
B.Sc. Microbiology/Biotech II Cell biology and Genetics Unit 5 microbial genetics
69
Linkage, crossing over and basic microbial genetics Cell biology and Genetics Unit – 5
-
Upload
rai-university -
Category
Science
-
view
112 -
download
3
Transcript of B.Sc. Microbiology/Biotech II Cell biology and Genetics Unit 5 microbial genetics

Linkage, crossing over and basic microbial genetics
Cell biology and Genetics
Unit – 5

Mapping of gene
• Genome mapping, is the creation of a genetic map assigning DNA fragments to chromosomes.
• Genetic linkage is the tendency of alleles that are located close together on a chromosome to be inherited together during meiosis. Genes whose loci are nearer to each other are less likely to be separated onto different chromatids during chromosomal crossover, and are therefore said to be genetically linked. In other words, the nearer two genes are on a chromosome, the lower is the chance of a swap occurring between them, and the more likely they are to be inherited together.

Linkage map
• A linkage map is a genetic map of a species or experimental population that shows the position of its known genes or genetic markers relative to each other in terms of recombination frequency, rather than a specific physical distance along each chromosome.
• Linkage mapping is critical for identifying the location of genes that cause genetic diseases.
• A genetic map is a map based on the frequencies of recombination between markers during crossover of homologous chromosomes.

• The greater the frequency of recombination(segregation) between two genetic markers, the furtherapart they are assumed to be. Conversely, the lower thefrequency of recombination between the markers, thesmaller the physical distance between them.
• Historically, the markers originally used were detectablephenotypes (enzyme production, eye color) derivedfrom coding DNA sequences; eventually, confirmed orassumed non coding DNA sequences such asmicrosatellites or those generating restriction fragmentlength polymorphisms (RFLPs) have been used.
• Genetic maps help researchers to locate other markers,such as other genes by testing for genetic linkage of thealready known markers.
• A genetic map is not a physical map (such as a radiationreduced hybrid map) or gene map.

Genome sequencing
• Genome sequencing are sometimesmistakenly referred to as "genome mapping"by non-biologists. While the scope, purposeand process are totally different, a genomeassembly can be viewed as the "ultimate"form of physical map, in that it provides allinformation that a traditional physical mapcan offer in a much better way.

Gene mapping• The essence of all genome mapping is to place a collection of molecular
markers onto their respective positions on the genome.
• Molecular markers come in all forms. Genes can be viewed as one specialtype of genetic markers in the construction of genome maps, and mappedthe same way as any other markers.
• In genetic mapping, any sequence feature that can be faithfullydistinguished from the two parents can be used as a genetic marker.Genes, in this regard, are represented by "traits" that can be faithfullydistinguished between two parents.
• In physical mapping, there are no direct ways of marking up a specificgene since the mapping do not include any information that concern traitsand functions. Genetic markers can be linked to a physical map byprocesses in situ hybridization. By this approach, physical map contigs canbe "anchored" onto a genetic map.

Physical Mapping
• In physical mapping, the DNA is cut by a restriction enzyme. Once cut, the DNA fragments are separated by electrophoresis. The resulting pattern of DNA migration (i.e., its genetic fingerprint) is used to identify what stretch of DNA is in the clone. By analyzing the fingerprints, contigs are assembled by automated (FPC) or manual means (Pathfinders) into overlapping DNA stretches. Now a good choice of clones can be made to efficiently sequence the clones to determine the DNA sequence of the organism under study (seed picking).
• Macrorestriction is a type of physical mapping wherein the high molecular weight DNA is digested with a restriction enzyme having a low number of restriction sites.
• There are alternative ways to determine how DNA in a group of clones overlap without completely sequencing the clones. Once the map is determined, the clones can be used as a resource to efficiently contain large stretches of the genome. This type of mapping is more accurate than genetic maps.

Recombination
• The exchange of DNA sequences between different molecules, occurring either naturally or as a result of DNA manipulation.
• A process by which pieces of DNA are broken and recombined to produce new combinations of alleles.

What is gene cloning?
1 A fragment of DNA, containing the gene to be cloned, is inserted into a circular DNA molecule called a vector, to produce a recombinant DNA molecule.
2 The vector transports the gene into a host cell, which is usually a bacterium, although other types of living cell can be used.
3 Within the host cell the vector multiplies, producing numerous identical copies, not only of itself but also of the gene that it carries.

4 When the host cell divides, copies of the recombinant DNA molecule are passed to the progeny and further vector replication takes place.
5 After a large number of cell divisions, a colony, or clone, of identical host cells is produced. Each cell in the clone contains one or more copies of the recombinant DNA molecule; the gene carried by the recombinant molecule is now said to be cloned.
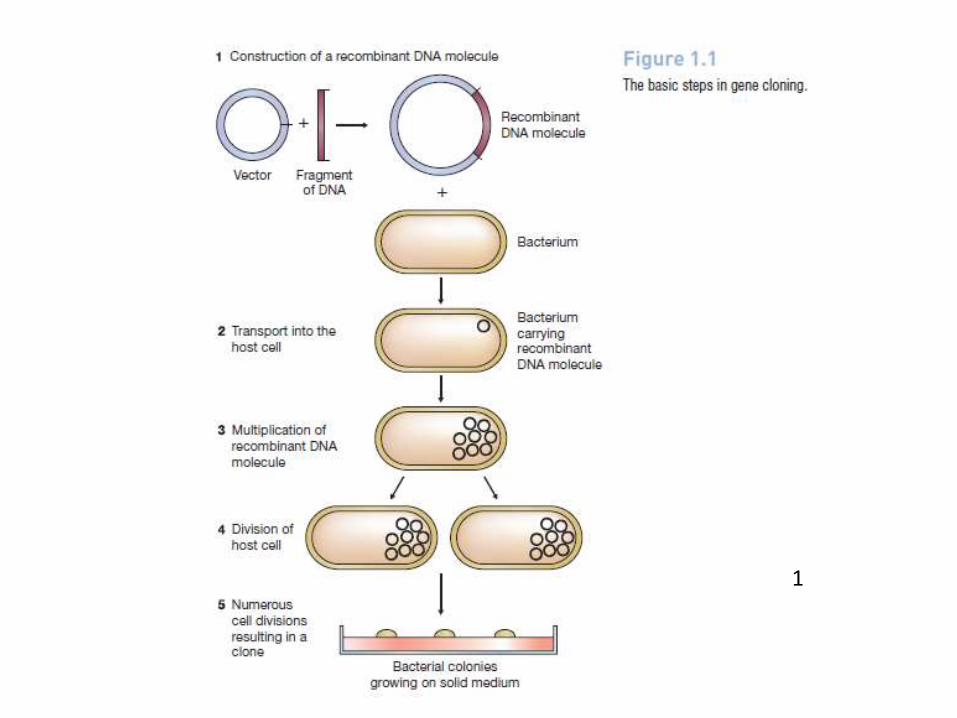
1

Crossing Over
• (i) Crossing over is a recombination of genes due to exchange of genetic material between two homologous chromosomes,
• (ii) It is the mutual exchange of segments of genetic material between non-sister chromatids of two homologous chromosomes, so as to produce re-combinations or new combinations of genes.
• The non-sister chromatids in which exchange of segments have occurred are called recombinants or cross-overs while the other chromatids in which crossing over has not taken place are known as parental chromatids or non cross-overs.

• Each parent cell has pairs of homologous chromosomes, one homolog from the father and one from the mother. In meiosis, the maternal and paternal chromosomes can be shuffled into the daughter cells in many different combinations (in humans there are 223 possible combinations!).

• This ensures genetic variation in sexually reproducing organisms. Further genetic variation comes from crossing over, which may occur during prophase I of meiosis.
• In prophase I of meiosis, the replicated homologous pair of chromosomes comes together in the process called synapsis, and sections of the chromosomes are exchanged. You can see that after crossing over, the resultant chromosomes are neither entirely maternal nor entirely paternal, but contain genes from both parents. Synapsis and crossing over occur only in meiosis.

Bacterial Sexual Processes
• Eukaryotes have the processes of meiosis to reduce diploids to haploidy, and fertilization to return the cells to the diploid state. Bacterial sexual processes are not so regular. However, they serve the same aim: to mix the genes from two different organisms together.
• The three bacterial sexual processes:
– 1. conjugation: direct transfer of DNA from one bacterial cell to another.
– 2. transduction: use of a bacteriophage (bacterial virus) to transfer DNA between cells.
– 3. transformation: naked DNA is taken up from the environment by bacterial cells.
15

Transformation
• The essence of recombinantDNA technology is to removeDNA from cells, manipulate itin the test tube, then put itback into living cells. In mostcases this is done bytransformation.
• In the case of E. coli, cells aremade “competent” to betransformed by treatmentwith calcium ions and heatshock. E. coli cells in thiscondition readily pick up DNAfrom their surroundings andincorporate it into theirgenomes.
16
2

Gene Transfer by Transformation
•Transformation is the process of importing free DNA into bacterial cells.
• the cells need to be competent.
• Many cells are capable of natural Transformation and naturally competent.
• others require artificial manipulations.
• Perturbing the membrane by chemical (CaCl2) or electrical (electroporation) methods
• Not all bacteria can take up free or naked DNA (<1%).
17

Gram positive Bacteria transform DNA using aTransformasome complex
Gram-negative bacteria do not use transformasomes
18
3

Conjugation
• Physical contact between two bacteria, usually associated with transfer of DNA from one cell to the other.

Conjugation
• Conjugation is the closest analogue in bacteria toeukaryotic sex.
• The ability to conjugate is conferred by the Fplasmid. A plasmid is a small circle of DNA thatreplicates independently of the chromosome.Bacterial cells that contain an F plasmid are called“F+”. Bacteria that don’t have an F plasmid arecalled “F-”.
• F+ cells grow special tubes called “sex pilli” fromtheir bodies. When an F+ cell bumps into an F-cell, the sex pilli hold them together, and a copy ofthe F plasmid is transferred from the F+ to the F-.Now both cells are F+.
20
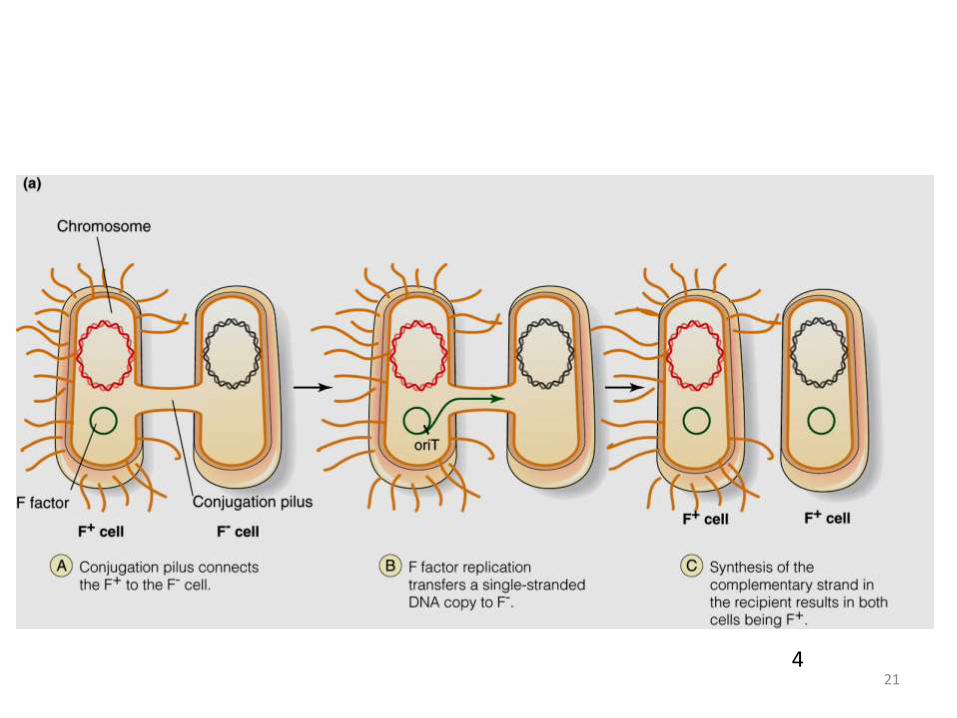
214

Hfr Conjugation
• When it exists as a free plasmid, the F plasmid can only transfer itself. This isn’t all that useful for genetics.
• However, sometimes the F plasmid can become incorporated into the bacterial chromosome, by a crossover between the F plasmid and the chromosome. The resulting bacterial cell is called an “Hfr”, which stands for “High frequency of recombination”.
• Hfr bacteria conjugate just like F+ do, but they drag a copy of the entire chromosome into the F-cell.
• Technique used in bacterial gene mapping
22

Intracellular Events in Conjugation
• The piece of chromosome that enters the F- form the Hfr is linear. It is called the “exogenote”.
• The F- cell’s own chromosome is circular. It is called the “endogenote”.
• Only circular DNA replicates in bacteria, so genes on the exogenote must be transferred to the endogenote for the F-to propagate them.
• This is done by recombination: 2 crossovers between homologous regions of the exogenote and the endogenote. In the absence of recombination, conjugation in ineffective: the exogenote enters the F-, but all the genes on it are lost as the bacterial cell reproduces.
23

F-prime (F’)
• The process of making an Hfr from an F+ involves a crossover between theF plasmid and the chromosome. This process is reversible: an Hfr canrevert to being F+ when the F plasmid DNA incorporated into the Hfrchromosome has a crossover and loops out of the chromosome formingan F plasmid once again.
• Sometimes the looping-out and crossing-over process doesn’t happen atthe proper place. When this happens, a piece of the bacterialchromosome can become incorporated into the F plasmid. This is calledan F’ (F-prime) plasmid.
• F’ plasmids can be transferred by conjugation. Conjugation with an F’ (or aregular F plasmid) is much faster and more efficient than with an Hfr,because only a very small piece of DNA is transferred. Since the F’ carriesa bacterial gene, this allele can be rapidly moved into a large number ofother strains. This permits its function to be tested rapidly. Also, tests ofdominance can be done.
• A cell containing an F’ is “merodiploid”: part diploid and part haploid. It isdiploid for the bacterial gene carried by the F’ (one copy on the F’ and theother on the chromosome), and haploid for all other genes.
24

Transduction
• Transduction is the process of moving bacterial DNA from one cell to another using a bacteriophage.
• Bacteriophage or just “phage” are bacterial viruses. They consist of a small piece of DNA inside a protein coat. The protein coat binds to the bacterial surface, then injects the phage DNA. The phage DNA then takes over the cell’s machinery and replicates many virus particles.
• Two forms of transduction:– 1. generalized: any piece of the bacterial genome can be transferred
– 2. specialized: only specific pieces of the chromosome can be transferred.
25

General Phage Life Cycle
• 1. Phage attaches to the cell and injects its DNA.
• 2. Phage DNA replicates, and is transcribed into RNA, then translated into new phage proteins.
• 3. New phage particles are assembled.
• 4. Cell is lysed, releasing about 200 new phage particles.
• Total time = about 15 minutes.
26

EM of Bacteriophages
27

Generalized Transduction
• Some phages, such as phage P1, break up the bacterial chromosome into small pieces, and then package it into some phage particles instead of their own DNA.
• These chromosomal pieces are quite small.• A phage containing E. coli DNA can infect a fresh host,
because the binding to the cell surface and injection of DNA is caused by the phage proteins.
• After infection by such a phage, the cell contains an exogenote (linear DNA injected by the phage) and an endogenote (circular DNA that is the host’s chromosome).
• A double crossover event puts the exogenote’s genes onto the chromosome, allowing them to be propagated.
28

Transduction Mapping
• Only a small amount of chromosome, a few genes, can be transferred by transduction. The closer 2 genes are to each other, the more likely they are to be transduced by the same phage. Thus, “co-transduction frequency” is the key parameter used in mapping genes by transduction.
• Transduction mapping is for fine-scale mapping only. Conjugation mapping is used for mapping the major features of the entire chromosome.
29

Mapping Experiment
• Important point: the closer 2 genes are to each other, the higher the co-transduction frequency.
• We are just trying to get the order of the genes here, not put actual distances on the map.
• Expt: donor strain is aziR leu+ thr+. Phage P1 is grown on the donor strain, and then the resulting phage are mixed with the recipient strain: aziS leu- thr-. The bacteria that survive are then tested for various markers
• 1. Of the leu+ cells, 50% are aziR, and 2% are thr+. From this we can conclude that azi and leu are near each other, and that leu and thr are far apart.
• But: what is the order: leu--azi--thr, or azi--leu--thr ?
30

Mapping Experiment, pt. 2
• 2. Do a second experiment to determine the order. Select the thr+ cells, then determine how many of them have the other 2 markers. 3% are also leu+ and 0% are also aziR.
• By this we can see that thr is closer to leu than it is to azi, because thr and azi are so far apart that they are never co-transduced.
• Thus the order must be thr--leu--azi.• Note that the co-transduction frequency for thr and
leu are slightly different for the 2 experiments: 2% and 3%. This is attributable to experimental error.
31

Intro to Specialized Transduction
• Some phages can transfer only particular genes to other bacteria.
• Phage lambda (λ) has this property. To understand specialized transduction, we need to examine the phage lambda life cycle.
• lambda has 2 distinct phases of its life cycle. The “lytic” phase is the same as we saw with the general phage life cycle: the phage infects the cell, makes more copies of itself, then lyses the cell to release the new phage.
32

Lysogenic Phase
• The “lysogenic” phase of the lambda life cycle starts the same way: thelambda phage binds to the bacterial cell and injects its DNA. Onceinside the cell, the lambda DNA circularizes, then incorporates into thebacterial chromosome by a crossover, similar to the conversion of an Fplasmid into an Hfr.
• Once incorporated into the chromosome, the lambda DNA becomesquiescent: its genes are not expressed and it remains a passiveelement on the chromosome, being replicated along with the rest ofthe chromosome. The lambda DNA in this condition is called the“prophage”.
• After many generations of the cell, conditions might get harsh. Forlambda, bad conditions are signaled when DNA damage occurs.
• When the lambda prophage receives the DNA damage signal, it loopsout and has a crossover, removing itself from the chromosome. Thenthe lambda genes become active and it goes into the lytic phase,reproducing itself, then lysing the cell.
33

Specialized Transduction
• Unlike the F plasmid that can incorporate anywhere in the E. coli genome, lambda can only incorporate into a specific site, called attλ. The gal gene is on one side of attλ and the bio gene (biotin synthesis) is on the other side.
• Sometimes when lambda come out of the chromosome at the end of the lysogenic phase, it crosses over at the wrong point. This is very similar to the production of an F’ from an Hfr.
• When this happens, a piece of the E. coli chromosome is incorporated into the lambda phage chromosome
• These phage that carry an E. coli gene in addition to the lambda genes are called “specialized transducing phages”. They can carry either the gal gene or the bio gene to other E. coli.
• Thus it is possible to quickly develop merodiploids (partial diploids) for any allele you like of gal or bio. Note that this trick can’t be used with other genes, but only for genes that flank the attachment site for lambda or another lysogenic phage.
34

Specialized Transduction
35
5

POPULATION GENETICS:
The study of the rules governing the maintenance and transmission of genetic variation in natural populations.

DARWINIAN EVOLUTION BY NATURAL SELECTION
Many more individuals are born than survive (COMPETITION).
Individuals within species are variable (VARIATION).
Some of these variations are passed on to offspring (HERITABILITY).
Survival and reproduction are not random. There must be a correlation between fitness and phenotype.

Gregor Mendel
The “rediscovery” of Mendel’s genetic studies in 1902 by William Bateson completed the missing model for the inheritance of genetic factors.
Mendel published his work in the Transactions of the Brunn Society of Natural History in 1866.

SEXUAL REPRODUCTION CONTRIBUTES TO VARIATION
Example – A Line Cross Experiment
Consider 2 diploid individuals with 3 loci and 2 alleles,
Parents: aabbcc x AABBCC
F1 progeny: AaBbCc
F2 progeny:AABBCC AABBCc AABBccAABbCC AABbCc AABbccAAbbCC AAbbCc AAbbccAaBBCC AaBBCc AaBBccAaBbCC AaBbCc AaBbccAabbCC AabbCc AabbccaaBBCC aaBBCc aaBBccaaBbCC aaBbCc aaBbccaabbCC aabbCc aabbcc
27 COMBINIATIONS

Mechanisms of Evolution: Mendelian Genetics in Populations
Genetic variation is the raw material of evolutionary change: how do we measure it?
What are the forces that cause genetic changes within populations? That is, what mechanisms cause evolutionary change?

Population Genetics
Evolution can be defined as a change in gene frequencies through time.
Population genetics tracks the fate, across generations, of Mendelian genes in populations.
Population genetics is concerned with whether a particular allele or genotype will become more or less common over time, and WHY.

A few things to keep in mind as we take an excursion into population genetic theory:
“Make things as simple as possible, but no simpler.”---Einstein
“All models are wrong, some are useful.”---Box
“No theory should fit all the facts because some of the facts are wrong.”---Bohr

Some Definitions:
Population: A freely interbreeding group of individuals.
Gene Pool: The sum total of genetic information present in a population at any given point in time.
Phenotype: A morphological, physiological, biochemical, or behavioral characteristic of an individual organism.
Genotype: The genetic constitution of an individual organism.
Locus: A site on a chromosome, or the gene that occupies the site.
Gene: A nucleic acid sequence that encodes a product with a distinct function in the organism.
Allele: A particular form of a gene.
Gene (Allele) Frequency: The relative proportion of a particular allele at a single locus in a population (a number between 0 and 1).
Genotype Frequency: The relative proportion of a particular genotype in a population (a number between 0 and 1).

The Gene Pool
•Members of a species can interbreed & produce fertile offspring
•Species have a shared gene pool
•Gene pool – all of the alleles of all individuals in a population
44
2

The Gene Pool
•Different species do NOT exchange genes by interbreeding
•Different species that interbreed often produce sterile or less viable offspring e.g. Mule
45
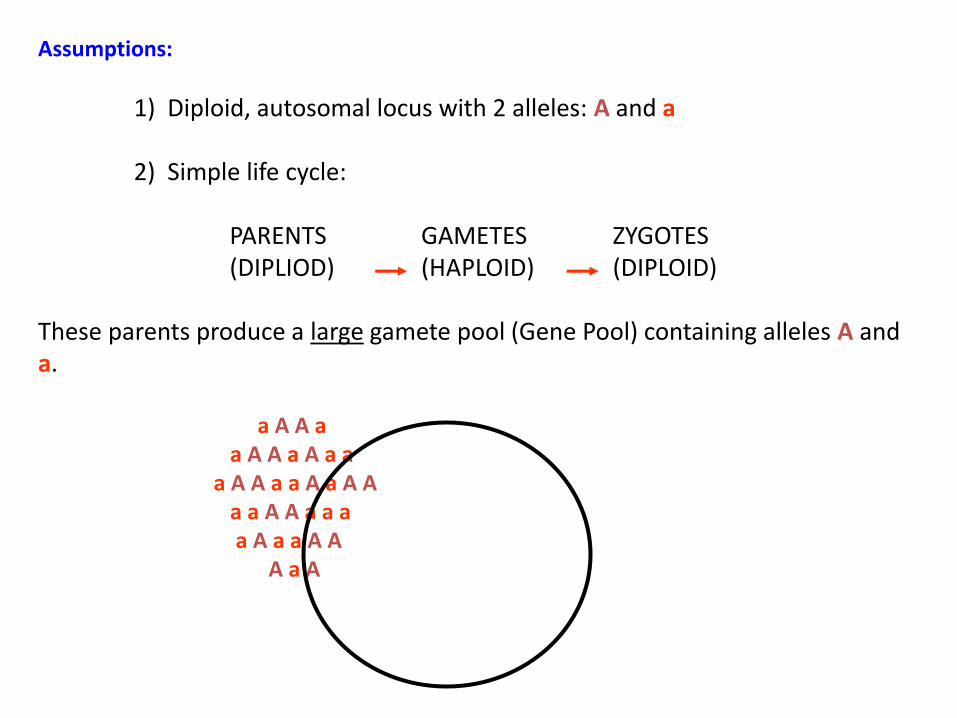
Assumptions:
1) Diploid, autosomal locus with 2 alleles: A and a
2) Simple life cycle:
PARENTS GAMETES ZYGOTES(DIPLIOD) (HAPLOID) (DIPLOID)
These parents produce a large gamete pool (Gene Pool) containing alleles A and a.
a A A aa A A a A a a
a A A a a A a A Aa a A A a a aa A a a A A
A a A

Gamete (allele) Frequencies:
Freq(A) = pFreq(a) = q
p + q = 1
Genotype Frequencies of 3 Possible Zygotes:
AA Aa aa
Freq (AA) = pA x pA = pA2
Freq (Aa) = (pA x qa) + (qa x pA) = 2pAqa
Freq (aa) = qa x qa = qa2
p2 + 2pq + q2 = 1

General Rule for Estimating Allele Frequencies from Genotype Frequencies:
Genotypes: AA Aa aa
Frequency: p2 2pq q2
Frequency of the A allele:
p = p2 + ½ (2pq)
Frequency of the a allele:
q = q2 + ½ (2pq)

Sample Calculation: Allele Frequencies
Assume N = 200 indiv. in each of two populations 1 & 2
Pop 1 : 90 AA 40 Aa 70 aa Pop 2 : 45 AA 130 Aa 25 aa
In Pop 1 :
p = p2 + ½ (2pq) = 90/200 + ½ (40/200) = 0.45 + 0.10 = 0.55 q = q2 + ½ (2pq) = 70/200 + ½ (40/200) = 0.35 + 0.10 = 0.45
In Pop 2 :
p = p2 + ½ (2pq) = 45/200 + ½ (130/200) = 0.225 + 0.325 = 0.55 q = q2 + ½ (2pq) = 25/200 + ½ (130/200) = 0.125 + 0.325 = 0.45

Main Points:
p + q = 1 (more generally, the sum of the allele frequencies equals one)
p2 + 2pq +q2 = 1 (more generally, the sum of the genotype frequencies equals one)
Two populations with markedly different genotype frequencies can have the same allele frequencies

Populations
•A group of the same species living in an area
•No two individuals are exactly alike (variations)
•More Fit individuals survive & pass on their traits
51

Speciation
•Formation of new species
•One species may splitinto 2 or more species
•A species may evolve into a new species
•Requires very long periods of time
52

Modern Evolutionary Thought

Modern Synthesis Theory
Combines Darwinian selection and Mendelian inheritance
Population genetics - study of genetic variation within a population
Emphasis on quantitative characters (height, size …)
54

Modern Synthesis Theory 1940s – comprehensive theory
of evolution (Modern Synthesis Theory)
Introduced by Fisher & Wright
Until then, many did not accept that Darwin’s theory of natural selection could drive evolution
55
S. Wright
A. Fisher

Modern Synthesis Theory
• TODAY’S theory on evolution
Recognizes that GENES are responsible for the inheritance of characteristics
Recognizes that POPULATIONS, not individuals, evolve due to natural selection & genetic drift
Recognizes that SPECIATION usually is due to the gradual accumulation of small genetic changes
56

Microevolution
• Changes occur in gene pools due to mutation, natural selection, genetic drift, etc.
• Gene pool changes cause more VARIATION in individuals in the population
• This process is called MICROEVOLUTION
• Example: Bacteria becoming unaffected by antibiotics (resistant)
57

Hardy-Weinberg Principle

The Hardy-Castle-Weinberg Law
A single generation of random mating establishes H-W equilibrium genotype frequencies, and neither these frequencies nor the gene frequencies will change in subsequent generations.
Hardy
p2 + 2pq + q2 = 1

The Hardy-Weinberg Principle
• Used to describe a non-evolving population.
• Shuffling of alleles by meiosis and random fertilization have no effect on the overall gene pool.
• Natural populations are NOT expected to actually be in Hardy-Weinberg equilibrium.
60

The Hardy-Weinberg Principle
• Deviation from Hardy-Weinberg equilibrium usually results in evolution
• Understanding a non-evolving population, helps us to understand how evolution occurs
61

5 Assumptions of the H-W Principle
1. Large population size- small populations have fluctuations in allele
frequencies (e.g., fire, storm).
2. No migration- immigrants can change the frequency of an allele by bringing in new alleles to a population.
3. No net mutations- if alleles change from one to another, this will change the frequency of those alleles
62

5 Assumptions of the H-W Principle
4. Random mating- if certain traits are more desirable, then individuals with those traits will be selected and this will not allow for random mixing of alleles.
5. No natural selection- if some individuals survive and reproduce at a higher rate than others, then their offspring will carry those genes and the frequency will change for the next generation.
63

The Hardy-Weinberg PrincipleThe gene pool of a NON-EVOLVING population remains
CONSTANT over multiple generations (allele frequency doesn’t change)
The Hardy-Weinberg Equation:
1.0 = p2 + 2pq + q2
Where:
p2 = frequency of AA genotype
2pq = frequency of Aa
q2 = frequency of aa genotype64

The Hardy-Weinberg Principle
Determining the Allele Frequency using Hardy-Weinberg:
1.0 = p + q
Where:
p = frequency of A allele
q = frequency of a allele
65

66
Allele Frequencies Define Gene Pools
As there are 1000 copies of the genes for color, the allele frequencies are (in both males and females):
320 x 2 (RR) + 160 x 1 (Rr) = 800 R; 800/1000 = 0.8 (80%) R160 x 1 (Rr) + 20 x 2 (rr) = 200 r; 200/1000 = 0.2 (20%) r
500 flowering plants
480 red flowers 20 white flowers
320 RR 160 Rr 20 rr

IMPLICATIONS OF THE H-W PRINCIPLE:
1) A random mating population with no external forces acting on it will reach the equilibrium H-W frequencies in a single generation, and these frequencies remain constant there after.
2) Any perturbation of the gene frequencies leads to a new equilibrium after random mating.
3) The amount of heterozygosity is maximized when the gene frequencies are intermediate.
2pq has a maximum value of 0.5 when p = q = 0.5

FOUR PRIMARY USES OF THE H-W PRINCIPLE:
1) Enables us to compute genotype frequencies from generation to generation, even with selection.
2) Serves as a null model in tests for natural selection, nonrandom mating, etc., by comparing observed to expected genotype frequencies.
3) Forensic analysis.
4) Expected heterozygosity provides a useful means of summarizing the molecular genetic diversity in natural populations.

References
• Images references:
• 1-6- Gene cloning and DNA analysis by TA Brown
• Reading references:
• Gene cloning and DNA analysis by TA Brown
1


















