
Bio Computing
31
Modeling Biochemical Systems using Molecular Orbital Calculations Prof. H. Bernhard Schlegel Department of Chemistry Wayne State University Current Research Group Hrant Hratchian John Knox Stan Smith Smriti
Transcript of Bio Computing

Modeling Biochemical Systems using Molecular Orbital Calculations
Prof. H. Bernhard SchlegelDepartment of ChemistryWayne State University
Current Research Group
Hrant Hratchian John KnoxStan Smith Smriti AnandJie Li

Research overview
With molecular orbital calculations, it is possible to investigate details of chemical reactions and molecular properties that are often difficult to study experimentally
Our group is involved in both the development and the application of new methods in ab initio molecular orbital (MO) methods.
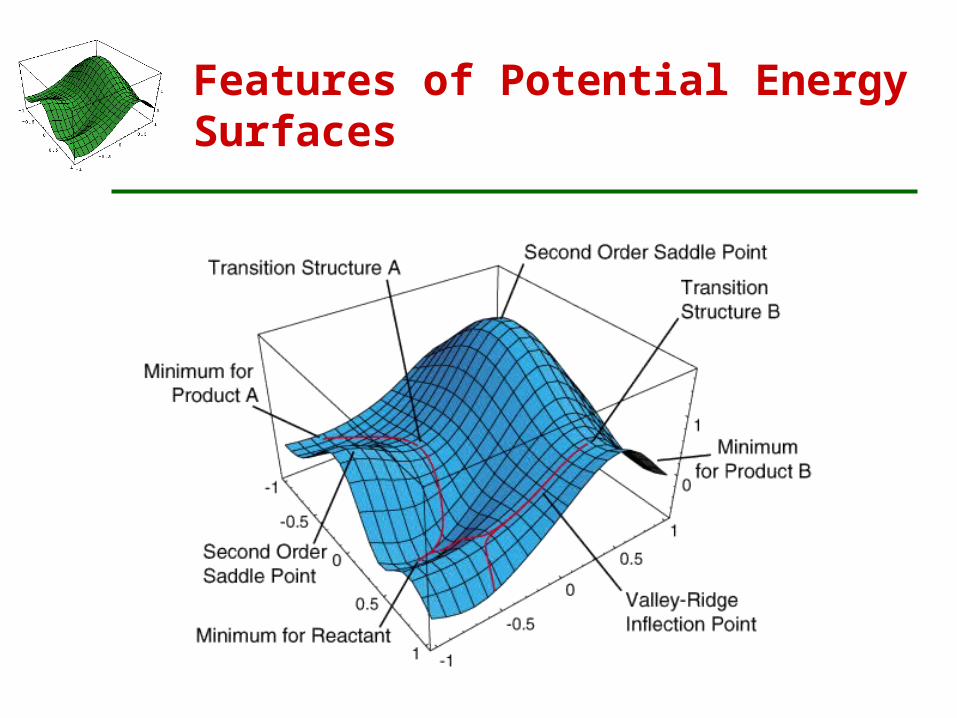
Features of Potential Energy Surfaces

Molecular Orbital Calculations on Biochemical systems
Conventional MO calculations scale as N3 or worse, where N is a measure of the size of the system (number of atoms, electrons, basis functions)
Recent developments have lead to linear scaling methods for semi-empirical, Hartree-Fock and density functional calculations
Similar improvements in geometry optimization facilitate the use of internal coordinates for finding local minima for small proteins

Optimization of Large Molecules
Calculations on larger molecules more practical because of faster software and hardware.
Traditional optimization methods require O(N3) computational effort, and O(N2) memory
Iterative O(N2) methods for coordinate transformation and quasi-Newton optimization.
O(N) methods using sparse matrix methods and GDIIS.

Coordinate transformations: dq = B dx and fx = BT fq
usually solved by generalized inverse ( O(N3) ) Newton Raphson and RFO steps: (H - S) q = fq
usually solved by diagonalization ( O(N3) ) Perturbative sequence of linear equations: yi = Mi xi,
iterative solution requires only O(N2) work speed-up convergence by updating Mi
-1 rapid convergence since changes are small from one
optimization step to next
Optimization of Large Molecules
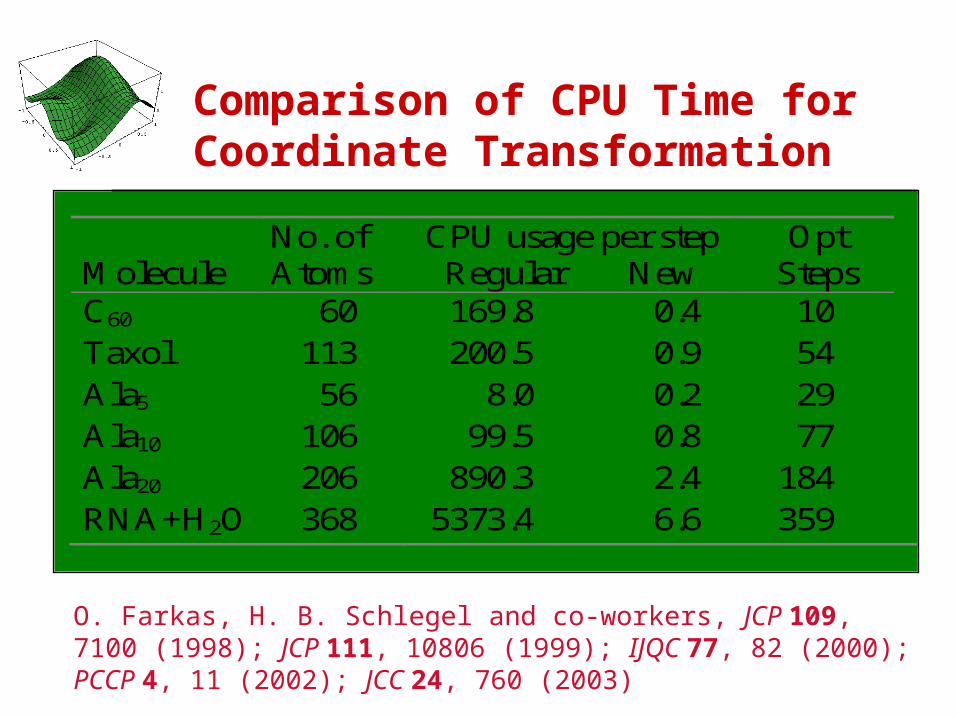
Comparison of CPU Time for Coordinate Transformation
O. Farkas, H. B. Schlegel and co-workers, JCP 109, 7100 (1998); JCP 111, 10806 (1999); IJQC 77, 82 (2000); PCCP 4, 11 (2002); JCC 24, 760 (2003)
Molecule
No. of Atoms
CPU usage per step Regular New
Opt Steps
C60 60 169.8 0.4 10 Taxol 113 200.5 0.9 54 Ala5 56 8.0 0.2 29 Ala10 106 99.5 0.8 77 Ala20 206 890.3 2.4 184 RNA+H2O 368 5373.4 6.6 359
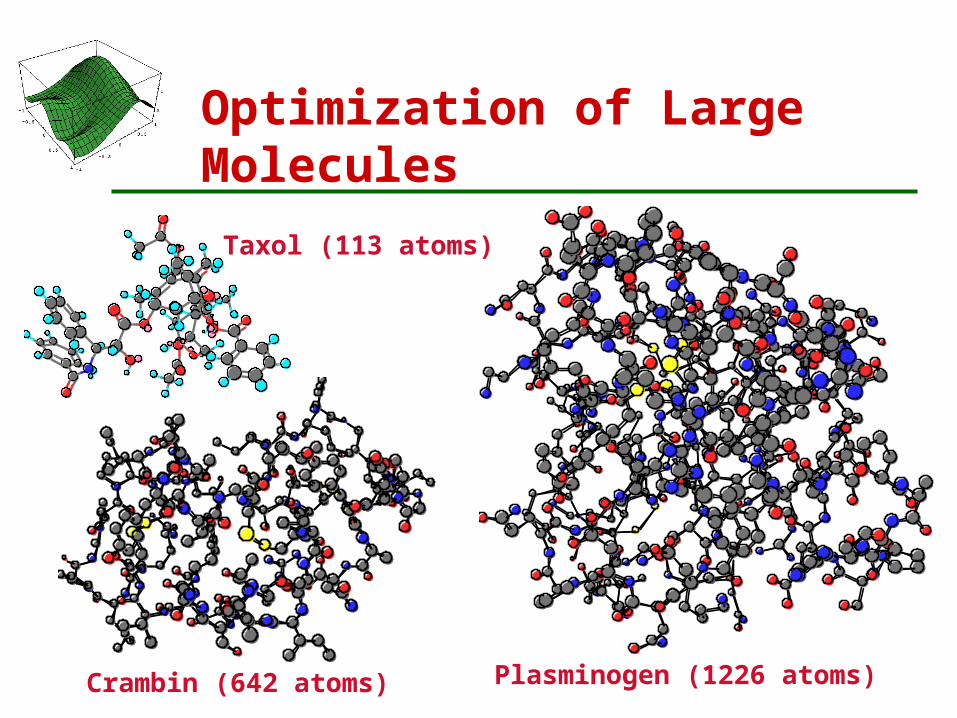
Taxol (113 atoms)
Crambin (642 atoms) Plasminogen (1226 atoms)
Optimization of Large Molecules
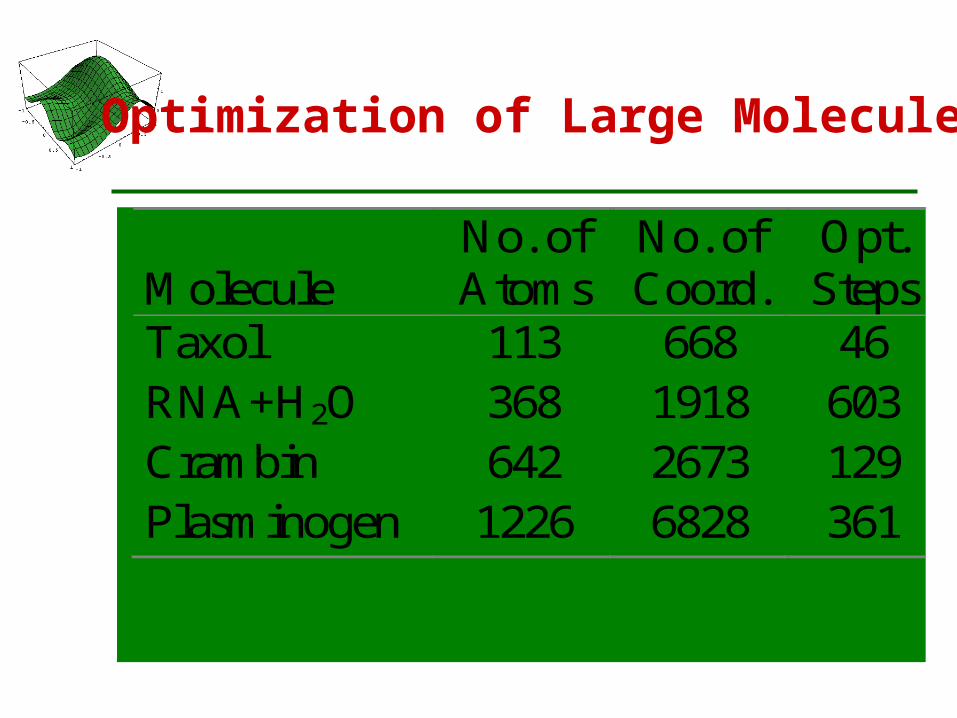
Optimization of Large Molecules
Molecule
No. of Atoms
No. of Coord.
Opt. Steps
Taxol 113 668 46 RNA+H2O 368 1918 603 Crambin 642 2673 129 Plasminogen 1226 6828 361

Hybrid Methods: MO/MO and QM/MM
The active site region is treated using high-level molecular orbital theory, while the most distant parts of the enzyme are treated using low-cost molecular mechanics.
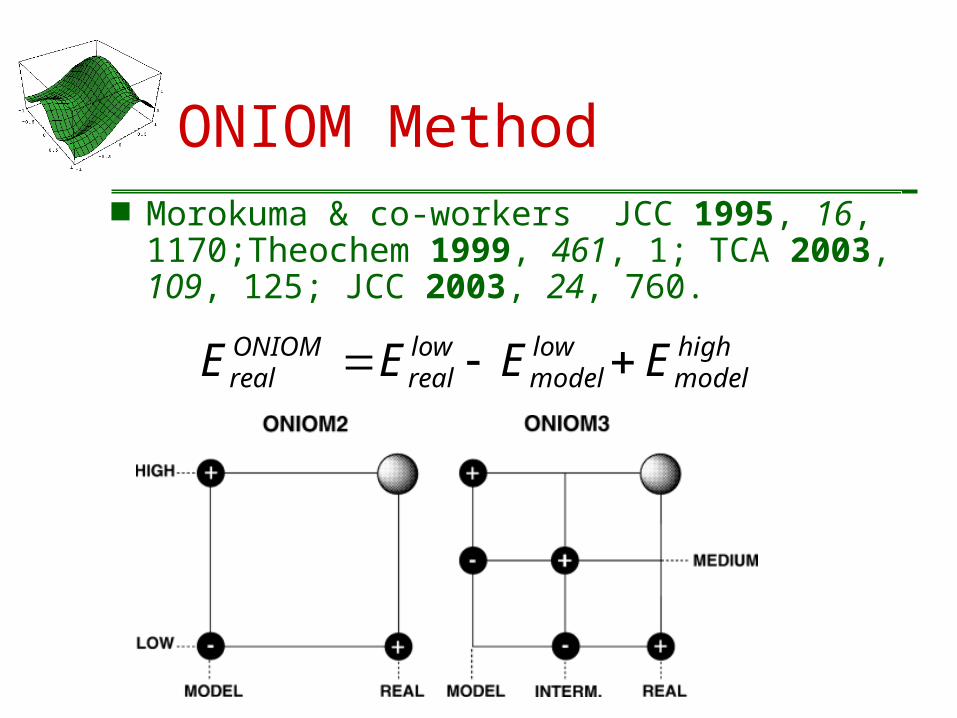
ONIOM Method Morokuma & co-workers JCC 1995, 16,
1170;Theochem 1999, 461, 1; TCA 2003, 109, 125; JCC 2003, 24, 760.
highmodel
lowmodel
lowreal
ONIOMreal EEEE
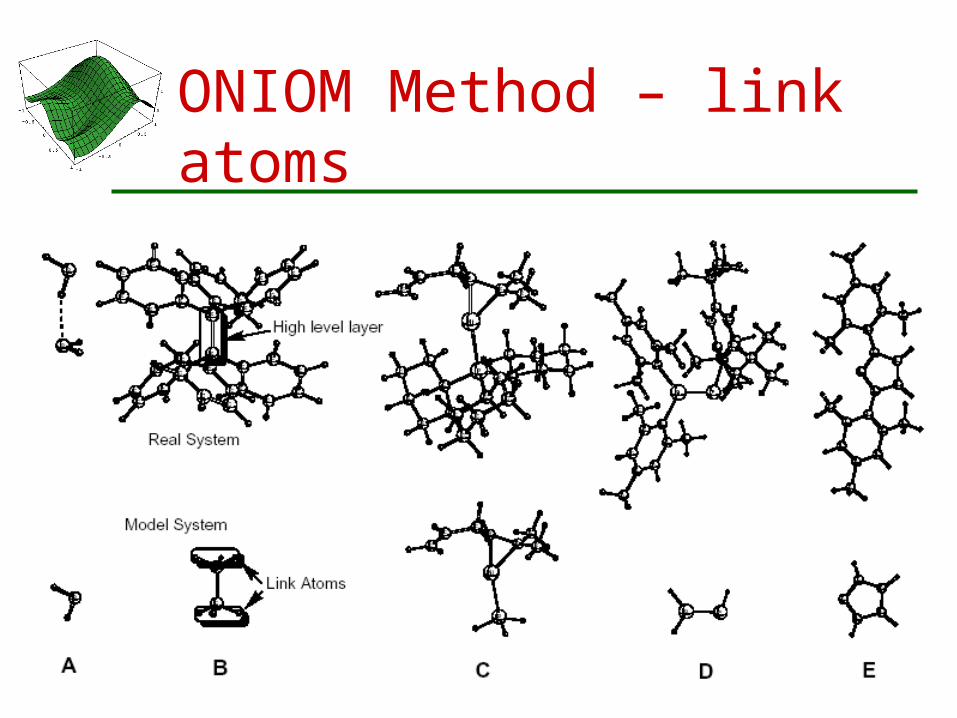
ONIOM Method – link atoms

Irreversible Inactivation of the Zinc-Dependent Protease Carboxypeptidase A (CPA)
Zinc ion plays a central function in the catalytic mechanism of many enzymes.
A novel class of mechanism-based inhibitors developed by Mobashery
Zinc promotes an SN2 reaction between the inhibitor and the carboxylate of Glu-270, resulting in irreversible inhibition
J. B. Cross, T. Vreven, S. O. Meroueh, S. Mobashery and H. B. Schlegel
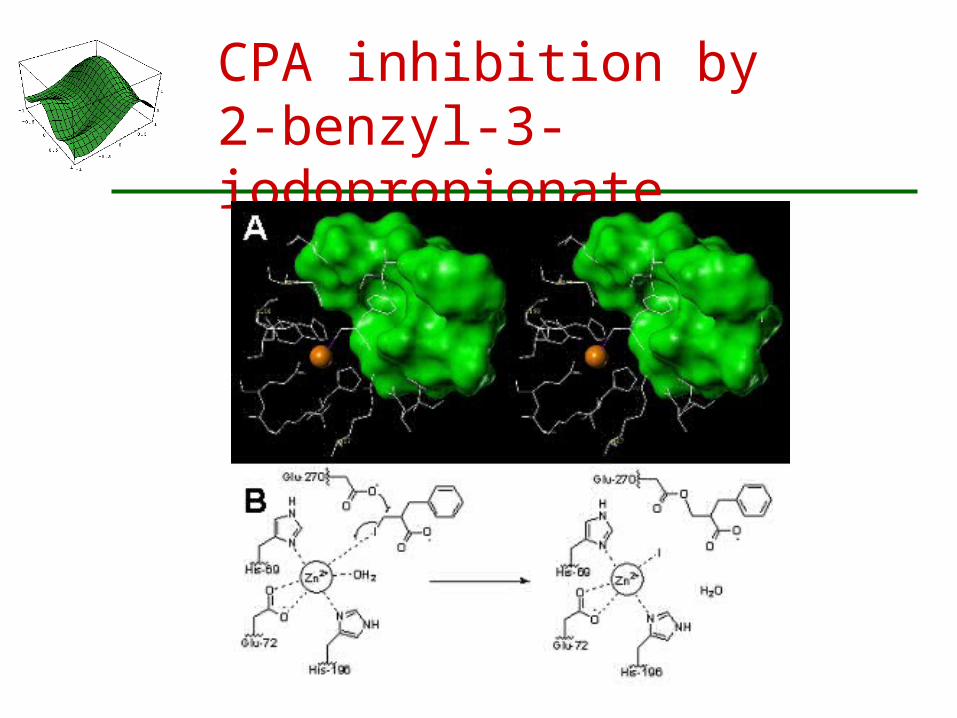
CPA inhibition by2-benzyl-3-iodopropionate

Effect of CPA Active Site Models on a Simple SN2 Reaction
Model A - a bare zinc ion ab initio calculation with HF/6-31G(d) barrier disappears for R(Zn-C) < 10 Å, entirely
due to electrostatics Model B – zinc ion with ligands
small barrier at R(Zn-C) = 5 Å ligand charge and polarization moderates the
electrostatics Model C – add residues within 10 Å of zinc
additional residues calculated by AM1 smaller barrier at 5 Å separation, due to
electrostatics from additional residues
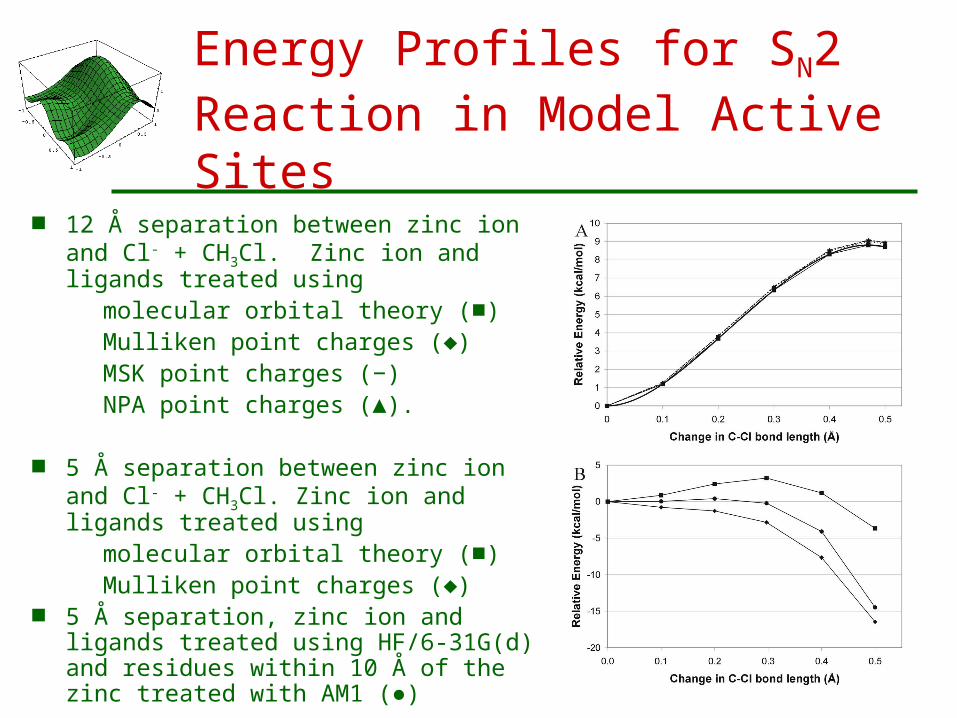
Energy Profiles for SN2 Reaction in Model Active Sites
12 Å separation between zinc ion and Cl- + CH3Cl. Zinc ion and ligands treated using
molecular orbital theory (■)Mulliken point charges (♦)MSK point charges (−)NPA point charges (▲).
5 Å separation between zinc ion and Cl- + CH3Cl. Zinc ion and ligands treated using
molecular orbital theory (■) Mulliken point charges (♦)
5 Å separation, zinc ion and ligands treated using HF/6-31G(d) and residues within 10 Å of the zinc treated with AM1 (●)
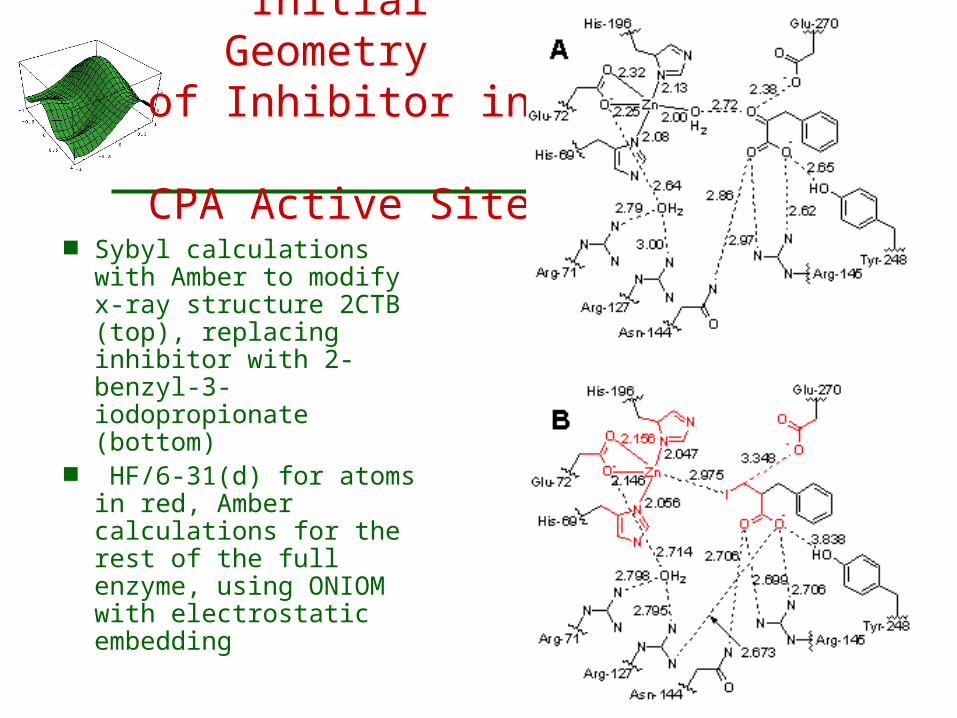
Initial Geometry of Inhibitor in
CPA Active Site
Sybyl calculations with Amber to modify x-ray structure 2CTB (top), replacing inhibitor with 2-benzyl-3-iodopropionate (bottom)
HF/6-31(d) for atoms in red, Amber calculations for the rest of the full enzyme, using ONIOM with electrostatic embedding
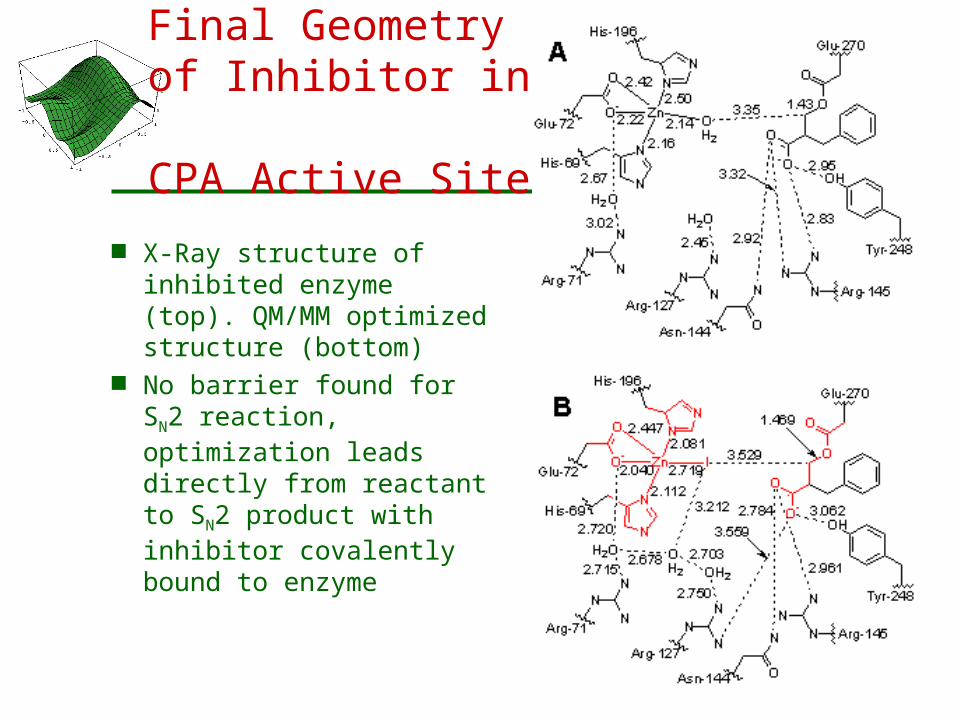
Final Geometry of Inhibitor in
CPA Active Site
X-Ray structure of inhibited enzyme (top). QM/MM optimized structure (bottom)
No barrier found for SN2 reaction, optimization leads directly from reactant to SN2 product with inhibitor covalently bound to enzyme
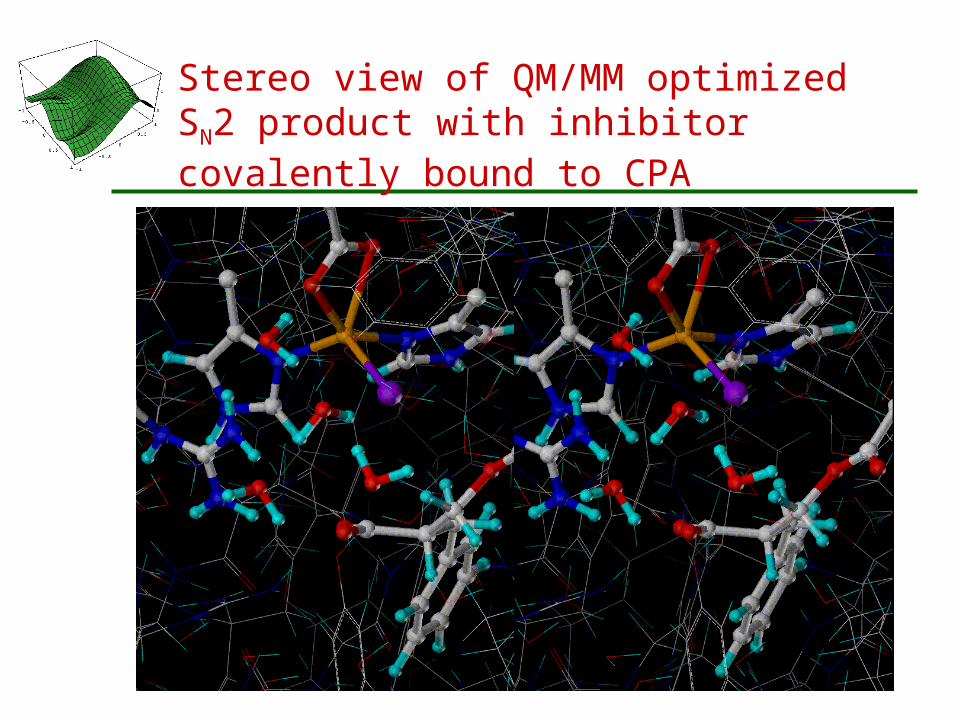
Stereo view of QM/MM optimized SN2 product with inhibitor covalently bound to CPA

OXA-10 β-lactamase X-ray structure shows
carboxylated Lys70 Modified Lys70 has
mechanistic role Removes proton
from Ser67 Leads to acylation of
Ser67 by substrate Enzyme shows biphasic
kinetics during substrate turnover
J. Li; J. B. Cross; T. Vreven; S. O. Meroueh; S. Mobashery; H. B. Schlegel.
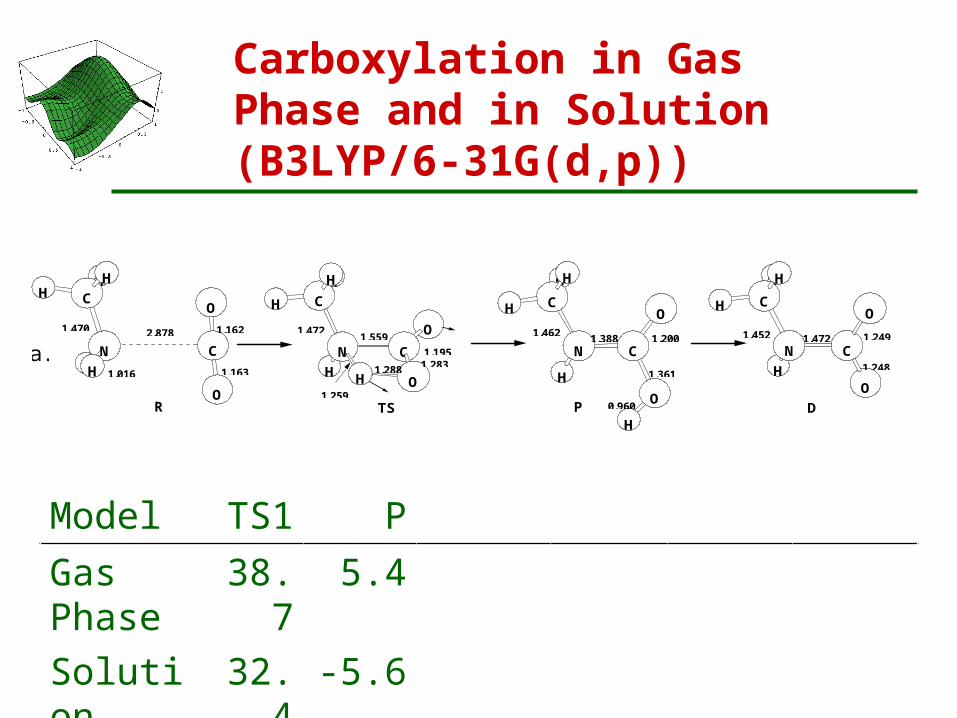
Carboxylation in Gas Phase and in Solution (B3LYP/6-31G(d,p))
Model TS1 PGas Phase 38.7 5.4
Solution 32.4 -5.6
H
H
O
C
O
N
C H
H
H
1.470 2.878 1.162
1.163 1.016
R
H
O
H C N
C H H
H O
1.472 1.559 1.195
1.283 1.288
1.259 TS
H
H O
C N
C
H
O H
H
1.462 1.388 1.200
1.361
0.960 P
H
H O
C N
C H O
H
1.452 1.472 1.249
1.248
D
a.
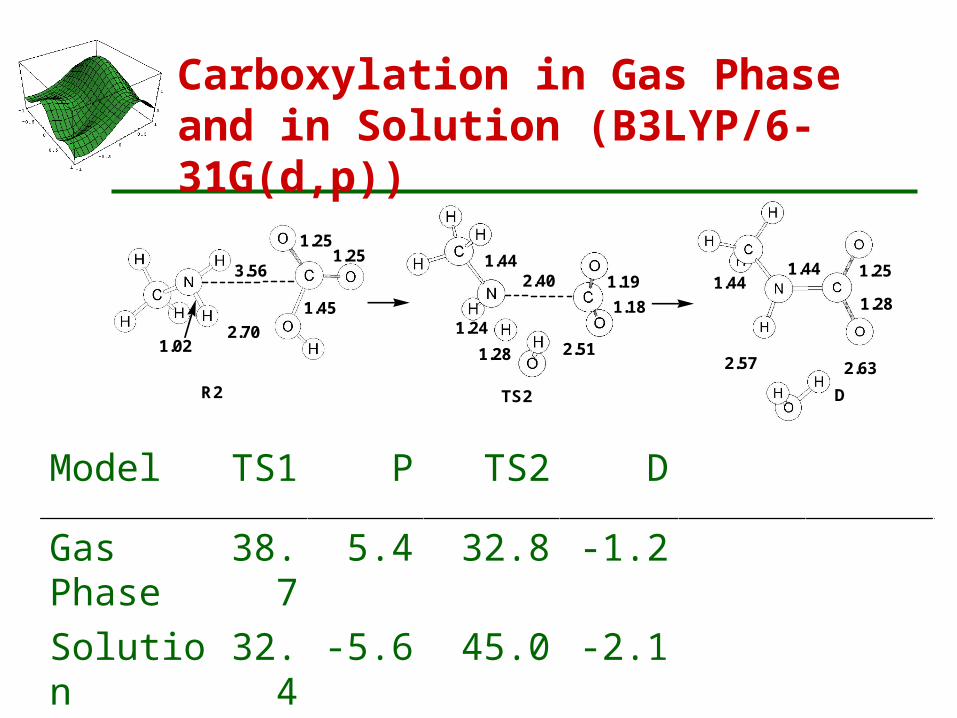
Carboxylation in Gas Phase and in Solution (B3LYP/6-31G(d,p))
Model TS1 P TS2 D
Gas Phase 38.7 5.4 32.8 -1.2
Solution 32.4 -5.6 45.0 -2.1
R2 (-9.7) TS2 (20.7)
3.56
1.022.70
1.251.25
1.452.40
1.241.28 2.51
1.191.18
1.44 1.44 1.25
1.28
2.57 2.63
1.44
D2 (-14.0)
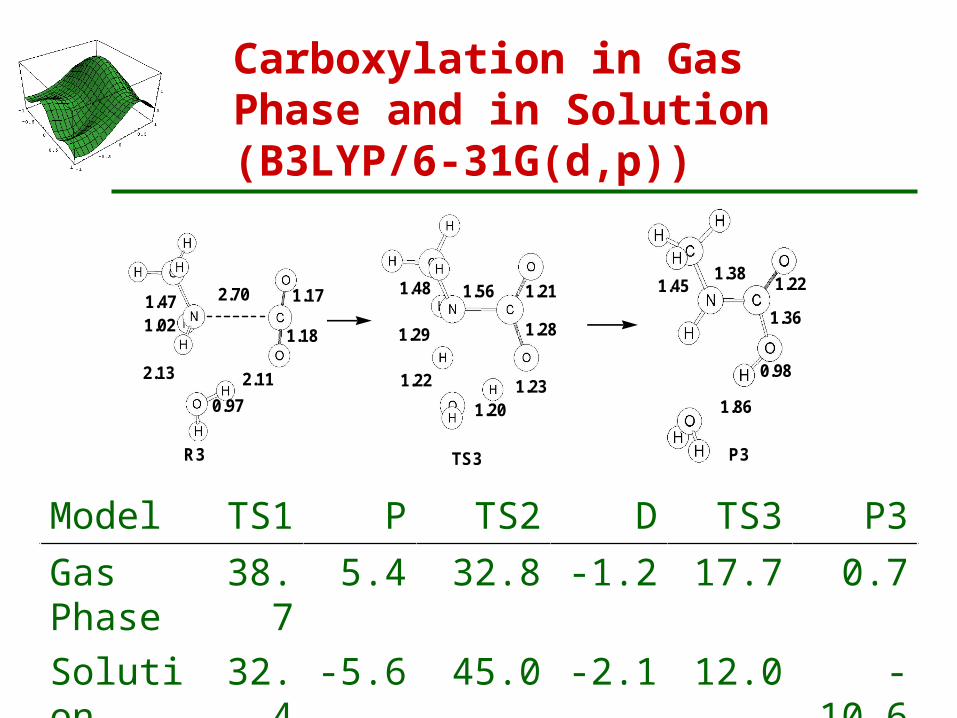
Carboxylation in Gas Phase and in Solution (B3LYP/6-31G(d,p))
R3 (-8.8)
2.70 1.17
1.181.021.47
2.112.13
0.97
TS3 (12.9)
1.29
1.221.20
1.23
1.28
1.211.561.48
P3 (-5.3)
1.38 1.22
1.36
0.98
1.86
1.45
Model TS1 P TS2 D TS3 P3Gas Phase 38.7 5.4 32.8 -1.2 17.7 0.7
Solution 32.4 -5.6 45.0 -2.1 12.0 -10.6

QM/MM Calculations of the Transition State for Lys-70 Carboxylation
Trp-154 Lys-70
Ser-67
Trp-154 Lys-70
Ser-67
Stereoview of the TS showing a molecule of water catalyzing the addition of carbon dioxide to the side chain of Lys-70

QM/MM Calculations of the Reactants, TS and Products for Lys-70 Carboxylation
N
O
O O
H
NH
H
H
OH
HO
H
Trp-154
Lys-70
2.732.95
2.87
2.802.84
QM/MM-R
1.18 1.18
2.26Ser-67
N
O
OO
H
N
H
H
HOH
HO
H
Trp-154
Lys-70QM/MM-TS
Ser-67
N
O
OO
H
N
H
H
H H
OH
HO
H
Trp-154
Lys-70
2.682.78
3.003.03
2.74N/A
2.68N/A2.87
2.98
QM/MM-P
1.221.23
1.351.25
1.381.37
Ser-67
H H
1.52
1.27
1.34
2.622.78
1.21
2.672.86
3.80 1.30
1.131.30
N
O
O O
H
N
H
H
H
OH
HO
H
Trp-154
Lys-70QM/MM-D
2.74N/A
3.063.03
2.70N/A
2.682.78
2.812.98
1.411.37
1.271.23
1.271.25
Ser-67
QM/MM values in normal text
X-ray values in italics

OXA-10 β-lactamase - Discussion A water molecule in the active site can catalyze
carboxylation of Lys70 with CO2
X-ray structure is most likely the deprotonated carboxylation product
Carboxylation is accompanied by deprotonation Re-protonation of carbamate nitrogen results in
barrierless loss of CO2, accounting for biphasic kinetics of enzyme

Selective Mechanism-Based Inhibition of human gelatinases, MMP-2 and MMP-9
A family of Zinc-dependent endopeptidases called MMPs (matrix metalloproteinases) carries out various cellular processes.
Excessive MMP activity has been implicated in many diseases such as cancer growth, tumor metastasis, etc.
A selective mechanism-based inhibitor (SB-3CT) for MMP-2 and MMP-9 developed by Mobashery
Zinc promotes an SN2 reaction between the inhibitor and the carboxylate of a glutamate in the active site, resulting in covalent attachment of the inhibitor to the enzyme
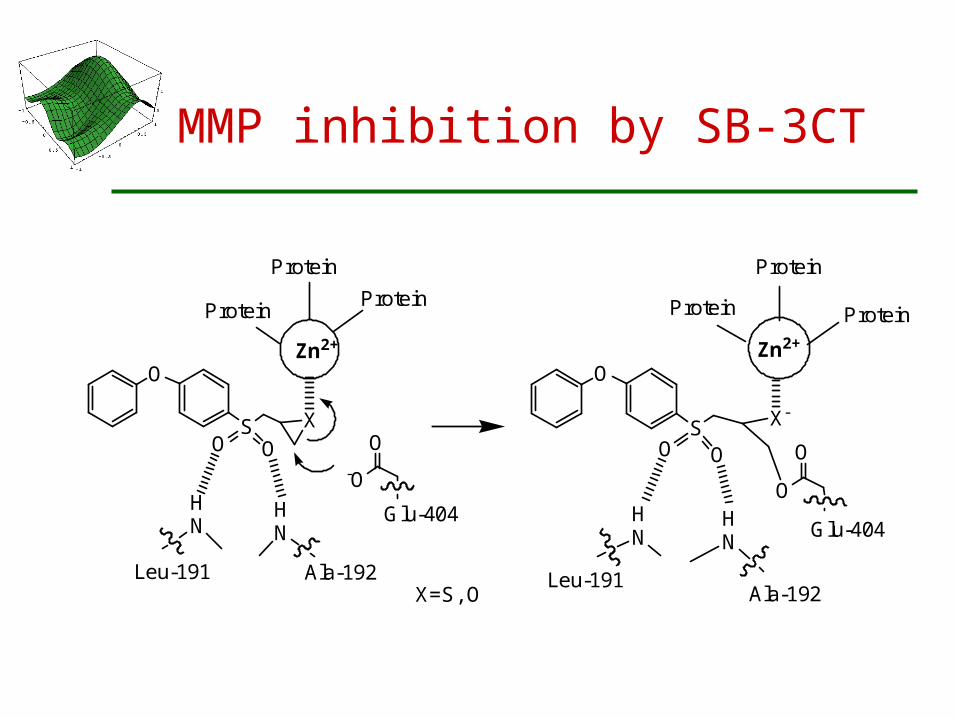
MMP inhibition by SB-3CT
O
S X
Zn2+
O O
Leu-191
HN
HN
Ala-192
O-O
Glu-404
O
S X-
Zn2+
O O
Leu-191
HN
HN
Ala-192
O
O
Glu-404
X=S, O
ProteinProtein
Protein Protein
Protein
Protein

Preliminary results for model systems
Method TS1 P1 TS2 P2HF/6-31G(d) 11.1 -9.2 26.9 11.4
MP2/6-31G(d) 12.2 -0.6 21.5 13.0
B3LYP/6-31G(d)
6.0 -3.3 16.3 10.0
CBS-QB3 10.4 -1.2 16.7 5.5
a
b

Schlegel Group

Acknowledgements
Collaborators: Prof. S.O. Mobashery, Notre Dame Prof. C.H. Winter, WSU Prof. J.W. Montgomery, WSU Prof. P.G. Wang, Ohio State Prof. R.J. Levis, Temple U Prof. W.L. Hase, Texas Tech Prof. E.M. Goldfield, WSU Prof. G. Scuseria, Rice Univ. Prof. G. Voth, U. of Utah Prof. Srini Iyengar, Indiana U Prof. O. Farkas, ELTE Prof. M.A. Robb, U. of London
Funding and Resources:
Wayne State U.
National Science Foundation
NCSA
Gaussian, Inc.


















