
Binding and uptake of trypan blue by developing oocytes of Locusta migratoria … · Developing...
11
/. Embryol. exp. Morph. Vol. 37, pp. 1-11, 1977 Printed in Great Britain Binding and uptake of trypan blue by developing oocytes of Locusta migratoria migratorioides By ESTHER WAJC, 1 TILLY BAKKER-GRUNWALD 2 AND SHALOM W. APPLEBAUM 1 From the Faculty of Agriculture of The Hebrew University, Department of Entomology, Rehovot, Israel SUMMARY Resorbing oocytes are heavily stained by trypan blue injected into the haemolymph; this selves as a basis for a quick and convenient method for measuring the degree of resorption. Oocytes in the beginning of their development are most susceptible towards resorptive tendencies. The uptake of trypan blue by normally developing oocytes is proportional to the oocyte surface. From 'double-marker' experiments, in which trypan blue is injected into the haemolymph together with [ 3 H]inulin (which does not bind to the oocyte membrane) it is estimated that the contribution of binding in the interiorization of trypan blue is in the order of 80 %, under the conditions given. In vitro incubations show the interaction of trypan blue with the membrane to be electro- static in nature. INTRODUCTION Developing oocytes of the African migratory locust Locusta migratoria migratorioides pinocytize vitellogenic proteins that are synthesized in the fat body and transferred in the haemolymph (Tobe & Loughton, 1967; Bentz, Girardie & Cazal, 1970; Bar-Zev et ah 1975). This uptake mechanism has been shown to be highly specific in the cecropia moth (Telfer, 1960), other proteins being barely incorporated. Factors contributing to this specificity may be the existence of specific receptor sites and/or charge effects on the oocyte plasma membrane, or the secretion of a follicle cell product which interacts with the vitellogenins and/or stimulates net pinocytotic uptake (Anderson & Telfer, 1970). Resorption of developing oocytes may occur until the final stage of vitello- genesis (Tobe & Pratt, 1975), and is influenced by environmental conditions (Highnam, Lusis & Hill, 1963). It has been shown (Anderson & Telfer, 1970) that the negatively charged vital 1 Author's address: Department of Entomology, Faculty of Agriculture, The Hebrew University of Jerusalem, Rehovot 76-100, P.O. Box 12, Israel. 2 Author's address: Department of Physiology, University of Colorado, Medical School, Denver. Colo. 80220.
Transcript of Binding and uptake of trypan blue by developing oocytes of Locusta migratoria … · Developing...

/ . Embryol. exp. Morph. Vol. 37, pp. 1-11, 1977
Printed in Great Britain
Binding and uptake of trypan blue by developingoocytes of Locusta migratoria migratorioides
By ESTHER WAJC,1 TILLY BAKKER-GRUNWALD2
AND SHALOM W. APPLEBAUM1
From the Faculty of Agriculture of The Hebrew University,Department of Entomology, Rehovot, Israel
SUMMARYResorbing oocytes are heavily stained by trypan blue injected into the haemolymph; this
selves as a basis for a quick and convenient method for measuring the degree of resorption.Oocytes in the beginning of their development are most susceptible towards resorptivetendencies.
The uptake of trypan blue by normally developing oocytes is proportional to the oocytesurface. From 'double-marker' experiments, in which trypan blue is injected into thehaemolymph together with [3H]inulin (which does not bind to the oocyte membrane) it isestimated that the contribution of binding in the interiorization of trypan blue is in the orderof 80 %, under the conditions given.
In vitro incubations show the interaction of trypan blue with the membrane to be electro-static in nature.
INTRODUCTION
Developing oocytes of the African migratory locust Locusta migratoriamigratorioides pinocytize vitellogenic proteins that are synthesized in the fatbody and transferred in the haemolymph (Tobe & Loughton, 1967; Bentz,Girardie & Cazal, 1970; Bar-Zev et ah 1975). This uptake mechanism has beenshown to be highly specific in the cecropia moth (Telfer, 1960), other proteinsbeing barely incorporated. Factors contributing to this specificity may be theexistence of specific receptor sites and/or charge effects on the oocyte plasmamembrane, or the secretion of a follicle cell product which interacts with thevitellogenins and/or stimulates net pinocytotic uptake (Anderson & Telfer,1970).
Resorption of developing oocytes may occur until the final stage of vitello-genesis (Tobe & Pratt, 1975), and is influenced by environmental conditions(Highnam, Lusis & Hill, 1963).
It has been shown (Anderson & Telfer, 1970) that the negatively charged vital1 Author's address: Department of Entomology, Faculty of Agriculture, The Hebrew
University of Jerusalem, Rehovot 76-100, P.O. Box 12, Israel.2 Author's address: Department of Physiology, University of Colorado, Medical School,
Denver. Colo. 80220.

2 E. WAJC AND OTHERS
stain trypan blue binds to the membrane of developing oocytes in cecropia, andis taken up in a way similar to the vitellogenic proteins.
In the following paper, we report on the use of trypan blue in the investigationof certain quantitative aspects of yolk deposition and its control in Locustamigratoria migratorioides.
MATERIALS AND METHODS
Locusts were reared under crowded conditions in a continuous light regime.Females were taken from the cages at various times after the adult moult.
For in vivo experiments, animals were injected with up to 100/d of a 1 % or1-5 % trypan blue solution (BDH; dialysed against distilled water for 48 h), and/or with a solution of [3H]inulin (Amersham; specific activity 300mCi/mmol);1 % or 10 % cold inulin (BDH) was added where indicated (in order to dissolvethis, the solution had to be heated). The animals were left in glass jars or smallcages under illumination (29-31 °C) until haemolymph sampling and dissection(after 15 min-13 h). At the end of the incubation period, haemolymph samples of10-20 iA were either added to 2-5 ml 1 % SDS solution for determination of try-pan blue content, or to 20 ml counting vials containing 0-2 ml Soluene (Packard)for determination of radioactivity (see below). Ovaries were removed andwashed twice in locust saline solution (NaCl 170 mM, KC1 6 mM, MgCl2 4 HIM,CaCl2 2 mM, NaH2PO4 7mM, NaHCO3 2 mM (Mordue & Goldsworthy, 1969)).
The length and diameter of the oocytes were measured with the aid ofa micrometer scale in the ocular of an Olympus stereoscopic microscope.Ovarioles were removed from the ovaries, cleaned of the ovarial sheath tissue,and the terminal oocytes were isolated. Follicle cells were still present at thatstage. Resorbed oocytes (detected as described in 'Results') and fully-grownoocytes in which 'cap' formation had started were discarded.
The amount of trypan blue associated with the oocytes was determinedessentially according to Anderson & Telfer (1970). For the determination of[3H]inulin content, 4-20 oocytes (depending upon the size) were solubilized in1 ml Soluene; 5 ml toluene-based scintillation fluid was added, and the sampleswere counted in a Packard Tri-Carb scintillation counter at optimal gain.
For in vitro experiments, the paired ovaries were removed and separated; oneovary served as a control and was incubated in 1 ml saline solution (NaCl80 mM, KC110mM, MgCl21 mM,CaCl2 lmM, Na-MES 5 mM, pH 6-5), and the otherin a modified solution, as indicated. After 30min in a shaking bath at 26 °C, theovaries were each transferred to 1 ml of the appropriate solutions, but nowcontaining 0-5 % trypan blue; in some cases, [3H]inulin (2 x 106 cpm) and coldinulin (1 %) were added. After an additional incubation, samples of theincubation medium were taken and the ovaries were washed. Trypan blue and[3H]inulin in the medium and that incorporated in the terminal oocytes weredetermined as described for the in vivo experiments.
In all cases less than 10 % of the trypan blue or inulin injected into the
Uptake of trypan blue by locust oocytes
0 1 2 3 4 5 6Oocyte length (mm)
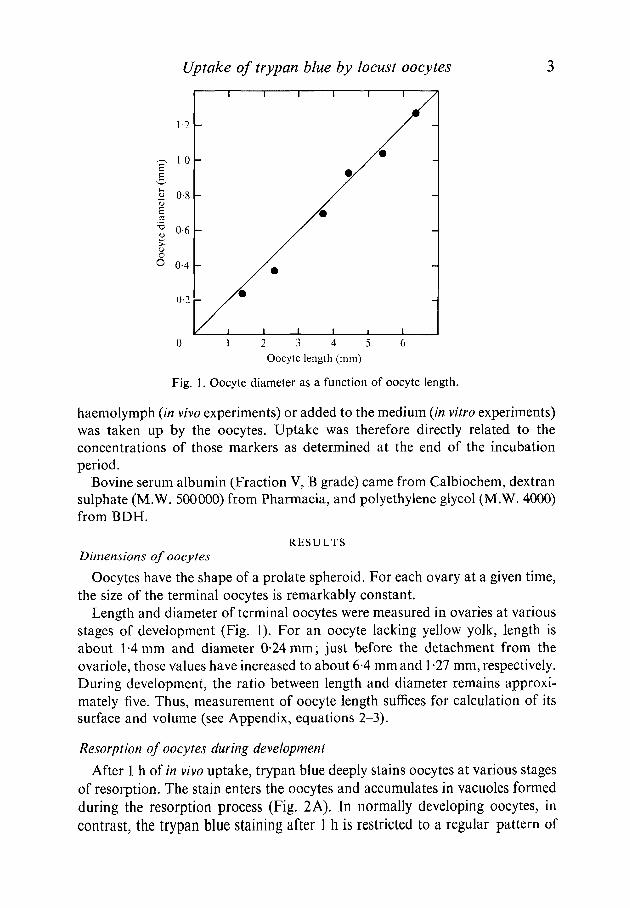
Fig. 1. Oocyte diameter as a function of oocyte length.
haemolymph (in vivo experiments) or added to the medium (in vitro experiments)was taken up by the oocytes. Uptake was therefore directly related to theconcentrations of those markers as determined at the end of the incubationperiod.
Bovine serum albumin (Fraction V, B grade) came from Calbiochem, dextransulphate (M.W. 500000) from Pharmacia, and polyethylene glycol (M.W. 4000)from BDH.
RESULTSDimensions of oocytes
Oocytes have the shape of a prolate spheroid. For each ovary at a given time,the size of the terminal oocytes is remarkably constant.
Length and diameter of terminal oocytes were measured in ovaries at variousstages of development (Fig. 1). For an oocyte lacking yellow yolk, length isabout 1-4 mm and diameter 0-24mm; just before the detachment from theovariole, those values have increased to about 6-4 mm and 1-27 mm, respectively.During development, the ratio between length and diameter remains approxi-mately five. Thus, measurement of oocyte length suffices for calculation of itssurface and volume (see Appendix, equations 2-3).
Resorption of oocytes during development
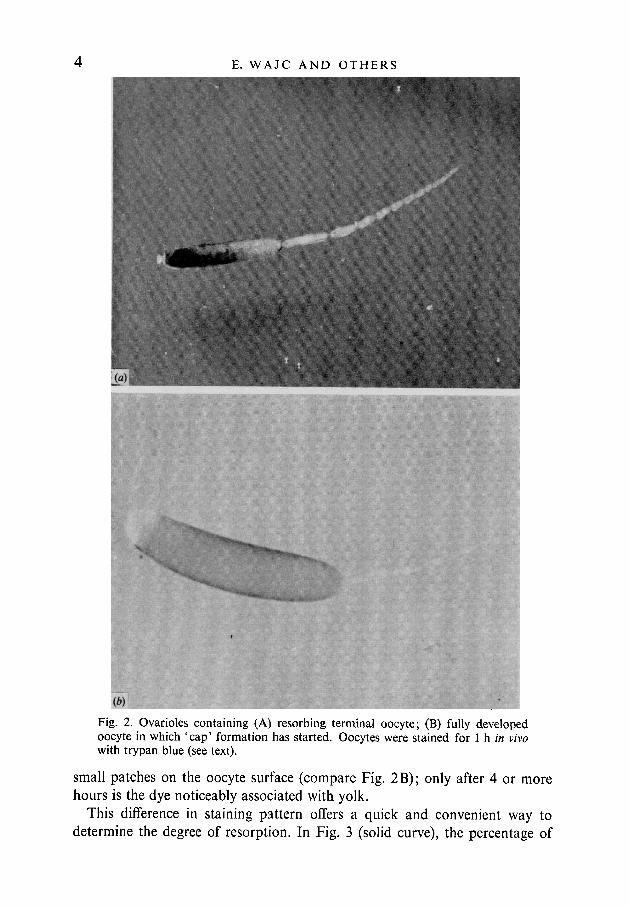
After 1 h of in vivo uptake, trypan blue deeply stains oocytes at various stagesof resorption. The stain enters the oocytes and accumulates in vacuoles formedduring the resorption process (Fig. 2A). In normally developing oocytes, incontrast, the trypan blue staining after 1 h is restricted to a regular pattern of
E. WAJC AND OTHERS
(b)
Fig. 2. Ovarioles containing (A) resorbing terminal oocyte; (B) fully developedoocyte in which 'cap' formation has started. Oocytes were stained for 1 h in vivowith trypan blue (see text).
moresmall patches on the oocyte surface (compare Fig. 2B); only after 4 orhours is the dye noticeably associated with yolk.
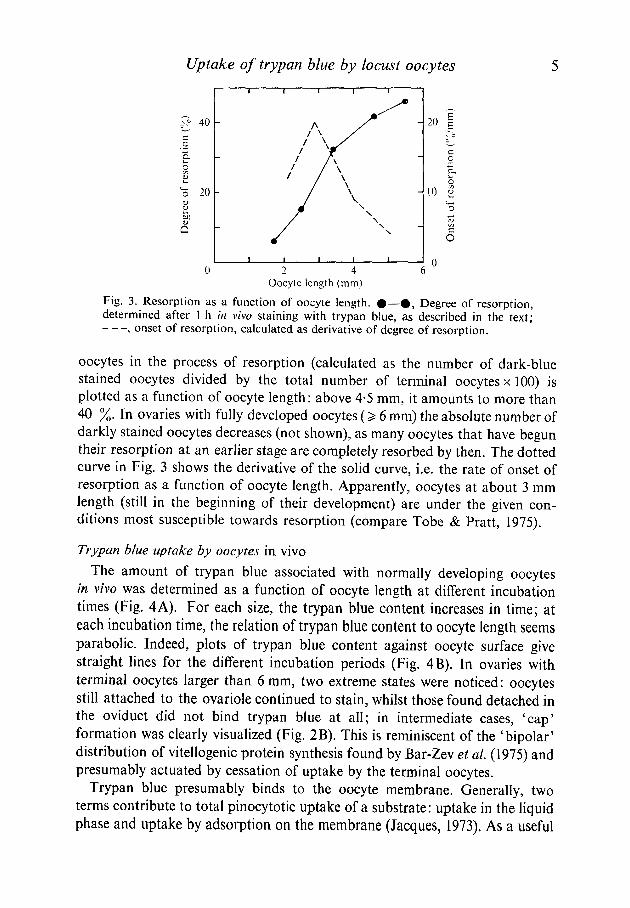
This difference in staining pattern offers a quick and convenient way todetermine the degree of resorption. In Fig. 3 (solid curve), the percentage of
Uptake of trypan blue by locust oocytes
x= 40 -
0 2 4Oocyte length (mm)
Fig. 3. Resorption as a function of oocyte length. #—ft, Degree of resorption,determined after 1 h in vivo staining with trypan blue, as described in the text;
, onset of resorption, calculated as derivative of degree of resorption.
oocytes in the process of resorption (calculated as the number of dark-bluestained oocytes divided by the total number of terminal oocytes x 100) isplotted as a function of oocyte length: above 4-5 mm, it amounts to more than40 %. In ovaries with fully developed oocytes (> 6 mm) the absolute number ofdarkly stained oocytes decreases (not shown), as many oocytes that have beguntheir resorption at an earlier stage are completely resorbed by then. The dottedcurve in Fig. 3 shows the derivative of the solid curve, i.e. the rate of onset ofresorption as a function of oocyte length. Apparently, oocytes at about 3 mmlength (still in the beginning of their development) are under the given con-ditions most susceptible towards resorption (compare Tobe & Pratt, 1975).
Trypan blue uptake by oocytes in vivo
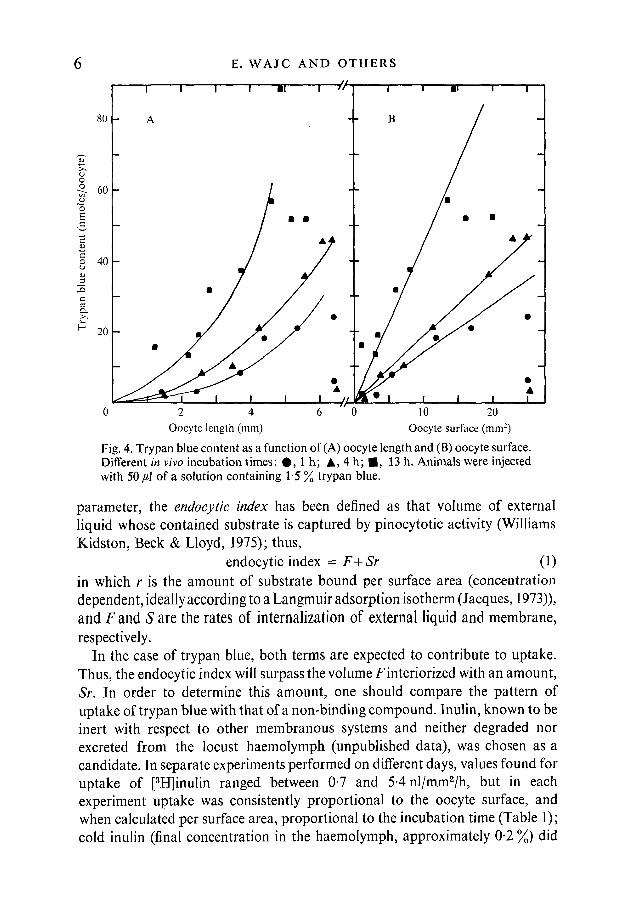
The amount of trypan blue associated with normally developing oocytesin vivo was determined as a function of oocyte length at different incubationtimes (Fig. 4A). For each size, the trypan blue content increases in time; ateach incubation time, the relation of trypan blue content to oocyte length seemsparabolic. Indeed, plots of trypan blue content against oocyte surface givestraight lines for the different incubation periods (Fig. 4B). In ovaries withterminal oocytes larger than 6 mm, two extreme states were noticed: oocytesstill attached to the ovariole continued to stain, whilst those found detached inthe oviduct did not bind trypan blue at all; in intermediate cases, 'cap'formation was clearly visualized (Fig. 2B). This is reminiscent of the 'bipolar'distribution of vitellogenic protein synthesis found by Bar-Zev et al (1975) andpresumably actuated by cessation of uptake by the terminal oocytes.
Trypan blue presumably binds to the oocyte membrane. Generally, twoterms contribute to total pinocytotic uptake of a substrate: uptake in the liquidphase and uptake by adsorption on the membrane (Jacques, 1973). As a useful
E. WAJC AND OTHERS
80
T I
A B
2 4Oocyte length (mm)
10 20Oocyte surface (mm2)
Fig. 4. Trypan blue content as a function of (A) oocyte length and (B) oocyte surface.Different in vivo incubation times: # , l h ; A, 4 h; • , 13 h. Animals were injectedwith 50/A of a solution containing 1 5 % trypan blue.
parameter, the endocytic index has been defined as that volume of externalliquid whose contained substrate is captured by pinocytotic activity (WilliamsKidston, Beck & Lloyd, 1975); thus,
endocytic index = F+Sr (1)in which r is the amount of substrate bound per surface area (concentrationdependent, ideally according to a Langmuir adsorption isotherm (Jacques, 1973)),and F and S are the rates of internalization of external liquid and membrane,respectively.
In the case of trypan blue, both terms are expected to contribute to uptake.Thus, the endocytic index will surpass the volume Finteriorized with an amount,Sr. In order to determine this amount, one should compare the pattern ofuptake of trypan blue with that of a non-binding compound. Inulin, known to beinert with respect to other membranous systems and neither degraded norexcreted from the locust haemolymph (unpublished data), was chosen as acandidate. In separate experiments performed on different days, values found foruptake of [3H]inulin ranged between 0-7 and 54 nl/mm2/h, but in eachexperiment uptake was consistently proportional to the oocyte surface, andwhen calculated per surface area, proportional to the incubation time (Table 1);cold inulin (final concentration in the haemolymph, approximately 0-2 %) did

Uptake of trypan blue by locust oocytes
Table 1. Inulin uptake in vivo
XI l^UUdtl^/IJ.time (h)
2
4
7-5
Coldinulin
_
+—+—
Oocytesize (mm)
6 04-53-85-22-53-8
Surface(mm2)
22128-7
163-78-7
f
nl/oocyte
805358
1305619
Uptake
nl/mm2
3-64-46-781
152-2
nl/mm2/h
1-92-21-82 0200-3
Animals were injected with 10/*1 of a solution containing 6x 105 cpm [3H]inulin, whereindicated preceded by 10 /tl 10 % cold inulin. After the indicated times, haemolymph samplesand oocytes were prepared for scintillation counting as described. Haemolymph volumesranged between 500-800/tl.
1 2 3 4 5Incubation time (h)
Fig. 5. Endocytic index for trypan blue and inulin as a function of incubation time.# , trypan blue uptake; O, inulin uptake. Animals were injected with 100/tl ofa solution containing 1 % trypan blue, 3 x 105 cpm [3H]inulin and 1 % cold inulin.
not significantly decrease label content. From the fact that the extrapolatedinulin content at zero time is zero, and that cold inulin in the given concen-tration does not act as a competitor, it can be inferred that inulin is inert withrespect to oocyte membranes too, and that its endocytic index at any momentrepresents the real volume of fluid interiorized.
In Fig. 5, the compiled results of some experiments are shown in which trypanblue and [3H]inulin were injected simultaneously. A certain amount of trypanblue (corresponding to 4 nl/mm2) appears to be associated with the oocytes atzero time, and the increase in trypan blue content is about six times as fast asthat in inulin content. Values for inulin uptake were rather scattered, but in
E. WAJC AND OTHERS
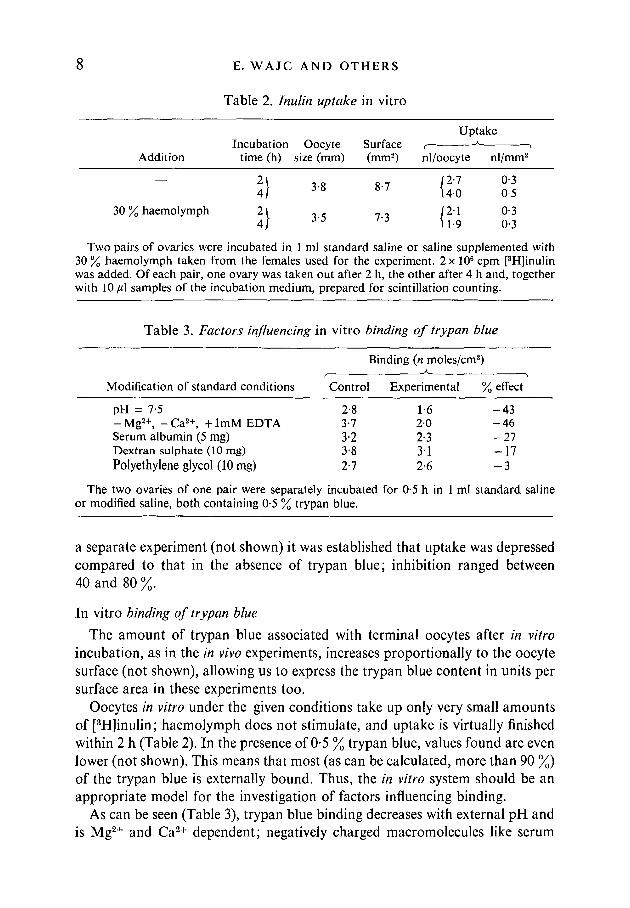
Table 2. Inulin uptake in vitro
Addition
30 % haemolymph
Incubationtime (h)
I)I)
Oocytesize (mm)
3-8
3-5
Surface(mm2)
8-7
7-3
Uptake
nl/oocyte
12-1\40
|2-111-9
nl/mm2
0-30-5
0-30-3
Two pairs of ovaries were incubated in 1 ml standard saline or saline supplemented with30 % haemolymph taken from the females used for the experiment. 2 x 106 cpm [3H]inulinwas added. Of each pair, one ovary was taken out after 2 h, the other after 4 h and, togetherwith 10 /tl samples of the incubation medium, prepared for scintillation counting.
Table 3. Factors influencing in vitro binding of trypan blue
Binding (n moles/cm2)
Modification of standard conditions Control Experimental % effect
pH = 7-5 2-8 1-6 - 4 3-Mg2 + , -Ca2 + , + lmMEDTA 3-7 20 - 4 6Serum albumin (5 mg) 3-2 2-3 - 2 7Dextran sulphate (10 mg) 38 3'1 - 1 7Polyethylene glycol (10mg) 27 2-6 - 3
The two ovaries of one pair were separately incubated for 0-5 h in 1 ml standard salineor modified saline, both containing 0-5 % trypan blue.
a separate experiment (not shown) it was established that uptake was depressedcompared to that in the absence of trypan blue; inhibition ranged between40 and 80 %
In vitro binding of trypan blue
The amount of trypan blue associated with terminal oocytes after in vitroincubation, as in the in vivo experiments, increases proportionally to the oocytesurface (not shown), allowing us to express the trypan blue content in units persurface area in these experiments too.
Oocytes in vitro under the given conditions take up only very small amountsof [3H]inulin; haemolymph does not stimulate, and uptake is virtually finishedwithin 2 h (Table 2). In the presence of 0-5 % trypan blue, values found are evenlower (not shown). This means that most (as can be calculated, more than 90 %)of the trypan blue is externally bound. Thus, the in vitro system should be anappropriate model for the investigation of factors influencing binding.
As can be seen (Table 3), trypan blue binding decreases with external pH andis Mg2+ and Ca2+ dependent; negatively charged macromolecules like serum

Uptake of trypan blue by locust oocytes 9albumin (probably as a serum albumin-trypan blue complex (Lloyd, Beck,Griffiths & Parry, 1968) and dextran sulphate act as competitors, whileneutrally charged polyethylene glycol has no significant effect.
DISCUSSION
In our experience, trypan blue proved to be a quick and convenient indicatorof resorbing oocytes, which could prove useful in other studies concerned withresorption.
Physiologically, trypan blue has toxic properties (Lloyd et al. 1968); as shownfor the cecropia moth (Anderson & Telfer, 1970) and especially relevant here, itinhibits pinocytotic uptake of vitellogenic proteins. In our experiments, thisexpresses itself in the fact that trypan blue suppresses the uptake of inulin, i.e. netvolume uptake. The reason for this may be either that vitellogenic proteins areessential for stimulation of pinocytosis and that trypan blue competes with themfor access to the membrane, or that trypan blue in itself inhibits pinocytosis.
Thus, trypan blue is not the best model compound one could choose forvitellogenin uptake studies. However, the fact that its uptake by the oocytes isproportional to the oocyte surface (Fig. 4B) is of general importance: it suggeststhat this may apply to vitellogenic proteins as well, which means that through-out development the pinocytotic activity per surface area would be constant,and volume increase would be proportional to the oocyte surface. (It should berealized that equation 2, Appendix, gives a value for the enveloping surface, andthat the actual surface involved in pinocytosis is many-fold larger, due to thepresence of microvilli; however, it seems reasonable to assume that the ratiobetween actual and enveloping surface area throughout development is constant.)In fact, via elementary calculus this can already be inferred to be the case fromthe experimental finding (see for instance Highnam & Haskell, 1964, Fig. 1,curve for flown-crowded animals, and Tobe & Pratt, 1975) that oocyte lengthincreases approximately linearly in time; derivation is given in Appendix,equations 4-5. After insertion of the appropriate constants it can be calculatedthat oocytes, in order to complete their development from 1-5 mm length till6-5 mm length in 7 days, should continuously interiorize about 5 nl/mm2
surface/h (Appendix, equation 6), and this is indeed the maximal value calculatedfrom [3H]inulin uptake. This again suggests that the endocytic index for [3H]-inuJin represents the real volume interiorized. Other labelled compounds havebeen used for quantitative determination of pinocytosis, such as [3H]sucrose(Wagner, Rosenberg & Estensen, 1971) and [125I]polyvinylpyrrolidone (Williamset al. 1975), but for our purposes [3H]inulin seems to be preferable because,contrary to sucrose, it is not metabolized in the locust haemolymph, andeventually it can be used in double-labelling experiments together witha [14C-]isotope.
The difference in slope in the trypan blue curve and the inulin curve in Fig. 5

10 E. WAJC AND OTHERS
indicates how much faster trypan blue is taken up than inulin; it also gives theconcentration factor for trypan blue. As the difference in slope is approximatelya factor of 6, the binding contribution in the uptake of trypan blue must beabout 5/6 = 80 % (compare equation 1). In principle, this contribution dependsupon the amount r bound per surface area, and thus upon the concentration oftrypan blue and the presence of competitors (such as vitellogenins).
The trypan blue content at zero time represents strong binding, both to theplasma membrane (functional in pinocytosis) and at other places not relevant foruptake (for instance, in between the follicle cells) (Anderson & Telfer, 1970).Probably, during pinocytosis an additional number of weak binding sites areoccupied, that lose their bound dye molecules upon washing.
From the in vitro experiments (Table 3), it appears that trypan blue is boundby virtue of its negative charge: cations such as Mg2+, Ca2+ and H+ enhancebinding, whilst poly-anions act as competitors (compare also Wallace & Ti,1972). Although our in vivo data show that in principle those electrostaticeffects could play a role in selective uptake, it should be realized that vitellogeninsrelatively are much stronger concentrated in the oocyte yolk than trypan blue;if, like in the cecropia moth (Telfer, 1960), the concentration factor is 20, theuptake of vitellogenins should be 20 x as fast as that of a non-binding com-pound, which means that the binding contribution should amount to 19/20= 95°/
yj / 0 .
Thus, in the binding and uptake of natural vitellogenins factors other thanelectrostatic ones might contribute, such as the presence of specific receptors orthe secretion of follicle cell product previously mentioned (Anderson & Telfer,1970). These factors will be evaluated with vitellogenic proteins which arepresently being isolated and radio-labelled.
This work was supported in part by a grant from the Volkswagenstiftung, Hannover.Drs Esther Wajc and Tilly Bakker-Grunwald were Lady Davis Fellows of The HebrewUniversity of Jerusalem.
A P P E N D I X
Oocytes have the form of a prolate spheroid, with length / = 5 timesdiameter d (Fig. 1). Thus,
Surface, s = n.d.l = 0-62/2 (2)
and volume, v = n {£)* (l ~ f ) = 0-03 /3 (3)
Experimentally (Jaques, 1973; Wagner et al. 1971) it is found that in firstapproximation oocyte length increases linearly in time, from 1-5 mm till6-5 mm in 7 days. Thus,
-T = constant = 0-03 mm/h. (4)

Uptake of trypan blue by locust oocytes 11
The volume increase in time is:
^ = ^ . ^ = 3.0-03 /2.0-03 mm/h = 0-003 /2 mm3/h (1 in mm), (5)
which is seen to be proportional to the surface.The volume increase in time per surface area can be calculated as:
dv 1 0-003/3 _ Ar.c ,, „,, . , „,, ,,.—.- = = 0-005 /d/mm2/h = 5nl mm2/h, (6)at s U-o 1
Compensatory water movements other than pinocytotic volume uptake, thatcould occur because of osmotic effects during the phase transition of solublevitellogenins to dispersed yolk proteins, are not taken into consideration.
REFERENCES
ANDERSON, L. M. & TELFER, W. H. (1970). Trypan blue inhibition of yolk deposition - a clueto follicle cell function in the cecropia moth. / . Embryol. exp. Morph. 23, 35-52.
BAR-ZEV, A., WAJC, E., COHEN, E., SAPIR, L., APPLEBAUM, S. W. & EMMERICH, H. (1975).Vitellogenin accumulation in the fat body and haemolymph of Locusta migratoria inrelation to egg maturation. / . Insect Physiol. 21, 1257-1263.
BENTZ, F., GIRARDIE, A. & CAZAL, M. (1970). Etude electrophoretique des variations de laproteinemie chez Locusta migratoria pendant la maturation sexuelle. / . Insect Physiol. 16,2257-2270.
HIGHNAM, K. C. & HASKELL, P. T. (1964). The endocrine systems of isolated and crowdedLocusta and Schistocerca in relation to oocyte growth, and the effects of flying uponmaturation. / . Insect Physiol. 10, 849-864.
HIGHNAM, K. C, LUSIS, O. & HILL, L. (1963). Factors affecting oocyte resorption in thedesert locust Schistocerca gregaria (Forskal). J. Insect Physiol. 9, 827-837.
JACQUES, P. J. (1973). Endocytosis. In Lysosomes in Biology and Pathology, vol. 2 (ed. J. T.Dingle & H. B. Fell), pp. 395^20. Amsterdam: North-Holland.
LLOYD, J. B., BECK, F., GRIFFITHS, A. & PARRY, L. M. (1968). The mechanism of action ofacid bisazo dyes. In Interaction of Drugs and Subcellular Components (ed. P. N. Campbell),pp. 171-202. London: Churchill Ltd.
MORDUE, W. & GOLDSWORTHY, G. L. (1969). The physiological effects of corpus cardiacumextracts in locusts. Gen. $omp. Endocr. 12, 360-369.
TELFER, W. H. (1960). The selective accumulation of blood proteins by the oocytes ofsaturniid moths. Biol. Bull. mar. biol. Lab., Woods Hole 118, 338-351.
TOBE, S. S. & LOUGHTON, B. G. (1967). The development of blood proteins in the Africanmigratory locust. Can. J. Zool. 45, 975-984.
TOBE, S. S. & PRATT, G. E. (1975). Corpus allatum activity in vitro during ovarian maturationin, the desert locust, Schistocerca gregaria. J. exp. Biol. 62, 611-627.
WAGNER, R., ROSENBERG, M. & ESTENSEN, R. (1971). Endocytosis in Chang liver cells.Quantitation by sucrose-3H uptake and inhibition by cytochalasin B. /. Cell Biol. 50,804-817.
WALLACE, R. H. & Ho, Ti (1972). Protein incorporation by isolated amphibian oocytes.II. A survey of inhibitors. / . exp. Zool. 181, 303-318.
WILLIAMS, K. E., KIDSTON, E. M., BECK, F. & LLOYD, J. B. (1975). Quantitative studies ofpinocytosis. I. Kinetics of uptake of [125I]polyvinylpyrrolidone by rat yolk sac culturedin vitro. J. Cell Biol. 64, 113-122.
(Received 27 November 1975, revised 6 August 1976)


















