambiente. Se colocó 1 L
de antígeno (AgO) sobre la membrana de nitrocelulosa (NC) formando un punto. Como control
negativo se utilizó PBS y como control positivo se utilizó la cepa homóloga al suero utilizado.
El revelado de la NC se realizó según el protocolo descrito en 5.4.5.

27
5.4.5. Revelado de la membrana de nitrocelulosa para dot-blot y western-blot
La membrana de nitrocelulosa de dot-blot o western-blot se colocó en solución de
bloqueo (TBS-gelatina 3%) por 45 min. Posteriormente, se realizaron 2 lavados con TBS-
Tween20 al 0,05% (v/v) por 3 min. cada uno. Se agregó los sueros policlonales de conejos y sus
respectivos sueros controles preinmunes, diluidos 1:1000 en solución de anticuerpo (TBS-
gelatina 1%). Se incubaron a 37ºC por 1h, con agitación suave. Las membranas de nitrocelulosa
se lavaron 5 veces con TBS-Tween 20 al 0,05%, luego se agregó el segundo anticuerpo anti-IgG
de conejo hecho en cabra, conjugado a fosfatasa alcalina (AP) o bien a peroxidasa (HRP),
diluido 1:3000 en solución de anticuerpo y se incubó 1 h a 37ºC, con agitación suave. Se lavó 4
veces con TBS-Tween 20 al 0,05% y el último lavado con TBS, para retirar el excedente de
detergente.
Para el conjugado unido a la enzima AP, se utiliza como sustrato NBT-BCIP, reacción
compuesta por 5-bromo-4-cloro-3-indolilfosfato toluidina 0,15 mg/mL de la sal (BCIP), disuelta
en 1mL de N,N- dimetilformamida, 0,3 mg/mL de nitroblue tetrazolium (NBT) en 700 L de
N,N- dimetilformamida y 300 L de agua. Ambas soluciones fueron mezcladas en 100 mL de
tampón carbonato 0,1 M, pH 9,8. La reacción se efectuó protegida de la luz, generando un color
violeta después de 5 a 15 min, y se detuvo agregando agua des-ionizada.
El revelado del anticuerpo unido a HRP se realizó con una solución de H2O2 0,015% y
de 4-cloro-1-naftol disuelto en TBS, que formó un color azulado después de 5 a 15 min. o hasta
ver coloración. El desarrollo de color se detuvo al colorearse el control negativo (background),
colocando las membranas en agua desionizada por 10 min. (Cipriano y cols., 1985).
5.4.6. Ensayo de Western-blot
Los ensayos se realizaron de acuerdo a Towbin y cols. (1979). Después de la separación
de los polipéptidos por electroforesis en condiciones desnaturantes o SDS-PAGE (LPS o
proteínas), las bandas fueron transferidas a una membrana de nitrocelulosa, por aplicación de un
campo eléctrico horizontal (60 V por 4 h o 350 mA por 1 h) en un tampón de transferencia
compuesto por Tris-HCl 25 mM pH 8,3; glicina 192 mM y 20% de metanol.

28
Una vez transferidas se procedió según 5.4.5. para el análisis de ambas técnicas de dot-
blot e inmunoblot (5.4.4. y 5.4.6.). Se utilizaron sueros absorbidos y no absorbidos (de cepas
homólogas y heterólogas).
5.5. Estudios de virulencia
La virulencia de los aislados de V. ordalii Au2, Au10, PF188, así como las cepas de L.
anguillarum PF2, PF7 y PF10 se realizó en ejemplares sanos de salmón del Atlántico (Salmo
salar) de 6 a 8 g de peso. Se realizó un control de lote previo consistente en análisis
microbiológicos a órganos específicos (riñón, hígado y bazo) y análisis por PCR de IPN y SRS.
. Los peces fueron distribuidos en una razón de 30 peces por estanque e inoculados vía
intraperitoneal (i.p.) con una concentración aproximada de 3,2 x 106 UFC por pez. Como control
se utilizó un grupo de peces inoculados con PBS. La experiencia tuvo una duración de 21 días,
durante los cuales la temperatura del agua fue de 16 1°C, salinidad de 15‰ y alimentación de
1% de la biomasa.
La mortalidad de los peces se registró dos veces al día. Éstos fueron analizados
microbiológicamente en el laboratorio, con el fin de determinar si la causa de muerte fue por la
bacteria inoculada. El reaislamiento se realizó a partir de los órganos internos riñón, hígado y
bazo, así como lesiones externas sobre placas de agar de Luria-Bertani. Además, se realizaron
exámenes microscópicos de branquias y aletas. Las colonias bacterianas se sometieron a la
caracterización morfológica y serológica para confirmar los aislados como V. ordalii y L.
anguillarum.

29
6. RESULTADOS
6.1. Identificación de los aislados de vibriosis
6.1.1. Análisis filogenético del gen ARNr 16S
Se analizaron 77 aislados bacterianos, los cuales habían sido identificados previamente
mediante pruebas bioquímicas (por los laboratorios de diagnósticos de peces), como Vibrio sp.,
Vibrio ordalii o Vibrio ordalii atípico. Estos aislados se fueron obtenidos entre octubre del 2004
y agosto del 2005, entre las regiones IX y XI del sur de Chile.
El análisis de la secuencia de aproximadamente 1490 pb del gen ARNr 16S de todos los
aislados comparados con las secuencias depositadas en la base de datos Genbank, permitió
identificar los aislados dentro de dos especies bacterianas del género Vibrio, específicamente
Listonella anguillarum (antes Vibrio anguillarum) y Vibrio ordalii (Figura 2). Además, al
comparar las secuencias de ambos grupos, fue posible observar diferencias en la composición de
bases ubicadas entre las 447 y 576 pb (Figura 2B). Así, estos aislados chilenos pueden ser
diferenciados en dos grupos filogenéticos, uno compuesto por 13 aislados que se relacionaron
genéticamente a la especie L. Anguillarum (acceso AY069971) y otro dado por un grupo
mayoritario compuesto por 61 aislados que presentan homología con la secuencia tipo de V.
ordalii (acceso AY628642).
Con los antecedentes obtenidos se realizó un dendograma (Figura 3), el que permitió
determinar la homología de los aislados bacterianos estudiados. De esta forma, las cepas de L.
anguillarum y V. ordalii aisladas a partir de salmónidos cultivados en Chile presentaron un 99%
de homología a la secuencia de su respectiva cepa tipo, mientras que al compararlos entre sí el
porcentaje de homología disminuyó a un 97%. Cabe destacar que tres de los aislados, los cuales
fueron codificados como PF18, PF86 y PF88, no correspondieron al género Vibrio y no fueron
incluidos en este estudio.
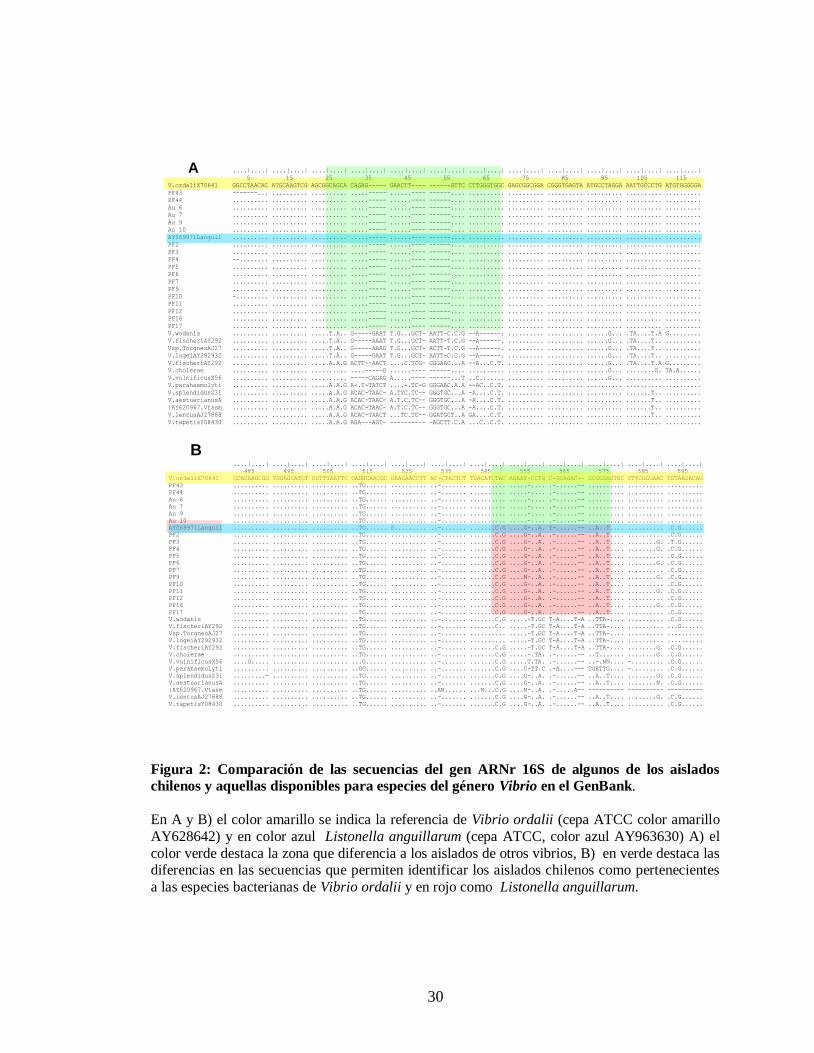
30
Figura 2: Comparación de las secuencias del gen ARNr 16S de algunos de los aislados
chilenos y aquellas disponibles para especies del género Vibrio en el GenBank.
En A y B) el color amarillo se indica la referencia de Vibrio ordalii (cepa ATCC color amarillo
AY628642) y en color azul Listonella anguillarum (cepa ATCC, color azul AY963630) A) el
color verde destaca la zona que diferencia a los aislados de otros vibrios, B) en verde destaca las diferencias en las secuencias que permiten identificar los aislados chilenos como pertenecientes
a las especies bacterianas de Vibrio ordalii y en rojo como Listonella anguillarum.
....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
5 15 25 35 45 55 65 75 85 95 105 115
V.ordaliX70641 GGCCTAACAC ATGCAAGTCG AGCGGCAGCA CAGAG----- GAACTT---- ------GTTC CTTGGGTGGC GAGCGGCGGA CGGGTGAGTA ATGCCTAGGA AATTGCCCTG ATGTGGGGGA
PF43 -------... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF44 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
Au 6 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
Au 7 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
Au 9 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
Au 10 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
AY069971Languil .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF2 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF3 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF4 --........ .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF5 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF6 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF7 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF9 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF10 -......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF11 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF12 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF16 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
PF17 .......... .......... .......... .....----- ......---- ------.... .......... .......... .......... .......... .......... ..........
V.wodanis .......... .......... .....T.A.. G-----GAAT T.G...GCT- AATT-C.C.G --A------. .......... .......... ......G... .TA....T.A G.........
V.fischeriAY292 .......... .......... .....T.A.. G-----AAAT T.G...GCT- AATT-T.C.G --A------. .......... .......... ......G... .TA....T.. ..........
Vsp.TorgnesAJ27 .......... .......... .....T.A.. G-----AAAG T.G...GCT- ACTT-T.C.G --A------. .......... .......... ......G... .TA....T.. ..........
V.logeiAY292932 .......... .......... .....T.A.. G-----GAAT T.G...GCT- AATT-C.C.G --A------. .......... .......... ......G... .TA....T.. ..........
V.fischeriAY292 .......... .......... .....A.A.G ACTT--AACT ....C.TCG- GGGAAC...A --A...C.T. .......... .......... ......G... .TA....T.A G.........
V.cholerae .......... .......... .......... ....-----G ......---- ------.... .......... .......... .......... ......G... ........G. TA.A......
V.vulnificusX56 .......... .......... .......... -----CAGAG A.....---- ------...T ..C....... .......... .......... ......G... .......... ..........
V.parahaemolyti .......... .......... .....A.A.G A-.T-TATCT ....-.TC-G GGGAAC.A.A --AC..C.T. .......... .......... .......... .......... ..........
V.splendidusZ31 .......... .......... .....A.A.G ACAC-TAAC- A.TYC.TC-- GGGTGC...A -A....C.T. .......... .......... .......... .......T.. ..........
V.aestuarianusA .......... .......... .....A.A.G ACAC-TAAC- A.T.C.TC-- GGGTGC...A -A....C.T. .......... .......... .......... .......T.. ..........
|AY620967.Vtasm .......... .......... .....A.A.G ACAC-TAAC- A.T.C.TC-- GGGTGC...A -A....C.T. .......... .......... .......... .......T.. ..........
V.lentusAJ27888 .......... .......... .....A.A.G ACAC-TAACT ...TC.TC-- GGATGCT..A GA....C.T. .......... .......... .......... .......T.. ..........
V.tapetisY08430 .......... .......... .....A.A.G AGA---AGT- ---------- -AGCTT.C.A ...C..C.T. .......... .......... .......... .......... ..........
A
....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
485 495 505 515 525 535 545 555 565 575 585 595
V.ordaliX70641 GCACAAGCGG TGGAGCATGT GGTTTAATTC GANNCAACGC GAAGAACCTT AC-CTACTCT TGACATCTAC AGAAT-CCTG C-GGAGAC-- GCGGGAGTGC CTTCGGGAAC TGTAAGACAG
PF43 .......... .......... .......... ..TG...... .......... ..-....... .......... .....-.... .-......-- .......... .......... ..........
PF44 .......... .......... .......... ..TG...... .......... ..-....... .......... .....-.... .-......-- .......... .......... ..........
Au 6 .......... .......... .......... ..TG...... .......... ..-....... .......... .....-.... .-......-- .......... .......... ..........
Au 7 .......... .......... .......... ..TG...... .......... ..-....... .......... .....-.... .-......-- .......... .......... ..........
Au 9 .......... .......... .......... ..TG...... .......... ..-....... .......... .....-.... .-......-- .......... .......... ..........
Au 10 .......... .......... .......... ..TG...... .......... ..-....... .......... .....-.... .-......-- .......... .......... ..........
AY069971Languil .......... .......... .......... ..TG...... S......... ..-....... .......C.G ....G-..A. Y-......-- ..A..T.... .......... .C.G......
PF2 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... .......... .C.G......
PF3 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... ........G. .T.G......
PF4 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... ........G. .C.G......
PF5 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... .......... .C.G......
PF6 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... ........G. .C.G......
PF7 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... .......... .C.G......
PF9 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....M-..A. .-......-- ..A..T.... ........G. .C.G......
PF10 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... .......... .C.G......
PF11 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... ........G. .C.G......
PF12 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... .......... .C.G......
PF16 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... ........G. .C.G......
PF17 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... .......... .C.G......
V.wodanis .......... .......... .......... ..TG...... .......... ..-....... .......C.G .....-T.GC T-A....T-A ..TTA-.... .......... .C.G......
V.fischeriAY292 .......... .......... .......... ..TG...... .......... ..-....... .......C.. .....-T.GC T-A....T-A ..TTA-.... .......... ...G......
Vsp.TorgnesAJ27 .......... .......... .......... ..TG...... .......... ..-....... .......... .....-T.GC T-A....T-A ..TTA-.... .......... ..........
V.logeiAY292932 .......... .......... .......... ..TG...... .......... ..-....... .......... .....-T.GC T-A....T-A ..TTA-.... .......... ..........
V.fischeriAY292 .......... .......... .......... ..TG...... .......... ..-....... .......C.G .....-T.GC T-A....T-A ..TTA-.... ........G. .C.G......
V.cholerae .......... .......... .......... ..TG...... .......... ..-....... .......C.G .....-.TA. .-......-- ..T....... ........G. .C.G......
V.vulnificusX56 ....G..... .......... .......... ...G...... .......... ..-....... .......C.G .....T.TA. .-......-- ..-.NN.... -......... .C.G......
V.parahaemolyti .......... .......... .......... ..GG...... .......... ..-....... .......C.G ....C-TT.C .-A....--- TGATTG.... -......... .C.G......
V.splendidusZ31 .........- .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... ........G. .C.G......
V.aestuarianusA .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... ........N. .C.G......
|AY620967.Vtasm .......... .......... .......... ..TG...... .......... ..AN...... ...N...C.G ....N-..A. .-.....A-- ---------- ---------- ----------
V.lentusAJ27888 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... ........G. .C.G......
V.tapetisY08430 .......... .......... .......... ..TG...... .......... ..-....... .......C.G ....G-..A. .-......-- ..A..T.... .......... .C.G......
B

31
PF84PF95PF94PF93PF92PF91PF90PF85PF89PF96PF98PF97
0.01
V.choleraeV.fischeriAY292942
V.wodanisVsp.TorgnesAJ278426
V.fischeriAY292949V.logeiAY292932
V.tapetisY08430V.rotiferiAJ440007.
V.parahaemolyticusV.equatorialisAJ440008
V.gallicusAJ440009V.vulnificusAY264936
V.vulnificusX56582.1V.splendidusAY046955V.tasmaniensisAJ514912
V.lentusAJ278880V.lentusAJ278881
V.lentusAJ536589V.pomeroyiAJ491290
V.aestuarianusAF172840V.splendidusAY129277
V.splendidusZ31657PF11PF3PF12
PF5AY662305.1 L anguillarum strai
PF4PF16
AY035897.1 L
anguillarumPF9PF6PF10PF7
PF2AY963630.1 L
anguillarumPF17PF 67PF 68
PF 48AY628645.1 Vibrio ordalii str
AY628642.1 Vibrio ordalii stAY628646.1 Vibrio ordalii sPF43PF44PF49PF50PF52PF53PF54PF55PF57PF70PF71PF72PF73PF81PF80PF79PF78PF77PF76PF74PF75PF83PF84PF95PF94PF93PF92PF91PF90PF85PF89PF96PF98PF97
PF84PF95PF94PF93PF92PF91PF90PF85PF89PF96PF98PF97
0.01
V.choleraeV.fischeriAY292942
V.wodanisVsp.TorgnesAJ278426
V.fischeriAY292949V.logeiAY292932
V.tapetisY08430V.rotiferiAJ440007.
V.parahaemolyticusV.equatorialisAJ440008
V.gallicusAJ440009V.vulnificusAY264936
V.vulnificusX56582.1V.splendidusAY046955V.tasmaniensisAJ514912
V.lentusAJ278880V.lentusAJ278881
V.lentusAJ536589V.pomeroyiAJ491290
V.aestuarianusAF172840V.splendidusAY129277
V.splendidusZ31657PF11PF3PF12
PF5AY662305.1 L anguillarum strai
PF4PF16
AY035897.1 L
anguillarumPF9PF6PF10PF7
PF2AY963630.1 L
anguillarumPF17PF 67PF 68
PF 48AY628645.1 Vibrio ordalii str
AY628642.1 Vibrio ordalii stAY628646.1 Vibrio ordalii sPF43PF44PF49PF50PF52PF53PF54PF55PF57PF70PF71PF72PF73PF81PF80PF79PF78PF77PF76PF74PF75PF83
0.01
V.choleraeV.fischeriAY292942
V.wodanisVsp.TorgnesAJ278426
V.fischeriAY292949V.logeiAY292932
V.tapetisY08430V.rotiferiAJ440007.
V.parahaemolyticusV.equatorialisAJ440008
V.gallicusAJ440009V.vulnificusAY264936
V.vulnificusX56582.1V.splendidusAY046955V.tasmaniensisAJ514912
V.lentusAJ278880V.lentusAJ278881
V.lentusAJ536589V.pomeroyiAJ491290
V.aestuarianusAF172840V.splendidusAY129277
V.splendidusZ31657PF11PF3PF12
PF5AY662305.1 L anguillarum strai
PF4PF16
AY035897.1 L
anguillarumPF9PF6PF10PF7
PF2AY963630.1 L
anguillarumPF17PF 67PF 68
PF 48AY628645.1 Vibrio ordalii str
AY628642.1 Vibrio ordalii stAY628646.1 Vibrio ordalii sPF43PF44PF49PF50PF52PF53PF54PF55PF57PF70PF71PF72PF73PF81PF80PF79PF78PF77PF76PF74PF75PF83
Figura 3: Árbol filogenético obtenido a partir de la comparación de secuencias del gen
ARNr 16S de los aislados chilenos causales de vibriosis en el sur de Chile.
Dendograma obtenido a partir del alineamiento de las secuencias nacionales con secuencias disponibles para especies del genero Vibrio. En color azul se destacan los aislados chilenos de L.
anguillarum, mientras que en color rojo las cepas pertenecientes a V. ordalii. La barra indica la
distancia filogenética equivalente al 0,01.

32
6.2. Caracterización genética
6.2.1. Amplificación de la región intergénica (PCR-ITS) entre los genes ARNr 16S y 23S
El estudio de los perfiles de ITS (Figura 4) mostró la existencia de dos patrones
electroforéticos, denotados como Patrón 1 y Patrón 2. Ambos patrones comparten 4 bandas, las
cuales tienen un peso molecular aproximado de 1000, 900, 650 y 580 pb, respectivamente. El
Patrón 1 muestra un producto de amplificación adicional de menor tamaño de aproximadamente
550 pb y se encuentra exclusivamente en los aislados de V. ordalii, mientras que el Patrón 2
agrupa a todos los aislados de L. anguillarum. Cabe señalar que sólo el aislado de V. ordalii
Au10 mostró un patrón distinto a P1 y P2 (ausencia de fragmento de 580 pb) (Figura 4B).
Figura 4: Análisis de los aislados mediante la técnica de PCR-ITS
Electroforesis en geles de acrilamida al 8% en SDS-PAGE y revelados con tinción con
plata. Muestras PF44–57, Au3, Au7 y Au10 corresponden a aislados de V. ordalii, muestras PF2, PF10 y PF17 corresponden a asilados de L. anguillarum. (A) Perfil de ITS característico de las
cepas de V. ordalii (P1, destacado en color azul) entre 1000 y 550pb y perfil de los aislados de L.
anguillarum (P2, señalados con color rojo) entre 1000 y 580pb. (B) detalle de la zona
característica entre 1000 y 400 pb. La flecha muestra la ausencia del fragmento de amplificación de 580 pb en el aislado de V. ordalii Au10, V.o: V. ordalii, L.a: L. anguillarum.
Patrón 2 Patrón 1 Patrón 1
A
B
3
P2
B
900
650
580
550
1000
P1
PF PF Au Au PF
2 52 2 10 10
A

33
6.2.2. Restricción del producto ITS (RFLP-ITS)
Al someter a restricción enzimática los productos de amplificación PCR-ITS con las
enzimas PvuII y HaeII, se observó perfiles de restricción idénticos dentro de la misma especie y
distinto al comparar entre especies, tanto con V. ordalii como con L. anguillarum (datos no
mostrados). Las enzimas de restricción Bst YI, Bam HI, Hpa I, Mlu I, Xcm I, Tth111 I, Eco0109
I, EcoR I, Hind III, Sal I y Xba I, no cortaron el producto de PCR- ITS (datos no mostrados).
6.2.3. Electroforesis de campo pulsado (PFGE)
Con el fin de realizar la caracterización del genoma completo de los dos grupos
bacterianos se realizó la electroforesis de campo pulsado. El genoma total se digirió usando la
enzima de restricción Not I, la cual permitió separar a los aislados en dos grupos genéticos. Un
grupo de aislados mostró un perfil homogéneo que presentó sólo diferencias menores, dadas por
un fragmento de amplificación adicional en el aislado Au10, así como la ausencia de una banda
en el aislado Au3. Los aislados que componen este patrón corresponden a las cepas de V. ordalii,
mostrando un grado de homología al menos de un 90% entre todos los aislados (Figura 5A).
Por el contrario, los aislados de L. anguillarum mostraron una gran heterogeneidad,
siendo posible observar la existencia de tres subgrupos genéticos con un porcentaje de similitud
de 60% (Figura 5B).
La Figura 6 muestra las diferencias entre los perfiles de campo pulsado de ambas
especies, observándose claramente las diferencias entre los patrones de restricción de las cepas
chilenas de V. ordalii y L. anguillarum.

34
Figura 5. Electroforesis de campo pulsado (PFGE) y dendograma de aislados causales de
vibriosis en Chile.
A) Perfil electroforético y dendograma de los aislados de V. ordalii chilenos. Con un círculo se
destaca las diferencias, presencia y ausencia de banda B) perfil electroforético y dendograma de
los aislados chilenos de L. anguillarum. Como control de restricción se incluyó la especie no relacionada Saccharomyces cerevisiae (Sac) y Vibrio parahaemoliticus (VpKX). La digestión
del genoma se realizó con la enzima Not I. Geles de agarosa al 1%, tinción con bromuro de
Etidio 0,5 g/mL.
A
B
PF
4
PF
17
PF
7
PF
12
PF
11
V
p
PF
8
PF
10
PF
9
PF
3
PF
2
PF
6
Vibrio ordali i
Listonella anguillarum
Sac
A
B
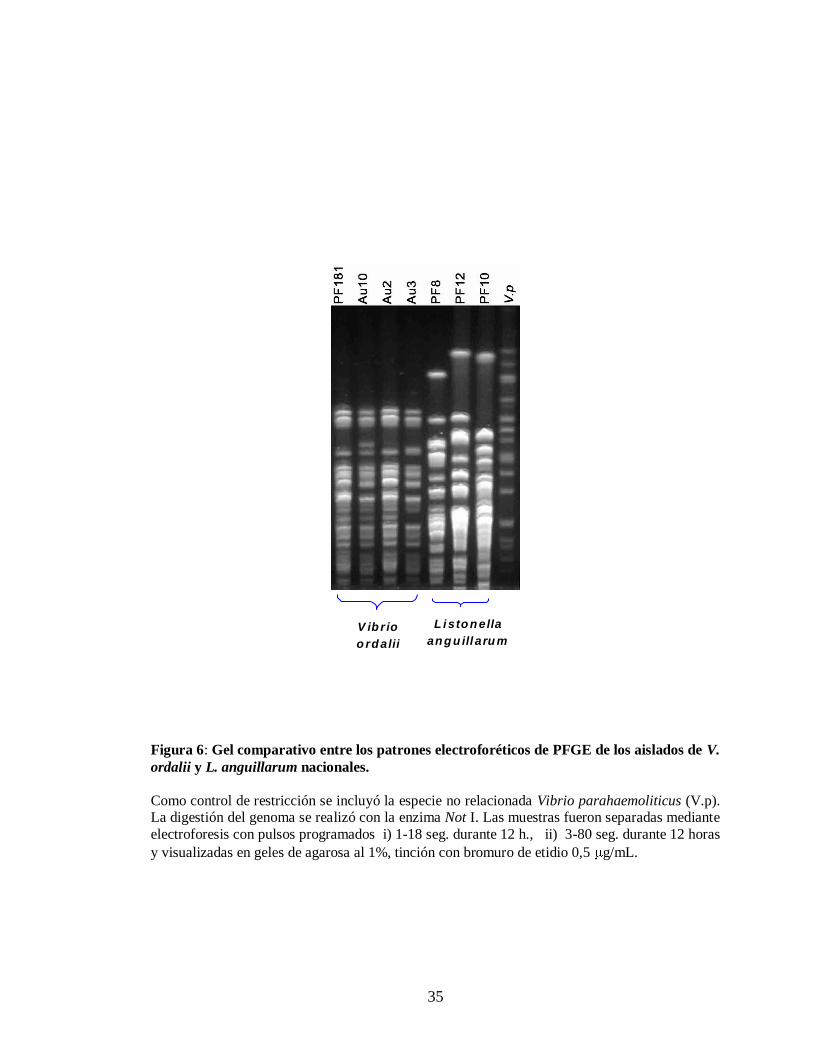
35
Figura 6: Gel comparativo entre los patrones electroforéticos de PFGE de los aislados de V.
ordalii y L. anguillarum nacionales.
Como control de restricción se incluyó la especie no relacionada Vibrio parahaemoliticus (V.p). La digestión del genoma se realizó con la enzima Not I. Las muestras fueron separadas mediante
electroforesis con pulsos programados i) 1-18 seg. durante 12 h., ii) 3-80 seg. durante 12 horas
y visualizadas en geles de agarosa al 1%, tinción con bromuro de etidio 0,5 g/mL.
PF
181
Au 1
0
Au 2
Au 3
PF
8
PF
12
PF
10
V.
p
L isto n ella
an g u ill aru m
V ib r io
o rd alii
C

36
6.2.4. Análisis de elementos extragénicos palindrómicos repetitivos (PCR-REP)
Con el fin de comparar resultados de genotipificación por otras técnicas alternativas a
PFGE, se realizó el análisis del genoma por amplificación por PCR de los elementos
palindrómicos repetitivos o PCR-REP, el cual se fundamenta en la amplificación de segmentos
extragénicos que se transcriben, pero que no se traducen y que son de un largo variables
permitiendo diferenciar en algunos casos a nivel de especie.
El análisis de 4 aislados representativos de V. ordalii (Au3, Au10, PF186 y PF187) y 3
cepas de L. anguillarum (PF2, PF7 y PF10), además de las cepas de referencia de V. ordalii
ATCC 33509 y L. anguillarum ATCC 43305 (serotipo O1) y ATCC 19264 (serotipo O2),
permitieron confirmar la homogeneidad genética de la especie bacteriana V. ordalii. Por otro
lado, los aislados de L. anguillarum nuevamente presentaron diversidad en los patrones de
amplificación, tanto entre ellos como con respecto a las dos cepas de referencias incluídas en el
estudio (Figura 7).
Figura 7: Electroforesis de PCR-REP de los aislados de vibriosis estudiados.
Las muestras Au10, PF186, PF187 y Au2 corresponden a aislados chilenos de V. ordalii, mientras que PF4, PF8, PF10 son las cepas de L. anguillarum; ATCC 33509 cepa tipo V. ordalii;
ATCC 19264 (serotipo O2), ATCC 43305 (serotipo O1) de L. anguillarum. Electroforesis en gel
de agarosa 1 % y tinción con bromuro de etidio. ST: estándar de peso molecular 100 pb.
1500
1000
800 700 600 500 400
200 300
Au PF PF Au ATCC PF . ATCC ATCC 10 186 187 2 33509 4 8 10 19264 43305
Vibrio ordalii Listonella anguillarum
ST ST PF PF PF 4 8 10

37
6.2.5. Análisis de secuencias intergénicas repetitivas de enterobacterias (PCR–ERIC)
Otra técnica de análisis alternativo es PCR-ERIC, que corresponde al análisis de las
secuencias ERIC o secuencias intergénicas repetitivas de enterobacterias, también llamadas
IRUs (unidades repetitivas intergénicas). Estas son secuencias altamente conservadas dentro del
genoma bacteriano y su distribución en el genoma varía según la especie.
Al realizar el análisis de los mismos ADN de las cepas descritas en el apartado anterior
usando la técnica de PCR-ERIC, nuevamente se observó homogeneidad en los patrones de los
aislados de V. ordalii y heterogeneidad en los patrones electroforéticos de las 5 cepas de L.
anguillarum analizadas (Figura 8).
Figura 8: Perfil electroforético del producto de amplificación PCR-ERIC de aislados de V.
ordalii y L. anguillarum.
Aislados de V. ordalii carriles: Au10, PF186, PF187 y Au2; aislados de L. anguillarum carriles:
PF4, PF7 y PF10. Además, se incluyeron las cepas de referencias de V. ordalii carril ATCC
33509 y L. anguillarum, carriles ATCC 19264 y ATCC 43305. ST: estándar de peso molecular 100 pb. Gel de agarosa 1 %, tinción con bromuro de etidio.
V. ordalii L. anguillarum
PF PF PF
2 7 10
Vibrio ordalii Listonella anguillarum

38
6.3. Caracterización molecular de componentes estructurales
6.3.1. Proteínas totales y externas
Con el fin de caracterizar los componentes de la envoltura celular de los aislados
estudiados se realizó un análisis de los patrones electroforéticos, tanto de proteínas totales
como de proteínas de membrana externa.
Vibrio ordalii
El análisis de ambos componentes proteicos permitió detectar un único patrón en todos
los aislados estudiados, independiente del huésped de aislamiento y origen geográfico. De esta
forma, todos los aislados comparten un número considerable de bandas entre los 25–70 kDa
(Figura 9). Además, utilizando el programa computacional GelCompar II se obtuvo un
porcentaje de similitud superior al 80% entre todos los aislados estudiados.
Figura 9. Electroforesis en SDS-PAGE de proteínas y dendograma para V. ordalii
Análisis de proteínas totales y externas de los aislados de V. ordalii en geles de SDS al 12%, tinción con azul de Coomassie (A) Perfiles electroforético y dendograma de proteínas totales
(B) perfil electroforético y dendograma obtenido de las proteínas externas de V. ordalii. PM: corresponde al estándar de peso molecular.
PF
185
PF
187
PF
181
Au2
PF
91
Au3
PF
181
PF
182
B
45
A
PF
185
PF
187
PF
18
Au
2
PF
91
Au
3
PF
181
PF
182
PM
PF
185
PF
187
Au
20
Au
2
PF
91
Au
3
PF
181
PF
182
Totales Externas
116
35
25
18,4
66,2
14,4
Au 2
PF1 8 5
PF 1 8 2
PF1 8 7
PF 9 1
100 90 80
PF1 8 8
A u 3
PF 1 8 1
85,4
75,3
92,5
81,3
87,5
92
96,6
P F 9 1
P F 1 88
P F 1 8 7
P F 1 8 2
2 8
7 3 P F 1 8 1
A u 2
P F 1 8 5
100 90 80
7 5
7 7, 4
80 ,6
83 ,7
8 6,6
86 ,4
8 9, 2
A u 3
B
45
A
PF
185
PF
187
PF
18
Au
2
PF
91
Au
3
PF
181
PF
182
PM
PF
185
PF
187
Au
20
Au
2
PF
91
Au
3
PF
181
PF
182
Totales Externas
116
35
25
18,4
66,2
14,4
Au 2
PF1 8 5
PF 1 8 2
PF1 8 7
PF 9 1
100 90 80
PF1 8 8
A u 3
PF 1 8 1
85,4
75,3
92,5
81,3
87,5
92
96,6
P F 9 1
P F 1 88
P F 1 8 7
P F 1 8 2
2 8
7 3 P F 1 8 1
A u 2
P F 1 8 5
100 90 80
7 5
7 7, 4
80 ,6
83 ,7
8 6,6
86 ,4
8 9, 2
A u 3
B
45
AP
F185
PF
187
PF
18
Au
2
PF
91
Au
3
PF
181
PF
182
PM
PF
185
PF
187
Au
20
Au
2
PF
91
Au
3
PF
181
PF
182
Totales Externas
116
35
25
18,4
66,2
14,4
Au 2
PF1 8 5
PF 1 8 2
PF1 8 7
PF 9 1
100 90 80
PF1 8 8
A u 3
PF 1 8 1
85,4
75,3
92,5
81,3
87,5
92
96,6
P F 9 1
P F 1 88
P F 1 8 7
P F 1 8 2
2 8
7 3 P F 1 8 1
A u 2
P F 1 8 5
100 90 80
7 5
7 7, 4
80 ,6
83 ,7
8 6,6
86 ,4
8 9, 2
A u 3 A)
Proteínas Totales
B)
Proteínas Externas

39
A)
Proteínas Totales
T o ta le s Ex te rn a s P
M
PF
2
PF
3
PF
4
PF
6
PF
7
PF
8
PF
9
PF
11
PM
1 1 6
4 5
3 5
1 8 , 4
1 4 , 4
6 6 ,2
PM
PF
2
PF
3
PF
4
PF
6
PF
7
PF
8
PF
9
PF
11
PM
1 1 6
4 5
3 5
1 8 ,4
1 4 , 4
6 6 ,2
T o t a le s E x t e rn a s
A
B
P F 3
PF 11
PF 6
PF 8
PF 4
P F 7
PF 2
PF 9
9 8, 3
100 8 0 6 0
9 6, 2
8 7 ,7
7 8 ,1 9 8, 5
81 ,7
8 3 ,4
PF 3
P F 4
P F 9
PF 7
PF 8
P F1 1
PF 6
PF 2
8 1,7
91 ,8
83
80 ,5
75 ,5
9 2, 1
100 80 60
T o ta le s Ex te rn a s
PM
PF
2
PF
3
PF
4
PF
6
PF
7
PF
8
PF
9
PF
11
PM
1 1 6
4 5
3 5
1 8 , 4
1 4 , 4
6 6 ,2
PM
PF
2
PF
3
PF
4
PF
6
PF
7
PF
8
PF
9
PF
11
PM
1 1 6
4 5
3 5
1 8 ,4
1 4 , 4
6 6 ,2
T o t a le s E x t e rn a s
A
B
P F 3
PF 11
PF 6
PF 8
PF 4
P F 7
PF 2
PF 9
9 8, 3
100 8 0 6 0
9 6, 2
8 7 ,7
7 8 ,1 9 8, 5
81 ,7
8 3 ,4
PF 3
P F 4
P F 9
PF 7
PF 8
P F1 1
PF 6
PF 2
8 1,7
91 ,8
83
80 ,5
75 ,5
9 2, 1
100 80 60
ST ST
T o ta le s Ex te rn a s
PM
PF
2
PF
3
PF
4
PF
6
PF
7
PF
8
PF
9
PF
11
PM
1 1 6
4 5
3 5
1 8 , 4
1 4 , 4
6 6 ,2
PM
PF
2
PF
3
PF
4
PF
6
PF
7
PF
8
PF
9
PF
11
PM
1 1 6
4 5
3 5
1 8 ,4
1 4 , 4
6 6 ,2
T o t a le s E x t e rn a s
A
B
P F 3
PF 11
PF 6
PF 8
PF 4
P F 7
PF 2
PF 9
9 8, 3
100 8 0 6 0
9 6, 2
8 7 ,7
7 8 ,1 9 8, 5
81 ,7
8 3 ,4
PF 3
P F 4
P F 9
PF 7
PF 8
P F1 1
PF 6
PF 2
8 1,7
91 ,8
83
80 ,5
75 ,5
9 2, 1
100 80 60
PF
2
PF
3
PF
4
PF
6
PF
7
PF
8
PF
9
PF
11
ST
B) Proteínas Externas
Listonella anguillarum
El análisis de los perfiles proteicos tanto de proteínas totales como proteínas externas
mostró una alta homología, pero con menor porcentaje respecto a V. ordalii. Los patrones de
proteínas totales presentan un porcentaje de similitud del 78,1%, mientras que en el caso de las
proteínas de membrana externa fue del 75,5% (Figura 10B), los dendogramas fueron realizados
usando el programa GelCompar II. Es interesante destacar que el 100% de los aislados de L.
anguillarum expresaron la banda característica de una proteína de membrana externa (porina) de
aproximadamente 40 kDa descrita para el género Vibrio, zona delimitada por el recuadro rojo .
Figura 10. Electroforesis en SDS-PAGE de proteínas y dendograma para L. anguillarum
Análisis de proteínas totales y externas de los aislados de L. anguillarum en geles de SDS al
12%, tinción con azul de Coomassie (A) Perfiles electroforético y dendograma de proteínas totales (B) perfil electroforético y dendograma obtenido de las proteínas externas. El recuadro
rojo destaca la zona donde se encontraría la proteína de membrana externa (banda negativa) de
aproximadamente 40 kDa, ST : estándar de peso molecular.

40
6.3.2. Lipopolisacáridos (LPS)
Los lipopolisacáridos o LPS son biomoléculas que definen muchas interacciones
patógeno-hospedero y posee inmunodominancia antigénica en la mayoría de las bacterias
Gram-negativas. Por esta razón, el estudio de estas estructuras es útil y nos entrega información
tanto de las caraterísticas de la complejidad estructural como de las características serológicas.
Los resultados del estudio de las cepas de V. ordalii detectaron una gran homología en la
composición del LPS (Figura 11A), siendo el porcentaje de similitud de un 90,8%. Por otra
parte, los perfiles de los aislados de L. anguillarum mostraron 2 patrones claramente definidos,
los que fueron codificados como P1, P2 Figura 11B). Las principales diferencias entre P1 y P2
fue la ausencia cadenas “O” de peso intermedio (ausencia de bandas ubicadas en la región
media.del perfil electroforetico) y una zona difusa de alto peso en P2.
Figura 11. Perfiles electroforéticos de LPS de los aislados de vibriosis en Chile.
Los LPS de los aislados se separaron mediante electroforesis en geles de acrilamida SDS-PAGE
al 12% y tinción con plata A) perfil electroforético para V. ordalii B) perfil electroforético para
las cepas de L. anguillarum pertenecientes a los 3 patrones señalados (P1, P2 y P3). PM:
estándar de peso molecular.
6.3. Estudios serológicos de agentes causales de vibriosis
De acuerdo a los resultados obtenidos en los ensayos anteriores se seleccionaron 20
aislados, como grupo representativo de L. anguillarum y V. ordalii. Para esta selección se
consideró además, el origen geográfico de las cepas y huésped de aislamiento (Tabla 2).
Listonella anguillarum Vibrio ordali i
PM
Au10
PF
185
PF
187
PF
186
Au20
Au2
Au9
Au3
PF
181
PF
182
PM
66,2
35
45
14,4
18,4
25
A
PM PM PMPF
2
PF
5
PF
9
PF
12
PF
6
PF
7
PF
10
PF
11
PF
16
PF
17
P1
B
P2 P1
PM PM PMPF
2
PF
5
PF
9
PF
12
PF
6
PF
7
PF
10
PF
11
PF
16
PF
17
P1
B
P2 P1
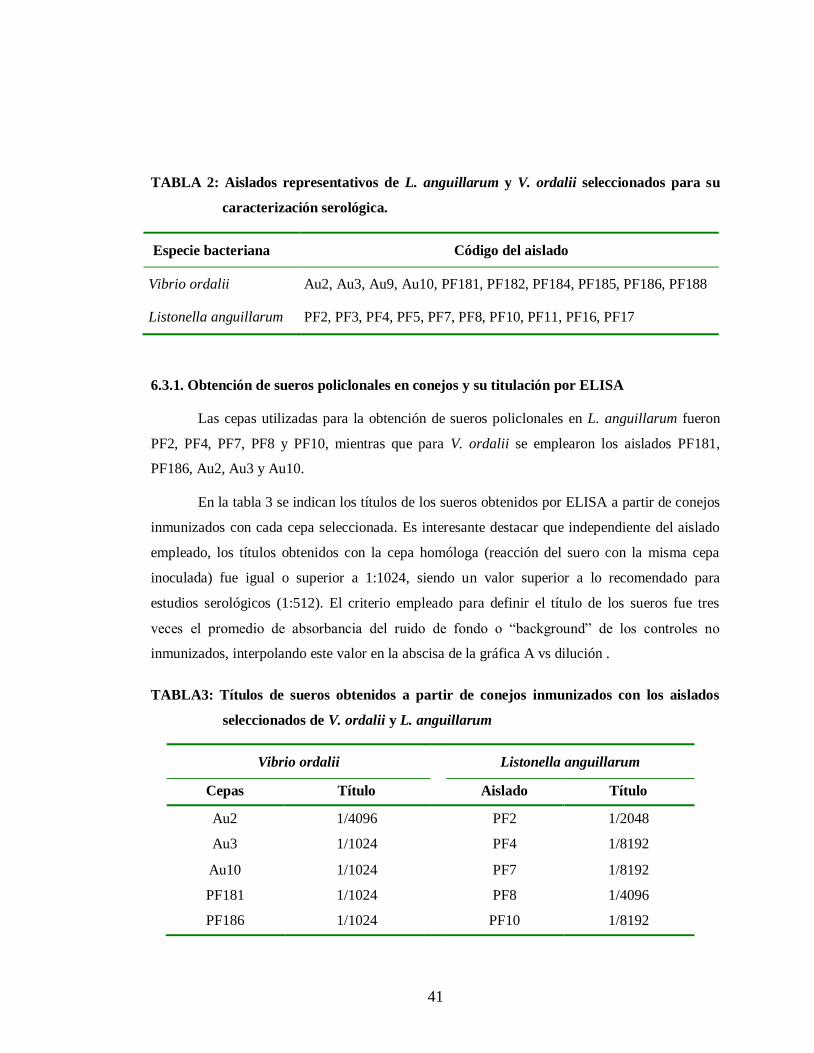
41
TABLA 2: Aislados representativos de L. anguillarum y V. ordalii seleccionados para su
caracterización serológica.
Especie bacteriana Código del aislado
Vibrio ordalii Au2, Au3, Au9, Au10, PF181, PF182, PF184, PF185, PF186, PF188
Listonella anguillarum PF2, PF3, PF4, PF5, PF7, PF8, PF10, PF11, PF16, PF17
6.3.1. Obtención de sueros policlonales en conejos y su titulación por ELISA
Las cepas utilizadas para la obtención de sueros policlonales en L. anguillarum fueron
PF2, PF4, PF7, PF8 y PF10, mientras que para V. ordalii se emplearon los aislados PF181,
PF186, Au2, Au3 y Au10.
En la tabla 3 se indican los títulos de los sueros obtenidos por ELISA a partir de conejos
inmunizados con cada cepa seleccionada. Es interesante destacar que independiente del aislado
empleado, los títulos obtenidos con la cepa homóloga (reacción del suero con la misma cepa
inoculada) fue igual o superior a 1:1024, siendo un valor superior a lo recomendado para
estudios serológicos (1:512). El criterio empleado para definir el título de los sueros fue tres
veces el promedio de absorbancia del ruido de fondo o “background” de los controles no
inmunizados, interpolando este valor en la abscisa de la gráfica A vs dilución .
TABLA3: Títulos de sueros obtenidos a partir de conejos inmunizados con los aislados
seleccionados de V. ordalii y L. anguillarum
Vibrio ordalii Listonella anguillarum
Cepas Título Aislado Título
Au2 1/4096 PF2 1/2048
Au3 1/1024 PF4 1/8192
Au10 1/1024 PF7 1/8192
PF181 1/1024 PF8 1/4096
PF186 1/1024 PF10 1/8192

42
Es interesante destacar que en el caso de L. anguillarum se detectaron claramente dos
grupos serológicos cuando se realizó la titulación de los sueros empleando el antígeno “O” de las
cepas heterólogas. Así, un grupo estaría conformado por los aislados PF2, PF4, PF7 y PF8, los
cuales mostraron un título similar con las cepas homólogas y heterólogas, excepto con el aislado
PF10 (Figura 12). El otro grupo, precisamente estaría constituido sólo por este aislado, ya que
cuando se empleó el suero PF10 se obtuvo una respuesta menor con todas las cepas heterólogas.
Figura 12: Gráfica de titulación por ELISA de sueros de conejo (anti-L.anguillarum)
Resultados de titulación con la técnica de ELISA de los sueros obtenidos en conejo con los aislados de L. anguillarum PF7 (A) y PF10 (B) frente a los antígenos homólogos y heterólogos.
Como control se utilizó el suero preinmune (triangulo verde). Entre la dilución 1/128 y 1/8192
del suero preinmune destaca el ruido de fondo (zona promedio) para definir el título de cada suero.
6.3.2. Ensayo de aglutinación en porta-objeto
Este ensayo se realizó sobre un portaobjeto, utilizando el antígeno termoestable “O” y
cada suero policlonal no absorbido. Al emplear los antisueros de salmónidos chilenos de V.
ordalii, claramente se observó una reacción fuerte con el antígeno “O” de todos los aislados
independiente del suero y antígeno empleado (Tabla 4). Como era de esperar, también se detectó
la reacción cruzada característica de esta especie con el suero policlonal anti-serotipo O2 de L.
anguillarum. Es interesante señalar que al emplear PBS y el suero pre-inmune ninguna cepa
mostró aglutinación ni reacción inespecífica.

43
En cuanto a las cepas de L. anguillarum, los resultados mostraron la existencia de dos
grupos serológicos similar a lo descrito anteriormente usando la técnica de ELISA, pues
nuevamente el aislado PF10 mostró un comportamiento diferente. Sorprendentemente, no existió
reacción antigénica de ninguna de las cepas de L. anguillarum con el antisuero pertenecientes a
los serotipo O1 y O2 de L. anguillarum, ambos reconocidos como los serotipos que afectan a
especies salmonídeas.
TABLA 4: Ensayo de aglutinación en portaobjeto usando los antisueros de L. anguillarum
y V. ordalii para los aislados chilenos.
+: aglutinación positiva, - : aglutinación negativa, O1: anti-serotipo O1 de L. anguillarum, O2:
anti-serotipo O2 de L. anguillarum, PI: suero pre-inmune. Se destaca en la diagonal la reacción
entre los sueros con su cepa homóloga.
6.3.3. Ensayos de Dot-blot
Para la caracterización serológica se utilizaron los 20 aislados y los sueros policlonales
obtenidos en conejo de raza Neozelandesa. Además, se incluyeron como controles y con fines
comparativos el suero preinmune, tampón fosfato salino y los sueros anti-serotipo O1 y anti-
serotipo O2 de L. anguillarum.
SUEROS
Vibrio ordalii Listonella anguillarum Controles
CEPAS Au10 PF186 PF188 Au2 Au3 PF2 PF4 PF7 PF8 PF10 O1 O2 PI
Au10 + + + + + - - - - - - + -
PF186 + + + + + - - - - - - + -
PF188 + + + + + - - - - - - + -
Au2 + + + + + - - - - - - + -
Au3 + + + + + - - - - - - + -
PF2 - - - - - + + + + - - - -
PF4 - - - - - + + + + - - - -
PF7 - - - - - + + + + - - - -
PF8 - - - - - + + + + - - - -
PF10 - - - - - - - - - + - - -

44
a) Vibrio ordalii
Con el fin de establecer una relación previa entre los sueros y los aislados seleccionados,
se realizó el ensayo de dot-blot según 5.4.4. Los resultados mostraron una reacción fuerte y clara
de todos los antisueros de V. ordalii y los antígenos de las cepas homólogas y heterólogas
empleadas, indicando la existencia de homogeneidad serológica dentro de las cepas de
salmónidos aislados en cultivos chilenos. Se observaron resultados similares fueron observados
cuando los ensayos se repitieron empleando el antisuero representativo del serotipo O2 de L.
anguillarum (Figura 13). Además, no se detectó reacción con los controles negativos incluídos
en este estudio, es decir, PBS, suero pre-inmune, antisueros de las cepas de L. anguillarum y sus
respectivos antígenos.
b) Listonella anguillarum
Los resultados de los ensayos de dot-blot fueron idénticos a aquellos obtenidos mediante
las técnicas serológicas de ELISA y aglutinación en porta-objetos. De esta forma, se evidenció la
existencia de dos grupos antigénicos, codificados como GI y GII. El Grupo I conformado por la
mayoria de los aislados de L. anguillarum estudiados, excepto las cepas PF10, PF11 y PF16, los
cuales reaccionan fuertemente con los antisueros obtenidos a partir de conejos inmunizados con
las cepas de salmónidos PF2, PF4, PF7 y PF8 (Figura 14A). Precisamente, los tres aislados
(PF10, PF11 y PF16) son los que contituyen el Grupo II, reaccionando fuertemente sólo con el
suero anti-PF10 (Figura 14B). Cabe destacar que cada uno de los grupos contiene aislados de
distinto origen geográfico y huésped de aislamiento.
Por otra parte, ninguno de los dos sueros representativos de los serotipos de L.
anguillarum (serotipos O1 y O2) logró reconocer algunos de los antígenos de las cepas chilenas
de L. anguillarum, lo cual sugiere la probabilidad que las cepas chilenas pertenezcan a los
restantes serotipos descritos para esta especie bacteriana.

45
PF3 PF6 PF9 PF11 Au10
PF2 PF4 PF7 PF8 PF10 A
PF3 PF6 PF9 PF11 Au10
PF2 PF4 PF7 PF8 PF10 B
Suero anti-PF10 (L. anguillarum) Suero anti-PF2 (L. anguillarum)
Figura 13: Ensayo de dot-blot de los aislados chilenos de Vibrio ordalii
Dot-blot obtenido con los sueros anti-Au2 de Vibrio ordalii (A) y pre-inmune (B) frente a los
antígenos “O” de las cepas chilenas de L. anguillarum (filas 1 y 2) y V. ordalii (filas 3 y 4).
Como control negativo se incluyo tampón fosfato salino (PBS).
Figura 14: Ensayo de dot-blot de los aislados chilenos de Listonella anguillarum
Resultados de dot-blot usando los sueros anti L. anguillarum PF2 (A) y PF10 (B) frente a los
antígenos “O” de las cepas chilenas de L. anguillarum, (A) muestra los aislados perteneciente al
Grupo I y (B) muestra los aislados perteneciente al Grupo II.
Suero anti-Au2 (V. ordalii) Suero Preinmune
A
B
A

46
6.3.4. Ensayos de Western-blot
En las bacterias Gram-negativas, es importante incluir un estudio de las estructuras
externas de la bacteria, como lo son las proteínas externas de membrana y el LPS. La
herramienta que nos entrega mayor información es el western-blot o inmuno-blot, ya que a
diferencia del dot-blot es posible establecer un perfil característico de las cadenas “O” que
componen el LPS y las bandas proteícas, que nos permiten determinar variabilidad o similitud
entre los aislados.
En este estudio se utilizaron las mismas cepas y antisueros usados en la técnica de dot-
blot, sin embargo, con el fin de determinar posibles diferencias antigénicas, los ensayos se
realizaron con los antisueros absorbidos, tanto con el aislado homólogo como heterólogo.
a) Proteínas de membranas externas
Los resultados del análisis de la envoltura celular externa de las cepas de V. ordalii,
demostró que antigénicamente existe una gran homología en los componentes peptídicos que
conforman la membrana celular de todos los aislados de V. ordalii estudiados, independiente del
suero empleado. Pese a esta similitud, es posible detectar diferencias en los patrones de
proteínas analizados mediante western-blot, obtenidos con los distintos antisueros (Figura 15).
En cuanto a los aislados de L. anguillarum, la heterogeneidad descrita en los análisis
anteriores se hace aún más evidente al usar la técnica de western-blot. Así según el suero
utilizado se distinguen los dos grupos antigénicos presentados en los ensayos anteriores, el
Grupo I (PF2, PF4, PF7, PF8) presenta un número mayor de bandas en su perfil que el Grupo II
dado por el aislado PF10 (Figura 16).
Al usar sueros absorbidos de L. anguillarum con aislados homólogos y heterólogos, se
logró establecer que dentro del Grupo I existen dos subgrupos, uno conformado por los aislados
PF2 y PF4 y otro constituído por las cepas PF7 y PF8, como se observa en la figura 16A y 16B,
donde se utilizó el suero anti-PF7 absorvido con el aislado PF4 (figura 16A), presentando
ausencia de bandas en los perfiles PF2 y PF4. En la figura 16B se puede observar que usando el
suero anti-PF4 absorbido con el antígeno del aislado PF7, también se pierden bandas proteicas
en los aislados PF7 y PF8 (Figura 16B).
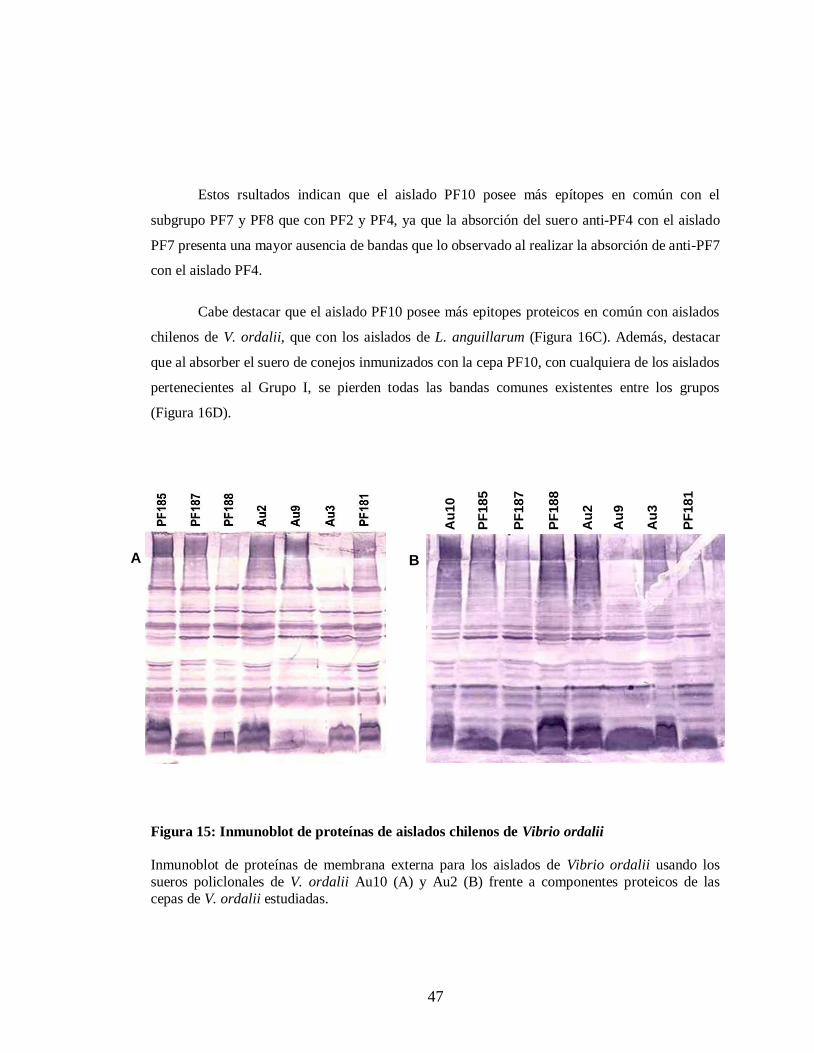
47
Estos rsultados indican que el aislado PF10 posee más epítopes en común con el
subgrupo PF7 y PF8 que con PF2 y PF4, ya que la absorción del suero anti-PF4 con el aislado
PF7 presenta una mayor ausencia de bandas que lo observado al realizar la absorción de anti-PF7
con el aislado PF4.
Cabe destacar que el aislado PF10 posee más epitopes proteicos en común con aislados
chilenos de V. ordalii, que con los aislados de L. anguillarum (Figura 16C). Además, destacar
que al absorber el suero de conejos inmunizados con la cepa PF10, con cualquiera de los aislados
pertenecientes al Grupo I, se pierden todas las bandas comunes existentes entre los grupos
(Figura 16D).
Figura 15: Inmunoblot de proteínas de aislados chilenos de Vibrio ordalii
Inmunoblot de proteínas de membrana externa para los aislados de Vibrio ordalii usando los
sueros policlonales de V. ordalii Au10 (A) y Au2 (B) frente a componentes proteicos de las
cepas de V. ordalii estudiadas.
A
PF
18
7
PF
18
8
PF
18
1
PF
18
5
Au
2
Au
3
Au
9
Au
10
B

48
Figura 16: Inmunoblot de proteínas de aislados chilenos de Listonella anguillarum
Inmunoblot de proteínas de membranas externas de L. anguillarum usando los sueros
absorbidos de la misma especie bacteriana. (A) anti-PF7 absorbido con el antígeno procedente
del aislado PF4; (B) anti-PF4 absorbido con el antígeno del aislado PF7; (C) anti-PF10 sin absorber y (D) anti-PF10 absorbido con el antígeno procedente del aislado PF4. Carril: 10, PF10;
8, PF8; 7, PF7; 4, PF4; 2, PF2 y Vo, aislado representativo de la especie V. ordalii.
b) Lipopolisacáridos
Como se mencionó anteriormente, los LPS son estructuras inmuno-estimuladoras y
cumplen un rol protector para los microorganismos. Además, estas estructuras han sido
utilizadas para la caracterización serológica de las bacterias, particularmente en base a los
patrones electroforéticos, los cuales se pueden divididir en tres zonas de reconocimiento,
facilitando así el análisis antigénico. Una zona superior o de alto peso (HMW, high molecular
weight), una zona media o IMW (intermediate molecular weight) y una zona de bajo peso o
LMW (low molecular weight).
10 8 7 4 2 10 8 7 4 2 Vo 10 8 7 4 2 10 8 7 4 2 St
A B C D
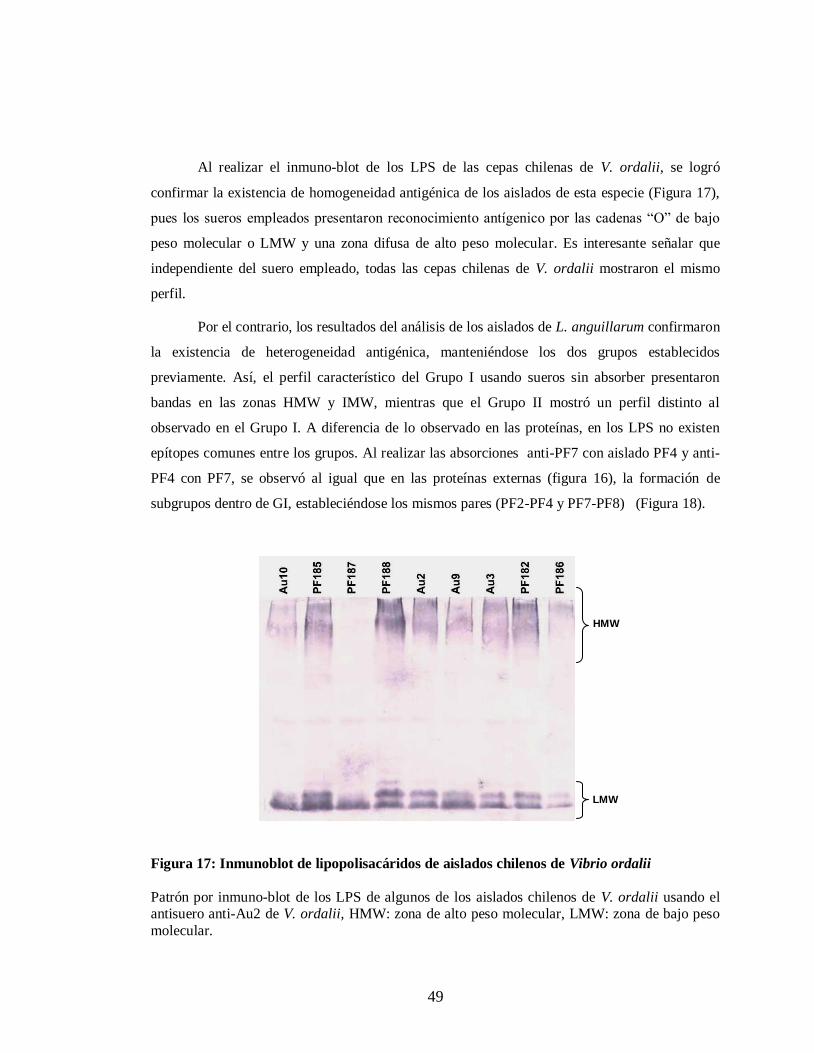
49
Al realizar el inmuno-blot de los LPS de las cepas chilenas de V. ordalii, se logró
confirmar la existencia de homogeneidad antigénica de los aislados de esta especie (Figura 17),
pues los sueros empleados presentaron reconocimiento antígenico por las cadenas “O” de bajo
peso molecular o LMW y una zona difusa de alto peso molecular. Es interesante señalar que
independiente del suero empleado, todas las cepas chilenas de V. ordalii mostraron el mismo
perfil.
Por el contrario, los resultados del análisis de los aislados de L. anguillarum confirmaron
la existencia de heterogeneidad antigénica, manteniéndose los dos grupos establecidos
previamente. Así, el perfil característico del Grupo I usando sueros sin absorber presentaron
bandas en las zonas HMW y IMW, mientras que el Grupo II mostró un perfil distinto al
observado en el Grupo I. A diferencia de lo observado en las proteínas, en los LPS no existen
epítopes comunes entre los grupos. Al realizar las absorciones anti-PF7 con aislado PF4 y anti-
PF4 con PF7, se observó al igual que en las proteínas externas (figura 16), la formación de
subgrupos dentro de GI, estableciéndose los mismos pares (PF2-PF4 y PF7-PF8) (Figura 18).
Figura 17: Inmunoblot de lipopolisacáridos de aislados chilenos de Vibrio ordalii
Patrón por inmuno-blot de los LPS de algunos de los aislados chilenos de V. ordalii usando el antisuero anti-Au2 de V. ordalii, HMW: zona de alto peso molecular, LMW: zona de bajo peso
molecular.
LMW
HMW

50
Figura 18: Inmunoblot de lipopolisacáridos de aislados chilenos de Listonella anguillarum
Perfil por Inmunoblot de los LPS de aislados chilenos de L. anguillarum usando los sueros no absorbidos anti-PF4 (A) y anti-PF10 (B), así como los antisueros PF7 absorbido con el antígeno
de la cepa PF4 (C) y el suero anti-PF4 absorbido con el antígeno de la cepa PF7 (D) donde GI:
grupo I, GII: grupo II, HMW: zona de alto peso molecular, IMW: zona de peso molecular
intermedio.
B A
C D PF2 PF4 PF7 PF8 PF10
C DC
PF2 PF4 PF7 PF8 PF10
GI GII
HMW
IMW

51
6.4. Estudios preliminares de virulencia
Con el propósito de evaluar la virulencia de las cepas chilenas de V. ordalii y L.
anguillarum se procedió a realizar un desafío de peces con estos patógenos. Los resultados
demostraron una alta mortalidad en los ejemplares de salmón del Atlántico, la cual comenzó 24
horas post-inoculación, alcanzando el 100% a los dos días de haber comenzado la experiencia.
Todos los peces mostraron los signos clásicos de vibriosis, incluyendo hemorragia en la base de
la aleta ventral, alrededor del poro anal y superficie externa (Figura 19). Además, en todos los
casos se recuperó en estado puro la cepa inoculada sembrada en placas de agar TSA a partir de
muestras de riñón.
Figura 19. Mortalidad en los ensayos preliminares de virulencia
(A) Ejemplares de salmón del Atlántico mostrando signos clínicos clásicos de vibriosis
particularmente zonas hemorragía en la superficie externa (indicada con cabeza de flecha) y (B)
control sin inoculación de bacteria (PBS).
A
B
A
B
A
B
A

52
7. DISCUSIÓN
La vibriosis está considerada como una de las patologías que causan grandes pérdidas
económicas en la industria acuícola mundial. A partir del año 2003, Chile no está ajena a esta
realidad detectándose las primeras mortalidades de salmónidos en el sur de Chile, por tanto
obteniéndose aislados de Vibrio ordalii y presuntamente de Listonella anguillarum. En este
estudio se analizaron un total de 77 aislados obtenidos a partir de casos clínicos de vibriosis
ocurridos entre los años 2004-2005.
7.1. Identificación
Los ácidos nucleicos y las proteínas son macromoléculas comunes en los organismos
vivos, que sufren cambios evolutivos en el tiempo, por lo cual pueden ser consideradas como
cronómetros moleculares y ser utilizadas para identificar y caracterizar a los microorganismos en
estudio.
Precisamente, una de las estructuras más empleadas para la caracterización e
identificación de diversas especies ha sido el ARN ribosomal. Esta macromolécula se ha
transformado hoy en día, en la estructura más comúnmente utilizada para estudios filogenéticos
y taxonómicos de microorganismos bacterianos, particularmente debido a que ha permanecido
altamente conservada a través de la evolución de organismos procariontes, arqueas y eucariontes
(Weisburg y cols., 1991, Rodicio y Mendoza, 2004), permitiendo diferenciar las especies que
habitan el planeta.
A través del análisis filogenético del gen ARNr 16S, en este estudio se estableció que
todos los aislados presentaron homología con dos microorganismos pertenecientes a la familia
Vibrionaceae. Se logró identificar que de los 77 aislados, 61 correspondieron a la especie V.
ordalii y 13 a la especie L. anguillarum, mientras que tres aislados mostraron similitud con
miembros del género Aeromona, que no fueron incluidas en este estudio.
Si bien, Colquhoun y cols., (2004) habían establecido la presencia de V. ordalii como un
patógeno emergente en cultivos de salmónidos de nuestro país, hasta ahora L. anguillarum sólo
había sido descrito como patógeno asociado a mortalidades de bivalvos (Riquelme y cols.,
1995). La aparición de esta especie en el país era mas o menos predecible, ya que el explosivo

53
crecimiento de la producción de salmón chileno en la última década (SalmonChile 2006),
sumado a que L. anguillarum es un microorganismo natural del medio marino y estuarino,
proporcionan las condiciones óptimas (Schiewe y Harrell, 1983) para la aparición de brotes de
vibriosis clásica (por L. anguillarum) en nuestro país. Probablemente, L. anguillarum no había
sido aislado a partir de peces muertos o moribundos debido a que la vibriosis a veces puede
preceder a otras patologías, particularmente a las causadas por la bacteria Pisciricketssia
salmonis o al virus de la necrosis pancreática infecciosa (IPNv). También pudo deberse a que la
especie no expresaba serotipos patogénicos, o bien existir una introducción o migración de la
bacteria desde otros lugares, estableciéndose un período de adaptación de la especie a las
condiciones de la región. Otra razón puede ser que en Chile no había mayor interés en estudiar
estas patologías, y prueba de ello es el bajo número de publicaciones nacionales respecto a este
tema. Además, el muestreo e identificación de los patógenos realizados por laboratorios de
diagnóstico para peces cuentan con metodologías de identificación de rutina que en ocasiones no
detectan diferencias entre los aislados, los que son identificados a nivel de género o como
especies o atípicas.
Esta dificultad está dada por la heterogeneidad bioquímica presentada, siendo necesario
el uso de un amplio rango de pruebas para su correcta identificación. Prueba de ello es que
muchos de los aislados nacionales fueron descritos como V. ordalii atípico o simplemente Vibrio
sp. También puede deberse a escasez de instrumental, metodologías o mal uso de ambas.
7.2 Caracterización genética
Los genes ribosomales están organizados en operones, es decir, conjuntos de genes que
se transcriben a partir de la misma región promotora. Cada operón ribosómico incluye genes
para los ARNr 23S, 16S y 5S, los que están separados por regiones intergénicas o espaciadores
intergénicos que pueden o no contener secuencias para la síntesis de ARN de transferencia
(Rodicio y Mendoza, 2004). Diversos estudios han determinado que la región espaciadora entre
el 16S-23S es menos conservada a través de la evolución que los genes ARNr 16S y 23S. Esta
mayor variabilidad, característica que convierte a las regiones espaciadoras en una zona útil para
detectar diferencias entre especies, lo cual han permitido realizar multiples estudios genéticos
comparativos dentro de una misma especie (Jensen y cols., 1993; Nagpal y cols., 1998).

54
El análisis de las secuencias intergénicas de los aislados estudiados en el presente
trabajo, presentó claramente la existencia de dos patrones electroforéticos. La diferencia entre
ambos perfiles fue la presencia de una banda adicional de bajo peso molecular
(aproximadamente 550 pb), la cual permitió agrupar las cepas chilenas dentro de las especies V.
ordalii y L. anguillarum. Coincidentemente, los tres aislados pertenecientes al género
Aeromonas mostraron un perfil distinto a los señalados previamente y fueron descartadas para
los estudios posteriores de caracterización genética, molecular y serológica. Hasta el inicio de
este trabajo, en la literatura no existían antecedentes relevantes sobre la secuencia de la región
espaciadora 16S–23S en las especies V. ordalii y L. anguillarum.
Recientemente ha sido incorporada a la base de datos GenBank la secuencia de V.
ordalii obtenida a partir de la cepa tipo, por lo que aún se realizan comparaciones genéticas de
estos aislados chilenos. Sin embargo, es interesante señalar que al igual que en otros estudios
(Jensen y cols., 1993; Gonzálezb y cols., 2003), la técnica de PCR-ITS permitió diferenciar los
aislados, los cuales fueron previamente en base a su secuencia del gen ARNr 16S, incluso de
especies relacionadas pertenecientes a la misma familia bacteriana, como es el caso de los tres
aislados de Aeromonas.
Al realizar la digestión del fragmento de amplificación entre los genes ARNr 16S y 23S
o RFLP-ITS, del total de las enzimas de restricción utilizadas, sólo dos enzimas (Pvu II y Hae
III) efectuaron cortes de los productos PCR-ITS. No se observó diferencias en los patrones
electroforéticos (RFLP).
Con el fin de evaluar otras técnicas de genotipado, se realizó un análisis del genoma
completo de la bacteria usando la técnica de campo pulsado (PFGE), detectando que los aislados
chilenos de V. ordalii son iguales, excepto el aislado Au10 mostró pequeñas diferencias en su
perfil, al repetir el ensayo, el perfil fue el característico para la mayoría de los aislados chilenos
de V. ordalii, demostrando que en experiencias anteriores la digestión de la enzima Not I fue
incompleta. Los perfiles de PFGE para V. ordalii podrían ser indistinguibles (según Tenover F.C
y cols., 1995) pero la ausencia de una banda de aproximadamente 65 pb, presentada por el
aislado Au3 puede ser considerada como una diferencia, siendo definidos los perfiles como
patrones estrechamente relacionados, diferencias producidas por un evento genético (Tenover
F.C y cols., 1995). Esta diferencia no es sorprendente, ya que Tainen y cols., (1995) señalan en

55
un estudio realizado con aislados de V. ordalii de diferentes orígenes geográficos, en el cual
mostró valores de similitud del orden del 95%.
En cuanto a la heterogeneidad genética mostrada por los aislados chilenos de L.
anguillarum, estudios previos de Skov y cols. (1995) utilizando la misma técnica y enzima que
en esta tesis, informan la existencia de hasta 35 perfilesdistntos. Cabe destacar que estos autores
analizaron un total de 75 aislados europeos de L. anguillarum, concluyendo que dentro de esta
especie bacteriana existe una importante heterogeneidad genética, concordante con lo presentado
por los aislados chilenos. Según Tainen y cols., (1995) los aislados serían considerados como no
relacionados a un brote específico, ya que sus patrones electroforéticos presentarían a lo menos
tres eventos genéticos, compartiendo entre ellos < 50% de las bandas.
Con el fin de confirmar si esta heterogeneidad depende de la técnica empleada, así como
validar otras herramientas moleculares basadas en amplificación aleatoria, se realizaron estudios
con REP-PCR y ERIC-PCR. Ambas técnicas reafirmaron lo detectado por PFGE, es decir, la
homogeneidad y heterogeneidad genética en los aislados de V. ordalii y L. anguillarum,
respectivamente.
En los últimos años, las técnicas moleculares basadas en la amplificación aleatoria han
progresado rápidamente, siendo más rápidas y requiriendo de menos equipamiento en
comparación al PFGE. Así, técnicas de amplificación dirigidas a elementos genéticos repetitivos
tales como REP y ERIC han sido usados para detectar “huellas digitales” de bacterias Gram-
negativas y positivas. En el caso del género Vibrio, Rivera y cols. (1995) proponen el uso de
ERIC-PCR para la tipificación de diversas especies como V. mimicus, V. vulnificus, V.
parahaemolyticus, V. campbelli, V. meditarranei y V. alginolyticus, mientras que la técnica de
PCR-REP también ha sido usada satisfactoriamente en la discriminación de cepas de V.
parahaemolyticus (Marshall y cols., 1999; Wong y Lin 2001; Khan y cols., 2002; Maluping y
cols., 2005), V. cholerae (Rivera y col., 1995; Shangkuan y cols., 1997) y V. tapenis (Rodríguez
y cols., 2006).

56
7.3. Caracterización de los componentes de la envoltura celular
El análisis de las características estructurales de los aislados bacterianos, es esencial para
el diseño de medios preventivos, particularmente aquellos que inciden directamente en la
elaboración de una vacuna como LPS y proteínas de membrana. Por ello, en la presente tesis se
estudiaron con detenimiento ambos componentes.
Los resultados han demostrado existencia de similitud en los componentes proteicos y
LPS que conforman la membrana de los aislados chilenos de V. ordalii, por otra parte los
aislados de L. anguillarum presentaron mayor variabilidad en las proteínas. Sin embargo, las
diferencias fueron más claras al observar los perfiles de los LPS, detectando en L. anguillarum
dos grupos mayoritarios
Al estudiar aislados representativos de L. anguillarum, Grisez y Ollevier (1995)
realizaron un estudio estableciendo 16 serotipos dentro de la especie. Los primeros 10 serotipos
(O1 - O10) representan a los grupos establecidos por Sorencen y Larsen y los serotipos O11-O16
fueron descritos como tipos D a I por Kitao y cols.1983 y 1984 (citados en Grisez y Ollevier,
1995). Las conclusiones obtenidas en el trabajo de Grisez y Ollevier mostraron que los 16
serotipos poseen patrones distintos de LPS, pudiéndose utilizar esta característica como un
método preliminar de diagnóstico. Basados en estos antecedentes, los dos patrones obtenidos en
nuestro estudio podrían corresponder a serotipos distintos y éstos a su ves, ser los serotipos
característicos de la vibriosis clásica, hipótesis que fue resuelta con los estudios serológicos
posteriores que se describen en este trabajo.
7.4. Caracterización antigénica
Los estudios serológicos entregan un complemento importante a la caracterización
genética y de moléculas estructurales, ya que permite asociar cada uno de estos resultados a la
expresión de la respuesta inmune, valorando lo realmente inmunogénico, obteniendo un
conocimiento clave en la búsqueda y elaboración de una vacuna contra una enfermedad.

57
Los antisueros utilizados para desarrollar estos estudios, preparados a partir de cepas de
L. anguillarum y V. ordalii nativas de nuestro país, mostraron títulos superiores a 1: 1024. Estos
valores se encuentran por sobre el título mínimo recomendado para este tipo de estudio (título 1:
512), siendo similares a aquellos empleados por Grisez y Ollevier (1995).
Al evaluar la respuesta antigénica de las proteínas constituyentes de la envoltura celular
tanto de V. ordalii como L. anguillarum, se detectó que todos los aislados comparten estructuras
antigénicas, siendo ésta una característica propia de la familia Vibrionaceae.
Al realizar la caracterización serológica de los aislados de V. ordalii, independiente de la
técnica empleada, se detectó la existencia de homogeneidad serológica, es decir, los 61 aislados
tienen estructuras antigénicas comunes (LPS y proteínas). Como era de esperar, los mismos
aislados mostraron reacción cruzada con suero anti-serotipo O2 de L. anguillarum (Schiewe y
cols., 1977; Mutharia y cols., 1993), contrario a lo descrito por Colqhoun (2004) quien usando
un número limitado de aislados chilenos de V. ordalii, no logró serotipificar sus muestras por
presentar propiedades autoaglutinantes. Cabe destacar que los sueros y cepas empleadas en este
estudio, son diferentes a las empleadas por el trabajo de Colqhoun (2004).
Los serotipos O1 y O2, son los principales responsables de los brotes de vibriosis en
salmónidos ocurridos en el mundo (Toranzo y Barja 1990; Toranzo y cols., 2005). Independiente
de las técnicas utilizadas, los aislados nacionales de L. anguillarum, no presentaron reacción
antigénica con ninguno de estos antisueros (serotipo O1 y O2 de L. anguillarum). Sin embargo,
el análisis serológico, con los sueros de conejos inmunizados con las cepas nativas de L.
anguillarum, permitieron agrupar los aislados nacionales dentro de dos grupos, denominados
como GI y GII, el primero compuesto por 10 aislados agrupa al 77% de las cepas, mientras que
el otro se encuentra constituido solamente por 3 aislados (23%).
De esta forma, los aislados nacionales de L. anguillarum no pertenecen al serotipo O1 ni
O2 descritos como patógenos de salmónidos, existiendo la posibilidad que estas cepas sean
microorganismos ambientales oportunistas representante de los ocho serotipos restantes
(Toranzo y cols., 2005) o bien los otros 21 serotipos según Pedersen y cols. (1999),
dependiendo del esquema empleado.

58
En los ensayos de virulencia se utilizó tres cepas representativas de cada especie. Los
ensayos demostraron que los aislados chilenos son altamente patogénicos para el salmón del
Atlántico. Además, en todos los ejemplares muertos los signos clínicos propios de la vibriosis
fueron evidentes y se logró re-aislar a los patógenos inoculados. Colquhoun y cols., (2004)
obtuvieron resultados similares, dado que empleando aislados de V. ordalii denotan valores de
mortalidad acumulada entre 91,9–94,3%, dependiendo de la concentración inoculada en los
peces (18 g). Cabe señalar que en este estudio el porcentaje de mortalidad acumulada fue
levemente superior, sin embargo, los peces eran más pequeños (<10 gramos de peso).
Las vacunas usadas en Chile para el control de la vibriosis no han obtenido los
resultados esperados, ya que para una efectiva protección, la vacuna debe ser dirigida contra el
organismo responsable de los brotes de la enfermedad. Por esta razón la naturaleza antigénica
del microorganismo es fundamental para el desarrollo de un efectivo método de control.
Con los resultados obtenidos en esta tesis, tanto en la caracterización molecular como
antigénica, se puede establecer que la formulación de una vacuna de tipo bacterina (vacuna
basada en el uso de la bacteria completa inactivada) contra vibriosis debería incluir, en el caso de
V. ordalii, al menos un aislado o sus componentes antigénicos, mientras que para L. anguillarum
debería contener un representante de cada grupo serológico (GI y GII).
Si bien en la presente tesis se demostró la existencia de patogenicidad de algunos de los
aislados estudiados, aun queda aún por seleccionar a aquellas cepas de V. ordalii y L.
anguillarum más adecuadas para la elaboración de una(s) vacuna(s) contra ambos patógenos. Por
ello, están en proceso estudios determinar variables imprescindibles en la formulación de una
vacuna tales como, dosis letal 50 (DL50), desafíos con cepas homólogas y heterólogas, dosis
mínima protectora, inocuidad, duración de la protección, entre otras. Los estudios de este tipo
son necesarios para determinar las características de la vacuna, como por ejemplo si esta será
mono o polivalente.

59
8. CONCLUSIONES
Sobre la base de los resultados obtenidos en esta tesis, se ha concluido que:
1. A partir de mortalidades por vibriosis en cultivos de salmónidos, en el sur de Chile, se
detectaron como agentes causales dos patógenos: Vibrio ordalii y Listonella anguillarum.
2. Las técnicas de genotipado empleadas en este estudio, excepto RFLP-ITS, son
herramientas eficaces para la caracterización y seguimiento de los aislados de Vibrio
ordalii y Listonella anguillarum.
3. Los aislados de Vibrio ordalii incluidos en este estudio, presentan homogeneidad genética,
molecular y antigénica, independiente del lugar geográfico y huésped de aislamiento.
4. Los aislados chilenos de Listonella anguillarum son genética, molecular y
antigénicamente heterogéneos.
5. A nivel antigénico todos los aislados de Vibrio ordalii reaccionan serológicamente con los
antisueros de los aislados chilenos de esta especie bacteriana, así como con el serotipo O2
de Listonella anguillarum.
6. Los aislados chilenos de Listonella anguillarum, no pertenecen a los serotipos O1 y O2;
causantes característicos de la vibriosis clásica en especies salmónidas. Sin embargo, los
aislados forman dos grupos serológicos.
7. Los conocimientos obtenidos en esta tesis sientan las bases genéticas y serológicas para la
elaboración de una futura vacuna o vacunas que permitan el control de la vibriosis en
Chile.

60
9. REFERENCIAS
Actis, L.A., Fish, W., Crosa, J.H., Kellerman, K., Ellenberger, S.R., Hauser, F.M., Sanders-
Loehr, J. (1986) Characterization of anguibactin, a novel siderophore from Vibrio
anguillarum 775(pJM1). Int. J. Bacteriol. 167:57-65.
Actis, L.A., Tolmasky, M.E, J.H. Crosa (1999) Vibriosis. In: fish diseases and disorders. Vol.3.
Bruno, P.T.K.W.a.D.W. (ed.): CAB international, pp. 523-557.
Austin, B., Austin D. A. (1999) Bacterial fish pathogens: disease of farmed and wild fish. Berlin,
Germany: Springer-Verlag KG.
Austin, B., Austin D.A. (1993) Bacterial fish pathogens: disease in farmed and wild fish.
Chichester: Ellis Horwood Ltd.
Bassler, B.L., Wright, M., Silverman, M.R. (1994) Multiple signalling systems controlling
expression of luminescence in Vibrio harveyi: sequence and function of genes encoding a
second sensory pathway. Mol. Microbiol. 13:273-286.
Bohle O.H., Olivares P., Bustos P., Fernández J., Silva-Rubio A., Jaureguiberry B., Pacheco T.
(2006). Listonella (V) anguillarum, the first isolation of reared salmonids in brackish and
seawater in Southest Chile. Bull. Eur. Assoc. Fish. Pathol. (enviado).
Bolinches, J., Lemos M., Fouz B., Cambra M., Larsen J. L., Toranzo A. E. (1990). Serological
relationship among Vibrio anguillarum strains. J. Aquat. Anim. Health. 2:21-29.
Chai, S., Welch, T.J., Crosa, J.H. (1998) Characterization of the interaction between Fur and the
iron transport promoter of the virulence plasmid in Vibrio anguillarum. Int. J. Biol. Chem.
273:33841-33847.
Chen, Q., Wertheimer, A.M., Tolmasky, M.E., Crosa, J.H. (1996) The AngR protein and the
siderophore anguibactin positively regulate the expression of iron-transport genes in
Vibrio anguillarum. Int. Mol. Microbiol. 22:127-134.
Cipriano, R.C., Pyle J.B., Starliper C.E. , Pyle S.W. (1985) Detection of Vibrio anguillarum
antigen by dot blot assay. J. Wildlife Dis. 21:211-218.

61
Colquhoun, DJ, Aase IL, Wallace C, Baklien A. , Gravningen (2004). First description of Vibrio
ordalii from Chile. Bull. Eur. Assoc. Fish Pathol. 24:185-188.
Crosa, J.H. (1980) A plasmid associated with virulence in the marine fish pathogen Vibrio
anguillarum specifies an iron-sequestering system. Nature. 284:566-568.
Croxato, A. (2006) VanT a central regulator of quorum sensing signalling in Vibrio anguillarum.
Tesis de Doctorado, Departament of Molecular Biology, Umea University, Umea-Sweden.
Donlan, R.M. (2002) Biofilms: microbial life on surfaces. Emerg. Infect. Dis. 8:881-890.
Ezura, Y., Tajima K., Yoshimizu M., Kimura T. (1980) Studies on the taxonomy and serology of
causative organisms of fish vibriosis. Fish Pathol. 14:167-179.
Fernández-Cuenca F. (2004) Aplicaciones de las técnicas de PCR a la epidemiología molecular
de las enfermedades infecciosas. Enferm. Infecc. Microbiol. Clin. 22:355-360.
Filip, C., Fletcher, G., Wulff, J., Earhart, C. F: (1973) Solubilization of the cytoplasmic
membrane of Escherichia coli by the ionic detergent sodium – lauryl sarcosinate. J.
Bacteriol. 115:717-722.
Gonzáleza, S.F., Osorio, C.R., Santos, Y. (2003) Development of a PCR-based method for the
detection of Listonella anguillarum in fish tissues and blood samples. Dis. Aquat. Org.
55:109-115.
Gonzálezb, N., Romero, J., Espejo, R. (2003). Comprehensive detection of bacterial populations
by PCR amplification of the 16S–23S ARNr spacer region. J. Microbiol. Met. 55: 91-97.
Grisez L., Ollevier F. (1995). Comparative Serology of the Marine Fish Pathogen Vibrio
anguillarum. Appl. Env. Microbiol. 61: 4367-4373.
Hill, B.J. (2005) The need for effective disease control in international aquaculture. Dev. Biol.
Basel. 121:3-12.
Hitchcock, P.J., Brown T.M. (1983) Morphological heterogeneity among Salmonella
lipopolysaccharide chemotypes in silver-stained polyacrilamide gels. J. Bacteriol.
154:269-277.

62
Instituto Tecnológico del Salmón (2000) Programa Nacional de Salud de la Industria
Salmonicultora. Bases del Programa.
Jensen, M.A., Webster J.A., Straus N. (1993) Rapid identification of bacteria on the basis of
polymerase chain reaction-amplified ribosomal DNA spacer polymorphisms. Appl.
Environ. Microbiol. 59:945-952.
Khan, A., McCarthy, S., Wang, R., Cerniglia, C. 2002. Characterization of United States
outbreak isolates of Vibrio parahaemolyticus using enterobacterial repetitive intergenic
consensus (ERIC) PCR and development of a rapid PCR method for detection of O3:K6
isolates. FEMS Microbiol. Lett. 206, 209-214.
Kitao, T., T. Aoki, M. Fukudome, K. Kawano, Y. Wada, Y. Mizuno. (19839 Serotyping of
Vibrio anguillarum isolated from diseased freshwater fish in Japan. J. Fish Dis. 6:175-181.
Koster, W.L., Actis, L.A., Waldbeser, L.S., Tolmasky, M.E., Crosa, J.H. (1991) Molecular
characterization of the iron transport system mediated by the pJM1 plasmid in Vibrio
anguillarum 775. J. Biol. Chem. 266:23829-23833.
Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature. 227:680-685.
MacDonell M.T., Colwell R.R. (1985) Phylogeny of the Vibrionaceae, and recommendation for
two new genera, Listonella and Shewanella. Syst. Appl. Microbiol. 6:171-182.
Maluping, R., Ravelo, C., Lavilla-Pitogo, C., Krovacek, K., Romalde, J.L. 2005. Molecular
typing of Vibrio parahaemolyticus strains isolated from the Philippines by PCR-based
methods. J. Appl. Microbiol. 99: 383-391.
Marshall, S., Clark, C.G., Wang, G., Mulvey, M., Kelly, M.T., Johnson, W.M. 1999. Comparison of molecular methods for typing Vibrio parahaemolyticus. J. Clin. Microbiol. 37:
2473-2478.
Milton, D. L., O’Toole, R., Hšrstedt, P., Wolf-Watz, H. (1996) Flagellin A is essential for the
virulence of Vibrio anguillarum. J. Bacteriol. 178:1310-1319.

63
Mutharia LW, Raymond BT, Dekievit TR, Stevenson RMW (1993) Antibody specificities of
polyclonal rabbit and rainbow trout ant~seraa gainst Vibrio ordalii and serotype 0:2 strains
of Vjbrio anguillarum. Can J Microbiol 39: 492-499
Nagpal, M. L., Fox, K., Fox, A. (1998) Utility of 16S – 23S rRNA spacer region methodology:
how similar are interspace regions within a genome and between strains for closely related
organisms?. J. Microbiol. Meth. 33: 211 – 219
Norqvist, A., Hagstrom, A., Wolf-Watz, H. (1989) Protection of rainbow trout against vibriosis
and furunculosis by the use of attenuated strains of Vibrio anguillarum. Appl. Environ.
Microbiol. 55:1400-1405.
Ormonde, P., Hörstedt, P., O'Toole, R. & Milton, D. L. (2000). Role of motility in adherence to
and invasion of a fish cell line by Vibrio anguillarum. J Bacteriol 182: 2326–2328
O'Toole, R., Milton, D.L., Wolf-Watz, H. (1996) Chemotactic motility is required for invasion
of the host by the fish pathogen Vibrio anguillarum. Mol. Microbiol. 19:625-637.
Pazos, P., Santos, Y., Magariños, B., Bandín, I., Núñez, S., Toranzo, A.E.(1993). Phenotypic
characteristics and virulence of Vibrio anguillarum-related organisms. Appl. Environ.
Microbiol. 59:2969-2976.
Pedersen, K., Larsen, J.L. (1998) Characterization and typing methods for the fish pathogen
Vibrio anguillarum. Recent Res. Dev. Microbiol. 2:17-93.
Pedersen K., Grisez L., Houdt R., Tiainen T., Ollevier F., Larsen J.L. (1999) Extended
serotyping scheme for Vibrio anguillarum with the definition and characterization of
seven provisional O-serogroups. Curr. Microbiol. 38: 183-198.
Powel, J. L., Loutit M. W., (1990) Isolation and characterisation of Vibrio anguillarum from
selected marine sites in New Zealand. N. Z. J. Marine Fresh. Res. 24:267-273.
Ransom, D.L., Rohovec, C.N., Fryer, JL (1984) Comparison of histopathology caused by Vibrio
anguillarum and Vibrio ordalii in three species of Pacific salmon. J. Fish Dis. 7:107-115.

64
Riquelme, C, Hayashida, G, Toranzo, A.E, Vilches, J., Chávez, P. (1995). Pathogenicity studies
on a Vibrio anguillarum-related (VAR) strain causing an epizootic in Argopecten
purpuratus larvae cultured in Chile. Dis. Aquat. Org. 22:135-141.
Rivera, I.G., Chowdhury, M. A. R., Huq, A., Jacobs, D., Martins, M.T., Colwell, R. R. (1995b).
Enterobacterial repetitive intergenic consensus sequences and the PCR to generate
fingerprints of genomic DNAs from Vibrio cholerae O1, O139 and non-O1. Appl.
Environ. Microbiol. 61: 2898-2904.
Rodicio, M., Mendoza, M. (2004) Identificación bacteriana mediante secuenciación del ARNr
16S: Fundamento, metodología y aplicaciones en Microbiología clínica. Enferm. Infec. y
Microbiol. Clín. 22 (4): 238 – 245
Rodríguez, J., Lopéz-Romalde, S., Beaz, R., Alonso, C., Castro, D., Romalde, JL. (2006).
Molecular fingerprinting of Vibrio tapetis strains using three PCR-based methods: ERIC-
PCR, REP-PCR and RAPD. Dis. Aquat. Org. 69: 175-183.
Romalde, J.L., Ravelo, S., Lopez-Romalde, S., Avendaño-Herrera, R., Magariños, B., Toranzo
A.E. ( 2005). Vaccination strategies to prevent emerging diseases for Spanish aquaculture.
Dev. Biol. Basel Karger 121: 85-95.
Saitou, N, Nei M. (1987). The neighbor-joining method: a new method for reconstructing
phylogenetic trees. Mol. Biol. Evol. 4:406-425.
Salinas, P.C., Tolmasky, M.E., Crosa, J.H. (1989) Regulation of the iron uptake system in Vibrio
anguillarum: evidence for a cooperative effect between two transcriptional activators.
Proc. Natl. Acad. Sci. USA. 86:3529-3533.
SalmonChile. (2006). Exportaciones chilenas totales, millones de dólares FOB Chile. En:
http://www.salmonchile.cl/
Schembri, M.A., Givskov, M., Klemm, P. (2002) An attractive surface: gram-negative bacterial
biofilms. Sci STKE 2002: RE6.
Schiewe, M.H., Crosa, J.H., Ordal, E.J..(1977). Deoxyribonucleic acid relationships among
marine vibrios pathogenic to fish. Can. J. Microbiol. 23: 954-958.

65
Schiewe, M.H., Trust, T.J., Courtice, I.D., Khouri, A.G., Crosa, J.H. (1981) Serum resistance
and hemagglutination ability of marine vibrios pathogenic for fish. Infect. Immun. 34:702-
707.
Schiewe, M.H.N., Harrell, A.J. (1983) Vibriosis of salmonids. Dev. Aquaculture Fisheries Sci.
17:323-327.
Shangkuan, Y.H., Lin, H.C., Wang, T.M. 1997. Diversity of DNA sequences among Vibrio
cholerae O1 and non-O1 isolates detected by whole-cell repetitive element sequence-
based polymerase chain reaction. J. Appl. Microbiol. 82: 335-344.
Skov, M.N., Pedersen, K., Larsen, J.L. (1995) Comparison of pulse-field gel electrophoresis,
ribotyping and plasmid profiling for typing of Vibrio anguillarum serovar O1. Appl.
Environ. Microbiol. 61:1540-1545.
Sorensen, U.B., Larsen, J.L. (1986) Serotyping of Vibrio anguillarum. Appl. Environ. Microbiol.
51:593-597.
Stork, M., Di Lorenzo, M., Welch, T.J., Crosa, L.M. , Crosa, J.H. (2002) Plasmid-mediated iron
uptake and virulence in Vibrio anguillarum. Plasmid 48: 222-228.
Tenover F.C., Arbeit R.D., Goering R. V., Mickelsen P. A., Murray B. E., Persing D. H.,
Swaminathan B. (1995) Interpreting Chromosomal DNA Restriction Patterns Produced by
Pulsed-Field Gel Electrophoresis: Criteria for Bacterial Strain Typing. J. Clin. Microb. 33:
2233-2239.
Tiainen, R., Pedersen, K., Larsen, J.L. (1995). Ribotyping and plasmid profiling of Vibrio
anguillarum serovar O2 and Vibrio ordalii. J. Appl. Bacteriol. 79:384-392.
Toranzo, A.E., Barja, J.L. (1990) A review of the taxonomy and seroepizootiology of Vibrio
anguillarum, with special reference to aquaculture in the Northwest of Spain. Dis. Aquat.
Org. 9:73-82.
Toranzo, A.E., Baya, A., Roberson, B.S., Barja, J.L., Grimes, D.J., Hetrick F.M. (1987)
Specificity of slide agglutination test for detecting bacterial fish pathogens. Aquaculture
61:81-97.

66
Toranzo, A.E., Combarro, P., Lemos, M., Barja, J.L. (1984) Plasmid coding for transferable drug
resistance in bacteria isolated from cultured rainbow trout. Appl. Environ. Microbiol. 48:
872-877.
Toranzo, A.E., Magariños, B., Romalde, J.L. (2005) A review of the main bacterial fish diseases
in mariculture system. Aquaculture 246:37-61.
Towbin, H., Staehelin, T., Gordon, J. (1979) Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl.
Acad. Sci.76:4350-4354.
Toyama, T., Kita-Tsukamoto, K. and Wakabayashi, H. (1996) Identification of Flexibacter
maritimus, Flavobacterium branchiophilum and Cytophaga columnaris by PCR targeted
16S ribosomal DNA. Fish Pathology. 31:25-31.
Trust, T. J., Chart, H. (1984). Characterization of the surface antigens of the marine fish
pathogens Vibrio anguillarum and Vibrio ordalii. Can. J. Microbiol. 30:703–710.
Trust, T.J., Courtice, I.D., Khouri, A.G., Crosa, J.H., Schiewe, M.H. (1981) Serum resistance
and hemagglutination ability of marine vibrios pathogenic for fish. Infect. Immun. 34:702-
707.
Tsai, C.M., Frasch, C.E. (1982) A sensitive silver stain for detecting lipopolysaccharides in
polyacrylamide gels. Anal. Biochem. 119:115-119.
Valerie, I., Roberts, R., Bromage, N. (1993) Bacterial diseases on fish. Blackwell Scientific
Publications, London, UK.
Vera M.R. (2005) Caracterización Patológica (Microscópica, microscópica y clínica) de la
vibriosis por Vibrio ordalii en salmón del Atlántico (Salmo salar) en el sur de Chile. Tesis
para optar al Grado de Licenciado en Ciencias Veterinarias. Universidad Católica de
Temuco.
Vesalovic J., Koeuth T. and R. Lupski J., (1991). Distribution of repetitive DNA sequences in
eubacteria and application to fingerprinting of bacterial genomes. Nucl. Aci. Res. 19, Nº24
6823-6831.

67
Wards, B.J., Patel, H.H. (1991) Characterisation by restriction endonuclease analysis and
plasmid profiling of Vibrio ordalii strains from salmon (Oncorhynchus tshawytscha and
Oncorhynchus nerka) with vibriosis in New Zealand. J. Mar. Freshwater Res. 25:345-350.
Watnick, P., Kolter, R. (2000) Biofilm city of microbes. J. Bacteriol. 182:2675-2679.
Weisburg, W., Barns, S., Pelletier, D., Lane, D. (1991) 16S ribosomal DNA amplification for
phylogenetic study. J. Bacteriol. 173:697-703.
Wertheimer, A.M., Verweij, W., Chen, Q., Crosa, L.M., Nagasawa, M., Tolmasky, M.E., Actis,
L.A., Crosa, J.H. (1999) Characterization of the angR gene of Vibrio anguillarum:
essential role in virulence. Infect. Immun. 67: 6496-6509.
Wong, H.C., Lin, C.H. 2001. Evaluation of typing Vibrio parahaemolyticus by three PCR
methods using specific primers. J. Clin. Microbiol. 39: 4233-4240.
Zorrilla, I., Chabrillon, M., Arijo, S., Daz-Rosales, P., Martnez-Manzanares, E., Balebona, M.C.,
Morinigo, M.A. (2003) Bacteria recovered from diseased cultured gilthead sea bream
(Sparus aurata L.) in southwestern Spain. Aquaculture. 218:11-20.
Páginas web (URL):
http://www.aapqis.org/main/path/viewpath_tax.asp?PathID=208&sec=tax&action=
http://www.the-icsp.org/taxa/vibriolist.htm
http://www.lsc.usgs.gov/FHB/leaflets/77.asp

68
ANEXO 2: Principales características fenotípicas, fisiológicas y bioquímicas de las especies Listonella anguillarum y Vibrio ordalii (Schiewe y cols., 1981).
+, prueba positiva; -, prueba negativa; d, resultado débil. *, indica diferencias entre ambas especies bacterianas.
Carácter Listonella anguillarum Vibrio ordalii
Tinción Gram - -
Motilidad + +
Sensibilidad agente vibriostático O/129 + +
Fermentación glucosa sin /gases + +
Reducción de nitratos + +
Citrato * + -
Vogues Proskauer * + -
Rojo de metilo - -
OPNG * + -
Producción de:
Oxidasa + +
Catalasa + +
H2S - -
Utilización de:
Arginina-alcalina * + -
Lisina decarboxilasa - -
Ornitina decarboxilasa - -
Degradación de:
Hidrólisis de gelatina + +
Hidrólisis de almidón * + -
Ureasa - -
DNasa + +
Lipasa * + -
Adonitol - -
Arabinosa + -
Celobiosa * + -
Eritritol - -
Galactosa + d
Glicerol * + -
Inositol - -
Lactosa - -
Maltosa + +
Manitol + +
Melibiosa - -
Rafinosa - -
Ramnosa - -
Sorbitol * + -
Sucrosa + +
Trealosa * + -
Xilosa - -
Rango de salinidad (% NaCl)
0 - -
0,5 + +
3 + +
7 - -
10 - -
Temperatura (ºC)
15 + +
25 + +
37 * + -

69
ANEXO 2: Aislados bacterianos estudiados en la presente tesis, los cuales fueron
asociados a mortalidades de salmónidos entre octubre 2004 y agosto2005
Código VQ Lugar aislamiento Fecha de aislamiento Especie
1 Au2 Castro 07/04 Salmo salar
2 Au3 Cisne 15/09/04 s.i.
3 Au4 Isla Tranqui, Chiloe 07/03 S. salar
4 Au 6 Pto. Chacab 10/04 S. salar
5 Au 7 Pto. Chacab 10/04 S. salar
6 Au8 Quemchi 09/04 S. salar
7 Au 9 Bahía Colulí 11/04 S. salar
8 Au 10 Isla Puluqui 01/04 S. salar 9 PF2 Chiloe Sur 08/03/05 S. salar
10 PF3 Chiloe Sur 08/03/05 Oncorhynchus kisutch
11 PF4 Castro 08/03/05 S. salar
12 PF5 Chiloe Sur 04/03/05 O. kisutch
13 PF6 Chiloe Sur 04/03/05 O. kisutch
14 PF7 Valdivia 19/02/05 S. salar
15 PF8 Castro 25/02/05 S. salar
16 PF9 Puntilla 19/02/05 Oncorhynchus mykiss
17 PF10 Valdivia 19/02/05 S. salar
18 PF11 Quenak 20/01/05 O. kisutch
19 PF12 Puntilla 19/02/05 O. mykiss
20 PF16 Pta Diamante 19/02/05 O. mykiss 21 PF17 Centinela/Pto. Montt 20/01/05 O. kisutch
23 PF28 Castro 02/03/05 S. salar
24 PF32 Castro 11/02/05 S. salar
25 PF42 Aysen s.i. O. mykiss
26 PF43 Castro 02/03/05 S. salar
27 PF44 Castro 26/02/05 S. salar
28 PF48 Nieves (Continente) 17/03/05 S. salar
29 PF49 Loncura/Quellón 16/02/05 S. salar
30 PF50 Bobe/Continente 17/03/05 S. salar
31 PF52 Nieves 17/03/05 S. salar
32 PF53 Nieves 17/03/05 S. salar 33 PF54 Leutepú/Queilén 17/03/05 S. salar
34 PF55 Ceres/Continente 17/03/05 S. salar
35 PF57 Bobe/Continente 17/03/05 S. salar
36 PF70 Estuario de Reloncaví/Ralún 07/01/05 S. salar
37 PF71 Calbuco/Abato 18/01/05 S. salar
38 PF72 Calbuco/Pilolcura 28/01/05 S. salar
39 PF73 Huar/Pto. Montt 08/02/05 S. salar
70
Continuación anexo 2.
s.i.: sin información
Nota: tres aislados fueron descartados por no pertenecer a agentes causales de vibriosis (PF18,
PF68 y PF88 ó 22, 52 y 54 del orden correlativo de la tabla), estos correspondieron a especies
del genero Aeromonas.
Código VQ Lugar aislamiento Fecha de aislamiento Especie
40 PF74 Calbuco 06/02/04 S. salar
41 PF75 Chillidque/Chiloe Sur 07/03/05 S. salar
42 PF76 Seno Reloncaví/Quillaipe 09/04/05 S. salar
43 PF77 Seno Reloncaví/Quillaipe 10/04/05 S. salar
44 PF78 Chaiten 10/04/05 S. salar
45 PF79 Chaiten 10/04/05 S. salar
46 PF80 Calbuco 11/04/05 S. salar 47 PF81 Calbuco 16/04/05 S. salar
48 PF82 Ralun del Este 16/04/05 S. salar
49 PF83 Seno Reloncaví/Quillaipe 16/04/05 S. salar
50 PF84 Seno Reloncaví/Quillaipe 17/04/05 S. salar
51 PF85 Isla Huar 01/05/05 S. salar
53 PF87 Fiordo Comau/Hornopiren 18/09/04 S. salar
55 PF89 Calbuco/Pilolcura 05/11/04 S. salar
56 PF90 Calbuco/Pilolcura 06/11/04 S. salar
57 PF91 Estuario de Reloncaví 08/11/04 S. salar
58 PF92 Chiloe Sur/Quetén 17/11/04 S. salar
59 PF93 Estuario de Reloncaví/Ralún 29/11/04 S. salar 60 PF94 Chiloe Sur/Chelín 01/12/04 S. salar
61 PF95 Calbuco 01/12/04 S. salar
62 PF96 Estuario de Reloncaví/Ralún 09/12/04 S. salar
63 PF97 Calbuco/Aptao 17/12/04 S. salar
64 PF98 Fiordo Comau/Hornopirén 17/12/04 O. mykiss
65 PF99 Calbuco/Huenquillahue 21/12/04 S. salar
66 PF158 Islote Quemado 06/04 S. salar
67 PF159 Ling I 10/04 S. salar
68 PF160 Ling I 10/04 S. salar
69 PF161 Renaico 10/04 S. salar
70 PF162 Chaculay 10/04 S. salar 71 PF181 Islote Quemado 05/05 S. salar
72 PF182 Puyuhuapi 10/04 S. salar
73 PF183 Pucheguin 26/07/05 S. salar
74 PF184 Leutepú 22/02/05 O. kisutch
75 PF185 Pta. Chequian 24/02/05 O. kisutch
76 PF186 08/05 S. salar
77 PF187 08/05 S. salar


















