
Brain-Penetrating IgG-Iduronate 2-Sulfatase Fusion...
39
DMD #42903 1 Brain-Penetrating IgG-Iduronate 2-Sulfatase Fusion Protein for the Mouse Qing-Hui Zhou Ruben J. Boado Jeff Zhiqiang Lu Eric Ka-Wai Hui William M. Pardridge Department of Medicine (Q.-H.Z., R.J.B., W.M.P.), UCLA, Los Angeles, California and ArmaGen Technologies, Inc. (R.J.B., E.K.-W.H., J.Z.L) Santa Monica, California DMD Fast Forward. Published on November 7, 2011 as doi:10.1124/dmd.111.042903 Copyright 2011 by the American Society for Pharmacology and Experimental Therapeutics. This article has not been copyedited and formatted. The final version may differ from this version. DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903 at ASPET Journals on June 12, 2018 dmd.aspetjournals.org Downloaded from
Transcript of Brain-Penetrating IgG-Iduronate 2-Sulfatase Fusion...

DMD #42903
1
Brain-Penetrating IgG-Iduronate 2-Sulfatase Fusion Protein
for the Mouse
Qing-Hui Zhou
Ruben J. Boado
Jeff Zhiqiang Lu
Eric Ka-Wai Hui
William M. Pardridge
Department of Medicine (Q.-H.Z., R.J.B., W.M.P.), UCLA, Los Angeles, California and
ArmaGen Technologies, Inc. (R.J.B., E.K.-W.H., J.Z.L) Santa Monica, California
DMD Fast Forward. Published on November 7, 2011 as doi:10.1124/dmd.111.042903
Copyright 2011 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
2
Running title: Brain Penetrating IgG-IDS Fusion Protein
Address correspondence to:
Dr. William M. Pardridge
UCLA Warren Hall 13-164
900 Veteran Ave.
Los Angeles, CA 90024
Ph: 310-825-8858
Fax: 310-206-5163
Email: [email protected]
Text pages: 20
Tables: 5
Figures: 6
References: 24
Abstract: 244 words
Introduction: 489 words
Discussion: 1126 words
Abbreviations: BBB, blood-brain barrier; IDS, iduronate 2-sulfatase; MAb, monoclonal antibody; TfR, transferrin receptor; cTfRMAb, chimeric MAb against the mouse TfR; HIR, human insulin receptor; HIRMAb, engineered MAb against the HIR; ID, injected dose; HC, heavy chain; LC, light chain; TV, tandem vector; CHO, Chinese hamster ovary; TCA, trichloroacetic acid; VD, volume of distribution: PS, permeability-surface area; AUC, area under the curve; IV, intravenous; TBS, Tris buffered saline; ELISA, enzyme linked immunosorbent assay; IDUA, iduronidase; AA, amino acid
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
3
Abstract
Mucopolysaccharidosis (MPS) Type II, Hunter’s syndrome, is caused by mutations in the
iduronate 2-sulfatase (IDS) fusion protein. MPS-II affects the brain, and enzyme replacement
therapy is not effective in the brain, because the enzyme does not cross the blood-brain barrier
(BBB). In order to treat mouse models of MPS-II with brain penetrating IDS, the lysosomal
enzyme was re-engineered as an IgG-IDS fusion protein. The mature human IDS was fused to
the carboxyl terminus of both heavy chains of the chimeric monoclonal antibody (MAb) against
the mouse transferrin receptor (TfR), and the fusion protein is designated cTfRMAb-IDS. The
purity and identity of the fusion protein was confirmed by electrophoresis and Western blotting
with antibodies to mouse IgG and human IDS. The EC50 of binding of the cTfRMAb-IDS fusion
protein to the mouse TfR, 0.85 ± 0.15 nM, was comparable to the EC50 of binding of the
cTfRMAb, 0.78 ± 0.05. The IDS enzyme activity of the cTfRMAb-IDS fusion protein was 126
± 1 nmol/hr/ug protein. After intravenous injection in the mouse, the cTfRMAb-IDS fusion
protein was rapidly removed from plasma and distributed to tissues, including brain and spinal
cord. The uptake of the fusion protein by brain or spinal cord was 1.3±0.1 and 2.2 ± 0.2 %
injected dose (ID)/gram, respectively, which is 100-fold greater than the brain uptake of IDS
alone. This work shows that a lysosomal sulfatase can be re-engineered as an IgG-enzyme fusion
protein that rapidly penetrates the brain after intravenous administration.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
4
Introduction
Most lysosomal storage disorders affect the brain, including Mucopolysaccharidosis
(MPS) Type II, or Hunter’s syndrome. MPS-II is caused by mutations in the gene encoding for
iduronate 2-sulfatase (IDS). MPS-II is treated with enzyme replacement therapy (ERT) using
recombinant human IDS (Muenzer et al, 2006). However, MPS-II severely affects the brain and
spinal cord (Al Sawaf et al, 2008), and ERT does treat the central nervous system (Wraith et al,
2008), because IDS, like other large molecule pharmaceuticals, does not cross the blood-brain
barrier (BBB). To provide a new treatment of the brain, the IDS enzyme was re-engineered as a
brain-penetrating IgG-enzyme fusion protein. The mature human IDS was fused to the carboxyl
terminus of each heavy chain of a genetically engineered monoclonal antibody (MAb) against
the human insulin receptor (HIR), and this fusion protein was designated the HIRMAb-IDS
fusion protein (Lu et al, 2010). The HIRMAb part of the fusion protein acts as a transport vector
to ferry the fused IDS across the BBB via receptor-mediated transport on the endogenous BBB
insulin receptor. The HIRMAb part of the fusion protein cross reacts with the insulin receptor in
Old World primates, such as the Rhesus monkey (Pardridge et al, 1995), and the HIRMAb-IDS
fusion protein rapidly penetrates the brain from blood in the Rhesus monkey (Lu et al, 2011).
However, the HIRMAb does not cross react with the insulin receptor in the mouse. There is no
known MAb against the murine insulin receptor that could be used as a BBB transport vector in
the mouse. BBB receptor-specific antibodies that can be used as brain transport vectors are
limited to those antibodies that bind an endocytosing exofacial epitope on the alpha subunit of
the murine insulin receptor. Therefore, a surrogate transport vector has been engineered for
mouse studies, which is a chimeric MAb against the mouse transferrin receptor (TfR), and
designated the cTfRMAb (Boado et al, 2009a). The variable regions of the cTfRMAb are
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
5
derived from a rat TfRMAb, and the constant regions of the heavy and light chain are derived
from mouse IgG1 and mouse kappa, respectively. Recently, a fusion protein of the cTfRMAb
and murine iduronidase (IDUA), the enzyme mutated in MPS-I, Hurler’s syndrome, was
engineered (Boado et al, 2011). Chronic, twice-weekly intravenous treatment of Hurler mice
with 1 mg/kg doses of the cTfRMAb-IDUA fusion protein resulted in a 73% decrease in brain
lysosomal inclusion bodies, as well as reduction in glycosoaminoglycan (GAG) levels in
peripheral tissues (Boado et al, 2011).
The purpose of the present study was to engineer a brain penetrating form of IDS that is
active in the mouse, and to measure the brain uptake of the enzyme in the mouse. The amino
terminus of mature IDS was fused to the carboxyl terminus of both heavy chains of the
cTfRMAb, and this fusion protein is designated cTfRMAb-IDS. The present study also
describes the cloning of an expression plasmid encoding human sulfatase modifying factor
(SUMF)-1. The generation of IDS enzyme activity requires a post-translational modification of
Cys59 to N-formyl glycine, which is mediated by SUMF1 (Zito et al, 2005). The role of co-
expression of the genes encoding SUMF1 and the cTfRMAb-IDS fusion protein on IDS enzyme
activity of the fusion protein was examined.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
6
Methods
Engineering of cTfRMAb-IDS expression vector. The human IDS cDNA, encoding
Ser26-Pro550 minus the 25 amino acid signal peptide (Genbank NP_000193) was produced by
reverse transcription (RT) as previously described (Lu et al., 2010) using forward (5’-phosphate-
CCTCCGAAACGCAGGCCAACTCG) and reverse (5’- phosphate-
TCAAGGCATCAACAACTGGAAAAGATC) oligodeoxynucleotides (ODN) synthesized at
Midland Certified Reagent Co. (Midland, TX). The PCR-generated ~1.5 kb IDS cDNA was
initially inserted at the HpaI site of the mouse IgG1 expression vector pCD-HC (Boado et al.,
2009a) to form a single expression vector for the cTfRMAb heavy chain-IDS fusion protein,
designated pCD-cTfRMAb-IDS. A tandem vector (TV) containing on a single plasmid DNA the
expression cassettes encoding the cTfRMAb heavy chain-IDS fusion protein, the cTfRMAb light
chain (LC), and the murine dihydrofolate reductase (DHFR), was engineered similar to a TV
described previously (Boado et al., 2010). The complete cTfRMAb-IDS heavy chain open
reading frame (orf) plus the bovine growth hormone (BGH) polyadenylation sequence was
obtained from pCD-cTfRMAb-IDS by NotI-AfeI digestion and inserted into the NotI-HpaI sites
of the TV. The TV coding for the cTfRMAb-IDS fusion protein was characterized by restriction
endonuclease mapping using XhoI. The DNA sequence of all expression plasmids was
confirmed by bi-directional DNA sequencing performed at Sequetech (Mountain View, CA)
using custom ODNs synthesized at Midland.
Engineering of pSUMF1 expression vector. The complete orf of the human sulfatase
modifying factor 1 (SUMF1) cDNA was produced by RT-PCR using ODNs generated from the
nucleotide sequence of the human SUMF1 mRNA (NM_182760), including the forward ODN
(AAAGAATTCTTGCCGCCACCATGGCTGCGCCCGCACTA) and reverse ODN
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
7
(CTTAAGCTTTCAGTCCATAGTGGG). The forward ODN introduces an EcoRI site and a
complete Kozak sequence followed by the methionine inititation codon of SUMF1. The reverse
ODN ends with the stop codon of SUMF1, which is followed by a HindIII site. The RT-PCR
was performed as previously described using human liver polyA+ RNA as template (Boado et al,
2008). The PCR-generated ~1.2 kb SUMF1 cDNA was eluted following agarose gel
electrophoresis and was digested with EcoRI and HindIII and inserted at the same restriction
endonuclease sites to form the expression vector designated pSUMF1. In the vector, the SUMF1
orf is 5’-flanked by the cytomegalovirus (CMV) promoter, and 3’-flanked by the BGH
polyadenylation sequence.
Expression in COS cells and affinity purification. COS cells were lipofected with the
cTfRMAb-IDS TV, with or without co-lipofection with the pSUMF1 vector, using
Lipofectamine 2000, with a ratio of 1:2.5 ug DNA:uL Lipofectamine. Following transfection,
the cells were cultured in 6-well cluster dishes in serum free VP-SFM (Invitrogen, Carlsbad,
CA). The conditioned medium (CM) was collected at 3 and 7 days. Transgene expression and
fusion protein secretion to the medium was assayed by measurement of mouse IgG in the
conditioned medium. Mouse IgG ELISA was performed in Immulon 2 high binding plates
(Dynex Tech., Chantilly, VA) with COS cell conditioned medium. For large scale production of
the COS-derived cTfRMAb-IDUA fusion protein, the lipofection was scaled up, and lipofected
cells were plated in 12xT500 flasks. The serum free CM was collected at 3 and 7 days and
pooled and ultra-filtered with a Sartopore-2 (0.45+0.22 um) filter (Sartorius Stedim North
America, Bohemia, NY). Following filtration, the CM was applied to a 25 mL protein G
Sepharose 4 Fast Flow (GE Life Sciences, Chicago, IL) column equilibrated in 25 mM Tris/25
mM NaCl/5 mM EDTA/pH=7.1. Following application of the sample, the column was washed
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
8
with 25 mM Tris/1 M NaCl/5 mM EDTA/pH=7.1, and the fusion protein was eluted with 0.1 M
glycine/pH=2.8. The acid eluate was pooled, Tris was added to raise the pH to 5.5, and NaCl was
added to 0.15 M. The volume of the pool was concentrated to 2 mL with an Ultra-15 centrifugal
filter unit with a 30,000 molecular weight cut-off (Millipore, Billerica, MA), and the solution
was stored sterile-filtered at -70ºC. The protein content of the final drug product was measured
with the bicinchoninic acid (BCA) assay (Pierce Chemical Co., Rockford, IL).
SDS-PAGE and Western blotting. The homogeneity of the cTfRMAb-IDS fusion
protein was evaluated with sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-
PAGE) under reducing and non-reducing conditions using 12% and 7.5% Tris-HCl gels (Biorad
Life Science, Hercules, CA), respectively. The prestained molecular weight (MW) standards
were obtained from Fermentas (Glen Burnie, MD) and from Invitrogen (Carlsbad, CA) for the
reducing and non-reducing gels, respectively. Western blot analysis was performed with a goat
anti-mouse IgG (H+L) antibody (Bethyl Labs, Montgomery, TX) for the mouse IgG Western,
and with a goat anti-human IDS antibody (R&D Systems, Minneapolis, MN) for the IDS
Western. The immunoreactivity of the cTfRMAb-IDS fusion protein was compared with the
cTfRMAb and with the HIRMAb-IDS fusion protein described previously (Boado et al, 2009a;
Lu et al, 2011). The HIRMAb-IDS fusion protein was produced in Chinese hamster ovary
(CHO) cells grown in serum free medium as described previously (Lu et al, 2011).
Mouse TfR binding ELISA. The affinity of fusion protein binding to the mouse TfR
extracellular domain (ECD) was determined with an ELISA. The Cys89-Phe763 mouse TfR ECD
(Sino Biologicals, Beijing, China) was plated on Nunc-Maxisorb 96 well dishes (200 ng/well) in
0.05 M NaHCO3/pH=8.3 overnight at 4C. The plates were washed with 0.01 M Tris/0.15 M
NaCl/pH=7.4 (TBS), and blocked with TBS/1% bovine serum albumin for 30 min. The fusion
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
9
protein or antibody solutions were added for 60 min at room temperature (RT). Following
washing with TBS/0.05% Tween-20 (TBST), the secondary antibody, a conjugate of a goat anti-
mouse IgG and alkaline phosphatase (Bethyl Labs, Montgomery, TX), was added for 45 min at
RT (100 ng/well). After washing with TBST, 100 uL/well of para-nitrophenylphosphate (Sigma
Chemical, St. Louis, MO), 1 mg/mL, was incubated at RT in the dark for 15 min. The reaction
was terminated with 1.2 M NaOH, and the absorbance (Abs) at 405 nM was measured with an
ELISA plate reader. The concentration of fusion protein that caused 50% of maximal binding,
the EC50, was determined by non-linear regression analysis by fitting the data to: Abs =
(Amax·S)/(EC50 + S), where Amax is the maximal absorbance and S=the concentration of IgG
or fusion protein. The proteins tested in the ELISA were the COS-derived cTfRMAb-IDS, the
CHO-derived cTfRMAb, and mouse IgG1κ (Sigma), which is the isotype control of the
cTfRMAb antibody (Boado et al, 2009a).
IDS enzyme assay. IDS enzyme activity was measured using a 2-step fluorometric
enzymatic assay (Voznyi et al, 2001), as described previously (Lu et al, 2010). The fusion
protein was tested at concentrations ranging from 0.2 to 0.5 ug/mL in a volume of 100 uL. The
IDS enzyme substrate, 4-methylumbelliferyl α-L-iduronide-2-sulphate (4-MUS), was purchased
from Moscerdam Substrates (Rotterdam, The Netherlands). In the first step of the reaction, IDS
converts 4-MUS to 4-methylumbelliferyl α-L-iduronide (MUBI). In the second step, iduronidase
(IDUA) converts MUBI to 4-methylumbelliferone (4-MU), and the 4-MU is detected
fluorometrically. The source of the IDUA was the HIRMAb-IDUA fusion protein produced from
CHO cells as described previously (Boado et al, 2009b).
Fusion protein uptake by mouse fibroblasts. 3T3 mouse fibroblasts (American Type
Culture Collection, Manassas, VA) were plated in 6-well Biocoat dishes (Becton-Dickinson,
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
10
Franklin Lakes, NJ) and grown to confluency in DMEM medium with 10% fetal bovine serum.
The medium was removed, and the cells washed with phosphate buffered saline (PBS), followed
by the addition of 1.0 mL/well of fresh DMEM containing 2 ug/mL cTfRMAb-IDS fusion
protein followed by incubation at 37C for 2 hours. The medium was then removed, and the wells
were washed with 1.5 mL/well of fresh serum free DMEM medium 3 times. Fresh DMEM
medium with 10% FBS, 1.0 mL/well, was added and the dishes were incubated at 37C for 0, 2,
24, 48, 72, or 96 hours. At the end of the incubation, the medium was removed and the wells
were washed with 1.5 mL/well cold PBS 3 times. The monolayer was lysed with the addition of
0.3 mL/well of 5 mM sodium acetate/pH=4.5/0.2% Triton X-100 followed by 3 successive
freeze-thaw cycles. The lysate was centrifuged at 8,000 g for 10 minutes at 4C, and the
supernatant was transferred to new tubes for measurement of intracellular protein with the
bicinchoninic acid assay (Thermo Fisher Scientific, Rockford, IL), and intracellular IDS enzyme
activity using the 2 step fluorometric assay described above. The intracellular IDS enzyme
activity was expressed as nmol/hr/mg cellular protein.
Radio-labeling of fusion protein. The cTfRMAb-IDS fusion protein, which was injected
into mice for a pharmacokinetics analysis, was radiolabeled with [3H]-N-succinimidyl propionate
from American Radiolabeled Chemicals (St. Louis, MO). The labeled protein was purified with
a 1x28 cm Sephadex G-25 gel filtration column, with an elution buffer of 0.01 M sodium
acetate/0.14 M NaCl/pH=6.5/0.2% Tween-20 (ABST), followed by 2 successive 1:15 buffer
exchanges with ABST using an Ultra-15 microconcentrator (Millipore, Billerica, MA). The
cTfRMAb-IDS fusion protein was labeled to a specific activity of 0.5 uCi/ug and a
trichloroacetic acid (TCA) precipitability of 95%. The murine OX26 MAb against the rat
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
11
transferrin receptor was purified from hybridoma generated ascites with protein G affinity
chromatography, and was iodinated with chloramine T as described previously (Lee et al, 2000).
Pharmacokinetics and brain uptake in the mouse. Adult male C57BL/6J mice,
weighing 28 grams, were obtained from Jackson Labs (Bar Harbor, ME). All procedures were
carried out in accordance with the Guide for the Care and Use of Laboratory Animals as adopted
and promulgated by the U.S. National Institutes of Health. Mice were anesthetized with intra-
peritoneal (IP) ketamine (100 mg/kg) and xylazine (10 mg/kg), and injected intravenously (IV)
in the tail vein with 60 uL (10 uCi) of [3H]-cTfRMAb-IDS fusion protein. The injection dose
(ID) in each mouse of the cTfRMAb-IDS fusion protein was 0.67 mg/kg. An aliquot (50 uL) of
venous blood was collected at 0.25, 2, 5, 15, 30, and 60 min after injection by sampling the
orbital vein. The blood was centrifuged for collection of plasma, which was analyzed for
radioactivity. At 60 min after injection, the mice were euthanized and the cerebral hemispheres
were removed and weighed for each mouse. One hemisphere was used for total radioactivity
after solubilization in Soluene-350 (PerkinElmer), and one hemisphere was homogenized for
capillary depletion analysis. Plasma and tissue samples were analyzed for 3H radioactivity with
Optifluor-O (PerkinElmer) and a Tricarb 2100TR liquid scintillation counter (PerkinElmer).
Brain uptake data was expressed as a volume of distribution (VD), which is the ratio of the 60
min organ radioactivity [disintegrations per minute (DPM)/gram) divided by the 60 min plasma
radioactivity (DPM/uL), or as % of injected dose (ID)/gram tissue. The plasma radioactivity that
was precipitable with cold 10% TCA was determined at each time point.
The plasma TCA-precipitable radioactivity, DPM/mL, was converted to % injected dose
(ID)/mL, and the %ID/mL was fit to a bi-exponential equation,
%ID/mL = A1e-k1t + A2e-k2t
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
12
The intercepts (A1, A2) and the slopes (k1, k2) were used to compute the pharmacokinetics
parameters, including the median residence time (MRT), the central volume of distribution (Vc),
the steady state volume of distribution (Vss), the area under the plasma concentration curve at 60
min (AUC), the steady state AUC (AUCss), and the systemic clearance (CL). Non-linear
regression analysis used the AR subroutine of the BMDP Statistical Software. Data were
weighted by 1/(%ID/mL)2.
The organ clearance (uL/min/g), also called the BBB permeability-surface area (PS)
product, is computed from the terminal brain uptake (%ID/g) and the 60 min plasma AUC
(%IDmin/mL) as follows:
PS product = [(%ID/g)/AUC]x1000
The brain uptake was determined from total brain 3H radioactivity, and the plasma AUC was
determined from TCA-precipitable 3H radioactivity.
Capillary depletion method. One cerebral hemisphere from each mouse was processed
for the capillary depletion method as described previously (Triguero et al, 1990), which separates
the brain into 3 fractions: the total brain homogenate, the microvascular capillary pellet, and the
post-vascular supernatant. The volume of distribution (VD) was determined for each of the 3
fractions from the ratio of total 3H radioactivity in the fraction, DPM/gram brain, divided by the
3H radioactivity in the 60 min terminal plasma, DPM/uL. A high VD in the post-vascular
supernatant, as compared to the VD in the capillary pellet, is evidence for transport of the protein
through the BBB into brain parenchyma.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
13
Results
Genetic engineering and amino acid sequence analysis of fusion protein. The
cTfRMAb-IDS tandem vector encoded the expression cassettes for the heavy chain fusion gene,
the light gene, and the DHFR gene on a single plasmid DNA. DNA sequence analysis showed
the 3 expression cassettes spanned 10,452 nucleotides (nt). The light chain (LC) was comprised
of 234 amino acids (AA), which included a 20 AA signal peptide, a 108 AA variable region of
the light chain of the cTfRMAb, and a 106 AA mouse kappa light chain constant region, which
is 100% identical to AA 113-218 of mouse kappa light chain (GenBank BAA06141). The
predicted molecular weight of the light chain is 23,554 Da with a predicted isoelectric point of
5.73. The fusion protein of the cTfRMAb heavy chain (HC) and the human IDS was comprised
of 988 AA, which included a 19 AA signal peptide. The predicted molecular weight of the heavy
chain, without glycosylation, is 108,229 Da with a predicted pI of 5.47. The domains of the
fusion heavy chain following the signal peptide include a 118 AA variable region of the heavy
chain of the cTfRMAb, a 324 AA mouse IgG1 constant region, which is 100% identical to AA
140-463 of mouse IgG1 heavy chain (GenBank BAC44885), a 2 AA linker (Ser-Ser), and the
525 AA human IDS, which is 100% identical to AA 26-550 of NP_000193. The HC contains 9
N-linked glycosylation sites: 1 site in the constant region of the IgG heavy chain and 8 sites in
the IDS domain. The predicted molecular weight, without glycosylation, of the hetero-tetrameric
cTfRMAb-IDS fusion protein is 263,566 Da with an isoelectric point of 5.50.
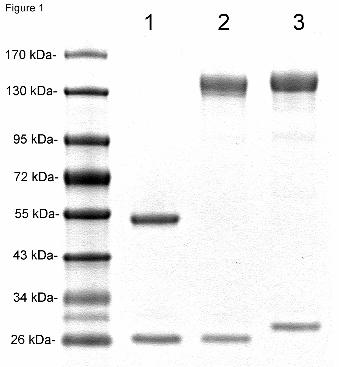
Purity and identity of fusion protein. The cTfRMAb-IDS fusion protein was purified
by protein G affinity chromatography following lipofection in COS cells and secretion into
serum free medium (Methods). Following reducing SDS-PAGE, the cTfRMAb-IDS fusion
protein was comprised of a HC, which migrated as a molecular weight (MW) of 136 kDa and a
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
14
LC that migrated with a MW of 26 kDa, which were comparable in size to the HC and LC of the
HIRMAb-IDS fusion protein (Figure 1). The MW estimates were made by linear regression from
the migration of the MW standards analyzed in the SDS-PAGE (Figure 1). The HC and LC of
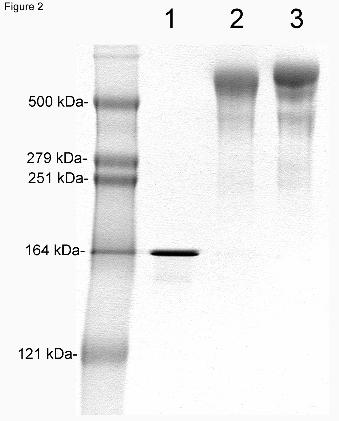
the cTfRMAb migrated at 56 kDa and 26 kDa, respectively (Figure 1). On non-reducing SDS-
PAGE, the cTfRMAb migrated at 156 kDa, and the cTfRMAb-IDS fusion protein and the
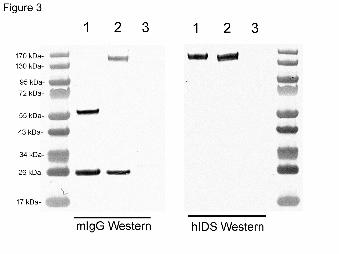
HIRMAb-IDS fusion protein migrated as a dimeric structure at 570 kDa (Figure 2). On Western
blotting, the anti-mouse IgG primary antibody reacted with the HC and LC of the cTfRMAb and
the cTfRMAb-IDS fusion protein, but did not react with the HIRMAb-IDS fusion protein (Figure
3, left panel). The anti-human IDS primary antibody reacted with the HC of the cTfRMAb-IDS
and HIRMAb-IDS fusion proteins, but did not react with the LC of either fusion protein and did
not react with the HC or LC of the cTfRMAb (Figure 3, right panel).
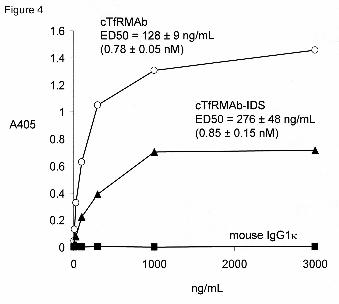
Fusion protein binding to mouse transferrin receptor. The binding of the cTfRMAb
or the cTfRMAb-IDS fusion protein to the ECD of the mouse TfR was determined by ELISA.
Both proteins bound to the mouse TfR via a saturable mechanism, whereas there was no binding
of mouse IgG1κ to the mouse TfR (Figure 4). The EC50 of binding was 128 ± 9 and 276 ± 48
ng/mL for the cTfRMAb and the cTfRMAb-IDS fusion protein, respectively. The MW of the
hetero-tetrameric protein for the TfRMAb and the cTfRMAb-IDS fusion protein, based on the
reducing SDS-PAGE (Figure 1), is 164 kDa and 324 kDa, respectively. Therefore, the EC50 of
binding to the mouse TfR for the cTfRMAb and the cTfRMAb-IDS fusion protein is 0.78 ± 0.05
and 0.85 ± 0.15 nM, respectively.
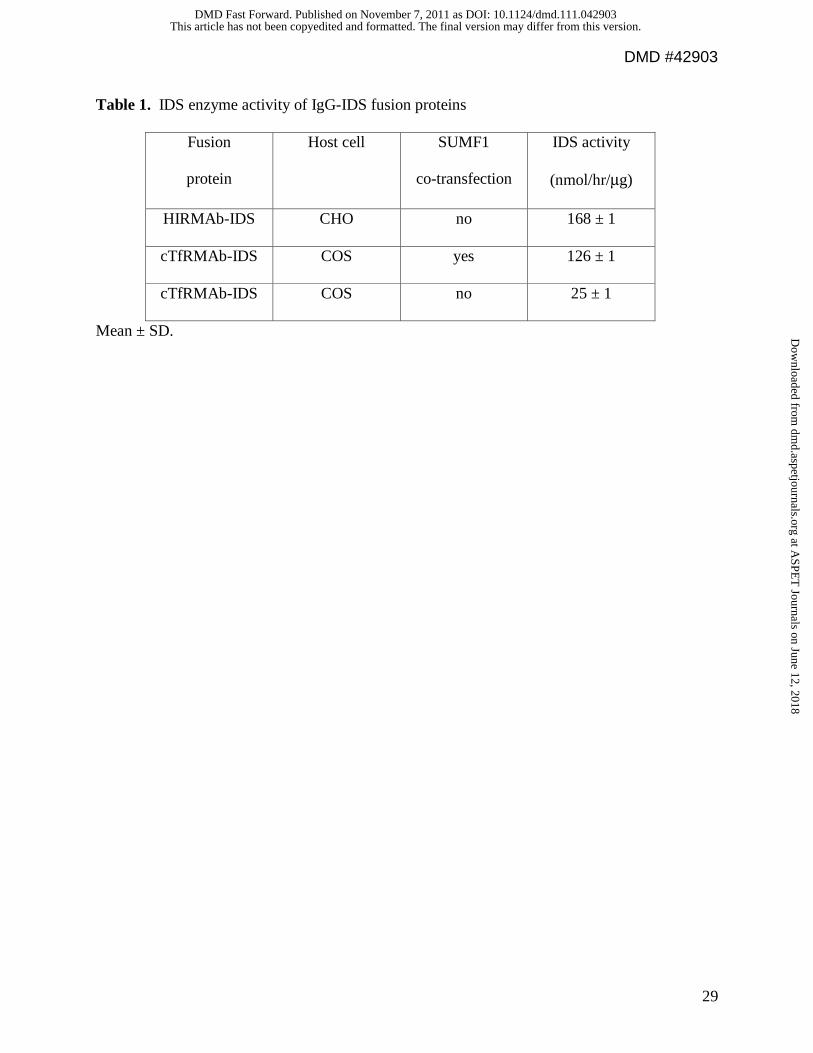
Fusion protein IDS enzyme activity. The cTfRMAb-IDS fusion protein was expressed
in COS cells with or without co-lipofection with the pSUMF1 expression plasmid. Following
fusion protein expression alone, the IDS enzyme activity of the purified cTfRMAb-IDS fusion
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
15
protein was 25 ± 1 nmol/hr/ug protein, whereas the IDS enzyme activity of the cTfRMAb-IDS
fusion protein was 126 ± 1 nmol/hr/ug protein following co-lipofection of the fusion protein and
SUMF1 (Table 1). The IDS enzyme activity of the HIRMAb-IDS fusion protein produced in
CHO cells without co-expression of SUMF1 was 168 ± 1 nmol/hr/ug protein (Table 1). The
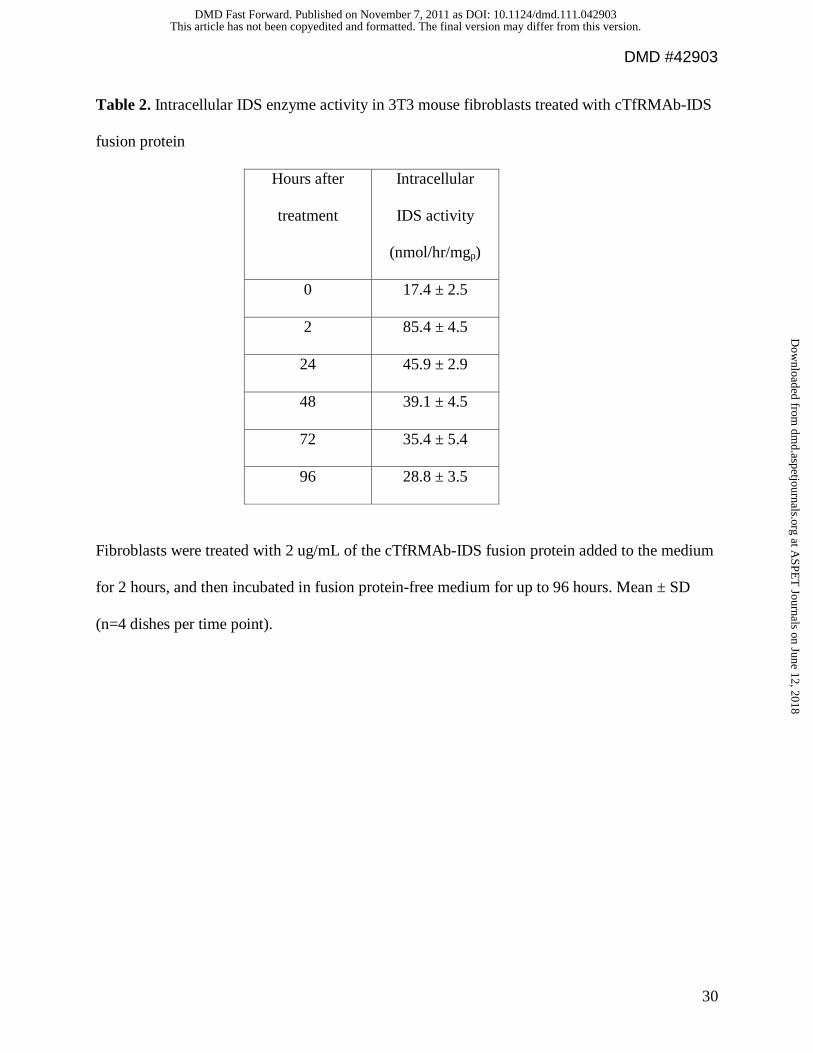
addition of the cTfRMAb-IDS fusion protein to the medium of mouse 3T3 fibroblasts resulted in
5-fold increase in intracellular IDS enzyme activity following a 2-hour exposure of the cells to
the fusion protein (Table 2). The cells were then incubated in fusion protein-free fresh medium,
and the increased IDS enzyme activity persisted for at least 96 hours after the initial single 2-
hour exposure of the cells to the fusion protein (Table 2). Linear regression analysis of the data
in Table 2 showed the IDS enzyme activity decayed with a half-time of 1.7 days (r=0.96).
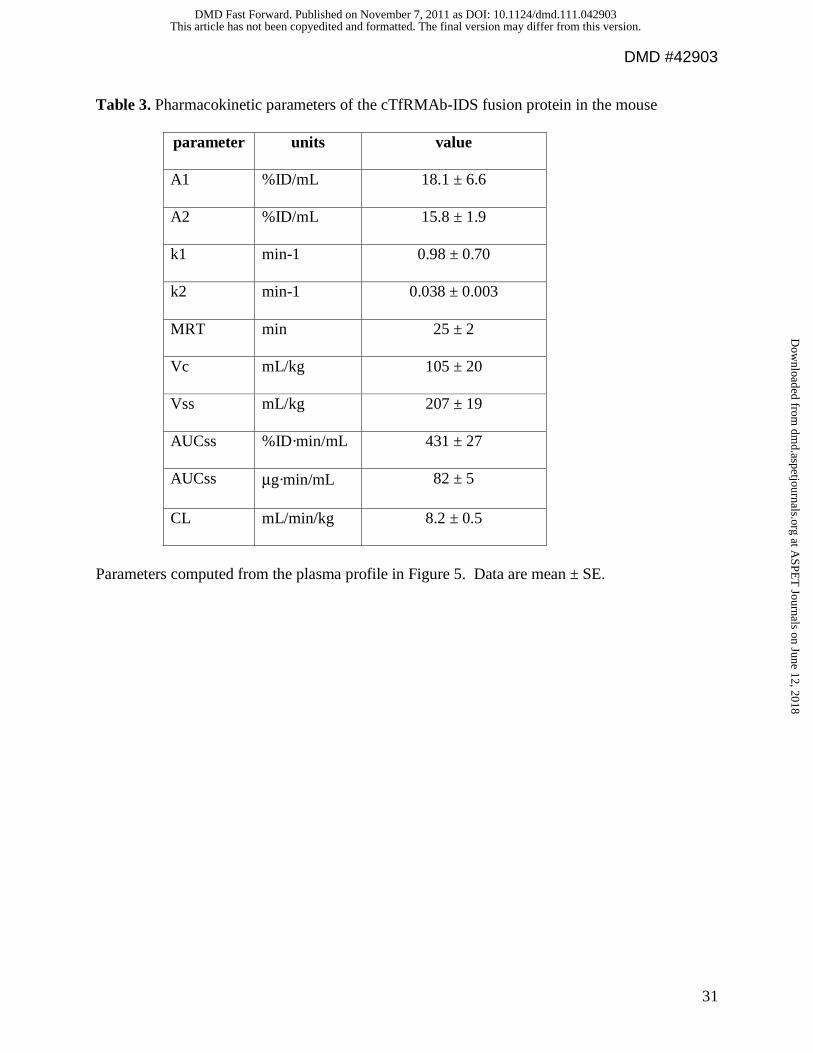
Fusion protein plasma pharmacokinetics in the mouse. The cTfRMAb-IDS fusion
protein was radiolabeled with the 3H-N-succinimidyl propionate reagent (Methods), and injected
into adult male C57Bl/6J mice via IV administration. The clearance of the [3H]-cTfRMAb-IDS
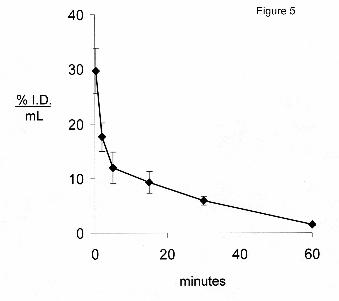
fusion protein from plasma after IV administration is plotted in Figure 5. The plasma
radioactivity decay curve was fit to a bi-exponential equation to yield the pharmacokinetics
parameters shown in Table 3. The cTfRMAb-IDS fusion protein was rapidly removed from
plasma (Figure 5). The plasma radioactivity that was precipitable by TCA was 88 ± 1, 83 ± 1, 79
± 1, 78 ± 3, 74 ± 2, and 50 ± 3 %, respectively at 0.25, 2, 5, 15, 30, and 60 min after IV injection.
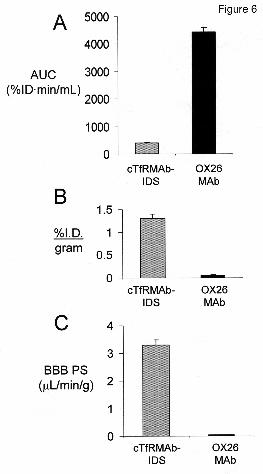
The rapid clearance of the fusion protein by plasma is also shown by the low 60 minute plasma
AUC compared to the AUC of the OX26 mouse MAb against the rat TfR (Figure 6A). The
OX26 MAb does not recognize the mouse TfR and is slowly cleared from plasma in the mouse
(Lee et al, 2000).
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
16
Fusion protein brain uptake in the mouse. The capillary depletion analysis showed the
VD of the cTfRMAb-IDS fusion protein in the brain homogenate was high, 377 ± 25 uL/gram
(Table 4). The VD of the fusion protein in the post-vascular supernatant, 102 ± 11 uL/gram, is
2.1-fold greater than the VD in the vascular pellet (Table 4), indicating the majority of the
cTfRMAb-IDS fusion protein had completed transcytosis through the BBB by 60 min after IV
injection.
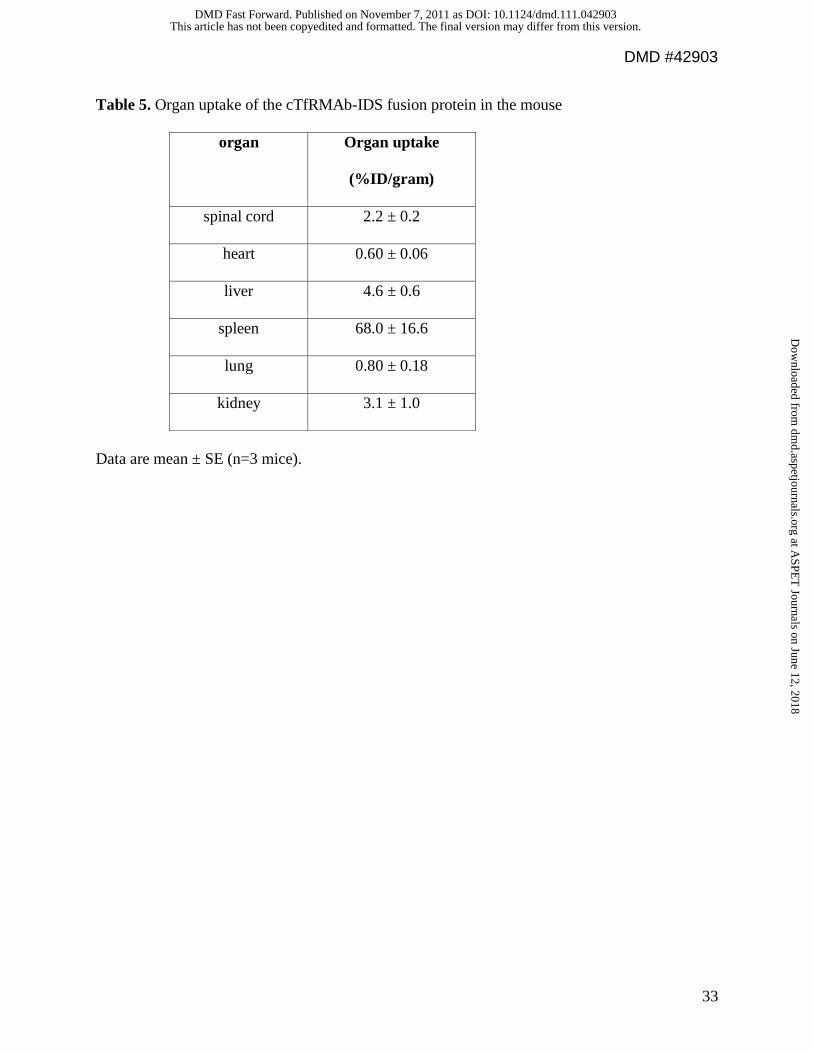
The uptake of the [3H]-cTfRMAb-IDS fusion protein by brain was determined at 60 min
after IV injection, and was 1.3 ± 0.1 %ID/gram brain (Figure 6B). The cTfRMAb-IDS fusion
protein was also taken up by spinal cord (thoracic-cervical), and the uptake was 2.2 ± 0.2
%ID/gram (Table 5). The BBB permeability-surface area (PS) product of the cTfRMAb-IDS
fusion protein in brain, which is equal to the ratio of the 60 min %ID/g (Figure 6B) and the 60
min plasma AUC (Figure 6A), is 3.3 ± 0.2 uL/min/g, which is high compared to the BBB PS
product for the OX26 MAb (Figure 6C). The cTfRMAb-IDS fusion protein was rapidly taken
up by peripheral tissues and the %ID/gram ranged from 0.60 ± 0.06 %ID/g for heart to 68 ± 17
%ID/g for spleen (Table 5).
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
17
Discussion
The results of these studies are consistent with the following conclusions. First, a new
IgG-IDS fusion protein has been engineered and expressed, the cTfRMAb-IDS fusion protein,
which is specific for the mouse and retains both high affinity binding to the mouse TfR (Figure
4) and high IDS enzyme activity (Tables 1-2). Second, the cTfRMAb-IDS fusion protein is
rapidly removed from blood (Figure 5, Table 3), and broadly distributed to brain, spinal cord,
and peripheral tissues (Figure 6, Table 5). Third, the cTfRMAb-IDS fusion protein undergoes
transcytosis across the BBB as shown by the capillary depletion method (Table 4), and
distributes to brain at rates that can restore a significant fraction of the IDS enzyme activity in
brain.
The cTfRMAb-IDS fusion protein is formed by fusion of the mature 525 AA human IDS
to the carboxyl terminus of each heavy chain of the cTfRMAb. Human IDS (Genbank
NP_000193) and murine IDS (Genbank NP_034628) have a high, 88%, AA identity. The
identity of the cTfRMAb-IDS fusion protein was confirmed by mouse IgG and human IDS
Western blotting (Figure 3), and the purity of the affinity purified fusion protein was confirmed
by reducing SDS-PAGE (Figure 1). Non-reducing SDS-PAGE shows the fusion protein exists as
a dimer at neutral pH. IDS and arysulfatase A (ASA) are members of a sulfatase family that form
dimers at neutral pH and octamers at acidic pH (Lukatela et al, 1998). These sulfatases require a
post-translational modification for enzyme activity, whereby SUMF1 converts Cys59 to N-formyl
glycine (Zito et al, 2005). Co-transfection with the SUMF1 gene and genes encoding the
HIRMAb-IDS fusion protein is not necessary to produce maximal enzyme activity in stably
transfected CHO cells (Lu et al, 2011). However, it is necessary to transiently co-transfect COS
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
18
cells with both the SUMF1 gene and the genes encoding the cTfRMAb-IDS fusion protein in
order to maximize IDS enzyme activity (Table 1).
The cTfRMAb-IDS fusion protein is rapidly removed from blood after IV injection with
a systemic clearance of 8.2 ± 0.5 mL/min/kg (Table 3) and a systemic volume of distribution,
Vss, that exceeds the central plasma volume, Vc (Table 3). The systemic clearance of the
HIRMAb-IDS fusion protein in the Rhesus monkey is 2.33 ± 0.04 mL/min/kg (Lu et al, 2011).
Similarly, lysosomal enzymes are rapidly removed from blood after IV injection. ASA, which
shares AA homology with IDS (Franco et al, 1995), has a plasma half-time of 40 minutes in the
mouse (Matzner et al, 2005). IDUA is rapidly removed from blood in the dog with a half-time of
<30 minutes (Shull et al, 1994). The rapid removal of the cTfRMAb-IDS fusion protein from
blood is associated with partial degradation of the fusion protein, as represented by the decrease
in plasma radioactivity that is precipitable by TCA (Results). However, previous work in the
primate used gel filtration chromatography of plasma removed 2 hours after intravenous
administration of the IgG-IDS to show the fusion protein in blood was intact (Lu et al, 2011).
The rapid clearance of the fusion protein from plasma parallels the widespread distribution of the
fusion protein into peripheral tissues (Table 5). The high uptake of the cTfRMAb-IDS fusion
protein by spleen (Table 5) is specific for the mouse, since the uptake of the HIRMAb-IDS
fusion protein in the Rhesus monkey by spleen is comparable to the uptake by liver (Lu et al,
2011). The weight of the spleen in the mouse is 81 mg (unpublished observations); therefore, the
total fraction of injected fusion protein taken up by the spleen in the mouse is <6 % of the
injected dose (Table 5). In contrast to the rapid removal of the cTfRMAb-IDS fusion protein
from blood in vivo (Figure 5), the clearance of the fusion protein from the intracellular
compartment is slow. The fusion protein-derived IDS enzyme activity remains elevated in
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
19
mouse fibroblasts for >96 hours after a single 2 hour exposure of the cells to the fusion protein
(Table 2), and the intracellular IDS enzyme activity decays with a half-time of 1.7 days
(Results).
The principal finding of this study is the high uptake of the fusion protein by brain. The
brain uptake of the cTfRMAb-IDS fusion protein by mouse brain is 1.3 ± 0.1 %ID/gram (Figure
6). In contrast, the brain uptake in the mouse of recombinant IDS is 100-fold lower, or 0.013%
ID (Garcia et al, 2007). The brain uptake in the mouse of the OX26 MAb, which is a brain blood
volume marker in the mouse, is 0.06% ID/g (Lee et al, 2000). Therefore, the mouse brain uptake
of IDS of only 0.013% ID is evidence that IDS does not cross the BBB, and is confined to the
blood volume of brain. The high rate of transport of the cTfRMAb-IDS fusion protein into brain
is caused by the high affinity binding of the fusion protein to the TfR on the BBB. The affinity of
the cTfRMAb-IDS fusion protein for the mouse TfR is equal to the affinity of the cTfRMAb
(Figure 4). Therefore, fusion of the IDS protein to the carboxyl terminus of the heavy chain of
the cTfRMAb has no effect on binding to the mouse TfR. The TfR expressed on the BBB is a
receptor-mediated transcytosis system, as demonstrated by electron microscopic immungold
studies (Bickel et al, 1994). In the present study, the capillary depletion method is used to
demonstrate BBB transcytosis (Table 4). Prior work has validated the capillary depletion method
as a measure of BBB transcytosis using emulsion autoradiography of brain (Skarlatos et al,
1995).
The brain uptake of the cTfRMAb-IDS fusion protein in the mouse is 1.3 ± 0.1
%ID/gram (Figure 6B) at an injection dose of 0.7 mg/kg (Methods). The IDS enzyme specific
activity of the fusion protein is 126 ± 1 units/ug protein, where 1 unit = 1 nmol/hr (Table 1).
Therefore, the IDS enzyme activity following the IV injection of a dose of 1 mg/kg of the
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
20
cTfRMAb-IDS fusion protein in a 30 gram mouse is 50 units/gram brain, which is equivalent to
0.5 units/mg protein, given 100 mg protein per gram brain. The endogenous IDS enzyme activity
in the mouse brain is 2.8 units/mg protein (Tomatsu et al, 2007). Therefore, a 1 mg/kg dose of
the cTfRMAb-IDS fusion protein is predicted to replace 18% of the endogenous IDS enzyme
activity in brain. This dosing is expected to be therapeutic in the brain, since the replacement of
only 1-2% of lysosomal enzyme activity reverses lysosomal inclusion lesions in tissues
(Muenzer and Fisher, 2004).
In summary, the cTfRMAb-IDS fusion protein is a new form of the IDS enzyme that is
brain penetrating in the mouse, and this study supports future treatment of the MPS-II mouse
model. Recent work described the treatment of the MPS-I mouse with the cTfRMAb-IDUA
fusion protein. Chronic treatment of 6 month old MPS-I mice with twice weekly IV injections of
1 mg/kg of the cTfRMAb-IDUA fusion protein caused a reduction in lysosomal inclusion bodies
in the brain, as well as reductions in GAG levels in peripheral tissues (Boado et al, 2011). The
MPS-I mice were treated twice weekly for 8 weeks with intravenous fusion protein, and this
treatment produced a low titer immune response without any limiting immune reactions (Boado
et al, 2011). The brains of humans born with MPS-I or MPS-II is potentially treatable with re-
engineered lysosomal enzymes that are brain-penetrating, such as the HIRMAb-IDUA and
HIRMAb-IDS fusion proteins.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
21
Acknowledgements
Winnie Tai and Phuong Tram provided technical assistance.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
22
Authorship Contributions
Participated in research design: Zhou, Boado, Lu, Hui, Pardridge
Conducted experiments: Zhou, Boado, Lu, Hui, Pardridge
Performed data analysis: Zhou, Boado, Lu, Hui, Pardridge
Wrote or contributed to the writing of the manuscript: Zhou, Boado, Lu, Hui, Pardridge
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
23
References
Al Sawaf S, Mayatepek E and Hoffmann B (2008) Neurological findings in Hunter disease:
pathology and possible therapeutic effects reviewed. J Inherit Metab Dis 31:473-480.
Bickel U, Kang YS, Yoshikawa T and Pardridge WM (1994) In vivo demonstration of
subcellular localization of anti-transferrin receptor monoclonal antibody-colloidal gold
conjugate within brain capillary endothelium. J Histochem Cytochem 42: 1493-1497.
Boado RJ, Zhang Y, Xia CF, Wang Y and Pardridge WM (2008) Genetic engineering of a
lysosomal enzyme fusion protein for targeted delivery across the human blood-brain
barrier. Biotechnol Bioeng 99:475-484.
Boado RJ, Zhang Y, Wang Y and Pardridge WM (2009a) Engineering and expression of a
chimeric transferrin receptor monoclonal antibody for blood-brain barrier delivery in the
mouse. Biotechnol Bioeng 102:1251-1258.
Boado RJ, Hui EK, Lu JZ and Pardridge WM (2009b) AGT-181: expression in CHO cells and
pharmacokinetics, safety, and plasma iduronidase enzyme activity in Rhesus monkeys. J
Biotechnol 144:135-141.
Boado RJ, Zhou QH, Lu JZ, Hui EK and Pardridge WM (2010) Pharmacokinetics and brain
uptake of a genetically engineered bifunctional fusion antibody targeting the mouse
transferrin receptor. Mol Pharm 7:237-244.
Boado RJ, Hui EK, Lu JZ, Zhou QH and Pardridge WM (2011) Reversal of lysosomal storage in
brain of adult MPS-I mice with intravenous Trojan horse-iduronidase fusion protein. Mol
PharmI 8: 1342-1350.
Franco B, Meroni G, Parenti G, Levilliers J, Bernard L, Gebbia M, Cox L, Maroteaux P,
Sheffield L, Rappold GA, Andria G, Petit C and Ballabio A (1995) A cluster of sulfatase
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
24
genes on Xp22.3: mutations in chondrodysplasia punctata (CDPX) and implications for
warfarin embryopathy. Cell 81:15-25.
Garcia AR, DaCosta JM, Pan J, Muenzer J and Lamsa JC (2007) Preclinical dose ranging studies
for enzyme replacement therapy with idursulfase in a knock-out mouse model of MPS II.
Mol Genet Metab 91:183-190.
Lee HJ, Engelhardt B, Lesley J, Bickel U and Pardridge WM (2000) Targeting rat anti-mouse
transferrin receptor monoclonal antibodies through blood-brain barrier in mouse. J
Pharmacol Exp Ther 292:1048-1052.
Lu JZ, Boado RJ, Hui EK, Zhou QH and Pardridge WM (2011) Expression in CHO cells and
pharmacokinetics and brain uptake in the Rhesus monkey of an IgG-iduronate-2-sulfatase
fusion protein. Biotechnol Bioeng 108: 1954-1964.
Lu JZ, Hui EK, Boado RJ and Pardridge WM (2010) Genetic engineering of a bifunctional IgG
fusion protein with iduronate-2-sulfatase. Bioconjug Chem 21:151-156.
Lukatela G, Krauss N, Theis K, Selmer T, Gieselmann V, von Figura K and Saenger W (1998)
Crystal structure of human arylsulfatase A: the aldehyde function and the metal ion at the
active site suggest a novel mechanism for sulfate ester hydrolysis. Biochemistry 37:3654-
3664.
Matzner U, Herbst E, Hedayati KK, Lullmann-Rauch R, Wessig C, Schroder S, Eistrup C,
Moller C, Fogh J and Gieselmann V (2005) Enzyme replacement improves nervous
system pathology and function in a mouse model for metachromatic leukodystrophy.
Hum Mol Genet 14:1139-1152.
Muenzer J and Fisher A (2004) Advances in the treatment of mucopolysaccharidosis type I. N
Engl J Med 350:1932-1934.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
25
Muenzer J, Wraith JE, Beck M, Giugliani R, Harmatz P, Eng CM, Vellodi A, Martin R,
Ramaswami U, Gucsavas-Calikoglu M, Vijayaraghavan S, Wendt S, Puga AC, Ulbrich
B, Shinawi M, Cleary M, Piper D, Conway AM and Kimura A (2006) A phase II/III
clinical study of enzyme replacement therapy with idursulfase in mucopolysaccharidosis
II (Hunter syndrome). Genet Med 8:465-473.
Pardridge WM, Kang YS, Buciak JL and Yang J (1995) Human insulin receptor monoclonal
antibody undergoes high affinity binding to human brain capillaries in vitro and rapid
transcytosis through the blood-brain barrier in vivo in the primate. Pharm Res 12:807-
816.
Shull RM, Kakkis ED, McEntee MF, Kania SA, Jonas AJ and Neufeld EF (1994) Enzyme
replacement in a canine model of Hurler syndrome. Proc Natl Acad Sci U S A 91:12937-
12941.
Skarlatos S, Yoshikawa T and Pardridge WM (1995) Transport of [125I]transferrin through the
rat blood-brain barrier. Brain Res 683:164-171.
Tomatsu S, Vogler C, Montano AM, Gutierrez M, Oikawa H, Dung VC, Orii T, Noguchi A and
Sly WS (2007) Murine model (Galns(tm(C76S)slu)) of MPS IVA with missense
mutation at the active site cysteine conserved among sulfatase proteins. Mol Genet Metab
91:251-258.
Triguero D, Buciak J and Pardridge WM (1990) Capillary depletion method for quantification of
blood-brain barrier transport of circulating peptides and plasma proteins. J Neurochem
54:1882-1888.
Voznyi YV, Keulemans JL, van Diggelen OP (2001) A fluorimetric enzyme assay for the
diagnosis of MPS II (Hunter disease). J Inherit Metab Dis 24: 675-680.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
26
Wraith JE, Scarpa M, Beck M, Bodamer OA, De Meirleir L, Guffon L, Meldgaard Lund A,
Malm G, Van der Ploeg AT and Zeman J (2008) Mucopolysaccharidosis type II (Hunter
syndrome): a clinical review and recommendations for treatment in the era of enzyme
replacement therapy. Eur J Pediatr 167: 267-277.
Zito E, Fraldi A, Pepe S, Annunziata I, Kobinger G, Di Natale PD, Ballabio A, and Cosma MP
(2005) Sulphatase activities are regulated by the interaction of sulphatase-modifying
factor 1 with SUMF2. EMBO Rep 6: 655-660.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
27
Legends to Figures
Figure 1. Reducing SDS-PAGE and Coomasie blue staining of 2 ug/lane of the cTfRMAb (lane
1), the cTfRMAb-IDS fusion protein (lane 2), and the HIRMAb-IDS fusion protein (lane 3). The
migration of MW standards is shown.
Figure 2. Non-reducing SDS-PAGE and Coomasie blue staining of 2 ug/lane of the cTfRMAb
(lane 1), the cTfRMAb-IDS fusion protein (lane 2), and the HIRMAb-IDS fusion protein (lane
3). The IgG-IDS fusion proteins migrate as dimers. The migration of MW standards is shown.
Figure 3. (Left panel) Western blotting examines the immunoreactivity of a primary antibody
against mouse IgG reacting with the cTfRMAb (lane 1), the cTfRMAb-IDS fusion protein (lane
2), and the HIRMAb-IDS fusion protein (lane 3). (Right panel) Western blotting examines the
immunoreactivity of a primary antibody against human IDS reacting with the HIRMAb-IDS
fusion protein (lane 1), the cTfRMAb-IDS fusion protein (lane 2), and cTfRMAb (lane 3). The
migration of MW standards is shown.
Figure 4. ELISA measures the absorbance at 405 nm, A405, relative to the concentration of the
cTfRMAb, the cTfRMAb-IDS fusion protein, or mouse IgG1κ. The EC50 was determined by
non-linear regression analysis (Methods). The EC50, in ng/mL, was converted to units of nM,
based on the MW of the cTfRMAb or cTfRMAb-IDS fusion protein determined by reducing
SDS-PAGE (Figure 1), which is 164 kDa and 324 kDa, respectively. Data are means of
triplicates that varied less than 10%.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
28
Figure 5. Plasma concentration, expressed as % of injected dose (I.D.)/mL, of the [3H]-
cTfRMAb-IDS fusion protein after intravenous injection in the mouse. Data are mean ± S.E.
(n=3 mice/point), and are derived from plasma TCA-precipitable radioactivity. A plasma
concentration of 30% I.D./mL corresponds to a plasma concentration of 5.6 ug/mL of the
cTfRMAb-IDS fusion protein, given an injection dose of 0.67 mg/kg (Methods).
Figure 6. Plasma area under the concentration curve (AUC) at 60 min after IV injection (A),
brain uptake or %I.D./gram (B), and the BBB permeability-surface area (PS) product (C) is
shown for the cTfRMAb-IDS fusion protein (gray bar) and the OX26 MAb against the rat TfR
(solid bar). The OX26 MAb does not recognize the murine TfR and is a blood volume marker
and does not penetrate the BBB in the mouse (Lee et al, 2000). The differences in the parameters
for the cTfRMAb-IDS fusion protein and the OX26 MAb were significant (P<0.005, Student’s t-
test).
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
DMD #42903
29
Table 1. IDS enzyme activity of IgG-IDS fusion proteins
Fusion
protein
Host cell SUMF1
co-transfection
IDS activity
(nmol/hr/μg)
HIRMAb-IDS CHO no 168 ± 1
cTfRMAb-IDS COS yes 126 ± 1
cTfRMAb-IDS COS no 25 ± 1
Mean ± SD.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
DMD #42903
30
Table 2. Intracellular IDS enzyme activity in 3T3 mouse fibroblasts treated with cTfRMAb-IDS
fusion protein
Hours after
treatment
Intracellular
IDS activity
(nmol/hr/mgp)
0 17.4 ± 2.5
2 85.4 ± 4.5
24 45.9 ± 2.9
48 39.1 ± 4.5
72 35.4 ± 5.4
96 28.8 ± 3.5
Fibroblasts were treated with 2 ug/mL of the cTfRMAb-IDS fusion protein added to the medium
for 2 hours, and then incubated in fusion protein-free medium for up to 96 hours. Mean ± SD
(n=4 dishes per time point).
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
DMD #42903
31
Table 3. Pharmacokinetic parameters of the cTfRMAb-IDS fusion protein in the mouse
parameter units value
A1 %ID/mL 18.1 ± 6.6
A2 %ID/mL 15.8 ± 1.9
k1 min-1 0.98 ± 0.70
k2 min-1 0.038 ± 0.003
MRT min 25 ± 2
Vc mL/kg 105 ± 20
Vss mL/kg 207 ± 19
AUCss %ID·min/mL 431 ± 27
AUCss μg·min/mL 82 ± 5
CL mL/min/kg 8.2 ± 0.5
Parameters computed from the plasma profile in Figure 5. Data are mean ± SE.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from

DMD #42903
32
Table 4. Capillary depletion analysis for brain uptake of the cTfRMAb-IDS fusion protein
Fraction VD
(μL/g)
Brain homogenate 377 ± 25
Post-vascular supernatant 102 ± 11
Vascular pellet 49 ± 10
Mean ± S.E. (n=3 mice). The fusion protein was administered by IV injection, and brain
measurements made 60 min following injection.
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
DMD #42903
33
Table 5. Organ uptake of the cTfRMAb-IDS fusion protein in the mouse
organ Organ uptake
(%ID/gram)
spinal cord 2.2 ± 0.2
heart 0.60 ± 0.06
liver 4.6 ± 0.6
spleen 68.0 ± 16.6
lung 0.80 ± 0.18
kidney 3.1 ± 1.0
Data are mean ± SE (n=3 mice).
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.DMD Fast Forward. Published on November 7, 2011 as DOI: 10.1124/dmd.111.042903
at ASPE
T Journals on June 12, 2018
dmd.aspetjournals.org
Dow
nloaded from









![The Biological Roles of Steroid Sulfonation...3. Steroid sulfatase In humans, 17 sulfatases have been identified [46], of which steroid sulfatase (aka STS or ar‐ yl sulfatase C)](https://static.fdocuments.in/doc/165x107/602528afd20a4009a77bb85b/the-biological-roles-of-steroid-sulfonation-3-steroid-sulfatase-in-humans.jpg)








