
Breeding for Different Flower Forms in Ornamental Crops
38
Indian Agricultural Research Institute, New Delhi Breeding for Different Flower Forms in Ornamental Crops Abhay Kumar Gaurav 10459 Ph. D. 1 st Year Seminar: FLA 691 Division of Floriculture and Landscaping Indian Agricultural Research Institute, New Delhi
-
Upload
abhay-kumar-gaurav -
Category
Education
-
view
66 -
download
3
Transcript of Breeding for Different Flower Forms in Ornamental Crops

Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
Breeding for Different Flower Forms in Ornamental Crops
Abhay Kumar Gaurav10459
Ph. D. 1st Year
Seminar: FLA 691
Division of Floriculture and LandscapingIndian Agricultural Research Institute, New Delhi

Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
Flower shape/form is one of the most important characteristics
in ornamental plants
Creation of new flower shapes in ornamental plants is a major
breeding target
Phenotype with unique forms of flower or, the double flower
has higher ornamental value than the single one
Key transcriptional factors for the identification of floral organs
have been clarified by analyzing model plants (Arabidopsis)
INTRODUCTION
Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
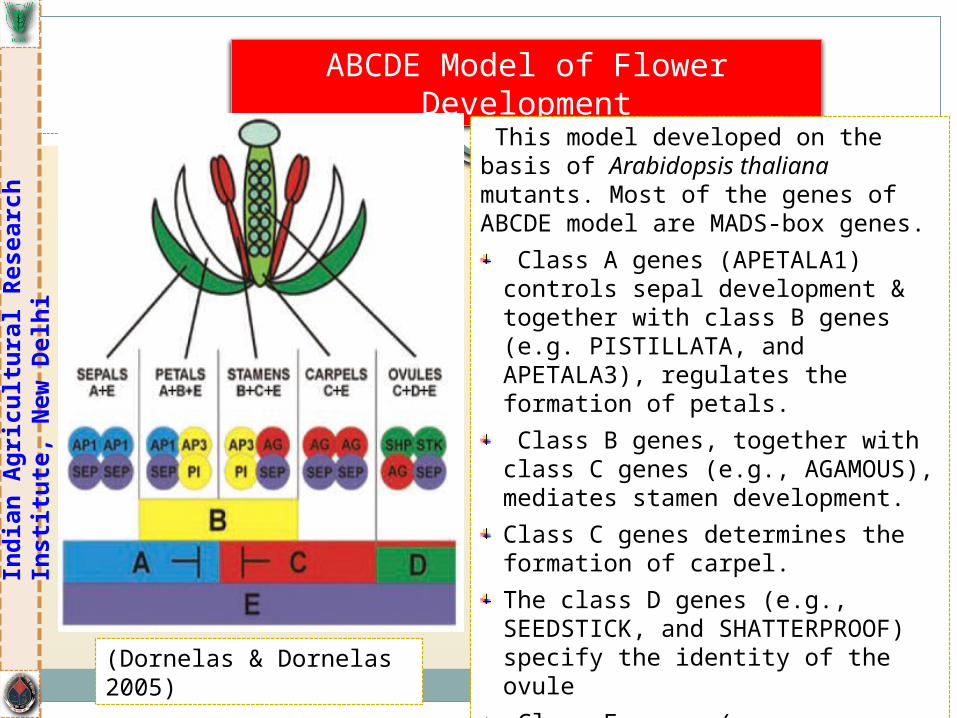
ABCDE Model of Flower Development
This model developed on the basis of Arabidopsis thaliana mutants. Most of the genes of ABCDE model are MADS-box genes.
Class A genes (APETALA1) controls sepal development & together with class B genes (e.g. PISTILLATA, and APETALA3), regulates the formation of petals.
Class B genes, together with class C genes (e.g., AGAMOUS), mediates stamen development.
Class C genes determines the formation of carpel.
The class D genes (e.g., SEEDSTICK, and SHATTERPROOF) specify the identity of the ovule
Class E genes (e.g., SEPALLATA), expressed in the entire floral meristem, & are necessary
(Dornelas & Dornelas 2005)

Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
c)
Sepals

Hybridization
Mutation
Polyploidy
Genetic Modification
RNAi or Gene silencing
Chimeric REpressor gene-Silencing Technology (CRES-T)
Micro RNA
Breeding Methods
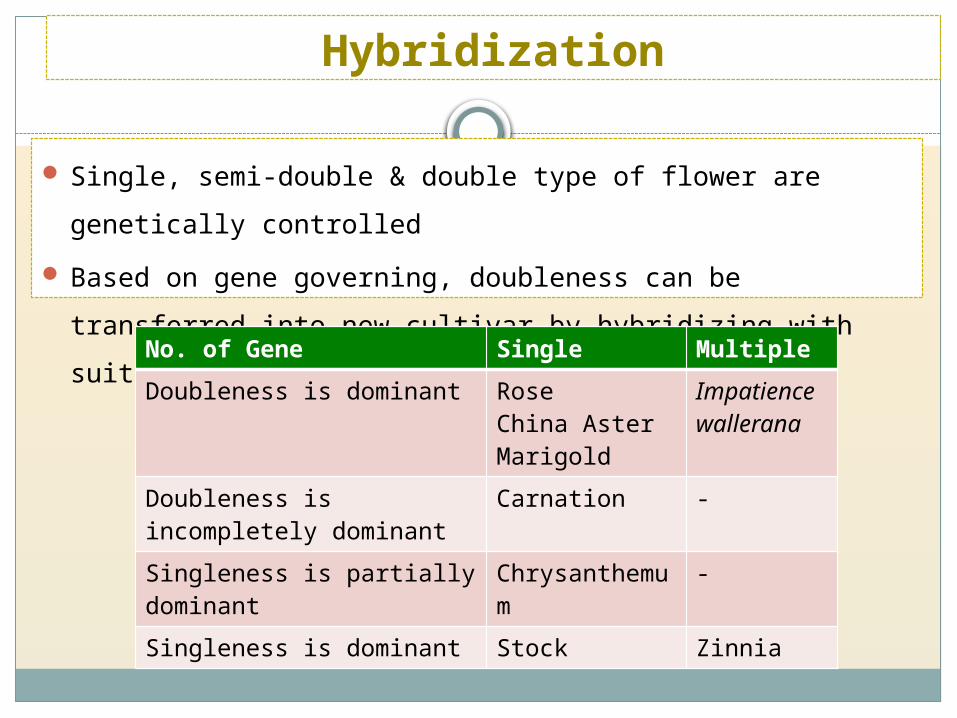
Single, semi-double & double type of flower are genetically controlled
Based on gene governing, doubleness can be transferred into new
cultivar by hybridizing with suitable parent
Hybridization
No. of Gene Single Multiple
Doubleness is dominant RoseChina AsterMarigold
Impatience wallerana
Doubleness is incompletely dominant
Carnation -
Singleness is partially dominant
Chrysanthemum
-
Singleness is dominant Stock Zinnia


Mutation BreedingIn
dia
n A
gri
cult
ura
l R
esea
rch
In
stit
ute
, N
ew D
elh
i
c)
The morphology of flowers & inflorescences can be affected by mutation
Mutation induction led to changes in flower size, petal shape, petal numbers
In Compositae, an increase in whorls of ligulate florets as well as a conversion from ligulate into tubular florets was described

Carnation varieties co-developed by Kirin Agribio and the JAEA using ion beams. The flower on the upper-left corner is the parent(var. .Vital.) and the others are mutants. Tanaka et al., 2010

Polyploidy Polyploidy breeding : Effective
method to double the chromosome number
Genetic variations created can be further used in breeding
Main consequences of induced polyploidy are increase in size and shape of plants/leaves/ branches, flower parts, fruits & seeds (Chopra, 2008)
Fig. Field performance of diploid and tetraploid Gerbera jamesonii Bolus cv. Sciella. a. Variation in plant characteristics between diploid (2x) and tetraploid (4x); b. variation in stalk length between 2x & 4x; c. variation in flower dia between 2x & 4x
Gantait et al., 2011

Genetic Modification of Ornamental Plants
It offer new opportunities for breeders of ornamental plants
Development of new ornamental varieties through gene transfer is possible by this technique
Genetic engineering can introduce traits not be generated by conventional breeding
Major traits amenable to manipulation by genetic modification include flower color, fragrance, abiotic stress resistance, disease resistance, pest resistance, manipulation of the form and architecture of plants and/or flowers, modification of flowering time, and post harvest life etc.
Ex: Chrysanthemum, Torenia: Fringed petal
Cyclamen, Petunia: Double flower

RNA interference technology
RNA interference (RNAi) is a naturally occurring mechanism that leads to the “silencing” of genes
In consequence, the respective protein is not synthesized
This technique can be used for loss-of-function studies where a gene is specifically silenced and character is not expressed

Chimeric REpressor Gene-Silencing Technology (CRES-T)
CRES-T is a recently developed technology
It induces a dominant loss-of-function phenotype of endogenous plant Transcription Factor by expression of a chimeric repressor
Transcription factors (TFs) are key regulators for the control of various plant phenomena
Here TF is fused with the plant-specific EAR-motif repression domain, SRDX (Mitsuda et al., 2011)
It suppresses target genes of a transcription factor dominantly
CRES-T has been successfully utilized to modify the shape of torenia (Shikata et al., 2011), chrysanthemum (Narumi et al., 2011), morning glory (Sage-Ono et al., 2011), cyclamen (Tanaka et al., 2011) and rose plants (Gion et al., 2011).

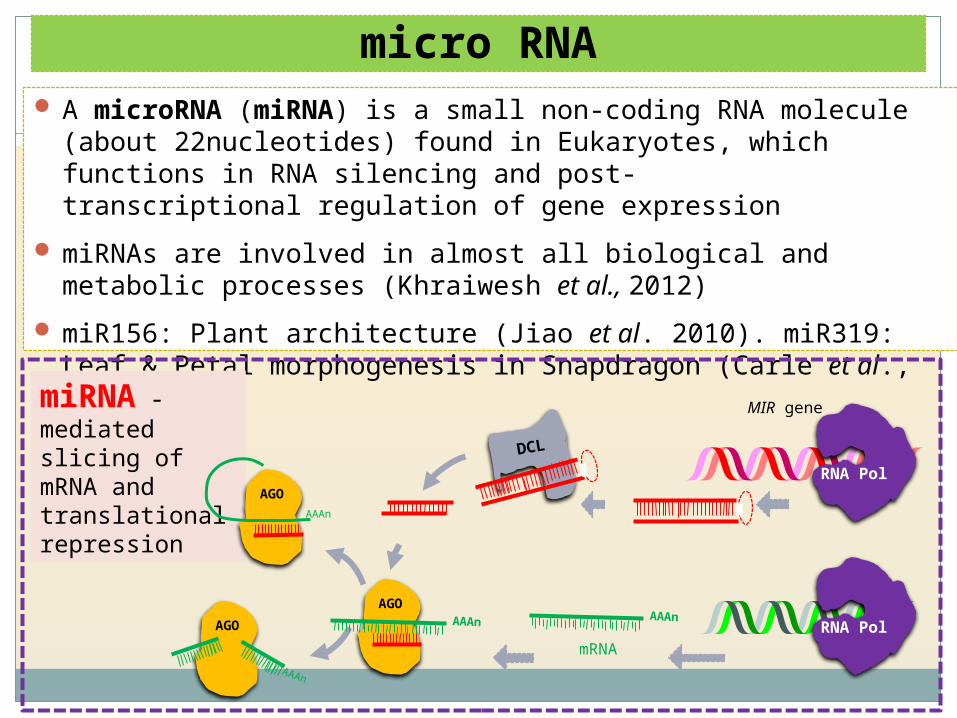
micro RNA A microRNA (miRNA) is a small non-coding RNA molecule (about 22nucleotides)
found in Eukaryotes, which functions in RNA silencing and post-transcriptional regulation of gene expression
miRNAs are involved in almost all biological and metabolic processes (Khraiwesh et al., 2012)
miR156: Plant architecture (Jiao et al. 2010). miR319: Leaf & Petal morphogenesis in Snapdragon (Carle et al., 2007)
DCL
MIR gene
RNA Pol
AGO
RNA Pol
miRNA -mediated slicing of mRNA and translational repression
mRNA
AGO
AGO
AAAn
AAAn
AAAnAAAn
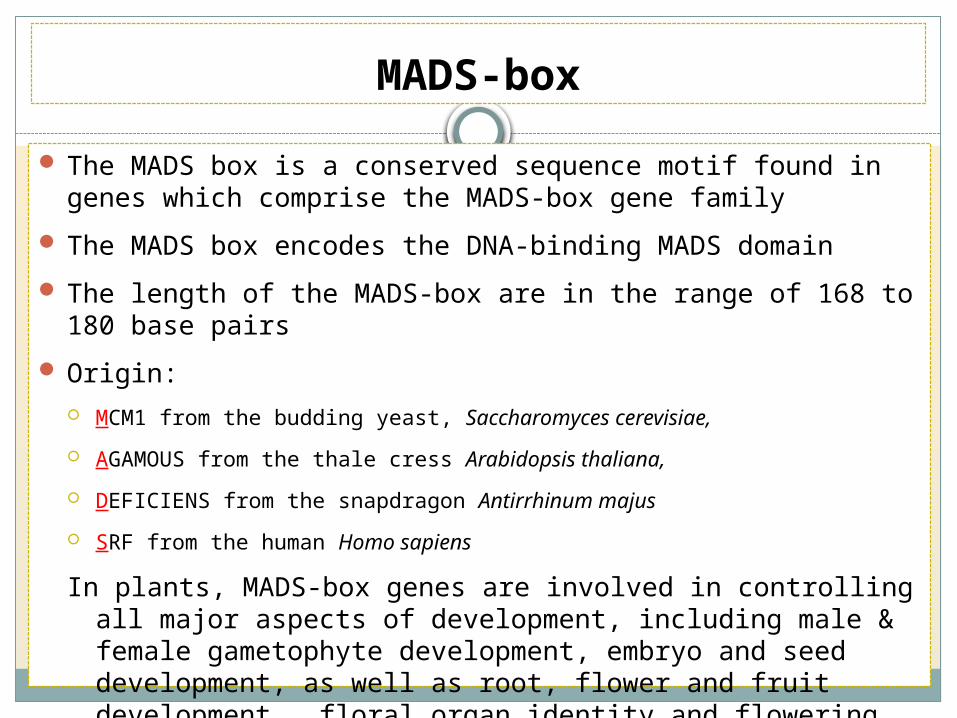
MADS-box
The MADS box is a conserved sequence motif found in genes which comprise the MADS-box gene family
The MADS box encodes the DNA-binding MADS domain
The length of the MADS-box are in the range of 168 to 180 base pairs
Origin:
MCM1 from the budding yeast, Saccharomyces cerevisiae,
AGAMOUS from the thale cress Arabidopsis thaliana,
DEFICIENS from the snapdragon Antirrhinum majus
SRF from the human Homo sapiens
In plants, MADS-box genes are involved in controlling all major aspects of development, including male & female gametophyte development, embryo and seed development, as well as root, flower and fruit development, floral organ identity and flowering time determination

CASE STUDIES
Ind
ian
Ag
ricu
ltu
ral
Res
earc
h
Inst
itu
te,
New
Del
hi

Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
c)
Case Study-1
Objectives:
1. Study of the effects of the gene silencing of C-class MADS-box genes by using a VIGS system on flower phenotypes in petunia cultivars.
2. Comparison between Large petaloid stamens induced by silencing both pMADS3 and FBP6 with small petaloid stamens induced by silencing only pMADS3.

IntroductionIn
dia
n A
gri
cult
ura
l R
esea
rch
In
stit
ute
, N
ew D
elh
i
c)
Double flowers enhances the commercial value of Petunia hybrida. As ornamental plants, double flowers with large petaloid stamens and/or new flowers at inner whorls are desired
Double flower formation: Mainly due to conversion of stamen and carpel into petal and new inflorescence
C-class genes along with B-class genes, specify stamen identity in whorl 3. A/C to ABC model of floral organ identity (Coen and Meyerowitz, 1991)
Suppressing C-class genes in whorl 3 results in the conversion of stamen into petal. C-class genes also specify carpel identity in whorl 4 and control floral meristem determinacy, their suppression induces the indeterminate development of flowers in whorl 4
C-class genes belong to AG-clade of the large MADS-box gene family

Petunia has two genes belonging to the AG-clade:
euAG- subclade gene PETUNIA MADS-BOX GENE3 (pMADS3) and
PLENA- subclade gene FLORAL BINDING PROTEIN6 (FBP6)
(Angenent et al., 2009; Tsuchimoto et al., 1993)
Silencing of either pMADS3 or FBP6 resulted in partial loss of stamen identity and slightly altered carpel morphology. No double flower
Flowers with both pMADS3 and FBP6 silenced exhibited near-complete loss of both stamen and pistil identities . They were completely converted into large petaloid tissues in whorl 3, new flowers were formed instead of carpels in whorl 4, and ornamental double flowers were produced

Materials and Methods
Plant materials:
VIGS treatments of each of the C-class MADS-box genes, pMADS3 and FBP6, and of pMADS3 & FBP6 conducted in four petunia cultivars, ‘Cutie Blue’, ‘Fantasy Blue’, ‘Picobella Blue’,and ‘Mambo Purple’
Plasmid construction:
The tobacco rattle virus (TRV)-based VIGS system (suppression of the anthocyanin pathway via chalcone synthase silencing as reporter as it produced white flower)
Vector: pTRV1 and pTRV2 VIGS
PhCHS was amplified and cloned into the EcoR1 site of pTRV2 vector
The non-conserved regions of petunia C-class genes, pMADS3 and FBP6, were amplified using the primers and cloned into the SmaI site of pTRV2 PhCHS vector individually to generate constructs for silencing pMADS3 and FBP6 separately and fused to generate a construct for silencing pMADS3 and FBP6 simultaneously

Agroinoculation of TRV vectors:Virus infection was carried out by
means of the Agrobacterium-mediated infection of petunias
Young leaves of 3-week old petunia plants were inoculated
Quantitative RT-PCR of C- and A-class MADS-box genes:Quantitative RT-PCR (qRT-PCR) of C- and A-class MADS-box genes in petals and stamens of VIGS-untreated control flowers and petaloid stamens of VIGS-induced flowers was performed.

Results and Discussion
In ‘Picobella Blue’ and ‘Mambo Purple’: No white flower was noted (Unknown genetic background, Chen et al., 2004)
In ‘CutieBlue’ and ‘Fantasy Blue’: Completely white double flowers were observed, indicating the strong and complete silencing
In flowers inoculated with either pMADS3-VIGS orFBP6-VIGS, morphologically significant but small conversions in whorls 3 & 4 were observed
In flowers of pMADS3-VIGS inoculated petunias, anthers converted into small petaloid tissues but filaments retained their original struc (Fig. 1c & d)
In flowers of FBP6-VIGS inoculated petunias, the stamens were almost unaffected
In petunias inoculated with pMADS3/FBP6-VIGS, prominent double flowers with highly ornamental appearance formed. Complete loss of stamen identity was observed. Both anthers and filaments were completely converted into petaloid tissues

Fig. 1. Morphological changes in flowers of P. hybrida cv ‘Cutie Blue’ inoculated with pTRV2-PhCHS/pMADS3 (pMADS3-VIGS) and pTRV2-PhCHS/pMADS3/FBP6 (pMADS3/FBP6-VIGS). (a) VIGS-untreated control flower; (b) Stamens and a carpel of non-VIGS
flower; (c) pMADS3-VIGS flower (white and
blue mixed color); (d) Petaloid stamens and a carpelof
pMADS3-VIGS flower; (e) pMADS3/FBP6-VIGS flower
(white); (f) Petaloid stamens and a carpel of
pMADS3/FBP6-VIGS flower (white).

Fig. 2. Morphological changes in flowers of P. hybrida cv ‘Fantasy Blue’, ‘Picobella Blue’, and ‘Mambo Purple’ inoculated with pTRV2-PhCHS/pMADS3/FBP6 (pMADS3/FBP6-VIGS). (a–c) ‘Fantasy Blue’; (d–f) ‘Picobella Blue’; (g–i) ‘Mambo Purple’; (a, d and g) VIGS-untreated control flowers; (b, e and h) pMADS3/FBP6-VIGS flowers; (c, f and i) stamens and carpels or converted new flowers of pMADS3/FBP6-VIGS flowers.

Fig. 3. New flower formation in whorl 4 and from axil of whorl 3 in a double flower of P. hybrida cv ‘Mambo Purple’ inoculated with (pMADS3/FBP6-VIGS). (a) An opened double flower with a
second new flower in whorl 4(b) An opened second new flower;(c) Fused corolla (left), a carpel
(center), and petaloid stamens (right) of the second flower;
(d) An ectopic new flower emerging from the axil of whorl 3;
(e) An unconverted stamen (left) and petal-like tissues of the ectopic new flower.
Flowers inoculated with pMADS3/FBP6-VIGS in whorl 4, carpels converted into new flower (Cultivar-dependent)In 50% of the double flowers of ‘Mambo Purple’, a 2nd new flower arose instead of a carpel. This process was repeated, generating 3rd & 4th new flowers. It exhibited a voluminous and decorative appearance with a high commercial value.

The surface areas of petaloid stamens in pMADS3/FBP6-VIGS plants were more than 10 times as large as those in pMADS3-VIGS plantsUpper limb-like region of the large petaloid stamens in pMADS3/FBP6-VIGS plants accounted for > 90% of the total area, so it was mostly due to the development of this regionThe average sizes of epidermal cells in plants inoculated with pMADS3/FBP6-VIGS were only 1.5 times as large as those in plants inoculated with pMADS3-VIGS

InferencesIn
dia
n A
gri
cult
ura
l R
esea
rch
In
stit
ute
, N
ew D
elh
i
c)
Double flowers can be induced by virus-induced gene silencing (VIGS) of two C-class MADS-box genes, pMADS3 and FBP6
Large petaloid stamens induced by pMADS3/FBP6-VIGS were compared with small petaloid stamens induced by pMADS3-VIGS
New flower formation in the inner whorl of flowers silenced in both pMADS3 and FBP6 gene is cultivar-dependent
They are valuable for future breeding of petunia cultivars bearing decorative double flowers with large petaloid stamens and inner new secondary flowers

Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
c)
Case Study-2
Objectives:
1. Isolation of flower color and shape mutants
2. Development of in vitro mutation technique to obtain new varieties of chrysanthemum by using gamma rays

Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
c)
Introduction
Chrysanthemum morifolium Ramat: Important ornamental flowers
For commercial floriculture: Demand for novel varieties
Radiation used for the development of new flower color/shape mutants in Chrysanthemums (Misra et al., 2003)
The selection of ornamentals is easy with visible characters (color, shape and size, or leaf form and growth habit).
The main bottleneck is formation of chimera where, the size of the mutant sector varies from a narrow streak on a petal to the entire flower
Many new flower color/shape mutants, lost due to the lack of a regeneration system from small-mutated sectors either in vivo or in vitro
Therefore, a regeneration system to establish mutant in pure form from a chimera and production of a wide range of new cultivars with novel flower colors and shapes is required

Work was done at China Agricultural University, Beijing, ChinaPlant materialsWhite C. morifolium Ramat cv. Youka plants regenerated from explants
of petals (spoon shape) used in the experimentCultureWhite petals (from Bud) 4 mm in length of the original cultivar were
excised and cultured on MS basal mediumThe calli was induced on a callus medium is comprised of MS medium
supplemented with 1.0 mg/l NAA, 2.0 mg/l BAP60CO radiation treatmentsCallus was exposed to gamma radiation using 60Co of gamma chamber
with doses of 0(Control), 10, 15 and 20 Gy and dose rate 1.02 Gy/min.
Materials and Methods

Fig. 1 In vitro regeneration C. morifolium ‘Youka’ from ray florets. A) White flower buds; B) Callus induction medium; C) Adventitious shoots formation after 4 weeks; D) In vitro Roots formations after 25 days; E) Plantlets in hardening chamber

Results and Discussion
Table: In vitro callus survival (%) and number of shoots(Mean ± SE) of white C. morifolium ‘‘Youka’’ as influenced by gamma ray dosesGamma
ray dose (Gy)
Callus survival
(%)
No. of shoots
0 86.67 7.22 ± 0.11a
10 62.43 7.67 ± 0.33a
15 30.33 7.89 ± 0.29a
20 17.23 3.00 ± 0.29b
Effect of gamma radiation on chrysanthemum callus
Table: Effect of in vitro treatment of C. morifolium ‘Youka’ with gamma radiation on flowering characteristics of the generated plantlets
Character Control 10 Gy 15 Gy 20 Gy
Flower No/plant 4.22 ± 0.29a
4.38 ± 0.12a
4.05 ± 0.41a
3.00 ± 0.00b
Flower dia (cm) 6.12 ± 0.23a
6.11 ± 0.15a
5.15 ± 0.18b
4.88 ± 0.06b
Petal length (cm) 3.03 ± 0.20a
3.14 ± 0.19a
3.00 ± 0.01a
3.01 ± 0.01a
Petal width (cm) 0.83 ± 0.07a
0.87 ± 0.12a
0.97 ± 0.03a
1.05 ± 0.05a
Petiole length (cm) 4.50 ± 0.17b
4.72 ± 0.18b
6.52 ± 0.29a
6.51 ± 0.30a
Petiole dia (mm) 2.50 ± 0.11a
1.89 ± 0.04b
1.89 ± 0.04b
1.85 ± 0.04b
Similar letters in the same row indicate that they were not significantly different from LSD005 test
Effect of gamma radiation on flower characteristics

Fig. Flower of tissue-raised plants . A) Control, white colored/spoon shaped petals; B) M1, white colored / tubular petals; C) M2, yellow colored/spoon shaped petals; D) M3, yellow colored/ flat shaped petals
The frequencies of flower color and shape mutations increased when the total dose was increased from 10 to 15 Gy, though it was not observed when the dose was increased from 15 to 20 Gy. Figure below illustrates the three mutants obtained in 15 Gy treated plants.

Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
c)
Inferences
Gamma radiation with 15 Gy dose can be used for in vitro induction of flower color and shape mutations of chrysanthemum cv. Youka
The isolated mutants on in vitro culture can be multiplied and rooted in vitro to produce new varieties of chrysanthemum

Conclusion:In
dia
n A
gri
cult
ura
l R
esea
rch
In
stit
ute
, N
ew D
elh
i
c)
Creation of new flower shapes in ornamentals plants is a major breeding target as increase its commercial value
Flower development is controlled by Gene (ABCDE-Model)
Different flower shape including double flower can be developed by different breeding techniques like Hybridization, Mutation, Polyploidy, Genetic engineering etc.
Now a days new techniques like RNAi, CRES-T, miRNA and other gene silencing techniques are being used to developed altered flower shape
Even though there are many techniques available but, very few variety has been developed for commercial purpose by Genetic transformation

Future thrust:In
dia
n A
gri
cult
ura
l R
esea
rch
In
stit
ute
, N
ew D
elh
i
c)
Identification of flower shape mutant and development of in-vitro protocol for their regeneration
Most of the genetic modification for flower form is done in Arabidopsis & Snapdragon only, it has be done in commercially important crop
Only few GM crop is released for commercial purpose i.e., Rose & Carnation (Flower colour). Effort is to be done to developed GM crop with modified flower architecture

Ind
ian
Ag
ricu
ltu
ral
Res
earc
h I
nst
itu
te,
New
Del
hi
c)


















