
BIOSYNTHESIS AND METABOLISM OF ASCORBIC ACID IN PLANTS
17
BIOSYNTHESIS AND METABOLISM OF ASCORBIC ACID IN PLANTS* Frank A. Loewus,t George Wagner,$ and Joan C. Yang Division of Cell and Molecular Biology State University of New York ut Buffalo Buffalo, New York 14214 INTRODUCTION Formation, function, and fate of L-ascorbic acid in plants and microorga- nisms remain topics of great challenge for today's biochemist. There has been some progress. Hexose is generally regarded as the ultimate source of the carbon chain, but neither intermediates nor enzymes operative in the conversion have been pinpointed and explored. Ascorbic acid appears to be ubiquitous in the plant world, yet a systematic survey of its presence and identity using modern methods of assay remains to be accomplished. The widespread occur- rence of ascorbic acid implies a functional role or roles of broad significance, yet the nature of such role(s) are matters of speculation rather than of fact. Information regarding the fate of ascorbic acid is just beginning to appear, an3 its appearance offers promise of fresh new ways in which to approach the other topics, biosynthesis and activity. I wish to review here our recent work on the relationship between L- ascorbic acid metabolism in plants and (+)-tartaric acid and oxalic acid forma- tion. This relationship appears to hold the key to ascorbic acid biosynthesis, which I regard as an essential matter if the functional role of ascorbic acid in plants is to be fully understood. TARTARIC ACID BIOSYNTHESIS FROM ASCORBIC ACID When Herbert et a1.l published the constitution of L-ascorbic acid in 1931, isolation of equivalent amounts of oxalate and tartrate following oxidation, as illustrated in FIGURE 1, supplied evidence of structure. All three, ascorbic acid, oxalate, and tartrate, occur in plants, often together. The latter two are also produced by certain bacteria. That L-ascorbic acid or a closely related sub- stance might be a precursor of (+) -tartaric acid was suggested by Hough and Jones,? who relied on the configurational similarity between carbons 4 and 5 of L-ascorbic acid and carbons 2 and 3 of (+)-tartaric acid. This possibility was first tested by feeding L-ascorbic acid-6-l-'C to a detached grape leaf.3 Tartaric acid was isolated after 25 hours of metabolism and found to be virtually i2 This work was supported by National Institutes of Health research grant GM- Present address: Department of Agricultural Chemistry, Washington State Uni- t Present address: Biology Department, Brookhaven National Laboratory, Upton, 12422 and a grant from Hoffmann-La Roche, Inc., Nutley, New Jersey. versity, Pullman, Washington 99163. New York 11973. 7
Transcript of BIOSYNTHESIS AND METABOLISM OF ASCORBIC ACID IN PLANTS

BIOSYNTHESIS AND METABOLISM OF ASCORBIC ACID IN PLANTS*
Frank A. Loewus,t George Wagner,$ and Joan C. Yang
Division of Cell and Molecular Biology State University o f New York ut Buffalo
Buffalo, New York 14214
INTRODUCTION
Formation, function, and fate of L-ascorbic acid in plants and microorga- nisms remain topics of great challenge for today's biochemist. There has been some progress. Hexose is generally regarded as the ultimate source of the carbon chain, but neither intermediates nor enzymes operative in the conversion have been pinpointed and explored. Ascorbic acid appears to be ubiquitous in the plant world, yet a systematic survey of its presence and identity using modern methods of assay remains to be accomplished. The widespread occur- rence of ascorbic acid implies a functional role or roles of broad significance, yet the nature of such role(s) are matters of speculation rather than of fact. Information regarding the fate of ascorbic acid is just beginning to appear, an3 its appearance offers promise of fresh new ways in which to approach the other topics, biosynthesis and activity.
I wish to review here our recent work on the relationship between L- ascorbic acid metabolism in plants and (+)-tartaric acid and oxalic acid forma- tion. This relationship appears to hold the key to ascorbic acid biosynthesis, which I regard as an essential matter if the functional role of ascorbic acid in plants is to be fully understood.
TARTARIC ACID BIOSYNTHESIS FROM ASCORBIC ACID
When Herbert et a1.l published the constitution of L-ascorbic acid in 1931, isolation of equivalent amounts of oxalate and tartrate following oxidation, as illustrated in FIGURE 1 , supplied evidence of structure. All three, ascorbic acid, oxalate, and tartrate, occur in plants, often together. The latter two are also produced by certain bacteria. That L-ascorbic acid or a closely related sub- stance might be a precursor of (+) -tartaric acid was suggested by Hough and Jones,? who relied on the configurational similarity between carbons 4 and 5 of L-ascorbic acid and carbons 2 and 3 of (+)-tartaric acid. This possibility was first tested by feeding L-ascorbic acid-6-l-'C to a detached grape leaf.3 Tartaric acid was isolated after 25 hours of metabolism and found to be virtually
i2 This work was supported by National Institutes of Health research grant GM-
Present address: Department of Agricultural Chemistry, Washington State Uni-
t Present address: Biology Department, Brookhaven National Laboratory, Upton,
12422 and a grant from Hoffmann-La Roche, Inc., Nutley, New Jersey.
versity, Pullman, Washington 99163.
New York 11973.
7

8 Annals New York Academy of Sciences
H- C COOH I I
HO-C-H HO-C-H H-C-OH HNO H-C-OH I I I
HO-C-H I
CHzOH CH20H
CH20H h0OH
Acid L- Ascorbic L- Threonic (+)-Tartaric
- -SHo- t -H
L Dehydro-
Acid Acid Acid FIGURE 1. Chemical conversion of L-ascorbic acid to (+)-tartaric acid and oxalic
acid?
devoid of label. Most of the carbon-14 was recovered as carbohydrate, mainly sucrose, glucose, and fructose.
In 1969, Saito and Kasaid made the interesting discovery that young grape berries converted L-ascorbic acid- 1 -1.C to tartaric acid in high isotopic yield, 72% of the soluble label was recovered as tartaric acid after 24 hours of metabolism. Most of the label appeared in carboxyl carbon of the acid. D- Glucuron0-6,3-lactone-6-~~C was one-third to one-half as effective and D-glu- curonate-6-IT only one-tenth as effective as L-ascorbic acid-1 -I4C. D-Glu- curono-6,3-lactone is readily converted 6 to L-ascorbic acid by plants, but D-glucuronate is poorly converted, if at all." Saito and Kasai's observations has been confirmed in this laboratory with detached grape berries obtained froin a closely related species of grape as well as detached leaves of a more distantly related species of the Vitaceae.7 A portion of our results is presented in TABLE 1. Comparison of distribution of label among plant constituents after a feeding of L-ascorbic acid-1-W or -6J4C revealed that only the former source of label
TABLE 1
FRACTION OF DETACHED Vitis Iabrusca BERRIES AND Parthenocissus insertu LEAVES 24 HOURS AFTER LABELING
DISTRIBUTION OF RADIOACTIVITY AMONG COMPONENTS IN WATER-SOLUBLE
Ascorbic Acid- 1-"C Ascorbic Acid-6-l'C V . lab. P. ins. V . lab. P . ins.
% of soluble radioactivity Tartaric acid 69.0 38.4 1.4 0.6 Oxalic acid 0.6 0.6 0.4 0.5 Other acids 10.2 30.2 33.8 50.8 Sugars and neutrals 3.9 11.4 45.3 38.4 Other compounds 16.3 19.4 19.5 9.7
_~ -- . -~ __________________

Loewus et al. : Biosynthesis and Metabolism 9
resulted in significant incorporation of radioisotope into tartaric acid. Virtually all of the ''C in ascorbic acid-l-lC-derived tartaric acid was in the carboxyl carbon. When ascorbic acid-6-1% was supplied to these plants, about 40% of the soluble label appeared as sugar, mainly sucrose, confirming the earlier report." Labeled oxalic acid was not found in these experiments. Our findings, which support those of Saito and Kasai, indicate that carbon 1 of L-ascorbic acid becomes a carboxyl function in (+)-tartaric acid, and, by inference, car- bons 1 through 4 of L-ascorbic acid provide the carbon chain of (+)-tartaric acid (FIGURE 2) .
This grape study prompted investigation of another type of (+)-tartaric acid accumulator, the geranium. We chose Pelargoniurn crispurn, a plant that accumulates about 10 mg of tartaric acid per g fresh weight of leaf. Experi- ments were designed in which detached vegetative tips (apices) imbibed the radioactive compound for a short period and then were left to metabolize the labeled material for a set period of time.h As before, comparison was made on distribution of label among plant constituents after feeding L-ascorbic acid-l- 14C and -6-14C. In addition, experiments were run with L-galactono-1 ,4-lactone- UL-l'C and - 6 - ' C The latter is a potent precursor of L-ascorbic acid in
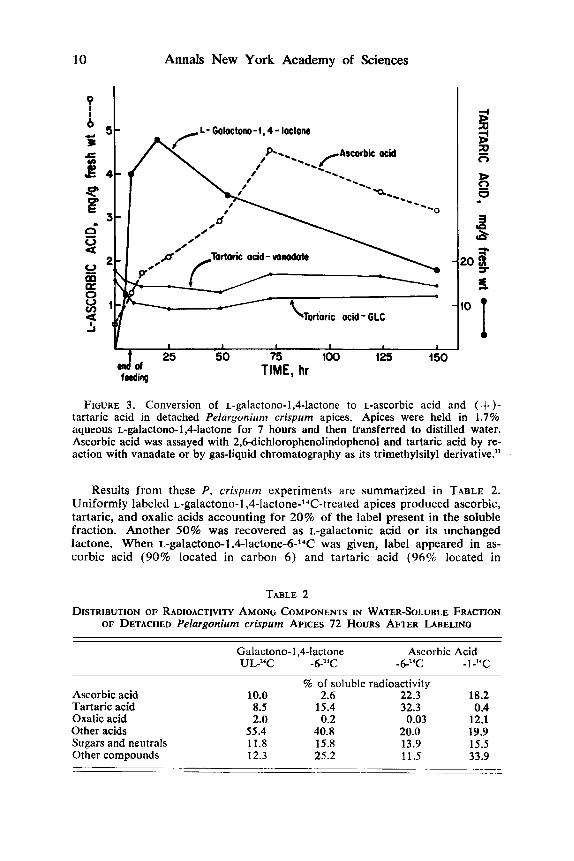
In P. crispurn apices that had been held in 1.7% L-galactono-l,4-lactone for 7 hours, then transferred to water, underwent a dramatic increase in ascorbic acid content, reaching nearly 10 times the normal level in 3 days then slowly declining over the next 3 days to a level still 6 times above the control (FIGURE 3 ) . In the same period, tartaric acid increased only slightly. It should be noted that the endogenous level of tartaric acid was about 10 times than of ascorbic acid at the start of the experiment, and any mctabolic contribution from ascorbic acid that had been generated from an exogenous source would be small compared to the endogenous tartaric acid pool. There may be some significance in the fact that the rapid decline in tartaric acid content following detachment was arrested by ascorbic acid feeding. Tartaric acid was assayed by 2 methods, vanadate colorimetric procedure and gas-liquid chromatography of the trimethylsilyl derivative.
'FOOH H-C-OH
HO-C-H I COOH
A I
(+)-Tartaric I
HO-f-H _ _ -*
.CH~ OH
L -Ascorbic acid
F I G U R E 2. Metabolism of L-ascorbic acid in the Vitaceae.
10 Annals New York Academy of Sciences
,L I .flC ,Tartaric acid- vanodok \
0
\Tortoric acid - GLC
I I 1 I I I I
andt feeding d TIME, hr 25 50 75 1 0 0 125 150
FIGURE 3. Conversion of ~-galactono-l,4-lactone to L-ascorbic acid and ( + )- tartaric acid in detached Prlurgoniurn crispum apices. Apices were held in 1.7% aqueous ~-galactono-l,4-lactone for 7 hours and then transferred to distilled water. Ascorbic acid was assayed with 2,f%dichlorophenolindophenol and tartaric acid by re- action with vanadate or by gas-liquid chromatography as its trimethylsilyl derivative.”
Results from these P. crispum experiments are summarized in TABLE 2. Uniformly labeled L-galactono-l,4-1a~tone-*~~C-treated apices produced ascorbic, tartaric, and oxalic acids accounting for 20% of the label present in the soluble fraction. Another 50% was recovered as L-galactonic acid or its unchanged lactone. When L-galactono-l+la~tone-6-’~C was given, label appeared in as- corbic acid (90% located in carbon 6 ) and tartaric acid (96% located in
TABLE 2 DISTRIBUTION OF RADIOACTIVITY AMONG COMPONENTS IN WATER-SOLUBLE FRACTION
OF DETACHED Pelurgonium crispum APICES 72 HOURS AFTER LABELING
Galactono-l,44actone Ascorbic Acid UL-I4C -fjF1’C -(j-”C - 1 - 1 4 ~
% of soluble radioactivity Ascorbic acid 10.0 2.6 22.3 18.2 Tartaric acid 8.5 15.4 32.3 0.4 Oxalic acid 2.0 0.2 0.03 12.1 Other acids 55.4 40.8 20.0 19.9 Sugars and neutrals 11.8 15.8 13.9 15.5 Other compounds 12.3 25.2 11.5 33.9

Loewus et al. : Biosynthcsis and Metabolism 1 1
carboxyl carbon) but none was found in oxalic acid, findings which suggested that the carbon chain of tartaric acid originated from the bottom 4 carbons of L-ascorbic acid. This was confirmed by an L-ascorbic acid-6-' 'C experiment. Of the label present as soluble constituents i n the lattcr, 32%) was (+)-tartaric acid (99% located in carboxyl carbon). Oxalic acid was devoid of label. Wheii L-ascorbic acid-l-*'C was supplied to f. crispirrn apices, appearance of label in (+)-tartaric acid and oxalic acid was reversed, now label appeared in oxalic acid but none in tartaric acid.
In a more detailed study, incorporation of label from L-ascorbic acid-6-"C into P. crispurri was examined at intervals from 4 to 72 hours. Profiles of the soluble radioactivity eluted from Dowex 1 (formate) exchange columns are given in FIGURE 4. Radioactivity in ascorbic acid decreased with time while that in tartaric acid increased. A small radioactive peak I1 appeared in the eluate in the region characterized by threonic acid. Other peaks, 1, 111, and IV, were also examined by paper chromatography, but their identities remain ten- tative.
- m 52 X v - E E \
Q V - >- c
v a 0
a Q a
0 to 0.1 N Formic acid *kFormic acid *I 0.1 to 4N
Tortoric "1 72hr A / ; r y b i c ~ , acid\ 4 1 4
0
1 ° 4 0
lo* 0
I 0 i 0 140 280 420 560 700 840
ELUATE, ml.
FIGURE 4. Profiles of radioactivity in eluates of Dowex-1 (formate) ion exchange chromatography after feeding L-ascorbic acid-6-l'C to Pelurgoniurn crispurn-a time- course study.
12 Annals New York Academy of Sciences
0 to 0.1 N Formic acid +Formic acid +I 0.1 to 4N
0 140 280 420 560 700 840
ELUATE, ml FIGURE 5 . Profiles of radioactivity in eluates of Dowex-1 (formate) ion-exchange
chromatography after feeding L-ascorbic acid- 1 -“C to Pelurgonium crispurn-+ time- course study.
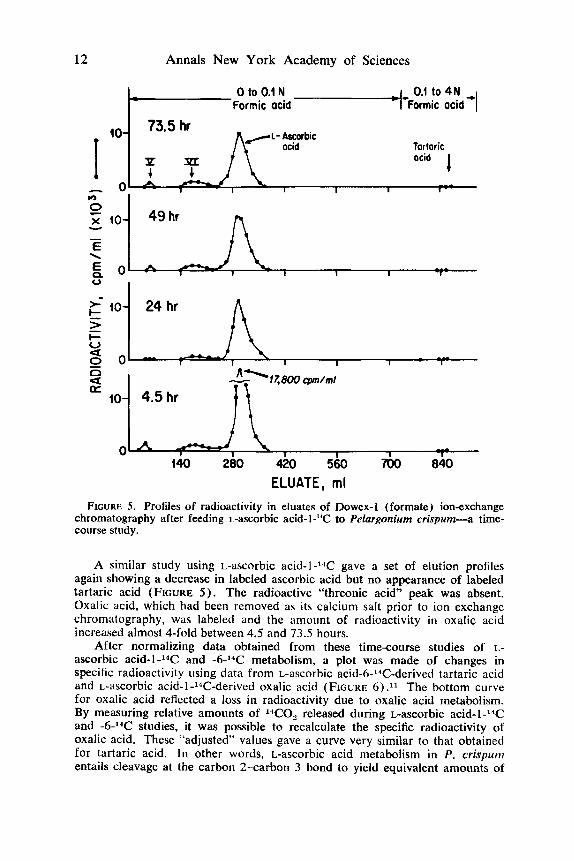
A similar study using L-ascorbic acid-I-”C gave a set of elution profiles again showing a decrease in labeled ascorbic acid but no appearance of labeled tartaric acid (FIGURE 5 ) . The radioactive “threonic acid” peak was absent. Oxalic acid, which had been removed as its calcium salt prior to ion exchange chromatography, was labeled and the amount of radioactivity in oxalic acid increased almost %fold between 4.5 and 73.5 hours.
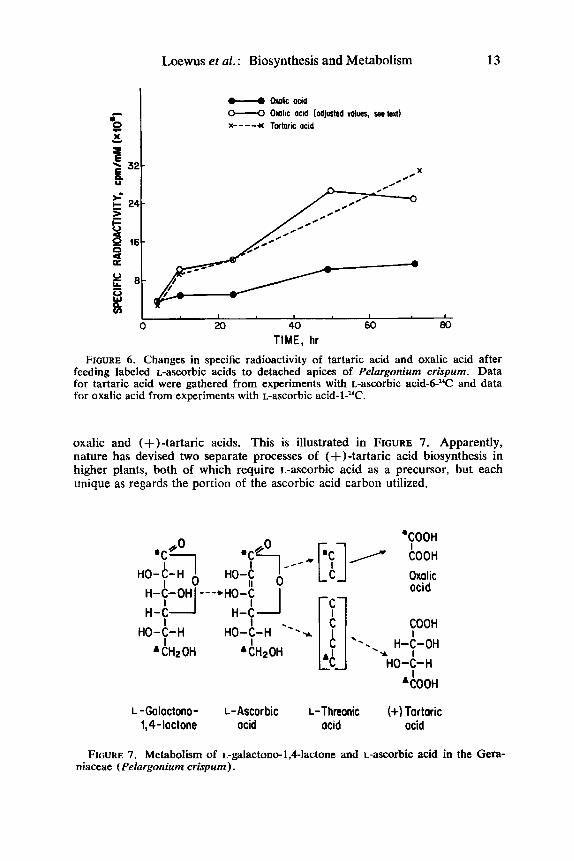
After normalizing data obtained from these time-course studies of L- ascorbic acid-1-l4C and -6-l‘C metabolism, a plot was made of changes in specific radioactivity using data from L-ascorbic acid-6-I4C-derived tartaric acid and L-ascorbic acid-1-*’C-derived oxalic acid (FIGURE 6) .ll The bottom curve for oxalic acid reflected a loss in radioactivity due to oxalic acid metabolism. By measuring relative amounts of I4C0, released during L-ascorbic acid-l-l’C and -6-l’C studies, it was possible to recalculate the specific radioactivity of oxalic acid. These “adjusted” values gave a curve very similar to that obtained for tartaric acid. In other words, L-ascorbic acid metabolism in P. crispum entails cleavage at the carbon 2-carbon 3 bond to yield equivalent amounts of
Loewus et al.: Biosynthesis and Metabolism
0- s x J E
-
13
C---. Orolic acid W Orplic acb (adjusted values, saetert) x----U Tartaric acid
TIME, hr FIGURE 6. Changes in specific radioactivity of tartaric acid and oxalic acid after
feeding labeled L-ascorbic acids to detached apices of Pelargonium crispum. Data for tartaric acid were gathered from experiments with L-ascorbic acid-6-"C and data for oxalic acid from experiments with L-ascorbic acid-1-"C.
oxalic and (+)-tartaric acids. This is illustrated in FIGURE 7. Apparently, nature has devised two separate processes of (+)-tartaric acid biosynthesis in higher plants, both of which require L-ascorbic acid as a precursor, but each unique as regards the portion of the ascorbic acid carbon utilized.
HO-C-H
H-C I
HO-C-H I CHz OH
--b
h] p] 4c I
'$OOH / COOH
Oxolic ocid
COOH I .--. H-C-OH
n I HO-C-H
+OOH
L-Goloctono- L-Ascorbic L-Threonic (+) Tortoric 1,440ctone ocid ocid ocid
FIGURE 7. Metabolism of ~-galactono-l,4-lactone and L-ascorbic acid in the Gera- niaceae (Pelargonium crispurn).

14 Annals New York Academy of Sciences
OXAZSC ACID BIOSYNTHESIS FROM ASCORBIC ACID
Conversion of L-ascorbic acid-1-W into labeled oxalic acid by P. crispum apices revealed a process of oxalic acid biosynthesis previously unconsidered in studies of oxalic acid formation in plants. Millard et al.'Z regarded glycolate and glyoxylate as intermediate precursors of oxalate in OxaIis pes-cuprue, con- sistent with formation of glyoxylate from isocitrate by action of isocitratase. Chang and Beevers l 3 obtained an enzyme that converted oxaloacetate to oxalate and acetate from extracts of red beet roots and young spinach leaves. Neither Millard et al. nor Chang and Beevers included ascorbic acid in their studies.
In animals, oxalate has been identified as a metabolic product of 1.-ascorbic, dehydroascorbic, and diketogulonic acids. This was first demonstrated in a conclusive way by Burns et al.I.1 during studies of L-ascorbic acid-l-l.*C metabo- lism in guinea pigs. Later, this observation was extended to include the rat and
Oxalate production was considered to be the result of nonenzymatic decomposition of diketogulonic acid, the end product of L-ascorbic acid oxida- tion.
Prompted by the P. crispmi results, recent efforts in this laboratory have been directed toward a study of the role of L-ascorbic acid in oxalate formation in certain plants known to be oxalate accumulators.l!'. 21-2:i Tissues chosen for this study included 12- to 14-day spinach seedlings (Spinuceu oleraceu, var. Bloomsdale), common yellow wood sorrel (Oxalis stricta L.), and whitc wood sorrel (Oxulis oregano). The latter was provided by Dr. Michael Cousens, University of Oregon. Methods were essentially those described earlier.', Greenhouse-grown spinach seedlings were detached from their roots and placed in vials containing the radioactive solution. Wood-sorrel leaves in their final stage of development or at the mature stage were detached from greenhouse- grown plants and placed in vials with petioles immersed in the radioactive solu- tion. Oxalic acid was recovered from the water-soluble fraction of plant extracts by precipitation as calcium oxalate. The crude oxalate was then acidified, dried, and purified by sublimation. The radioisotopic purity of labeled ascorbic, de- hydroascorbic, and diketogulonic acids used in these experiments was determined by means of thin-layer chromatography on cellulose or polyamide.2'
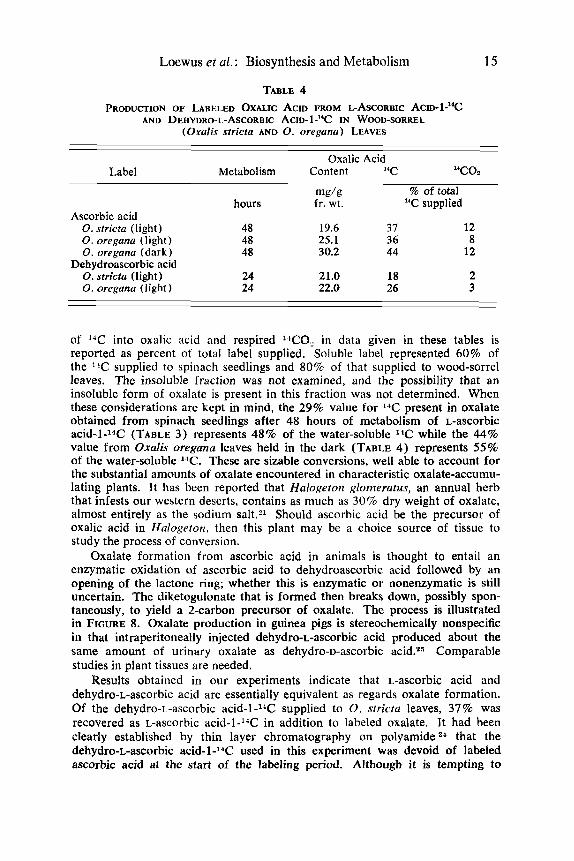
Labeled oxalic acid was a major product of i,-ascorbic acid-l-*C and dehydro-L-ascorbic acid-1-"C in both spinach seedlings and wood-sorrel leaves. Results from several experiments are given in TABLES 3 and 4. Incorporation
TABLE 3 PRODIJCIION OF LABELED OXALIC ACID FROM L-ASCORBIC A c l ~ l - ~ ' C ,
DEHYDRO-L-ASCORRIC ACID-1 -32, AND DIKETO-L-GULONATE-I-~~C IN SPINACH (Spinuceu olcruceu) SEEDLINGS
Oxalic Acid Label Metabolism Content "C 14c02
mg/g % of total hours fr. wt. "C supplied
Ascorbic acid 24 20 23 1.3 17 48 1.8 29 24
Dehydroascorbic acid 24 1.3 25 14 Diketogulonate 24 1.8 1.7 19
. _ _ _ _ _ ~ ~
Loewus et al.: Biosynthesis and Metabolism 15
TABLE 4 PRODUCTION OF LABELED OXALIC ACID FROM L-ASCORBIC ACID-~-"C
AND DEHYDRO-L-ASCORBIC ACID- 1 -"C IN WOOD-SORKEL (Oxalis srricta AND 0. orrgana) LEAVES
Oxalic Acid Label Metabolism Content 14C 1 4 ~ 0 ~
m d g % of total hours fr. wt. "C supplied
Ascorbic acid 0. stricta (light) 48 19.6 37 12
0. orrgana (dark) 48 30.2 44 12
0. stricta (light) 24 21.0 18 2 0. oregana (light) 24 22.0 26 3
0. oregana (light) 48 25.1 36 8
Dehydroascorbic acid
of "C into oxalic acid and respired lC0, in data given in these tables is reported as percent of total label supplied. Soluble label represented 60% of the I'C supplied to spinach seedlings and 80% of that supplied to wood-sorrel leaves. The insoluble fraction was not examined, and the possibility that an insoluble form of oxalate is present in this fraction was not determined. When these considerations are kept in mind, the 29% value for 14C present in oxalate obtained from spinach seedlings after 48 hours of metabolism of L-ascorbic acid-1-l4C (TABLE 3) represents 48% of the water-soluble IIC while the 44% value from Oxalis oregana leaves held in the dark (TABLE 4) represents 55% of the water-soluble "C. These are sizable conversions, well able to account for the substantial amounts of oxalate encountered in characteristic oxalate-accumu- lating plants. It has been reported that Halogeton glomeratus, an annual herb that infests our western deserts, contains as much as 30% dry weight of oxalate, almost entirely as the sodium salt.?l Should ascorbic acid be the precursor of oxalic acid in Halogeton, then this plant may be a choice source of tissue to study the process of conversion.
Oxalate formation from ascorbic acid in animals is thought to entail an enzymatic oxidation of ascorbic acid to dehydroascorbic acid followed by an opening of the lactone ring; whether this is enzymatic or nonenzymatic is still uncertain. The diketogulonate that is formed then breaks down, possibly spon- taneously, to yield a 2-carbon precursor of oxalate. The process is illustrated in FIGURE 8. Oxalate production in guinea pigs is stereochemically nonspecific in that intraperitoneally injected dehydro-L-ascorbic acid produced about the same amount of urinary oxalate as dehydro-D-ascorbic acid.gs Comparable studies in plant tissues are needed.
Results obtained in our experiments indicate that L-ascorbic acid and dehydro-L-ascorbic acid are essentially equivalent as regards oxalate formation. Of the dehydro-L-ascorbic acid-l-l'C supplied to 0. stricta leaves, 37% was recovered as L-ascorbic acid-l-l'c in addition to labeled oxalate. It had been clearly established by thin layer chromatography on polyamide z* that the dehydro-L-ascorbic acid-1-1 1C used in this experiment was devoid of labeled ascorbic acid at the start of the labeling period. Although it is tempting to

16 Annals New York Academy of Sciences
HO-$-H CH2OH
fOOH
c=o f= 0 7'0
I - - H- $ H-$-OH
fOOH
c= 0 I I
H- $ H-$-OH
7'0 - - H O - ~ - H HO-C- t i
I CH2 OH CH20H
COOH
I I I I I I
L- Ascorbic Dehydro- 2,3-Diketo- I L-T hreonic Acid L-Ascorbic L-Gulonic I Acid or other
Acid Acid products I I _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ I
FIGURE 8. Oxalic acid formation from ascorbic acid as postulated in animal sys- tems. The bypass indicated by a dashed line has been proposed by Baker et al."
suggest that dehydro-L-ascorbic acid is intermediate in the conversion of L- ascorbic acid to oxalic acid, such speculation is premature.
2,3-Diketo-~-gulonate-l-~'C, prepared from dehydro-L-ascorbic acid-1 -l'C just prior to use by titration with NaOH, failed to produce labeled oxalate in spinach seedlings (TABLE 3). Recently, the experiment was repeated in detached 0. oreguna leaves, with similar results. Plant tissues may be unable to utilize diketogulonate as an oxalate precursor, but the possibility also exists that de- tached plants have a vascular or cellular barrier that prevents diketogulonate from reaching the site of oxalate synthesis.
Although these experiments do not provide a clear picture of the path of conversion from ascorbic acid and dehydroascorbic acid to oxalic acid, they do demonstrate the existence of a conversion process from the former 2 compounds to the latter and as such provide an experimental approach for the study of ascorbic acid biosynthesis by way of this metabolic product.
CONVERSION OF DGLUCOSE TO L-ASCORBIC ACID AND ITS METABOLIC PRODUCTS, (+)-TARTARIC ACID AND OXALIC ACID
L-Ascorbic acid is a product of hexose phosphate metabolism in plants, but its path of biosynthesis is still unresolved.2* Earlier proposals,27 which compare ascorbic acid formation in plants to that in animals in which ~-giucurono-6,3- lactone and L-gulono-l,4-lactone are regarded as intermediates between D glucose and L-ascorbic acid,28 fall short in that these schemes do not take into consideration a dichotomy encountered when D-glucose is oxidized to the level of a sugar acid by plants. This is illustrated in FIGURE 9. On the one hand, higher plants convert D-glucose to D-glucuronic acid and its metabolic products by oxidation of carbon 6. Oxidation occurs in one of two possible ways, either the sugar nucleotide oxidation pathway with UDP-Dglucose as an intermedi- ate 29 or the myo-inositol oxidation pathway.3o On the other hand, it is carbon 1 of Dglucose or a related intermediate that is oxidized during L-ascorbic acid biosynthesis.20.31,32 When labeling patterns in pectin-derived D-galacturonate, L-ascorbic acid, and sucrose-derived D-glucose were compared after feeding

Locwus et al. : Biosynthesis and Metabolism 17
I
I -G-
H&-0- Hob-0- 6OOH
Ascorbic Acid Hexose Precursor Uronic Acid Products Products
FIGURE 9. Oxidation of hexose to uronic acid products or ascorbic acid products. (From Loewtis.'E By permission of P/iytoc/io,zi.slr.y.)
rrglucose-6-1'C to detached strawberry fruits, it was found that all 3 products retained about 80% of their incorporated label in carbon 6.:12 In another set of experiments involving the feeding of D-glucose-1 - I 'C to detached parsley leaves, equivalent results were obtained,:':) that is, about 80% of the incorporated label appeared in carbon I of pectin-derived D-galacturonate, L-ascorbic acid, and sucrose-derived wglucose. It should be emphasized that all products were isolated from 1 source of labeled tissue in each example. These experiments clearly illustrate the dichotomy seen in FIGURE 9.
Probably the most disturbing feature of L-ascorbic acid formation in plants has been the knowledge that ~-glucurono-6,3-lactone. n-galacturonate, L-gulono- 1 ,Clactone, and especially ~-galactono-l,4-lactone are readily converted to L- ascorbic acid .ll, 27;. :I1 when thcse compounds are provided exogenously. At- tempts to fit this information into a scheme of L-ascorbic acid biosynthesis analogous to the one proposed for animals have met with little success. For the reasons just discussed, D-glucuronic acid or ~glucurono-6,3-lactone is prob- ably not normally required in conversion of hexose to ascorbic acid even though plants contain an oxidoreductase capable of reducing ~-gluciirono-6,3- lactone to ~-giilono-l,4-lactone. Part of the preoccupation with such schemes in which a D - ~ I U C O or D-galacto configuration is inverted to a L-gulo or L-galacto configuration has been the need to provide an accounting for the I. configuration of naturally occurring ascorbic acid. This matter has been discussed in a recent
Hannincn et al."' have reported that hen kidney slices were able to syn- thesize L-ascorbic acid from myo-inositol as well as from D-glucuronic acid, D-g~ucurono-6,3-Iactone, and L-gulono- I ,4-lactonc. Their report further high- lights a basic difference between animals and plants. In plant tissues, a major portion of exogenously supplicd rnyo-inositol was oxidized to n-glucuronic acid. The latter was rapidly converted to other products, particularly cell-wall uronic acid and pentose unjts.21;. Since myo-inositol is formed from D-glucose without disturbing the carbon chain sequence R ( i and oxidative cleavage of rriyo-inositol opens the cyclitol ring at the samc carbon-carbon bond formed during cyclization of D - ~ ~ U C O S C to inyo- inositol,37 it follows that wglucuronic acid produced in the myo-inositol oxida-
:W L-Ascorbic acid was not among those
18 Annals New York Academy of Sciences
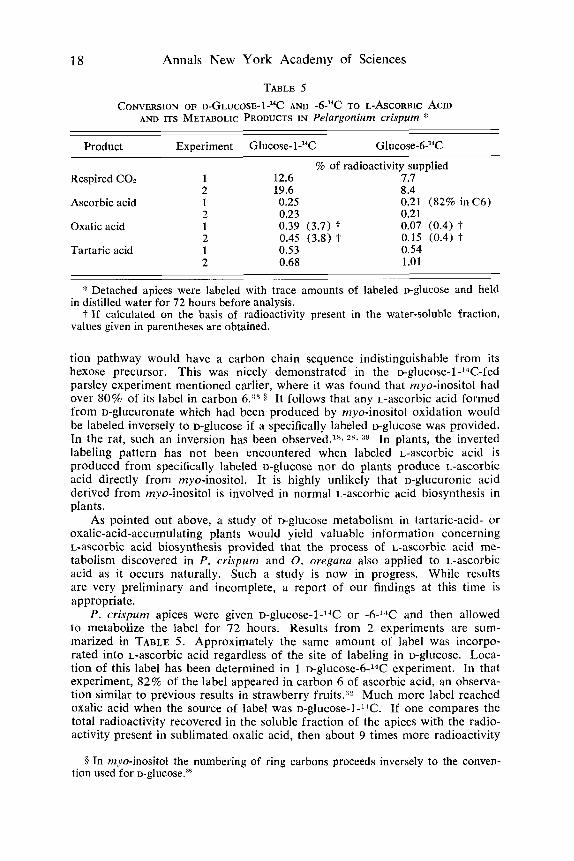
TABLE 5
AND ITS METABOLIC PRODUCTS IN Pelargonium crispum * CONVERSION OF D-GLUCOSE- 1-l4C AND -6-"C TO L-ASCORBIC ACID
Product Experiment Glucose- 1J4C GIuco~e-6-~~C
% of radioactivity supplied Respired CO, 1 12.6 7.7
2 19.6 8.4 Ascorbic acid 1 0.25 0.21 (82% inC6)
2 0.23 0.21 Oxalic acid 1 0.39 (3.7) ? 0.07 (0.4) I'
2 0.45 (3.8) t 0.15 (0.4) -1- Tartaric acid 1 0.53 0.54
2 0.68 1.01 -
'$ Detached apices were labeled with trace amounts of labeled u-glucose and held
?. If calculated on the basis of radioactivity present in the water-soluble fraction, in distilled water for 72 hours before analysis.
values given in parentheses are obtained.
tion pathway would have a carbon chain sequence indistinguishable from its hexose precursor. This was nicely demonstrated in the D-glucose-1 -'"C-fed parsley experiment mentioned earlier, where it was found that myo-inositol had over 80% of its label in carbon 6. 5 It follows that any L-ascorbic acid formed from D-glucuronate which had been produced by myo-inositol oxidation would be labeled inversely to D-gluCOSe if a specifically labeled D-glucose was provided. In the rat, such an inversion has been observed.lx* Z R , 39 In plants, the inverted labeling pattern has not been encountered when labeled L-ascorbic acid is produced from specifically labeled n-glucose nor do plants produce L-ascorbic acid directly from myo-inositol. It is highly unlikely that D-glucuronic acid derived from myo-inositol is involved in normal L-ascorbic acid biosynthesis in plants.
As pointed out above, a study of D-glucose metabolism in tartaric-acid- or oxalic-acid-accumulating plants would yield valuable information concerning L-ascorbic acid biosynthesis provided that the process of L-ascorbic acid me- tabolism discovered in P. crispum and 0. oregana also applied to L-ascorbic acid as it occurs naturally. Such a study is now in progress. While results are very preliminary and incomplete, a report of our findings at this time is appropriate.
P . crispurn apices were given D-glUCOSe-l-14C or -W4C and then allowed to metabolize the label for 72 hours. Results from 2 experiments are sum- marized in TABLE 5. Approximately the same amount of label was incorpo- rated into L-ascorbic acid regardless of the site of labeling in D-glucose. Loca- tion of this label has been determined in l ~ - g h c o s e - 6 - ~ ~ c experiment. In that experiment, 82% of the label appeared in carbon 6 of ascorbic acid, an observa- tion similar to previous results in strawberry fruits.3L Much more label reached oxalic acid when the source of label was ~ -g lucose - l -~~C. If one compares the total radioactivity recovered in the soluble fraction of the apices with the radio- activity present in sublimated oxalic acid, then about 9 times more radioactivity
5 In myo-inositol the numbering of ring carbons proceeds inversely to the conven- tion used for D-glucose.m

Loewus et al.: Biosynthesis and Metabolism 19
TABLE 6 DISTRIBUTION OF CARBON-14 IN TARTARIC ACID FROM Pclurgoniuin crispum
AFTER LABELING WITH D-GLUCOSE-~-"C OR -6-"C "-
Carbons 1 f 4 Carbons 2+3 Source of Label Exper. 1 Exper. 2 Exper. 1 Exper. 2
% of incorporated label 6.8 67 32 33 90 87 10 13
:E See Wagner and Loewus " for experimental details.
was present in D-glucose-1 'C-derived oxalic acid than in ~-glucose-6-l-'C- derived oxalic acid.
The amount of label incorporated into (+)-tartaric acid was similar in 1 set of studies with i>-glucose-l-llC and -6-IlC but showed a significant increase with C6 label over C I label in the other. After careful recrystallization to constant specific radioactivity, all samples were degraded with periodate to determinc the distribution of label between carboxyl and internal carbons. Re- sults are given in TABLE 6. About 3 times more I'C redistributed into internal carbons, C2 + C3, in D-glucose-l-llC experiments than in D-glucose-6-11C ones. Clearly, label located in the C6 position of o-glucose followed a more direct path of conversion. Maroc-Gyr If) has also noted that more label appeared in tartaric acid of Pelargoniurn zonale L. after detached leaves were fed D-glucose- 6-14C than after a feeding of D-glucose-l-lT. She did not examine the internal labeling pattern of tartaric acid.
When 0. oreganci leaves were labeled with D-glucose-l-llC, -2-"C, or -6°C for 48 or 72 hours, respired CO, and thc water-soluble extract accounted for a major portion of the labcl, 60 to 70% in D-glucose-I-*l-C- and -2-*Glabeled leaves and 80 to 90% in D-glucosc-6-1~C-labeled leaves. A significant part of the water-soluble label was oxalic acid. As in the P . crispuriz experiments, D- glucose-1 -"C was a better source of label than ~-gIucose-h-~~C. Strangely, D-ghICOSe-2-14c was no better than ~-glucose-6-' IC. Even when the incorpora- tion into oxalic acid was calculated as percent of soluble label, this observation persisted. Expectation had been that ~-glucose-2-' IC would furnish as much labcl as D-~Iucosc - I -~~C. At present we have n o explanation to offer regarding these results. (TABLE 7) .
TABLE 7
A N D RESPIRED CO, IN GLUCOSE-LABELFD Owulis o r e p i i n LEAVES ~NCORI'ORATI0N OF CARBON- 14 INTO WATER-SOLUBI E CONSTITUENTS, OXALlC ACID,
48 Hours' Metabolism 72 Hours' Metabolism Label Solubles Oxalic acid COX Solubles Oxalic acid COP
% of radioactivity supplied Glucose-1-"C 33 11.8 (36):b 24 29 13.7 (47)': 26 Glucose-2-"C 29 4 (14)" 32 33 7 (21)" 40 Gluco~e-6-~~C 40 6 (15)" 40 45 1 (16)* 45
* Calculated as percent of water-soluble radioactivity.

20 Annals New York Academy of Sciences
Although these glucose-labeling studies have turned up interesting new problems regarding tartaric acid and oxalic acid biosynthesis, they are con- sistent with our earlier findings, which support a scheme of L-ascorbic acid biosynthesis in plants that conserves the carbon chain of hexose and includes oxidation of carbon 1 of hexose to form the carboxyl carbon of ascorbic acid.
The structural relationship of the carbon chain of D-glucose to that of L-ascorbic acid in plants and the metabolic relationship of the latter to (+)- tartaric acid should bring renewed interest regarding the proposed role of 5-keto-~-gIucose or 5-keto-D-gluconic acid in plant systems."' Recently, Kodama et al:*l and Kotera et al.". ~3 examined 5-keto-~-gluconic acid metabolism in a high-( +) -tartaric-acid-excreting mutant of Gluconohacter suboxydans and have constructed a biosynthetic scheme of (+)-tartaric acid production based on their findings (FIGURE 10). In this scheme, pretaric acid, formed by a Baeyer- Villiger-type oxidation, provides the intermediate that breaks down to (+) - tartaric acid and a 2-carbon fragment.
SUMMARY
In plants, the biosynthetic relationship of &glucose, L-ascorbic acid, and products of L-ascorbic acid metabolism is regarded as distinct from the bio- synthetic relationship of D-glucose, D-glucuronic acid, and products of D-glU- curonic acid metabolism. Plants that accumulate (+)-tartaric acid and/or oxalic acid can form these compounds from r--ascorbic acid. Oxalic acid derives its carbon from C1 + C2 of L-ascorbic acid. (+)-Tartaric acid derives its carbon chain from C1 through C4 of 1-ascorbic acid when it occurs in the Vitaceae but from C3 through C6 when from the Geraniaceae. Attempts to find a biosynthetic relationship between D-glucose, L-ascorbic acid, (+)-tartaric acid, and oxalic acid are now in progress.
D- Glucose - 0- Gluconic - 5-Ketagluconic acid
__* 4,s - Dienol of - 4 - Ketoglucanic acid glucanic acid
COOH COOH I I
H-C-OH H-C-OH I
HO-C-H I COOH
acid C HO COOH
0 (+)-Tartaric I
I &OH-+ CHIOH
H-C-OH I CH20H
Pretaric acid Glycooldehyde Glycolic acid
FIGURE 10. Conversion of o-glucose to (+)-tartaric acid via 5-ketogluconic acid as proposed by Kotera et alP'

Loewus et al. : Biosynthesis and Metabolism 21
REFERENCES
1. HERBERT, R. W., E. L. HIRST, G. V. PERCIVAL, R. J. W. RENOLDS & F. SMITH. 1933. The constitution of ascorbic acid. J. Chem. SOC. London. 1270-1290.
2. HOUGH, L. & J. K. N. JONES. 1956. The biosynthesis of the monosaccharides. Advances Carbohydr. Chem. 11: 185-262 (see p. 240).
3. LOEWUS, F. A. & H. A. STAFFORD. 1958. Observations on the incorporation of C" into tartaric acid and the labeling pattern of D-glucose from an excised grape leaf administered ~-ascorbic-acid-6-C'". Plant Physiol. 33: 155-156.
4. SAITO, K. & Z. KASAI. 1969. Tartaric acid synthesis from ~-ascorbic-acid-l-~'C in grape berries. Phytochemistry 8: 2177-2182.
5. FINKLE, B. J., S. KELLY & F. A. LOEWUS. 1960. Metabolism of D-[l-''C]- and D-[6-'4C]glucuronolactone by the ripening strawberry. Biochim. Biophys. Acta
6. LOEWUS, F. A,, S. KELLY & E. F. NEUFELD. 1962. Metabolism of myo-hositol in plants: conversion to pectin, hemicellulose, o-xylose and sugar acids. Proc. Nat. Acad. Sci. U S A . 48: 421-425.
7. WAGNER, G. & F. A. LOEWUS. 1974. L-ascorbic acid metabolism in the Vita- ceae. Conversion to (+)-tartaric acid and hexoses. Plant Physiol. 54: 784- 787.
8 . WAGNER, G. & F. LOEWUS. 1973. The biosynthesis of (+)-tartaric acid in Pelargonium crispum. Plant Physiol. 52: 651-654.
9. BAIG, M. M., S. KELLY & F. LBEWUS. 1970. L-ascorbic acid biosynthesis in higher plants from ~-gulono-1,4-lactone and ~-galactono-1,4-lactone. Plant Physiol. 46: 277-280.
10. JACKSON, G. A. D., R. B. WOOD & M. V. PROSSER. 1961. Conversion of L- galactono-y-lactone into L-ascorbic acid by plants. Nature 191: 282-283.
1 1. WAGNER, G. 1974. Carbohydrate interconvenions involving the biosynthesis and metabolism of L-ascorbic acid and tartaric acid in plants. Ph.D. Thesis. State Univ. of New York at Buffalo.
12. MILLERD, A., R. K. MORTON & J. R. E. WELLS. 1963. Oxalic acid biosynthesis in shoots of Oxalis pes-cuprac L. Biochem. J. 8 6 57-62.
13. CHANG, C-C. & H. BEEVERS. 1968. Biogenesis of oxalate in plant tissues. Plant Physiol. 43: 1821-1828.
14. BURNS, J. J., H. B. BURCH & C. G. KING, 1951. The metabolism of l-C''-L- ascorbic acid in guinea pigs. J. Biol. Chem. 191: 501-514.
15. BURNS, J. J., P. G. DAYTON & S. SCHULENBURG. 1956. Further observations on the metabolism of L-ascorbic acid in guinea pigs. J. Biol. Chem. 218 15-21.
16. HELLMAN, L. & J. J. BURNS. 1958. Metabolism of L-ascorbic acid-l-I4C in man. J. Biol. Chem. 230: 923-930.
17. DAYTON, P. G., F. EISENBERG, JR. & J. J. BURNS. 1959. Metabolism of C"- labeled ascorbic, dehydroascorbic and diketogulonic acids in guinea pigs. Arch. Biochem. Biophys. 81: 1 11-1 18.
18. KING, C. G. & R. R. BECKER. 1959. The biosynthesis of vitamin C (ascorbic acid). Wld. Rev. Nutr. Diet. 1: 61-72.
19. OE, 0. L. 1969. Oxalic acid in plants and in nutrition. Wld. Rev. Nutr. Diet.
20. HAGLER, L. & R. H. HERMAN. 1973. Oxalate metabolism. I. Amer. J . Clin.
21. FASSETT, D. W. 1973. Oxalates. In Toxicants Occurring Naturally in Foods.
22. RANSON, S. L. 1965. Plant acids. I n Biosynthetic Pathways in Higher Plants.
23. Bum, M. L. 1960. A Bibliography of Organic Acids in Higher Plants. Agric.
38: 332-339.
1 0 262-303.
Nutr. 26: 758-765.
2nd edit. : 346-362. Nat. Acad. Sci., Washington, D.C.
J. B. Pridham, Ed. : 179-198. Academic Press. New York, N.Y.
Handbook No. 164, U.S. Dept. Agric. Washington, D.C.

22 Annals N e w York Academy of Sciences
24. HuTTERnAucH, R. & 1. KEINER. 1968. Separation of ascorbic acid and dehydro-
25. DAYTON, P. G., M. McM. SNELL & J. M. PEREL. 1966. Ascorbic and dehydro-
26. LOEWUS, F. I97 1 . Carbohydrate interconversions. Annu. Rev. Plant Physiol.
27. MAPSON, L. W. 1967. Biogenesis of L-ascorbic acid in plants and animals. In The Vitamins. W. H. Sebrell, Jr. & R. S. Harris, Eds. 1: 369-385. Academic Press. New York, N.Y.
28. KING, C. G. 1973. The biological synthesis of ascorbic acid. Wld. Rev. Nutr. Diet. 18: 47-59.
29. HASSID, W. Z . 1970. Biosynthesis of sugars and polepaccharides. In The Carbo- hydrates. W. Pigman & D. Horton, Eds. 2nd edit., 2A: 301-373.
30. Loswus, F., M-S. CHEN & M. W. LOEWUS. 1973. The myo-inositol oxidation pathway to cell wall polysaccharides. Iri Biogenesis of Plant Cell Wall Poly- saccharides. F. Loewus, Ed. : 1-27. Academic Press. New York, N.Y.
31 . LOEWUS, F. A. 1961. Aspects of ascorbic acid biosynthesis in plants. Ann. N.Y. Acad. Sci. 92: 57-78.
32. LOEWUS, F. A. 1963. Tracer studies on ascorbic acid formation in plants. Phy- tochemistry 2: 109-128.
33 . LOEWUS, F. A. 1965. lnositol metabolism and cell wall formation in plants. Fed. Proc., FASEB. 24: 855-862.
34. HANNINEN, O., R. RAUNIO & J . MARNIEMI. 1971. Biosynthesis of L-ascorbic acid from myo-inositol. Carbohydr. Res. 16: 343-351.
35. LOEWUS, F. 1974. The biochemistry of myo-inositol in plants. I n Metabolism and Regulation of Secondary Plant Products. V. C . Runeckles and E. E. Conn, Eds. Recent Advan. Phytochemistry 8: 179-207. Academic Press, New York, N.Y.
36. Loewus, F. A. & S. KELLY. 1962. Conversion of glucose to inositol in parsley leaves. Biochem. Biophys. Res. Commun. 7: 204-208.
37. LOEWUS, F. A. & S. KELLY. 1963. Inositol metabolism in plants. I. Labeling patterns in cell wall polysaccharides from detached plants given myo-inositol- 2-t or -2-"C. Arch. Biochem. Biophys. 102: 96-105.
IUPAC-IUB COMMISSION ON BIOCHEMICAL NOMENCLATURE. 1968. J. Biol. Chem. 243: 5809-58 19.
39. LOEWUS, F. A., S. KELLY & H. H. HIATT. 1960. Ascorbic acid synthesis from D-glUCOSe-2-'"C in the liver of the intact rat. J . Biol. Chem. 235: 937-939.
40. MAROC-GYR, J . 1965. The metabolism of glucose and gluconate in the leaves of Pelargonirrm zonale L. in relation to the formation of organic acids and in particular tartaric acid. Physiol. Veg. 3: 167-180.
41. KODAMA, T., U. KOTERA & K. YAMADA. 1972. Induction of mutants from the tartaric producing bacterium, Gluconobacter sriborydans and their properties. Agr. Biol. Chem. 36: 1299-1305.
42. KOTERA, U., K. UMEHARA, T. KODAMA & K. YAMADA. 1972. Isolation method of highly tartaric acid producing mutants of Glrrconobacter suboxydans. Agr. Biol. Chem. 36: 1307-1313.
43. KOTERA, U., T. KODAMA, Y. MINODA & K. YAMADA. 1972. Isolation and chemi- cal structure of new oxidation product of 5-ketogluconic acid, and a hypotheti- cal pathway from glucose to tartaric acid through this compound. Agr. Biol. Chem. 3 6 1315-1325.
44. BAKER, E. M., J . C. SAARI & B. M. TOLBERT. 1966. Ascorbic acid metabolism in man. Amer. J . Clin. Nutr. 19: 371-378.
ascorbic acid on polyamide plates. Pharmazie 23: 157.
ascorbic acids in guinea pigs and rats. J. Nutr. 88: 338-344.
22: 337-364.
38. IUPAC COMMISSION ON THE NOMENCLATURE OF ORGANIC CHEMISTRY AND

Loewus et al. : Biosynthesis and Metabolism 23
DISCUSSION
DR. J. J . BURNS: In the data that you showed for the plants that form oxalate, would you care to estimate how much oxalate may come via ascorbate in these plants or can it come from other sources?
DR. LOEWUS: In the oxalate biosynthesis in plants, two other pathways have been proposed. One in which oxaloacetic acid is broken to form acetic acid and oxalic acid has been shown in vivo to occur in certain plants such as beet and spinach. The other pathway in which glycolate or glyoxalate is oxidized has been studied by Millard and others in Australia. Both of these proposals offer alternatives for oxalate biosynthesis, but neither has shown the dramatic conversion that we find now with ascorbic acid. Since ascorbic acid is being synthesized naturally at a substantial rate in plants, I think the best candidate for study at this particular time might be biosynthesis of ascorbic acid and its ultimate metabolism. DR. JULIUS BERGER (Hoffmann-La Roche, Nutley, N.J.): Has there been any
confirmed demonstration of the production of ascorbic acid by any micro- organisms?
DR. LOEWUS: There are no definitive reports of ascorbic acid biosynthesis by bacteria. However, we find that the algae are very good at producing ascorbic acid. We have been examining one particular diatom that converts glucose to ascorbic acid but fragments it into such a redistribution that we are at a loss to explain the biosynthetic pathway at this particular time.
DR. KING: Have you found any organisms too far down the biological scale to have ascorbic acid present in them? Is there any indication at all of living organisms that do not require ascorbic acid either by synthesis or by intake?
DR. LOEWUS: Ascorbic acid seems to be ubiquitous in the plant world, hut it is not well established whether this extends to bacteria. Some species of fungi apparently have ascorbic acid. Modern methods of analysis such as gas chromatography should answer these questions. Up to now, for the most part, ascorbic acid has been identified by its oxidation-reduction properties, which is not a clear criterion.


















