
BIOLOGY ESSAY PACK · biology essay pack i. carbohydrates 2 ii. lipids 5 iii. proteins 8 iv. ... x....
97
COMPILED BY: GABRIELLA CRESSIDA BOONE (RAFFLES INSTITUTION, 2015) BIOLOGY ESSAY PACK I. CARBOHYDRATES 2 II. LIPIDS 5 III. PROTEINS 8 IV. ENZYMES 13 V. CELL STRUCTURE 18 VI. CELL MEMBRANE 21 VII. MITOSIS & MEIOSIS 23 VIII. DNA & GENOMICS 28 IX. VIRUSES 34 X. BACTERIA 39 XI. PROKARYOTIC & EUKARYOTIC GENOMES 45 XII. GENETICS 49 XIII. RESPIRATION 53 XIV. PHOTOSYNTHESIS 55 XV. ISOLATING, CLONING & SEQUENCING DNA 60 XVI. STEM CELLS & GENE THERAPY 72 XVII. GENETICALLY MODIFIED ORGANISMS 78 XVIII. PLANT CLONING 80 XIX. DIVERSITY & EVOLUTION 83 XX. HOMEOSTASIS 90 XXI. CELL SIGNALING 91 XXII. NERVOUS SYSTEM 94
Transcript of BIOLOGY ESSAY PACK · biology essay pack i. carbohydrates 2 ii. lipids 5 iii. proteins 8 iv. ... x....

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
BIOLOGYESSAYPACK
I.CARBOHYDRATES 2
II.LIPIDS 5
III.PROTEINS 8
IV.ENZYMES 13
V.CELLSTRUCTURE 18
VI.CELLMEMBRANE 21
VII.MITOSIS&MEIOSIS 23
VIII.DNA&GENOMICS 28
IX.VIRUSES 34
X.BACTERIA 39
XI.PROKARYOTIC&EUKARYOTICGENOMES 45
XII.GENETICS 49
XIII.RESPIRATION 53
XIV.PHOTOSYNTHESIS 55
XV.ISOLATING,CLONING&SEQUENCINGDNA 60
XVI.STEMCELLS&GENETHERAPY 72
XVII.GENETICALLYMODIFIEDORGANISMS 78
XVIII.PLANTCLONING 80
XIX.DIVERSITY&EVOLUTION 83
XX.HOMEOSTASIS 90
XXI.CELLSIGNALING 91
XXII.NERVOUSSYSTEM 94

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
I.CARBOHYDRATES
1. Describethestructureofglycogen.[5m]
1 HELICAL1.Glycogenismadeupofα-glucosemonomers*linkedbyα(1-4)glycosidicbonds*withinabranch.2.Sinceeachresidueisbent inonedirectionwithrespecttothepreviousresidue,theglycogenmoleculeiscoiledintoahelix*.3.Hydrogenbonding*ispresentbetweentheinward-projectinghydroxylgroups*oftheresidues,allowingthehelixtomaintainitsstructure.(Note:glycosidicbondcanbebrokenbyadditionofwaterinhydrolysisreaction,enzymaticactionofmaltase,oradditionofhydrochloricacidinacidhydrolysis)
2 BRANCHING4.Atbranchpoints,α(1-6)glycosidicbonds*arepresent.5.Branching*ismoreextensiveinglycogenthaninthatofamylopectin,occurringataboutevery8-10glucoseresidues,optimisingpackingofmanyglucosesubunitsperunitvolume.6.Extensivebranchinggivesalotmoreends*fromwhichmultipleenzymes*canworkontohydrolysetheglycosidicbonds,increasingenergygenerationperunittime.
2. Explainhowthestructureofcelluloseissuitedtoitsfunction.[6m]
1 HIGHTENSILESTRENGTH1.Celulloseismadeupofβ-glucosemonomers*linkedbyβ(1-4)glycosidicbonds*.2.Asalternateβ-glucosemonomersarerotated180°withrespecttoeachother,straight*moleculesareformed,allowingthemtolieparallel*toeachother.3.Outward-projectinghydroxylgroups*oftheresiduesineitherdirectionallowextensivehydrogenbonding*betweenadjacentparallelchains,formingmicrofibrils*.4.Theassociationofnumerouscellulosemoleculesthroughintermolecularhydrogenbondinggivesrisetohightensilestrength*,conferringstrengthtothecellwall*.
2 INSOLUBLE5.Celluloseisalargemolecule/macromolecule*.6. Since the hydroxyl groups projecting from the parallel* cellulose molecules have already been used inintermolecular hydrogen bonding, only the surface of the microfibril is exposed to water. Thus, there arerelatively fewer free hydroxyl groups* available for hydrogen bonding* with the surrounding watermolecules*.7.Cellulosecanthereforeremaininsoluble*,andtheintegrityofthecellwall*ismaintainedinanaqueousenvironment.
3 POROUS8.Throughcriss-crossing,ameshwork*ofmicrofibrilsformsthecellulosecellwall.9.Themeshworkhasaporous*structureduetogapsbetweenthemicrofibrils,makingitfreelypermeabletowaterandsolutes.Itallowsfreemovementofsubstances*inandoutofthecell,butitselfdoesnotaffectthewaterpotential*ofthecellasitisinsoluble.
4 STRONG&RIGID10. Themeshwork alsodistributes the stresses* in all directions. It serves to enclose the plant cell andprotectitfromphysicaldamageandburstingduetoosmoticstress.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. Comparethestructures,functionsandpropertiesofcellulose,starch,andglycogen.[9m]
FEATURE CELLULOSE STARCH GLYCOGEN1 MONOMERS β-glucose α-glucose2 BONDS β(1-4)glycosidicbond
Amylose:α(1-4)glycosidicbondAmylopectin:α(1-4)+α(1-6)glycosidicbond
α(1-4)glycosidicbondα(1-6)glycosidicbond(evenmorebranchedthanamylopectin)
Intermolecularhydrogenbonds
Intramolecularhydrogenbonds
3 STRUCTUREOFMOLECULES
UnbranchedlinearLong,straightchain
Amylose:unbranchedhelicalAmylopectin:branchedhelical(every20residues)
Branchedhelical(every8-10residues)
4 FUNCTION Structural Energystorage(plants) Energystorage(animals)5 PROPERTIES Insoluble*inwater:
-Largesize-IntermolecularH-bondsèDoesnotaffectwaterpotential*èIntegrityofthecellwall*ismaintainedinaqueousenvironment
Insoluble*inwater:-Largesize-IntramolecularH-bondsèDoesnotaffectwaterpotential*
Insolubleinwater:-Largesize-IntramolecularH-bondsèDoesnotaffectwaterpotential*
Hightensilestrength*:-Enzymes*thatrecogniseandhydrolyseβ(1-4)glycosidic*bondsarerare*-Alternatemonomersrotated180°withrespecttoeachother-Forminglongstraightchains-Hydroxylgroups*projectedoutwardsineitherdirection-Allowingnumeroushydrogenbonds*toformbetweenadjacentparallelchains(intermolecularhydrogenbonding)formingmicrofibrils*-Meshworkofcriss-crossingmicrofibrilsformcellulosecellwall*èConferstrengthtocellwall*
Compact*:-Enzymes*(amylase)thatrecogniseandhydrolyseα(1-4)glycosidic*bondscommonlyavailable-Monomersbondedinsameorientation-Forminghelicalstructure-Branchingincreasesaccessibilityforhydrolyticactionbyenzymes*likeamylases,manyofwhichworkfromthetipsofthebranches-OptimisingpackingofmanyglucoseunitsperunitvolumeèMoreglucoseunitscanbeoxidizedtoproducemoreenergyperunittime
Compact*:-Monomersbondedinsameorientation-Forminghelicalstructure-Branchingincreasesaccessibilityforhydrolyticactionbyenzymes*likemaltases,manyofwhichworkfromthetipsofthebranches-OptimisingpackingofmanyglucoseunitsperunitvolumeèMoreglucoseunitscanbeoxidizedtoproducemoreenergyperunittime

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
II.LIPIDS1. Explainhowthemolecularstructureoftriglyceridesisrelatedtotheirfunction.[6m]
1 MOLECULARSTRUCTURE1. Triglycerides aremade up of3 long non-polar*, hydrophobic* hydrocarbon tails/chains* joined to aglycerol*molecule.2.Eachchainisconnectedtotheglycerolmoleculeviaesterlinkages*throughaprocessofcondensation*.
2 ENERGY3.Duetothelonghydrocarbontails,therearenumeroushydrogenandcarbonatomscomparedtooxygenatoms,more so in a triglycerideas compared toanequivalentmass of carbohydrates.Agramof fat storesmorethantwiceasmuchenergyasagramofglycogenorstarch.Thus,itcanstoremoreenergyperunitmass*thanotherrespiratorysubstrates.4.ThisveryhighproportionofC-Hbondsallowsmoreoxidation*totakeplace,formingmoreenergy*intheformofATP*.
3 METABOLICWATER5.Oxidation*alsoreleasesmetabolicwater*intheprocess.6.Thisoccursbecausethehydrogenatomswillcombinewiththeoxygensuppliedduringoxidationtoformwatermolecules.7. It isusedasasourceofwater foranimalsespeciallydesertanimals, suchas thekangaroorat,asexternalwatersupplyislimiting.
4 INSOLUBILITYOFHYDROCARBONTAILS8. Due to their long hydrophobic hydrocarbon tails that make up the most of these lipid molecules,triglyceridesareinsolubleinwater*.9.These tails arenon-polar and so cannot form hydrogen bonds* with water, thusnot affecting waterpotentialwhileservingasenergystorages.
2. Suggestwhyplantcellsmainlystorecarbohydratesandanimalcellsmainlystorelipids.[4m]
1 TRIGLYCERIDESLipids,intheformoftriglycerides,arestoredbyanimalscells.Triglyceridesaremadeupof3longnon-polar*,hydrophobic*hydrocarbontails/chains*joinedtoaglycerol*molecule.Duetothelonghydrocarbontails,therearenumerous hydrogen and carbon atomscompared tooxygen atoms,moreso ina triglycerideascompared to an equivalent mass of carbohydrates. This very high proportion of C-H bonds allows moreoxidation*totakeplace,formingmoreenergy*intheformofATP*.
2 LIPIDSVSCARBOHYDRATESAnimalsaremobileandneedtocarryenergy storeswiththemsothereisanadvantagetohavingamorecompact reservoir of fuel, which are lipids. These lipids are stored in their adipose cells. Plants arerelativelyimmobilesotheycanfunctionwithbulkyenergystorageintheformofstarch.Oilsaregenerallyfoundinseedswherecompactstorageisanassettotheplant.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. Outlinetherolesofphospholipids,cholesterol,andglycolipidsinmembranestructure.[6m]1 PHOSPHOLIPIDS
Phospholipidsareformedwhen2non-polar*,hydrophobic*hydrocarbontails/chains*andahydrophilicphosphatehead*attachtoaglycerol,resultinginanamphipathicmolecule.Thephospholipidsformabilayer*,themaincomponentofthecellmembrane.Itisafluidlayerinwhichproteinsmove/float.Thephospholipidbilayeractsasabarrier*againstthemovementofions,polarandlargemolecules*.Itservesasabarriertoretaincellcontentsandtocreatecompartmentswithinthecell.
2 CHOLESTEROLCholesterol is a hydrophobic* four-ringed structure with a hydrophilic* end making it an amphipathicmolecule.Cholesterolalignswithphospholipidstoregulatefluidityofthemembrane.Cholesterol has the ability topreventphospholipids fromgetting too close together and thereforepreventssolidificationofmembraneatlowtemperatures;itcanalsopreventthemembranefrombeingexcessivelyfluidandunstableathighertemperaturesasitcanpreventphospholipidsfromdriftingaparttoorapidly.
3 GLYCOLIPIDSGlycolipidsareoligosaccharides(carbohydrates)covalentlybondedtolipids,soeachlipidcanhaveoneormorecarbohydratesattachedtoit.Incellmembranes,glycolipidsarefoundwhenacarbohydratechainisattachedtoaphospholipidmoleculeinthebilayer,helpingtomaintainthestabilityofthemembrane.Theyalsohelpwithcell-cell recognition* andcell-cell binding*. Glycolipids aremadeupofdiverse sugarcomponentsofuniqueshapesthatallowthemtoactasmarkersforcell-cellrecognition*todistinguishcellsas ‘self’ or ‘non-self’ as thebasis of the immune system; cell-cell recognition also result incell adhesion,allowingcellstobeattachedtooneanothertoformtissuesandorgans.Glycolipidsactasreceptors*forcertainbacterialtoxinstobindtohostcells.Glycolipidsarefoundinthemyelinsheath*usedforelectricalinsulationofnervecells.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
4. Explainthedifferencesinmolecularstructurebetweentriglyceridesandphospholipids.[6m] FEATURE TRIGLYCERIDES PHOSPHOLIPIDS1 RESIDUES Triglyceride is made up of a glycerol*
backboneand3non-polar*,hydrophobic*hydrocarbontails*.
Phospholipid is made up of 2 non-polar*,hydrophobic* hydrocarbon tails/chains*and a hydrophilic phosphate head*attachedtoaglycerol.Phospholipids may contain a nitrogen-containingcholinewhichisattachedtothephosphategroup.
2 LINKAGES Theoriginalfattyacidchainsarelinkedtotheglycerolmoleculeviaesterlinkages*.3 POLARITY&
INTERACTIONWITHWATER
The entire molecule is non-polar* and isthereforeentirelyhydrophobic*.
Thenegativelychargedphosphate*makesit hydrophilic* while the hydrocarbontails* are hydrophobic*, resulting in anamphipathicmolecule.
5. Describethefunctionsoftriglyceridesandphospholipidsinlivingorganisms.[6m]
1 LIPIDS–GENERAL
Lipids form aprotective layer* around delicate internal organs* ofmammals. Hence, they act as shockabsorbersandcushionorganssuchaskidneysfrommechanicaldamage.Lipids foundbeneaththe layerofskin(subcutaneous fat)canprovidethermal insulation*as theyarepoorconductors of heat. The layer of fat is especially thick in seals and other marinemammals living in coolerclimates.Lipidsare less dense thanwaterandhence improve buoyancy* inmammals,especiallymarinemammalslikethewhale.Lipidscanfunctionasareservoirforstorageoffatsolublevitamins*likevitaminsA,DandK.Lipidsare insoluble inwater*andhenceareosmotically inactive,meaning theydonot affect thewaterpotential*ofmammaliancells.
2 LIPIDS–TRIGLYCERIDESTriglyceridescanstoremoreenergyperunitmass* thanotherrespiratorysubstrates.Agramoffatstoresmorethantwiceasmuchenergyasagramofglycogenorstarch.They areoxidised* in respiration* to yield significantlymore energy than carbohydrates. Due to the longhydrocarbontails,therearenumeroushydrogenandcarbonatomscomparedtooxygenatoms,moresoinatriglyceride as compared to anequivalent mass of carbohydrates. This very high proportion of C-H bondsallowsmoreoxidation*totakeplace,formingmoreenergy*intheformofATP*.Oxidation* also releases metabolic water* in the process. This occurs because thehydrogen atoms willcombinewiththeoxygensuppliedduringoxidationtoformwatermolecules.Itisusedasasourceofwaterforanimalsespeciallydesertanimals,suchasthekangaroorat,asexternalwatersupplyislimiting.
3 LIPIDS–PHOSPHOLIPIDSPhospholipidsareamphipathic.Thus,thenegatively-charged,hydrophilicphosphateheadsfaceoutwardsand interact with the aqueous environment of the cell interior or exterior, forming hydrogen bonds withwater.Thenon-polar,hydrophobichydrocarbontailstailsfaceinwards,awayfromthewater.Thetailsalsoformhydrophobic interactionswith thehydrocarbon tails of adjacent phospholipidmolecules to form thehydrophobiccoreofthephospholipidbilayer*.Thephospholipidbilayersisamajorcomponentofthecellmembrane*.Thehydrophobiccoreregionisonlypermeable to small hydrophobic solutes and has a very low permeability to and repel polar and chargedmolecules. Thus the bilayer naturally serves as a barrier* to movement of ions, polar and largemolecules*.Passageof thesemoleculesacross themembranearecontrolledbytransmembrane transportproteinsorionchannels.Thephospholipidbilayerhenceallowscompartmentalisationtotakeplacebyactingasaboundarybetweentheintracellularandextracellularaqueousenvironments.Specialisedprocessescanthenbeisolatedineachcompartmenttotakeplaceatoptimalconditions.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
Phospholipids in the cell membrane are also not locked in place but canmove laterally. This allows themembranetobefluidandalsothemovementofincorporatedproteinswithinthebilayer.
III.PROTEINS1. Describe the structure of an amino acid and how a peptide bond is formed with another
aminoacid.[6m]
(DIAGRAM:STRUCTUREOFAMINOACID)
1A STRUCTUREThereare20aminoacidsthatareusedinthebiosynthesisofproteinsincells.Theyarethebasicstructuralunitsormonomersofproteins.Anaminoacidconsistsofacarbonatomthatiscovalentlybondedto4groups:ahydrogenatom,anaminogroup(–NH2),acarboxylgroup(–COOH)andavariableRgroup,alsocalledasidechain.Note:prolinedoesnothaveadistinctRgroupandaminogroupastheyarebondedtooneanother.Amino acids exist as zwitterions, in which they can carry both positive and negative charges. As they aregenerallysolubleinwater,theyionize.Insolution,theaminogroupreceivesanH+andbecomespositivelycharged–NH3+.Thecarboxylgroupdissociates,releasinganH+andbecomesnegativelycharged–COO-.Due to their existence as zwitterions, the amino and carboxyl groups can therefore act as bases and acidsrespectively,andtheiramphotericnatureallowthemtobebuffers.WhenaminoacidsinasolutiondonateoracceptH+, theyminimise changes in the pHof theirsurroundingwhenasmallamountofacidoralkali isadded to it. Such a property is essential where any sudden changes in pH could adversely affect enzymeactivity.
1B Eachofthe20aminoacidshavedifferentRgroupsthatvaryincharge,hydrogen-bondingcapacity,chemical
reactivity, size and shape. Hence, the physical and chemical properties of the R group determines theuniquenessofeachaminoacidandhencethepolypeptideitisapartof.Underthecategoryofneutralaminoacids,therearethosewithnon-polarRgroupswhicharehydrophobic,andalso thosewithpolarR-groupswhicharehydrophilic.Theyarebothelectricallyneutralas thesumofpositiveandnegativechargesareequal.Under the categoryofelectrically-charged amino acids, there are thosewithnegatively-chargedR groupsand those with positively-charged R groups. Negatively-charged R groups have carboxyl groups thatdissociatetoformCOO-,thusbeingacidicaminoacids.Positively-chargedRgroupshaveaminogroupsthatacceptH+to formNH3,thisbeingabasicaminoacid.Astheyarebothcharged,acidicandbasicaminoacidsarehydrophilic.
RGROUPS
NEUTRALAMINOACIDS
NON-POLAR
HYDROPHOBIC
POLAR
HYDROPHILIC
ELECTRICALLY-CHARGEDAMINO
ACIDS
NEGATIVELY-CHARGED
HYDROPHILIC,ACIDIC
POSITIVELY-CHARGED
HYDROPHILIC,BASIC

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
(DIAGRAM:CONDENSATIONREACTIONTOFORMPEPTIDEBOND)
3 PEPTIDEBONDAminoacidsarejoinedviaacondensationreactionthatlinksthecarboxylgroupofoneaminoacidtotheaminogroupofanother,withtheremovalofonewatermolecule.Thisformedapeptidebondjoiningthe2aminoacidsandeachaminoacidmonomeristhenknownasaresidue.
2. Outlinethestructureofaproteinmolecule.[4m]1 OVERALL
Thereare4 levelsoforganisation in thestructureofproteins.Apolypeptidechainhasdirectiondue to its2specific ends, the amino-terminus (N-terminus) and the carboxyl-terminus (C-terminus). The regularlyrepeating part of the chain, excluding theR groups, is referred to as thepolypeptide backbone.When thechainfoldsintoaspecific3Dconformation,itformsaprotein.
2 PRIMARYSTRUCTURETheprimarystructureofaproteinmoleculereferstothenumber*andsequence*ofaminoacidsinasinglepolypeptidechain,anditismaintainedbypeptidebonds*betweenaminoacidresidues.Eachpolypeptidechainisuniqueinitssequence*,number*andtype*ofaminoacids.ItisthissequenceofaminoacidsandtheirRgroups*thatdeterminethetypeandlocationofchemical interactions,andhencethe pattern of folding*. The pattern of folding in turn determines the conformation* and uniquecharacteristicoftheprotein.
3 SECONDARYSTRUCTUREThe secondary structure is formed by regular coiling* or pleating* of a single polypeptide chain. Thestructure ismaintainedbyhydrogen bonds*, formedbetween the CO andNH groupsof thepolypeptidebackbone(note:Rgroupsarenotyetinvolvedinhydrogenbondinginthisstructure).
A-HELIX B-PLEATEDSHEETSA single polypeptide chain is wound into acoiled/spiraledstructure*.Turns of helix are linked by hydrogen bonds*betweentheOoftheCOgroupofoneturn*andtheN of the NH group of the next turn* at regularintervals* of every 4th peptide bond*/four aminoacidsaway.Thisconfersstabilityinthestructure.Thereare3.6aminoacidresiduesineveryturn.(e.g.keratinfoundinhair,woolandnails.)
Two or more segments/regions of a singlepolypeptide chain lie side by side, either runningparallel* or anti-parallel*, forming a flat sheet*whichbecomesfolded.Hydrogen bonds* are formed at irregularintervals* between the CO (or NH) group of onesegment/region* and theNH (or CO) group of anadjacent* segment/region* of a single polypeptidechain.(e.g.fibroinfoundinsilkproducedbysilkwormsandspiders.)
4 TERTIARYSTRUCTUREThe tertiary structure is formedby further extensive folding and bending* of a singlepolypeptide chain,usuallyformingacompact,globular/sphericalmolecule,givingrisetothespecific3Dconformation*ofaprotein.Thestructureismaintainedbyall4typesof interactionsnamelyhydrogenbonds*, ionicbonds*,hydrophobic interactions* and disulfide bonds*, and these are formedbetween theR groups* of aminoacidresidues.
HYDROGENBONDS IONICBONDSHydrogen bonds* are formed between theelectropositive (H) and electronegative (O/N)atoms* of polar R groups*. They are collectivelystrong* andare able to support the conformationofthechain.
Ionic bonds* are formed between oppositely-charged* R groups*. A change in pH* of thesurroundingmediumcanalter thesechargesontheRgroupsandhencetheionicbonds.
HYDROPHOBICINTERACTIONS DISULFIDEBONDSHydrophobic interactions* are formed betweennon-polarRgroups*whicharehydrophobic*.Mosthydrophobic*Rgroupspoint inwards*towardsthecentre/core* of the roughly spherical molecule,shielded from the aqueous environment; mosthydrophilic* R groups face outwards* into theaqueousenvironment,makingtheproteinsoluble*.
Disulfidebonds*areformedbetweentwocysteine*amino acid residues by oxidation of sulfydryl (–SH) groups. They are covalent bonds, and thestrongest*of the4interactions,remainingintactathigher temperatures*. This confers stability of theproteintoheatdenaturation*.
5 QUATERNARYSTRUCTUREThequaternarystructurereferstotheassociationoftwoormore*polypeptidechainsintoonefunctionalproteinmolecule.Eachfoldedpolypeptidechainisreferredtoasasubunit*.Thesubunitsareheldtogether
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
againbyhydrogenbonds,ionicbonds,hydrophobicinteractionsanddisulfidebonds.Proteins with 2 subunits are dimers, those withmore than 2 are oligomers. Constituent chains of themultimericproteincanbeidenticalordifferent.Note: not all proteins have a quaternary structure, monomeric proteins like lysosomes have only onepolypeptideunitandarehenceorganiseduptothetertiarylevelonly.
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
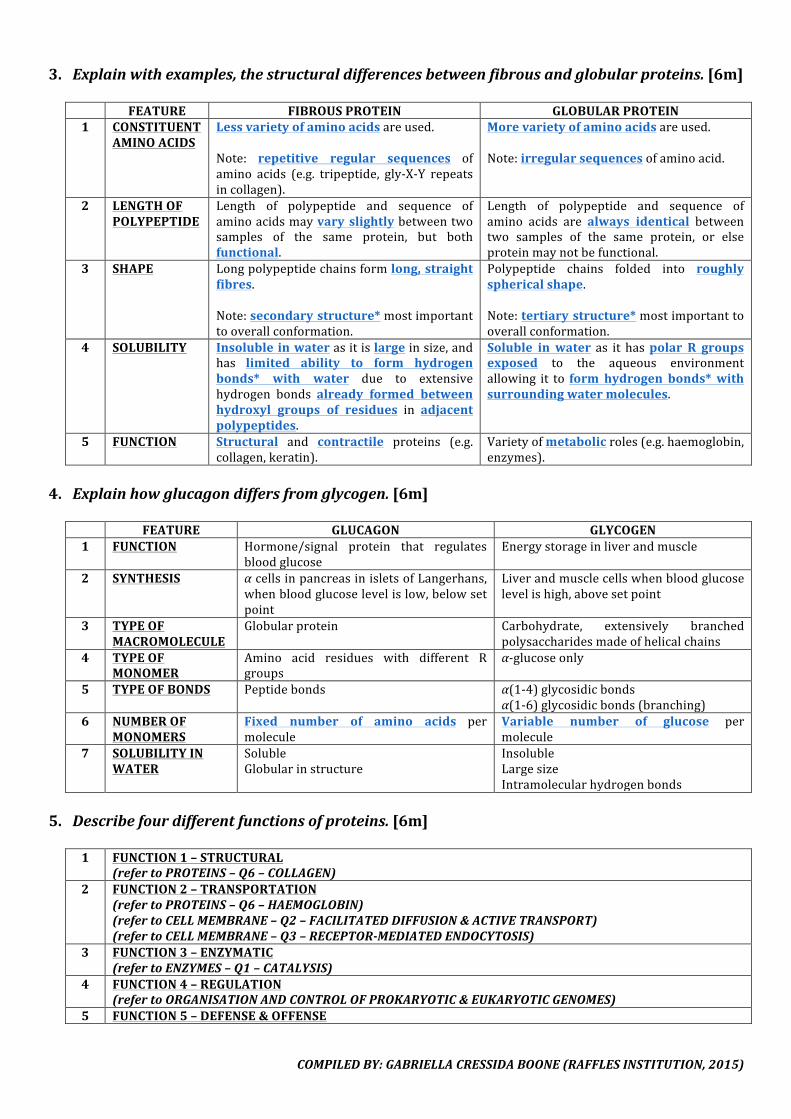
3. Explainwithexamples,thestructuraldifferencesbetweenfibrousandglobularproteins.[6m] FEATURE FIBROUSPROTEIN GLOBULARPROTEIN1 CONSTITUENT
AMINOACIDSLessvarietyofaminoacidsareused.Note: repetitive regular sequences ofamino acids (e.g. tripeptide, gly-X-Y repeatsincollagen).
Morevarietyofaminoacidsareused.Note:irregularsequencesofaminoacid.
2 LENGTHOFPOLYPEPTIDE
Length of polypeptide and sequence ofaminoacidsmayvary slightlybetweentwosamples of the same protein, but bothfunctional.
Length of polypeptide and sequence ofamino acids are always identical betweentwo samples of the same protein, or elseproteinmaynotbefunctional.
3 SHAPE Longpolypeptidechainsformlong,straightfibres.Note:secondarystructure*mostimportanttooverallconformation.
Polypeptide chains folded into roughlysphericalshape.Note:tertiarystructure*mostimportanttooverallconformation.
4 SOLUBILITY Insolubleinwaterasitislargeinsize,andhas limited ability to form hydrogenbonds* with water due to extensivehydrogen bonds already formed betweenhydroxyl groups of residues in adjacentpolypeptides.
Soluble in water as it haspolar R groupsexposed to the aqueous environmentallowing it to form hydrogen bonds*withsurroundingwatermolecules.
5 FUNCTION Structural and contractile proteins (e.g.collagen,keratin).
Varietyofmetabolicroles(e.g.haemoglobin,enzymes).
4. Explainhowglucagondiffersfromglycogen.[6m]
FEATURE GLUCAGON GLYCOGEN1 FUNCTION Hormone/signal protein that regulates
bloodglucoseEnergystorageinliverandmuscle
2 SYNTHESIS αcellsinpancreasinisletsofLangerhans,whenbloodglucoselevelislow,belowsetpoint
Liverandmusclecellswhenbloodglucoselevelishigh,abovesetpoint
3 TYPEOFMACROMOLECULE
Globularprotein Carbohydrate, extensively branchedpolysaccharidesmadeofhelicalchains
4 TYPEOFMONOMER
Amino acid residues with different Rgroups
α-glucoseonly
5 TYPEOFBONDS Peptidebonds α(1-4)glycosidicbondsα(1-6)glycosidicbonds(branching)
6 NUMBEROFMONOMERS
Fixed number of amino acids permolecule
Variable number of glucose permolecule
7 SOLUBILITYINWATER
SolubleGlobularinstructure
InsolubleLargesizeIntramolecularhydrogenbonds
5. Describefourdifferentfunctionsofproteins.[6m]
1 FUNCTION1–STRUCTURAL
(refertoPROTEINS–Q6–COLLAGEN)2 FUNCTION2–TRANSPORTATION
(refertoPROTEINS–Q6–HAEMOGLOBIN)(refertoCELLMEMBRANE–Q2–FACILITATEDDIFFUSION&ACTIVETRANSPORT)(refertoCELLMEMBRANE–Q3–RECEPTOR-MEDIATEDENDOCYTOSIS)
3 FUNCTION3–ENZYMATIC(refertoENZYMES–Q1–CATALYSIS)
4 FUNCTION4–REGULATION(refertoORGANISATIONANDCONTROLOFPROKARYOTIC&EUKARYOTICGENOMES)
5 FUNCTION5–DEFENSE&OFFENSE

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
6. Explainhowthestructureofcollagenandhaemoglobinarerelatedtotheirfunctions.[10m]Explainhow the generalised structure of a proteinmolecule canbe varied to suit differentfunctions.[6m]Describehowthestructureofanamedfibrousproteinisrelatedtoitsfunction.[6m]Explainwhatismeantbyprimary,secondary,tertiaryandquaternarystructuresofanamedprotein.[8m]1 COLLAGEN[1/2m]
Collagenisafibrous,structuralprotein*.Itisthemostabundantfibrousproteininthehumanbody.Itisanessentialcomponentofconnectivetissuesintendons,bones,skin,andteeth.
2 COLLAGEN–OVERALLSTRUCTURE[1/2m]Atropocollagenmolecule*consistsofthreehelicalpolypeptidechainswoundaroundeachotherinarope-likestructure,makingitthequaternarystructure.Cross-linkingofstaggeredmoleculesformacollagenfibril.Bundlesoffibrilsformcollagenfibres.
3 COLLAGEN–HELICALCHAIN[1m]Eachofthethreepolypeptidechainscontainabout1000aminoacidsthatformloosehelixes*.Thesequenceishighly regular, usually a repeating tripeptide* unit ofglycine-X-Y* whereX is usuallyproline*, andY isusuallyhydroxyproline*.Numeroushydrogenbonds*areformedwithineachhelicalpolypeptidechain*,specificallybetweenthehydroxylgroupsofdifferenthydroxyprolineresidues,stabilisingeachchain(note:differentfromaproteinwithα-helixsecondarystructureastheformerhasirregularhydrogenbonding,whilethelatterhashydrogenbondsbetweenCOandNHgroupsatevery4thpeptidebond).
4 COLLAGEN–TROPOCOLLAGEN[2m]Hydrogenbonds*arealso formedbetween the CO groups andNH groups in thedifferent residues*ofadjacent*polypeptidechains.Theyfurtherstabilisethetropocollagenandincreaseitsrigidityandtensilestrength.Theextensive*hydrogenbondingalsolimittheinteractionofthetropocollagenwithsurroundingwatermolecules,thusrenderingitinsoluble*inanaqueousenvironment.Almosteverythird*aminoacid isglycine*,thesmallest*aminoacid. Itfits intotherestrictedspace* inthecenterofthetriplehelixstructure.Thisallowsthetropocollagentoformaninelasticcompactcoil*.Thenumerousbulky*andrelatively inflexibleprolineandhydroxyproline*residuesconferrigidity* totheentiretropocollagenassembly.
5 COLLAGEN–FIBRE[2m]Each tropocollagen cross-links* with a neighbouring tropocollagen running parallel* to it. Cross-links areunusual typesofcovalent bonds* involving lysine* residuesofadjacent* tropocollagen, forming fibrils*.Thisarrangementgreatlyincreasesthetensilestrength*ofthefibrils.The staggered/overlapping* arrangement of the tropocollagen structures minimises points ofweaknesses*alongthelength*ofthefibrils,againincreasingthestrengthofthefibrils.Thefibrilsinturnunitetoformfibres*,whicharelargerbundlesofcollagenfibrils,increasingitsstrength.
6 HAEMOGLOBIN–STRUCTUREHaemoglobin isaglobularprotein*withametabolic* role. It isfoundinredbloodcellsof vertebrates,transportingoxygeninblood.
7 HAEMOGLOBIN–OVERALLSTRUCTUREIntheadult,ahaemoglobinmoleculehasaquaternarystructurewith4polypeptidesubunits,namely2α-globin subunits*and2β-globin subunits*.Eachsubunit ismadeupofapolypeptide component* calledglobin*andaprostheticcomponent*calledhaemgroup*.
8 HAEMOGLOBIN–POLYPEPTIDECOMPONENT(GLOBIN)Each subunit is arranged so that most of its hydrophilic* amino acid side chains of the polypeptidecomponentsareontheexternalsurface*,whilemostofitsnon-polar*,hydrophobic*aminoacidsidechainsareburiedintheinterior*,awayfromtheaqueoussurrounding.Thismakeshaemoglobinsolubleinwater*,allowingittobetransportedtocarryoxygenfromthelungstotissuesandviceversa.
9 HAEMOGLOBIN–PROSTHETICCOMPONENT(HAEMGROUP)Eachhaemgroupconsistsofaporyphyrinring*andaniron ion (Fe2+)*.TheFe2+ofthehaemgroupbindstemporarily/reversiblytooxygen*,so1haemoglobinmoleculecancarryupto4oxygenatatime,formingoxyhaemoglobin*.
10 HAEMOGLOBIN–SUBUNITSThe4 polypeptide subunits are held together by ionic bonds*,hydrophobic interactions*, andhydrogenbonds*,givingthemoleculeflexibility*instructuralchanges.Thisallowssubunitstomovewithrespect toeachother.Hence,cooperativebinding*ofoxygencanhappen,inwhichthebindingof1oxygenmoleculeto

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
1haemoglobinsubunitinduces*aconformationalchange*intheremaining3subunitssothattheiraffinity*foroxygenincreases.
IV.ENZYMES1. Describethemodeofactionofanenzymeintermsofspecificityandactivationenergy.[8m]
1 SPECIFICITY–INTERACTION/BINDINGAn enzyme has a specific* active site* which is complementary in conformation and charge* to thesubstrate.Basedonthelockandkeyhypothesis*,theenzymeisthelock*andthesubstrateisthekey*.Basedontheinduced fithypothesis*, thesubstrateinducesa change in the conformationoftheenzymeand therefore its active site, so that the active site is amore precise fit for the substrate for effectivecatalysis.Effectivecollisionsbetweentheenzymeandsubstrateformatemporaryenzyme-substratecomplex*.Theenzyme-substratecomplexisheldtogetherbecausethesubstratemoleculesformweakinteractions,suchashydrogenbonds, ionicbonds,andhydrophobicinteractions*,withthecontactresidues*oftheactivesite.
2 SPECIFICITY–CATALYSISAspecificreactioncatalysedbyanenzymeisdeterminedbytheRgroups*ofthecatalytic residues* intheactivesite,whichhelptocatalysetheconversionofthesubstratetotheproduct.Some enzymes may have absolute specificity, meaning it only catalyses a single specific reaction (e.g.maltaseonlycatalysestheglycosidicbondinmaltosetoyieldglucose,andnootherreaction).Some enzymes have group specificity, meaning it can attack one type of chemical bond in a variety ofsubstances(e.g.proteasedigeststhepeptidebondinvarioussubstrates).
3 ACTIVATIONENERGY–CATALYSISIn a chemical reaction, the reactantsmust absorb enough energy (Ea) to reach the transition state beforebondswithinthereactantscanbebrokenandthereactioncanproceed.During catalysis, the enzymeprovides an alternate pathway with lower activation energy barrier*. Byprovidinganalternatepathwayrequiringloweractivationenergy,moremoleculeswillhaveenergyequaltoorexceedingactivationenergy,allowingthereactiontoproceedatahigherrate.Activationenergycanbeloweredby:
§ Temporaryaligningorbindingofthesubstratesnexttoeachotherintheactivesite,increasingthechancesforthereactiontooccur
§ Distortingthesubstrates,strainingbondswhicharetobebroken,increasingthechancesforbondstobreak
§ Orientingthesubstratessuchthatitsbondsareexposedtoattack§ Providingafavourablemicroenvironmentsuitableforthereactiontooccur(e.g.awater-freezone
wherebyanon-polarorhydrophobicreactantmayreactmoreeasily)§ Presence of acidic/basicR groupsofaminoacidresidues in theactive sitewhich facilitatedirect
catalysis4 SPECIFICITY–RELEASE
Aftercatalysis,theproductsnolongerfittheactivesiteandarereleased.Theenzymeremainsunchangedafterthereactionandcanbeusedagain.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
2. Explainhowtemperatureaffectstherateofanenzyme-catalysedreaction.[7m]
(DIAGRAM:GRAPHOFRATEOFREACTIONAGAINSTTEMPERATURE)
1 INCREASEINTEMPERATUREBELOWOPTIMUMTEMPERATURE
Beginningatalowtemperature,anincreaseintemperatureresultsinanincreaseinkineticenergy*oftheenzymeandsubstratemolecules.Thisincreasesthefrequencyofeffectivecollisions*betweenthesubstrateand the enzymes’ active sites. This results in an increase in the rate of formation of enzyme-substratecomplexes*.Increasedkineticenergyalsoincreasesthenumberofmoleculeshavingsufficientenergytoovercometheactivationenergybarrier*toformtheproductsofthereaction.Therateofreactiondoublesforevery10°Cincreaseintemperature.
2 OPTIMUMTEMPERATURETherateof thereaction increaseswithtemperatureonlyuntil theoptimum temperature*of theenzymeisreached,atwhichtherateofenzymereactionproceedsatamaximumrate,withthehighestamountofproductsformed.Some enzymes have a higher optimum temperature. They tend to have a higher proportion of cysteineresidues and thusstrong covalent,disulfide bonds, ornumerous intramolecular interactions thatholdthetertiarystructureoftheenzymetogether.
3 INCREASEINTEMPERATUREABOVEOPTIMUMTEMPERATUREThe increase in kinetic energy at temperatures beyond the optimum temperature causes intramolecularvibrations to increase. Thisbreaks hydrogen bonds, ionic bonds and otherweak interactions, such ashydrophobicinteractions,thatdeterminetheconformation*oftheenzyme.Denaturation*occurswhenboththespecificconformationsoftheactivesiteandenzymearelost.Thesubstrate isno longer complementary* in conformation to the enzyme’s active site.Failureof thesubstratetofitintotheactivesiteresultsinfewerenzyme-substratecomplexes*beingformed,resultinginthedecreasingrateofreaction.
3. ExplainhowpHaffectstherateofanenzyme-catalysedreaction.[7m]
(DIAGRAM:GRAPHOFRATEOFREACTIONAGAINSTpH)
1 OPTIMUMpHEachenzymehasanoptimumpHatwhichitismostactive.TherateofreactionismaximumatthisoptimalpH.
2 DEVIATIONTherateofreactiondecreasesasthepHdeviatesfromtheoptimumpH.Excess [H+]or [OH-]ionsmayaffect the ionisationofRgroups/sidechains*ofaminoacids,neutralisingthechargesontheRgroups.ExcessH+resultsin–COO-groupsbecoming–COOH.ExcessOH-resultsin–NH3+groupsbecoming–NH2.
3 EFFECTSOFDEVIATIONpHdeviationmayaffectthestructuralaminoacidresidues.Ionicbonds*andhydrogenbonds*thatstabilisethespecificconformationoftheactivesite*maythusbedisrupted.Thisresultsinthelossofconformationoftheactivesite,resultinginthedenaturation*oftheenzyme.pHdeviationmayaffectthecontact/bindingaminoacidresiduesaswell.Whenthespecificcharge*oftheRgroups*ofthecontact/bindingresidues*intheactivesite*areneutralised,temporarybindingbetweenthe enzyme and substratemoleculesmay not occur. If no binding occurs, theenzyme-substrate complex*cannotbeformedandcatalysismaynottakeplace.pHdeviationmayalsoaffectthecatalyticaminoacidresidues.Whenthespecificcharge*oftheRgroups*of

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
thecatalyticresidues*intheactivesite*areneutralised,thecatalyticactivityoftheenzymemaybelost,andthecatalyticprocessitselfcannotbecarriedout.
4. Explainhowsubstrateconcentrationaffectstherateofanenzyme-catalysedreaction.[4m]
1 INCREASEINSUBSTRATECONCENTRATIONBELOWSATURATIONPOINTAtlowsubstrateconcentration,theactivesites*oftheenzymesarenotsaturated,andreadilyavailabletocatalysethereaction.Substrateconcentrationisthelimitingfactor.When substrate concentration increases, it increases the frequency of effective collisions* between thesubstrate and the enzymes’ active sites. This results in an increase in the rate of formation of enzyme-substratecomplexes*.Kmiswhenthereactionrateis½Vmax.
2 SATURATIONPOINTAthighersubstrateconcentration,thereissaturationoftheactivesites*atanyonetime.Thus,enzymeconcentrationisthelimitingfactor.Furtherincreaseinsubstrateconcentrationwillnotcausetherateofreactiontoincreasefurther.
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
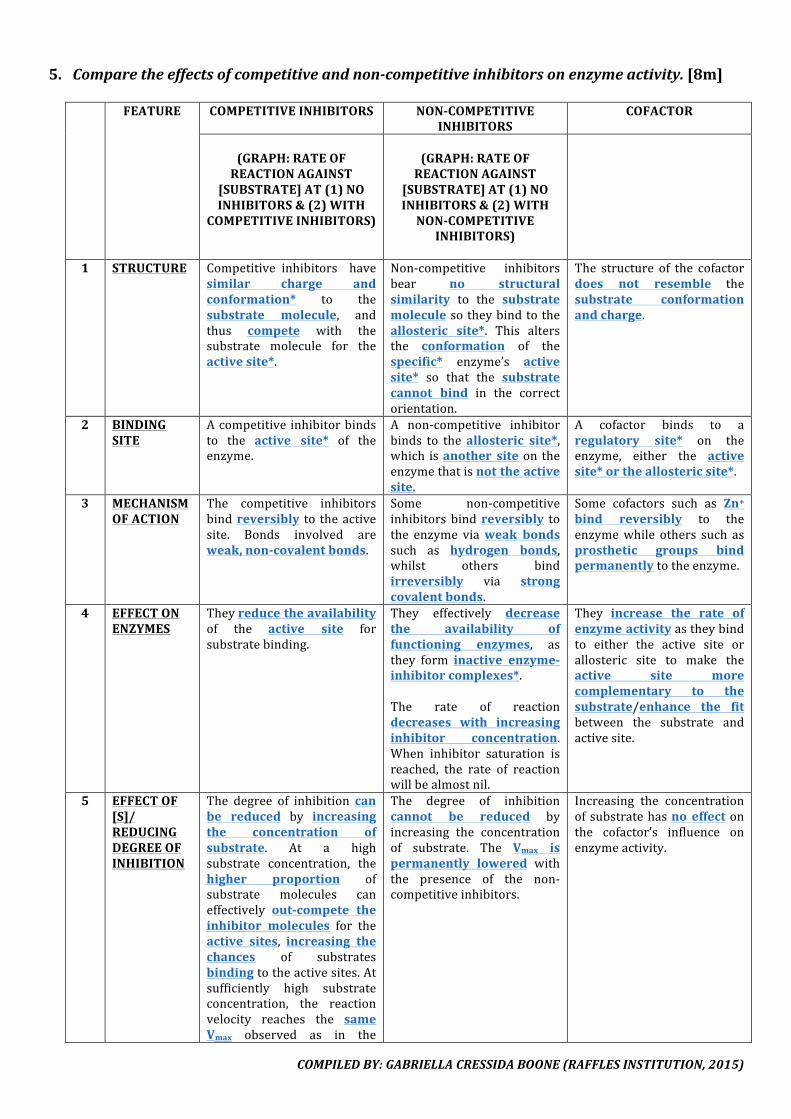
5. Comparetheeffectsofcompetitiveandnon-competitiveinhibitorsonenzymeactivity.[8m]
FEATURE COMPETITIVEINHIBITORS NON-COMPETITIVEINHIBITORS
COFACTOR
(GRAPH:RATEOFREACTIONAGAINST
[SUBSTRATE]AT(1)NOINHIBITORS&(2)WITH
COMPETITIVEINHIBITORS)
(GRAPH:RATEOFREACTIONAGAINST
[SUBSTRATE]AT(1)NOINHIBITORS&(2)WITHNON-COMPETITIVE
INHIBITORS)
1 STRUCTURE Competitive inhibitors havesimilar charge andconformation* to thesubstrate molecule, andthus compete with thesubstrate molecule for theactivesite*.
Non-competitive inhibitorsbear no structuralsimilarity to the substratemoleculesotheybindtotheallosteric site*. This altersthe conformation of thespecific* enzyme’s activesite* so that the substratecannot bind in the correctorientation.
The structure of the cofactordoes not resemble thesubstrate conformationandcharge.
2 BINDINGSITE
Acompetitiveinhibitorbindsto the active site* of theenzyme.
A non-competitive inhibitorbinds to theallosteric site*,which isanother siteon theenzymethatisnottheactivesite.
A cofactor binds to aregulatory site* on theenzyme, either the activesite*ortheallostericsite*.
3 MECHANISMOFACTION
The competitive inhibitorsbindreversibly to theactivesite. Bonds involved areweak,non-covalentbonds.
Some non-competitiveinhibitorsbindreversibly tothe enzyme viaweak bondssuch as hydrogen bonds,whilst others bindirreversibly via strongcovalentbonds.
Some cofactors such as Zn+bind reversibly to theenzymewhile others such asprosthetic groups bindpermanentlytotheenzyme.
4 EFFECTONENZYMES
Theyreducetheavailabilityof the active site forsubstratebinding.
They effectively decreasethe availability offunctioning enzymes, asthey form inactive enzyme-inhibitorcomplexes*.The rate of reactiondecreases with increasinginhibitor concentration.When inhibitor saturation isreached, the rate of reactionwillbealmostnil.
They increase the rate ofenzymeactivityastheybindto either the active site orallosteric site to make theactive site morecomplementary to thesubstrate/enhance the fitbetween the substrate andactivesite.
5 EFFECTOF[S]/REDUCINGDEGREEOFINHIBITION
The degree of inhibition canbe reduced by increasingthe concentration ofsubstrate. At a highsubstrate concentration, thehigher proportion ofsubstrate molecules caneffectively out-compete theinhibitor molecules for theactive sites, increasing thechances of substratesbindingtotheactivesites.Atsufficiently high substrateconcentration, the reactionvelocity reaches the sameVmax observed as in the
The degree of inhibitioncannot be reduced byincreasing the concentrationof substrate. The Vmax ispermanently lowered withthe presence of the non-competitiveinhibitors.
Increasing the concentrationofsubstratehasno effectonthe cofactor’s influence onenzymeactivity.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
absence of competitiveinhibitors.
6. Howcantwoaminoacids,‘A’and‘B’,thatarefarapartalongthepolypeptidechainsomehowendupnexttoeachotherinanactivesite?[4m]
1 STRUCTURES
The primary structure of the enzyme, which consists of the sequences and lengths of its amino acids,determinethesecondaryandtertiarystructures.
2 BONDS&INTERACTIONSThe polypeptide chain is folded into its secondary structure, either as and α-helix or β-pleated sheet,throughhydrogenbondingbetweentheCOandNHgroups,formingthepolypeptidebackbone.The secondary structure is further folded into its tertiary structure via hydrogen bonds, ionic bonds,hydrophobic interactionsanddisulfide linkages,whichareformedbetweenthedifferentRgroupsoftheaminoacids.
3 3DCONFORMATIONThefoldingofthepolypeptidechainthusresultsinaglobularstructuresuchthattheaminoacidresiduesarebroughtclosertogetherintheactivesite.
7. Describetheprocessofallostericregulation,withtheuseofanexample.[8m]
1 ALLOSTERICENZYMESAllosteric enzymesusuallyconsistoftwoormoresubunitswhereeachsubunithastheirownactive site*that binds to substrates, and allosteric site* that binds to activators* or inhibitors*. These sites are atdifferentlocationswithinthesamesubunit.
2 CONFORMATIONALSTATESAllosteric enzymes can be regulated by inhibitors* and activators*. A single inhibitor or activator issufficienttoinhibitoractivatetheactivityoftheenzyme.Theenzymescanthusexistintwoconformationalstates.Thebindingofanallostericinhibitorstabilisestheinactiveconformationoftheenzyme.Thebindingofanallostericactivator at theallosteric site or thebindingofasubstrate at theactive site,stabilisesthefunctionallyactiveconformation.
3 COOPERATIVITYThebindingof substrates in allosteric enzymesexhibitcooperativity*. So thebinding of a substrate to thefirst subunit changes the conformation of the other subunits such that it becomes easier to acceptsubsequent substrates. InarateVagainst[S]plot,anS-shaped sigmoid curveoftenindicatescooperativebindingofsubstratetoactivesite.
4 COOPERATIVITY–HAEMOGLOBINCooperativebindingcanbeobservedduringthebindingofoxygenmoleculestohaemoglobin.Thebinding of one oxygen molecule to one haemoglobin subunit induces a structural change in theremaining3subunits.Thiscausestheiraffinity foroxygentoincrease.This,theinitial“hesitant”loadingofthefirstoxygenmoleculeresultsintherapidloadingoftheother3oxygenmolecules.Conversely,whenonesubunitunloadsitsoxygenmolecule,theother3quicklyfollowsuit,astheconformationalchangealsolowerstheiraffinityforoxygen.
5 COOPERATIVITY–THREONINE&ISOLEUCINECooperativebindingcanalsobeobservedduringtheproductionofaminoacid isoleucine fromthreonine,specificallyoccurringasend-product inhibition*.Duringend-product inhibition*, ametabolicpathway isinhibitedbythebindingof theendproductofabiochemicalpathwaytoanenzyme thatactsearly in thepathway.Asisoleucineaccumulatesasanendproduct,itinhibitstheenzymethreoninedeaminaseinthefirststepof the reactionbybinding to its allosteric site*.Hence, the endproductalters the conformation* of theenzyme’s active site*. The substrate threonine can no longer bind to the active site in the correctorientation, so therate of reaction decreases. This prevents the cell fromwasting resources in producingexcessisoleucine.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
V.CELLSTRUCTURE
1 NUCLEOID>DNAtranscribedtorRNA(ribosomal)butnotmRNA(messenger)
2 NUCLEUS>DNAtranscription
3 RIBOSOMES>ComplexesofribosomalRNAandproteins>Consistsofsmallsubunitandlargesubunit>Prokaryotes:70s=small30s+large50s>Eukaryotes:80s=small40s+large60s>Ribosomeshaveamino-acyltRNAbindingsite,peptidyl-tRNAbindingsite,exitsite
4 MITOCHONDRIA CHLOROPLAST>ContainsDNAthatcanbetranscribedtomRNA>ContainsrRNAthatcantranslatemRNA>DNAiscircular>Largesurfaceareaofinnermembrane>ATPsynthesis>Only1outermembrane(total2membranes) >Chloroplastenvelope
>Doubleoutermembrane(total3membranes)>Intergranallamella>Granum>Thylakoid
1. Describethestructureandfunctionoftheendoplasmicreticulum.[6m]
1 ROUGHER–STRUCTURE
Theroughendoplasmicreticulum(rER)consistsofflattenedmembrane-boundsacscalledcisternae*.Itsmembraneiscontinuouswiththeoutermembraneofthenuclearenvelope.TherERhasribosomes*thatstudtheoutersurfaceofthemembrane,givingitagranularappearance.
2 ROUGHER–FUNCTIONTheribosomesontherERareinvolvedinthetranslation*ofthemRNAintopolypeptides.Theseproteinsareeithermeantforsecretionoutofthecellortargetedforinsertionintotheplasmamembrane.The polypeptide chains formed enter the cisternal space, where they fold into their specific 3Dconformationtoformproteinsandundergomodifications.Vesiclescarryingtheproteinsthenbudoff fromtherERtothecis faceof theGolgi apparatus forfurthermodifications.
3 SMOOTHER–STRUCTUREThe smooth endoplasmic reticulum (sER) comprises of interconnectedmembrane-bound tubules calledcisternae*withoutboundribosomes.
4 SMOOTHER–FUNCTIONThesERisinvolvedinlipid synthesis(e.g.steroidhormones,phospholipids),carbohydratemetabolismanddetoxificationofdrugsandpoisons.Vesicles carrying the lipids thenbud off from the sER to the cis face of theGolgi apparatus for furthermodifications.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
2. ExplainhowthestructuresoftheGolgiapparatusandlysosomearerelatedtotheirfunctionsinthecell.[10m]1 GOLGIAPPARATUS–STRUCTURE
TheGolgiapparatusconsistsofastackofflattenedmembrane-boundsacscalledcisternae*,togetherwithGolgivesicles.
2 GOLGIAPPARATUS–FUNCTIONVesicles* from the rough and smooth endoplasmic reticulum carrying proteins and lipid productsrespectivelywillfuse*withthecis faceoftheGolgiapparatus,releasingtheircontents intothecisternalspace. The products are further modified by glycosylation* (e.g. oligosaccharide on glycoproteins can bemodifiedtogiveavarietyofglycoproteinproducts).Golgi vesicles carrying the finished products bud off from the trans face of the Golgi apparatus. Thesemodifiedproductsaresortedintodifferenttypesofvesiclesandtransportedtovariouspartsofthecellorfor secretion (e.g. to the plasma membrane for insertion into the membrane or secretion throughexocytosis*).TheGolgiapparatusisalsoinvolvedintheformationoflysosomes*.Inplantcells,theGolgibodyisthesiteforsynthesisofpolysaccharidessuchaspectinandthentransportedinvesiclestothecellmembrane.
3 LYSOSOMES–STRUCTURELysosomesaremembrane-boundvesiclescontaininghydrolytic/digestiveenzymes.
4 LYSOSOMES–FUNCTIONLysosomesfuse*withendocyticvesiclestodigesttheingestedmaterialswithinthem.Lysosomesalso fuse* (asexocytic vesicles)with theplasmamembrane toreleasedigestive enzymesviaexocytosisforbreakdownofextracellularstructures.Lysosomes also fuse* with phagocytic vesicles to destroy bacteria. The products of digestion may beabsorbedandassimilatedintothecytoplasm.Unwantedproductsarereleasedthroughexocytosis*.Lysosomesalsofuse*withautophagicvesiclescontainingold/obsolete/unwantedorganellestobreakthemdownandtheorganicproductsofdigestionarerecycled.Whenthedigestiveenzymesinthelysosomearereleasedwithinthecell,andautolysis*,whichiscelldeath,occurs(e.g.breakdownofobsoletecellsduringmetamorphosisintadpoles).

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. Relate the structure of theGolgi apparatus to its function inmodificationand transport ofcellularproteins.[8m]1 CISTERNAE
Stacksof cisternawithcis and trans faces functiontoreceiveproteins fromthetransportvesiclesof theroughendoplasmicreticulumandleadproteinsthroughaprocessofmodificationandmaturationfromthecistothetransfaceoftheGolgiapparatusandintosecretoryvesicles.
2 HIGHSURFACEAREAOFMEMBRANESThe high surface area of membranes provided by the cisternae allow for a higher concentration ofembeddedenzymesandtheformationofmorevesicles.
3 ENZYMESEnzymes either in the lumen of the cisternae or embedded in the Golgi apparatus bilayer function toglycosylateorfurthermodifyproteins.ThefunctionalpartsoftheenzymesfacethelumenandproteinsaremodifiedwithinthelumenintheGolgiapparatus.
4 PHOSPHOLIPIDBILAYERMEMBRANEThe phospholipid bilayer membrane, which is similar to other membranes in components such as thephospholipids, cholesterol and some other membrane proteins, functions to be continuous with theendomembranesystem*.VesiclesfromtheendoplasmicreticulumcanthusfusetotheGolgiapparatus,andvesiclesfromtheGolgiapparatuscanformlysosomesorfusewiththeplasmamembraneforexocytosis.The bilayer membrane functions to create a suitable environment within the cisternal space for themodificationofproteins,byprovidingasuitablepHoroptimalconditionsforenzymesthatmodifyproteins.
5 PLASMAMEMBRANEPROTEINSPlasma membrane proteins embedded in themembrane of the cisternae function in cell recognition.These proteins face the cisternal space, so their position can be inverted to face the extracellularenvironmentwhensecretoryvesiclesfusewiththeplasmamembraneafterleavingtheGolgiapparatus.
6 SECRETORYVESICLESSecretoryvesiclesthatremaininthecytoplasmaslysosomesfunctiontodigestforeignsubstancessuchasbacteriaorfoodparticles.
7 LIPIDBILAYEROFSECRETORYVESICLESThe lipid bilayer that binds to the secretory vesicles from the Golgi apparatus function to allowincorporationintothecytoplasmicmembrane,enablingittoreleasecontentsviaexocytosis.
8 VESICLESATTACHEDTOCYTOSKELETONVesicles from the Golgi apparatus that attach to the cytoskeleton/microtubules function to direct themovementofsecretoryvesiclestothecellmembrane.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
VI.CELLMEMBRANE1. Explainthetermfluidmosaicmodel.[3m]
1 FLUID
Phospholipidsandembeddedproteinsarefreetomove,theyaredynamic.Phospholipidsmovewithintheirownlayer,transversely.
2 MOSAICVariousproteinsareembeddedinthephospholipidbilayer.Theyarescatteredandpositionedrandomlyacrossthebilayer.
2. Describetherolesofcholesterol,glycolipidsandglycoproteinsinthecellmembrane.[8m]
1 CHOLESTEROL
(refertoLIPIDS–Q2–CHOLESTEROL)2 GLYCOLIPIDS
(refertoLIPIDS–Q2–GLYCOLIPIDS)3 GLYCOPROTEINS
Glycoproteins areproteins with covalently attached sugar* molecules, often projected on the extracellularsideofthemembrane.Togetherwithglycolipids,theyformtheglycoalyx*(i.e.sugarcoat)locatedontheexteriorofthecell.Theyusuallyactasmarkers/recognitionsitesincell-cellrecognition*.
3. Describehowbulktransportoccurs.[7m]
1 INTRODUCTION
Bulktransportinvolvesmovementofsubstancesinlargequantitiesintoandoutofthecell.ATP*isneededfortherearrangementofmicrofilaments/microtubulesforthemovementofvesiclesandthecellmembrane.
2 ENDOCYTOSIS–PHAGOCYTOSISPhagocytosis* iswhen large, solid or insolubleparticlesare takenup into cells (e.g.bacteriaandviruses,foodparticles).Outwardextensionsoftheplasmamembrane,calledpseudopodia*,areformed,eventuallyenclosingsolidparticlesinavesicle/endosomewhenthepseudopodiafuse*.
3 ENDOCYTOSIS–PINOCYTOSISPinocytosis*bringsinliquidorsolubleparticlesintocells(e.g.proteins).A small area of the plasma membrane invaginates, eventually enclosing the soluble particles in avesicle/endosome.
4 ENDOCYTOSIS–RECEPTOR-MEDIATEDENDOCYTOSISReceptor-mediated*endocytosisoccurswhenspecificmoleculesorligandsbindtoreceptorproteinsonthemembrane.Theplasmamembraneinvaginates,enclosingtheligand-receptorcomplexinavesicle(e.g.ironboundtotransferrin,growthfactors,antibodies,lipoprotein).
5 EXOCYTOSISExocytosis* istheexportofsubstancesoutofacell(e.g.wastematerials,proteinsforexportsuchasinsulin,enzymes).TheGolgi/secretory vesicle* carrying substances for export,moves to theplasmamembrane and fuses*withit,releasingthecontentsintotheextracellularenvironment.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
4. Explainhowcompoundsaretakenupbythecell.[8m]
1 SIMPLEDIFFUSIONSimple diffusion consists of small, non-polar* moleculesmoving from a region of high concentration to aregionoflowconcentration,downaconcentrationgradient*andwithouttheuseofATP*.Thesemoleculescanpassthroughthehydrophobic*coreofthephospholipidbilayerreadily.
2 FACILITATEDDIFFUSIONFacilitateddiffusioninvolvesthepassivetransportofpolar,charged*moleculesorionsviasolute-specificproteinchannelsdownaconcentrationgradient,acrossthemembranewithouttheuseofATP*.Polar,charged*moleculesorionsareunabletodiffusethroughandrepelledbythehydrophobiccore*ofthemembrane.Hence, the interiorofchannelproteins*,madeupofpolar or chargedR groupsof amino acid residues,provideahydrophilicpore*formovementacrossthemembrane,whichisselectiveforspecificsolute*.Moreover, carrier proteins* undergo conformational change to bring the solute from one side of themembranetotheotherwhenaspecificsolute*bindstoit.Membranecanholdonto theseproteinsasnon-polar hydrocarbon chainsof thephospholipidbilayercanform hydrophobic interactions with the non-polar or hydrophobic R groups of amino acid residuesfoundontheexteriorsurfaceofthechannelprotein,facingthehydrophobiccore.Glucosemovesfromtheplasmaintoredbloodcellsviafacilitateddiffusion.
3 OSMOSISOsmosisisthediffusionofwatermoleculesfromaregionofhigherwaterpotential*toaregionoflowerwaterpotential*throughtheselective,semi-permeablemembrane*.Water,althoughpolar, issmall enough todiffusedirectlyacrossthemembranethroughtransientpores*thatarisewhenthephospholipidsareinlateralmotion.Watermoleculescanalsodiffuse throughaquaporin* proteins that facilitate theirdiffusionofwateracrossthemembranedowntheconcentrationgradient.
4 ACTIVETRANSPORTActive transport is needed when polar, charged* molecules or ions are to be transported against aconcentrationgradient*acrossthemembrane,withtheuseofATP*.Itinvolvestheuseofproteinpumpsanditisalsosolute-specific*.Inansodium-potassiumpump,foreachATPthatishydrolysed,3Na+ionsarepumpedoutwhile2K+ionsarepumpedin.Thisunbalancedchargetransferiscrucialinpropagatingactionpotentialsinnervecells.
5 ENDOCYTOSIS(refertoCELLMEMBRANE–Q3–BULKTRANSPORT)

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
VII.MITOSIS&MEIOSIS1. Outlinetheroleofcentriolesinmitosis.[2m]
1 POLARITYOFCELL
Apairofcentriolesateachpoleandtheirpositiondeterminesthepolarityofthecell.2 ORGANISINGSPINDLEFIBRES
Centrioles formpartof themicrotubule-organising centre, andorganise thesynthesis of spindle fibres.Thisleadstotheseparationofchromosomesafterthecentromeredivides.
2. DescribethedifferencesinthebehaviourofchromosomesinmitosisandmeiosisI.[5m]
PHASE MITOSIS MEIOSISI1 PROPHASE
Homologouschromosomesremainseparated.
Homologous chromosomes* pair upduringsynapsis*,formingbivalents*.
Noformationofchiasmata,nocrossingoverbetween non-sister chromatids ofhomologouschromosomes.
Chiasmata formation* may occur,crossing over* may occur between non-sister chromatids* of homologous pairsofchromosomes.
2 METAPHASE Chromosomes, each consisting of a pair ofsisterchromatids,alignindividually*alongthe metaphase plate*, forming a singlerow.
Homologous chromosomesalign in pairs*as tetrads along the metaphase plate*,formingtworows.
Independentassortmentdoesnotoccur. Independent assortment* of homologouspairs of chromosomes occurs along themetaphase plate, where randomarrangement of one pair of homologouschromosomes at the metaphase plate isindependent of the arrangement of theotherpairsofhomologues
3 ANAPHASE Centromeresdivide*. Centromeresdonotdivide.Sister chromatids* separate and eachmovetooppositepoles*.
Homologous pairs* of chromosomesseparate and each migrate to oppositepoles* (i.e. non-identical sister chromatidsremainjoinedandmovetothesamepole).
4 NO.OFCHROMOSOMES
Maintainsdiploidnumber Haploidnumber
5 NUCLEARDIVISIONS
1 2
6 NUCLEARENVELOPE
During prophase, disintegrates to allowchromosomestomovetometaphaseplateDuring telophase, reforms to enclosechromosomesinsidethenucleus
Duringprophase I,disintegrates to allowchromosomestomovetometaphaseplateDuring telophase II, reforms to enclosechromosomesinsidethenucleus

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. Describetheroleofmitosisinmaintaininggeneticstability.[6m]
1 REPLICATIONMitosisproducestwodaughtercellsandnucleiwiththesamenumberofchromosomesandsameallelesastheparentcell.During S phase* of interphase*, semi-conservative replication of DNA occurs to produce genetically-identicalsisterchromatids*.The double-stranded DNA unwinds and each strand acts as a template* for adding nucleotides bycomplementarybasepairing*.
2 ALIGNMENT&SEPARATIONDuringmetaphase*,chromosomesalignsinglyonthemetaphaseplate*.Duringanaphase*, thecentromere*dividesandthekinetochoremicrotubules fromeitherpoleattachedtoeachsisterchromatidshortens.Duringanaphase*,genetically-identical sister chromatids*areseparatedandmovedtooppositepoles*ensuringequaldistributionofchromosomes.Thenewdaughtercellshavethesamenumberandtypeofchromosomes*withthesamealleles*.(Note:shorteningthenlengtheningofspindlefibresduetoslidingactionduringthese2stages.)
3 ASEXUALREPRODUCTIONInasexual reproductionmitosis occurs toproducegenetically-identical daughter cellswhich areclones ofeachother.
4. Explaintheneedforareductiondivisioninmeiosis.[6m]
1 MEIOSISReductiondivisionistheproductionof4haploid*gametesfromadiploid*cell.
2 PURPOSE1–GENETICVARIATION–GAMETESDuring prophase I*, synapsis* occurs and homologous chromosomes* pair up, forming bivalents. Theformationofchiasmatabetweenthenon-sister*chromatidsofhomologouschromosomes, isfollowedbycrossingover*wherelinkagegroupsbreakandrejoinwitheachother,resultinginanexchangeofequalamountsofgeneticmaterial,formingnon-identicalsisterchromatidswithnewcombinationsofalleles.
3 PURPOSE2–GENETICVARIATION–GAMETESIndependent assortment* of chromosomes occur duringmetaphase I*. This leads to a great variety ofgameteswithdifferent combinations ofmaternal andpaternal chromosomes (e.g.223possible typesofgametesinahuman).
4 PURPOSE3–GENETICVARIATION–OFFSPRINGThehaploidnumberofagameteallowsfortherandomfusion*ofgametes,whichcombinesgeneticmaterialfrom two individuals. This gives rise to further variation in the offspring (e.g. 223 x 223 possible types ofoffspring).
5 PURPOSE4–GENETICSTABILITY–CHROMOSOMENUMBERUponfertilisation,arestorationofchromosomenumberoccurs(e.g.23chromosomesineggsandspermseach,whichfusetogive46chromosomesinthehumanoffspring).Inthisway,thechromosomenumber*ofthespeciesremainthesameaftermanygenerations.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
5. Describe how the behaviour of chromosomes during nuclear division result in geneticstabilityandgeneticvariation.[8m]
1 MITOSIS
Mitosis* achieves genetic stability, by resulting in 2 genetically identical* daughter cells, each with thesamenumberandtypeofchromosomes.During S phase* of interphase*, semi-conservative replication of DNA occurs to produce genetically-identicalsisterchromatids*.Duringmetaphase*,chromosomes*,eachwith2geneticallyidentical*sisterchromatids,lineupsinglyattheequatorialplane/metaphaseplate.During anaphase*, the centromeres divide*. Sister chromatids*, now individually known as daughterchromosomes*, are pulled to opposite poles* with the centromeres leading, by the shortening ofkinetochoremicrotubules*.
2 MEIOSISIDuring prophase I*, synapsis* occurs and homologous chromosomes* pair up, forming bivalents. Theformationofchiasmatabetweenthenon-sister*chromatidsofhomologouschromosomes, isfollowedbycrossingover*wherelinkagegroupsbreakandrejoinwitheachother,resultinginanexchangeofequalamountsofgeneticmaterial,formingnon-identicalsisterchromatidswithnewcombinationsofalleles.Duringmetaphase I*, independent assortment of homologous chromosomes*occurswhentherandomarrangement of one pair of homologous chromosomes at the metaphase plate is independent of thearrangementoftheotherpairsofhomologues.Duringanaphase I*, chromosomesofeach homologous pairseparate independently fromeachother tooppositepolesofthecell.Thereisthusanewcombinationofmaternalandpaternalchromosomesintheresultantdaughtercells.
3 MEIOSISIIDuring metaphase II*, there is also the random/independent arrangement of non-identical sister-chromatidsatthemetaphaseplate.DuringanaphaseII*,thereisalsothesubsequentseparationofnon-identicalsisterchromatids.Meiosis*resultsingameteswithnewcombinationsofallelesthatdifferfromparentalcombinationofalleles.Thisgivesrisetogeneticvariationwith2n*possiblecombinationsofchromosomesinagametewherenisthenumberofhomologouspairsofchromosomes.Duringfertilisation*,randomfusion*ofgametesresultsingreaternumberofgenotypiccombinationsofazygote,with22ndifferentoffspringgenotypesandpossiblephenotypes.
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
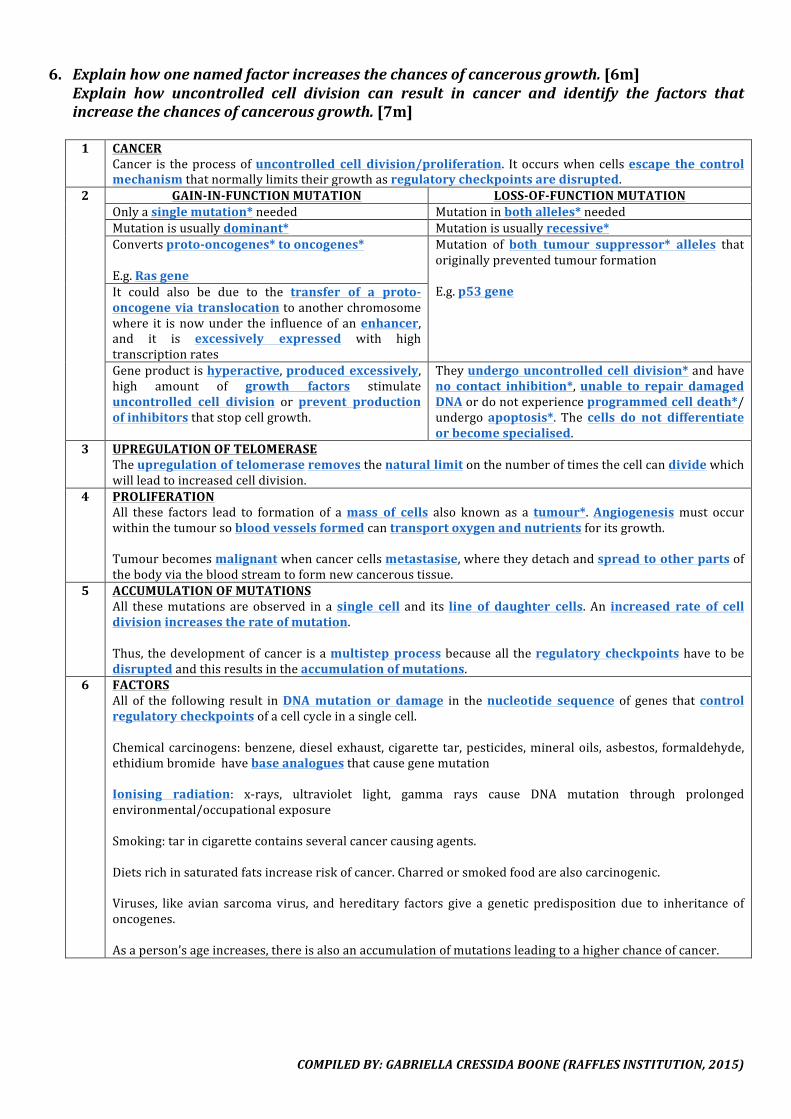
6. Explainhowonenamedfactorincreasesthechancesofcancerousgrowth.[6m]Explain how uncontrolled cell division can result in cancer and identify the factors thatincreasethechancesofcancerousgrowth.[7m]
1 CANCER
Cancer is theprocessofuncontrolled cell division/proliferation. Itoccurswhencellsescape the controlmechanismthatnormallylimitstheirgrowthasregulatorycheckpointsaredisrupted.
2 GAIN-IN-FUNCTIONMUTATION LOSS-OF-FUNCTIONMUTATIONOnlyasinglemutation*needed Mutationinbothalleles*neededMutationisusuallydominant* Mutationisusuallyrecessive*Convertsproto-oncogenes*tooncogenes*E.g.Rasgene
Mutation of both tumour suppressor* alleles thatoriginallypreventedtumourformationE.g.p53geneIt could also be due to the transfer of a proto-
oncogeneviatranslocationtoanotherchromosomewhere it isnowunder the influenceof anenhancer,and it is excessively expressed with hightranscriptionratesGeneproductishyperactive,producedexcessively,high amount of growth factors stimulateuncontrolled cell division or prevent productionofinhibitorsthatstopcellgrowth.
Theyundergouncontrolledcelldivision*andhaveno contact inhibition*,unable to repair damagedDNAordonotexperienceprogrammedcelldeath*/undergoapoptosis*. The cells do not differentiateorbecomespecialised.
3 UPREGULATIONOFTELOMERASETheupregulationoftelomeraseremovesthenaturallimitonthenumberoftimesthecellcandividewhichwillleadtoincreasedcelldivision.
4 PROLIFERATIONAll these factors lead to formation of amass of cells also known as a tumour*.Angiogenesismust occurwithinthetumoursobloodvesselsformedcantransportoxygenandnutrientsforitsgrowth.Tumourbecomesmalignantwhencancercellsmetastasise,wheretheydetachandspreadtootherpartsofthebodyviathebloodstreamtoformnewcanceroustissue.
5 ACCUMULATIONOFMUTATIONSAll thesemutations areobserved in asingle cell and its line of daughter cells. An increased rate of celldivisionincreasestherateofmutation.Thus, thedevelopmentofcancer isamultistepprocessbecauseall theregulatory checkpointshavetobedisruptedandthisresultsintheaccumulationofmutations.
6 FACTORSAll of the following result inDNAmutation or damage in thenucleotide sequence of genes thatcontrolregulatorycheckpointsofacellcycleinasinglecell.Chemicalcarcinogens:benzene,dieselexhaust, cigarette tar,pesticides,mineraloils,asbestos, formaldehyde,ethidiumbromidehavebaseanaloguesthatcausegenemutationIonising radiation: x-rays, ultraviolet light, gamma rays cause DNA mutation through prolongedenvironmental/occupationalexposureSmoking:tarincigarettecontainsseveralcancercausingagents.Dietsrichinsaturatedfatsincreaseriskofcancer.Charredorsmokedfoodarealsocarcinogenic.Viruses, like avian sarcoma virus, and hereditary factors give a genetic predisposition due to inheritance ofoncogenes.Asaperson’sageincreases,thereisalsoanaccumulationofmutationsleadingtoahigherchanceofcancer.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
7. Distinguishbetweengeneandchromosomalmutation.[6m]
POINTOFCOMPARISON
GENEMUTATION CHROMOSOMALMUTATION
1 TYPEOFCHANGE
Change in the sequence of bases in theDNAofonegene
Change in chromosomal structure usuallyinvolving several gene loci ornumber ofchromosomes
2 MECHANISM One (pointmutation) or several nucleotidebasesSubstitutionDeletionInsertionInversion
AchromosomalsegmentDeletionDuplicationInversionTranslocationEntirechromosome(s)duetonon-disjunction*Polyploidy(3n,4n,etc.)Aneuploidy(2n+1,2n–1)
3 CONSEQUENCE/EXAMPLE
Changeinbase/nucleotidesequenceChangeincodonChangeinsequenceofaminoacidinpolypeptideChangeinRgroups,changeintypeandlocationofbondsformedChangeinsecondaryandtertiarystructure,changein3DconformationofproteinBindingsiteofproteinnolongercomplementaryinchargeandconformationtotargetDNAsequences/substrates,cannotbindChangeinphenotype
E.g.Burkitt’slymphoma(cancer):Translocationofasegmentcontainingaproto-oncogenetoanotherchromosomewithregionofactivegenetranscription,upregulatinggeneexpression;becomesoncogenicgene.E.g.Downsyndrome/Trisomy21:Non-disjunction*ofchromosome21homologouspairduringmeioisisresultinginaneuploidy*
Silent:similar/nochangeinaminoacidMissense:differentaminoacidNonsense:prematurestopcodon
8. Explainhowgeneticvariationarisesinanaturalpopulation.[7m]
1 MEIOSIS(refertoMITOSIS&MEIOSIS–Q4–MEIOSISI&MEIOSISII)
2 GENEMUTATIONSGenemutations,throughsubstitution,deletionorinsertionofasinglebase,canchangethetripletcodesandhencepolypeptidetoo,resultinginphenotypicvariation.
3 CHROMOSOMALMUTATIONSPolyploidyiswhenmorethan2homologoussetsofchromosomesarepresent(e.g.triploid=3n,tetraploid=4n).Aneuploidyiswhenaparticularchromosomeisover-representedorunder-represented(e.g.Trisomy21).Deletioniswhenasegmentofachromosomeismissing(e.g.cri-du-chatdisease).Duplicationiswhenanextrasegmentofachromosomeispresent.Inversioniswhenachromosomesegmentisdetached,flippedaround180degreesandattachedtotherestofthe chromosome. Crossovers between the inversion and normal chromosomes lead to chromosomes withdeletions,deficienciesorabnormalstructure.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
Translocation is when a segment from one chromosome is detached and reattached to a differentchromosome,thereforelinkagerelationshipsarealtered.
VIII.DNA&GENOMICS1. Outlinethemainfeaturesofthegeneticcode.[4m]
1 INTRODUCTION
Thegeneticcodereferstotherelationshipbetweennucleotidebasesandaminoacids.Itdescribesthemannerinwhichaparticularnucleotidesequenceistranslatedintoitscorrespondingaminoacidsequence.
2 TRIPLETCODEThecodeisatripletcode,wherebyeachtripletcode,calledacodon*,codesforoneaminoacid*.
3 NON-OVERLAPPINGCODEThe code is non-overlapping*, only being read as successive groups of 3 nucleotides, whereby eachnucleotideinatripletcodeisonlyusedonce.
4 CONTINUOUSCODEThere are no nucleotides skipped between the codons, so the code is read as a continuous* sequence ofnucleotidebases.
5 UNIVERSALCODEThe code is universal*, in that the same triplet of nucleotides codes for the same amino acid in allorganisms(e.g.GGUwillalwayscodeforglycine,regardlessoftheorganism).
6 DEGENERATECODEThe code isdegenerate*, meaning that a givenamino acid*may be coded for bymore than one tripletcode/codon* (e.g.GAAandGAGbothspecifyglutamicacid). Inmostcases, thedifference in the tripletcodeliesinthethirdbase,whichwouldbeknownasthewobblebase.
7 START&STOPCODONSThecodeincludesstartandstopsequences.ThestartcodonisAUG*,whichcodesforaminoacidsmethionine*.ThiscodonsignalstheinitiationsiteoftranslationofthemRNA(downstreamofthestartcodon)intoasequenceofaminoacids.Thestop codonsareUAA/UAG/UGA*.Thesecodonsdonot code for anyaminoacid,asthere isnotRNAwithanticodoncomplementarytothese3codons.Theyactasstopsignalsfortheterminationofpolypeptidechainsynthesisduringtranslation.
2. Explaintheeffectsofsubstitutingasinglebaseinthegenecodingforaprotein.[5m]
1 NOEFFECT–DEGENERACY
AsinglebasesubstitutionwillnotresultinaframeshiftandwillonlychangeonecodoninthemRNA.Duetothedegeneracy*ofthegeneticcode,morethanonecodoncancodeforthesameaminoacid.Hence,thesamefunctionalproteinisproduced.
2 LITTLEEFFECT–SIMILARPROPERTIESAsinglebasesubstitutionwillnotresultinaframeshiftandwillonlychangeonecodoninthemRNA.Thenewcodonmaycodeforadifferentaminoacid,butitsRgroup*hassimilarpropertiestotheoriginalaminoacidbeingsubstituted.Hence,asimilarfunctionalproteinisproduced.
3 MAJOREFFECT–DIFFERENTPROPERTIESIfoneofthebasesintheDNAischanged,theresultingcodoninthemRNA*willchange.Thenewcodonwillcodeforadifferentaminoacid*withadifferentRgroup*.Thiswill result indifferent R group interactions ofvariousaminoacids, thusaltering the folding of thepolypeptidechain,andsubsequentlychangingthe3Dconformationandpropertiesoftheprotein.
4 MAJOREFFECT–PREMATURETERMINATIONIfoneofthebasesintheDNAischanged,theresultingcodoninthemRNA*willchange.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
ThenewcodonmaybeoneofthestopcodonsUAA/UAG/UGA*.Sincethesecodonsdonot code for any amino acid,as there isnotRNAwithanticodoncomplementarytothese3codons.theyresultinprematureterminationofpolypeptidechainsynthesisduringtranslation.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. OutlinethemainfeaturesofDNAreplication.[8m]
1 DNAREPLICATIONReplicationofDNAissemi-conservative*wheretheoriginalstrandsofthedoublehelixseparates*andeachstrand acts as a template* for the synthesis of twonew strands. This gives rise to two new/daughterDNAmolecules,eachconsistingofoneoriginalandonenewlysynthesisstrand.
2 STARTOFDNAREPLICATIONReplicationofDNAbeginsattheoriginofreplication*wheretheenzymehelicase*willbindtoandunzip*theDNAmoleculebybreakingthehydrogenbonds*betweencomplementarybasepairs.Separated strandsofDNA interactwith thesingle-strandedDNA binding proteins* so that itwillremainsingle-strandedandeachstrandcanserveasatemplate*forreplication.Topoisomerase*relievesoverwindingstrainaheadofreplicationforkbybreaking,swivelingandrejoiningDNAstrands.Note: To speed up the replication process, there aremultiple origins of replicationwheremanymore DNApolymerasescanworksimultaneously.Thisoccursineukaryotes.
3 SYNTHESISOFLEADINGSTRANDEnzymeprimase*catalysesthesynthesisofashortRNAprimer*,providingafree3’hydroxylend*whichisrequiredforDNApolymerase,whichhasaunidirectional function*,toinitiateDNAsynthesisandaddmorefreenucleotides.Complementary base pairing* occursbetween template* strandand free incomingdeoxyribonucleosidetriphosphate*.Adenine* forms 2 hydrogen bonds* with thymine*, while guanine* forms 3 hydrogen bonds* withcytosine*.DNA polymerase* catalyses the formationofphosphodiester bonds* linkingDNAnucleotides to form thesugarphosphatebackbone.ThenewDNAstrandissynthesizedinthe5’to3’direction*.Oneofthedaughterstrandsisknownastheleadingstrand*andissynthesisedcontinuouslytowardsthereplicationfork.
4 SYNTHESISOFLAGGINGSTRANDSince theparentalstrandsareanti-parallel*, the2newstrandsaresynthesized inoppositedirections,withrespecttothereplicationfork.Hence,theotherstrandisknownasthelaggingstrand*andissynthesiseddiscontinuouslyawayfromthereplicationfork,givingrisetoOkazakifragments*.The synthesis of each Okazaki fragment is also initiated by an RNA primer before the addition of DNAnucleotides.
5 ENDOFREPLICATIONRNAprimerswillthenbeexcised/removedandreplacedbyDNAbyanotherDNApolymerase.DNAligase*catalysestheformationofphosphodiesterbonds*betweentheOkazakifragments,joiningthe3’hydroxylendofeachnewDNAfragmenttothe5’phosphateendofthegrowingchain,sealingthenicks*.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
4. Outlinethemaineventsoftranscription.[7m]
1 INITIATIONRNA polymerase* will attach to the promoter* region of a gene on the DNAmolecule with thehelp ofgeneraltranscriptionfactors*.
2 ELONGATIONRNA polymerase will then unzip* the DNA double helix by breaking hydrogen bonds* betweencomplementarybasepairs*.OneoftheDNAstrandswillbeusedasthetemplate*strandreadinthe3’to5’*directionforthesynthesisofacomplementary*mRNAstrand.Free ribonucleotides*willbindbycomplementarybasepairing* to thenucleotidesontheDNAtemplatestrand.Adenine*onthetemplatestrandforms2hydrogenbonds*withuracil*,thymine*onthetemplatestrandforms2hydrogenbonds*withadenine*,cytosine*forms3hydrogenbonds*withguanine*.RNApolymerasewillcatalyse the formationof phosphodiester bonds*between free ribonucleotides toformthesugarphosphatebackbone.PolymerisationoftheribonucleotideswillresultintheformationofanewmRNAstrandthatissynthesizedinthe5’to3’direction*.
3 TERMINATIONRNApolymerasewilldissociatefromthetemplateDNAstrandwhenitreachestheterminationsequence*.
5. Describetheprocessoftranslation.[8m]
1 AMINO-ACYLtRNA
AspecificaminoacidandatRNAwithaspecificanticodonbindtotheirrespectivespecificactivesitesonanaminoacyl-tRNAsynthetase*,whicharerespectivelycomplementaryinconformationandcharge.ThisdoublespecificityensuresthereisacorrectmatchbetweenaminoacidandtRNAwithspecificanticodon.Theaminoacyl-tRNAsynthetasecatalysesthecovalentbondbetweenthespecificaminoacidandthespecificanticodonatthetRNA’s3’CCAstem.Thisformstheamino-acyltRNA.[Note:templateDNA(3’AAG5’)àsensemRNA(5’UUC3’)àtRNAanticodon(3’AAG5’)]
2 BINDINGOFRIBOSOMETOmRNAThesmallsubunitoftheribosomebindstothe5’end/AUGcodonofthemRNAwiththehelpofinitiationfactors.
3 INITIATIONThe initiator tRNA carrying methionine* pairs with the AUG*/start codon* on the mRNA throughcomplementarybasepairing*.3nucleotidescodefor1codonwhichdeterminesthespecificaminoaciditpairswith.Thelarge ribosomal subunitbindswiththeinitiator tRNA inthepeptidyl-tRNA (P) site*,completingtheribosome,formingthetranslation-initiationcomplex*.
4 ELONGATIONAsecondamino-acyl tRNAenterstheaminoacyl-tRNA(A)site*oftheribosomewhereitsanticodonbindsthroughcomplementarybasepairing*tothecodononthemRNA.Apeptidebond*isformedbetweenthetwoaminoacids,catalysedbypeptidyltransferase*.
5 TRANSLOCATIONTheribosomeshiftsonecodondowninthe5’to3’direction.ThefirsttRNAnowoccupiestheexit(E)site*whereitisreleasedandrecycled.
6 TERMINATIONTheemptyAsiteisnowreadytoreceivethenextaminoacyl-tRNA.Theprocessisrepeateduntilthestop/terminationcodon*(i.e.UAA/UAG/UGA*)enterstheAsite.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
ReleasefactorswillhydrolysethebondbetweenthepolypeptidechainandthetRNAatthePsite.6. DescribetheroleofmessengerRNAinproteinsynthesis.[4m]
1 TRANSCRIPTION
mRNA, a single stranded polynucleotide, formed from transcription* of a specific region of DNA acts as acarrierofgeneticcode.mRNAissynthesisedbycomplementarybasepairing*withDNAservingsasatemplate*whereadenine*basepairswiththymine*toform2hydrogenbondsandguanine*bondswithcytosine*toform3hydrogenbonds.mRNA being freely mobile, conveys the genetic message from the gene in the nucleus* out through thenuclear pore* to the ribosomes* in the cytoplasm* or on the rough endoplasmic reticulum* wheretranslationtakesplace.
2 TRANSLATIONmRNAcontainsaspecificsequenceoftripletbasesknownascodons*,andtheycodeforaspecificaminoacidsequenceofasinglepolypeptide*.Codons in themRNA interactwith anticodons*ofaspecifictRNA* carryingaspecificaminoacid(aminoacyltRNA)throughcomplementarybasepairing*.mRNAhasrecognitionsitessuchasAUGstartcodonthatallowsittobindtothesmallribosomalsubunitwiththehelpofinitiationfactors.
3 GENEEXPRESSIONREGULATIONGeneexpressioncanberegulatedeitherbyvaryingthefrequencyofmRNAsynthesis/transcriptionoritsrateofbreakdown.
7. DescribetheroleofribosomalRNAinproteinsynthesis.[3m]
1 STRUCTURErRNAassociateswithasetofproteinstoformribosomesandtheirsubunits.Itissingle-strandedandfoldsintomanyhairpinloopswithsomeareasofbasepairing.
2 BINDINGTOmRNArRNA is the main constituent of the interface between the large and small ribosomal subunits. The smallribosomalsubunitcanbindtothemRNA,viacomplementarybasepairingbetweenmRNAandtherRNAinthemRNAbindingsiteofthesmallribosomalsubunit.rRNA is themainconstituentof theP site (peptidyl-tRNAbinding site)andAsite (amino-acyl tRNAbindingsite).Hence,rRNAenablesthebindingofamino-acyltRNAstothePsiteandAsite.
3 PEPTIDEBONDrRNAonthe largeribosomalsubunitalsocatalyses the formationof thepeptide bond,betweentheaminogroupofthenewaminoacidintheAsiteandthecarboxylendofthegrowingpolypeptideinthePsite.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
8. Explain, using a named example, how a gene mutation may affect the phenotype of anorganism.[8m]1 SICKLECELL–MUTATION
Sicklecellanaemia*isaresultofapointsubstitutionmutation*.A single base substitution, where thymine* is replaced by adenine*, in the gene coding for the B-globinchain*, 6th triplet codon is changed from GAG to GUG, resulting in the amino acid glutamic acid* beingreplacedbyvaline*.
2 SICKLECELL–PHENOTYPEValine being hydrophobic* and non-polar, compared with glutamic acid which is hydrophilic* andcharged,resultsinachangeinpropertiesofthepolypeptidechain.Theprimary, secondary, and tertiary structures change because theway the polypeptide chain folds isaffectedbythechangeinRgroups*andbondsformed.Duetothischange in conformation*,normalhaemoglobinorHbAbecomessickle cellhaemoglobinorHbS*.
3 SICKLECELL–EFFECTSWhenoxygenlevelsarelowinblood,anunusualHbSconformationalchangecausesahydrophobicpatchtostickoutwhereHbSpolymerises*orcrystallises*toformrigidfibres*.Thiscausesredbloodcells*tochangefromacircularbiconcaveshapetoasickleshape.Sickle-shaped redblood cells aremore fragile, resulting in themhaving ashorter life span. This causes ashortage of red blood cells andpoor oxygen transport aswell as reduced ATP production, resulting inanaemia*.Sickle-shapedredbloodcells,beingpointedandelongated,mayalsogetlodgedinsmallbloodvesselsandthereforeinterferewithbloodcirculation.Thismayresultinorgandamageorlocalisedpain.It is a recessive* condition, requiring both alleles of the B-globin chains to be mutated in order for thesymptomstoappear.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
IX.VIRUSES1. Describethecharacteristicsofviruses.[7m]
1 STRUCTURE
Virusescanbeconsideredaslivingornon-living.Virusescanbeconsideredaslivingastheycontaingeneticmaterial*suchasviralRNAorDNA*butnotboth.Theycanbeconsideredasnon-livingastheyareacellular*.Theydonothaveorganelles,cytoplasmandarenotenclosedbyplasmamembranes.Instead,theyareenclosedbyaproteincoatcalledcapsid*thatismadeofsmallerunitsknownascapsomeres.Theyhavegeometricshapesandmayexistascrystallineformsoutsidethehostcells.Virusesareverysmallwithsizesrangingfrom10nmto300nm.
2 PROCESSESOutsidethehostcell,virusesdonotcarryoutmetabolicprocessessuchasrespiration.Once inside thehostcell,virusescanreproduce and replicatemanycopiesof theirviralgenomeusing thehostcellmachinery.Viruses use the host cell’s enzymes such as RNA polymerase for transcription and ribosome fortranslation toproduceviralproteinsanduseotherhost cell resources suchasnucleotides, amino acids,tRNAsandATPtoreplicate.Thus,virusesareknownasobligate parasites*as theyrequire thehostcell tocomplete their lifecycleandreproduce.
2. Explainhowavirusisreplicated.[8m]
1 ATTACHMENTViralreplicationbeginswiththevirusinvadingthehostcellandtakingoverthehost’smetabolicmachinery.Eachvirushasaspecifichostthatitinfects.Itrecognisesitshostcellviaspecifichostcellantigensorspecificreceptors(e.g.glycoproteinatcellsurfacemembrane).[Thehostrangeofavirusisdeterminedbytheproteinsonitssurface,andthoseonthehostcell’ssurface.]
2 PENETRATION&UNCOATINGThegenetic material, eitherDNAorRNA, is injected into the host cell, or the entire virusmayenter anddisassembleinsidethehostcelltofreethegeneticmaterial.
3 REPLICATIONThevirususesthehostcellmachinerytosynthesiseitsnucleicacidsuchasDNAvirus.TheDNAofthehostcellishydrolysedandthenucleotidesareused.ThehostDNApolymerase*isusedtoreplicateitsDNA.ThevirususesthehostcellRNApolymerase*totranscribe*itsgenesandhostribosome*totranslate*itsmRNA* to viral coat proteins and enzymes.Alsosuppliedby thehostcellarenucleotides, amino acids,tRNAsandATP.Itcontainsonlyafewgeneswhichcodefortheviralstructuralcomponentslikethecapsidproteinsandviralenzymesthatareinvolvedinthevirallifecycle.
4 MATURATIONOncealltheviralcomponentsaresynthesised,itwillselfassembletoformnewvirusparticlesorvirions.
5 RELEASE

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
Cellmembrane containingembedded viral glycoproteinssurround thevirus particle, and thenexit thecellviabudding*orexocytosis*orthroughlysisofthehostcell.
3. OutlinehowtheHIVandinfluenzavirusesareabletobypassthehumandefensemechanism.[3m]1 ANTIGENICDRIFT
For HIV, the fast/high rate of replication of the virus account for the high mutation rate in theribonucleotidesequencewhichcauseshighantigenicdrift/changeinantigenfrequency,withthefrequentconformationalchangeoftheviralglycoproteinsontheviralenvelope.
2 ANTIGENICSHIFTForinfluenza,antigenic shift,wherethereisgeneticreassortmentof theRNAsegmentsbetweentwoormore strains of influenza viruses that infect the same single host cell, results innew glycoproteins (i.e.haemagglutininandneuraminidase)ontheviralenvelope.
3 LACKOFPROOF-READINGThere is also a lack of proof-reading ability of RNA-dependent RNA polymerase in influenza, and reversetranscriptaseinHIV.
4 LACKOFRECOGNITIONBYANTIBODIESThus,theycannotbe inhibitedorrecognisedbyantibodiesthatwereoriginallytargetedagainstpreviousstrains.
4. ExplainhowanewcombinationofRNAsegmentscouldhaveariseninaninfluenzavirus,and
whatchangesmayhaveoccurredtogiveitgreaterabilitytoinfecthumans?[3m]1 ANTIGENICSHIFT
3strainsofvirusesinfect thesamecellofananimal.RNAsegmentsfromthe3differentorganismsareassortedtogetherinavirionthatformsanewinfluenzavirus.
2 MUTATIONAmutationtothegenecodingforHX,whichishaemagglutinin,sotheviruscannowbindtothereceptorofhumanrespiratoryepitheliumcelltoenteritbyendocytosis,thusinfectingthecell.AmutationtothegenecodingforNX,whichisneuraminidase,soitcannowfacilitatethereleaseofnewlyassembledvirusesfromthehostcellsviabudding/exocytosis,thusre-infectingothercells.
5. Describethestructuresoftwodifferenttypesofanimalviruses.[8m]
1 INFLUENZAVIRUS
Theinfluenzavirusisusuallysphericalorovoidinshape.ItcontainsasegmentedRNAgenomeof8*single-strandedRNA*,whichareallnegativesensestrands.(Minor point: each segment is also bound by 3 polymerase proteins which can combine to form anRNA-dependentRNApolymeraseforthetranscriptionandreplicationoftheviralgenome.)TheRNAgenomeisboundtoproteins,forminganucleoprotein.Thenucleoproteinissurroundedbyaproteincapsid*,madeupofsubunitscalledcapsomeres*,servingasaprotectivelayer.Thenucleoproteintogetherwiththecapsid,iscalledanucleocapsid*.Thenucleocapsidisinturnenclosedinaphospholipidenvelopederivedfromthehostcellplasmamembraneandisstuddedwithviralglycoproteins,haemagglutinin*andneuraminidase*.
2 HIVRETROVIRUSTheHIVvirusissphericalinshape.ThevirushasanRNAgenomecomprisingof2single-strandedRNA*,whicharebothpositivesense.TheRNAgenome,reversetranscriptase*,protease*andintegrase*areencapsulatedinaconicalproteincapsid*.Thenucleocapsid*isthenenclosedinaphospholipidenvelopederivedfromthehostcellplasmamembrane

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
andisstuddedwithviralglycoproteins,gp120*andgp41*ontheexteriorsurface.(Minor point: also at the envelope isa layer of matrix protein that lies along its interior to stabilise theattachmentofgp41tothemembrane.)
6. Describehowtheinfluenzavirusinfectslivingcellsandreproduces.[8m]1 INFLUENZAVIRUS–ATTACHMENT
Haemagglutinin*ontheviralsurfacebindstospecificsialicacidreceptor*foundonepithelialcellsliningtherespiratorytractofthehost.
2 INFLUENZAVIRUS–PENETRATION&UNCOATINGThevirusentersthehostcellbyreceptor-mediatedendocytosis*,wherethehostcellmembraneinvaginatesandpinchesoff,forminganendocyticvesicle.Endocyticvesiclefuseswithalysosome*andpHislowered/acidification*.Thistriggersthefusion*oftheviralenvelopewiththemembraneoftheendocyticvesicle.Thenucleocapsidisreleasedintothecytoplasm.Thecapsid layeristhendegradedbycellularenzymestoreleasetheviralRNAstrandswhichwillenterthenucleusofthehostcell.
3 INFLUENZAVIRUS–REPLICATION&MATURATIONWithin thenucleus, the negative senseRNA strands are transcribed* to their complementary strands,whicharepositivesense,byRNA-dependentRNApolymerase.Thepositivesensestrandsactastemplates*forthesynthesisofnewcopiesofviralRNAaswellastoserveasmRNA*.The mRNA* leaving the nucleus of host cells may be translated* in the cytoplasm or at the roughendoplasmicreticulum.At the rough endoplasmic reticulum, glycoproteins namely haemagglutinin* and neuraminidase*, areformedandpackedintovesiclesfortransportationtotheplasmamembranewheretheywillbeembedded.mRNA*translatedinthecytoplasmwillgiverisetocapsidproteinswhichwillassociatewiththeproteinsatthecytoplasmicsideofthecellmembrane.Assemblyofthevirioninvolvestheviralgenomeassociatingwithproteinstoformnucleoproteinswhichwillinteractwiththecapsidproteinsatthecellmembranetoinitiatethebuddingprocess.
4 INFLUENZAVIRUS–RELEASETheviruswillbudoff*fromthecell,acquiringitsenvelopefromthehostcellmembrane,nowembeddedwiththeviralglycoproteins.Thereleaseofvirus/buddingprocessisfacilitatedbyneuraminidase*whichcleavessialicacid*.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
7. DescribehowtheHIVvirusinfectslivingcellsandreproduces.[8m]
1 HIV–ATTACHMENTTheviralglycoprotein*gp120*recognisesandbindstotheCD4receptor*onthemembraneofthehostcell,suchasaThelpercellormacrophage.
2 HIV–PENETRATION&UNCOATINGTheviralenvelopefuses*withthehost cellmembranetoreleasethenucleocapsidintothecell, leavingtheenvelopebehind.Thecapsid*isdegradedbycellularenzymes.TheviralRNAanditsaccompanyingviralenzymesarereleasedintothecytoplasm.
3 HIV–REPLICATIONTheviral reverse transcriptase*will firstsynthesise a DNA strand complementary to the two single-strandedviralRNAstrandstoformRNA-DNAhybrids.TheRNAstrandswillbedegradedandasecondDNAstrandissynthesisedusingthefirstcDNAasatemplatetoformdouble-strandedviralDNA.The viral double-stranded DNA enters the nucleus of the host cell to be integrated into the host’schromosomebytheviralenzymeintegrase*.TheintegratedviralDNAisnowknownasaprovirus*,whichremainlatentforyearsuntilactivation.Uponactivation,theprovirus*istranscribedintomRNAwhichexitsthenucleus.TheseRNAcanalsoserveastheviralgenomesforthenewgenerationofviruses.Afterbeingtransportedtothecytoplasm,viralmRNAistranslated into viralpolyproteins*,usingthehostcell’s rough endoplasmic reticulum. Viral polyproteins are then cleaved into smaller functional proteins byprotease*.Glycoproteins,namelygp120 and gp41,aresynthesisedandpackaged intovesiclesandtransportedto thecellsurfacemembranewheretheyareembedded.Capsidproteinsarealsosynthesisedattheroughendoplasmicreticulum.
4 HIV–MATURATIONThecapsid* formsaroundnucleoproteins andviral enzymes, andassembles at the cellmembrane into anewvirus.
5 HIV–RELEASEThe new viruses will bud off/evaginate from the host cell acquiring the host cell membrane, nowembeddedwiththeviralglycoproteins,toinfectanothercell.
6 HIV–AIDSUpon infection by HIV, a patient’s immune system responds by destroying the CD4/T helper cell*population.The patient develops Acquired Immunodeficiency Syndrome (AIDS)when the immune system isunable tomountanimmuneresponseagainstcommoninfections/becomessusceptibletoopportunisticdiseases.
8. Suggestadvantagesofusingvirusesfortreatmentofbacteriainfectionoverantibiotics.[3m]
1 SPECIFIC
Viruses are very specific and will only attack a particular bacteria, while antibiotic kills different types ofbacteria.
2 QUANTITYOnlyasmallquantityofvirusisneededasviruscanreplicateonceitinfectsthebacteriumtoproducemoreviruses.Theystopreplicatingoncethespecificbacteriatheytargetaredestroyed.
3 RESISTANCEAsbacteriaevolveresistance,viruscanalsoevolvetoovercometheresistance.
4 SIDEEFFECTSTheyarelesslikelytocausesideeffectsorallergiestothepatient.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
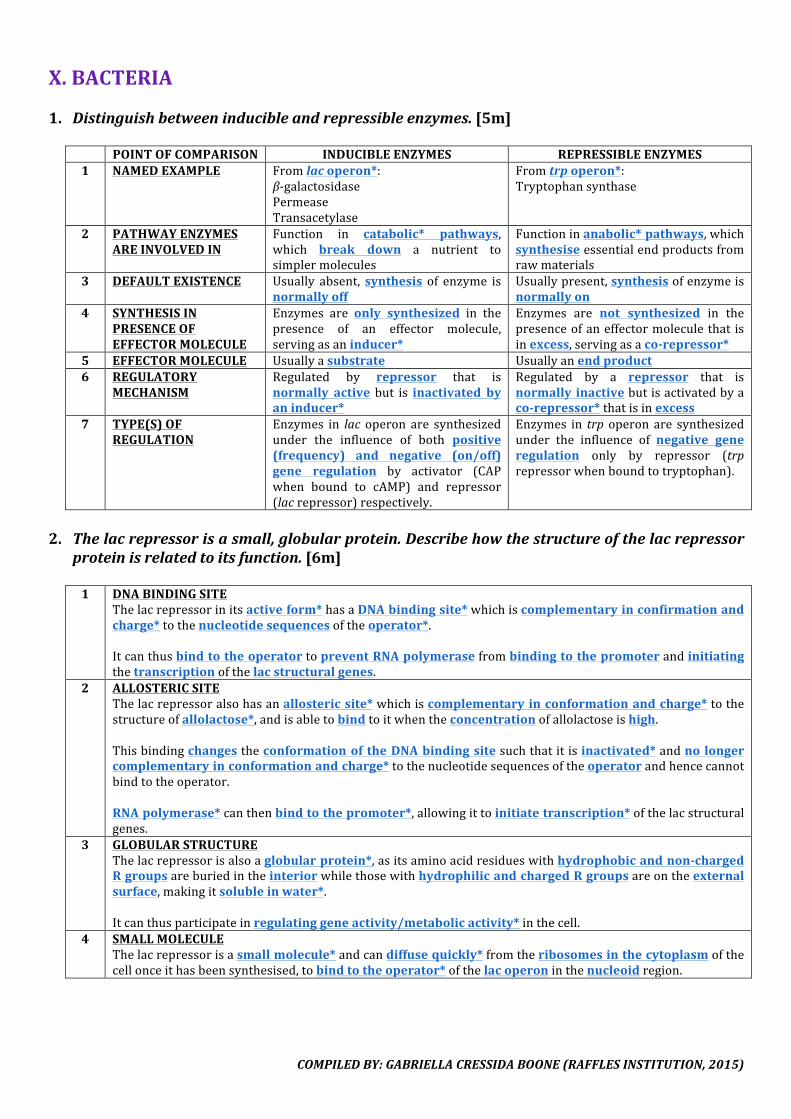
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
X.BACTERIA1. Distinguishbetweeninducibleandrepressibleenzymes.[5m]
POINTOFCOMPARISON INDUCIBLEENZYMES REPRESSIBLEENZYMES1 NAMEDEXAMPLE Fromlacoperon*:
β-galactosidasePermeaseTransacetylase
Fromtrpoperon*:Tryptophansynthase
2 PATHWAYENZYMESAREINVOLVEDIN
Function in catabolic* pathways,which break down a nutrient tosimplermolecules
Functioninanabolic*pathways,whichsynthesiseessentialendproductsfromrawmaterials
3 DEFAULTEXISTENCE Usually absent, synthesis of enzyme isnormallyoff
Usuallypresent,synthesisofenzymeisnormallyon
4 SYNTHESISINPRESENCEOFEFFECTORMOLECULE
Enzymes are only synthesized in thepresence of an effector molecule,servingasaninducer*
Enzymes are not synthesized in thepresenceofaneffectormoleculethat isinexcess,servingasaco-repressor*
5 EFFECTORMOLECULE Usuallyasubstrate Usuallyanendproduct6 REGULATORY
MECHANISMRegulated by repressor that isnormally active but is inactivated byaninducer*
Regulated by a repressor that isnormally inactivebutisactivatedbyaco-repressor*thatisinexcess
7 TYPE(S)OFREGULATION
Enzymes in lac operon are synthesizedunder the influence of both positive(frequency) and negative (on/off)gene regulation by activator (CAPwhen bound to cAMP) and repressor(lacrepressor)respectively.
Enzymes in trp operonare synthesizedunder the influence of negative generegulation only by repressor (trprepressorwhenboundtotryptophan).
2. Thelacrepressorisasmall,globularprotein.Describehowthestructureofthelacrepressor
proteinisrelatedtoitsfunction.[6m]1 DNABINDINGSITE
Thelacrepressorinitsactiveform*hasaDNAbindingsite*whichiscomplementaryinconfirmationandcharge*tothenucleotidesequencesoftheoperator*.ItcanthusbindtotheoperatortopreventRNApolymerasefrombindingtothepromoterandinitiatingthetranscriptionofthelacstructuralgenes.
2 ALLOSTERICSITEThelacrepressoralsohasanallostericsite*whichiscomplementaryinconformationandcharge*tothestructureofallolactose*,andisabletobindtoitwhentheconcentrationofallolactoseishigh.Thisbindingchangestheconformationof theDNAbindingsitesuchthatitisinactivated*andnolongercomplementaryinconformationandcharge*tothenucleotidesequencesoftheoperatorandhencecannotbindtotheoperator.RNApolymerase*canthenbindtothepromoter*,allowingittoinitiatetranscription*ofthelacstructuralgenes.
3 GLOBULARSTRUCTUREThelacrepressorisalsoaglobularprotein*,asitsaminoacidresidueswithhydrophobicandnon-chargedRgroupsareburiedintheinteriorwhilethosewithhydrophilicandchargedRgroupsareontheexternalsurface,makingitsolubleinwater*.Itcanthusparticipateinregulatinggeneactivity/metabolicactivity*inthecell.
4 SMALLMOLECULEThelacrepressorisasmallmolecule*andcandiffusequickly*fromtheribosomesinthecytoplasmofthecellonceithasbeensynthesised,tobindtotheoperator*ofthelacoperoninthenucleoidregion.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. DiscusshowdualcontrolofgeneexpressionisachievedinEscherichiacoli.[8m]Explainhowthepresenceoflactoseinthegrowthmediumswitchesonthelacoperon.[8m]
1 NEGATIVECONTROL–ABSENCEOFLACTOSE
Negativegeneregulationisbythelacrepressor*protein.Intheabsenceoflactose,noallolactoseisboundtotheallostericsite*onthelacrepressor.Thelacrepressorremainsinitsactiveconformation,anditbindstotheoperator*attheDNA-bindingsite*.RNA polymerase cannot bind to the promoter*, and the structural genes of the lac operon are nottranscribed.However,abasallevelofβ-galactosidaseandpermeaseisstillpresentwithinthecell.Thisisbecauserepressionof the lacoperonbytherepressoris‘leaky’.TheweakinteractionsbetweentherepressorandoperatormeansthattherepressormaydissociatefromtheoperatoroccasionallytoallowtheRNApolymeraseaccesstothelacoperon’spromoter.
2 NEGATIVECONTROL–PRESENCEOFLACTOSEAsmallnumberofpermeases*isthereforepresenttotransportthelactosecurrentlypresentinthegrowthmediumintothecell.Somelactosewillbeconvertedtoitsisomerallolactose*byβ-galactosidase*inthecell.Allolactosethenactsasaninducer*moleculewhichbindstotherepressorproteinatitsallostericsite.ThisalterstheconformationoftheDNA-bindingsiteoftherepressor.Therepressorisnowinactivatedandcannolongerrecogniseandbindtotheoperator.RNA polymerase* is now free to bind to the promoter* and can move downstream to transcribe* thestructuralgenes.
3 POSITIVECONTROL–ABSENCEOFGLUCOSEPositivegeneregulationisbytheCataboliteActivatorProtein(CAP)*.Whenglucoseisabsent,thereishighlevelofcAMP*.cAMP binds toallosteric site* ofCAP,activating CAPwhichbinds to the promoter* at theCAP-bindingsite*.This strengthens the affinity of RNA polymerase* to the promoter*, increasing the frequency oftranscriptionofthestructuralgenesinthelacoperon.
4 OVERALLOperonisnowturnedonwithincreasedsynthesisofβ-galactosidase*,permease*andtransacetylase*formetabolismoflactose.
5 DECREASEDEXPRESSIONInthepresenceofglucose,bacteriacanutilise/absorbglucosefromthesurroundingmedium.Itdoesnotneedtosynthesiseβ-galactosidase*tobreakdownlactose.Hence,decreasedexpressioninthepresenceofglucosehelpstoconserveresourcesthatcouldbedivertedforotherusestopreventwastageofresources.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
4. Statethedifferentpartsofaphage.[2m]1 T4BACTERIOPHAGE
>Capsidhead>Contractiletailsheath>Tailpins>Tailfibres:recognisesandbindstoreceptorsonsurfaceofbacterialcell>Baseplate:changesconformationinresponsetomoleculesreleasedfrombacterialcell,causingcontractionofcontratilesheathforinjectionofviralDNA>ViralDNA
5. DescribetheinfectionprocessoftheT4bacteriophageandthelambdaphage.[7m]
1 T4BACTERIOPHAGETheT4bacteriophagetailfibres*recogniseandbindtoreceptorsonsurfaceofbacterialcell.Itreleaseslysosyme*,anenzymethatdigeststhebacterialcellwall.Moleculesreleasedfrombacterialcellinduceachangeintheconformationofthebaseplate*.Thetailsheathcontractsandthruststhehollowcoretubethroughthebacterialcellwall.ViralDNAisinjectedintothebacterialhostcell.ViralDNAistranscribedandtranslatedtosynthesiseviralenzymesandphagecomponentsimmediatelyafterinfection.Enzymescodedbyphagegenometerminatethebacterium’smacromolecularsynthesis.Phageproteinsassembleandformthetailfibresandbaseplate,andrestofthephage.Itundergoesthelyticpathwaywherethelysozymebreaksdownthebacterialcellwallandthebacterialysesandthenewlyassembledbacteriophageinfectsnewbacterialcells.
2 LAMBDAPHAGEOntheotherhand,thelambdaphageundergoesalysogenicpathway.ThelambdaphageusesintegrasetointegrateitsDNAintothebacterialhostDNA,nowcalledaprophage*.ProphageDNAisreplicatedalongwithhostcellgenome.Thisallowscontinuousreplicationoftheprophagewithoutkillingthehostbacteria.Prophage gene expresses two repressor proteins which block the expression of other phage genesinvolvedinphagereplication,undergoingalatentstage.Afteraninductionevent,theprophageisexcisedfromthehostgenome.ViralDNAistranscribedandtranslatedtosynthesiseviralenzymesandphagecomponents.Lysozymebreaksdownthebacterialcellwallandthebacterialysesandthenewlyassembledbacteriophageinfectsnewbacterialcells.Lambdaphagecouldbepreventedfromenteringlysogeniccycleif:Thereisaloss-of-functionmutationintheviralgenecodingforintegrase,therebynotbeingabletointegratetheviralDNAasprophage.Thereisadeletionofnucleotidesatthesiteofphageintegrationonthebacterialchromosome.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
6. Outlinethedifferencesbetweenthelyticcycleandthelysogeniccycle.[4m]
LYTICCYCLE LYSOGENICCYCLE1 Norepressorproteinssynthesised Two repressor proteins systhesised, block the
expressionofotherphagegenes involvedinphagereplication,undergoingalatentstage
2 PhageDNAnotintegrated Phage DNA integrated by integrase into host cellgenome,formingprophage
3 PhageDNAreplicatedindependently ProphagereplicatedalongwithhostcellgenomeContinuousreplicationoflambdaphageanditsDNAwithoutkillingthehostcell
4 Enzymes coded for by phage DNA shuts down hostcellmacromoleculesynthesis
No production of enzymes, host cell macromoleculesynthesiscanproceed

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
7. Describe how DNA is transferred from one bacteria to another via transformation,transduction,andconjugation.[6m]
1 TRANSFORMATION
Transformation involves the taking up of foreign naked DNA fragments by a bacterial cell from thesurroundingmedium.Theyaresubsequentlyincorporatedintothebacterialgenomebycrossingoverathomologousregions*,resultinginhomologousrecombination.IftheforeignDNAcontainsadifferentallelethatisnowexpressed,thebacterialcellhasbeentransformed.
2 GENERALISEDTRANSDUCTION SPECIALISEDTRANSDUCTIONA phage* infects a bacterium*, injecting its viralgenomeintothehostcell.The bacterial DNA is degraded into smallfragments, a segment of which may be randomlypackaged into a capsid head* during thespontaneousassembly*ofnewviruses.Upon cell lysis, the defective phage* will infectanotherbacteriumandinjectbacterialDNAfromtheprevioushostcellintothenewbacterium.The foreign bacterial DNA can replace thehomologous region in the recipient cell’schromosome when crossing over at homologousregions* take place. This results in a change ingenotypeandthusphenotype.
A temperate phage* infects abacterium*, injectingitsviralgenomeintothehostcell.The viral DNA is integrated by integrase into thebacterialchromosomeformingaprophage*.Theprophagemaybeimproperlyexcisedtoincludeadjacent segments of bacterial DNA during aninduction*event.The excised bacterial DNA may be randomlypackaged into a capsid head* during thespontaneousassembly*ofnewviruses.Upon cell lysis, the defective phage will infectanotherbacteriumandinjectbacterialDNAfromtheprevioushostcellintothenewbacterium.The foreign bacterial DNA can replace thehomologous region in the recipient cell’schromosome when crossing over at homologousregions* take place. This results in a change ingenotypeandthusphenotype.
3 CONJUGATIONConjugationinvolvesthedirecttransferoftheFplasmid.Thesexpilus*ofanF+cellmakescontactwithanF-cell,andretractstobringtheF-cellcloser.Acytoplasmicmatingbridge*isformedbetweenthe2cells.Oneofthe2strandsoftheFplasmidintheF+cellisnickedbyanuclease,andtransferredfromtheF+celltotheF-cellthroughthematingbridgeviaarollingcirclemechanism.[The free 3’ end of the nick isextended by DNA polymerase for synthesisof acomplementary strand,usingtheotherintactstrandasatemplate.(Note:thereisnoend-replicationproblem)Thenewly synthesisedstranddisplaces thenickedstrand,whichisconcurrently transferredviathe5’endfromtheF+cell,acrossthematingbridge,intotherecipientF-cell.Anothernickoccurstoreleasetheoriginalstrandandendthereplicationofthenewlysynthesisedstrand.]Thetransferredsingle-strandedFplasmidcircularisesintheF-cellandisusedasatemplate*tosynthesiseacomplementarystrand*foradouble-strandedFplasmidDNA.ThisresultsintwoF+cells.Thisresultsinachangeingenotypeandthusphenotype.CONJUGATION–FPLASMIDItcontainsgenesthatcodeforproteinsnecessaryfortheformationofsexpilliandthecytoplasmicmatingbridge, allowing for conjugation to occur, and thus transfer of bacterial genes between bacteria,increasinggeneticvariationbetweenbacteria.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
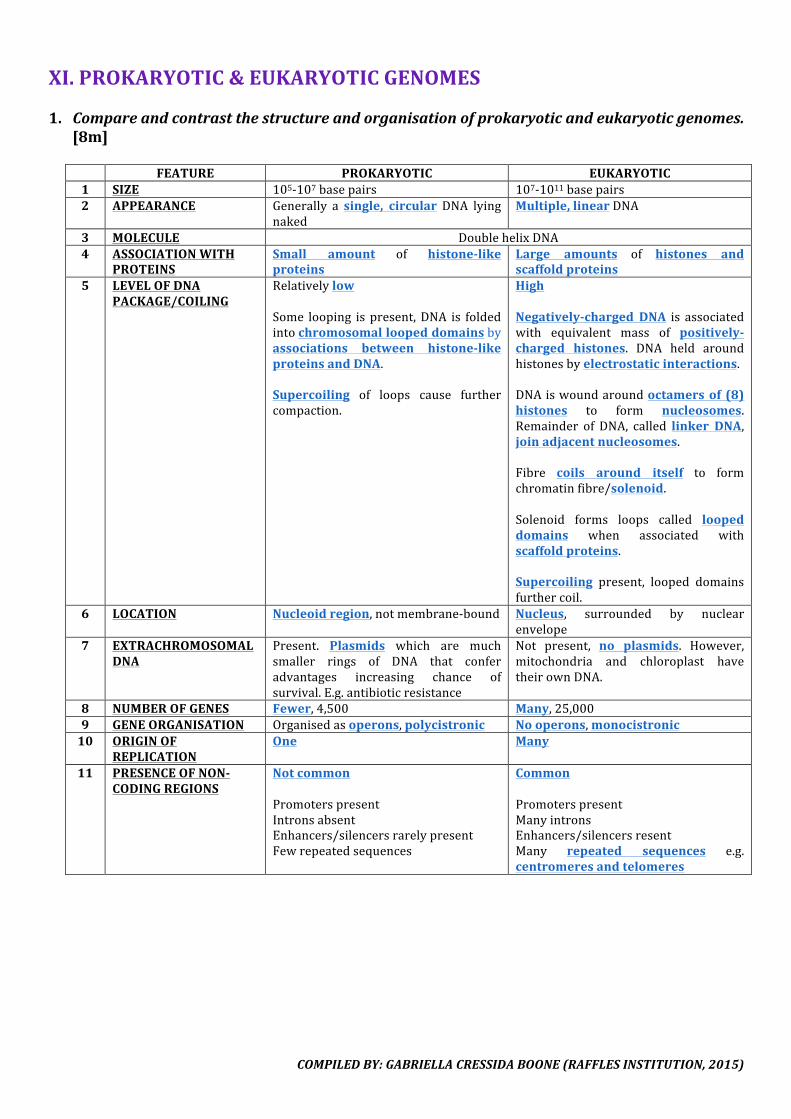
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
XI.PROKARYOTIC&EUKARYOTICGENOMES1. Compareandcontrastthestructureandorganisationofprokaryoticandeukaryoticgenomes.
[8m] FEATURE PROKARYOTIC EUKARYOTIC1 SIZE 105-107basepairs 107-1011basepairs2 APPEARANCE Generally a single, circular DNA lying
nakedMultiple,linearDNA
3 MOLECULE DoublehelixDNA4 ASSOCIATIONWITH
PROTEINSSmall amount of histone-likeproteins
Large amounts of histones andscaffoldproteins
5 LEVELOFDNAPACKAGE/COILING
RelativelylowSome looping is present,DNA is foldedintochromosomalloopeddomainsbyassociations between histone-likeproteinsandDNA.Supercoiling of loops cause furthercompaction.
HighNegatively-charged DNA is associatedwith equivalent mass of positively-charged histones. DNA held aroundhistonesbyelectrostaticinteractions.DNAiswoundaroundoctamersof (8)histones to form nucleosomes.Remainder of DNA, called linker DNA,joinadjacentnucleosomes.Fibre coils around itself to formchromatinfibre/solenoid.Solenoid forms loops called loopeddomains when associated withscaffoldproteins.Supercoiling present, looped domainsfurthercoil.
6 LOCATION Nucleoidregion,notmembrane-bound Nucleus, surrounded by nuclearenvelope
7 EXTRACHROMOSOMALDNA
Present. Plasmids which are muchsmaller rings of DNA that conferadvantages increasing chance ofsurvival.E.g.antibioticresistance
Not present, no plasmids. However,mitochondria and chloroplast havetheirownDNA.
8 NUMBEROFGENES Fewer,4,500 Many,25,0009 GENEORGANISATION Organisedasoperons,polycistronic Nooperons,monocistronic10 ORIGINOF
REPLICATIONOne Many
11 PRESENCEOFNON-CODINGREGIONS
NotcommonPromoterspresentIntronsabsentEnhancers/silencersrarelypresentFewrepeatedsequences
CommonPromoterspresentManyintronsEnhancers/silencersresentMany repeated sequences e.g.centromeresandtelomeres
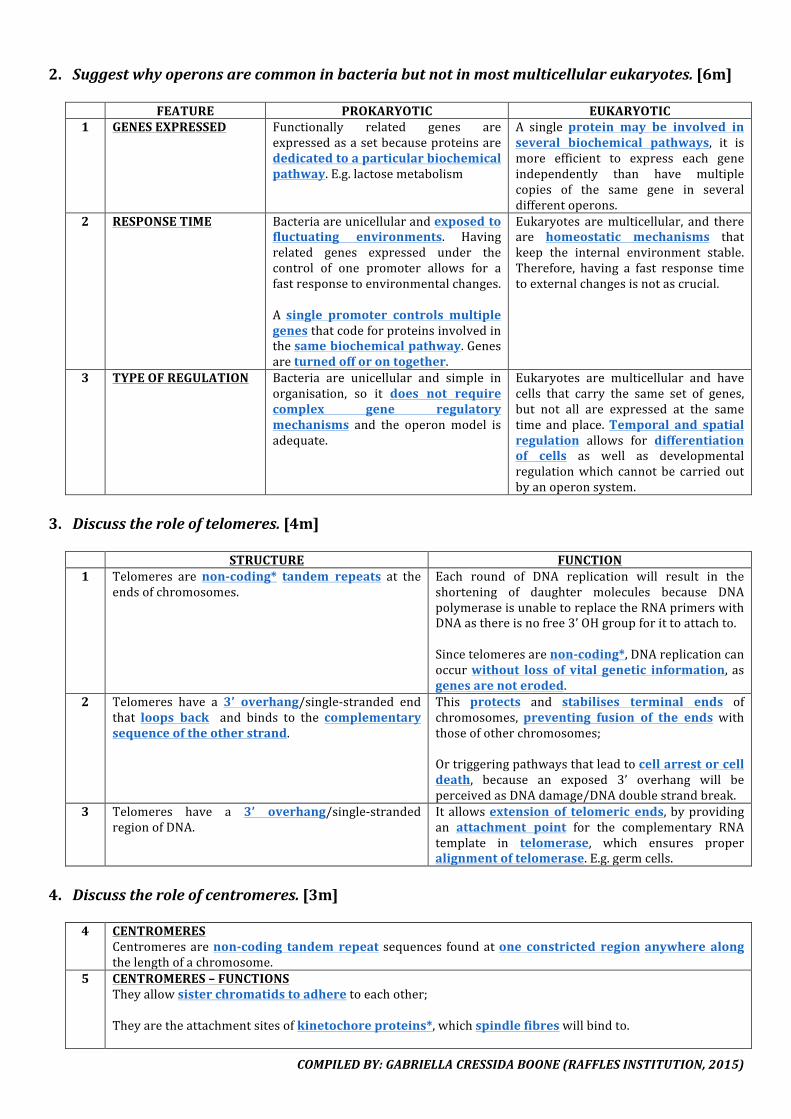
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
2. Suggestwhyoperonsarecommoninbacteriabutnotinmostmulticellulareukaryotes.[6m]
FEATURE PROKARYOTIC EUKARYOTIC1 GENESEXPRESSED Functionally related genes are
expressedasasetbecauseproteinsarededicatedtoaparticularbiochemicalpathway.E.g.lactosemetabolism
A single protein may be involved inseveral biochemical pathways, it ismore efficient to express each geneindependently than have multiplecopies of the same gene in severaldifferentoperons.
2 RESPONSETIME Bacteriaareunicellularandexposedtofluctuating environments. Havingrelated genes expressed under thecontrol of one promoter allows for afastresponsetoenvironmentalchanges.A single promoter controls multiplegenesthatcodeforproteinsinvolvedinthesamebiochemicalpathway.Genesareturnedofforontogether.
Eukaryotes aremulticellular, and thereare homeostatic mechanisms thatkeep the internal environment stable.Therefore, having a fast response timetoexternalchangesisnotascrucial.
3 TYPEOFREGULATION Bacteria are unicellular and simple inorganisation, so it does not requirecomplex gene regulatorymechanisms and the operon model isadequate.
Eukaryotes are multicellular and havecells that carry the same set of genes,but not all are expressed at the sametime andplace.Temporal and spatialregulation allows for differentiationof cells as well as developmentalregulationwhich cannot be carriedoutbyanoperonsystem.
3. Discusstheroleoftelomeres.[4m]
STRUCTURE FUNCTION1 Telomeres are non-coding* tandem repeats at the
endsofchromosomes.Each round of DNA replication will result in theshortening of daughter molecules because DNApolymeraseisunabletoreplacetheRNAprimerswithDNAasthereisnofree3’OHgroupforittoattachto.Sincetelomeresarenon-coding*,DNAreplicationcanoccurwithout loss of vital genetic information, asgenesarenoteroded.
2 Telomeres have a 3’ overhang/single-stranded endthat loops back and binds to the complementarysequenceoftheotherstrand.
This protects and stabilises terminal ends ofchromosomes, preventing fusion of the ends withthoseofotherchromosomes;Ortriggeringpathwaysthatleadtocellarrestorcelldeath, because an exposed 3’ overhang will beperceivedasDNAdamage/DNAdoublestrandbreak.
3 Telomeres have a 3’ overhang/single-strandedregionofDNA.
Itallowsextension of telomeric ends,byprovidingan attachment point for the complementary RNAtemplate in telomerase, which ensures properalignmentoftelomerase.E.g.germcells.
4. Discusstheroleofcentromeres.[3m]
4 CENTROMERESCentromeresarenon-coding tandem repeat sequences foundatone constricted regionanywhere alongthelengthofachromosome.
5 CENTROMERES–FUNCTIONSTheyallowsisterchromatidstoadheretoeachother;Theyaretheattachmentsitesofkinetochoreproteins*,whichspindlefibreswillbindto.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
So that sister chromatids or homologous chromosomes can align along the metaphase plate duringmetaphase,andbeseparatedtooppositepolesofacellduringanaphase.
5. Distinguish between structural and regulatory genes. Explain how the structure of specifictranscriptionfactorsislinkedtotheirrolesincontrolofeukaryoticgeneexpression.[8m]1 STRUCTURALGENES
AregionofDNAthatcodesforaproteinorRNAmoleculethat formspartofastructureorhasanenzymaticfunction.
2 REGULATORYGENESAregionofDNA that codes fora specificprotein that regulates theexpressionof thestructuralgenes/othergenes.
3 SPECIFICTRANSCRIPTIONFACTORSSpecifictranscriptionfactorsconsistofactivatorsandrepressors.The2bindingregionstheyhavearetheDNAbindingsiteandtheproteinbindingsite.Thereisthuscoordinatedgeneexpression,associatedwithgroupsofgeneswithrelatedfunctions.
ACTIVATORS REPRESSORSTheDNAbindingsiteiscomplementaryinconformationandchargeforspecificDNAsequences.Bindtoenhancers BindtosilencersTheproteinbindingsiteiscomplementaryinconformationandchargeforproteinstheyinteractwith.SpacerDNAbends, recruitsRNApolymerase/generaltranscriptionfactorstohelpassemblyoftranscriptioninitiationcomplexatthepromoter.Chromatin remodeling complexes and histoneacetylases loosen the coiling of negatively-chargedDNA around positively-charged histones or increaseaccessibilityofDNA.
SpacerDNAbends,preventsRNApolymerase/generaltranscription factors from assembly of transcriptioninitiationcomplexatthepromoter.Chromatin remodeling complexes, histonedeacetylases, DNAmethylation tighten the coiling ofnegatively-charged DNA around positively-chargedhistonesordecreaseaccessibilityofDNA.
6. Describetheeukaryoticprocessingofpre-mRNAintermsofintronsplicing,polyadenylation
and5’capping,andexplaintheirsignificance.[6m]
1 5’CAPPING7-methylguanosineisaddedtothe5’endofthepre-mRNA(beforeAUGcodon).ThishelpsthecelltorecognisemRNAfromotherRNAsforsplicing&polyadenylation.
2 INTRONSPLICINGSplicingofpre-mRNAisaprocesswhereby introns* are excised,andexons* are joined together to formmaturemRNA.Itiscarriedoutbyspliceosomes*.Intronsarenon-codingregionsthatareremoved.Exonsarecodingregionsforaminoacidsinaprotein.
3 POLYADENYLATIONPolyadenylationisaprocesswherebymultiple adenosinemonophosphatesareaddedtothe3’ endofthepre-mRNAbypoly-Apolymerase*,forminga3’poly-Atail*.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
7. Explain how gene expression can be controlled at the translational level and describe themechanismspresentinprokaryotesandeukaryotes.[6m]
FEATURE PROKARYOTES EUKARYOTES1 mRNAHALF-LIFE BothinvolveregulationofmRNAhalf-life
Longerhalf-lifeallowsfortranslationofmoreproteinsfrommRNAmRNA rapidly degraded, has short half lifetoallowforfastchangesingeneexpressionBinding of anti-sense mRNA can targetmRNA for degradation, further decreasingits half-life (note: use anti-sense mRNA topreventtranslationofundesiredproducts.)
mRNA half-life influenced by length of 3’poly-A tail, longer 3’ poly-A tail = morestable, longer half-life; when poly-A tailshortens to a critical length, 5’ cap isremoved andmRNA isdegradedbysiRNAthatbindstoit
2 AVAILABILITYOFINITIATIONFACTORS
Initiationfactorsrequiredforproperpositioningofsmallribosomalsubunittogetherwith initiator tRNA on the mRNA, and recruitment of large ribosomal subunit forassemblyoftranslationinitiationcomplex
3 BINDINGTOSMALLRIBOSOMALUNIT
Small ribosomal subunit recognises andbinds to the Shine Dalgarno region onmRNA for proper positioning of startcodon
Interactionbetween5’capand3’poly-Atailisrequiredfortranslationinitiation
Translational repressor protein or anti-sense mRNA binds to or near ShineDalgarno region on mRNA to preventbindingofsmallribosomalsubunit
Translational repressor protein binds tothe 5’ cap or 3’ poly-A tail to preventinteraction
TranslationcanbeblockedbypreventingthebindingofthesmallribosomalsubunittothemRNAforassemblyofthetranslationinitiationcomplex
8. Describetheprocessandsignificanceofgeneamplificationineukaryotes.[8m]
1 GENEAMPLIFICATION
Geneamplificationrefertothereplicationofaspecificgenemultipletimestocreatemorecopiesofthegene.Allothergeneshavenormalnumbersexceptforthegeneofinterestwhichexistsinhighcopynumber.Itisanexample of gene regulation, and consists ofupregulation only. This process isuncommon as othermethodsaltertherateoftranscriptionortranslation.
2 SIGNIFICANCEDuring some stages ofdevelopment of a eukaryotic organism, some gene products are required in largerquantities.Asinglecopyofagenecannotbe transcribed,norcan itsmRNAbetranslated,rapidly enough.Geneamplificationhelpstomeetthehighdemand.
FEATURE X.laevisSOUTHAFRICANCLAWEDTOAD
Drosophila
3 SIGNIFICANCE Oocytesneed to contain large numberof ribosomes so that rapid proteinsynthesis can occur after fertilizationforrapidgrowthofembryo.Precursor cells that differentiate intoeggsonlycontainfewerthan1,000copiesofrRNAgenecluster.Gene amplification causes number ofrRNA genes to increase from 0.2% ofgenome to 68% in just a few days,increasingsynthesisofribosomes.
Chorion (eggshell) protein needed inlargeamountstoenvelopeandprotectzygote. Protein is made specifically incells of the female insect’s reproductivetract.Amplifying chorion protein gene bymultiple rounds of DNA replicationmeets the demand for high levels ofchorionmRNA.
4 PROCESS/MECHANISM Genomic chromosomes first give rise toan extrachromosomal circular DNAcarrying therRNA genes.Fromthis firstring, many more copies of circular DNAare synthesized through the rolling-circlemodel.
Origin of replication in the DNAinitiates replication repeatedly andterminates randomly. This leavesreplication bubbles nested within largerreplicationbubbles.
5 LOCATIONOFREPLICATEDGENE
Millions of extra copies of ribosomalgenesexistascircularextrachromosomal
Forked structures due to replicationbubbles nestedwithin larger bubbles.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
DNA in the hundreds of nucleoli innucleoplasmofoocytes.
Inchromosomesinovariancells.
XII.GENETICS1. Casestudies
1 DEFINITIONSLocus=positionofagenealongachromosome,allelesofthesamegeneoccupysamelocusofchromosomesAllele = alternative form of a gene, alleles of same gene occupy same loci of homologous chromosomes,recessive/dominant/codominant
2 MONOHYBRIDRATIO=3:13 DIHYBRIDRATIO=9:3:3:14
MULTIPLEALLELES–BLOODGROUPIAIA=typeA IBIB=typeB IAIB=typeABIAIO=typeA IBIO=typeB IOIO=typeO
MULTIPLEALLELES–COATCOLOUR
C(grey)>cch(silverygrey)>ch(whitewithblackextremities)>c(albino)
5 CODOMINANCEMonohybrid,heterozygoteXheterozygote=1:2:1
6 LETHALALLELES(MANXCATS)Monohybrid,heterozygoteXheterozygote=2:1T=dominantalleleforshorttailst=recessiveallelefornormaltailsTT=lethalgenotype
T tT TT
(lethal)Tt
(shortenedtails)t Tt
(shortenedtails)tt
(normaltails)7 AUTOSOMALLINKAGE
Parentalphenotypicratio=1:1Recombinantphenotypicratio=1:1
𝑅𝑒𝑐𝑜𝑚𝑏𝑖𝑛𝑎𝑡𝑖𝑜𝑛 𝑓𝑟𝑒𝑞𝑢𝑒𝑛𝑐𝑦 = 𝑁𝑜. 𝑜𝑓 𝑜𝑓𝑓𝑠𝑝𝑟𝑖𝑛𝑔 𝑤𝑖𝑡ℎ 𝑟𝑒𝑐𝑜𝑚𝑏𝑖𝑛𝑎𝑛𝑡 𝑝ℎ𝑒𝑛𝑜𝑡𝑦𝑝𝑒𝑠
𝑇𝑜𝑡𝑎𝑙 𝑛𝑜. 𝑜𝑓 𝑜𝑓𝑓𝑠𝑝𝑟𝑖𝑛𝑔 × 100
SEXLINKAGE>Haemophilia,sex-linkedrecessive>Colourblindness,sex-linkedrecessive

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
>MalesinheritXchromosomecarryingdiseasedrecessiveallelefrommother

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
8 DISCONTINUOUSVARIATION>Notalwayscausedbyonegeneonly>Sometimesinfluencedbyenvironment
9 CONTINUOUSVARIATION>Alwayspolygenic/multiplegenesinvolved>Eachgenehassmalloveralleffect>Additiveeffectofeachgene>Phenotypeexpressionmaybeinfluencedbyenvironment
10 GENES&ENVIRONMENT–PHENYLKETONURIA>Autosomalrecessivegeneticdisorder>Deficiencyinenzymephenylalaninehydroxylase>Normal:phenylalaninemetabolisedtotyrosine>Diseased:phenylalanineaccumulatesandconvertedtotoxicsubstance>Effect:problemswithbraindevelopment,progressivementalretardationandseizures>Solution:dietlowinphenylalanineGENES&ENVIRONMENT–HIMALAYANRABBITS&TEMPERATURE>DeterminescoatcolourofHimalayanrabbits>Hightemperatureinhibitsenzymeactivitythatcodesforblackpigment>Mostlywhitebodywithblackears,nose,feetandtail>Iffuronthebackisshavedandicepackisfixed,blackhairwilldevelopGENES&ENVIRONMENT–NILECROCODILES&TEMPERATURE>Averagetemperatureduringmiddlethirdofincubationperioddeterminessex>Males:between31.7°Cand34.5°C>Females:outsideoftheaboverangeGENES&ENVIRONMENT–UVLIGHT>Melanocytesinskinaremelanin-producingcellslocatedinbottomlayersofskin’sepidermis>ExposureofUVlightincreasesmelaninproduction>DarkcolouredmelaninabsorbsandblocksUV-Blightfrompassingskinlayer>GreaterexposuretoUVlightresultsindarkercolouredskin
11 EPISTASIS>Formofgeneinteractioninwhichageneatonelocusaltersthephenotypicexpressionofanothergeneatasecondlocus>Considerbiochemicalpathway>Usuallygenesforlightercoloursneededasprecursorfordarkercolours>SohomozygousrecessivegenotypeforlightercoloursareepistaticovertheA/alocusofdarkercolours
CHICKENCOMBSHAPE>R_P_walnut>R_pprose>rrP_pea>rrppsingle
AGOUTIMICE>1genecontrolspigmentproduction,1genecontrolsbanding>Black,unbanded>Agouti,banded>White
12 PEAPLANTS>Purplevswhiteflowers>Roundvswrinkledseeds>Suitableforexperimentsbecause:(1)growfast,shortdurationofstudyformultiplegenerations+(2)manyoffspringsproducedinonegenerationtoobtainratio+(3)manyobservabletraitswithcontrastingforms+(4)easeinmanipulatingpollination(crossorself)

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
2. State the conditions of a chi-squared test. State what conclusionsmay be drawn from theprobability.[3m]1 CONDITIONS
>Measurementvariablesmustbeindependent>Valuesofvariablesmustbemutuallyexclusive>Samplesmustberandomlydrawnfromthepopulation
2 P>0.05Atalevelofsignificanceof5%,sincep>0.05,wedonotrejectthenullhypothesis,andwecanconcludethatthedifferencebetweenobservedandexpectedresultsisnotsignificant,andduetochancealone.
3 P<0.05At a level of significance of 5%, since p < 0.05,we reject the null hypothesis, andwe can conclude that thedifferencebetweenobservedandexpectedresultsissignificant,andnotduetochancealone.ThedifferencebetweentheobservedandexpectedratiosdonotfollowMendel’sLaw.There is no independent assortment of genes. The genes for phenotype 1 and phenotype 2 are probablylinked/locatedonthesamechromosome.ORTheobservedratioisclosetoX:Y,amodified9:3:3:1ratio,whichindicatesepistasis.
3. Tests
1 TESTCROSSTestcrosstodeterminegenotypeoforganismthatexhibitsdominanttrait(homozygousorheterozygous).Crossorganismwithanindividualexpressinghomozygousrecessivetrait.Ifoffspringphenotypicratioisalldominant,parentorganismishomozygousdominant.Ifoffspringphenotypicratiois1dominant:1recessive,parentorganismisheterozygous.
2 RECIPROCALCROSSReciprocalcrosstodetermineifcoatcolourisautosomalorsex-linked.MateAfemalewithBmaleandAmalewithBfemale.Ifoffspringofbothcrossesshowthesamephenotypicratio,coatcolourisautosomalinheritance.Ifoffspringofbothcrossesshowdifferentphenotypicratios,coatcolourissex-linked.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
XIII.RESPIRATION1. Explain how the mitochondrion is adapted to its role in carrying out aerobic respiration
efficiently.[6m]
1 DOUBLEMEMBRANE–COMPARTMENTALISATIONThrough compartmentalisation, the double membrane of the mitochondrion isolates the Krebs cycle andoxidativephosphorylation from reactions occurring in the cytoplasm, such as glycolysis. This improves theefficiency of the different reactions by concentrating the enzymes needed in a fixed compartment andprovidingoptimal conditionsrequiredforeachreactiontotakeplace.Forexample,theKrebs cycle inthemitochondrialmatrixandoxidativephosphorylationinthecristae.
2 DOUBLEMEMBRANE–SELECTIVEPERMEABILITYThe selective permeability of the mitochondrial double membrane to oxygen and pyruvate enables aconstantsupplyofthesesubstratestoenterthemitochondrion,andalsoforcarbondioxidetoleaveasaby-product.
3 INTERMEMBRANESPACE–ESTABLISHMENTOFPROTONGRADIENTIn between the double membrane is the enclosed intermembrane space (or thylakoid space inphotosynthesis). The phospholipid bilayer making up the membranes that enclose this region areimpermeabletochargedions.Theenclosedintermembranespacethereforeallowsthebuildupofprotonshere by the protonpumps found on the innermembrane, enabling the establishment of aproton gradientacosstheinnermembrane.ThisprotonmotiveforceisthencoupledtotheproductionofATPfromADPandinorganic phosphate byATP synthase. Therefore this compartmentalisation allows chemiosmosis to takeplace.
4 CRISTAE–INCREASEDSURFACEAREAThe inner mitochondrial membrane is highly folded into cristae, which increases the surface area forattachment ofmany electron carriers involved in the electron transport chain, enzymes such as ATPsynthase,andprotonpumps(andphotosystemsformaximumlightabsorptioninphotosynthesis).By being membrane-bound, these enzymes involved in oxidative phosphorylation can be attached in anorderedsequencethatfacilitatesthetransportofelectrons.Thisimprovesefficiencyofthereactions.(Note: electron carriers alternate between reduced and oxidised states as they accept and donateelectronsrespectively.Theyhelptofacilitatetheflowofelectronsintheelectron transport chain foundintheinnermitochondrialmembraneduringoxidativephosphorylation.)
2. OutlinethemainfeaturesoftheKrebsCycle.[8m]
1 LOCATION&CONDITIONKrebscycletakesplaceinthemitochondrialmatrixandwhenoxygenispresent.
2 2C+4C=6CAcetylcoenzymeA(2C)formedinthelinkreactioncombineswithoxaloacetate(4C)toformcitrate(6C).
3 6C-1C=5CCitrate(6C)isdecarboxylatedanddehydrogenatedtoforma-ketoglutarate (5C)andNADH.Thelossofacarbonresultsin1CO2moleculebeingreleasedasaby-product.
4 5C-1C=4C/REGENRATIONOFOXALOACETATERegeneration of oxaloacetate (4C) involves onemoredecarboxylation step and threedehydrogenationstepstoyield2NADH,1FADH2and1CO2.
5 NAD+&FADè NADH&FADH2HighenergyelectronsoriginallyfromtheglucosemoleculehavenowbeentransferredtotheelectroncarriersNAD+andFAD.NAD++2H++2e-èNADH+H+FAD+2H++2e-èFADH2
6 ADPè ATP1ATPisalsoproducedthroughsubstrate-levelphosphorylationduringthisregenerationprocess.
7 Cè CO2Allthecarbonoriginallyinglucoseislostascarbondioxide.
8 1MOLECULEOFGLUCOSEAltogether,1molecule of glucosewill yield 6 NADH, 2 FADH2, and 2 ATP through theKrebs cycle.Thecoenzymeswith their reducing powerwill next be transported to the electron transport chain in the innermitochondrialmembranewherethebulkofATPisgenerated.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. Explainhow the regenerationofNAD+andFADduringoxidativephosphorylationresults in
theproductionofATP.[4m]1 MOBILEELECTRONCARRIERS
NADHand FADH2 are reoxidisedwhen they transfer high energy electrons* toelectron carriers* of theelectrontransportchain*.
2 PROTONGRADIENTAstheelectronsarepasseddownelectroncarriers*ofdecreasingenergylevels,energyisreleasedthroughredoxreactionsalongtheelectrontransportchain.TheenergyisusedtoactivelypumpH+donatedbyNADHandFADH2fromthemitochondrialmatrixtotheinnermembranespace,togenerateaprotonmotiveforceacrosstheinnermitochondrialmembrane.AsH+diffusethroughATPsynthase*,downtheprotongradient*,backintothemitochondrialmatrix,ATPisproducedfromADPandinorganicphosphateviachemiosmosis*.
3 REGENERATIONReoxidationofNADHandFADH2allowedregenerationofNAD+andFAD*.Thisallowsthemtopickupmoreprotonsandelectronsfromglycolysis, linkreactionandKrebscycle*sothesereactionscancontinue.
4 ATPGENERATED1NADHà2.5ATP1FADH2à1.5ATP
4. ExplainthesmallyieldofATPfromanaerobicrespiration.[6m]
1 ANAEROBICVSAEROBICOnly2ATPisgeneratedperglucosemoleculeduringanaerobicrespirationascomparedto36-38ATPduringaerobicrespiration.
2 ABSENCEOFO2ASFINALELECTRONACCEPTORIn theabsence of O2, there isno final electron acceptor to remove electrons from the electron transportchain,thusdisrupting it,andoxidativephosphorylationcannotoccur.Hence,noATPcanbeproducedbythereactionsinthemitochondria,namelytheKrebscycleandoxidativephosphorylation,whichgeneratethebulkoftheATP.
3 GLYCOLYSISOnlyglycolysis cancontinuetoproducethe2ATPper glucosemolecule in thecytosolviasubstrate levelphosphorylation.Inaerobic respiration,complete oxidation of glucose produces carbon dioxide. Inanaerobic respiration,incompleteoxidationproducespyruvate.Asglycolysisproceeds,NADHaccumulatesbecauseitisunabletodonateitselectronstotheelectrontransportchain.Inorderforglycolysistocontinue,NADHhastobereoxidisedtoregenerateNAD+.
4 FERMENTATIONAsglycolysisalsoproducespyruvate(3C),pyruvateactsasthefinalelectronacceptor.
MAMMALIANCELLS YEASTCELLSUnder lactic acid fermentation, pyruvate is reducedto lactate by NADH, which is catalysed by lactatedehydrogenase.Lactate can be converted back to pyruvate by livercells.
Under alcohol fermentation, pyruvate is firstdecarboxylated to ethanal, and then reduced toethanolbyalcoholdehydrogenase.Ethanolcannotbeconvertedbacktopyruvate.
Ineitheroneoftheprocesses,NAD+isregeneratedsothatitmayreturntotakepartinglycolysistocontinueproducing2ATP.Inaerobicrespiration,oxidationofNADHyields34-36ATP.Inanerobicrespiration,noextraATPisproducedduringtheregenerationofNAD+.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
XIV.PHOTOSYNTHESIS1. Outlinethefeaturesofphotophosphorylation.[8m]
1 PHOTOSYSTEM
Aphotosystemconsistsofareactioncentresurroundedbymanylightharvestingcomplexes.Photosyntheticpigmentsboundtoproteinsarearrangedinthelightharvestingcomplexes.Areactioncentrecontains2specialchlorophyllamoleculesandaprimaryelectronacceptor.
2 EXCITATIONOFELECTRONSLight energy is absorbed* byaccessory pigments molecules such as chlorophylls a and b in the lightharvestingcomplexesofPhotosystems(PS)IandII*.Theelectronsinthesepigmentmoleculesbecomeexcitedandwhenreturningtotheirgroundstates,passon the released energy to the next pigment molecule and excite the electrons in them. This resonancetransfer of energyoccursuntil it reachesoneof the twospecial chlorophyll amolecules (P680 inPS II&P700inPSI)inthereactioncentre.Whenthespecialchlorophyllamoleculeabsorbstheenergy,anexcitedelectronisdisplaced(chlorophyllaàchlorophylla++e-),leavinganelectron“hole”inPSIandII.
3 NON-CYCLICPHOTOPHOSPHORYLATION CYCLICPHOTOPHOSPHORYLATION(P700inPSI)AtPSII,theelectron“hole”isfilledbythephotolysisof water*, donating a pair of electrons and yieldingoxygenasaby-product.Astheexcitedelectronsflowdownachainofelectroncarriers of the electron transport chain ofincreasing electronegativity/progressively lowerenergylevels,energylossthroughtheredoxreactioniscoupledtotheactivepumpingofH+ionsfromthestroma, across the thylakoid membrane, into thethylakoidspace.H+accumulatesinthethylakoidspacewhichservesas the H+ reservoir, generating a proton gradient*formedacrossthethylakoidmembrane.AstheH+diffusedowntheconcentrationgradientback into the stroma via the ATP synthasecomplex*, ADP is phosphorylated to ATP viachemiosmosis*.Innon-cyclicphotophosphorylation,NADPisthefinalelectron acceptor and is reduced to NADPH byNADPreductase.
Excited electrons are then passed on to the middlepart of the first electron transport chain whichlinks PS II to PS I, and they flow down a chain ofcarriers of increasing electronegativity/progressively lower energy levels. Energy lossthrough the redox reaction is coupled to the activepumping of H+ ions from the stroma, across thethylakoidmembrane,intothethylakoidspace.H+accumulatesinthethylakoidspacewhichservesas the H+ reservoir, generating a proton gradient*formedacrossthethylakoidmembrane.AstheH+diffusedowntheconcentrationgradientback into the stroma via the ATP synthasecomplex*, ADP is phosphorylated to ATP viachemiosmosis*.In cyclic photophosphorylation, the electron fillsbacktheelectron“hole”leftinPSI.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
2. Listhownon-cyclicphosphorylationdiffersfromcyclicphosphorylation.[6m]
FEATURE NON-CYCLICPHOTOPHOSPHORYLATION
CYCLICPHOTOPHOSPHORYLATION
1 PHOTOSYSTEM(S)INVOLVED
PSII&I PSIonly
2 ENDPRODUCT(S) ATP,NADPH&O2(by-product) ATPonly3 SOURCEOFELECTRONS H2O, splitting of water by an enzyme
releases H+ and e- (although directdonorisP680)
P700
4 PATHWAYOFELECTRONS
One direction: from water as theelectron donor, through PS II, to firstelectron transport chain, through PS I,to second electron transport chain, toNADP+
Cyclical: passing from PS I as theelectron donor, to first electrontransportchain,andbacktoPSI
5 FINALELECTRONACCEPTOR
NADP+ P700
6 MAINTENANCEOFHIGHH+CONCENTRATIONINTHYLAKOIDSPACE
Accumulation of H+ from splitting ofH2O in thylakoid space, and activetransportofH+ionsfromstromaintothylakoid space by electron transportchain proteins in the thylakoidmembrane
Active transport of H+ ions fromstroma into thylakoid space byelectrontransportchainproteinsinthethylakoidmembrane
3. ExplainbrieflyhowreducedNADPisformedinthelight-dependentstageandhowitisusedin
thelight-independentstage.[7m]
1 NON-CYCLICLIGHT-DEPENDENTSTAGEInthepresenceoflight,H2OissplitbyanenzymeandreleasesH+.H+ diffuses from the thylakoid space back into the stroma via ATP synthase* integrated in the thylakoidmembrane.Electronsarepasseddownanelectrontransportchainandaretransferred toNADPwhichcombineswithH+toformreducedNADPinthestroma.ThisreductioniscatalysedbyNADPreductase.NADPisthusthefinalelectronacceptor*inthenon-cycliclightdependentreaction*,formingNADPH.
2 LIGHT-INDEPENDENTSTAGE/CALVINCYCLEReducedNADP,NADPH,isusedintheCalvincycle*inthestromaofthechloroplast.6moleculesofNADPHfromthelight-dependentreactionserveasthereducingpowertoreduce*glyceratephosphate(GP)to3Cglyceraldehyde-3-phosphate(G3P).6moleculesofATPfromthelight-dependentreactionactasthesourceofenergyrequiredforthereduction.WhenGPisreducedtoG3P,NADP is regenerated tocarryoutitsroleasamobileelectroncarrierfromthelightdependentreaction.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
4. DescribethemainstagesoftheCalvincycle.[8m]
1 CARBONFIXATIONDuring the carbon fixation stage, CO2 is combined with 5C ribulose bisphosphate (RuBP)* to form anunstable6Cmolecule.TheenzymecatalysingthisreactionisRuBPcarboxylase(Rubisco).Theunstable6Cintermediateimmediatelysplitsinto2moleculesofglyceratephosphate(GP).
2 REDUCTIONBYNADPH6moleculesofNADPHfromthelight-dependentreactionserveasthereducingpowertoreduce*GPto3Cglyceraldehyde-3-phosphate(G3P).6moleculesofATPfromthelight-dependentreactionactasthesourceofenergyrequired.G3P is the first sugar formed inphotosynthesisandtheendproductof theCalvincycle.2moleculesofG3Pmaybeusedtoform1moleculeofglucose(hexosesugar).
3 REGENERATIONOFRuBP5moleculesofG3Pareusedtoregenerate*3RuBPsothatthecycleofcarbondioxidefixationcancontinue.Thisrequires3moleculesofATPfromthelight-dependentreaction.Thenetsynthesisof1moleculeofG3Prequires3CO2tobefixed.
5. Explaintheeffectofcarbondioxideandlightaslimitingfactorsontherateofphotosynthesis.
[6m]1 CARBONDIOXIDE
Carbon dioxide is a raw material for the Calvin cycle, needed for carbon fixation of CO2 to ribulosebisphosphate.Increaseintheconcentrationofcarbondioxidewillleadtoanincreaseintherateofphotosynthesis.However,attheatmosphericconcentrationofcarbondioxide,itbecomesalimitingfactor.
2 LIGHTLightplaysanimportantroleinthelightdependentstageofphotosynthesisasitexciteselectrons.Photosynthesisoccursundercertainwavelengthoflight–red(about700nm)andblue(about470nm).Lightcompensationisthepointwherephotosyntheticrateisequaltotherespirationrate.Lightcompensationpoints are unique to plants. For plants with higher light compensation points, higher light intensities arerequired.Whenlightintensityisnolongerthelimitingfactor,lightsaturationisachieved.However,lightisrarelythelimitingfactorduringdaylighthours.
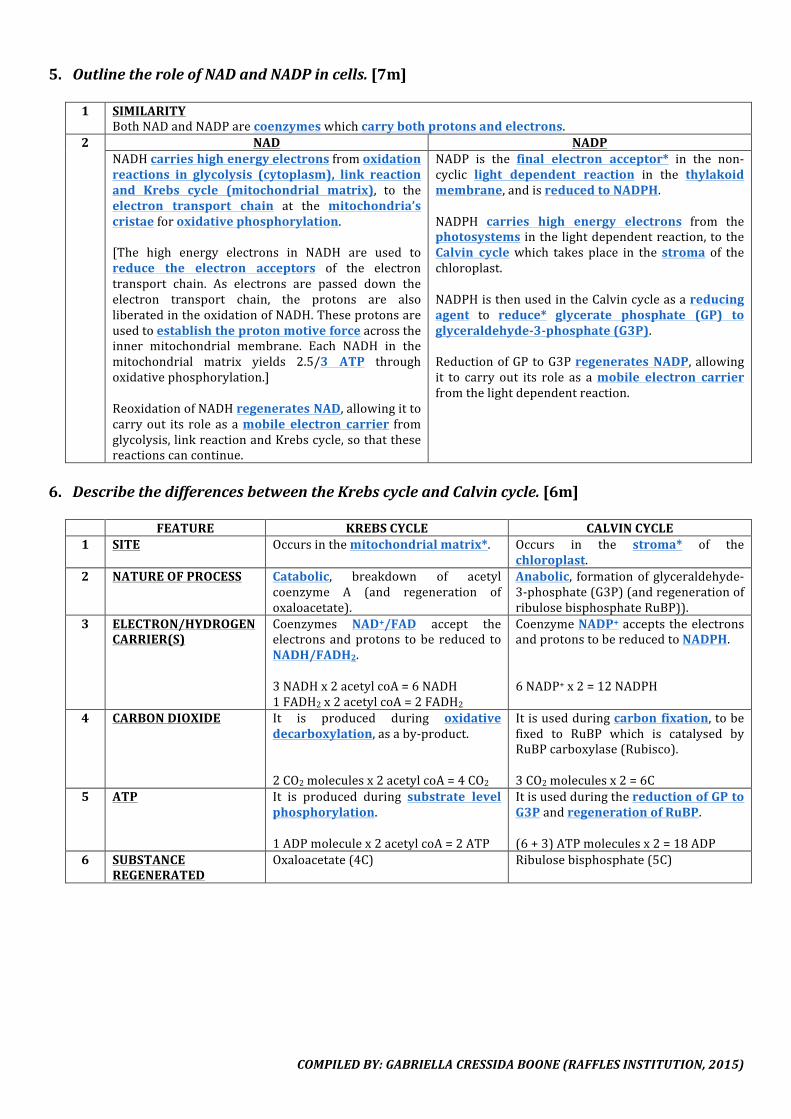
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
5. OutlinetheroleofNADandNADPincells.[7m]
1 SIMILARITYBothNADandNADParecoenzymeswhichcarrybothprotonsandelectrons.
2 NAD NADPNADHcarrieshighenergyelectronsfromoxidationreactions in glycolysis (cytoplasm), link reactionand Krebs cycle (mitochondrial matrix), to theelectron transport chain at the mitochondria’scristaeforoxidativephosphorylation.[The high energy electrons in NADH are used toreduce the electron acceptors of the electrontransport chain. As electrons are passed down theelectron transport chain, the protons are alsoliberatedintheoxidationofNADH.Theseprotonsareusedtoestablishtheprotonmotiveforceacrosstheinner mitochondrial membrane. Each NADH in themitochondrial matrix yields 2.5/3 ATP throughoxidativephosphorylation.]ReoxidationofNADHregeneratesNAD,allowingittocarryout its roleasamobile electron carrier fromglycolysis,linkreactionandKrebscycle,sothatthesereactionscancontinue.
NADP is the final electron acceptor* in the non-cyclic light dependent reaction in the thylakoidmembrane,andisreducedtoNADPH.NADPH carries high energy electrons from thephotosystems inthelightdependentreaction,totheCalvin cyclewhich takes place in thestroma of thechloroplast.NADPHisthenusedintheCalvincycleasareducingagent to reduce* glycerate phosphate (GP) toglyceraldehyde-3-phosphate(G3P).ReductionofGPtoG3PregeneratesNADP,allowingit to carry out its role as amobile electron carrierfromthelightdependentreaction.
6. DescribethedifferencesbetweentheKrebscycleandCalvincycle.[6m]
FEATURE KREBSCYCLE CALVINCYCLE1 SITE Occursinthemitochondrialmatrix*. Occurs in the stroma* of the
chloroplast.2 NATUREOFPROCESS Catabolic, breakdown of acetyl
coenzyme A (and regeneration ofoxaloacetate).
Anabolic, formationofglyceraldehyde-3-phosphate(G3P)(andregenerationofribulosebisphosphateRuBP)).
3 ELECTRON/HYDROGENCARRIER(S)
Coenzymes NAD+/FAD accept theelectronsandprotons tobe reduced toNADH/FADH2.3NADHx2acetylcoA=6NADH1FADH2x2acetylcoA=2FADH2
CoenzymeNADP+acceptstheelectronsandprotonstobereducedtoNADPH.6NADP+x2=12NADPH
4 CARBONDIOXIDE It is produced during oxidativedecarboxylation,asaby-product.2CO2moleculesx2acetylcoA=4CO2
Itisusedduringcarbon fixation,tobefixed to RuBP which is catalysed byRuBPcarboxylase(Rubisco).3CO2moleculesx2=6C
5 ATP It is produced during substrate levelphosphorylation.1ADPmoleculex2acetylcoA=2ATP
ItisusedduringthereductionofGPtoG3PandregenerationofRuBP.(6+3)ATPmoleculesx2=18ADP
6 SUBSTANCEREGENERATED
Oxaloacetate(4C) Ribulosebisphosphate(5C)
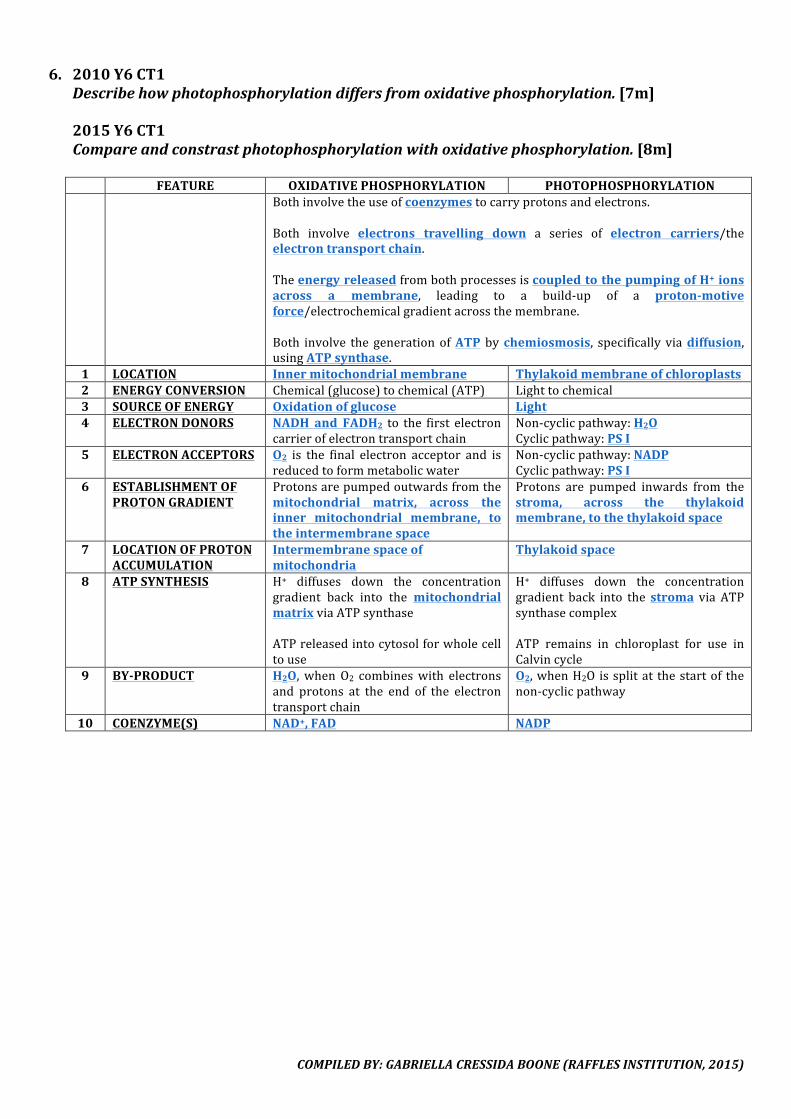
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
6. 2010Y6CT1Describehowphotophosphorylationdiffersfromoxidativephosphorylation.[7m]2015Y6CT1Compareandconstrastphotophosphorylationwithoxidativephosphorylation.[8m]
FEATURE OXIDATIVEPHOSPHORYLATION PHOTOPHOSPHORYLATION Bothinvolvetheuseofcoenzymestocarryprotonsandelectrons.
Both involve electrons travelling down a series of electron carriers/theelectrontransportchain.TheenergyreleasedfrombothprocessesiscoupledtothepumpingofH+ionsacross a membrane, leading to a build-up of a proton-motiveforce/electrochemicalgradientacrossthemembrane.Both involve the generationofATP bychemiosmosis, specifically viadiffusion,usingATPsynthase.
1 LOCATION Innermitochondrialmembrane Thylakoidmembraneofchloroplasts2 ENERGYCONVERSION Chemical(glucose)tochemical(ATP) Lighttochemical3 SOURCEOFENERGY Oxidationofglucose Light4 ELECTRONDONORS NADH and FADH2 to the first electron
carrierofelectrontransportchainNon-cyclicpathway:H2OCyclicpathway:PSI
5 ELECTRONACCEPTORS O2 is the final electron acceptor and isreducedtoformmetabolicwater
Non-cyclicpathway:NADPCyclicpathway:PSI
6 ESTABLISHMENTOFPROTONGRADIENT
Protonsarepumpedoutwardsfromthemitochondrial matrix, across theinner mitochondrial membrane, totheintermembranespace
Protons are pumped inwards from thestroma, across the thylakoidmembrane,tothethylakoidspace
7 LOCATIONOFPROTONACCUMULATION
Intermembranespaceofmitochondria
Thylakoidspace
8 ATPSYNTHESIS H+ diffuses down the concentrationgradient back into themitochondrialmatrixviaATPsynthaseATPreleasedintocytosolforwholecelltouse
H+ diffuses down the concentrationgradient back into the stroma via ATPsynthasecomplexATP remains in chloroplast for use inCalvincycle
9 BY-PRODUCT H2O,whenO2 combineswith electronsand protons at the end of the electrontransportchain
O2,whenH2Oissplitat thestartof thenon-cyclicpathway
10 COENZYME(S) NAD+,FAD NADP
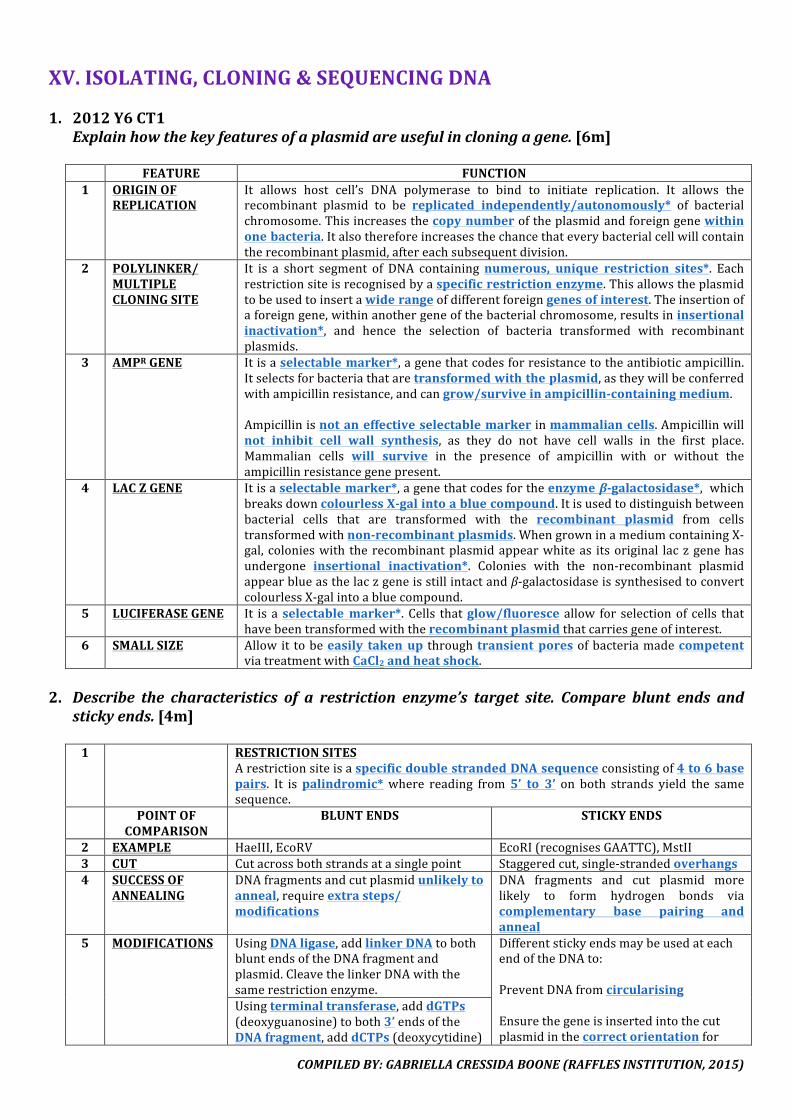
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
XV.ISOLATING,CLONING&SEQUENCINGDNA
1. 2012Y6CT1Explainhowthekeyfeaturesofaplasmidareusefulincloningagene.[6m] FEATURE FUNCTION1 ORIGINOF
REPLICATIONIt allows host cell’s DNA polymerase to bind to initiate replication. It allows therecombinant plasmid to be replicated independently/autonomously* of bacterialchromosome.Thisincreasesthecopynumberoftheplasmidandforeigngenewithinonebacteria.Italsothereforeincreasesthechancethateverybacterialcellwillcontaintherecombinantplasmid,aftereachsubsequentdivision.
2 POLYLINKER/MULTIPLECLONINGSITE
It is a short segment of DNA containingnumerous, unique restriction sites*. Eachrestrictionsiteisrecognisedbyaspecificrestrictionenzyme.Thisallowstheplasmidtobeusedtoinsertawiderangeofdifferentforeigngenesofinterest.Theinsertionofaforeigngene,withinanothergeneofthebacterialchromosome,resultsininsertionalinactivation*, and hence the selection of bacteria transformed with recombinantplasmids.
3 AMPRGENE Itisaselectablemarker*,agenethatcodesforresistancetotheantibioticampicillin.Itselectsforbacteriathataretransformedwiththeplasmid,astheywillbeconferredwithampicillinresistance,andcangrow/surviveinampicillin-containingmedium.Ampicillinisnotaneffectiveselectablemarkerinmammaliancells.Ampicillinwillnot inhibit cell wall synthesis, as they do not have cell walls in the first place.Mammalian cells will survive in the presence of ampicillin with or without theampicillinresistancegenepresent.
4 LACZGENE Itisaselectablemarker*,agenethatcodesfortheenzymeβ-galactosidase*,whichbreaksdowncolourlessX-galintoabluecompound.Itisusedtodistinguishbetweenbacterial cells that are transformed with the recombinant plasmid from cellstransformedwithnon-recombinantplasmids.WhengrowninamediumcontainingX-gal, colonieswith therecombinantplasmidappearwhiteas itsoriginal laczgenehasundergone insertional inactivation*. Colonies with the non-recombinant plasmidappearblueasthelaczgeneisstillintactandβ-galactosidaseissynthesisedtoconvertcolourlessX-galintoabluecompound.
5 LUCIFERASEGENE It is aselectablemarker*. Cells thatglow/fluoresce allow for selectionof cells thathavebeentransformedwiththerecombinantplasmidthatcarriesgeneofinterest.
6 SMALLSIZE Allowit tobeeasily takenup throughtransient poresofbacteriamadecompetentviatreatmentwithCaCl2andheatshock.
2. Describe the characteristics of a restriction enzyme’s target site. Compare blunt ends and
stickyends.[4m]
1 RESTRICTIONSITESArestrictionsiteisaspecificdoublestrandedDNAsequenceconsistingof4to6basepairs. It ispalindromic* where reading from5’ to 3’ on both strands yield the samesequence.
POINTOFCOMPARISON
BLUNTENDS STICKYENDS
2 EXAMPLE HaeIII,EcoRV EcoRI(recognisesGAATTC),MstII3 CUT Cutacrossbothstrandsatasinglepoint Staggeredcut,single-strandedoverhangs4 SUCCESSOF
ANNEALINGDNAfragmentsandcutplasmidunlikelytoanneal,requireextrasteps/modifications
DNA fragments and cut plasmid morelikely to form hydrogen bonds viacomplementary base pairing andanneal
5 MODIFICATIONS UsingDNAligase,addlinkerDNAtobothbluntendsoftheDNAfragmentandplasmid.CleavethelinkerDNAwiththesamerestrictionenzyme.
DifferentstickyendsmaybeusedateachendoftheDNAto:PreventDNAfromcircularisingEnsurethegeneisinsertedintothecutplasmidinthecorrectorientationfor
Usingterminaltransferase,adddGTPs(deoxyguanosine)toboth3’endsoftheDNAfragment,adddCTPs(deoxycytidine)

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
toboth3’endsoftheplasmid. directionalcloning.3. OutlinehowrestrictionenzymesareusedforremovingsectionsofDNAfromachromosome.
[6m]
1 NATURALROLERestriction enzymes cut and destroy foreign DNA carried by bacteriophages attacking the bacteria, bybreaking phosphodiester bondswithin these foreignDNA. Itconfers protection to the bacteria againstbacteriophages;itsownDNAismethylatedforprotection.(Note:methylationofcytosineintheDNAchangesconformationoftheDNAattherestrictionsitesuchthatitisno longer complementary in conformationand charge totherestrictionenzyme’sactive site.HencetherestrictionenzymeisunabletorecogniseandcleavetheDNA.)
2 RECOGNITIONAsDNAisusuallycomplexedwithhistoneproteinsinachromosome,digestionofhistonesbyproteaseisneededtoreleaseDNAfromproteinforattackbyrestrictionenzymes.Arecognitionenzyme,beinghighlyspecificwillbeabletorecogniseaspecificsequenceof4-6nucleotidebasepairsthatispalindromic.Thisiscalledarecognitionsite(e.g.EcoRIrecognisesGAATTC).Differentrestrictionenzymesrecognisedifferentrestrictionsites.Itsspecificityisduetocomplementarityofconformationbetweenauniquerestrictionsiteandtheactivesiteoftheenzyme.
3 CLEAVINGIfaparticularsectionofDNA is toberemoved,chooseenzymeswithrestriction sites flanking thespecificsection.Uponrecognitionandbinding to restriction site, theenzymewillhydrolyse the phosphodiester bonds onbothstrandsoftheDNAbetweenspecificbases.Restrictionwillresultineitherbluntends(e.g.HaeIII)orstickyends(e.g.EcoRI).
4 ANNEALINGStickyendsallowannealingofcomplementarybaseson the2separateDNAmolecules,allowingrejoiningofDNAmolecules.DNA ligase then seals the nick between the DNA molecules by catalysing the formation of covalentphosphodiesterbondsbetweenadjacentnucleotidesofbothstrands.
4. Explain, giving an example, what ismeant by recombinant DNA. Explainwhy only a small
proportionofrecombinantplasmidsofthedesiredconfigurationisobtained.[3m]1 RECOMBINANTDNA
RecombinantDNAismadeupofDNAfrommorethanonesource.When making recombinant DNA, the fragment of interest and the plasmid need to be cut with the samerestrictionenzymeandthennicksaresealedbyDNAligase.
2 INSULINHumaninsulingenecanbeinsertedintoaplasmidofbacterialorigintoformrecombinantDNA.
3 PROBLEMSPlasmidmayrecircularisethroughself-annealingofcomplementarystickyendsoneitherend.Geneofinterestmaycircularisethroughself-annealingofcomplementarystickyendsoneitherend.Plasmidsmayformconcatamers.Multiplecopiesofthegeneofinterestmayannealtoformtandemrepeats.
4 SOLUTIONDifferentstickyendsmaybeusedateachendoftheDNAto:PreventDNAfromcircularising.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
Ensurethegeneisinsertedintothecutplasmidinthecorrectorientationfordirectionalcloning.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
5. Explainhowhumaninsulincanbesynthesisedbybacteria.[8m]
1 ISOLATIONOFHUMANINSULINmRNAIsolatemRNAfrombetacellsofisletsofLangerhansinpancreascodingforhumaninsulin.Thisisdonebythe restriction enzyme that cuts at restriction sites flanking the gene of interest, breaking thephosphodiesterbondsbetweenadjacentnucleotidesonthemRNAmolecule.UsingmRNAas a template andreverse transcriptase*, carry out reverse transcription toproducesingle-strandedcDNA*viacomplementarybasepairing.UsingRNase*,degradethemRNAafterthefirststrandofcDNAisproduced.UsingDNApolymerase*,synthesisethesecondstrandofDNAcomplementarytothefirststrand.Double-strandedcDNA*isobtained.UsepolymerasechainreactiontoamplifythecopyofinsulincDNA.
2 FORMATIONOFRECOMBINANTDNACuttheplasmidwitharestrictionenzyme*thatleavesbluntends*(e.g.EcoRV).
LINKERS TERMINALTRANSFERASEAddlinkerDNA*tobothbluntendsofthecDNAandplasmidusingDNAligase.CleavethelinkerDNAwiththesamerestrictionenzyme*tocreatecomplementarystickyends*.
Useterminaltransferase.AdddGTPs(deoxyguanosine)toboth3’endsoftheDNAfragment.AdddCTPs(deoxycytidine)toboth3’endsoftheplasmid.
Mix the cut plasmid with the cDNA to allow complementary sticky ends* of the gene of interest andplasmid to anneal through formation of hydrogen bonds. Adenine pairs with thymine, and cytosine withguanine.AddDNAligase*toseal thenicksthroughformationofcovalentphosphodiesterbondsbetweenadjacentnucleotidesofbothstrands.Arecombinantplasmidisthenobtained.
3 TRANSFORMATIONWITHRECOMBINANTPLASMIDBacteriaismadecompetentandtransformedwiththerecombinantplasmid,donebyheatshocktreatmentinthepresenceofCaCl2soitcanpickuptherecombinantplasmidthroughtransientpores.
4 SELECTIONOFSUCCESSFULLYTRANSFORMEDCELLSAMPICILLIN+X-GAL+LACTOSE AMPICILLIN+TETRACYCLINE
Plate bacteria in a selection medium containingampicillinantibiotic,lactoseandX-gal*toselectforbacteriathataretransformed.Bacteria that possessplasmid will exhibit antibioticresistanceandsurviveintheselectionmedium.Bacteria thatpossess therecombinant plasmidwillappearaswhitecoloniesbecause the lac z gene*onthe plasmid would have been insertionallyinactivated*bythehumaninsulincDNA.Bacteriathatdonotpossesstherecombinantplasmidwillappearblueasthelaczgeneisstillintactandinthepresenceoflactose,β-galactosidaseissynthesisedtoconvertcolourlessX-galintoabluecompound.
Plate bacteria in a selection medium containingampicillin antibiotic* to select for bacteria that aretransformed with plasmid carrying an intactampicillinresistancegene.Bacteria that possessplasmidwill exhibit ampicillinresistanceandsurviveintheselectionmedium.Carry out replica plating* to do a second antibioticselection for transformed bacteria that contain therecombinant plasmid, in a selection medium withtetracycline antibiotic*. [During replica plating,velvet surface is used as bacteria can stick to itwithoutdamage.]Bacteria thatpossessrecombinant plasmidwillnotexhibittetracyclineresistanceandasthetetracyclineresistance gene would have been insertionallyinactivated.
5 SCALINGUP&PURIFICATIONOFPRODUCTExpressionofgenes/insulinissynthesisedbybacteriagrowninnutrientbrothoffermenter/bioreactor.Extractandpurifythepro-insulin*andenzymaticallycleaveoffchainCpeptideandchemicallycombinechainsAandBpeptideviadisulfidebonds.
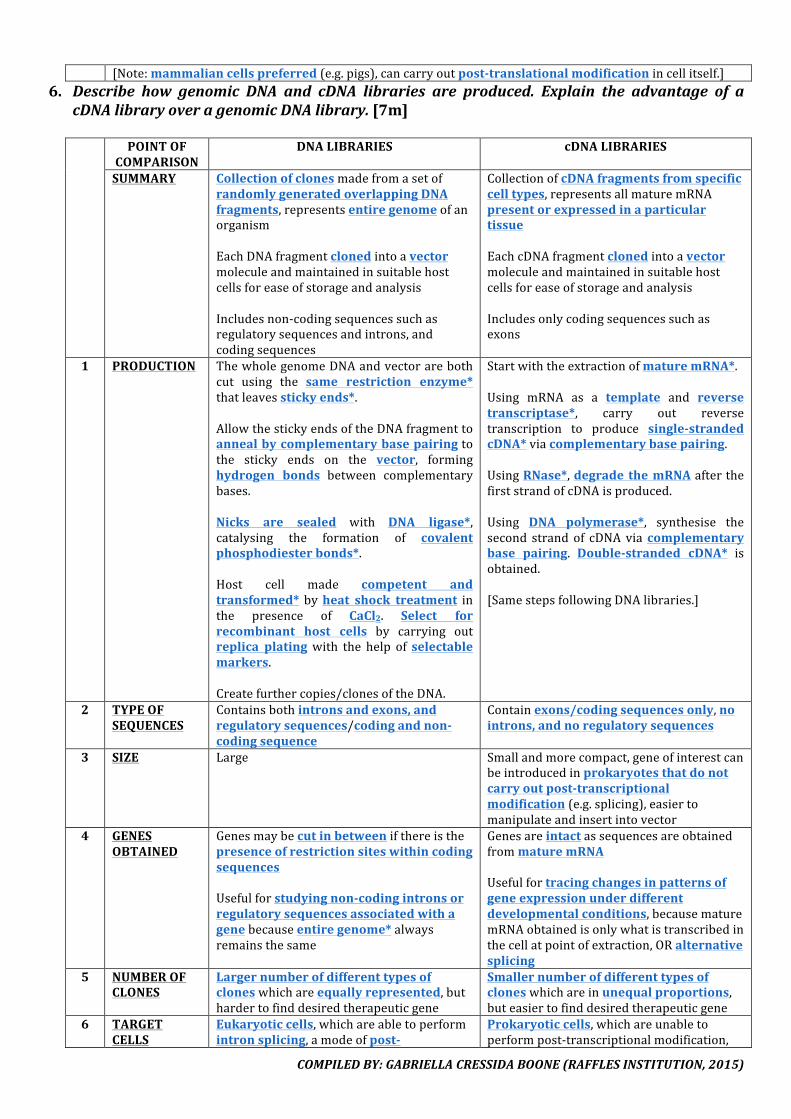
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
[Note:mammaliancellspreferred(e.g.pigs),cancarryoutpost-translationalmodificationincellitself.]6. Describe how genomic DNA and cDNA libraries are produced. Explain the advantage of a
cDNAlibraryoveragenomicDNAlibrary.[7m] POINTOF
COMPARISONDNALIBRARIES cDNALIBRARIES
SUMMARY CollectionofclonesmadefromasetofrandomlygeneratedoverlappingDNAfragments,representsentiregenomeofanorganismEachDNAfragmentclonedintoavectormoleculeandmaintainedinsuitablehostcellsforeaseofstorageandanalysisIncludesnon-codingsequencessuchasregulatorysequencesandintrons,andcodingsequences
CollectionofcDNAfragmentsfromspecificcelltypes,representsallmaturemRNApresentorexpressedinaparticulartissueEachcDNAfragmentclonedintoavectormoleculeandmaintainedinsuitablehostcellsforeaseofstorageandanalysisIncludesonlycodingsequencessuchasexons
1 PRODUCTION ThewholegenomeDNAandvectorarebothcut using the same restriction enzyme*thatleavesstickyends*.AllowthestickyendsoftheDNAfragmenttoannealbycomplementarybasepairingtothe sticky ends on the vector, forminghydrogen bonds between complementarybases.Nicks are sealed with DNA ligase*,catalysing the formation of covalentphosphodiesterbonds*.Host cell made competent andtransformed* byheat shock treatment inthe presence of CaCl2. Select forrecombinant host cells by carrying outreplica platingwith the help of selectablemarkers.Createfurthercopies/clonesoftheDNA.
StartwiththeextractionofmaturemRNA*.Using mRNA as a template and reversetranscriptase*, carry out reversetranscription to produce single-strandedcDNA*viacomplementarybasepairing.UsingRNase*,degrade themRNAafterthefirststrandofcDNAisproduced.Using DNA polymerase*, synthesise thesecond strandof cDNAviacomplementarybase pairing. Double-stranded cDNA* isobtained.[SamestepsfollowingDNAlibraries.]
2 TYPEOFSEQUENCES
Containsbothintronsandexons,andregulatorysequences/codingandnon-codingsequence
Containexons/codingsequencesonly,nointrons,andnoregulatorysequences
3 SIZE Large Smallandmorecompact,geneofinterestcanbeintroducedinprokaryotesthatdonotcarryoutpost-transcriptionalmodification(e.g.splicing),easiertomanipulateandinsertintovector
4 GENESOBTAINED
GenesmaybecutinbetweenifthereisthepresenceofrestrictionsiteswithincodingsequencesUsefulforstudyingnon-codingintronsorregulatorysequencesassociatedwithagenebecauseentiregenome*alwaysremainsthesame
GenesareintactassequencesareobtainedfrommaturemRNAUsefulfortracingchangesinpatternsofgeneexpressionunderdifferentdevelopmentalconditions,becausematuremRNAobtainedisonlywhatistranscribedinthecellatpointofextraction,ORalternativesplicing
5 NUMBEROFCLONES
Largernumberofdifferenttypesofcloneswhichareequallyrepresented,buthardertofinddesiredtherapeuticgene
Smallernumberofdifferenttypesofcloneswhichareinunequalproportions,buteasiertofinddesiredtherapeuticgene
6 TARGETCELLS
Eukaryoticcells,whichareabletoperformintronsplicing,amodeofpost-
Prokaryoticcells,whichareunabletoperformpost-transcriptionalmodification,

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
transcriptionalmodification andsorequirecDNAfrommaturemRNA

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
7. Describe the three steps of polymerase chain reaction. Explain how it facilitates geneticfingerprinting.[6m]1 STAGE1–DENATURATION
DenaturationofdoublestrandedDNAintosinglestrandedDNAbyheatingto95°C*.Weak hydrogen bonds* between complementary bases of DNA molecule are broken due to increasedmolecularvibrations.
2 STAGE2–ANNEALINGTemperatureisloweredto64°C*toallowforaDNAprimer*toanneal.TheDNAprimerbindsspecificallytothe3’endsof theregionsflankingthegeneofinterestonthesinglestrandedDNAbyformationofhydrogenbondsbetweencomplementarybasepairs.(Note:single-strandedDNAstrandsnolongerannealtoeachotherastheconcentrationofprimersexceedtheconcentrationofdenaturedDNA.
3 STAGE3–EXTENSIONTaqpolymerase*performssynthesisofcomplementaryDNAstrandat72°C*.Chain extension occurs from 3’ end of the DNA primer which provides a free 3’ OH required by thepolymerase.
4 TaqPOLYMERASETaqpolymeraseisthermostable*,evenathightemperaturesof95°C*whenDNAisdenatured.Theyhaveahigher proportion of cysteine residues and thus strong covalent, disulfide bonds, or numerousintramolecularinteractionsthatholdthetertiarystructureoftheenzymetogether.TaqpolymeraseremainsfunctionalandcanbereusedafteracycleofPCRiscompleted,allowingPCRtobeautomated*.MammalianDNApolymerasewouldbedenaturedandnewenzymeswillhavetobeaddedforeverycycle.
5 GENETICFINGERPRINTINGPCRamplifiesDNAfromalimitedsourceofDNA,whichisespeciallyimportantinacrimescenewheretissueevidenceislimited.ThisensuresthatthereissufficientamountofDNAforproceduresinvolvedinanalysis,asthebandsmaynotbevisibleifDNAislimited.
8. DescribethelimitationsofPCR.[6m]
1 LACKSPROOFREADINGABILITYTaqpolymeraselacks3’to5’proofreadingability.ErrorsoccurringearlyinthePCRreactionwillgetcompoundedwitheachreplicationcycleandalldaughtermoleculesresultingfromtheearlyerrorwillbeexponentiallyaffected.
2 SEQUENCEINFORMATIONRequiressequenceinformationflankingtargetregionofamplification.TheDNAprimerbinds specifically to the3’ ends of the regions flanking the geneof interest. If flankingsequencesareunknown,noproperprimerscanbesynthesisedtoamplifygeneofinterest.
3 LIMITTOSIZEDNAfragmentstobeamplifiedarelimitedtoabout3kb.Further increase in length of target sequence decreases efficiency of amplification. This is because TaqpolymerasetendstofallofftheDNAtemplatebeforechainextensioniscomplete.
4 CONTAMINANTDNAPoorlaboratoryskillsmayresultinPCRmixturecontainingminuteamountsofcontaminantDNA.SuchunwantedDNAsequencesmaybeamplifiedtosignificantamounts,alongsidethetargetDNAsequences.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
9. ExplainhowgelelectrophoresisisusedtoseparatefragmentsofDNA.[6m]
1 LOADINGFragments of DNA arepipetted intowells at the top of the agarose gel, furthest away from the positiveelectrode.Buffersolutioncontainsionswhichallowsconductionofelectriccurrent,allowingnegativelychargedDNAmoleculestomovefromthenegativeelectrodetothepositiveelectrode.Denseloadingdyecontainsglycerol,makingDNAsampledenserthanthebuffersoitcansinktothebottomofthewell.Tracking dye isadded to thecolourless DNA sample toallowvisualisation of theseparation process.2coloured dyes act as visual markers, one typically runs ahead of the sample, the other runs after thesample.
2 MIGRATIONWhen the direct current is switched on, thenegatively-charged DNAmigrates out of thewell towards thedirectionofthepositiveelectrode/anode.Fragmentsmigratethroughtheagarosegelmatrix,madeupofameshworkofpolysaccharides.Themeshworkimpedesmovementoflargerfragmentsmorethantheshorterfragments.Thus,largerfragmentsmigrateslowerthanshorterfragments.
3 DNALADDERMarkers,whichmakeup theDNA ladder, are run in oneof the lanes. These arepreparedmixtures ofDNAfragmentsofknownsizes,whichformthebasisofcomparisonfortheunknownfragmentsinthesample.
10. Possiblebandingpatterns.[6m]
1 POSITIVERESULT
NbandsofsizeXcoincideswithmolecularsizeofXgeneofXbases.2 NEGATIVERESULT
Mbandsobservedaresmaller/largerthanexpectedmolecularsizeofXgene.3 NORESULT
NobandsobservedasprimerscouldnotannealtoDNA,Xgenenotpresent.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
11. ExplainwhatismeantbyRestrictionFragmentLengthPolymorphism(RFLP)andhowitcanbe detected. State three of its applications and suggest one of its limitations. Explain thepurposeofthecontrolDNAwithknownnumberoftandemrepeats.[10m]
1 RFLP
RestrictionFragmentLengthPolymorphism(RFLP)referstotheuniquebandingpatternamongindividualswhentheirgenomicDNAaredigestedbyrestrictionenzymesandseparatedbygelelectrophoresis.Due to thepresenceofDNA polymorphisms indifferent individuals, therewill bevariations in firstly, thenumber and location of restriction sites (SNPs), and secondly, the number of tandemly repeatednucleotidesequencesamongindividuals(STRs&VNTRs).
STRs&VNTRs SNPsAfter gel electrophoresis, to detect polymorphisms, the double-stranded DNA fragments are denatured to single-stranded DNAusinganalkalinesolution likeNaOH.Theyarethentransferredtoanitrocellulosemembrane.Nucleic acid hybridisation is carried out by incubating themembrane with single-stranded radioactive DNA probecomplementarytotheDNA fragmentof interest.Theradioactively-labeled probe hybridises by complementary base pairing andformationofhydrogenbonds.When the nitrocellulose membrane is placed on an X-ray film,radioactive areas expose the film, appearing as bands in theautoradiograph, highlighting the differences in restriction sites andhence,theDNApolymorphismsindifferentindividuals.
CarryoutPCRthendigestwithrestrictionenzymethengelelectrophoresis.The DNA fragments are stainedwithethidium bromide, a DNA bindingdye, causing them to fluoresceunderUVlight.Band patterns will highlight thedifferences in restriction sites andhence, the DNA polymorphisms indifferentindividuals.
2 APPLICATIONSItfacilitateslinkagemapping,detectionofdisease,andDNAfingerprinting(forensics&paternitytesting).Linkagemapping:1.Nucleotidesequenceofdiseasealleleisnotknown.2.Mutationinthediseasealleleisnotknown.3.Mutationinthediseasealleledidnotintroduceorremovearestrictionsite.DNAfingerprinting:1.MultiplelociareusuallycomparedastheywillgiverisetouniqueDNAfingerprinting/bandingpatterns.TheprobabilityoftwoindividualswithidenticalDNAbandingprofileisextremelylow.Therefore,individualscanbedistinguishedfromeachother,allowingaccurateidentification.2.Whenusingasingle locus, it ispossiblefor2individualstohavethesamebandingpattern,especiallyiftheyarerelated.
3 LIMITATIONOFRFLPMARKERThere is a limitationwhen studyingRFLPs flanking a disease gene.An individualmay suffer froma diseaseeventhoughgeneticscreeningusingtheRFLPmarkeridentifiedhimasbeingonlyacarrier.Thismayhappenduetocrossingover*occurringatalocusbetweenthediseasealleleandtheRFLPmarkerduringmeiosis,such that the disease allele has become linked to the healthy RFLP allele marker. So although he isheterozygousfortheRFLPallelemarkers,heisinfacthomozygousrecessivefortheactualdiseaseallele.However,suchRFLPsarestillusedbecausethey lie very close to the geneof interest,andcrossing overfrequenciestendtobeverylow.Therefore,RFLPmarkerallelemustbetightlylinkedtothediseaseallelesothattheyhaveahigherchanceofco-segregating/beinginheritedtogether.ItminimisesthechanceofcrossingoverattheregionbetweentheRFLPmarkeralleleanddiseaseallele,preventingtheirlinkagefrombeingdisrupted.
4 LIMITATIONOFGENEMARKERGenes code for functional polypeptide. Hence, their DNA sequences are highly conserved* due to theirimportantbiologicalfunction,sotheirmutationrateislowerthannon-codingregions.Genesusuallydonothavesufficientdifferencestoproduceuniquebandingpatternsfordifferentiationbetweenindividuals.
5 CONTROLDNADNAcontainingthelocusofstudywithknownnumberoftandemrepeatsisusedasacontrol.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
Ifabandofexpectedsizeisnotseenonthegel,itshowsthatthereagentsarenotworkingproperly(viceversa).
12. ExplainhowRFLPanalysishashelpedtheprocessofdetectinganamedgeneticdisease.[8m]
1 GENETICDISEASE1.SicklecellanaemiaisanautosomalrecessivegeneticdiseasewhichiscausedbyasubstitutionmutationintheDNAcodingfortheβ-globinchain*ofhaemoglobin,wherethymineissubstitutedbyadenine,the6thtriplet codon ischangedfromGAGtoGUG,resulting intheaminoacidglutamatic acid*beingreplacedbyvaline*.[Redbloodcellsassumeanabnormal rigid, sickle shape andcan lead toanemia,obstructionofcapillariesduetosickling,organdamage,dropsinhaemoglobinlevelsandsubsequentlyevendeath.]2.ThismutationislocatedwithinarestrictionsitefortherestrictionenzymeMstII*,suchthattherestrictionsiteisnoweliminatedinthedisease-causingalleleHbS,sonorestrictionsitecanberecognisedbyMstII.3.ItispossibletoinferthegenotypeofindividualsusingRFLPanalysis.4.WhennormalanddiseaseallelesaredigestedbyMstII,adifferentmixtureof fragmentswillbeproducedfromeachallele.
2 RFLPANALYSIS5.ThedigestedDNAisseparatedbasedonsizeusinggelelectrophoresis.6.Thefragmentsarethenmadesingle-strandedthroughdenaturationusingNaOHandtransferredontoanitrocellulosemembrane.7.Nucleicacidhybridisationiscarriedoutbyincubatingthemembranewithradioactively-labeledprobe,complementarytotheβ-globingeneonbothsidesoftherestrictionsite.Theyannealviacomplementarybasepairingwithtargetfragments.8.WhenthenitrocellulosemembraneisplacedonanX-rayfilm,radioactiveareasexposethefilm,revealingthebandingpatternintheautoradiograph.9.SincetheHbSallelehaslosttheMstIIrestrictionsitethroughsubstitution,itisnotdigestedandwillhenceyieldthelargestfragmentwhenvisualisedintheautoradiograph.10. A single large band indicates the presence of 2 HbS alleles and hence, a sufferer; 2 bands, oneintermediate and one small indicates the presence of 2 HbA alleles and hence, a normal individual; 3bands,onelarge,oneintermediate,andonesmall,indicatesthepresenceof1HbSalleleand1HbAalleleandhence,aheterozygote,whoisphenotypicallynormal.(Note:adiagrammayhelp.)
13. Explain what a chromosome map is and how RFLP can be used with linkage mapping to
constructachromosomemap.[5m]1 CHROMOSOMEMAP
Adiagramshowingpositionsofwhereknowngenesoccuralongachromosomeandtheirrelativedistancesapartfromeachother.
2 RFLPGenesarelinkedtoandthusdetectedbyRFLPs.ParentswithknowngenotypesattwoRFLPlociarecrossedtofindrecombinationfrequency.Recombinationfrequency(%)oflinkedRFLPsindicatesrelativedistancebetween2RFLPloci.1%=1centiMorgan(cM)Highrecombinationfrequencymeans2RFLPsarefarapart;lowrecombinationfrequencymeans2RFLPsareclosetogether.
3 LIMITATIONS(MAPPINGOFHUMANGENOME)Linkagemapping:Requirescrossingovertoobtainrecombinants,unethicalforhumanbeings.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
Requiresshortgenerationtimeinthestudiedorganism,buthumanbeings’generationtimeistoolong.Dependsonobservablephenotypes,buthumangenomeconsistsofcodingfornon-observablephenotypes.Involvesheritablediseases,buthumangenomeconsistsofcodingfornon-inheritablediseases.
14. Discuss the benefits and ethical concerns that have arisen regarding the human genomeproject.[6m]
1 IMPROVEDDIAGNOSIS
Diagnostictestscanbedevelopedtoscreencarrierswithfaultygenestoinformthemofpossiblerisks/tomakechanges in lifestyle to minimise risks. OR To screen embryos during prenatal testing to avoid the birth ofaffectedchildren.
2 DESIGNOFTREATMENTSDesignoftreatmentsforgeneticdisorderswherethemutationsofthegeneisidentifiedsothatacuremaybepossiblethrough:(1)genetherapyOR(2)drugscanbedevelopedtoblocktheactionoftheenzymesOR(3)switchingofffaultygenewithgeneknockouttechniquessuchasanti-senseRNAtechnology.
3 PHARMACOGENOMICSTailoringofdrugprescriptiontofitpatient’sconditionforgreaterefficacyandtoavoiddangeroussideeffectsduetoaperson’sgeneticmakeup.
4 FAIRNESSINUSEOFGENETICINFORMATIONInsurers,employers,courts,schoolsandmilitaryandotherstakeholdersmayhaveaccesstopersonalgeneticinformationandmayusesuchinformationinanethicallyquestionableway.Insurers:raisingpremiumsbasedongeneticpredispositiontodiseaseEmployers:hiringandpromotingbasedongeneticsuperiorityratherthanonmeritCourts:judgeadefendantbasedonageneticpredispositiontoviolenceratherthantheseverityofthecrime
5 PRIVACYANDCONFIDENTIALITYOFGENETICINFORMATIONRiskprofiles fordiseasesmaybeaccessible to insurers, employers, courts, schools andmilitary. Individual’sprivacymaybeinfringedandusedinadiscriminatorymanner.
6 PSYCHOLOGICALIMPACTANDSTIGMATISATIONDUETOGENETICDIFFERENCESPersonal genetic information may affect an individual and society’s perception of them in a negative way.Individualmaywithdrawfromsocietyorsocietymaytreatthepersonasanoutcastdependingonaperson’sgeneticpredispositiontoillness,intellect.
7 REPRODUCTIVEDECISIONMAKINGReproductive issues would include having adequate informed consent for complex and potentiallycontroversialprocedures,useofgeneticinformationinreproductivedecisionmaking,andreproductiverights.Thereisachancethatfoetalgenetictestingmaynotbereliableininstancesinvolvingcomplexdiseaseswheregenetic predisposition and actual occurrence of a disease may not correlate, so parents may abort healthyfoetusesunknowingly.Theremaybeelementsofsocietythatmaypracticeeugenicsbasedonaccesstogeneticinformation.
8 CLINICALISSUESEducation of doctors, other health service providers, patients and general public in genetic capabilities,scientific limitations and social risks, is important. Proper counselingmustbedone in conjunctionwith riskprofiling, and where there is no treatment for certain diseases. The public may be unprepared for suchinformationandthatitisirresponsibletoprovideitwithoutanexperttohelpputinincontext.Implementationofstandardsandqualitycontrolmeasuresintestingproceduresarealsoneeded.
9 UNCERTAINTIESASSOCIATEDWITHGENETESTSTheremay be uncertainties associatedwith gene tests, for susceptibilities and complex conditions, that arelinkedtomultiplegenesandgene-environmentconditions.
10 HUMANRESPONSIBILITY/FREEWILL/GENETICDETERMINISMThereareconceptualandphilosophical implicationsregardinghumanresponsibility, freewillversusgeneticdeterminism.Knowledgeofwhethergenesaloneaffectbehaviourandhumanphysiologyarenotwellstudied,andwithsomanygenesandthecomplexityofmultigenicinteractions,wemaynotfullyunderstandsuchissuesenoughtodeterminebehaviourandphysiologybasedonjustthegeneticcode.
11 COMMERCIALISATION/PROPERTYRIGHTSGeneticinformationofindividualsmaybecomeprivateintellectualpropertyofbiotechonologycompanies.Commercialisationofrelatedtechnologiesderivedfromsuchresearch,butpossiblynoreciprocaldirectbenefitforindividualscontributingtheirgeneticinformationtotheresearch.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
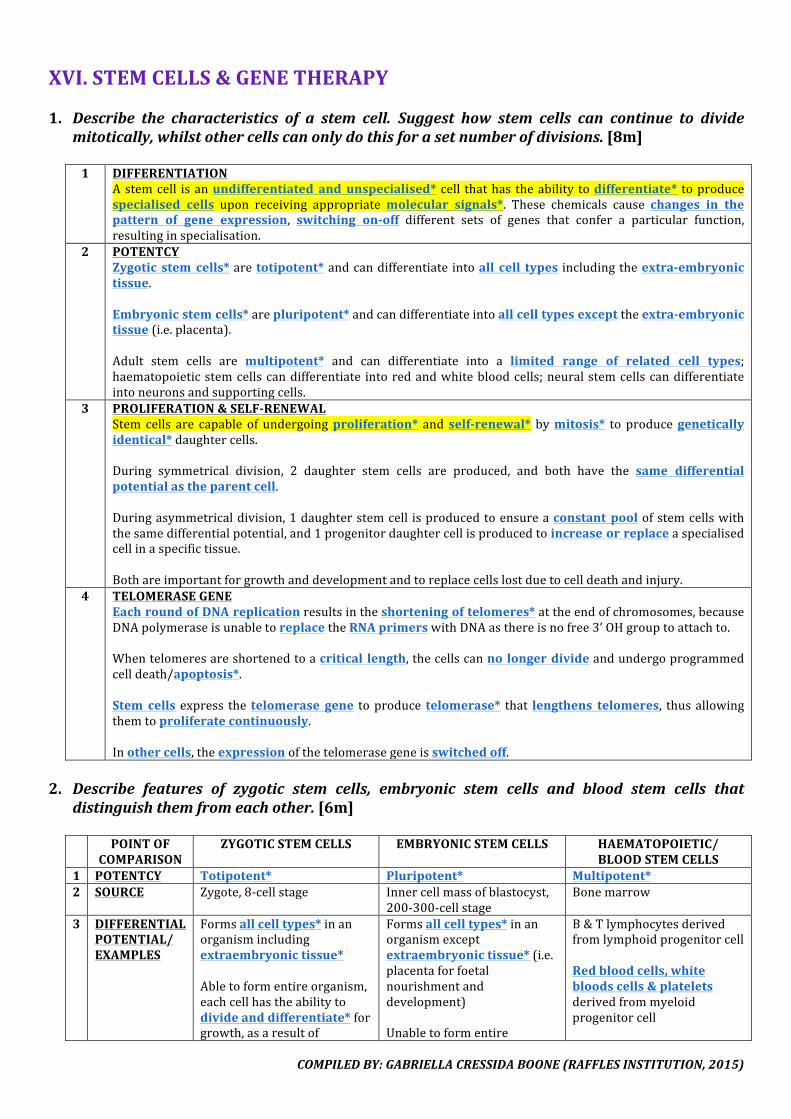
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
XVI.STEMCELLS&GENETHERAPY1. Describe the characteristics of a stem cell. Suggest how stem cells can continue to divide
mitotically,whilstothercellscanonlydothisforasetnumberofdivisions.[8m]
1 DIFFERENTIATIONAstemcell isanundifferentiated andunspecialised* cell thathas theability todifferentiate* toproducespecialised cells upon receiving appropriatemolecular signals*. These chemicals cause changes in thepattern of gene expression, switching on-off different sets of genes that confer a particular function,resultinginspecialisation.
2 POTENTCYZygotic stemcells*aretotipotent*andcandifferentiate intoall cell types includingtheextra-embryonictissue.Embryonicstemcells*arepluripotent*andcandifferentiateintoallcelltypesexcepttheextra-embryonictissue(i.e.placenta).Adult stem cells are multipotent* and can differentiate into a limited range of related cell types;haematopoieticstemcellscandifferentiate intoredandwhitebloodcells;neuralstemcellscandifferentiateintoneuronsandsupportingcells.
3 PROLIFERATION&SELF-RENEWALStem cells are capable of undergoingproliferation* andself-renewal* bymitosis* to producegeneticallyidentical*daughtercells.During symmetrical division, 2 daughter stem cells are produced, and both have the same differentialpotentialastheparentcell.Duringasymmetricaldivision,1daughterstemcell isproducedtoensureaconstantpoolofstemcellswiththesamedifferentialpotential,and1progenitordaughtercellisproducedtoincreaseorreplaceaspecialisedcellinaspecifictissue.Bothareimportantforgrowthanddevelopmentandtoreplacecellslostduetocelldeathandinjury.
4 TELOMERASEGENEEachroundofDNAreplicationresultsintheshorteningoftelomeres*attheendofchromosomes,becauseDNApolymeraseisunabletoreplacetheRNAprimerswithDNAasthereisnofree3’OHgrouptoattachto.Whentelomeresareshortenedtoacritical length,thecellscanno longerdivideandundergoprogrammedcelldeath/apoptosis*.Stem cells express the telomerase gene toproduce telomerase* that lengthens telomeres, thusallowingthemtoproliferatecontinuously.Inothercells,theexpressionofthetelomerasegeneisswitchedoff.
2. Describe features of zygotic stem cells, embryonic stem cells and blood stem cells that
distinguishthemfromeachother.[6m] POINTOF
COMPARISONZYGOTICSTEMCELLS EMBRYONICSTEMCELLS HAEMATOPOIETIC/
BLOODSTEMCELLS1 POTENTCY Totipotent* Pluripotent* Multipotent*2 SOURCE Zygote,8-cellstage Innercellmassofblastocyst,
200-300-cellstageBonemarrow
3 DIFFERENTIALPOTENTIAL/EXAMPLES
Formsallcelltypes*inanorganismincludingextraembryonictissue*Abletoformentireorganism,eachcellhastheabilitytodivideanddifferentiate*forgrowth,asaresultof
Formsallcelltypes*inanorganismexceptextraembryonictissue*(i.e.placentaforfoetalnourishmentanddevelopment)Unabletoformentire
B&TlymphocytesderivedfromlymphoidprogenitorcellRedbloodcells,whitebloodscells&plateletsderivedfrommyeloidprogenitorcell

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
differentialswitchingonofgenesuponreceivingmolecularsignals
organism Unabletoformentireorganism
3. Nameonegeneticdiseasewhichhasbeentreatedwithstemcelltransplantation.Explainwhystemcellissuitableforthispurpose.[4m]1 GENETICDISEASE
Bonemarrowhaematopoieticstemcellstransplantsfromnormalhealthybonemarrowdonorstoleukemiapatients.AdultneuralstemcelltransplantsforParkinson’sdisease/multiplesclerosispatients.
2 SUITABILITYAdult stem cells aremultipotent* and can differentiate into a range of related specialised cell types,restoringfunctionofdamagedordiseasestissues.Stem cells are capable of undergoingproliferation* andself-renewal* bymitosis* to producegeneticallyidentical*daughtercells,tomaintainaconstantpoolofstemcells.Thesehealthystemcellwillcarrythenormalandfunctionalallelethus,theycanproducenormallevelsoffunctionproteinandbeusedtotreatgeneticdiseases.
4. Explainwhy a nucleus taken from a somatic cell from an animalwas used to produce the
embryonicstemcellsinatherapy.[3m]1 SOURCE
Nucleustakenfromthesomaticcellisdiploid*,whilenucleusremovedfromthegamete/oocyte/spermitselfishaploid*.
2 PURPOSEBydoingso,itensurestheembryonicstemcellsproducedfromthegametewouldbegeneticallyidentical*tothecellsoftheanimal.Therefore,whentheseembryonicstemcellsareinjectedintotheanimalagainaftergeneticengineering,theywouldnotberejectedorevokeanimmuneresponse,unlikeforeigncells.
5. Stateargumentsinfavourofandagainsttheuseofanimalgametesfortherapeuticcloningof
humancells.[2m]1 INFAVOUR
Animalgametesaremoreeasilyattainablethanhumangametes.Humangametesarelimitedinsupplycomparedtoanimalgametes.
2 AGAINSTMitochondrialDNAofanimalsisdifferentfromthatofhumans,andthelongtermeffectsofthemixtureofDNAfromtwodifferentspeciesarenotpredictable.NotallproteinsrequiredforproperfunctioningofmitochondriaareencodedbythemitochondrialDNA,becausesomeareencodedforbynuclearDNA.NuclearDNAfromhumansmaynotbeabletosupportthefullfunctioningofmitochondriaofanimalorigins.
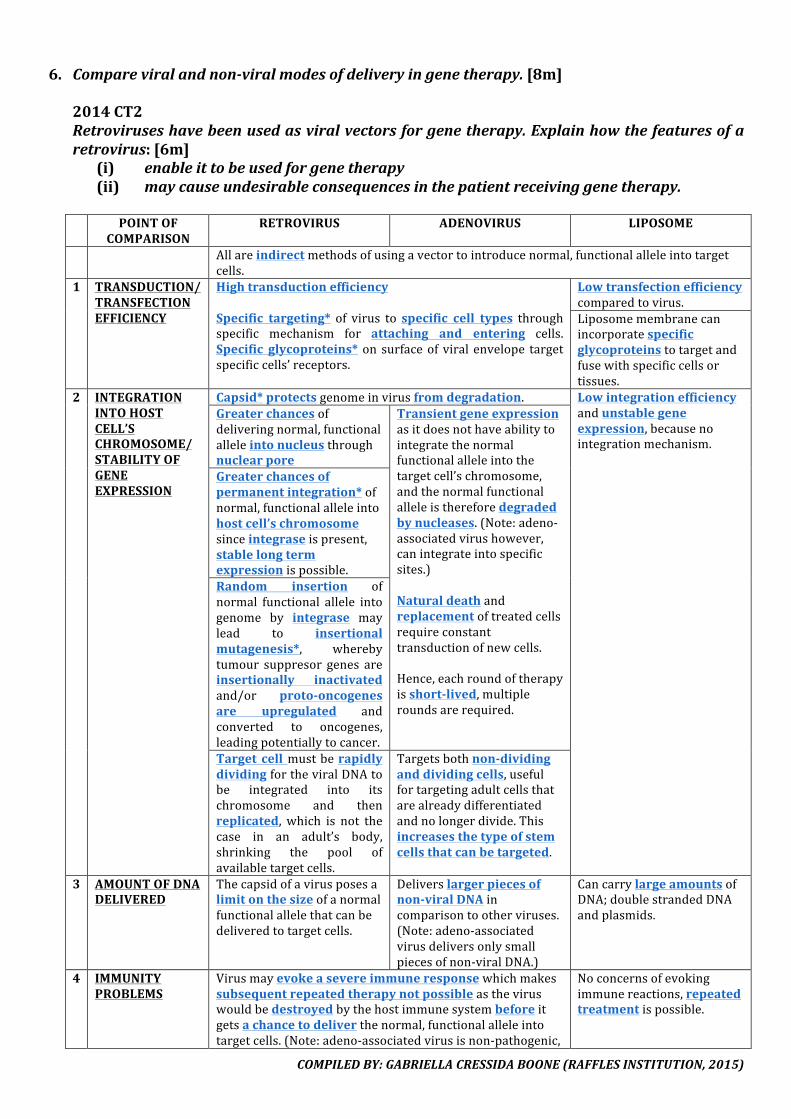
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
6. Compareviralandnon-viralmodesofdeliveryingenetherapy.[8m]2014CT2Retroviruseshavebeenusedasviralvectorsforgenetherapy.Explainhowthefeaturesofaretrovirus:[6m]
(i) enableittobeusedforgenetherapy(ii) maycauseundesirableconsequencesinthepatientreceivinggenetherapy.
POINTOF
COMPARISONRETROVIRUS ADENOVIRUS LIPOSOME
Allareindirectmethodsofusingavectortointroducenormal,functionalalleleintotargetcells.
1 TRANSDUCTION/TRANSFECTIONEFFICIENCY
HightransductionefficiencySpecific targeting* of virus to specific cell types throughspecific mechanism for attaching and entering cells.Specific glycoproteins* on surfaceof viral envelope targetspecificcells’receptors.
Lowtransfectionefficiencycomparedtovirus.Liposomemembranecanincorporatespecificglycoproteinstotargetandfusewithspecificcellsortissues.
2 INTEGRATIONINTOHOSTCELL’SCHROMOSOME/STABILITYOFGENEEXPRESSION
Capsid*protectsgenomeinvirusfromdegradation. Lowintegrationefficiencyandunstablegeneexpression,becausenointegrationmechanism.
Greaterchancesofdeliveringnormal,functionalalleleintonucleusthroughnuclearpore
Transientgeneexpressionasitdoesnothaveabilitytointegratethenormalfunctionalalleleintothetargetcell’schromosome,andthenormalfunctionalalleleisthereforedegradedbynucleases.(Note:adeno-associatedvirushowever,canintegrateintospecificsites.)Naturaldeathandreplacementoftreatedcellsrequireconstanttransductionofnewcells.Hence,eachroundoftherapyisshort-lived,multipleroundsarerequired.
Greaterchancesofpermanentintegration*ofnormal,functionalalleleintohostcell’schromosomesinceintegraseispresent,stablelongtermexpressionispossible.Random insertion ofnormal functional allele intogenome by integrase maylead to insertionalmutagenesis*, wherebytumour suppresor genes areinsertionally inactivatedand/or proto-oncogenesare upregulated andconverted to oncogenes,leadingpotentiallytocancer.Target cellmustberapidlydividingfortheviralDNAtobe integrated into itschromosome and thenreplicated, which is not thecase in an adult’s body,shrinking the pool ofavailabletargetcells.
Targetsbothnon-dividinganddividingcells,usefulfortargetingadultcellsthatarealreadydifferentiatedandnolongerdivide.Thisincreasesthetypeofstemcellsthatcanbetargeted.
3 AMOUNTOFDNADELIVERED
Thecapsidofavirusposesalimitonthesizeofanormalfunctionalallelethatcanbedeliveredtotargetcells.
Deliverslargerpiecesofnon-viralDNAincomparisontootherviruses.(Note:adeno-associatedvirusdeliversonlysmallpiecesofnon-viralDNA.)
CancarrylargeamountsofDNA;doublestrandedDNAandplasmids.
4 IMMUNITYPROBLEMS
Virusmayevokeasevereimmuneresponsewhichmakessubsequentrepeatedtherapynotpossibleastheviruswouldbedestroyedbythehostimmunesystembeforeitgetsachancetodeliverthenormal,functionalalleleintotargetcells.(Note:adeno-associatedvirusisnon-pathogenic,
Noconcernsofevokingimmunereactions,repeatedtreatmentispossible.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
willnotcauseimmunityproblems.)5 SIDEEFFECTS Virusmayregainvirulenceinthehostandcausedisease. Non-pathogenic.
7. Suggestwhyexpressionofanormalalleleisdifferentinthestemcellsgiventotwodifferentgroupsinanexperiment.[3m]
1 COPIESOFNORMAL,FUNCTIONALALLELE
TransfectedstemcellsfromanunaffecteddonoringroupAislikelytohave2copiesofthenormal,functionalalleleinitstargetcell,hencethehigherlevelofexpression;transducedstemcellsingroupBarelikelytohaveatmost1copyofthenormal,functionalalleleintegratedinitsgenomeasviralvectorsposea limitonthesizeofallelethatcanbedelivered,hencethelowerlevelofexpression.
2 INTEGRATIONThenormal,functionalalleleinthetransducedstemcellsingroupBisnotstablyintegratedintothegenomeand hence is gradually lost with each subsequent round of cell division, resulting in lower level ofexpression;stableorpermanentintegrationisnotanissuewithstemcellsfromgroupA,henceresultinginhigherlevelofexpression.
3 CONTROLELEMENT–ENHANCERThenormal,functionalalleleforstemcellsingroupAwasintegratedatapositionundertheinfluenceofanenhancerelement,resultinginhigherlevelofexpression;thenormal,functionalalleleforstemcellsingroupB was integrated at a position not under the influence of an enhancer element, hence a lower level ofexpression.
4 CONTROLELEMENT–SILENCERThenormal,functionalalleleforstemcellsingroupBwasintegratedatapositionunder the influenceofasilencerelement,resultinginlowerlevelofexpression;thenormal,functionalalleleforstemcellsingroupAwasintegratedatapositionnotundertheinfluenceofasilencerelement,henceahigherlevelofexpression.
8. Suggest why genetic therapy is usually attempted for recessive genetic diseases. State the
factorswhichpreventgenetherapyfrombecominganeffectivetreatment.[5m]
1 RECESSIVEGENETICDISEASESMutations toageneusuallyresult inanon-functional proteinorno proteinbeingsynthesisedatall.Suchloss-of-functionmutationscanbemaskedbyintroducingasinglenormal,functionalallelethatcodesforthefunctionalprotein.
2 INEFFECTIVENESS–GENERALTherapyistypicallyshort-livedbecauseofdifficultyinintegratingnormal,functionalalleleintothehostcellchromosomeandnaturaldeathoftreatedcellsmeansthatnewcellsneedtobeconstantlytransformed.Manydiseasesarearesultofmultiplegenesand/ormultiplefactors(e.g.Alzheimer’sdisease,diabetes,etc.)andassuch,willbedifficulttotreatwithgenetherapywhichcurrentlytreatsonlysinglegenedisorders.Some disease such as thalassemia require aprecise level of expression of the normal gene in order fortherapytobeeffective.Regulationoflevelsofgeneexpressioniscurrentlydifficult.
3 INEFFECTIVENESS–MODESOFDELIVERY(refertoSTEMCELLS&GENETHERAPY–Q3)
9. Discussthesocialandethicalconcernsfortheuseofgenetherapy.[6m]
1 AReferto2013prelimpaper3

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
10. DescribeSCIDanditsgenetherapytreatment.Explainwhytheex-vivoapproachisadopted.[8m]
1 SCID
Severe combined immunodeficiency (SCID) is aheritable immunodeficiency diseasewhere childrenwiththisconditionarebornwithoutafunctioningimmunesystem.Theyarevulnerabletosecondary/opportunisticinfectionsfrompeopletheycomeintocontactwith. X-LINKED AUTOSOMALTYPEOFGENETICDISORDER
X-linkedrecessivedisorderX-(linked)SCID*
Autosomalrecessivedisorder
LOCATIONOFMUTATION
Mutation*incommongammachain*gene
Mutation*inadenosinedeaminaseenzyme(ADA)*gene
LOCATIONOFGENE
X-chromosome Chromosome20*
RESULTANTPROTEIN
Non-functionalornocommongammachainsynthesised/interleukinreceptor
Non-functionalornoADA
EFFECTONIMMUNESYSTEM
FailureofbothBandTlymphocytestoproliferateanddifferentiate*
NobreakdownofadenosineAccumulationofdeoxyadenosineToxiccompoundcausesTandBlymphocytestodie
2 SCIDTREATMENT1.Treatmentusuallyinvolvesuseofaviral-mediatedgenedeliverysystem,employinganex-vivo*approach.2.Aretrovirussuchasalentivirus,isdisarmedbyremovinggenesresponsibleforviralreplication.3.Normal, functionalADAallele foundonchromosome20 is isolatedfromahealthy individual,andtheninsertedintotheviralvector.4.Recombinantretrovirusisusedtoinfecthaematopoieticstemcells*fromthebonemarrow,whichwouldhavebeenremovedpreviouslyfromthepatientwithSCID.5. The retrovirus enters the cell and releases the normal, functional ADA allele which is then reverse-transcribed toadouble strandedDNAbyreverse transcriptaseandDNApolymerase,and integrated intothehostcell’schromosomebyintegrase.6. Cells are grown inculture to screen for correct ADA gene expression, tumour formation, or presence ofinfectiousvirusesbeforebeingreintroducedbackintotheSCIDpatient.7.Thesestemcellsdevelopintonormal,functioningBandTlymphocytes.
3 EX-VIVOTargetcellsareeasilyremovedfrombloodandreturned.Ex-vivo approach allows screening of recombinant cells before reintroduction; this allowsmonitoring ofgeneexpression,andscreeningforwrongly-insertedgenewhichcouldcauseinsertionalmutagenesis.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
11. Describecysticfibrosisanditsgenetherapytreatment.[6m]
1 CYSTICFIBROSISCysticfibrosisisaresultofanautosomaldeletionmutation*of3nucleotides*onexon10ofchromosome7*,resultinginapolypeptidemissingaphenylalanine*.Thegenethatismutatedcodesforcystic fibrosis transmembraneconductanceregulator (CFTR)*thatisresponsiblefortransportofCl-acrosstheepithelialcellmembrane.[ThelossofthephenylalanineaminoacidchangestheprimarystructureoftheCFTRprotein.Theaminoacidisno longerable to formbonds involvedinmaintenanceofthesecondaryand tertiary structuresoftheprotein.Thisresults inachange in theoverall 3D conformationof theCFTRprotein,particularly theATP-bindingdomain. Itnowhasan inability to bind toATPandhence,noopeningof the channel protein’sporeforCl-todiffuseout.Failure to transportCl- results in retentionofNa+.Water potential in thecellbecomesmore negative andconsequently,thereisretentionofwater.Mucus lining inthelumenthenbecomesundiluted,resultinginthickmucus inthelungsandpancreaticduct.Asitaccumulates,itreducesgaseousexchange,causingseverebreathingdifficulties.Bacteriagrowthleadstolungcongestionandinfections,possiblyafatalcondition.Blockedpancreaticductspreventthereleaseofenzymes,leadingtochronicindigestion.Oneofthesymptomsthatisusedtodiagnosetheconditioniscopiussaltysweat.
2 CYSTICFIBROSISTREATMENT1.Treatmentmayinvolvetheuseofanon-viralgenedeliverysystem,employingthein-vivo*approach.2.Liposometransfection*maybeused.3.Normal,functionalCFTRallelefoundonchromosome7isisolatedfromahealthyindividual.4. It is thenencapsulatedbyaphospholipidbilayer spherecalleda liposome that is thenpackagedinanaerosoldispenser.5. The aerosol dispenser delivers aerosol of liposomes into the lungs of patients, and the liposomemembrane fuseswith themembraneof lungepithelial cells,deliveringthenormal, functionalalleleintoaffectedcells.6.TheCFTRgenemaybetranscribedand translated,andtheCFTRmaybecorrectlyincorporatedon theplasmamembraneoftheepithelialcells.7.Cl-maymoveout,andmayallowwatertomoveoutviaosmosis,dilutingthemucus.Lackof integrationofnormal,functionalalleleusingthenon-viralgenedeliverysystemresultsintransient,short-livedtherapy.Thisissuitable,astargetlungepithelialcellsarereplacedfrequently.Repeatedtherapyisneeded.
3 TRIVIAGut cellswith a single copyof themutant allele takeup80% fewerSalmonellatyphi thannormal cells. Theselectiveadvantageoftheheterozygotespreventtheselectivepressureagainstindividualswithcysticfibrosisfromremovingtherecessiveallelefromthepopulation.
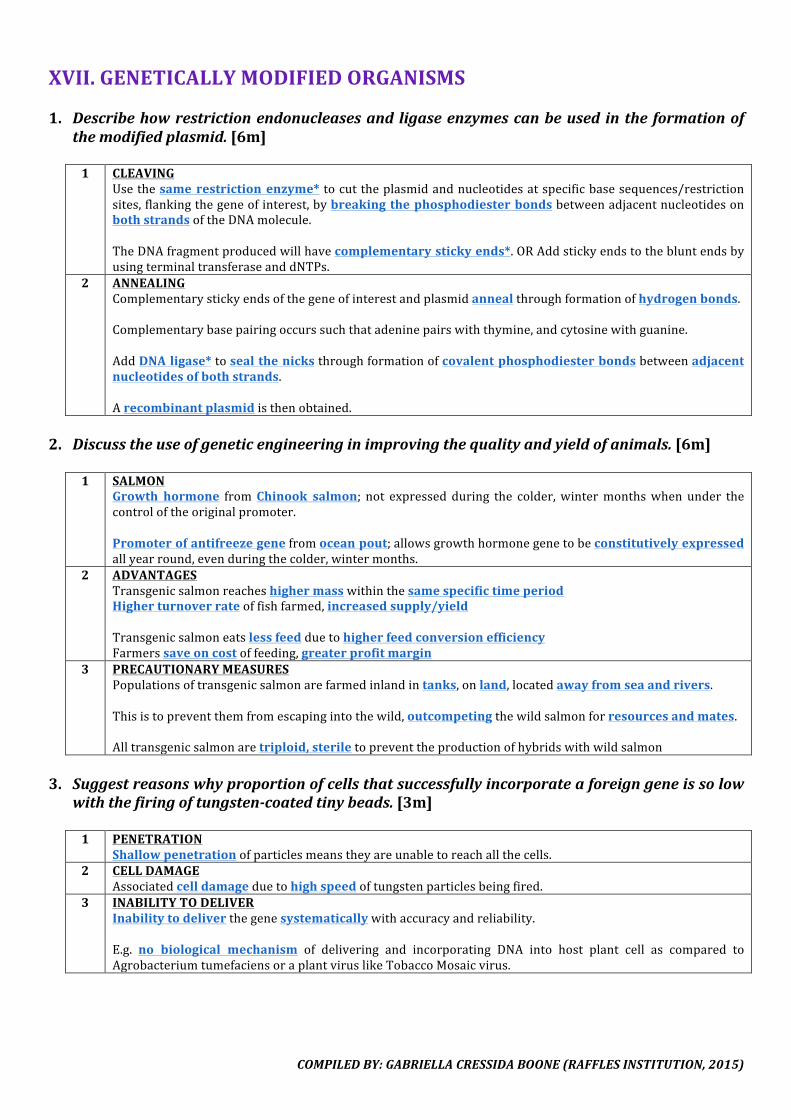
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
XVII.GENETICALLYMODIFIEDORGANISMS1. Describehowrestrictionendonucleasesand ligaseenzymescanbeused inthe formationof
themodifiedplasmid.[6m]1 CLEAVING
Usethesamerestrictionenzyme* tocuttheplasmidandnucleotidesatspecificbasesequences/restrictionsites,flankingthegeneofinterest,bybreakingthephosphodiesterbondsbetweenadjacentnucleotidesonbothstrandsoftheDNAmolecule.TheDNAfragmentproducedwillhavecomplementarystickyends*.ORAddstickyendstothebluntendsbyusingterminaltransferaseanddNTPs.
2 ANNEALINGComplementarystickyendsofthegeneofinterestandplasmidannealthroughformationofhydrogenbonds.Complementarybasepairingoccurssuchthatadeninepairswiththymine,andcytosinewithguanine.AddDNAligase*tosealthenicksthroughformationofcovalentphosphodiesterbondsbetweenadjacentnucleotidesofbothstrands.Arecombinantplasmidisthenobtained.
2. Discusstheuseofgeneticengineeringinimprovingthequalityandyieldofanimals.[6m]
1 SALMONGrowth hormone fromChinook salmon; not expressed during the colder,wintermonthswhenunder thecontroloftheoriginalpromoter.Promoterofantifreezegenefromoceanpout;allowsgrowthhormonegenetobeconstitutivelyexpressedallyearround,evenduringthecolder,wintermonths.
2 ADVANTAGESTransgenicsalmonreacheshighermasswithinthesamespecifictimeperiodHigherturnoverrateoffishfarmed,increasedsupply/yieldTransgenicsalmoneatslessfeedduetohigherfeedconversionefficiencyFarmerssaveoncostoffeeding,greaterprofitmargin
3 PRECAUTIONARYMEASURESPopulationsoftransgenicsalmonarefarmedinlandintanks,onland,locatedawayfromseaandrivers.Thisistopreventthemfromescapingintothewild,outcompetingthewildsalmonforresourcesandmates.Alltransgenicsalmonaretriploid,steriletopreventtheproductionofhybridswithwildsalmon
3. Suggestreasonswhyproportionofcellsthatsuccessfullyincorporateaforeigngeneissolow
withthefiringoftungsten-coatedtinybeads.[3m]1 PENETRATION
Shallowpenetrationofparticlesmeanstheyareunabletoreachallthecells.2 CELLDAMAGE
Associatedcelldamageduetohighspeedoftungstenparticlesbeingfired.3 INABILITYTODELIVER
Inabilitytodeliverthegenesystematicallywithaccuracyandreliability.E.g. no biological mechanism of delivering and incorporating DNA into host plant cell as compared toAgrobacteriumtumefaciensoraplantviruslikeTobaccoMosaicvirus.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
4. 2012Y6CT1Discusstheuseofgeneticengineeringinimprovingthequalityandyieldofcropplants.[6m]1 YIELD–PEST-RESISTANTBTCORN
Insect pests like theEuropean corn borer feeds on the corn plant, destroying the plant parts leading todetrimentaleffectsonplantgrowthandhence,yieldofcorn.Using the Agrobacterium vector, containingmodified Ti plasmid with theBt toxin gene from Bacillusthuringiensis,cornisgeneticallyengineered.ThetransgeniccornexpressestheBttoxingenetoproducetoxinthatkillsinsectpestswhentheyfeedonit.Thetransgeniccorncropsarethereforenotdamagedasbadlybyinsectpests,henceincreasingyield.Additionally,thereisnoneedtosprayinsecticides,andfarmerscansavecosts,higherprofitfromhigheryieldThereisnodangerofinsecticiderunoffwhichcouldleadtobioaccumulationandaffectnon-targetorganisms.
2 YIELD–HERBICIDE-RESISTANTROUNDUPREADYSOYBEANHerbicides tendtobebroad-spectrum intheireffectsandwouldharmboththeweedsandthecropplants,leadingtolossofyield.UsingtheAgrobacteriumvector,containingmodifiedTiplasmidwiththeherbicide/glyphosate-resistantgenefromagrobacterium,soybeanisgeneticallyengineered.Thetransgenicsoybeancropsarethusnotaffectedbyherbicidecontainingglyphosate,andtheherbicidecanbefreelyusedtoeliminateweeds.Soybeancropnolongerhavetocompetewithweedfornutrients,henceincreasingyield.LIMITATIONSResistance gene may be passed on to nearby weed relatives through cross-pollination/horizontal genetransfer,creatingasuperweed.Createalargebufferzonebyplantingunrelatedcrops.Intense use of same herbicide will eventually select for resistant weeds, and Round-Up will no longer beeffective.AlternateuseofRound-Upwithothertypesofherbicide.Excessiveuseofherbicidemaycauserunoffswhicharetoxictofishandamphibians.Promoteresponsibleuseofherbicideamongfarmers.
3 QUALITY–NUTRITIONALGOLDENRICENon-transgenicrice ispoor inmicronutrients like vitamin A.This isbecauseenzymes required for thesynthesisofbeta-carotene,aprecursorofvitaminA,arenotexpressedintheendospermofriceplants.UsingtheAgrobacteriumvector,containingmodifiedTiplasmidwiththephytoenesynthase(psy)gene*fromdaffodils andcarotene desaturase (crtl) gene* frombacteriaErwiniauredovora, rice is geneticallyengineered.The transgenic rice can therefore produce beta-carotene, a precursor of vitamin A, in its endosperm, andeventuallysynthesisevitaminA,increasingthenutritionalqualityofrice,whichmaypreventmalnutritionandnightblindness.
4 QUALITY–FULLERFLAVOUREDFLAVRSAVRTOMATOESNormaltomatoestendtoripenrapidlyandsoften,leadingtomushyfruitsandconsequentdamageandlossofquality.UsingtheAgrobacteriumvector,containingmodifiedTiplasmidwiththecomplementaryDNAsequenceforpolygalacturonase(PG)enzyme,tomatoisgeneticallyengineered.ComplementaryPGDNAsequenceproducesantisensemRNA,whichbindstothenormalmRNAtranscriptforPG,sotranslationofpolygalactonurasemRNAintotheenzymeisprevented.Since polygalacturonase is responsible for ripening of fruits, the transgenic tomatoes undergo delayedripening.Theyripenonthevine,thusproducingfullerflavouredfruit.Theyalsohavealongershelf-lifeandareabletobeshippedoverlongerdistanceswithoutdeteriorationinquality.
5 GENERAL

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
GMcrops likedrought-resistant transgenicwheat cangrowin less favourable environments, increasingarableland.
XVIII.PLANTCLONING1. Explainwhyplantsderivedfromatissueculturearedescribedasclones.Describetheprocess
ofcloningofplantsfromtissueculture.[8m]
2012Y6CT1Outline and explain one named method of tissue culture that can produce disease-freeplantletsfromavirusinfectedplant.[6m]
2013Y6CT1RoundupReadysoybeanisatransgeniccropthatcarriestheherbicideresistantgene.Giventhe recombinant plasmid that carries the herbicide resistant gene, describe tissue culturetechnique(s)involvedintheproductionofthetransgenicplant.[9m]
1 CLONES
They are derived fromone parent*. They are products ofmitotic division*, and are thereforegeneticallyidentical*totheparentduetosemi-conservativeDNAreplication*.
2 TISSUECULTURE–EXPLANT1.Usingaseptictechniques,chooseandexciseapicalmeristems*,likeroot/shoottips.Theseexplantsfromyoungpartsofplantsaredisease-freeandwillallowforpropagationofdisease-freeplants.[Apicalmeristems have inhibitors of viral replication, not present in othermeristems. Also, thevascularbundleswhichusuallytransmitdiseasesarenotyetestablishedatstemtips,butarepresentinleavesandflowers.]2.Surfacesterilise*theexplantswithdilutesodiumhypochlorite/bleach.Thiswillkillbacteriaandfungiwhichmaycontaminateanddepletenutrientsfromtheculture.
3 TISSUECULTURE–ASEPTICTRANSFER3. Cut explants into small fragments and transfer explants aseptically to sterile agar medium [withnutrients].4.Useplantgrowthregulators*suchasauxins*andcytokinins*inintermediateamounts.Thisstimulatesmitosis*forcallusformation*.5. Agar medium contains nutrients. The sugars are for energy as callus cells cannot photosynthesise, thenitratesandphosphatesforproteinandnucleicacidsynthesis,themagnesiumforchlorophyllproduction.
4 TRANSFORMATION–SOYBEANTransform Agrobacterium tumefaciens withmodified Ti plasmid containing the herbicide/glyphosate-resistantgene,usingCaCl2andheatshock*.ThetransformedAgrobacteriaisusedtoinfect*andtransfertherecombinantplasmidtothecalluscells.Calluscellsaregrownonagarplatecontainingherbicideforscreening,andonlytransformedcellsthatcarrytherecombinantplasmidwiththeherbicide-resistantgenecansurviveandareselected.
5 TISSUECULTURE–SUBCULTURING6.Thenumberofcallicanbeincreasedbysubculturing*.Itinvolvessubdividingthecallusandgrowingtheminseparateculturevessels.Thisenablesthemassproductionofgeneticallyidentical*clonesbecausecalluscellsdivideviamitosis*andtheirDNAundergoessemi-conservativereplication*.
6 TISSUECULTURE–PLANTLETFORMATION7. Callus cells are totipotent* and can be induced to undergoorganogenesis* with varying ratios of plantgrowthregulatorsthatallowthemtodifferentiate intoplantlets.Calluscellsaretransferredtotheanothernutrient medium with: (1) high cytokinin:auxin ratio to induce shoot formation, and then to (2) highauxin:cytokininratiotoinducerootformation.8.Plantletsarethensoakedinfungicideandgrowninsterilesoil,exposedtoincreasinglightintensityand

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
lowering humidity. This to acclimatise* the plantlets to farming conditions and reduce transplantingshock.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
2. Explaintheadvantagesofcallusandtissuecultureovermoretraditionalmethodsofcloningplants,suchastakingcuttingsorgrafting.[4m]
1.Calluscellshavethinnercellwalls,makingiteasierforDNAtopenetrateandenterthenucleus,geneticmodificationcanbecarriedoutviagenetransfertoprotoplast/callus.2.Calluscellsdividerapidly,soalargernumberofthegenetically identicalplantscanbeproducedinthesameamountoftime.3.Calluscanbesubculturedtogiverisetomorecallusandultimately,moreplants,allowingmassproduction.4.Lessspaceisrequiredforproduction.5.Cultureiseasiertotransport,withreferencetoairfreight,ascomparedtoadultplants.6.Culturingisnotlimitedbyexternalconditionssuchasseasons.7.Calluscellsaretotipotent*andcanbeinducedtoundergoorganogenesistogrowintoanentireplant.
3. Identifydifferentingredientsofgrowthmediaandwhateachisusedfor.[4m]
INGREDIENT USE1 CARBONSOURCE/GLUCOSE Energysource,respiratorysubstrate2 PHOSPHATE Nucleicacidsynthesis,ATPsynthesis3 NITRATES Nucleicacidsynthesis4 SULFUR Aminoacidsynthesis5 AMINOACIDS Proteinsynthesis6 TRACEELEMENTSe.g.Fe/Zn Enzymeactivators/cofactors7 VITAMINS Enzymeactivators/cofactors8 MAGNESIUM Chlorophyllsynthesis9 CALCIUM Cellwallformation(middlelamella)10 GIBBERELLIN Cellelongation
4. Explain briefly why different tissue culture techniques are used in the propagation of two
agriculturalplants.[8m]
1 PROTOPLASTCULTURE(e.g.tomato,tobacco)Inprotoplastculture,thecellwallisremovedwithenzymescellulaseandpectinase.Withoutthecellwall,itiseasiertointroduceforeignDNA/plasmidcontaininggeneofinterestintotheplantcellofcrops.Protoplast fusion fromdifferent species results in ahybridwith desirable traits frombothparents.ORProtoplastfusioncangiverisetopolyploidplantswithincreasedvigourorsizeoftheplants.[Note: it ismore susceptible tomicrobial contamination. It also needs to bemaintained in a solution of thesamewaterpotentialtopreventlysis.]
2 MERISTEMATICTISSUECULTURE(e.g.soyabean,potatoes)Inmeristematicculture,explantsareobtainedfromfastgrowingsectionsliketheshoot tip/root tip/apicaldomethatcontainthemeristematictissue.Theyareusuallyfreefromvirusesandpathogens,thusallowingthepropagationofdisease-free*plants.
3 EMBRYOCULTURE(e.g.peaches)Inembryoculture,embryosareremovedandcultured.Itallowspropagationofhybridsthatfail todevelopnormally,orfail toovercomeseeddormancywhereseedscannotorhavedifficultygerminating.
4 ANTHERCULTURE(e.g.rice,maize)Inantherculture,antherscontaininghaploidpollengrainsarecultured.Pollengrainsdevelopintohaploidplantlets.Sincetheyonlyhaveoneallele/onecopyofeachgene,theycanbeeasilyselectedforfavourabletraitsduetonon-maskingofphenotypes.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
5 SUSPENSIONCULTUREToextractusefulsecondarymetabolites.
XIX.DIVERSITY&EVOLUTION1. Describehowclassificationdiffersfromphylogeny.[7m]
POINTOF
COMPARISONCLASSIFICATION
(LINNAEANTAXONOMY)PHYLOGENY
1* GROUPINGOFORGANISMS
Basedonoverall/morphologicalsimilaritiesanddoesnottakeintoaccountevolutionaryhistoryoforganisms
Tracesevolutionaryhistorybasedonancestor-descendentrelationship/evolutionaryrelationshipSpeciesmustbeabletointerbreedtoformfertile,viableoffsprings
2* SYSTEMOFORGANISINGORGANISMS
HierarchicalclassificationbasedonevolutionaryrelationshipsderivedfromphylogenyHierarchicalclassification*namingsystem,groupingorganismsinto:kingdom,phylum,class,order,family,genusandspecies
Arrangedbasedonevolutionaryrelationshipwitheachother,eachorganismassignedapositiononabranchingtreerelativetootherorganisms,relatedorganismsinthesamebranch
3* HOWSPECIESAREPRESENTED
Binomialnomenclature* Phylogenetictree*
4 NATUREOFCHARACTERISTICS
Doesnotdiscriminatebetweenanalogousorhomologouscharacters
Homologouscharacters*thatarederivedfromacommonancestor
5 TYPESOFCHARACTERSUSED
Onlymorphologicalcharacteristics Morphological,anatomical,embryological,andmolecularcharacteristicssuchasDNA,RNA,aminoacidsequences
6 STRENGTHS Easycategorisationandnaming Rarelyclassifieswronglyduetoextensivesourcesdifferentiatingspecies
7 WEAKNESSES Maywronglyclassifyorganismsthatarenotrelatedbutlooksimilarduetoconvergentevolution*Unabletoinferevolutionaryrelationshipbetweenmembersofcategoryorhowdistantlyrelatedtheyaretooneanother
Cannotimmediatelyplaceanorganismintothephylogramasevolutionaryhistoryneedtoestablishedfrommultiplesources
8* INFERENCEOFSPECIATIONEVENTS
Doesnotallowinferenceofhistoricalspeciationevents
Indicatesspeciationeventsasnodesonthephylogenetictree
9*
INFERENCEOFCOMMONANCESTORS
Doesnotallowinferenceofcommonancestors
Allowsinferenceofcommonancestors,descendantsarerepresentedinthesamebranch
10* INFERENCEOFRELATIONSHIPS
Doesnotallowinferenceofhowclosely2speciesaresincetheyaregroupedtogetherinthesamehierarchy
Allowsaccurateinferenceofhowcloselyrelated2speciesarebyidentifyingbranchpointandlookingathowrecentlytheydivergedfromtheircommonancestor
11 APPLICATIONOFMOLECULARCLOCK
Notpossibletoapplymolecularclocktodatespeciationevents
Possibletoapplywithmolecularevidence,timeofspeciationcanbeinferred
2. Calculatetherateofmutation.[1m]
𝑁𝑢𝑚𝑏𝑒𝑟 𝑜𝑓 𝑠𝑢𝑏𝑠𝑡𝑖𝑡𝑢𝑡𝑖𝑜𝑛𝑠𝑁𝑢𝑚𝑏𝑒𝑟 𝑜𝑓 𝑙𝑖𝑛𝑒𝑎𝑔𝑒 × 𝑛𝑢𝑚𝑏𝑒𝑟 𝑜𝑓 𝑦𝑒𝑎𝑟𝑠

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
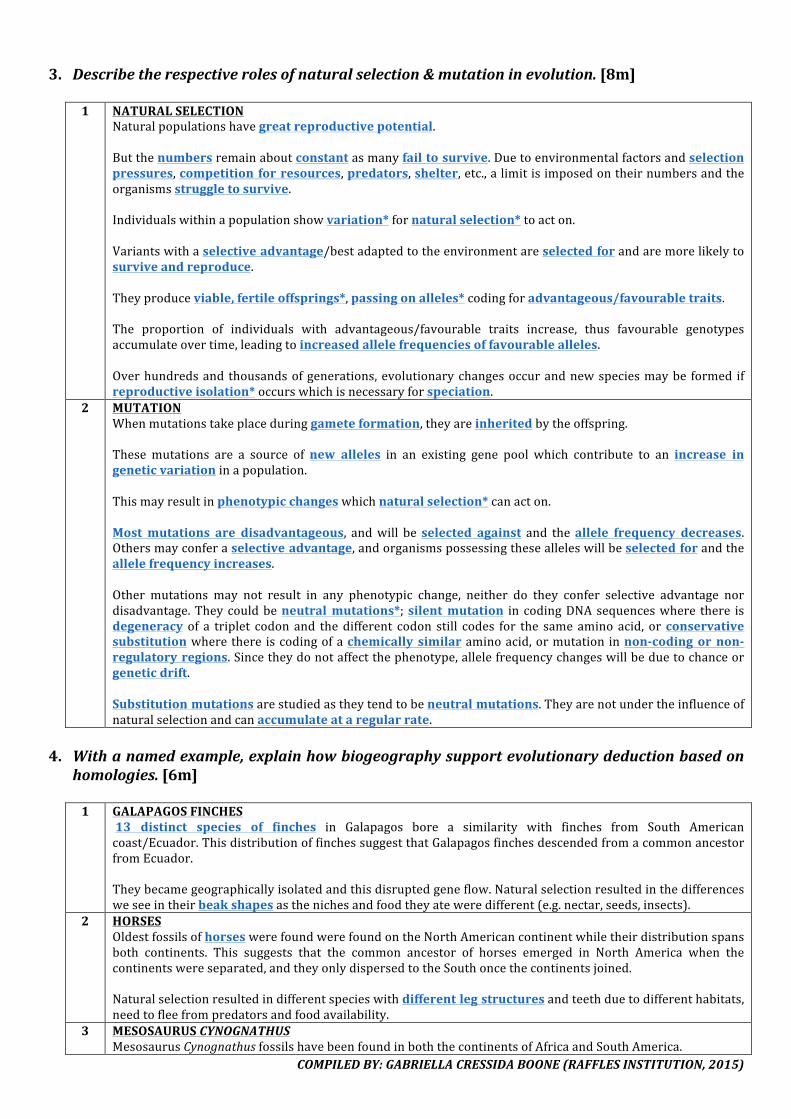
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. Describetherespectiverolesofnaturalselection&mutationinevolution.[8m]1 NATURALSELECTION
Naturalpopulationshavegreatreproductivepotential.Butthenumbersremainaboutconstantasmanyfailtosurvive.Duetoenvironmentalfactorsandselectionpressures,competition forresources,predators,shelter,etc.,alimitisimposedontheirnumbersandtheorganismsstruggletosurvive.Individualswithinapopulationshowvariation*fornaturalselection*toacton.Variantswithaselectiveadvantage/bestadaptedtotheenvironmentareselectedforandaremorelikelytosurviveandreproduce.Theyproduceviable,fertileoffsprings*,passingonalleles*codingforadvantageous/favourabletraits.The proportion of individuals with advantageous/favourable traits increase, thus favourable genotypesaccumulateovertime,leadingtoincreasedallelefrequenciesoffavourablealleles.Overhundredsand thousandsofgenerations,evolutionarychangesoccurandnewspeciesmaybe formed ifreproductiveisolation*occurswhichisnecessaryforspeciation.
2 MUTATIONWhenmutationstakeplaceduringgameteformation,theyareinheritedbytheoffspring.These mutations are a source of new alleles in an existing gene pool which contribute to an increase ingeneticvariationinapopulation.Thismayresultinphenotypicchangeswhichnaturalselection*canacton.Mostmutations are disadvantageous, andwill beselected against and theallele frequency decreases.Othersmayconferaselectiveadvantage,andorganismspossessingthesealleleswillbeselectedforandtheallelefrequencyincreases.Other mutations may not result in any phenotypic change, neither do they confer selective advantage nordisadvantage.Theycouldbeneutralmutations*;silentmutation in codingDNAsequenceswhere there isdegeneracy of a triplet codonand thedifferent codon still codes for the sameamino acid, orconservativesubstitutionwherethere iscodingofachemically similaraminoacid,ormutation innon-codingornon-regulatoryregions.Sincetheydonotaffectthephenotype,allelefrequencychangeswillbeduetochanceorgeneticdrift.Substitutionmutationsarestudiedastheytendtobeneutralmutations.Theyarenotundertheinfluenceofnaturalselectionandcanaccumulateataregularrate.
4. Withanamedexample,explainhowbiogeographysupportevolutionarydeductionbasedon
homologies.[6m]
1 GALAPAGOSFINCHES13 distinct species of finches in Galapagos bore a similarity with finches from South Americancoast/Ecuador.ThisdistributionoffinchessuggestthatGalapagosfinchesdescendedfromacommonancestorfromEcuador.Theybecamegeographicallyisolatedandthisdisruptedgeneflow.Naturalselectionresultedinthedifferencesweseeintheirbeakshapesasthenichesandfoodtheyateweredifferent(e.g.nectar,seeds,insects).
2 HORSESOldestfossilsofhorseswerefoundwerefoundontheNorthAmericancontinentwhiletheirdistributionspansboth continents. This suggests that the common ancestor of horses emerged in North America when thecontinentswereseparated,andtheyonlydispersedtotheSouthoncethecontinentsjoined.Naturalselectionresultedindifferentspecieswithdifferentlegstructuresandteethduetodifferenthabitats,needtofleefrompredatorsandfoodavailability.
3 MESOSAURUSCYNOGNATHUSMesosaurusCynognathusfossilshavebeenfoundinboththecontinentsofAfricaandSouthAmerica.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
4 OTHERSHaemoglobin;whaleflipper+batwing+birdwing
5. Using named examples, explain how anatomical, embryological and molecular homologysupportDarwin’stheoryofevolutionbynaturalselection.[12m]
1 BASISOFHOMOLOGY
1.Verydifferentorganismsshareanunderlyingsimilaritybecauseoftheirsharedcommonancestry,calledhomology.2. These characteristics present in an ancestral organismdeveloped into different forms as the result ofnaturalselection,astheyfaceddifferentenvironmentalconditions(descentwithmodification).
2 ANATOMICALHOMOLOGY1.Anatomicalhomologyreferstosimilaritiesinthebonearrangement*ofdifferentspecies.2.Anexampleisthepentadactyl(five-digit)limb*structureoftetrapods,4-limbedanimals,likeamphibians,reptiles,birdsandmammals.2.Theforelimbsofhumans,cats,batsandwhalesshowthesamearrangementofbonesfromtheshouldertothetipsofthedigits,eventhoughtheseappendagesdonotresembleeachotherandhaveverydifferentfunctions:lifting/grasping,walking,flyingandswimming.3. Natural selection had resulted in the different forms of pentadactyl limb to suit their specialisedenvironmentsandfunctions.4.And that itwasamodification of the five-digit forelimbof thecommon ancestorwhich theydescendedfrom.
3 EMBRYOLOGICALHOMOLOGY1. Embryological homology refers to similarity in embryonic development* of different species such ashumans,cats,fishesandturtles.2.Thelonger theembryologicaldevelopmentremainssimilar,themorecloselyrelatedtheorganismsare.3. At the early stage of embryological development, all vertebrates have gill slits, long bony tail, twochamberedheartandnotochord.4. However, these gill slits ultimately develop into structures with very different functions, such as gills infishesandpartsoftheearsandthroatinhumansandothermammals.5.Basicembryologicalbodyplanintheancestralspecieswasmodifiedinthedifferentdescendantspeciesastheselectionpressureschangesduetodifferencesinhabitats.
4 MOLECULARHOMOLOGY1.MolecularhomologyreferstosimilarityinDNA,RNA&aminoacidsequences*,suchasthecytochromecgeneorthep53protein.2.Thesearetypically importantenoughthatevery organismpossesses themandtheycarryoutthesamefunction in different organisms.Theirsequences are significantlyconserved suggestingsharedcommonancestry.3.Anancestralgenenucleotidesequencewouldbemodifiedovermanygenerationsduetoaccumulationofmutations.4. Homologous protein provides direct correlation to the relatedness of the species from which they arederived–sequencesofcloselyrelatedspeciestendtobemoresimilartoeachother.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
6. Explainhownaturalselectionorislandscanresultintheformationof(i)newspeciesand(ii)sub-species.[6m]
1 GEOGRAPHICALISOLATION
Physicalbarriersresultingeographical isolation*thatprevents inter-breeding,resultingindisruptionofgeneflow*.(i) Differing habitats present many niches for the species to fill, exposing them to different selectionpressures*suchaspredation,habitats,andnaturalselection*actsonthem.Adaptiveradiation*occurred.(ii) Divided populations are exposed to different environments and are under different selectionpressures*suchaspredation,habitats,availablefoodsources,andnaturalselection*actsonthem.
2 VARIATION&ALLELEFREQUENCIESVariation* exists in populations, and those best adapted to the local conditions due tofavourable/advantageoustraitsareselectedfor,andaremorelikelytosurvive,reproduceandoffspringinherittheiralleles*.Allelefrequencieschangebecauseofnaturalselection*andgeneticdrift*.
3 SPECIES/SUB-SPECIESOver hundreds and thousands of generations, sub-populations evolve independently and accumulatedifferentgeneticmutations.(i)Hence, theybecomereproductively isolated* andcanno longer interbreed* toproduceviable, fertileoffspring*.Thisgivesrisetoanewspeciesthroughallopatricspeciation*.(ii)Hence,theydevelopdistinctphenotypicdifferenceswithintheirsub-populationsandformsub-species.
7. Describe how bacteria evolve through natural selection and the implication in antibiotic
resistance.[8m]
1 NATURALSELECTIONDue to spontaneous mutation, there is existing variation* in the population of bacteria with antibiotic-resistantstrainsandnon-resistantstrains.Thealleleforantibioticresistancecanalsobeacquiredthroughconjugation/transduction/transformation(horizontalgenetransfer)(refertoBACTERIA–Q5).When exposed to antibiotic, antibiotic acts as a selection pressure* to select against the non-resistancebacteria.Thosewiththeantibioticresistancealleleareselectedforandsurvive.Theypass on the allele for antibiotic resistance to subsequent generations of bacteria cells during binaryfission,thusmostofthebacteriaareresistanttoantibiotic(verticalgenetransfer).Allelefrequencieschangeovertimeasaresultofnaturalselection.
2 ANTIBIOTICRESISTANCEOvermany generations, bacteria cells evolve by natural selection and the bacteria become resistance to theantibiotic,renderingtheantibioticineffective.Thus, other types/different combinations of antibiotics have to be used, but this could lead to bacteriadevelopingmultipleantibioticresistance.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
8. Explaintheadvantagesofmolecularmethods,andspecifically,cytochromebinmitochondrialDNA,indeterminingphylogeny.[6m]
1 MOLECULARMETHODS
Theyareobjective*. MolecularcharacterstatesareunambiguousasnitrogenousbasesA,T,C,Gareeasilyrecognisableandcannotbeconfused.Theyarequantitative*.Moleculardataareeasily converted tonumerical form andhenceareamenable tomathematicalandstatisticalanalysisandhence,computation.Thedegreeofrelatednesscanbeinferredandquantifiedbycalculatingthenucleotidedifferencesbetweenspecies.All known life is based on nucleic acids* and would have DNA. Remotely related organisms can becomparedbecausetheysharesomeproteinssuchascytochromec.The nucleotide data can be used to compare specieswhich aremorphologically indistinguishable due toconvergentevolution.
2 CYTOCHROMEB–ADVANTAGESCytochromebisahomologousgene*foundinthemitochondrialDNA.Itisconservedinallspeciesbeingcompared,foundincommonancestorsandthusformsabasisofcomparison.Itdoesnot undergo recombinationasmitochondrialDNAisderiveddirectly fromthematernal line.AnychangestotheDNAissolelyduetoaccumulationofmutationsovertimeataregularrate.Thismakesitanidealcandidateforthemolecularclock*.However,itstillhasafastermutationratethannuclearDNAandso,hassufficient differences/variability todistinguish theclosely related species*, anddeterminewhenthedifferentkingdomsoflifedivergedfromeachother.
3 CYTOCHROMEB–MUTATIONSThemutationsarebasesubstitutions.TheDNAmaybedamagedby reactiveoxygen radicalsgeneratedinthemitochondrion.ThemitochondriaDNArepairmechanismsmaynotberobusttoo.
4 CYTOCHROMEB–NEUTRALTHEORYCytochromebisanevidencefortheneutraltheory.Sinceitschangesoccurinalinearfashion,itgivesrisetoaconstantrateofaccumulationofmutations.Itsmutationsarealsoselectivelyneutral,sincetheyconfernoselectivedisadvantageoradvantageontheorganism’sphenotype.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
9. Explainhowgelelectrophoresiscanbeusedtoprovideevidenceofmolecularhomology.[8m]1 TECHNIQUE
DNA*fromvarious/differentorganisms*beingcomparedareextractedusingabufferandpurified.Using specific forward and reverse primers that hybridise to the 3’ flanking regions of a potentialhomologousgene,polymerasechainreaction*isthencarriedout.Theamplifiedfragmentisthencutusingarestrictionenzyme*,e.g.EcoRI.TheDNAisloadedintoanagarosegel*andsubjectedtoanelectricfield;DNAbeingnegatively-charged,willmigratetowardsthepositiveelectrode.Themeshworkofpolymerfibersimpedethemovementoflongerfragmentsmorethanshorterfragments;fragmentsarethusseparatedbasedontheirsize,with larger fragmentsmigrating furtheralongthegel thanshorterfragments.The DNA fragments are stained with ethidium bromide, a DNA binding dye, causing them to fluoresceunderUVlight.
2 ANALYSISThePCRprimerswouldhybridisetoregionsflankingthehomologousgeneitwastargetingandamplify thegene.ORThePCRprimerswouldnotamplifythegeneiftherewerenocomplementarysequencesitcouldbindto.If therewas indeedmolecular homology amongst theorganismsbeingcompared, thegelwould reveal thesamebandingpatternforalltheorganismscompared.ORIfanyoneormoreofthelanesdidnothaveanybands,thereisnomolecularhomology.Once it is established that the different organisms show molecular homology, their DNA can then besequenced todeterminethedegreeof nucleotide similarity,withtheDNAfromcloselyrelatedorganismsbeingthemostsimilar.
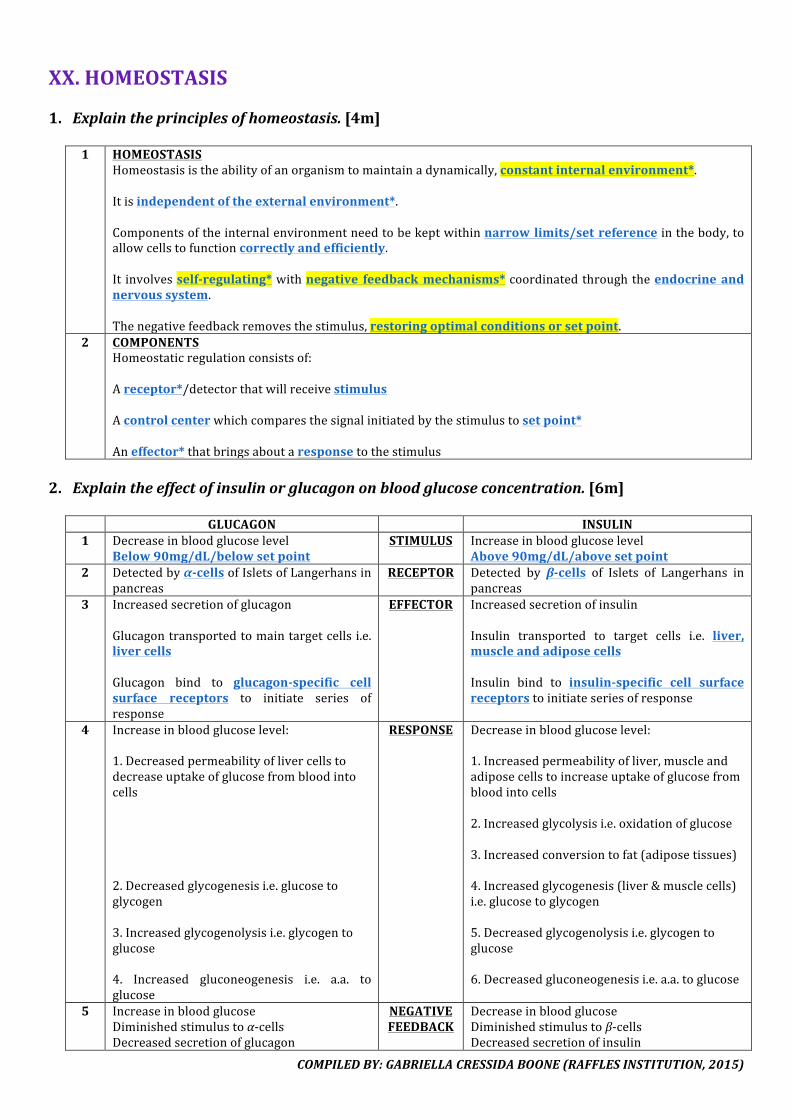
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
XX.HOMEOSTASIS1. Explaintheprinciplesofhomeostasis.[4m]
1 HOMEOSTASIS
Homeostasisistheabilityofanorganismtomaintainadynamically,constantinternalenvironment*.Itisindependentoftheexternalenvironment*.Componentsoftheinternalenvironmentneedtobekeptwithinnarrowlimits/setreferenceinthebody,toallowcellstofunctioncorrectlyandefficiently.It involvesself-regulating*withnegative feedbackmechanisms*coordinatedthroughtheendocrineandnervoussystem.Thenegativefeedbackremovesthestimulus,restoringoptimalconditionsorsetpoint.
2 COMPONENTSHomeostaticregulationconsistsof:Areceptor*/detectorthatwillreceivestimulusAcontrolcenterwhichcomparesthesignalinitiatedbythestimulustosetpoint*Aneffector*thatbringsaboutaresponsetothestimulus
2. Explaintheeffectofinsulinorglucagononbloodglucoseconcentration.[6m]
GLUCAGON INSULIN1 Decreaseinbloodglucoselevel
Below90mg/dL/belowsetpointSTIMULUS Increaseinbloodglucoselevel
Above90mg/dL/abovesetpoint2 Detectedbyα-cellsofIsletsofLangerhansin
pancreasRECEPTOR Detected by β-cells of Islets of Langerhans in
pancreas3 Increasedsecretionofglucagon
Glucagontransportedtomaintargetcellsi.e.livercellsGlucagon bind to glucagon-specific cellsurface receptors to initiate series ofresponse
EFFECTOR IncreasedsecretionofinsulinInsulin transported to target cells i.e. liver,muscleandadiposecellsInsulin bind to insulin-specific cell surfacereceptorstoinitiateseriesofresponse
4 Increaseinbloodglucoselevel:1.Decreasedpermeabilityoflivercellstodecreaseuptakeofglucosefrombloodintocells2.Decreasedglycogenesisi.e.glucosetoglycogen3.Increasedglycogenolysisi.e.glycogentoglucose4. Increased gluconeogenesis i.e. a.a. toglucose
RESPONSE Decreaseinbloodglucoselevel:1.Increasedpermeabilityofliver,muscleandadiposecellstoincreaseuptakeofglucosefrombloodintocells2.Increasedglycolysisi.e.oxidationofglucose3.Increasedconversiontofat(adiposetissues)4.Increasedglycogenesis(liver&musclecells)i.e.glucosetoglycogen5.Decreasedglycogenolysisi.e.glycogentoglucose6.Decreasedgluconeogenesisi.e.a.a.toglucose
5 IncreaseinbloodglucoseDiminishedstimulustoα-cellsDecreasedsecretionofglucagon
NEGATIVEFEEDBACK
DecreaseinbloodglucoseDiminishedstimulustoβ-cellsDecreasedsecretionofinsulin

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
Returntosetpoint Returntosetpoint
XXI.CELLSIGNALING1. Outlinethemainstagesofcellsignaling.Explainhowthestructureofareceptorallowsitto
carryoutitsfunction.[6m]1 LIGAND-RECEPTORINTERACTION
Signal molecule/ligand complementary in conformation* binds to ligand-binding site on extracellulardomainofaspecificreceptor*.
2 SIGNALTRANSDUCTIONBindingofsignalmoleculecausesintracellulardomain*toundergoconformationalchange*.PHOSPHORYLATIONCASCADESignalcascadeoftenmediatedbykinasesandphosphorylasesActivatedkinasephosphorylatesandactivatesarelayprotein,usuallyanotherkinaseActivatedphosphatasedephosphorylatesandinactivatesarelayprotein
SIGNALAMPLIFICATIONOccursviasignalingcascadeEachstepyieldsanumberofactivatedproductthatismorethanaprecedingstep
3 CELLULARRESPONSETransducedsignaltriggersaspecificcellularresponse:Catalysisbyanenzyme(e.g.glycogenphosphorylasebreakingdownglucagontoglucose)Physicalresponse(e.g.rearrangementofcytoskeleton)Activationofspecificgenesinnucleus
4 STRUCTUREOFRECEPTORAreceptorisatransmembraneproteinwithanextracellulardomainthatbindstoaligand.Ithasaregionthatspansthehydrophobiccoreofthemembrane,consistingofaminoacidswithnon-polarRgroups, enabling hydrophobic interaction with the non-polar hydrocarbon tails of the phospholipidsmakingupthemembrane,andtobeembeddedwithinthemembrane.Polar/hydrophilic R groups of the amino acids interact with the hydrophilic phosphate head of thephospholipidbilayerandaqueousenvironment.
2. Describetheadvantagesofacellsignalingpathway.[5m]
1 SIGNALAMPLIFICATIONFacilitatessignalamplification*,onlyasmallnumberofsignalmoleculesisneededtoelicitalargecellularresponse.Phosphorylationcascadehappenswhenasequential activationofkinases is initiatedbyactivePKA.Thenumberofactivatedproduct/kinasesisalwaysgreaterthaninprecedingsteps.
2 MULTIPLECHECKPOINTSProvidesmultiplecheckpointsforregulation,cellularresponsecanberegulatedandcontrolledateachstep.
3 SPECIFICITYEnsuresspecificityasaspecificsignalmoleculebindstoaspecificreceptor,toelicitaspecificreactionviaaspecificpathwayineachcelltype.
4 MULTIPLERESPONSESMultiple responses to 1 signal molecule as 1 signal molecule can triggermultiple signal transductionpathwaystoelicitdifferentresponses.Thisisduetotheabilityofthesignalmoleculetoactivatemanydifferentcellssimultaneouslytoo.
5 ACTIVATEGENESINNUCLEUSAbilityofsignalmoleculetoactivategenesinnucleusuponbindingtocellsurfacereceptorwithouttheneed
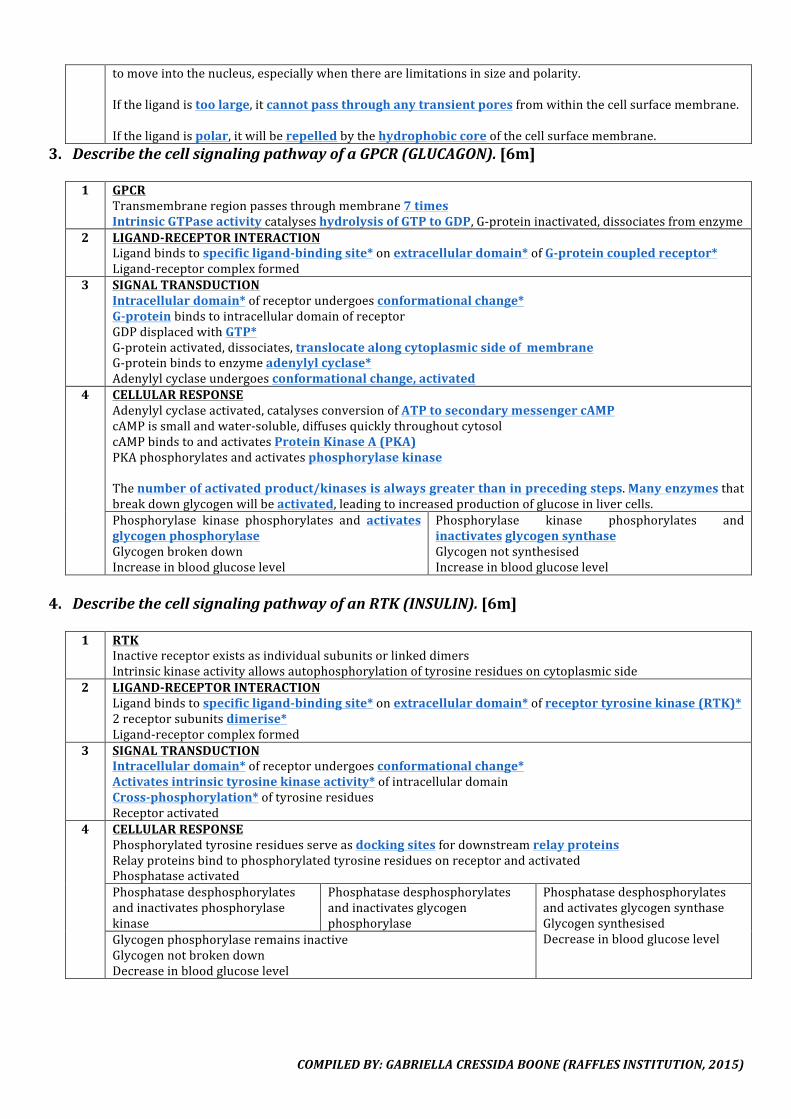
COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
tomoveintothenucleus,especiallywhentherearelimitationsinsizeandpolarity.Iftheligandistoolarge,itcannotpassthroughanytransientporesfromwithinthecellsurfacemembrane.Iftheligandispolar,itwillberepelledbythehydrophobiccoreofthecellsurfacemembrane.
3. DescribethecellsignalingpathwayofaGPCR(GLUCAGON).[6m]1 GPCR
Transmembraneregionpassesthroughmembrane7timesIntrinsicGTPaseactivitycatalyseshydrolysisofGTPtoGDP,G-proteininactivated,dissociatesfromenzyme
2 LIGAND-RECEPTORINTERACTIONLigandbindstospecificligand-bindingsite*onextracellulardomain*ofG-proteincoupledreceptor*Ligand-receptorcomplexformed
3 SIGNALTRANSDUCTIONIntracellulardomain*ofreceptorundergoesconformationalchange*G-proteinbindstointracellulardomainofreceptorGDPdisplacedwithGTP*G-proteinactivated,dissociates,translocatealongcytoplasmicsideofmembraneG-proteinbindstoenzymeadenylylcyclase*Adenylylcyclaseundergoesconformationalchange,activated
4 CELLULARRESPONSEAdenylylcyclaseactivated,catalysesconversionofATPtosecondarymessengercAMPcAMPissmallandwater-soluble,diffusesquicklythroughoutcytosolcAMPbindstoandactivatesProteinKinaseA(PKA)PKAphosphorylatesandactivatesphosphorylasekinaseThenumberofactivatedproduct/kinasesisalwaysgreaterthaninprecedingsteps.Manyenzymesthatbreakdownglycogenwillbeactivated,leadingtoincreasedproductionofglucoseinlivercells.Phosphorylase kinase phosphorylates and activatesglycogenphosphorylaseGlycogenbrokendownIncreaseinbloodglucoselevel
Phosphorylase kinase phosphorylates andinactivatesglycogensynthaseGlycogennotsynthesisedIncreaseinbloodglucoselevel
4. DescribethecellsignalingpathwayofanRTK(INSULIN).[6m]
1 RTKInactivereceptorexistsasindividualsubunitsorlinkeddimersIntrinsickinaseactivityallowsautophosphorylationoftyrosineresiduesoncytoplasmicside
2 LIGAND-RECEPTORINTERACTIONLigandbindstospecificligand-bindingsite*onextracellulardomain*ofreceptortyrosinekinase(RTK)*2receptorsubunitsdimerise*Ligand-receptorcomplexformed
3 SIGNALTRANSDUCTIONIntracellulardomain*ofreceptorundergoesconformationalchange*Activatesintrinsictyrosinekinaseactivity*ofintracellulardomainCross-phosphorylation*oftyrosineresiduesReceptoractivated
4 CELLULARRESPONSEPhosphorylatedtyrosineresiduesserveasdockingsitesfordownstreamrelayproteinsRelayproteinsbindtophosphorylatedtyrosineresiduesonreceptorandactivatedPhosphataseactivatedPhosphatasedesphosphorylatesandinactivatesphosphorylasekinase
Phosphatasedesphosphorylatesandinactivatesglycogenphosphorylase
PhosphatasedesphosphorylatesandactivatesglycogensynthaseGlycogensynthesisedDecreaseinbloodglucoselevelGlycogenphosphorylaseremainsinactive
GlycogennotbrokendownDecreaseinbloodglucoselevel

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
5. Describehowcellularresponsescanbeterminatedandtheimportanceofsignaltermination.[6m]
1 SIGNALTERMINATION
Signal termination allows the cell torespond quickly tonew ligands released as a result of new stimuli. Itprevents thepathwayfrombeingactive longerthannecessary, leadingto inappropriateorexcessivecellularresponse(e.g.uncontrolledcellgrowth).Acellsignalingpathwayhascheckpointsforregulation.
2 CHECKPOINT1–RECEPTIONRegulationcanoccuratreception.Degradationofligandbyligand-degradingenzyme.Endocytosisofcellsurfacereceptortopreventligand-receptorinteraction.Endocytosisofligand-receptorcomplextopreventsignaltransduction.Inhibitorthatbindstoreceptortopreventligand-receptorbinding.Inhibitor that binds to intracellular domain of ligand-receptor complex or other downstream relayproteinstopreventsignaltransduction.
3 CHECKPOINT2–PHOSPHORYLATIONCASCADERegulationcanoccurduringphosphorylationcascade.Removalofphosphategroupfromactivatedrelayproteinbyphosphatasetoinactivaterelayproteinandpreventfurthersignaltransduction.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
XXII.NERVOUSSYSTEM1. Describehowrestingmembranepotential(-70mV)ismaintainedacrossmembranes.[4m]
1 LEAKCHANNELS
Na+ and K+ leak channels* allow these ions to diffuse down their concentration gradient across themembrane.More K+ leak channels than Na+ leak channels are present, so there ismore K+ leaving the cell than Na+entering.Thereisanetdiffusionofpositiveionsoutofthemembrane,andanetnegativeinterior.
2 Na+-K+PUMPThroughactivetransport*,theNa+-K+pump*usesATP*topumpionsagainsttheconcentrationgradient*.3Na+arepumpedout* forevery2K+pumped in*tomaintaintheconcentrationofhighNa+outsideandhighK+insidethecell,andthusanetnegativeinterior.
3 ORGANICANIONSLargeorganicanions*suchasproteinsandnucleic acidswithinthecellcontributetothenegativechargeinsidethecell.
2. ExplaintheroleofNa+andK+,themyelinsheathandacetylcholineinimpulsetransmission.
[6m]
1 MOVEMENTThesechargedionsmovethroughspecificionchannelsembeddedinthemembraneofthenervecells,suchasleakchannels, voltage-gated ionchannels, and ligand-gated ionchannels,by facilitated diffusion*, downanelectrochemicalgradient*.They also move through ion pumps where ions are actively transported* against their concentrationgradient*withtheuseofATP*.
2 Na+Whenthemembraneisstimulatedanddepolarised to thresholdpotential*,voltage-gatedNa+ channels*ontheneuronopentoallowaninfluxofNa+*intotheneuron.Themembranedepolarisesfrom-70mVto+40mV,generatinganactionpotential.Na+intheaxonspreadsaslocalcircuits*toadjacentnodesofRanvier*toinitiateanotheractionpotentialatthenextregion.
3 K+When themembrane is stimulated anddepolarised to threshold potential*,voltage-gated K+ channels*willslowlyopentoallowaneffluxofK+*outoftheneuron.Thisresultsinrepolarisation*ofmembranetorestoretherestingmembranepotential.
4 MYELINSHEATHThemyelinsheathcontainshigh levelof lipidsthatwraparoundtheaxon,toactasanelectrical insulatoragainstthemovementofNa+andK+acrossit.Thus,action potentials canonlybegeneratedat thenodes of Ranvier*,where therearehighdensitiesofvoltage-gatedNa+andK+channels,andareexposedtotheextracellularfluid.Localcircuit*producedbyactionpotentialinonenode,travelstothenextnode.Hence, the action potential “jumps” from node to node, enabling impulses to spread by saltatorypropagation*alongtheaxon,whichspeedsuptheimpulsetransmission.
5 ACETYLCHOLINEAcetylcholineincreasestheselectivepermeabilityofthecellmembrane.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
3. Statethedifferencebetweenagradedpotentialandanactionpotential.[2m]
1 GRADEDPOTENTIALWhendepolarisationisbelowthresholdpotential,onlyagradedpotentialisproduced.Noactionpotentialisproduced,andthusnoimpulseispropagatedtotheadjacentregion.
2 ACTIONPOTENTIALWhendepolarisationisexceedsthresholdpotential,anactionpotentialisproduced.Voltage-gatedNa+channels*intheaxonmembraneopentoallowaninfluxofNa+*intotheneuron.Animpulseisthuspropagatedtotheadjacentregion.
4. Describethetransmissionofanactionpotentialalongamyelinatedneuron.[8m]
1 RESTINGPOTENTIAL
Theneuronhasarestingmembranepotentialof -70mV*,largelydeterminedbyitshighpermeabilitytoK+ionsandmaintainedbythesodium-potassiumpumpwhichisinvolvedinthemovementofNa+andK+ions.
2 DEPOLARISATIONWhenastimulusarrivesatthefirstnodeofRanvierandisabovethethresholdpotential*,ittriggerstheopeningofthevoltage-gatedNa+channels*atthenode.InfluxofNa+* intotheneuronleadstoarapiddepolarisation*of themembraneforthegenerationof anactionpotentialatthenode.
3 REPOLARISATIONAt thepeak of the action potential at+40mV*, voltage-gated Na+ channels will close, while thevoltage-gatedK+channelswillopen.EffluxofK+*outoftheneuronleadstorepolarisation*ofthemembrane.
4 HYPERPOLARISATIONAsthevoltage-gatedK+channelsareslowtoclose,thereishyperpolarisation*ofthemembrane.
5 RESTINGPOTENTIALWhenthevoltage-gatedK+channelshaveclosed,restingpotentialisrestoredagain.
6 UNIDIRECTIONALTheNa+ influxalsocreateslocalcircuits*ofNa+whichdiffusesidewayswithintheaxoplasm,drivenbyitselectrochemicalgradient.AlthoughlocalcurrentsofNa+ionscantravelupstream,thevoltage-gatedNa+channelsinthepreviousnodeofRanvierwouldbeintherefractoryperiod*,sotheywillnotrespondtothestimulus.
7 PROPAGATIONDiffusionofNa+downtheaxoncausesdepolarisationthatisabovethethresholdpotentialatthenextnodeofRanvier,causingthevoltage-gatedNa+channelsinthenextnodetoopenandgenerateanactionpotential.Thisresultsinpropagationofactionpotentials*alongthemyelinatedaxonoftheneuron.Thepresenceofmyelinatedsheathspeedsupthetransmissionofimpulsebysaltatorypropagation*.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
5. Describe how a nerve impulse is transmitted across the synapse. Explain why synaptictransmissionisrelativelyslowerandonlytravelsinonedirection.[8m]
1 PRE-SYNAPTICMEMBRANE
Whentheactionpotentialreachesthepresynapticknob,thereisdepolarisationofthemembranethatleadstoinfluxofCa2+ions*.Suddenrise inCa2+concentrationstimulatessynaptic vesicles*carryingacetylcholine(neurotransmitter)tomigrateandfusewiththepresynapticmembrane*.Acetylcholineisreleasedintothesynapticcleftbyexocytosis*.
2 SYNAPTICCLEFTAcetylcholine*moleculesdiffuseacrossthesynapticcleft*andbindstotheligand-gatedNa+channels*onthepost-synapticmembrane*ofanotherneuron.
3 POST-SYNAPTICMEMBRANELigand-gatedNa+channelsopen*onthepost-synapticmembrane.InfluxofNa+ions*depolarises*thepost-synapticmembrane.Ifthedepolarisationexceedsthethresholdpotential*,anactionpotentialisgenerated.Thedepolarisationtoreachthresholdpotentialcanbeduetothetemporalorspatialsummation*ofseverallocalised depolarisation, since the post-synaptic neuron can be receiving several impulses from differentsynapses.Intensityofstimulusispassedasfrequency*ofactionpotentials/impulses.Theacetylcholineishydrolysedbyacetylcholinesterase*intoacetateandcholine.
4 RELATIVELYSLOWERSPEEDSynaptictransmissioninvolvesthediffusionofneurotransmitters*acrossthesynapticcleftwhichisslowcomparedtotherapidsaltatorypropagationduringthedepolarisationofnerveimpulses.Thediffusionofneurotransmittersisdrivenonlybytheirconcentrationgradient,whilethemovementof Na+ during electrical transmission is driven by both its concentration gradient and electrostaticattractionwithintheaxoplasm.
5 UNIDIRECTIONALCa2+andsynapticvesicles*containingneurotransmittersareonlyfoundinthepresynapticknob.Receptors/ligand-gated Na+ ion channels* for neurotransmitters are only found on the post-synapticmembrane.
6. Suggestwhateffectatoxinwillhaveonthenervoussystemifitsmodeofactionisinhibiting
acetylcholinesterase.[3m]
1 INHIBITIONInhibitedacetylcholinesterasecannotbreakdowntheneurotransmitteracetylcholine.Acetylcholineaccumulatesandremainsboundtothepost-synapticreceptors.
2 EFFECTLigand-gatedNa+ionchannelsremainopen*,sothereiscontinuousdepolarisation*ofthepost-synapticmembrane*,andrepeatedgenerationofactionpotentials*.Repeatedactionpotentialscauseparalysisinthevictim.

COMPILEDBY:GABRIELLACRESSIDABOONE(RAFFLESINSTITUTION,2015)
7. Explaintheneedformitochondriaalongtheaxonintermsofnerveimpulseconduction.[3m]
1 ATPMitochondriaiswhereaerobicrespirationoccurstoproduceATP*.
2 Na+-K+PUMPATP is required for active transport*, where the Na+-K+ pump* pump ions against the concentrationgradient*.3Na+arepumpedout*forevery2K+pumpedin*tomaintaintheconcentrationofhighNa+outsideandhighK+ insidethecell,andthusanetnegative interior,whichistherestingmembranepotentialsothatthenextactionpotentialcanbegenerated.
3 SYNAPTICVESICLESATPisalsorequiredbymicrotubulestotransportthesynapticvesiclessynthesisedatthecellbodyalongtheaxon,tothesynapticterminalsattheendoftheaxon.
8. State the differences between the depolarisation of the pre-synaptic and post-synaptic
membrane.[3m]
POINTOFCOMPARISON
PRE-SYNAPTICMEMBRANE POST-SYNAPTICMEMBRANE
1 TYPEOFSTIMULUS
Electrical/localcircuits Chemicalintheformofneurotransmitters
2 TIMEWHENDEPOLARISATIONOCCURS
Earlier Shortdelayafterdepolarisationatpresynapticmembrane
3 SHARPNESSOFPEAK
Sharperpeak Lesssharppeak
4 CHANGEINMAGNITUDEOFPOTENTIALDIFFERENCE
Largerchangeinpotentialdifference Smallerchangeinpotentialdifference
5 DURATIONOFDEPOLARISATION
Shorter Longer


















