
Biochemistry 201 Biological Regulatory Mechanisms...
26
1 Biochemistry 201 Biological Regulatory Mechanisms Lecturer: Geeta Narlikar February 2, 2015 Chromatin Structure and Its Regulation-1 Key Points 1. Packaging of eukaryotic DNA into nucleosomes and higher order folded states provides a means to functionally compartmentalize genes. The DNA in nucleosomes is tightly bound but very dynamic, unpeeling and rebinding on the order of milliseconds. 2. The accessibility of DNA within a nucleosome is very sensitive to its location and can be reduced relative to free DNA from 10-fold to 500,000 fold. 3. The intrinsic preferences of DNA for histone octamers can play a role in positioning nucleosomes in vivo but there is more regulation of nucleosome positions than just sequence 4. There are at least two ways in which arrays of chromatin can fold into the 30 nm fiber. This process is likely regulated by other proteins so that the chromatin fiber may adopt additional packing conformations in vivo. References BOOKS: Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K. & Walter, P. Molecular Biology of the Cell (5 th ed.) Garland Publishing Inc., New York, Chapter 4. ARTICLES: Nucleosome Structure and Dynamics 1. Luger, K., et al., Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature, 1997. 389(6648): p. 251-60. 2. Li, G., et al., Rapid spontaneous accessibility of nucleosomal DNA. Nat Struct Mol Biol, 2005. 12(1): p. 46-53. 3. Polach, K.J. and J. Widom, Mechanism of protein access to specific DNA sequences in chromatin: a dynamic equilibrium model for gene regulation. J Mol Biol, 1995. 254(2): p. 130-49.
Transcript of Biochemistry 201 Biological Regulatory Mechanisms...

1
Biochemistry 201 Biological Regulatory Mechanisms Lecturer: Geeta Narlikar February 2, 2015 Chromatin Structure and Its Regulation-1 Key Points 1. Packaging of eukaryotic DNA into nucleosomes and higher order folded states provides a means to functionally compartmentalize genes. The DNA in nucleosomes is tightly bound but very dynamic, unpeeling and rebinding on the order of milliseconds. 2. The accessibility of DNA within a nucleosome is very sensitive to its location and can be reduced relative to free DNA from 10-fold to 500,000 fold. 3. The intrinsic preferences of DNA for histone octamers can play a role in positioning nucleosomes in vivo but there is more regulation of nucleosome positions than just sequence 4. There are at least two ways in which arrays of chromatin can fold into the 30 nm fiber. This process is likely regulated by other proteins so that the chromatin fiber may adopt additional packing conformations in vivo. References BOOKS: Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K. & Walter, P. Molecular Biology of the Cell (5th ed.) Garland Publishing Inc., New York, Chapter 4. ARTICLES:
Nucleosome Structure and Dynamics 1. Luger, K., et al., Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature, 1997. 389(6648): p. 251-60. 2. Li, G., et al., Rapid spontaneous accessibility of nucleosomal DNA. Nat Struct Mol Biol, 2005. 12(1): p. 46-53. 3. Polach, K.J. and J. Widom, Mechanism of protein access to specific DNA sequences in chromatin: a dynamic equilibrium model for gene
regulation. J Mol Biol, 1995. 254(2): p. 130-49.

2
In vivo mapping of nucleosome positions and occupancy 1. Kaplan N et al. The DNA-encoded nucleosome organization of a eukaryotic genome. Nature. 2009 458:362-6. 2. Zhang Y et al. Intrinsic histone-DNA interactions are not the major determinant of nucleosome positions in vivo. Nat Struct Mol Biol.
2009;16:847-52. 3. Albert I et al. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature. 2007
446:572-6. 4. Zhang Z, Pugh BF. High-resolution genome-wide mapping of the primary structure of chromatin. Cell. 2011; 144:175-86. 5. Kaplan N et al. Nucleosome sequence preferences influence in vivo nucleosome organization.Nat Struct Mol Biol. 2010; 17:918-20 6. Zhang et al. Evidence against a genomic code for nucleosome positioning, Nat Struct Mol Biol. 2010;17: 920-22 7. Pugh BF. A preoccupied position on nucleosomes. Nat Struct Mol Biol. 2010; 17:923 8. Sasaki S et al. Chromatin-associated periodicity in genetic variation downstream of transcriptional start sites.Science. 2009 323: 401-4. 9. Floer M et al. A RSC/nucleosome complex determines chromatin architecture and facilitates activator binding. Cell. 2010;141:407-18. 10. Zhang Z,et al. A packing mechanism for nucleosome organization reconstituted across a eukaryotic genome. Science. 2011;332:977-80. 11. Vera D L et al. Differential Nuclease Sensitivity Profiling of Chromatin Reveals Biochemical Footprints Coupled to Gene Expression and Functional DNA Elements in Maize. Plant Cell 2014;26:3883-3893 Higher Order Chromatin folding 1. Tremethick DJ. Higher-order structures of chromatin: the elusive 30 nm fiber. Cell. 2007. 128:651-4.
2. Bassett A, et al. The folding and unfolding of eukaryotic chromatin. Curr Opin Genet Dev. 2009 19:159-65. 3. Dorigo B, et al. Nucleosome arrays reveal the two-start organization of the chromatin fiber. Science. 2004. 306:1571-3. 4. Schalch, T., et al., X-ray structure of a tetranucleosome and its implications for the chromatin fibre. Nature, 2005. 436(7047): p. 138-41. 5. Routh A, Sandin S and Rhodes D, Nucleosome repeat length and linker histone stoichiometry determine chromatin fiber structure. Proc Natl Acad Sci. 2008 105:8872-7. 6. Horn, P.J., et al., The SIN domain of the histone octamer is essential for intramolecular folding of nucleosomal arrays. Nat Struct Biol,
2002. 9(3): p. 167-71.

Regulating the DNA Template

Organized nature of eukaryo1c DNA observed more than a 100 years ago
1882: Salivary gland cell from Bloodworm(?) larvae observed by Walther Flemming
Lighter stains = euchroma?n Darker stains = heterochroma?n

A few decades ago: Drosophila salivary glands polytene chromosomes stained with similar dyes
1882: Salivary gland cell from Bloodworm(?) larvae observed by Walther Flemming
Lighter stains = euchroma?n Darker stains = heterochroma?n
‘puffs” associated with transcrip?on suggested major changes in chroma?n organiza?on
Outline
Func1ons of Chroma1n
Packing material Complex regulatory plaCorm
Replica1on Transcrip1on RNA processing
coordina1on/coupling
Outline
Func1ons of Chroma1n
Packing material Complex regulatory plaCorm
Replica1on Transcrip1on RNA processing
coordina1on/coupling
Intrinsic proper1es Regula1on of intrinsic proper1es
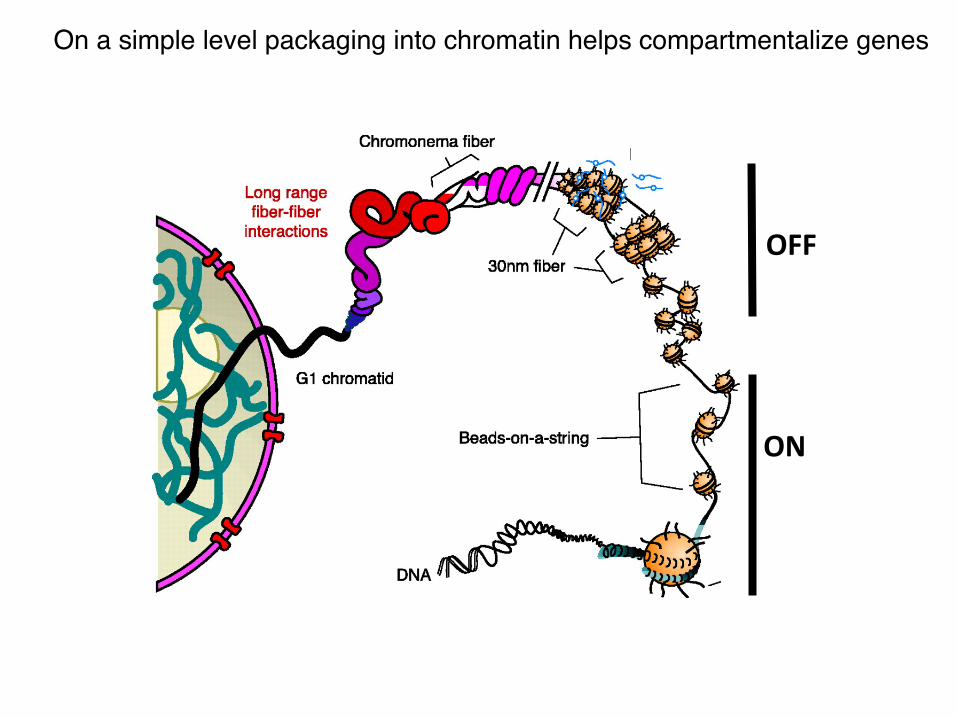
ON
OFF
On a simple level packaging into chromatin helps compartmentalize genes !

A closer look at the nucleosome
• 4 histones: 2 copies of each form an octamer that wraps 147 bp of DNA. • N-‐terminal tails of histones extend out (~ 1/4th the histone mass). • DNA is highly bent. Histone-‐DNA interac?ons stabilize this unfavorable conforma?on. • DNA becomes less accessible to DNA binding proteins • Almost all the histone-‐DNA contacts are sequence independent. Most interac?ons are between the -‐vely charged phosphate backbone of DNA and +vely charged histone residues. Yet some DNA sequences can bind histone octamers 100-‐fold more strongly than others. Why?

DNA sequence can affect energe?cs of DNA bending
minor groove compressed
major groove compressed
GC base-‐pairs more easily allow major groove compression
5 bp
5 bp
Proper placement of AT and GC base-‐pairs can give DNA sequences with high histone octamer binding affini?es rela?ve to bulk genomic DNA This pa_ern is seen in naturally occurring “nucleosome posi?oning sequences”
AT base-‐pairs more easily allow minor groove compression

~ 4 s-‐1
~ 90 s-‐1
Nucleosomal DNA is intrinsically dynamic
closed open
Above numbers are for a site that is ~10 bp in from one end
Keq= 1/20 = [open]/[closed]
closed open Keq= 1/20
+ +
Kd= 10 nM
Overall Kd for nucleosomal site = 10/(1/20) = 200 nM
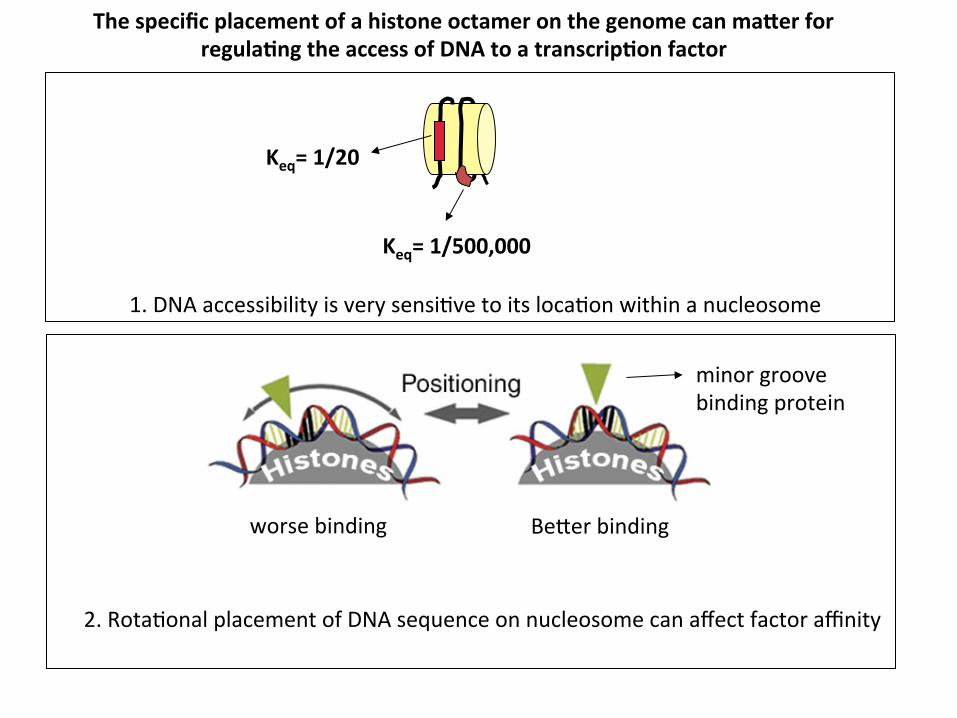
Keq= 1/20
Keq= 1/500,000
The specific placement of a histone octamer on the genome can maUer for regula1ng the access of DNA to a transcrip1on factor
1. DNA accessibility is very sensi?ve to its loca?on within a nucleosome
minor groove binding protein
Be_er binding worse binding
2. Rota?onal placement of DNA sequence on nucleosome can affect factor affinity

Coopera?ve binding through protein-‐protein interac?ons
1. A B A and B directly interact

The architecture of the nucleosome can give coopera?ve binding behavior for non-‐interac?ng proteins
+ +
1.
2.
A B
+ A B
A and B directly interact
A and B do not directly interact

Debate in the field: does the genome encode its own packaging?
Placement of AT and GC base-‐pairs every 10 bp can increase histone octamer binding affini?es rela?ve to bulk genomic DNA The specific transla?onal and rota?onal posi?on of nucleosomes can have a large impact on accessibility of regulatory sequences

Do intrinsic proper?es of DNA sequences play a significant role in posi?oning nucleosomes in vivo?
In vitro
purified yeast DNA + purified histones
assemble into nucleosomes
map loca?ons by MNase and deep sequencing or micro-‐array
In vivo
compare to in vivo posi?ons

Comparison of in vivo and in vitro maps in yeast suggested in vivo role for DNA sequence preferences
Average occupancy at each base-‐pair
number of reads that cover the bp
average number of reads per bp across genome
=
Kaplan et al. Nature. 2009 458:362-‐6.

But, it can’t be that simple…
1. How can one nucleosome map explain mul?ple transcrip?onal states? If nucleosome placement affects transcrip?on factor binding and transcrip?on factor binding regulates expression of specific genes, then when gene expression pa_erns change upon changes in environment, differen?a?on etc., the nucleosome posi?ons and occupancy should change as well.
2. What happens to nucleosome posi?ons and occupancy in the coding region when gene expression levels change?

The devil is in the details and the differences may reveal more than the similari1es
1. Correla?on between in vivo and in vitro maps is greater in non-‐promoter intergenic regions (R=0.83) and lower in promoter and coding regions (0.69) 2. At coding regions deple?on levels of nucleosomes in vivo rela?ve to in vitro increases with the expression levels of the associated genes 3. Both maps show nucleosome deple?on at transcrip?on start sites, but level of deple?on in vivo is greater 4. Level of nucleosome deple?on around transcrip?on factor binding sites generally correlates between the two maps but there are interes?ng differences.
Nucleosome occupancy near different transcrip1on factor binding sites

Nucleosome Occupancy vs. Posi1oning
Pugh BF. Nat Struct Mol Biol. 2010; 17:923
(Example: # reads)
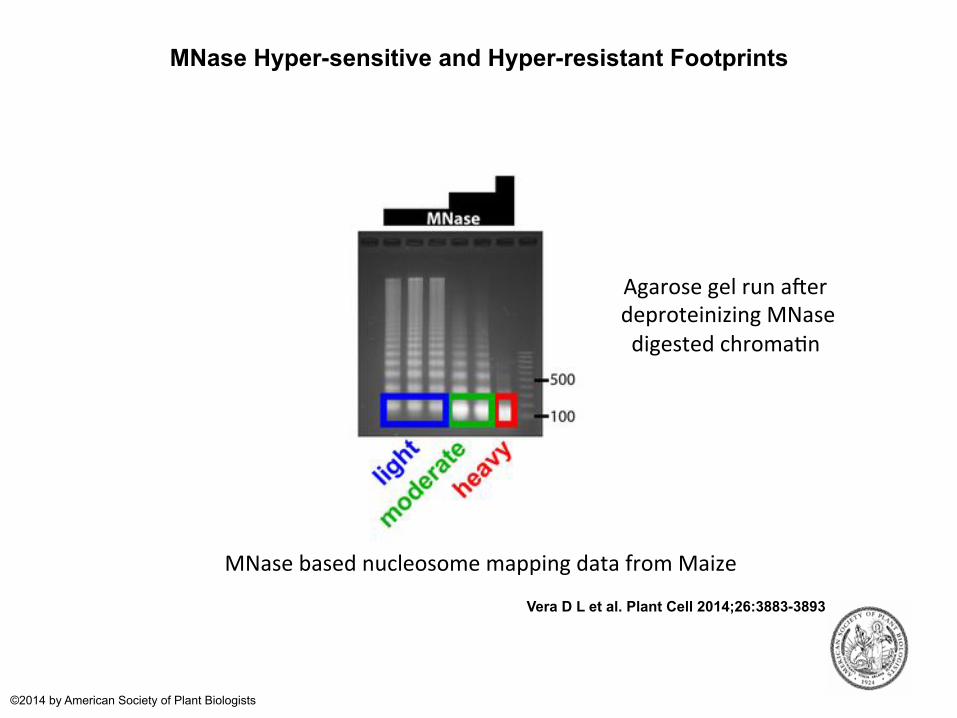
MNase Hyper-sensitive and Hyper-resistant Footprints
Vera D L et al. Plant Cell 2014;26:3883-3893
©2014 by American Society of Plant Biologists
MNase based nucleosome mapping data from Maize
Agarose gel run aqer deproteinizing MNase digested chroma?n

Vera D L et al. Plant Cell 2014;26:3883-3893
©2014 by American Society of Plant Biologists
MNase based nucleosome mapping data from Maize
MNase-Hypersensitive Footprints Correlate with regulatory elements like transcription factor binding sites

Higher order compac1on of chroma1n
One start Two start
30 nm Electron micrographs

Regula?on of higher order chroma?n folding
1. Short inter-‐nucleosomal spacing (<20 bp) may favor two-‐start structure while longer spacing may favor one start structure
EM micrographs
Is this process regulated by non-‐histone proteins in vivo? Perhaps chroma?n fiber adopts addi?onal packing conforma?ons in vivo.

H2A-‐H2B acidic patch
viral LANA pep?de bound to acidic patch
H4 N-‐terminal tail bound to acidic patch
Histone tails mediate inter-‐nucleosomal contacts through electrosta1c interac1ons
Highly basic histone tails interact with DNA of neighboring nucleosomes
Histone H4 tail interacts with an acidic patch formed by H2A-‐H2B
Kaposi's sarcoma–associated herpesvirus latency-‐associated nuclear an?gen (LANA) mediates viral genome a_achment to mito?c chromosomes by binding nucleosomes through H2A-‐H2B acidic patch

Linker histones (Histone H1) promote chroma1n folding


















