
Biomass production and nitrate metabolism of
12
Oecologia (Berlin) (1987) 72:303-314 Oecologia Springer-Verlag 1987 Biomass production and nitrate metabolism of Atriplex hortensis L. (C3 plant) and Amaranthus retrotlexus L. (C4 plant) in cultures at different levels of nitrogen supply* G. Gebauer 1, M.I. Schuhmacher 2, B. Krsti~ 3, H. Rehder 1, and H. Ziegler 1 1 Institut ffir Botanik und Mikrobiologie der Technischen Universitfit Mtinchen, Arcisstrasse 21, D-8000 Miinchen 2, Federal Republic of Germany 2 Centro de Ecologia, Universidad National de Cordoba, Cordoba, Argentina 3 Institute of Biology, University of Novi Sad, 21000 Novi Sad, Yugoslavia Summary. Pure and mixed cultures of the dicotyledons Atriplex hortensis L. (C3 plant) and Amaranthus retroflex- us L. (C4 plant) were maintained under open air conditions in standard soil at low and high nitrogen supply levels. A comparison of shoot dry weight and shoot length in the various series shows that the growth of the above- ground parts of both species was severely reduced under low N conditions. In both pure and mixed cultures the differences resulting from low N vs. high N conditions was less pronounced with Atriplex (C3 plant) than with Amaran- thus (C4 plant). The root dry weight of the two species was not reduced so much under low N conditions as was the shoot dry weight. The low N plants were found to contain a larger proportion of their biomass in the roots than did the high N plants. In general the root proportion of Atriplex was greater than that of Amaranthus. The con- tents of organic nitrogen and nitrate and the nitrate reduc- tase activity (NRA) per g dry weight of both species de- creased continually throughout the experiments. With the exception of young plants, the low N plants always had lower contents of organic nitrogen and nitrate and nitrate reductase activities than did the high N plants. The highest values of NRA were measured in the leaf laminae. The leaves also exhibited the highest concentrations of organic nitrogen. The highest nitrate concentrations, however, were observed in the shoot axis, and in most cases the lowest nitrate values were found in the laminae. At the end of the growing season this pattern was found to have been reversed with Atriplex, but not with Amaranthus. Thus Atriplex was able to maintain a higher NRA in the laminae than Amaranthus under low N conditions. The transpiration per leaf area of the C4 plant Amaran- thus during the course of a day was substantially lower than that of the C3 plant Atriplex. There were no significant differences in transpiration between the low N and high N series of Amaranthus. The low N plants of Atriplex, how- ever, clearly showed in most cases higher transpiration rates than the corresponding high N plants. These different tran- spiration rates of the high N and the low N Atriplex plants were also reflected in a distinct 13C discrimination. The sum of these results points to the conclusion that the C3 plant Atriplex hortensis can maintain a better inter- nal inorganic nitrogen supply than the C4 plant Amaranthus * Dedicated to Prof. Dr. Karl M/igdefrau, Deisenhofen, on the occasion of his 80th birthday Offprint requests to: G. Gebauer retroflexus under low N conditions and an ample water supply, due to the larger root proportion and the more pronounced and flexible transpiration of the C3 plant. Key words: Nitrogen use efficiency- C3 plant- C4 plant - Biomass production - Nitrate metabolism The hypothesis of Brown (1978) that C4 plants can utilize nitrogen more efficiently than C3 plants and are therefore better adapted to low N conditions (cf. reviews by Osmond et al. 1982; Pearcy and Ehleringer 1984) has been generaUy accepted for some time. This hypothesis was based on the fact that C4 plants require less of the enzyme ribulose-l,5- bisphosphate carboxylase (RuBP carboxylase) to maintain photosynthetic rates comparable to those evidenced by C3 plants (Bj6rkman et al. 1976; Ku et al. 1979). Recent findings, however, necessitate a reassessment of the problem of the adaptation of C3 and C4 plants to the essen- tial and in many cases growth limiting factor nitrogen nutri- tion. Doliner and Jolliffe (1979) employed various statistical techniques to reveal that differences in the distribution of C3 and C4 species could be related to the factors tempera- ture and moisture availability, but not significantly to the factor soil nitrogen. In experiments incorporating various C3 and C4 plants in water culture at high and low nitrogen nutrition levels, Krsti6 and Sari6 (1983) observed species- specific differences in biomass production, in the shoot/root ratio, and in nitrogen content which were, however, not related to the pathway of photosynthesis (C3 or C4). Ge- bauer et al. (1987) even observed a superiority of C3 plants under low N conditions in competition experiments em- ploying two C3 and two C4 grasses in pure and in mixed cultures at high and low N supply levels. Christie and De- tling (1982) and Edwards et al. (1985) also describe superior growth of C3 plants under low N conditions, but only at low temperatures (20 ~ C/12 ~ C and 18 ~ C/18 ~ C day/night regimes, respectively). Under high temperature conditions (30 ~ C/15 ~ C and 32 ~ C/18 ~ C day/night regimes, respective- ly), however, these authors observed better growth of C4 plants irrespective of nitrogen nutrition. Gebauer et al. (1987) discuss the possibility that the reduced competitive ability of C4 plants under low N conditions is related to the different transpiration rates of C3 and C4 plants (cf. Osmond et al. 1982). Possibly the low transpiration rates and the high water use efficiency of the C4 plants result
Transcript of Biomass production and nitrate metabolism of

Oecologia (Berlin) (1987) 72:303-314 Oecologia �9 Springer-Verlag 1987
Biomass production and nitrate metabolism of Atriplex hortensis L. (C3 plant) and Amaranthus retrotlexus L. (C4 plant) in cultures at different levels of nitrogen supply* G. Gebauer 1, M.I . Schuhmacher 2, B. Krsti~ 3, H. Rehder 1, and H. Ziegler 1 1 Institut ffir Botanik und Mikrobiologie der Technischen Universitfit Mtinchen, Arcisstrasse 21, D-8000 Miinchen 2, Federal Republic of Germany 2 Centro de Ecologia, Universidad National de Cordoba, Cordoba, Argentina 3 Institute of Biology, University of Novi Sad, 21000 Novi Sad, Yugoslavia
Summary . Pure and mixed cultures of the dicotyledons Atriplex hortensis L. (C3 plant) and Amaranthus retroflex- us L. (C4 plant) were maintained under open air conditions in standard soil at low and high nitrogen supply levels.
A comparison of shoot dry weight and shoot length in the various series shows that the growth of the above- ground parts of both species was severely reduced under low N conditions. In both pure and mixed cultures the differences resulting from low N vs. high N conditions was less pronounced with Atriplex (C3 plant) than with Amaran- thus (C4 plant). The root dry weight of the two species was not reduced so much under low N conditions as was the shoot dry weight. The low N plants were found to contain a larger proportion of their biomass in the roots than did the high N plants. In general the root proportion of Atriplex was greater than that of Amaranthus. The con- tents of organic nitrogen and nitrate and the nitrate reduc- tase activity (NRA) per g dry weight of both species de- creased continually throughout the experiments. With the exception of young plants, the low N plants always had lower contents of organic nitrogen and nitrate and nitrate reductase activities than did the high N plants. The highest values of NRA were measured in the leaf laminae. The leaves also exhibited the highest concentrations of organic nitrogen. The highest nitrate concentrations, however, were observed in the shoot axis, and in most cases the lowest nitrate values were found in the laminae. At the end of the growing season this pattern was found to have been reversed with Atriplex, but not with Amaranthus. Thus Atriplex was able to maintain a higher NRA in the laminae than Amaranthus under low N conditions.
The transpiration per leaf area of the C4 plant Amaran- thus during the course of a day was substantially lower than that of the C3 plant Atriplex. There were no significant differences in transpiration between the low N and high N series of Amaranthus. The low N plants of Atriplex, how- ever, clearly showed in most cases higher transpiration rates than the corresponding high N plants. These different tran- spiration rates of the high N and the low N Atriplex plants were also reflected in a distinct 13C discrimination.
The sum of these results points to the conclusion that the C3 plant Atriplex hortensis can maintain a better inter- nal inorganic nitrogen supply than the C4 plant Amaranthus
* Dedicated to Prof. Dr. Karl M/igdefrau, Deisenhofen, on the occasion of his 80th birthday
Offprint requests to: G. Gebauer
retroflexus under low N conditions and an ample water supply, due to the larger root proportion and the more pronounced and flexible transpiration of the C3 plant.
Key words: Nitrogen use efficiency- C3 p l a n t - C4 plant - Biomass production - Nitrate metabolism
The hypothesis of Brown (1978) that C4 plants can utilize nitrogen more efficiently than C3 plants and are therefore better adapted to low N conditions (cf. reviews by Osmond et al. 1982; Pearcy and Ehleringer 1984) has been generaUy accepted for some time. This hypothesis was based on the fact that C4 plants require less of the enzyme ribulose-l,5- bisphosphate carboxylase (RuBP carboxylase) to maintain photosynthetic rates comparable to those evidenced by C3 plants (Bj6rkman et al. 1976; Ku et al. 1979). Recent findings, however, necessitate a reassessment of the problem of the adaptation of C3 and C4 plants to the essen- tial and in many cases growth limiting factor nitrogen nutri- tion. Doliner and Jolliffe (1979) employed various statistical techniques to reveal that differences in the distribution of C3 and C4 species could be related to the factors tempera- ture and moisture availability, but not significantly to the factor soil nitrogen. In experiments incorporating various C3 and C4 plants in water culture at high and low nitrogen nutrition levels, Krsti6 and Sari6 (1983) observed species- specific differences in biomass production, in the shoot/root ratio, and in nitrogen content which were, however, not related to the pathway of photosynthesis (C3 or C4). Ge- bauer et al. (1987) even observed a superiority of C3 plants under low N conditions in competition experiments em- ploying two C3 and two C4 grasses in pure and in mixed cultures at high and low N supply levels. Christie and De- tling (1982) and Edwards et al. (1985) also describe superior growth of C3 plants under low N conditions, but only at low temperatures (20 ~ C/12 ~ C and 18 ~ C/18 ~ C day/night regimes, respectively). Under high temperature conditions (30 ~ C/15 ~ C and 32 ~ C/18 ~ C day/night regimes, respective- ly), however, these authors observed better growth of C4 plants irrespective of nitrogen nutrition. Gebauer et al. (1987) discuss the possibility that the reduced competitive ability of C4 plants under low N conditions is related to the different transpiration rates of C3 and C4 plants (cf. Osmond et al. 1982). Possibly the low transpiration rates and the high water use efficiency of the C4 plants result

304
in a sub-optimal supply with mineral nitrogen from the soil, in particular under conditions of low N and low tem- perature. A strong dependency of the uptake and transport of nitrate from the intensity of transpiration is reported for the C4 plant maize (Shaner and Boyer 1976).
The aim of the present work was to verify the competi- tive advantage of C3 plants under low N conditions in com- petition experiments incorporating dicotyledonous C3 and C4 plants. Such experiments have hitherto been restricted to monocotyledons. Moreover, mutual communities and differences were investigated with regard to some ecologi- cally significant aspects of the nitrogen nutrition of C3 and C4 plants during a growing season in relation to the transpi- ration rate.
Materials and methods
Plant material
The experiments were carried out with Atriplex hortensis L. (C3 plant) and Amaranthus retroflexus L. (C4 plant). These two species are annual weeds found in nitrogen-rich habitats and are of similar size and exhibit - according to Ellenberg (1979) - comparable ecological behaviour. Amaranthus has a C~-typical high temperature figure (cf. Ellenberg 1979). The seeds were germinated in the green- house and transplanted into Eternit (=asbestos cement)- boxes (150 x 80 x 80 cm) in the open air when 4-6 cm tall. Only equally tall seedlings were employed. Natural climatic conditions were influenced only during dry periods by addi- tional watering of the plants. The course of the mean tem- perature during the growing season is indicated in Fig. 1.
Substrate
The plants were cultured in soil containing 70% "Fruh- storfer Erde T " (Industrieerdenwerk Erich Archut, Lauter- bach/Fulda, FRG) and 30% sand. "Fruhstorfer Erde T " is a peaty soil with a 40% clay content. At the beginning of the experiments the substrate in each of the 28 boxes was analyzed respective of the following parameters: - pH value (in aqueous solution): 7.52 (standard deviation
s: 0.08); - total nitrogen content: 0.13% of the dry weight (s: 0.02); - total carbon content: 3.18 % of the dry weight (s: 0.56); - p l a n t available inorganic nitrogen ( N O ~ + N H 2 ) :
10.7 gg N/g dry wt (s: 5.0).
Experimental series
Two different groups of experiments were carried out. In experiment A the growth and the transpiration rate of the plants were investigated, as well as some aspects of the nitrogen metabolism of the two species during the growing season (mid May - mid September). Experiment A com- prised the following series: (1) High N set: Each box was planted with 72 plants of either Atriplex or Amaranthus at regular spacings. At the beginning of the experiments (when the seedlings were transplanted to the open air boxes) and after 37 days (Amaranthus) or 45 days (Atriplex) the plants of each box were fertilized with 50 g o f " Calnitro Kalkammonsalpeter" (Hoechst). This fertilizer contains 26% nitrogen (13% in the form of NO~- and 13% in the form of N H +) and besides the nitrogen compounds only 22% of CaCO3. Due to im-
Temperature [ 'c]
2 5
2 0
15
10
r
r J
-J
i
I I I I I I �9 20 40 60 80 100 120
T [doys]
Fig. l. Mean temperature values of ten days periods during the growing season (Munich, mid May mid September, 1984)
mediate nitrification, however, ammonium plays only a mi- nor role as a nitrogen source in our substrate (cf. Melzer et al. 1984). A third fertilization was carried out with the Amaranthus set after 107 days. (2) Low N set: 72 plants of either Atriplex or Amaranthus per box were fertilized once with only 10 g of "Calnitro Kalkammonsalpeter" at the beginning of the experiments.
At intervals of from two to four weeks three plants per box were selected at random and separated into their different organs (laminae, midribs and petioles, segments of the shoot axis, roots, reproductive organs). During the growing season Atriplex developed only weakly pro- nounced branches of the shoot axis. These axes were thus not investigated as separate organs (as they were in the case of Amaranthus), but were rather combined with the fraction of petioles and midribs. The plants were always harvested between 1145 and 1230 h (Central European Sum- mer Time). For each of these samples dry weight, total nitrogen content (with the exception of the initial harvests), nitrate content and nitrate reductase activity were deter- mined. The leaf material of one harvest was also used to analyze 613C values. Soil samples were taken from the root area of each plant harvested and analyzed for their momen- tary nitrate content. In the middle of the growing season the daily transpiration courses of the high N and low N plants of each species were investigated repeatedly. Transpi- ration and photosynthetically active radiation (PAR) were each measured in five repetitions from 6 oo to 22~176 at intervals of 2 h, Only the youngest fully developed and un- shaded leaves were selected for these measurements. The transpiration rates of Atriplex leaves of different ages were also compared. At the time of this comparison the Atriplex plants had eight (high N set), or seven (low N set) distin- guishable leaf storeys. The transpiration of one leaf of each of these storeys was observed in ten repetitions between 1030 and 1530 h. The data obtained from the various series were treated statistically using Student's t-test.
In experiment B the competitive behaviour of the high N and low N plants of each species was investigated in detail in pure cultures and in mixed cultures. For this pur- pose the following series were chosen:
(I) High N set: For pure cultures 60 seedlings of either Atriplex or Amaranthus were planted per box at regular spacings. The pure cultures were run in three replicates. For mixed cultures 30 seedlings of the C3 species were planted per box alternating with 30 seedlings of the C4 species in a regular manner. These cultures were run in

six replicates. Each box of this set was fertilized with 25 g of "Calnitro Kalkammonsalpeter" at the beginning of the experiment and after 28 days.
(2) Low N set: Pure and mixed cultures were planted in the same manner as in the high N set, but each box was fertilized twice with only 5 g o f " Calnitro Kalkammon- salpeter ".
After ten weeks of cultivation (July-September) ten re- presentative plants (pure cultures) or ten plants per species (mixed cultures) were harvested from each box. Dry weight and length of the above-ground and below-ground parts of each plant were measured. For the determination of total nitrogen content the ten segments of the shoot and of the root material from each species and box were ground and homogenized.
Methods
Plant-available inorganic nitrogen of the soil was analyzed according to the method of Bremner and Keeney (1965) using a distillation apparatus described by Gerlach (1973). For dry weight determinations the plant material was dried at 105 ~ C until weight constancy. The total nitrogen content of plant samples and the nitrogen and carbon contents of soil samples were measured by analysis of the homogenized dry material using an automatic nitrogen analyzer (Mod. 1400) from CARLO ERBA. The content of organic nitrogen in the plant samples of experiment A was calcu- lated by subtracting the nitrate nitrogen values from the total nitrogen contents. Nitrate in the plant and soil samples was determined by HPLC using a modification of the meth- od of Thayer and Huffaker (1980) as described by Gebauer
305
et al. (1984). Soil sample extracts were prepared according to the method of Bremner and Keeney (1965). Nitrate re- ductase activity (NRA) was measured according to a modi- fied version of the in vivo-test described by Hageman and Hucklesby (1971) or Jaworski (1971). For details of the test and the optimization procedures of the incubation buffer see Gebauer et al. (1984). An incubation buffer of the following composition was found to be optimal for the determination of NRA in Atriplex hortensis as well as in Amaranthus retroflexus: 0.08 M KNOa, 0.25 M KH2PO4, 1.5% (v/v) propanol, pH 7.5. Transpiration and radiation were measured with the Steady State Porometer LI-1600 from LI-COR. The 13C/12C isotope ratios of the leaf sam- ples were analyzed according to Osmond et al. (1975).
Results
Dry weight and length of the above-ground and below-ground parts
The growth of the two plant species examined differed as indicated in Fig. 2. Dry weight of Atriplex in the high N set increased nearly linearly until the measurement carried out after 86 days. The growth rate of Amaranthus attained a comparable value about 20 days later than in the case of Atriplex, but from day 51 to day 83 an erratic increase of the Amaranthus biomass was observed, followed by stag- nation. The initial delay of growth observed with Amaran- thus appears to be correlated with the relatively low temper- atures during this period (see Fig. 1) and the high tempera- ture demand of the C4 plant Amaranthus. At the end of the growth period Amaranthus (high N set) had produced
dry weight [g]
50 A
LO
30
20
10
2'0 ~'o i' 6o
! 8'0 100 T [days]
dr weight [g]
50
B 4O
30
20
lo
2~ 60 8o 16o T
T [days] 120
dry weight [g]
ZO[ C
Io i
2; 4'o 60 do 16o T [days]
dr weight [g]
D 2c
lO
z'o - - i m
t.O 6~0 810 100 120
T [days]
Fig. 2A-D. Dry weight development of the total plants and their different organs. A Atriplex high N set; B Amaranthus high N set; C Atriplex low N set; D Amaranthus low N set. ~ reproductive organs, [ ~ laminae, ~ midribs + petioles ( + branches of the shoot axis in the case of Atriplex), ~ branches of the shoot axis (in the case of Amaranthus), ~ shoot axis, [ ~ roots, T fertilization of the high N sets
306
more biomass than Atriplex (high N set). Ripening of the seeds was found to have been completed about 20 days earlier in the case of Atriplex. Biomass production of both species was reduced severely in the low N sets, when com- pared with the high N versions. The low N plants of Atrip- lex and Amaranthus produced about the same biomass. During the plant development the proportions of the differ- ent organs contributing to the total biomass shifted drasti- cally, but this took place in all series in the same manner. The absolute dry weight of the laminae increased contin- ually until day 58 (Atriplex) or day 83 (Amaranthus). During the same period, however, the dry weight proportion of the laminae decreased from 60-68% to 25-30%. This shift took place especially in favour of the shoot axis portion. During the following phase of senescence even absolute de- creases in leaf dry weight were observed. In this regard the proportion of the laminae was reduced to only 5-10% of the total dry weight at the end of the experiments. The dry weight loss due to leaf senescence was almost compen- sated for by a vast increase in the dry weight of the repro- ductive organs.
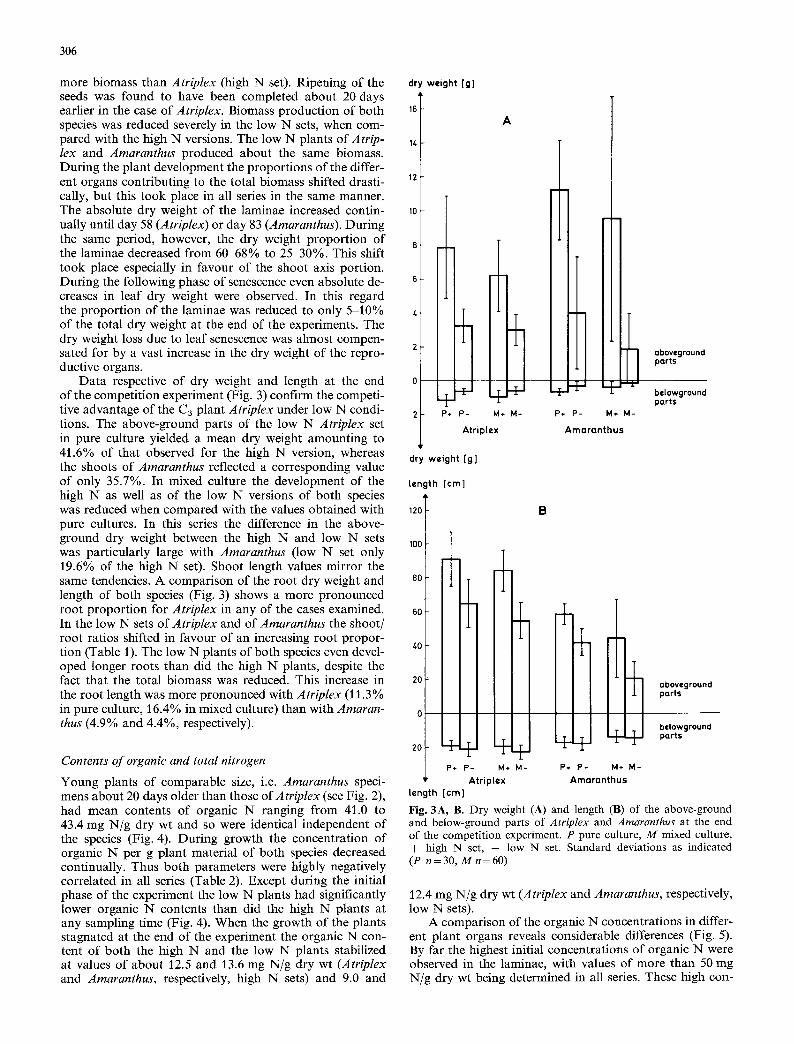
Data respective of dry weight and length at the end of the competition experiment (Fig. 3) confirm the competi- tive advantage of the C3 plant Atriplex under low N condi- tions. The above-ground parts of the low N Atriplex set in pure culture yielded a mean dry weight amounting to 41.6% of that observed for the high N version, whereas the shoots of Amaranthus reflected a corresponding value of only 35.7%. In mixed culture the development of the high N as well as of the low N versions of both species was reduced when compared with the values obtained with pure cultures. In this series the difference in the above- ground dry weight between the high N and low N sets was particularly large with Amaranthus (low N set only 19.6% of the high N set). Shoot length values mirror the same tendencies. A comparison of the root dry weight and length of both species (Fig. 3) shows a more pronounced root proportion for Atriplex in any of the cases examined. In the low N sets of Atriplex and of Amaranthus the shoot/ root ratios shifted in favour of an increasing root propor- tion (Table J). The low N plants of both species even devel- oped longer roots than did the high N plants, despite the fact that the total biomass was reduced. This increase in the root length was more pronounced with Atriplex (11.3% in pure culture, 16.4% in mixed culture) than with Amaran- thus (4.9% and 4.4%, respectively).
Contents of organic and total nitrogen
Young plants of comparable size, i.e. Amaranthus speci- mens about 20 days older than those of Atriplex (see Fig. 2), had mean contents of organic N ranging from 41.0 to 43.4 mg N/g dry wt and so were identical independent of the species (Fig. 4). During growth the concentration of organic N per g plant material of both species decreased continually. Thus both parameters were highly negatively correlated in all series (Table 2). Except during the initial phase of the experiment the low N plants had significantly lower organic N contents than did the high N plants at any sampling time (Fig. 4). When the growth of the plants stagnated at the end of the experiment the organic N con- tent of both the high N and the low N plants stabilized at values of about 12.5 and 13.6 mg N/g dry wt (Atriplex and Amaranthus, respectively, high N sets) and 9.0 and
dry weight [g]
16 A
1/-
12
10
6
2 P§ P- M§ M-
Atripiex
t aboveground parts
I I i J - - betowground parts
P* P- M~ M-
Amaranthus
dry weight [g]
[ength [crn]
120
100
8O
~0
20
0
20 I + ~J~ P§ P- M§ M-
Atriplex length [cm]
+ I
I br T T parts J. J.
P. P- M* M- Amaranthus
Fig. 3A, B. Dry weight (A) and length (B) of the above-ground and below-ground parts of Atriplex and Amaranthus at the end of the competition experiment. P pure culture, M mixed culture, + high N set, -- low N set. Standard deviations as indicated (P n = 30, M n = 60)
12.4 mg N/g dry wt (Atriplex and Amaranthus, respectively, low N sets).
A comparison of the organic N concentrations in differ- ent plant organs reveals considerable differences (Fig. 5). By far the highest initial concentrations of organic N were observed in the laminae, with values of more than 50 mg N/g dry wt being determined in all series. These high con-

Table 1. Shoot/root ratios of Atriplex hortensis and Amaranthus retroflexus at the end of the competition experiment
307
A triplex Amaranthus
Pure culture Mixed culture Pure culture Mixed culture
High N set Low N set High N set Low N set High N set Low N set High N set Low N set
Shoot dry wt Root dry wt 6.70 5.34 6.59 4.91 16.78 12.34 23.91 12.87
Shoot dry wt Root length 0.42 0.16 0.32 0.13 0.65 0.22 0.69 0.13
organic N (mg/g dry w t l
50 A
~0
30
20
10
210 /~lot 610 810 1;0 T [days)
organic N [rng/g dry wt ]
d
50
/,0
30
20
I �9
120
B
I I i �9
2; f/*0 610 810 100 t 120 T [days ]
Fig. 4A, B. Development of the organic N content in the total plants. A Atriplex; B Amaranthus. �9 high N set, o low N set, * P < 0.05, ** P<0.01, *** P<0.001, ~ fertilization of the high N sets
centrat ions in the laminae began to decrease more slowly in the high N plants - especially with respect to Atriplex - than in the low N plants. The late fertilization of the high N Amaranthus after 107 days resulted in only slightly increasing organic N contents of the laminae. Only at the beginning of the flowering stage were the N contents of the laminae exceeded by those o f the reproductive organs. Already at the first harvest the other organs examined (mid- ribs and petioles, shoot axis segments, roots) had organic
.N contents of only 30.5 to 20.5 mg N/g dry wt. The values of the high N plants for these organs were again higher
Table 2. Linear correlations between dry weight and organic nitro- gen content and dry weight and nitrate content with Atriplex hor- tensis and Amaranthus retroflexus of various ages
Correlated Series r Slope y- df P < parameters inter-
cept
Dry weight/ Organic N content
Atriplex High N set --0.9988 --0.969 45.07 3 0.001 Low N set --0.9335 -2.055 42.28 3 0.05
Amaranthus High N set -0.9588 -0.471 41.93 2 0.05 LowN set --0.9980 -1.812 42.57 2 0.01
Dry weight/ Atriplex Nitrate High N set -0.7440 -13.69 569.9 4 n.s. content Low N set -0.9323 --25.20 376.1 4 0.01
Amaranthus High N set --0.6083 - 4.83 390.3 3 n.s. Low N set --0.9088 --18.2/ 320.5 3 0.05
than those of the corresponding low N plants. The rate of value decrease with increasing age of these organs, how- ever, was quite similar in all series (Fig. 5).
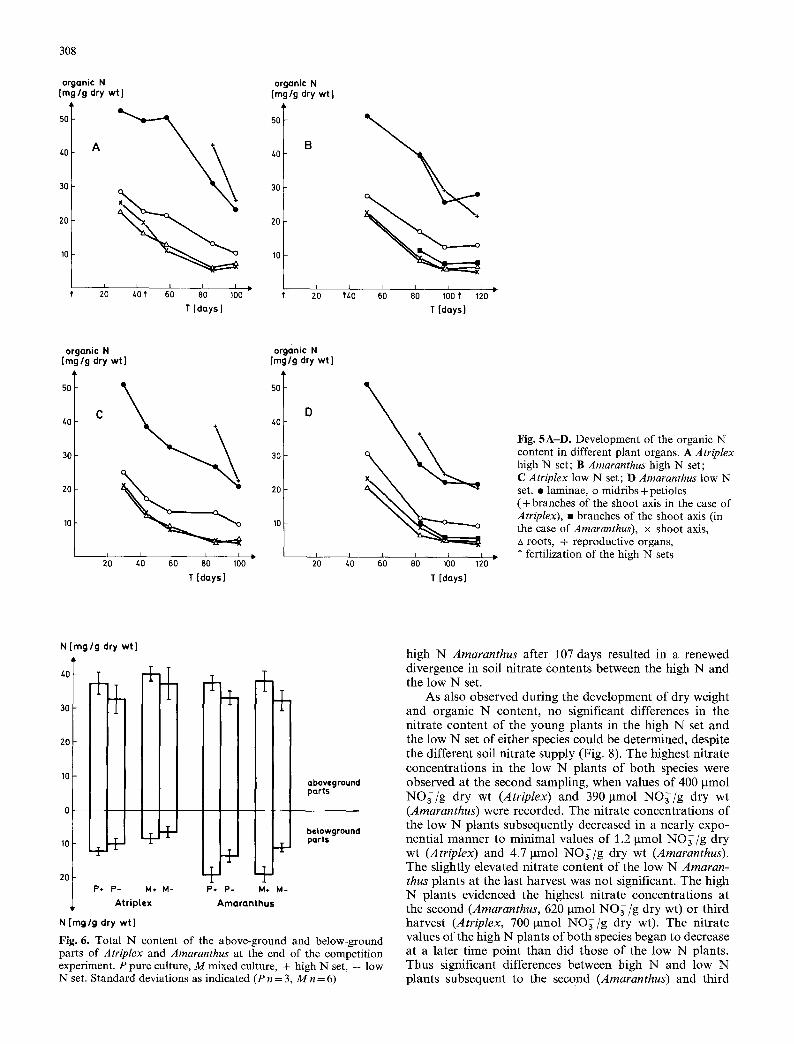
At the end of experiment B the C3 and the C4 plants showed hardly any differences in the total ni trogen contents of their above-ground parts (Fig. 6). In pure culture the shoots of both species had mean total N contents of 37 mg N/g dry wt (high N set) and 33 mg N/g dry wt (low N set). In mixed culture the total N values of the above- ground parts of the high N and the low N Atriplex plants were slightly higher than those of Amaranthus. In contrast , the roots of the two species exhibited clear differences with respect to total N content. Amaranthus always evidenced significantly higher total root N concentrat ions than did Atriplex in the corresponding sets (Fig. 6). The difference between the high N and the low N set was only 10% (mixed culture) and 15% (pure culture) with Atriplex on the one hand and 30% (pure culture) and over 40% (mixed culture) with Amaranthus on the other.
Nitrate content o f soil and plants
Whereas the soil ni trate content o f the high N sets of both species was essentially higher than that of the low N sets at the first harvests, it progressively approximated the ni- trate values in the soil of the low N plants during the course of the growing season (Fig. 7). The late fertilization o f the
I ?40
organic N [mg/g dry wt]
50
40 A
30
2o
10
2; l,'0? 6; 8'0 I;0' T ldays ]
organic N [rng/g dry wt]
50
B 40
30
2O
10
2=0
410
i i | tO 8tO 100 't 120
T [days]
organic N [mg/g dry wt]
L
5O
~o C.
30
20
10
2L0 4L0 6=0 8t0 100 ~
T [days]
organic N [mg/g dry wt]
50
D 4O
30
20
10
I I II 610 8~0 10 0 1 2 0
T [days]
Fig. 5A-D. Development of the organic N content in different plant organs. A Atriplex high N set; B Amaranthus high N set; C Atriplex low N set; D Amaranthus low N set. �9 laminae, o midribs + petioles (+ branches of the shoot axis in the case of Atriplex), �9 branches of the shoot axis (in the case of Amaranthus), x shoot axis, ~x roots, + reproductive organs, T fertilization of the high N sets
N [mg/g dry wt ]
,o + T 30 '
, I
zo I
10
0
I I -I" I I
20 P* P- M* M-
Atriptex
N [mg/g dry wt ]
308
+§ +-In
t +
P* P- M* M- Amaranthus
abovcground parts
betowground parts
Fig. 6. Total N content of the above-ground and below-ground parts of Atriplex and Arnaranthus at the end of the competition experiment. P pure culture, M mixed culture, + high N set, -- low N set. Standard deviations as indicated (P n = 3, M n = 6)
high N Amaranthus after 107 days resulted in a renewed divergence in soil ni trate contents between the high N and the low N set.
As also observed during the development of dry weight and organic N content, no significant differences in the ni trate content of the young plants in the high N set and the low N set of either species could be determined, despite the different soil ni trate supply (Fig. 8). The highest nitrate concentrat ions in the low N plants of both species were observed at the second sampling, when values of 400 pmol N O ~ / g dry wt (Atriplex) and 390 gmol N O ~ / g dry wt (Amaranthus) were recorded. The nitrate concentrat ions of the low N plants subsequently decreased in a nearly expo- nential manner to minimal values of 1.2 pmol NO;- /g dry wt (AtripIex) and 4.7 pmol N O ~ / g dry wt (Amaranthus). The slightly elevated nitrate content of the low N Amaran- thus plants at the last harvest was not significant. The high N plants evidenced the highest ni trate concentrat ions at the second (Amaranthus, 620 pmol NO~-/g dry wt) or third harvest (Atriplex, 700 pmol N O ~ / g dry wt). The nitrate values of the high N plants of both species began to decrease at a later time point than did those of the low N plants. Thus significant differences between high N and low N plants subsequent to the second (Amaranthus) and third
309
N i t r a t e c o n t e n t [pmo( NO:~/g dry w t ]
10
1
0.1
2 ; I I I IL ,o, 6; 8'0 100 120 T [days]
N i t r a t e c o n t e n t [ ,mot NO3"/g dry w t ]
10
1
0.1
;o ' t~o 6'o 8'o ' , D 100 f 120 T [days]
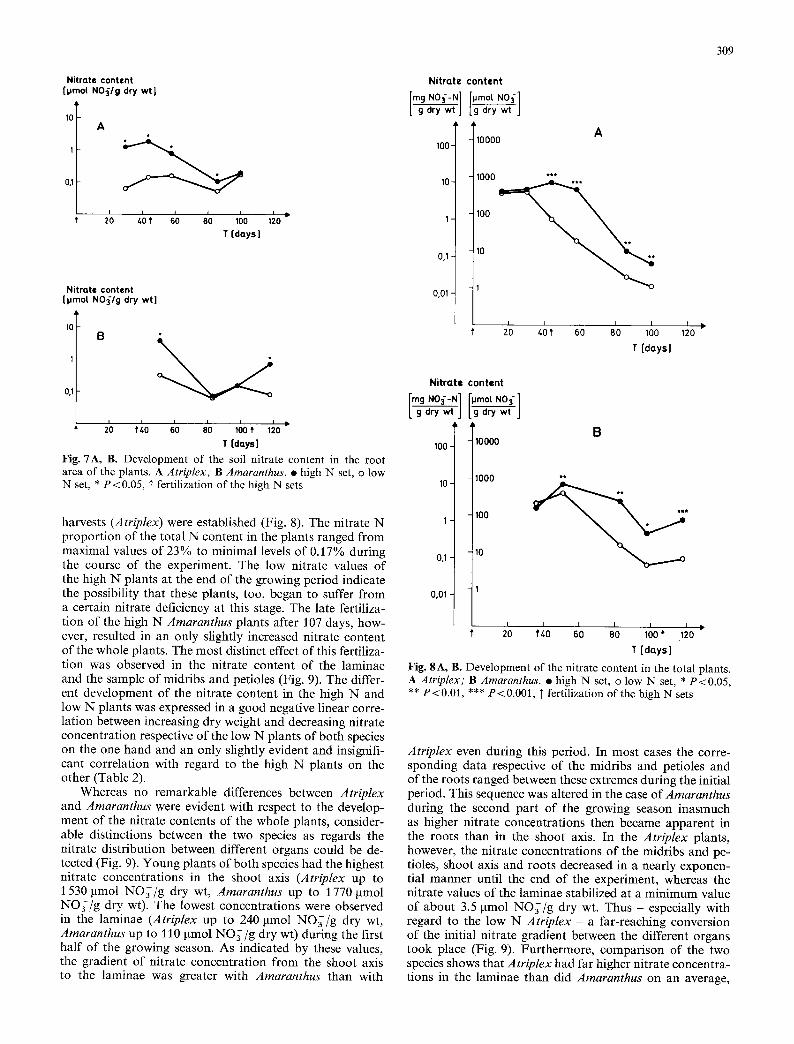
Fig. 7A, B. Development of the soil nitrate content in the root area of the plants. A Atriplex; B Amaranthus. �9 high N set, o low N set, * P<0.05, ]" fertilization of the high N sets
harvests (Atriplex) were established (Fig. 8). The nitrate N proportion of the total N content in the plants ranged from maximal values of 23% to minimal levels of 0.17% during the course of the experiment. The low nitrate values of the high N plants at the end of the growing period indicate the possibility that these plants, too, began to suffer from a certain nitrate deficiency at this stage. The late fertiliza- tion of the high N Amaranthus plants after 107 days, how- ever, resulted in an only slightly increased nitrate content of the whole plants. The most distinct effect of this fertiliza- tion was observed in the nitrate content of the laminae and the sample of midribs and petioles (Fig. 9). The differ- ent development of the nitrate content in the high N and low N plants was expressed in a good negative linear corre- lation between increasing dry weight and decreasing nitrate concentration respective of the low N plants of both species on the one hand and an only slightly evident and insignifi- cant correlation with regard to the high N plants on the other (Table 2).
Whereas no remarkable differences between Atriplex and Amaranthus were evident with respect to the develop- ment of the nitrate contents of the whole plants, consider- able distinctions between the two species as regards the nitrate distribution between different organs could be de- tected (Fig. 9). Young plants of both species had the highest nitrate concentrations in the shoot axis (Atriplex up to 1 530 gmol NO~/g dry wt, Amaranthus up to 1 770 gmol NO3/g dry wt). The lowest concentrations were observed in the laminae (Atriplex up to 240 gmol NO~/g dry wt, Amaranthus up to 110 I~mol NO~/g dry wt) during the first half of the growing season. As indicated by these values, the gradient of nitrate concentration from the shoot axis to the laminae was greater with Amaranthus than with
Nitrate content
g dry wt I [g dr-"~y-wi-J
10000 100
IOO0 10-
100 1-
10 0,1-
0.01 - 1
t 210
A
= I 810 i I I- 4 0 t 60 100 120
[days]
Nitrate content
. , , NO;-. 1 r.mo,.03-1 g d~-~-~wT-J [g dry wt J
100- 100oo
lOOO lO -
lOO 1
10 0.1
1 O,01
2'0
B
t
I I I I 1 I. t/.0 60 80 100 T 120
[days ]
Fig. 8A, B. Development of the nitrate content in the total plants. A Atriplex; B Amaranthus. �9 high N set, o low N set, * P<0.05, �9 * P<0.01, *** P<0.O01, T fertilization of the high N sets
Atriplex even during this period. In most cases the corre- sponding data respective of the midribs and petioles and of the roots ranged between these extremes during the initial period. This sequence was altered in the case of Amaranthus during the second part of the growing season inasmuch as higher nitrate concentrations then became apparent in the roots than in the shoot axis. In the Atriplex plants, however, the nitrate concentrations of the midribs and pe- tioles, shoot axis and roots decreased in a nearly exponen- tial manner until the end of the experiment, whereas the nitrate values of the laminae stabilized at a minimum value of about 3.5 gmol NO~-/g dry wt. Thus - especially with regard to the low N Atriplex - a far-reaching conversion of the initial nitrate gradient between the different organs took place (Fig. 9). Furthermore, comparison of the two species shows that A triplex had far higher nitrate concentra- tions in the laminae than did Amaranthus on an average,
3:10
Nitrate content
mg 1 [.mo= ,o; 1 9 d---d~7-~i-I L g dry wt j
10000 lOO-
lO-
1
o,1
1 O,Ol
A
1000
100
10
I I I I I
20 40 t 60 80 100
T [days]
Nitrate content
mg .o ; - . 1 .03-1 g d r y w t j [g d r y w t j
loooo lOO.
1ooo 1o-
IOO I-
IO o,1-
I o.oi -
J
2O
B
I I I I I t/,0 60 80 100 t 120
T [ days ]
Nitrate content
mg NO3--N ] pmoL Noi l g d - - - ~ - ~ - l g d~y wt j
1000o lOO-
lO-
1-
o,1-
O, Ol -
lOOO
lOO
lO
1
I I I I 2 l,O 60 80 100
T [days}
Nitrate content
Ira. No -. 1 [.moL No,- 1 [ g d r y w t ] [ g d r y w t J
k
100 10000
10 - 1000
lOO 1
lO o,1
1 O, Ol
2=0
D
/,i 0 I ; I I 60 8 100 120
T [days]
Fig. 9A-D. Development of the nitrate content in different plant organs. A Atriplex high N set; B Amaranthus high N set; C Atriplex low N set; D Amaranthus low N set. �9 laminae, o midribs + petioles (+ branches of the shoot axis, in the case of A triplex), �9 branches of the shoot axis (in the case of Amaranthus), x shoot axis, zx roots, + reproductive organs, T fertilization of the high N sets
whereas Amaranthus exhibited higher mean nitrate values in the roots.
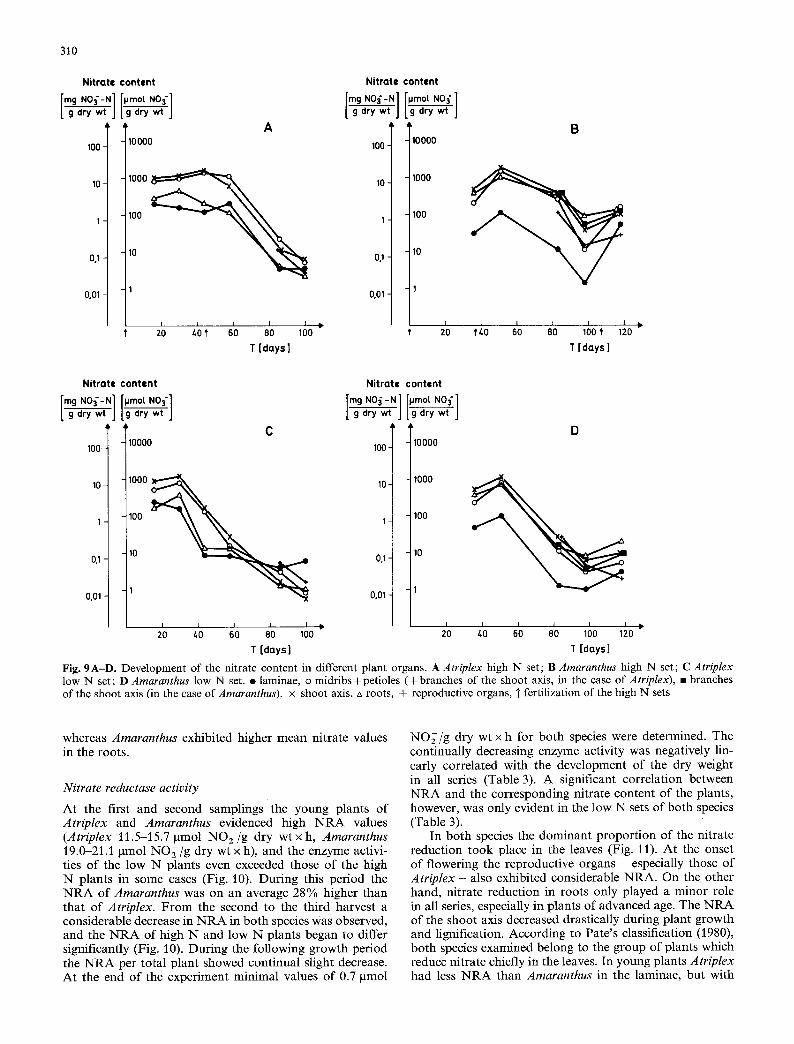
Nitrate reductase activity
At the first and second samplings the young plants of Atriplex and Amaranthus evidenced high N R A values (Atriplex 11.5-15.7 gmol N O z / g dry w t x h , Amaranthus 19.0-21.1 pmol N O 2 / g dry wt x h), and the enzyme activi- ties of the low N plants even exceeded those of the high N plants in some cases (Fig. 10). During this period the N R A of Amaranthus was on an average 28% higher than that o f Atriplex. From the second to the third harvest a considerable decrease in N R A in both species was observed, and the N R A of high N and low N plants began to differ significantly (Fig. 10). During the following growth period the N R A per total plant showed continual slight decrease. At the end of the experiment minimal values of 0.7 pmol
N O ; / g dry wt x h for both species were determined. The continually decreasing enzyme activity was negatively lin- early correlated with the development of the dry weight in all series (Table 3). A significant correlation between N R A and the corresponding nitrate content of the plants, however, was only evident in the low N sets of both species (Table 3).
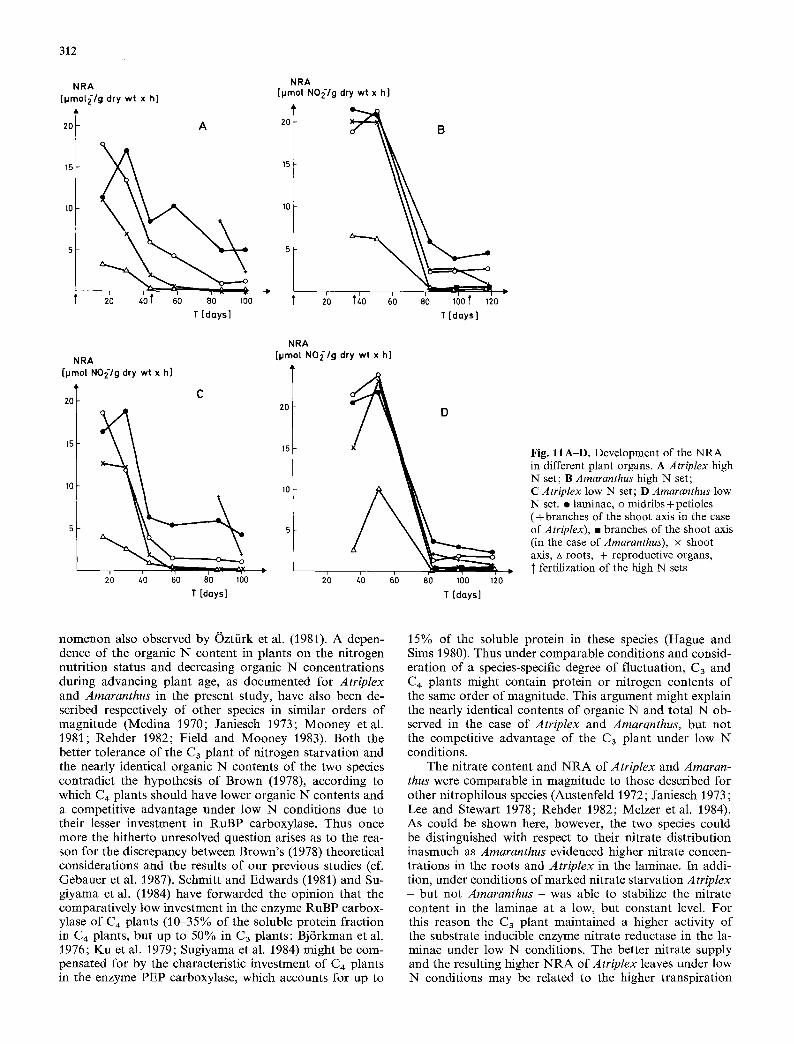
In both species the dominant proport ion of the nitrate reduction took place in the leaves (Fig. 11). At the onset of flowering the reproductive organs - especially those of Atriplex - also exhibited considerable NRA. On the other hand, nitrate reduction in roots only played a minor role in all series, especially in plants of advanced age. The N R A of the shoot axis decreased drastically during plant growth and lignification. According to Pate's classification (1980), both species examined belong to the group of plants which reduce nitrate chiefly in the leaves. In young plants Atriplex had less N R A than Amaranthus in the laminae, but with

NRA [pmol NO2/g dry wt x hi
2~ T
20 /,0 t 60 80 100
T [days ]
NRA [pmol NO2-/g dry wt x h]
A
20 B
15
10
5
'o ' ' 2 t/.0 60 80 100 t 120
T [days]
Fig. 10A, B. Development of the NRA in the total plants. A Atrip- lex; B Amaranthus. �9 high N set, o low N set, * P<0.05, ** P< 0.01, T fertilization of the high N sets
Table 3. Linear correlations between dry weight and NRA and nitrate content and NRA with Atriplex hortensis and Arnaranthus retroflexus of various ages
Correlated Series r Slope y- df P< parameters inter-
cept
Dry weight/ Atriplex NRA High N set -0.9368 -0.339 11.98 4 0.01
Low N set --0.9431 --0.968 15.33 4 0.01
Arnaranthus High N set --0.9964 --0.331 19.80 3 0.001 Low N set --0.9938 --1.209 20.45 3 0.001
Nitrate A triplex content/ High N set 0.5144 0.010 2.79 4 n.s. NRA Low N set 0.9991 0.038 0.96 4 0.001
Amaranthus High N set 0.6465 0.028 2.16 3 n.s. Low N set 0.9455 0.057 1.50 3 0.05
311
advancing age high N and low N Atriplex plants were able to maintain higher NRA in this important organ (Fig. 11). The late fertilization of Amaranthus resulted in only a slight increase of NRA in the laminae.
Transpiration
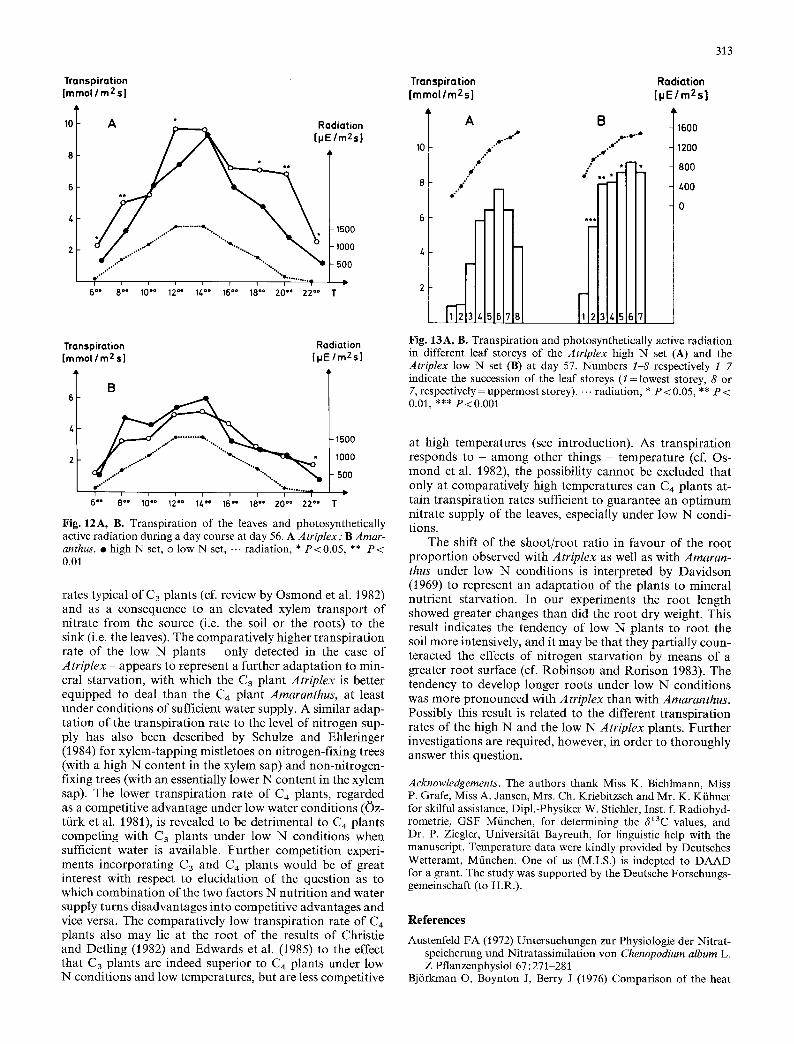
With the exception of the early morning (6 oo and 8 ~176 h) and the late evening hours (22 oo h), substantially lower transpiration values were determined for the youngest fully developed leaves of the C4 plant Amaranthus than for those of the C3 plant Atriplex during the course of a day (Fig. 12). When the measurement at 22 oo h is disregarded, no signifi- cant differences between the high N and the low N set of Amaranthus were observed. On the other hand the tran- spiration of the low N Atriplex was clearly and in most cases significantly higher than that of the corresponding high N plants. This result was verified in several repetitions (not illustrated).
The above results are confirmed by ~13C data obtained from the high N and low N plants of both species. The average ~13C values of the leaves in the high N and the low N set of the C4 plant Amaranthus were exactly identical (-12.97%o; high N set s: 0.16; low N set s: 0.19). With Atriplex a C3 plant-typical increased 13C discrimination was observed, i.e. more negative ~3C values, and further- more a significant difference between the high N set (6a 3C = -28.01%o; s: 0.33) and the low N set (fi13C=-28.95%o; s: 0.40) of the C3 plant was evident. The more negative 6a3C value of the low N Atriplex indicates a more intense transpiration (i.e. reduced stomatal diffusion resistance and thus increased internal CO/concentrations) of the C3 plant under low N conditions integrated over the growing season.
As indicated in Fig. 13, Atriplex leaves of various ages also usually reflected the distinct transpiration patterns of high N and low N plants. In both sets increasing transpira- tion values were observed to progress from the oldest (storey 1) up to the youngest fully developed leaves (storey 6). Not fully developed leaves (storeys 7 and 7 and 8, respec- tively), however, showed reduced transpiration rates. Whereas the mean radiation level available to the upper- most leaf storeys of the high N and low N plants was equal, the average radiation reaching the lower storeys decreased more pronouncedly with the high N plants, due to the denser leaf cover resulting intensified self-shading (Fig. 13). For the same reason the transpiration gradient from storey to storey was steeper with the high N plants than with the low N plants.
Discussion
As previously observed in competition experiments with the monocotylous C3 plants Hordeum vulgare and Arena sativa and the C4 plants Panieum miliaceurn and P. crus- galli (Gebauer et al. 1987), the dicotylous C3 plant Atriplex hortensis was also better able tolerate low N nutrition levels than was the C4 plant Amaranthus retroflexus under our experimental conditions. This superiority of the C3 plant in pure culture was enhanced under the competitive strain of mixed culture. During no phase of the plant growth period were any pronounced differences found respective of the organic N or the total N content in corresponding sets of the two species. The roots of the C4 plant Amaran- thus even tended to possess higher total N contents, a phe-
312
NRA [prno[2-/g dry wt x hi
2O
15
10
2O
A
4c) '~ 60 80 lOO T [days]
NRA [pmol NO2-/g dry wt x hi
2O
2'0 40 6'0 80 100 T [days]
NRA [pmol NO2-/g dry wt x h]
20
15
10
5
2'o ~'J,o 6'o 8'o ~ ' loo ~ ~2o T [days]
NRA [proof NO;~/g dry wt x hi
20
15
10
D
2'o 4.O 6'0 80 1~o 12o T [days]
F i g . llA-D. Development of the NRA in different plant organs. A Atriplex high N set; B Amaranthus high N set; C Atriplex low N set; D Amaranthus low N set. �9 laminae, o midribs + petioles (+branches of the shoot axis in the case of Atriplex), �9 branches of the shoot axis (in the case of Amaranthus), x shoot axis, zx roots, + reproductive organs, T fertilization of the high N sets
nomenon also observed by Oztfirk et al. (1981). A depen- dence of the organic N content in plants on the nitrogen nutrition status and decreasing organic N concentrations during advancing plant age, as documented for Atriplex and Amaranthus in the present study, have also been de- scribed respectively of other species in similar orders of magnitude (Medina 1970; Janiesch 1973; Mooney etal. 1981; Rehder 1982; Field and Mooney 1983). Both the better tolerance of the C3 plant of nitrogen starvation and the nearly identical organic N contents of the two species contradict the hypothesis of Brown (1978), according to which C4 plants should have lower organic N contents and a competitive advantage under low N conditions due to their lesser investment in RuBP carboxylase. Thus once more the hitherto unresolved question arises as to the rea- son for the discrepancy between Brown's (1978) theoretical considerations and the results of our previous studies (cf. Gebauer et al. 1987). Schmitt and Edwards (1981) and Su- giyama et al. (1984) have forwarded the opinion that the comparatively low investment in the enzyme RuBP carbox- ylase of C4 plants (10-35% of the soluble protein fraction in C4 plants, but up to 50% in C3 plants: Bj6rkman et al. 1976; Ku et al. 1979; Sugiyama et al. 1984) might be com- pensated for by the characteristic investment of C4 plants in the enzyme PEP carboxylase, which accounts for up to
15% of the soluble protein in these species (Hague and Sims 1980). Thus under comparable conditions and consid- eration of a species-specific degree of fluctuation, C3 and C4 plants might contain protein or nitrogen contents of the same order of magnitude. This argument might explain the nearly identical contents of organic N and total N ob- served in the case of Atriplex and Amaranthus, but not the competitive advantage of the C3 plant under low N conditions.
The nitrate content and NRA of Atriplex and Amaran- thus were comparable in magnitude to those described for other nitrophilous species (Austenfeld 1972; Janiesch 1973 ; Lee and Stewart 1978; Rehder 1982; Melzer et al. 1984). As could be shown here, however, the two species could be distinguished with respect to their nitrate distribution inasmuch as Amaranthus evidenced higher nitrate concen- trations in the roots and Atriplex in the laminae. In addi- tion, under conditions of marked nitrate starvation A triplex - but not Amaranthus - was able to stabilize the nitrate content in the laminae at a low, but constant level. For this reason the C3 plant maintained a higher activity of the substrate inducible enzyme nitrate reductase in the la- minae under low N conditions. The better nitrate supply and the resulting higher NRA of Atriplex leaves under low N conditions may be related to the higher transpiration
Transpiration [mmollm2s]
10
8
.~176 ~176176176 "*~176176176176 "L~176 ,,~176176
r i r r i "~176176176 . . . . . . . . ' ~ i i 6 " " 8 " " 10 -0 12 ~176 1/, ~ 16 ~ 18"" 2 0 " * 22 ~
Radiation [pE/m2s]
1500
- 1000
500
Transpiration Radiation [mmoi/m2s] [pE/rn2s]
B
- 1500
2 - 1000
500
i i i i i i i IT" r "~ 6 . . 80o 10 o, 12.0 1/,o* 16 ~ 18 ~ 200= 22 .0
Fig. 12A, B. Transpiration of the leaves and photosynthetically active radiation during a day course at day 56. A Atriplex; B Amar- anthus. �9 high N set, o low N set, -.. radiation, * P<0.05, ** P < 0.01
rates typical of C3 plants (cf. review by Osmond et al. 1982) and as a consequence to an elevated xylem t ranspor t of nitrate from the source (i.e. the soil or the roots) to the sink (i.e. the leaves). The comparat ively higher t ranspirat ion rate of the low N plants - only detected in the case o f Atriplex appears to represent a further adapta t ion to min- eral starvation, with which the C3 plant Atriplex is better equipped to deal than the C4 plant Amaranthus, at least under condit ions of sufficient water supply. A similar adap- tat ion of the t ranspira t ion rate to the level of ni trogen sup- ply has also been described by Schulze and Ehleringer (1984) for xylem-tapping mistletoes on nitrogen-fixing trees (with a high N content in the xylem sap) and non-ni trogen- fixing trees (with an essentially lower N content in the xylem sap). The lower t ranspira t ion rate of C , plants, regarded as a competi t ive advantage under low water condit ions (C)z- ti irk et al. 1981), is revealed to be detr imental to C4 plants competing with Ca plants under low N condit ions when sufficient water is available. Fur ther compet i t ion experi- ments incorporat ing C3 and C4 plants would be of great interest with respect to elucidation of the question as to which combinat ion of the two factors N nutr i t ion and water supply turns disadvantages into competit ive advantages and vice versa. The comparat ively low t ranspira t ion rate of C4 plants also may lie at the root of the results of Christie and Detl ing (1982) and Edwards et al. (1985) to the effect that C3 plants are indeed superior to C4 plants under low N condit ions and low temperatures, but are less competi t ive
313
Transpiration [mmol/m2s]
A .e..-e "e
1C ~ e
8 ..g" -~ o"
6
/,
2
F
B e . o �9
.o" f . . e " -
3 / , 5 6
Radiation [pE/mZs]
1600
1200
* 800
/,00
0 I I I
!
I
Fig. 13A, B. Transpiration and photosynthetically active radiation in different leaf storeys of the Atriplex high N set (A) and the Atriplex low N set (B) at day 57. Numbers 1-8 respectively 1-7 indicate the succession of the leaf storeys (1 : lowest storey, 8 or 7, respectively = uppermost storey). -.- radiation, * P < 0.05, ** P < 0.01, *** P<0.001
at high temperatures (see introduction). As t ranspira t ion responds to - among other things - temperature (cf. Os- mond et al. 1982), the possibil i ty cannot be excluded that only at comparat ively high temperatures can C4 plants at- tain t ranspira t ion rates sufficient to guarantee an op t imum nitrate supply of the leaves, especially under low N condi- tions.
The shift of the shoot / root ratio in favour of the root p ropor t ion observed with Atriplex as well as with Amaran- thus under low N condit ions is interpreted by Davidson (1969) to represent an adapta t ion of the plants to mineral nutrient starvation. In our experiments the root length showed greater changes than did the root dry weight. This result indicates the tendency of low N plants to root the soil more intensively, and it may be that they par t ia l ly coun- teracted the effects of ni trogen starvat ion by means o f a greater root surface (cf. Robinson and Ror ison 1983). The tendency to develop longer roots under low N condit ions was more pronounced with Atriplex than with Amaranthus. Possibly this result is related to the different t ranspira t ion rates of the high N and the low N Atriplex plants. Fur ther investigations are required, however, in order to thoroughly answer this question.
Acknowledgements. The authors thank Miss K. Bichlmann, Miss P. Grafe, Miss A. Jansen, Mrs. Ch. Kriebitzsch and Mr. K. Kfihner for skilful assistance, Dipl.-Physiker W. Stichler, Inst. f. Radiohyd- rometrie, GSF Miinchen, for determining the 6~aC values, and Dr. P. Ziegler, Universitfit Bayreuth, for linguistic help with the manuscript. Temperature data were kindly provided by Deutsches Wetteramt, Mfinchen. One of us (M.I.S.) is indepted to DAAD for a grant. The study was supported by the Deutsche Forschungs- gemeinschaft (to H.R.).
R e f e r e n c e s
Austenfeld FA (1972) Untersuchungen zur Physiologie der Nitrat- speicherung und Nitratassimilation von Chenopodium album L. Z Pflanzenphysiol 67: 271-281
Bj6rkman O, Boynton J, Berry J (1976) Comparison of the heat

314
stability of photosynthesis, chloroplast membrane reactions, photosynthetic enzymes, and soluble protein in leaves of heat- adapted and cool-adapted C4 species. Carnegie Inst Washing- ton Yearb 75:400-407
Bremner JM, Keeney DR (1965) Steam distillation methods for determination of ammonium, nitrate and nitrite. Anal Chim Acta 32:485-495
Brown RH (1978) A difference in the N use efficiency in C3 and C4 plants and its implications in adaptation and evolution. Crop Sci 18 : 93-98
Christie EK, Detling JK (1982) Analysis of interference between C3 and C4 grasses in relation to temperature and soil nitrogen supply. Ecology 63 : 1277-1284
Davidson RL (1969) Effects of soil nutrients and moisture on root/ shoot ratios in Loliurn perenne L. and Trifolium repens L. Ann Bot (London) 33:571-577
Doliner LH, Jolliffe PA (1979) Ecological evidence concerning the adaptive significance of the C4 dicarboxylic acid pathway of photosynthesis. Oecologia (Berlin) 38:23-34
Edwards GE, Ku MSB, Monson RK (1985) C4 photosynthesis and its regulation. In: Barber J, Baker NR (eds) Photosynthetic mechanisms and the environment. Elsevier, Amsterdam New York, pp 288-327
Ellenberg H (1979) Zeigerwerte der Gef/il3pflanzen Mitteleuropas. Scripta Geobotanica, Bd 9. Goltze, G6ttingen
Field C, Mooney HA (1983) Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a California shrub. Oecolo- gia (Berlin) 56: 348-355
Gebauer G, Melzer A, Rehder H (1984) Nitrate content and nitrate reductase activity in Rumex obtusifolius L. I. Differences in or- gans and diurnal changes. Oecologia (Berlin) 63:136-142
Gebauer G, Schubert B, Schuhmacher MI, Rehder H, Ziegler H (1987) Biomass production and nitrogen content of C3- and C4-grasses in pure and mixed culture with different nitrogen supply. Oecologia (Berlin) 71:613-617
Gerlach A (1973) Methodische Untersuchungen zur Bestimmung der Stickstoffnettomineralisation. Scripta Geobotanica, Bd 5. Goltze, G6ttingen
Hageman RH, Hucklesby DP (1971) Nitrate reductase from higher plants. In: San Pietro A (ed) Methods in enzymology, vol 23. Academic Press, London New York, pp 491-503
Hague DR, Sims TL (1980) Evidence for light-stimulated synthesis of phosphoenolpyruvate carboxylase in leaves of maize. Plant Physiol 66 : 505-509
Janiesch P (1973) Beitrag zur Physiologic der Nitrophyten. Nitrat- speicherung und Nitratassimilation bei Anthriscus sylvestris Hoffm. Flora 162:47%491
Jaworski EG (1971) Nitrate reductase assay in intact plant tissues. Biochem Biophys Res Commun 43:1274-1279
Krsti6 B, Sari6 MR (1983) Efficiency of nitrogen utilization and photosynthetic rate in C3 and C4 plants. In: Sari6 MR, Lough- man BC (eds) Genetic aspects of plant nutrition. Nijhoff/Junk, The Hague Boston Lancaster, pp 255-260
Ku SB, Schmitt MR, Edwards GE (1979) Quantitative determina- tion of RuBP-carboxylase-oxygenase protein in leaves of sever- al C3 and C4 plants. J Exp Bot 30:89-98
Lee JA, Stewart RG (1978) Ecological aspects of nitrogen assimila- tion. Adv Bot Res 6:2-43
Medina E (1970) Relationships between nitrogen level, photosyn- thetic capacity and carboxydismutase activity in A triplex patula leaves. Carnegie Inst Washington Yearb 69: 655-662
Melzer A, Gebauer G, Rehder H (1984) Nitrate content and nitrate reductase activity in Rumex obtusifolius L. II. Responses to nitrate starvation and nitrogen fertilization. Oecologia (Berlin) 63 : 380-385
Mooney HA, Field C, Gulmon SL, Bazzaz FA (1981) Photosyn- thetic capacity in relation to leaf position in desert versus old- field annuals. Oecologia (Berlin) 50:109-112
Osmond CB, Winter K, Ziegler H (1982) Functional significance of different pathways of CO2 fixation in photosynthesis. In: Lange OL, Nobel PS, Osmond CB, Ziegler H (eds) Encyclope- dia of plant physiology, New Series, vol 12B. Springer, Berlin Heidelberg New York, pp 480-547
Osmond CB, Ziegler H, Stichler W, Trimborn P (1975) Carbon isotope discrimination in alpine succulent plants supposed to be capable of Crassulacean acid metabolism (CAM). Oecologia (Berlin) 18:209-217
Oztfirk M, Rehder H, Ziegler H (1981) Biomass production of C3- and C4-plant species in pure and mixed culture with differ- ent water supply. Oecologia (Berlin) 50 : 73-81
Pate JS (1980) Transport and partitioning of nitrogenous solutes. Ann Rev Plant Physiol 31:313-340
Pearcy RW, Ehleringer J (1984) Comparative ecophysiology of C3 and C4 plants. Plant Cell Environ 7:1-13
Rehder H (1982) Nitrogen relations of ruderal communities (Rum# eion alpini) in the northern calcareous alps. Oecologia (Berlin) 55:120-129
Robinson D, Rorison JH (1983) Relationships between root mor- phology and nitrogen availability in a recent theoretical model describing nitrogen uptake from soil. Plant Cell Environ 6: 641-647
Schmitt MR, Edwards GE (1981) Photosynthetic capacity and ni- trogen use efficiency of maize, wheat, and rice: A comparison of C3 and C4 photosynthesis. J Exp Bot 32:459-466
Schulze ED, Ehleringer JR (1984) The effect of nitrogen supply on growth and water-use efficiency of xylem-tapping mistletoes. Planta 162 : 268-275
Shaner DL, Boyer JS (1976) Nitrate reductase activity in maize (Zea mays L.) leaves. II. Regulation by nitrate flux at low leaf water potential. Plant Physiol 58 : 505-509
Sugiyama T, Mizuno M, Hayashi M (1984) Partitioning of nitrogen among ribulose-l,5-bisphosphate carboxylase/oxygenase, phos- phoenolpyruvate carboxylase, and pyruvate orthophosphate dikinase as related to biomass productivity in maize seedlings. Plant Physiol 75 : 665-669
Thayer JR, Huffaker RC (1980) Determination of nitrate and ni- trite by high-pressure liquid chromatography: comparison with other methods for nitrate determination. Analyt Biochem 102:110-119
Received October 28, 1986


















