
Behavioral Pharmacology of Cannabinoids with a Focus on ...
25
350 Current Drug Abuse Reviews, 2008, 1, 350-374 1874-4737/08 $55.00+.00 © 2008 Bentham Science Publishers Ltd. Behavioral Pharmacology of Cannabinoids with a Focus on Preclinical Models for Studying Reinforcing and Dependence-Producing Properties George Panagis *,1 , Styliani Vlachou 2 and George G. Nomikos 3 1 Laboratory of Behavioral Neuroscience, Department of Psychology, School of Social Sciences, University of Crete, 74100 Rethymno, Crete, Greece 2 Department of Psychiatry, M/C 0603, School of Medicine, University of California, San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0603, USA 3 Cannasat Therapeutics, Toronto, Ontario, MCJ 1C9, Canada Abstract: Cannabis preparations as recreational drugs are the most widely used illicit drugs in the world. Although can- nabis derivatives produce clear subjective motivational responses in humans leading to drug-seeking behavior and in a specific proportion in repeated drug use, the reinforcing/rewarding attributes of these subjective effects are difficult to de- fine in experimental animals. This led to the notion of cannabinoids being considered as "atypical" or "anomalous" drugs of abuse. To this end, our knowledge and understanding of the way cannabis and its main psychoactive constituent, 9 - tetrahydrocannabinol ( 9 -THC), act in the central nervous system to exert their reinforcing/rewarding effects is far from complete. The aim of the present article is to review from a preclinical perspective the current status of what is known about the behavioral pharmacology of cannabinoids including the recently identified cannabinoid neurotransmission modifiers with a particular emphasis on their motivational/reinforcing and dependence-producing properties. We conclude that cannabinoids exhibit reinforcing/rewarding properties in experimental animals mostly under particular experimental conditions, which is not the case for other drugs of abuse, such as opiates, psychostimulants, alcohol and nicotine. The paper will discuss these findings critically and also point to open questions that should be addressed in the future in order to improve our understanding of these specific actions of cannabinoids that will also impact drug discovery and develop- ment efforts of related compounds as therapeutics in the clinic. INTRODUCTION Cannabis derivatives are the most widely used illegal drugs in the world. The popularity of cannabis as a recrea- tional drug is due to its ability to alter sensory processing as well as producing clear subjective motivational responses accompanied with a sense of mild euphoria and well-being. Indeed, this elation along with the sense of relaxation pro- duced by cannabis, are among the most cited subjective psy- chotropic properties of the drug [1]. It has been suggested that in a proportion of regular users, these effects lead to drug-seeking behavior and repeated drug use [2-4]. On the other hand, the ability of cannabis preparations to cause a variety of medicinal effects unrelated to its psycho- tropic effects has been recognized for more than 3000 years now [5]. The last years in many countries, public debate has focused on the possible legalization of cannabis and 9 -THC for therapeutic uses. Because the largest impediment to the development of therapeutic cannabinoid medications is the occurrence of euphoric side effects, which may lead to their abuse in humans, understanding the mechanisms through which these actions occur as well as their neuronal substrate may be essential to developing rational approaches to can- nabinoid medication development. In spite of the well estab- lished abuse properties of cannabis preparations in humans, knowledge about the abuse liability of 9 -THC and other *Address correspondence to this author at the University of Crete, School of Social Sciences, Department of Psychology, Laboratory of Behavioral Neu- roscience, 74100 Rethymno, Crete, Greece; Fax: (30) (28310) 77548; E-mail: [email protected] cannabinoid analogs in experimental animals is still limited, and undoubtedly this issue remains controversial. Conse- quently, our understanding of the way marijuana and its main psychoactive constituent, 9 -THC, act to exert their pleasurable effects in humans is far from complete. In this review, evidence on the actions of cannabis, its main psychoactive ingredient 9 -THC, and more potent syn- thetic analogs and cannabinoid neurotransmission modifiers on brain reward circuitry will be provided and the possible underlying mechanisms will be discussed. CANNABINOIDS AND THE ENDOCANNABINOID SYSTEM Herbal cannabis contains over 400 compounds including over 60 terpeno-phenols, unique to the plant of cannabis, known as plant cannabinoids. One of these constituents, 9 - THC, has been the object of intense investigation [6, 7], as it is, by far, the major psychoactive constituent of the plant. From the other plant cannabinoids, cannabidiol, a non- psychoactive ingredient, has also been widely investigated as it exhibits interesting anti-inflammatory, anti-schizophrenic and antiepileptic actions [8]. The pharmacology and the be- havioral actions of other plant cannabinoids are largely un- known. 9 -THC and other cannabinoids are highly lipophilic and were originally thought to act by simple diffusion through cell membranes, resulting in relatively non-selective disrup- tion of neurotransmission [9]; the last years, however, our knowledge about the cannabinoid system has increased enormously [5]. During the past decade, two cannabinoid
Transcript of Behavioral Pharmacology of Cannabinoids with a Focus on ...

350 Current Drug Abuse Reviews, 2008, 1, 350-374
1874-4737/08 $55.00+.00 © 2008 Bentham Science Publishers Ltd.
Behavioral Pharmacology of Cannabinoids with a Focus on Preclinical Models for Studying Reinforcing and Dependence-Producing Properties
George Panagis*,1
, Styliani Vlachou
2 and George G. Nomikos
3
1Laboratory of Behavioral Neuroscience, Department of Psychology, School of Social Sciences, University of Crete,
74100 Rethymno, Crete, Greece
2Department of Psychiatry, M/C 0603, School of Medicine, University of California, San Diego, 9500 Gilman Drive, La
Jolla, CA 92093-0603, USA
3Cannasat Therapeutics, Toronto, Ontario, MCJ 1C9, Canada
Abstract: Cannabis preparations as recreational drugs are the most widely used illicit drugs in the world. Although can-
nabis derivatives produce clear subjective motivational responses in humans leading to drug-seeking behavior and in a
specific proportion in repeated drug use, the reinforcing/rewarding attributes of these subjective effects are difficult to de-
fine in experimental animals. This led to the notion of cannabinoids being considered as "atypical" or "anomalous" drugs
of abuse. To this end, our knowledge and understanding of the way cannabis and its main psychoactive constituent, 9-
tetrahydrocannabinol (9-THC), act in the central nervous system to exert their reinforcing/rewarding effects is far from
complete. The aim of the present article is to review from a preclinical perspective the current status of what is known
about the behavioral pharmacology of cannabinoids including the recently identified cannabinoid neurotransmission
modifiers with a particular emphasis on their motivational/reinforcing and dependence-producing properties. We conclude
that cannabinoids exhibit reinforcing/rewarding properties in experimental animals mostly under particular experimental
conditions, which is not the case for other drugs of abuse, such as opiates, psychostimulants, alcohol and nicotine. The
paper will discuss these findings critically and also point to open questions that should be addressed in the future in order
to improve our understanding of these specific actions of cannabinoids that will also impact drug discovery and develop-
ment efforts of related compounds as therapeutics in the clinic.
INTRODUCTION
Cannabis derivatives are the most widely used illegal drugs in the world. The popularity of cannabis as a recrea-tional drug is due to its ability to alter sensory processing as well as producing clear subjective motivational responses accompanied with a sense of mild euphoria and well-being. Indeed, this elation along with the sense of relaxation pro-duced by cannabis, are among the most cited subjective psy-chotropic properties of the drug [1]. It has been suggested that in a proportion of regular users, these effects lead to drug-seeking behavior and repeated drug use [2-4].
On the other hand, the ability of cannabis preparations to cause a variety of medicinal effects unrelated to its psycho-tropic effects has been recognized for more than 3000 years now [5]. The last years in many countries, public debate has focused on the possible legalization of cannabis and
9-THC
for therapeutic uses. Because the largest impediment to the development of therapeutic cannabinoid medications is the occurrence of euphoric side effects, which may lead to their abuse in humans, understanding the mechanisms through which these actions occur as well as their neuronal substrate may be essential to developing rational approaches to can-nabinoid medication development. In spite of the well estab-lished abuse properties of cannabis preparations in humans, knowledge about the abuse liability of
9-THC and other
*Address correspondence to this author at the University of Crete, School of
Social Sciences, Department of Psychology, Laboratory of Behavioral Neu-
roscience, 74100 Rethymno, Crete, Greece; Fax: (30) (28310) 77548;
E-mail: [email protected]
cannabinoid analogs in experimental animals is still limited, and undoubtedly this issue remains controversial. Conse-quently, our understanding of the way marijuana and its main psychoactive constituent,
9-THC, act to exert their
pleasurable effects in humans is far from complete.
In this review, evidence on the actions of cannabis, its main psychoactive ingredient
9-THC, and more potent syn-
thetic analogs and cannabinoid neurotransmission modifiers on brain reward circuitry will be provided and the possible underlying mechanisms will be discussed.
CANNABINOIDS AND THE ENDOCANNABINOID SYSTEM
Herbal cannabis contains over 400 compounds including over 60 terpeno-phenols, unique to the plant of cannabis, known as plant cannabinoids. One of these constituents,
9-
THC, has been the object of intense investigation [6, 7], as it is, by far, the major psychoactive constituent of the plant. From the other plant cannabinoids, cannabidiol, a non-psychoactive ingredient, has also been widely investigated as it exhibits interesting anti-inflammatory, anti-schizophrenic and antiepileptic actions [8]. The pharmacology and the be-havioral actions of other plant cannabinoids are largely un-known.
9-THC and other cannabinoids are highly lipophilic and
were originally thought to act by simple diffusion through cell membranes, resulting in relatively non-selective disrup-tion of neurotransmission [9]; the last years, however, our knowledge about the cannabinoid system has increased enormously [5]. During the past decade, two cannabinoid

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 351
receptors, named CB1 [10] and CB2 [11], were cloned and their distribution and transduction pathways were described [12]. The CB1 receptor is highly expressed in the central nervous system whereas the CB2 receptor is primarily found in cells of the immune system [13]. However, recently the CB2 receptor has also been located in specific brain regions [14, 15]. In addition to the CB1 and CB2 receptors, pharma-cological evidence has been accumulating over the years to support the existence of more cannabinoid receptors [16].
The presence of cannabinoid receptors in mammalians suggested the existence of endogenous ligands that bind and affect the function of these receptors. In 1992, the first en-dogenous ligand for these receptors, named anandamide (N-arachidonoyl ethanolamide or AEA), was isolated [17]. Anandamide could be synthesized by neurons upon stimula-tion and it mimicked many biochemical, pharmacological and behavioral properties of cannabinoids [18, 19]. Three years after the discovery of anadamide, another endogenous ligand for the cannabinoid receptor, named 2-arachidonoyl glycerol (2-AG), was identified [20]. More recently, other molecules with cannabinoid receptor binding activity termed 2-arachidonoyl glyceryl ether (noladine ether), O-arachido-noyl ethanolamine (virodhamine), and N-arachidonoyl-dopamine (NADA) have been discovered (for a recent re-view, see [21]) and their physiological actions remain to be elucidated. Endogenous cannabinoids are produced from membrane-associated precursors and exert their neurochemi-cal actions by binding and stimulating cannabinoid and va-nilloid receptors [22]. Their concentrations are regulated in a strict manner through a reuptake process into cells and a rapid enzymatic inactivation [23, 24].
The cannabinoid receptors, the endocannabinoids, the proteins that synthesize endocannabinoids and the proteins of the inactivation process together constitute what is now referred to as the “endocannabinoid system”. This system modulates various physiological functions not only in the central nervous system but also in the autonomic nervous system, and in the gastrointestinal, the endocrine, the repro-ductive and the immune systems, as well as in the circulation [19].
The development of potent and selective agonists and antagonists for the cannabinoid receptors or agents that block the reuptake or degradation of endocannabinoids has played a major role in the recent advances obtained in behavioral and pharmacological research of the cannabinoid system [25, 26]. Genetically modified mice have also been used as re-search tools to study the behavioral and pharmacological actions of cannabinoids (reviewed by [27, 28]).
PHARMACOLOGICAL AND MOLECULAR TOOLS TO STUDY ENDOCANNABINOID NEUROTRANS-MISSION
The discovery of the cannabinoid receptors, the endocan-nabinoid ligands and associated enzymes which are respon-sible for their metabolism and deactivation has generated a great interest in developing novel cannabinoid therapeutic drugs. Many of them have been used extensively as pharma-cological tools to study endocannabinoid neurotransmission and pharmacology. Currently, five major classes of cannabi-noid analogues have been identified based on chemical struc-ture [26, 29-31].
Classical cannabinoids are tricyclic terpenoid derivatives bearing a benzopyran moiety. This group includes cannabis-derived compounds (phytocannabinoids) and synthetic ana-logues. The major phytocannabinoids included in this group are
9-THC,
8-THC and cannabinol. From the synthetic
analogues notable examples are the non-selective CB1/CB2
receptor agonists, 11-hydroxy-8-THC-dimethylheptyl (HU-
210), levonantradol, nabilone, AMG-3 and the selective CB2
receptor agonists L-759633, L-759656 and JWH-133.
Non-classical cannabinoids consist of bicyclic and tri-cyclic analogues of
9-THC lacking the pyran ring of classi-
cal cannabinoids. Notable examples include the potent non-selective CB1/CB2 receptor agonists CP-55,940, CP-47,497, CP-55,244 and the selective CB2 receptor agonist HU-308. [
3H]CP-55,940 was the key compound that led to the discov-
ery of the CB1 receptor [32].
The class of aminoalkylindoles has a completely different structure from the other cannabinoids. They also differ in their lipophilicity (they are considerable less lipophilic) as well as in the way they activate cannabinoid receptors [33]. The most widely investigated compound from the aminoal-kylindoles is the potent CB1/CB2 receptor agonist WIN55,212-2. This compound has high stereoselectivity and slight preference for the CB2 receptor. Other compounds of this class include the potent and selective CB2 receptor ago-nist AM1241 and the selective CB2 receptor antagonist AM630.
The eicosanoid group consists of cannabinoid receptor agonists that have markedly different structures both from aminoalkylindoles and from classical and non-classical can-nabinoids. Notable members of this group are the endocan-nabinoids, anandamide, 2-AG, virodhamine, noladin ether and several synthetic analogues of anandamide, including R-(+)-methanandamide, arachidonyl-2’-chloroethylamide (A CEA), and arachidonylcyclopropylamide (ACPA).
The fifth class of 1,5 biarylpyrazoles includes cannabi-noid-related compounds that act as cannabinoid receptor antagonists. SR141716A (rimonabant) was developed at Sanofi as the first CB1 receptor antagonist and has since been used extensively as pharmacological tool in many studies. This compound shows selectivity for the CB1 receptor and has often been reported to act as an inverse agonist rather than a neutral antagonist [34]. The follow-on synthesis of AM251 with a selectivity for the CB1 receptor presented re-searchers with a similarly potent antagonist/inverse agonist. SR144528 was also developed at Sanofi as a CB2 receptor antagonist.
The finding that the actions of anandamide and 2-arachidonoyl glycerol are terminated by cellular uptake and intracellular enzymatic hydrolysis has been followed by the development of several drugs that inhibit these processes [35, 36]. Several of these drugs have been used as pharma-cological tools in animal experiments aimed at elucidating the pathophysiological roles of endocannabinoids. Important members of this group are the FAAH inhibitors PMSF, palmitylsulphonyl fluoride (AM374), stearylsulphonyl fluo-ride (AM381), O-1887, URB-532 and URB-597. Among the anandamide transport inhibitors, AM-404 is the most exten-sively studied. Other compounds of this group, with a more
352 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
selective action in the transporter than AM-404 are OMDM-1, OMDM-2, and VDM-11
One important advance has also been the development of transgenic mice that lack CB1, CB2 or both CB1 and CB2 re-ceptors [12]. The availability of such animals provide a use-ful additional tool to study whether or not responses to can-nabinoid compounds are CB1 receptor and/or CB2 receptor mediated as well as the physiological roles of CB1 and CB2
receptors. FAAH-KO mice represent another powerful model system in the study of the neurochemical and behav-ioral consequences of endocannabinoid catabolism [36].
CANNABINOID EFFECTS ON BRAIN STIMULA-
TION REWARD
The intracranial self-stimulation (ICSS) paradigm offers the most direct measurement of drug effects on brain reward substrates [37-39]. This behavioral paradigm is based on the discovery by Olds and Milner that rats will repeatedly press a lever to stimulate components of the brain reward circuit. It has been shown that most drugs of abuse are able to lower brain stimulation reward threshold [48-51], which support the notion that they activate the same substrate with electri-cal stimulation in a synergistic manner [39, 40].
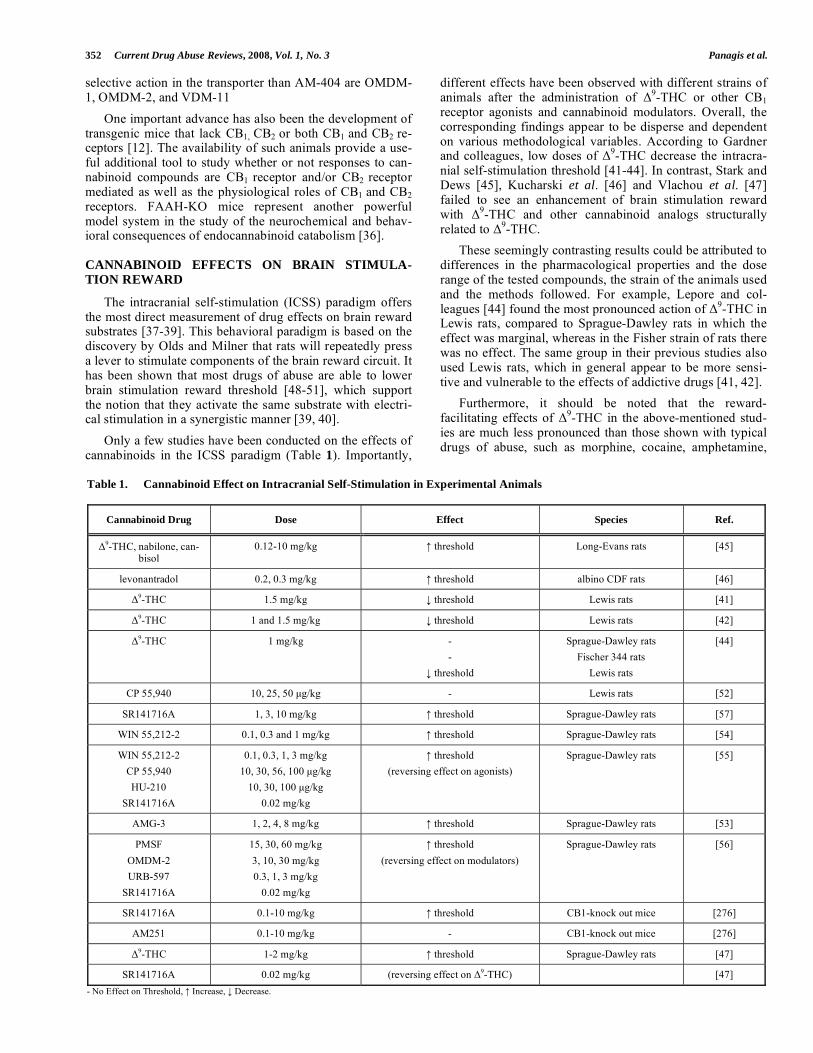
Only a few studies have been conducted on the effects of cannabinoids in the ICSS paradigm (Table 1). Importantly,
different effects have been observed with different strains of animals after the administration of
9-THC or other CB1
receptor agonists and cannabinoid modulators. Overall, the corresponding findings appear to be disperse and dependent on various methodological variables. According to Gardner and colleagues, low doses of
9-THC decrease the intracra-
nial self-stimulation threshold [41-44]. In contrast, Stark and Dews [45], Kucharski et al. [46] and Vlachou et al. [47] failed to see an enhancement of brain stimulation reward with
9-THC and other cannabinoid analogs structurally
related to 9-THC.
These seemingly contrasting results could be attributed to differences in the pharmacological properties and the dose range of the tested compounds, the strain of the animals used and the methods followed. For example, Lepore and col-leagues [44] found the most pronounced action of
9-THC in
Lewis rats, compared to Sprague-Dawley rats in which the effect was marginal, whereas in the Fisher strain of rats there was no effect. The same group in their previous studies also used Lewis rats, which in general appear to be more sensi-tive and vulnerable to the effects of addictive drugs [41, 42].
Furthermore, it should be noted that the reward-facilitating effects of
9-THC in the above-mentioned stud-
ies are much less pronounced than those shown with typical drugs of abuse, such as morphine, cocaine, amphetamine,
Table 1. Cannabinoid Effect on Intracranial Self-Stimulation in Experimental Animals
Cannabinoid Drug Dose Effect Species Ref.
9-THC, nabilone, can-bisol
0.12-10 mg/kg threshold Long-Evans rats [45]
levonantradol 0.2, 0.3 mg/kg threshold albino CDF rats [46]
9-THC 1.5 mg/kg threshold Lewis rats [41]
9-THC 1 and 1.5 mg/kg threshold Lewis rats [42]
9-THC 1 mg/kg -
-
threshold
Sprague-Dawley rats
Fischer 344 rats
Lewis rats
[44]
CP 55,940 10, 25, 50 μg/kg - Lewis rats [52]
SR141716A 1, 3, 10 mg/kg threshold Sprague-Dawley rats [57]
WIN 55,212-2 0.1, 0.3 and 1 mg/kg threshold Sprague-Dawley rats [54]
WIN 55,212-2
CP 55,940
HU-210
SR141716A
0.1, 0.3, 1, 3 mg/kg
10, 30, 56, 100 μg/kg
10, 30, 100 μg/kg
0.02 mg/kg
threshold
(reversing effect on agonists)
Sprague-Dawley rats [55]
AMG-3 1, 2, 4, 8 mg/kg threshold Sprague-Dawley rats [53]
PMSF
OMDM-2
URB-597
SR141716A
15, 30, 60 mg/kg
3, 10, 30 mg/kg
0.3, 1, 3 mg/kg
0.02 mg/kg
threshold
(reversing effect on modulators)
Sprague-Dawley rats [56]
SR141716A 0.1-10 mg/kg threshold CB1-knock out mice [276]
AM251 0.1-10 mg/kg - CB1-knock out mice [276]
9-THC 1-2 mg/kg threshold Sprague-Dawley rats [47]
SR141716A 0.02 mg/kg (reversing effect on 9-THC) [47]
- No Effect on Threshold, Increase, Decrease.

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 353
MDMA or nicotine [48-51]. These later drugs can reliably shift the rate-frequency function to the left by between 0.2 and 0.5 log units. However in the study by Lepore et al. [44],
9-THC could only shift the rate-frequency function to the
left by approximately 0.05 log units. Additionally, Lepore et al. [44] adopt a very strict criterion of stable responding (i.e., 0.01 log units for three consecutive days), which indicates that the observed effect of
9-THC in their study could re-
flect normal baseline variation over days. In most of the studies utilizing the ICSS procedure the criterion of stable responding is defined within 0.1 log units over three con-secutive days. At least some of this apparent “discrepancy” can be also explained by the method Lepore et al. [44] used to analyze their data. For example, depending on the crite-rion used to obtain the critical measurement of threshold in the ICSS paradigm their results appear different. Thus, in Sprague-Dawley rats,
9-THC exhibits reward facilitating
effects only with the 0 criterion, and not the M50 criterion. It is worth noting that Vlachou et al. [47] analyzed their data with a criterion that resembles the M50 criterion. In this way, their data are in agreement with the data of Lepore et al. in the Sprague-Dawley strain of rats [44].
A number of studies have examined the effects of various synthetic cannabinoid agonists and antagonists on brain stimulation reward. Arnold and colleagues [52] have re-ported that the CB1 receptor agonist CP55,940 did not affect the reinforcing properties of medial forebrain bundle (MFB) self-stimulation. Similarly, in a series of studies from our laboratory we have shown that the CB1 receptor agonists WIN55,212-2, CP55,940, HU-210 and AMG-3 either do not affect or increase intracranial self-stimulation threshold, de-pending on the dose used [53-55]. In an analogous manner, we have recently shown that the indirect cannabinoid ago-nists PMSF, AM-404, OMDM-2 and URB-597 either do not affect or increase intracranial self-stimulation threshold, de-pending on the dose used [56]. Low doses of the CB1 recep-tor antagonists SR141716A and AM-251 do not appear to affect brain reward threshold [53, 55, 56]. On the other hand, it has been shown that high and likely non-selective doses of the CB1 receptor antagonist SR141716A increase brain stimulation reward threshold [52, 57]. However, in such high doses it is possible that SR141716A could act as a partial or inverse agonist at cannabinoid receptors, as it has been ob-served in other studies [58-60]. This could be a plausible explanation for its anhedonic effects observed with high doses on brain stimulation reward. Interestingly, clinical studies indicate that SR141716A (rimonabant) may induce anhedonic states and depressed mood [61].
In summary, the majority of the studies using the ICSS paradigm failed to show any reward facilitating effects for
9-THC or other cannabinoids, or on the contrary, they pre-
sent data for anhedonic actions of these compounds, at least in the high dose range. In the studies by Gardner’s group the reward facilitating effect of
9-THC in the ICSS paradigm
was only found in rats of the Lewis strain. Although one can speculate that the reinforcing effect of cannabinoids may only be obtained in certain strains of rat, suggesting an im-portant genetic component in this action, other studies have shown that
9-THC produces aversion in the conditioned
place preference paradigm [62] in the same strain of rats, as discussed below.
CANNABINOID SELF-ADMINISTRATION
Human subjects and laboratory animals will self-administer addictive drugs by a variety of routes, including oral, intragastric, intraperitoneal, and intracranial routes. Intravenous drug self-administration has been one of the most direct approaches to study the rewarding properties of abused drugs in experimental animals, whether rodents or primates. In this behavioral paradigm, animals are given the opportunity to self-administer intravenously a drug by mak-ing an operant response, such as pressing a lever in an oper-ant chamber or inserting their nose into a hole [63-66]. In other words, the model studies directly drug-taking behavior in sub-human subjects. Reinforcing effects of a drug as-sessed by intravenous self-administration procedures in ex-perimental animals are considered one of the most reliable predictors of abuse potential in human subjects.
Generally, most drugs abused by humans, including psy-chostimulants, opiates, ethanol, and nicotine, support reliable and persistent self-administration behavior in drug naïve experimental animals [67-69]. However, for many years, it has been rather difficult to show self-administration of can-nabis or
9-THC in rodents or primates [70-76]. Interest-
ingly, different research groups have varied the parameters of self-administration procedure in order to demonstrate reli-able and persistent self-administration of
9-THC or other
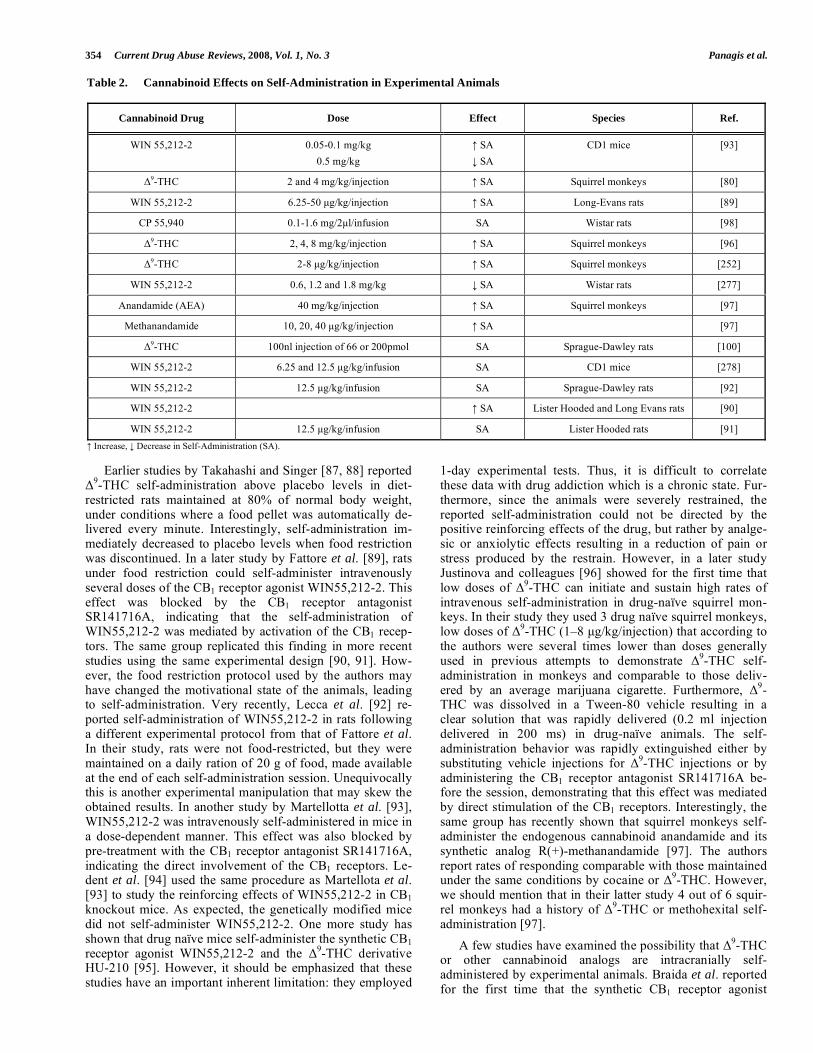
synthetic cannabinoids in experimental animals (Table 2).
The first successful self-administration of cannabis is reported by Deneau and Kaymakcalan [77] and Kaymakca-lan [78], who demonstrated acquisition of
9-THC self-
administration behavior in two monkeys out of six studied, but only after withdrawal from forced automatic i.v. injec-tions of
9-THC, when signs of physical dependence on
9-
THC occurred. Furthermore, one monkey exhibited 9-THC
self-administration behavior following cocaine self-administration. Naïve monkeys did not self-administer
9-
THC. Other studies also have shown self-administration of 9-THC in experimental animals pretreated with phencycli-
dine [79] or cocaine [80]. Furthermore, in the study by Pick-ens et al. [79] rates of responding were relatively low and there was no clear evidence that responding for
9-THC
could persist above vehicle control levels over repeated daily sessions. After all, as it has been shown, previous drug expo-sure can be an important factor in modulating acquisition and maintenance of self-administration of various abused drugs in experimental animals [67, 81]. In other words, the functional state as well as the motivational state in naïve animals compared with animals that self-administer other drugs could be different, and therefore their corresponding response could also vary accordingly. Interestingly, other studies have failed to show robust
9-THC self-
administration, even in animals pre-exposed to other drugs of abuse, such as cocaine, phenobarbital, phencyclidine, ethanol or amphetamine [72, 75].
In another frequently used procedure to study the self-administration of cannabinoids food deprivation was used to initiate and subsequently maintain drug self-administration. However, this manipulation may also alter the motivational state of the animal, which per se is an inherent limitation. Indeed, as it has been repeatedly shown food restriction can facilitate the initiation of drug self-administration [82-86].
354 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
Earlier studies by Takahashi and Singer [87, 88] reported 9-THC self-administration above placebo levels in diet-
restricted rats maintained at 80% of normal body weight, under conditions where a food pellet was automatically de-livered every minute. Interestingly, self-administration im-mediately decreased to placebo levels when food restriction was discontinued. In a later study by Fattore et al. [89], rats under food restriction could self-administer intravenously several doses of the CB1 receptor agonist WIN55,212-2. This effect was blocked by the CB1 receptor antagonist SR141716A, indicating that the self-administration of WIN55,212-2 was mediated by activation of the CB1 recep-tors. The same group replicated this finding in more recent studies using the same experimental design [90, 91]. How-ever, the food restriction protocol used by the authors may have changed the motivational state of the animals, leading to self-administration. Very recently, Lecca et al. [92] re-ported self-administration of WIN55,212-2 in rats following a different experimental protocol from that of Fattore et al. In their study, rats were not food-restricted, but they were maintained on a daily ration of 20 g of food, made available at the end of each self-administration session. Unequivocally this is another experimental manipulation that may skew the obtained results. In another study by Martellotta et al. [93], WIN55,212-2 was intravenously self-administered in mice in a dose-dependent manner. This effect was also blocked by pre-treatment with the CB1 receptor antagonist SR141716A, indicating the direct involvement of the CB1 receptors. Le-dent et al. [94] used the same procedure as Martellota et al. [93] to study the reinforcing effects of WIN55,212-2 in CB1 knockout mice. As expected, the genetically modified mice did not self-administer WIN55,212-2. One more study has shown that drug naïve mice self-administer the synthetic CB1 receptor agonist WIN55,212-2 and the
9-THC derivative
HU-210 [95]. However, it should be emphasized that these studies have an important inherent limitation: they employed
1-day experimental tests. Thus, it is difficult to correlate these data with drug addiction which is a chronic state. Fur-thermore, since the animals were severely restrained, the reported self-administration could not be directed by the positive reinforcing effects of the drug, but rather by analge-sic or anxiolytic effects resulting in a reduction of pain or stress produced by the restrain. However, in a later study Justinova and colleagues [96] showed for the first time that low doses of
9-THC can initiate and sustain high rates of
intravenous self-administration in drug-naïve squirrel mon-keys. In their study they used 3 drug naïve squirrel monkeys, low doses of
9-THC (1–8 μg/kg/injection) that according to
the authors were several times lower than doses generally used in previous attempts to demonstrate
9-THC self-
administration in monkeys and comparable to those deliv-ered by an average marijuana cigarette. Furthermore,
9-
THC was dissolved in a Tween-80 vehicle resulting in a clear solution that was rapidly delivered (0.2 ml injection delivered in 200 ms) in drug-naïve animals. The self-administration behavior was rapidly extinguished either by substituting vehicle injections for
9-THC injections or by
administering the CB1 receptor antagonist SR141716A be-fore the session, demonstrating that this effect was mediated by direct stimulation of the CB1 receptors. Interestingly, the same group has recently shown that squirrel monkeys self-administer the endogenous cannabinoid anandamide and its synthetic analog R(+)-methanandamide [97]. The authors report rates of responding comparable with those maintained under the same conditions by cocaine or
9-THC. However,
we should mention that in their latter study 4 out of 6 squir-rel monkeys had a history of
9-THC or methohexital self-
administration [97].
A few studies have examined the possibility that 9-THC
or other cannabinoid analogs are intracranially self-administered by experimental animals. Braida et al. reported for the first time that the synthetic CB1 receptor agonist
Table 2. Cannabinoid Effects on Self-Administration in Experimental Animals
Cannabinoid Drug Dose Effect Species Ref.
WIN 55,212-2 0.05-0.1 mg/kg
0.5 mg/kg
SA
SA
CD1 mice [93]
9-THC 2 and 4 mg/kg/injection SA Squirrel monkeys [80]
WIN 55,212-2 6.25-50 μg/kg/injection SA Long-Evans rats [89]
CP 55,940 0.1-1.6 mg/2μl/infusion SA Wistar rats [98]
9-THC 2, 4, 8 mg/kg/injection SA Squirrel monkeys [96]
9-THC 2-8 μg/kg/injection SA Squirrel monkeys [252]
WIN 55,212-2 0.6, 1.2 and 1.8 mg/kg SA Wistar rats [277]
Anandamide (AEA) 40 mg/kg/injection SA Squirrel monkeys [97]
Methanandamide 10, 20, 40 μg/kg/injection SA [97]
9-THC 100nl injection of 66 or 200pmol SA Sprague-Dawley rats [100]
WIN 55,212-2 6.25 and 12.5 μg/kg/infusion SA CD1 mice [278]
WIN 55,212-2 12.5 μg/kg/infusion SA Sprague-Dawley rats [92]
WIN 55,212-2 SA Lister Hooded and Long Evans rats [90]
WIN 55,212-2 12.5 μg/kg/infusion SA Lister Hooded rats [91]
Increase, Decrease in Self-Administration (SA).

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 355
CP55,940 is self-administered intracerebrally by rats in a free-choice procedure [98]. This effect was antagonized by the CB1 receptor antagonist SR141716A, indicating that it was specifically mediated by the CB1 receptors. However, one limitation of this study is that the animals were water-deprived and water was concurrently delivered with each infusion. This may have altered the motivational state of the animals, provoking the self-administration response. It should be emphasized that CP55,940 was not self-adminis-tered by rhesus monkeys in the study by Mansbach et al. [75]. In a follow-up study, Braida and colleagues reported intracerebroventricular self-administration of
9-THC in the
rat, although under water deprivation conditions, as in their previous study [99]. Recently, Zangen et al. [100] identified two possible brain sites for the rewarding effects of the re-ported intracerebral self-administration of
9-THC: the pos-
terior ventral tegmental area and the shell of the nucleus ac-cumbens. In their study, animals self-administered
9-THC
directly into the posterior ventral tegmental area and the shell of the nucleus accumbens, but not in the core of the nucleus accumbens or the anterior ventral tegmental area or the re-gion dorsal to this.
To conclude, most attempts to obtain a robust self-admin-istration of
9-THC or other synthetic cannabinoids, under
normal experimental conditions (i.e., drug naïve animals, not restrained and without previous drug exposure or food depri-vation), have been negative. Only a limited number of stud-ies report a robust procedure for cannabinoid self-admin-istration either in a limited number of squirrel monkeys or intracerebrally in rodents. This illustrates the differential status of cannabinoids as atypical drugs of abuse.
CANNABINOIDS EFFECTS ON CONDITIONED PLACE PREFERENCE/AVERSION
The conditioned place preference (CPP) procedure pro-vides an animal model of the subjective, motivational effects of drugs as well as non-drug stimuli [101-104]. A drug is injected and the animal is placed in a test chamber with dis-tinctive environmental cues (e.g., tactile, visual, and/or olfac-tory). This procedure is repeated for several days. During these conditioning trials the animal learns an association between the subjective state produced by the drug (e.g., a heightened feeling of euphoria comparable to pleasure in humans) and the environmental cues present during the drug state. When the subject is tested in an apparatus that contains the drug-related environmental cues in one compartment and neutral cues in another, it voluntarily spends more time in the compartment associated with the drug-related cues. In other words, in the CPP procedure neutral environmental cues acquire incentive salience which is closely related to the subjective state produced by the drug. Although the CPP paradigm does not directly measure drug reinforcement, it can be used to infer the motivational values of addictive drugs. Indeed, the concordance with intracranial self-stimulation and self-administration studies is fairly good [102, 104, 105]. Thus, CPP produced by a drug indicates that it exhibits rewarding properties and that it might therefore have abuse potential in humans [105]. A similar place condi-tioning procedure can also be used to study the aversive properties of a drug or a non-drug stimulus. In this case, the animal will avoid staying in a compartment previously asso-ciated with a drug producing aversive or dysphoric effects.
While most drugs abused by humans are able to increase the time spent in the drug-associated compartment, produc-ing a clear place preference, the corresponding data on can-nabinoids are not very consistent. Indeed,
9-THC and other
synthetic cannabinoid agonists can induce both appetitive and aversive effects in the CPP paradigm under various ex-perimental conditions (Table 3). Notably, in most of the studies reporting place preference of cannabinoids these ef-fects are usually dependent upon the particular dose used and the preference is connected to a single dose. However, this is not the case with typical drugs of abuse, in which place pref-erence can be obtained over a range of doses [101-104]. Fur-thermore, the timing between injections has been suggested as another important factor determining whether cannabi-noids produce preference or aversion [106], since pharma-cokinetic factors (i.e. factors related to bioavailability of the drug formulation used, lipophilicity, distribution and storage in fat tissue, metabolism and elimination rate) seem to be responsible in part for the subjective effects of cannabinoids.
A number of studies in rodents failed to show that 9-
THC produces conditioned place preference, or revealed clear aversive phenomena [62, 107-113]. In an attempt to show CPP on
9-THC or other cannabinoids many groups
manipulated the experimental conditions, and more specifi-cally the timing of injections as well as the range of the doses used.
The first report in which 9-THC produced conditioned
place preference was published in 1995 by Lepore et al. [106]. In their study, 2-4 mg/kg
9-THC produced condi-
tioned place preference in Long Evans rats when the CPP pairing interval was 24 h; using the same experimental de-sign, 1 mg/kg of
9-THC did not produce any preference.
However, in the same study, when the schedule of daily in-jections was changed, allowing a longer wash-out interval between injections (48 h),
9-THC produced a clear place
aversion in the dose of 2-4 mg/kg, but place preference in the dose of 1 mg/kg. The authors attributed these differential responses to the different methodological conditions fol-lowed, such as the duration and the number of sessions. In this regard, increasing the time interval between
9-THC
injections might have changed the effects of 9-THC due to
a post-drug dysphoric rebound effect. In other words, at the shorter pairing interval the post-
9-THC rebound dysphoria
might have attenuated 9-THC’s rewarding effect, while at
the longer pairing interval, where the post-9-THC rebound
effect subsided, the effects of all doses were accentuated, allowing the 1 mg/kg to be rewarding and the highest doses to be aversive. In any case, these unusual results emphasize for one more time that
9-THC is an atypical, if not an
anomalous, drug of abuse.
Similarly, in a study by Braida et al. [99] 9-THC pro-
duced conditioned place preference in Wistar rats in a dose range between 0.075 and 0.75 mg/kg. In the same study, higher doses of
9-THC (6 mg/kg) produced clear aversive
effects. The authors suggested that the low doses of 9-THC
used in their experiment could have minimized any dysphoric effects of the drug that might lead to aversion. More recently, Le Foll et al. [114] reported that a low 0.1 mg/kg dose of
9-THC produced conditioned place prefer-
ence. Doses lower (i.e., 0.01 mg/kg) or higher (i.e., 1 mg/kg) than this did not produce any preference.

356 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
Valjent and Maldonado in their study [115] bypassed the dysphoric/aversive effects of
9-THC that has been reported
in naïve animals by giving the animals a priming injection 24h before the first conditioning session. In this way,
9-
THC induced clear CPP in mice. However, a more recent study by Vlachou et al. [47], in which the authors used the same experimental manipulation, no conditioned place pref-erence was observed with the 1 mg dose of
9-THC. This
inconsistency in results could be attributable to the different strain of animals used, the number of pairings or the periods of conditioning and administration of the drugs. Mal-donado’s group replicated their findings in a more recent
study [116]. However, it is worth noting that in the latter study 5 mg/kg
9-THC produced conditioned place aversion.
Conflicting results have also been reported in the litera-ture for synthetic direct and indirect cannabinoid agonists. McGregor et al. [117] reported aversive effects of the syn-thetic CB1 receptor agonist CP55,940 in the CPP paradigm both in a very low dose of 10 μg/kg as well as in a higher dose of 100 μg/kg. In the same study, only the dose of 100 μg/kg was aversive in the conditioned taste aversion para-digm. It is worth noting that in the ICSS paradigm, Vlachou et al. [55] observed a clear anhedonic effect for the CP55,940 compound when administered in the dose of 100
Table 3. Cannabinoid Effects on Conditioned Place Preference in Experimental Animals
Cannabinoid Drug Dose Effect Species Ref.
9-THC 1mg/kg
2 and 4 mg/kg
2 and 4 mg/kg (wash-out period)
1 mg/kg
CPA
CPP
CPA
CPP
Long-Evans rats [106]
9-THC 1 mg/ml CPA Lewis and Sprague-Dawley rats [62]
CP 55,940 100 μg/kg CPA Wistar rats [117]
9-THC 1.5 mg/kg
15 mg/kg
-
CPA
Sprague-Dawley rats [112]
WIN 55,212-2
SR141716A
0.3-1 mg/kg
up to 10mg/kg
CPA
-
(reversing effect on
WIN 55,212-2)
Wistar rats [118]
9-THC 20 mg/kg CPA CD1 mice [109]
9-THC
Anandamide (AEA)
1 and 1.5 mg/kg
up to 16 mg/kg
CPA
-
Wistar rats [110]
HU-210 9-THC
20, 60, 100 μg/kg
1.5 mg/kg
CPA
CPA
Lister Hooded rats [107]
9-THC 5 mg/kg
1 mg/kg
5 mg/kg (not standard protocol-
pretreatment)
1 mg/kg
CPA
-
-
CPP
CD1 mice [115]
CP 55,940
SR141716A
20 μg/kg
0.5 mg/kg
CPP
-
(reversing effect on CP
55,940)
Wistar rats [98]
9-THC 5 mg/kg - dynorphin deficient mice [113]
9-THC
SR141716A
0.075-0.75 mg/kg
0.25-1 mg/kg
CPP
(reversing effect on 9-THC)
Wistar rats [99]
9-THC 1 mg/kg
5 mg/kg
CPP
CPA
A2AKO and wild-type mice [116]
URB-597 0.03-0.3 mg/kg - Wistar, Sprague-Dawley rats [121]
AM-404
1.25-10 mg/kg CPP
-
Rats (anxiety models) [122]
9-THC 0.1 mg/kg CPP Sprague-Dawley rats [114]
9-THC 1 mg/kg - Sprague-Dawley rats [47]
CPA Conditioned Place Aversion (Avoidance), - No Effect, Increase, Decrease.

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 357
μg/kg. Similarly, Chaperon et al. [118] using the synthetic CB1 receptor agonist WIN55,212-2 in a wide dose range between 0.003-1 mg/kg did not observe any CPP. To the contrary, in the doses of 0.3-1 mg/kg the animals exhibited a clear conditioned place aversion, which was reversed by pretreatment with the CB1 receptor antagonist SR141716A. A later study by Robinson et al. [111] using a conditioned place preference protocol in the open-field water maze did not report any preference or aversion for WIN55,212-2 in the dose range of 1 and 3 mg/kg, extending and confirming the results of the study by Chaperon et al. It is worth noting that in the same study, using the same experimental design,
9-
THC produced conditioned place aversion, but diazepam produced a clear and robust conditioned place preference. Interestingly in another study by Cheer et al. [107] both
9-
THC and its analog HU-210 produced conditioned place aversion in the rat, although cocaine and the CB1 receptor antagonist SR141716A produced a clear preference. The fact that SR141716A produced place preference opens the possi-bility that an endogenous cannabinoid tone might be present in the brain, as a physiological system to suppress reward or induce aversion [107]. However, in other studies SR141716A failed to produce either place conditioning or place aversion [118, 119]. To the contrary with the studies cited above, Braida et al. [119] reported that CP55,940 elic-its a conditioned place preference only in the dose of 20 μg/kg. The authors used a broader time interval between the cannabinoid injections, compared with the study by McGre-gor et al. [117], in which a place aversion was shown using a narrow time interval. These findings corroborate those ob-served in the Lepore et al. study [106] with
9-THC. Thus,
these authors suggested that in the study by McGregor et al. [117] the narrow interval between the cannabinoid injections might lead to a rebound anhedonic or dysphoric state mask-ing the appetitive effects of the drug. The conditioned place preference induced by CP55,940 was fully antagonized by pretreatment with the CB1 receptor antagonist SR141716A. Recently, Castané et al. [120] also reported that WIN55,212-2 produced conditioned place preference in mice pre-exposed to a priming injection of the drug.
A limited number of studies have examined the effects of endogenous cannabinoids or compounds increasing their levels in the brain. Mallet and Beninger [110] were the first to report that the administration of the endogenous cannabi-noid anandamide did not produce any significant effects in place conditioning, although using the same experimental protocol
9-THC produced conditioned place aversion.
Gobbi et al. [121] in a study investigating the antidepressant properties of URB-597, a compound that blocks anandamide hydrolysis, did not find any rewarding effects in the CPP paradigm. Interestingly, in a later study examining various properties of the anandamide transport inhibitor AM-404, Bortolato et al. [122] showed that AM-404 produced condi-tioned place preference only in rats housed under enriched conditions, but not in rats kept in standard cages.
In conclusion, while almost all drugs abused by humans are able to produce a clear and reliable place preference (i.e., increase the time spent in the drug paired compartment) over a limited range of doses, results with
9-THC and other can-
nabinoids have not always been consistent. The studies re-porting a conditioned place preference associated with
administration of a cannabinoid have used particular experi-mental methodology, and the preference often occurs at only a single dose.
CANNABINOID DISCRIMINATION
The pharmacological effects of a drug can include dis-tinct interoceptive cueing effects. These cueing effects can play an important role in the addiction process, such as prompting additional drug-seeking behavior. The standard preclinical rodent model for studying the cueing (discrimina-tive stimulus) effects of a drug is referred to as drug dis-crimination [123, 124]. In this procedure, animals faced with two possible responses one of which is reinforced (usually with food, under some schedule) while the other is not rein-forced. The animals are trained to detect whether they re-ceived an active drug or vehicle (no drug) injection, in order to determine through the specific drug effects which re-sponse is the correct. Rats (like humans) can readily learn to discriminate between a drug versus a non-drug state pressing the reinforcement-associated lever. The discrimination be-tween drug and non-drug conditions is based upon the pres-ence or absence of subjective and central effects. Although the drug discrimination procedure does not measure directly the reinforcing/rewarding effects of drugs, it can be used to infer if an experimental drug may have abuse potential in humans [125, 126].
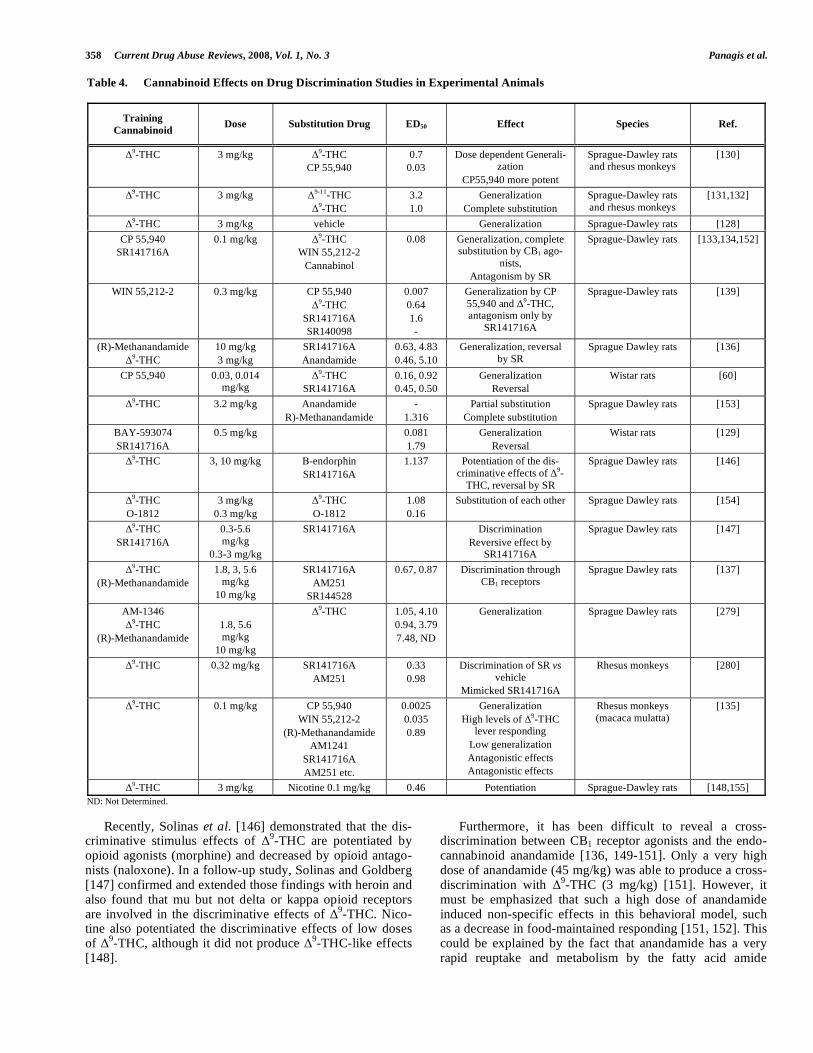
A significant number of drug discrimination studies have shown that cannabinoids produce subjective effects in ex-perimental animals [60,127-134] (Table 4). In particular, cannabinoids show a pharmacological specificity in this be-havioral paradigm. In animals trained to discriminate injec-tions of
9-THC from injections of saline, only drugs that
activate CB1 receptors generalize to the 9-THC training
stimulus [60, 128-133, 135]. Furthermore, synthetic CB1 receptor agonists generalize to the
9-THC training stimulus
with a potency that corresponds with their in vitro affinity for the CB1 receptors [127, 130, 134]. Likewise, CB1 recep-tors seem to be selectively involved in these actions, since they can be blocked by SR141716A administration [60, 129, 133-139]. Moreover, it seems very likely that the cannabi-noid cue effect is centrally mediated through the CB1 recep-tor subtype, since Pério et al. [139] have shown that the CB1 receptor antagonist SR140098, which does not cross the blood brain barrier, could not antagonize the discriminative stimulus properties of cannabinoids, and Kallman et al. [140] have shown that intracerebral administration of
9-THC in-
duced drug appropriate responding in rats trained to dis-criminate it from vehicle after systemic injections.
In general, experimental animals trained to discriminate a CB1 receptor agonist from saline showed complete generali-zation only if they were tested with other CB1 receptor ago-nists and not compounds from other pharmacological classes, such as opioids, barbiturates, psychostimulants, hal-lucinogens, anticonvulsants, neuroleptics and antidepressants [127-129, 141-143]. Only a partial overlap in the discrimina-tive stimulus properties between
9-THC and diazepam,
phencyclidine or 3,4-methylenedioxymethamphetamine has been reported [128, 141, 144, 145]. However, in a more re-cent study the substitutions of both pentobarbital and diaze-pam for
9-THC were found to be considerably less pro-
nounced than those reported previously [128, 144].
358 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
Recently, Solinas et al. [146] demonstrated that the dis-criminative stimulus effects of
9-THC are potentiated by
opioid agonists (morphine) and decreased by opioid antago-nists (naloxone). In a follow-up study, Solinas and Goldberg [147] confirmed and extended those findings with heroin and also found that mu but not delta or kappa opioid receptors are involved in the discriminative effects of
9-THC. Nico-
tine also potentiated the discriminative effects of low doses of
9-THC, although it did not produce
9-THC-like effects
[148].
Furthermore, it has been difficult to reveal a cross-discrimination between CB1 receptor agonists and the endo-cannabinoid anandamide [136, 149-151]. Only a very high dose of anandamide (45 mg/kg) was able to produce a cross-discrimination with
9-THC (3 mg/kg) [151]. However, it
must be emphasized that such a high dose of anandamide induced non-specific effects in this behavioral model, such as a decrease in food-maintained responding [151, 152]. This could be explained by the fact that anandamide has a very rapid reuptake and metabolism by the fatty acid amide
Table 4. Cannabinoid Effects on Drug Discrimination Studies in Experimental Animals
Training
Cannabinoid Dose Substitution Drug ED50 Effect Species Ref.
9-THC 3 mg/kg 9-THC
CP 55,940
0.7
0.03
Dose dependent Generali-zation
CP55,940 more potent
Sprague-Dawley rats and rhesus monkeys
[130]
9-THC 3 mg/kg 9-11-THC 9-THC
3.2
1.0
Generalization
Complete substitution
Sprague-Dawley rats
and rhesus monkeys
[131,132]
9-THC 3 mg/kg vehicle Generalization Sprague-Dawley rats [128]
CP 55,940
SR141716A
0.1 mg/kg 9-THC
WIN 55,212-2
Cannabinol
0.08
Generalization, complete substitution by CB1 ago-
nists,
Antagonism by SR
Sprague-Dawley rats [133,134,152]
WIN 55,212-2 0.3 mg/kg CP 55,940 9-THC
SR141716A
SR140098
0.007
0.64
1.6
-
Generalization by CP
55,940 and 9-THC, antagonism only by
SR141716A
Sprague-Dawley rats [139]
(R)-Methanandamide 9-THC
10 mg/kg
3 mg/kg
SR141716A
Anandamide
0.63, 4.83
0.46, 5.10
Generalization, reversal
by SR
Sprague Dawley rats [136]
CP 55,940
0.03, 0.014 mg/kg
9-THC
SR141716A
0.16, 0.92
0.45, 0.50
Generalization
Reversal
Wistar rats [60]
9-THC 3.2 mg/kg Anandamide
R)-Methanandamide
-
1.316
Partial substitution
Complete substitution
Sprague Dawley rats [153]
BAY-593074
SR141716A
0.5 mg/kg 0.081
1.79
Generalization
Reversal
Wistar rats [129]
9-THC
3, 10 mg/kg -endorphin
SR141716A
1.137 Potentiation of the dis-
criminative effects of 9-
THC, reversal by SR
Sprague Dawley rats [146]
9-THC
O-1812
3 mg/kg
0.3 mg/kg
9-THC
O-1812
1.08
0.16
Substitution of each other Sprague Dawley rats [154]
9-THC
SR141716A
0.3-5.6
mg/kg
0.3-3 mg/kg
SR141716A Discrimination
Reversive effect by SR141716A
Sprague Dawley rats [147]
9-THC
(R)-Methanandamide
1.8, 3, 5.6
mg/kg
10 mg/kg
SR141716A
AM251
SR144528
0.67, 0.87 Discrimination through
CB1 receptors
Sprague Dawley rats [137]
AM-1346 9-THC
(R)-Methanandamide
1.8, 5.6
mg/kg
10 mg/kg
9-THC
1.05, 4.10
0.94, 3.79
7.48, ND
Generalization Sprague Dawley rats [279]
9-THC 0.32 mg/kg SR141716A
AM251
0.33
0.98
Discrimination of SR vs
vehicle
Mimicked SR141716A
Rhesus monkeys [280]
9-THC 0.1 mg/kg CP 55,940
WIN 55,212-2
(R)-Methanandamide
AM1241
SR141716A
AM251 etc.
0.0025
0.035
0.89
Generalization
High levels of 9-THC lever responding
Low generalization
Antagonistic effects
Antagonistic effects
Rhesus monkeys
(macaca mulatta)
[135]
9-THC 3 mg/kg Nicotine 0.1 mg/kg 0.46 Potentiation Sprague-Dawley rats [148,155]
ND: Not Determined.
Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 359
hydrolase (FAAH). Indeed, the metabolically more stable analogs of anandamide, 2-methyl-arachidonyl-2-fluoroethyl-amide, R-methanandamide and O-1812, can produce a com-plete generalization to
9-THC [136, 137, 142, 149, 150,
153, 154], although the dose of 9-THC seems to be an im-
portant determinant for the degree of this generalization. However, anandamide has been shown to produce a cross-discrimination with methanandamide [136]. The discrepancy between the ability of anandamide to produce a cross-discrimination between methanandamide and not
9-THC
might be related to the different affinity of 9-THC and
methanandamide for cannabinoid receptors, or the possibility that anandamide and methanandamide, but not
9-THC, in-
teract with a different cannabinoid receptor than the CB1. Recently it has been shown, however, that the cross-discrimination between methanandamide and
9-THC is
mediated through the CB1 receptor [137]. Very recently, Solinas et al. [155] showed that methanandamide when in-jected i.v., produced clear dose-related
9-THC-like dis-
criminative effects, but anandamide produced 9-THC-like
discriminative effects only at a high dose that almost elimi-nated lever-press responding. The FAAH inhibitor URB-597 produced no
9-THC-like discriminative effects alone but
dramatically potentiated discriminative effects of anan-damide. The
9-THC-like discriminative stimulus effects of
anandamide after URB-597 and methanandamide were blocked by the CB1 receptor antagonist SR141716A. How-ever, surprisingly in the same study, the anandamide trans-port inhibitors AM-404 and UCM-707 did not potentiate
9-
THC-like discriminative effects of anandamide. Hence, anandamide substitution for other cannabinoids is, at best, unreliable. This may be due to the fact that anandamide, un-like direct cannabinoid receptor agonists, acts as an endova-nilloid and affects also vanilloid receptors.
In summary, drug discrimination with cannabinoids is pharmacologically selective, since
9-THC fully substituted
for other synthetic cannabinoids with affinity for the CB1 receptor. In contrast, compounds with anandamide-like
structure exhibit differences from classical cannabinoids, since in a number of studies they failed to substitute for
9-
THC or other CB1 receptor agonists. Since 9-THC-like dis-
criminative stimulus effects can predict cannabis-like intoxi-cation in humans, anandamide-like compounds and drugs that directly affect the endocannabinoid levels may prove promising therapeutics in the future, with less unwanted side effects and minimal abuse potential.
BEHAVIORAL SENSITIZATION TO CANNABI-NOIDS
Behavioral sensitization is a progressive and enduring enhancement in a behavioral response (usually motor behav-ior) to the administration of constant or incrementally en-hanced doses of a drug. Sensitization processes are subject to complex neurobiological alterations that persist for a long time and are thought to play an important role in the devel-opment and maintenance of drug addiction by a process in which a simple liking of the drug becomes sensitized up to the point where loss of control and drug craving is mani-fested [156]. Basically, drugs that humans abuse also pro-duce behavioral sensitization [157]. Accordingly, if a drug produces behavioral sensitization upon repeated administra-tion it might have abuse potential in humans. Whether can-nabinoids, and in particular
9-THC, produce similar effects
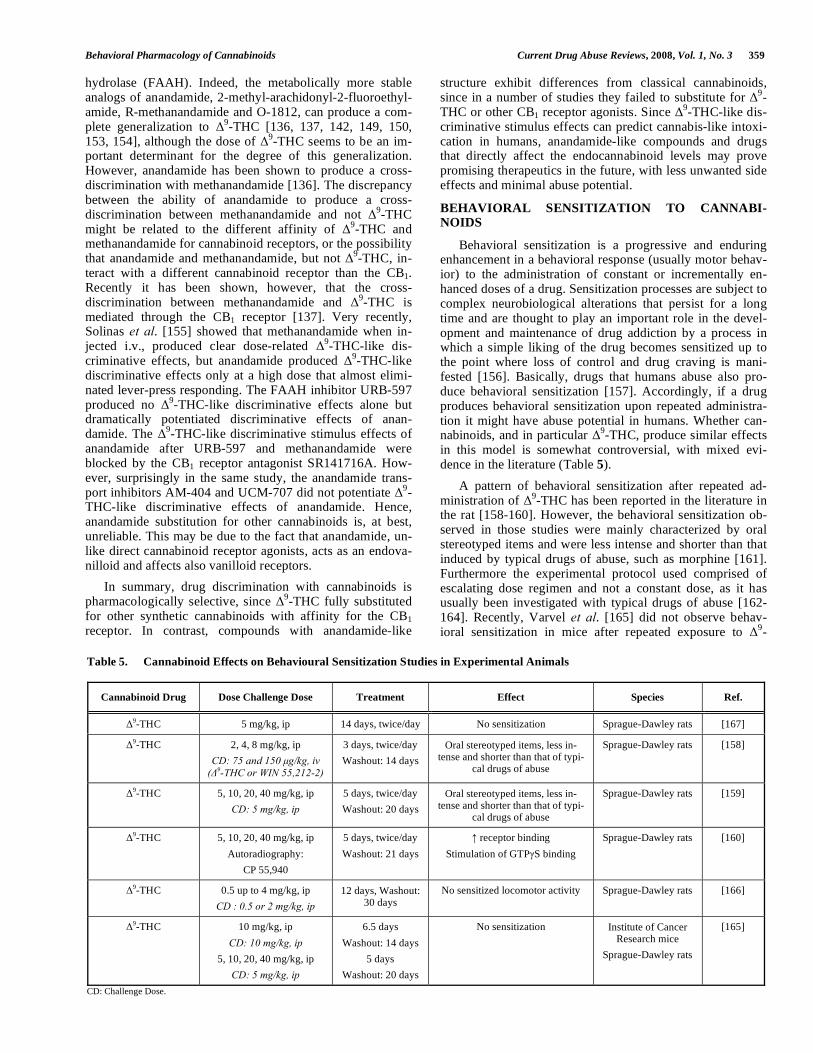
in this model is somewhat controversial, with mixed evi-dence in the literature (Table 5).
A pattern of behavioral sensitization after repeated ad-ministration of
9-THC has been reported in the literature in
the rat [158-160]. However, the behavioral sensitization ob-served in those studies were mainly characterized by oral stereotyped items and were less intense and shorter than that induced by typical drugs of abuse, such as morphine [161]. Furthermore the experimental protocol used comprised of escalating dose regimen and not a constant dose, as it has usually been investigated with typical drugs of abuse [162-164]. Recently, Varvel et al. [165] did not observe behav-ioral sensitization in mice after repeated exposure to
9-
Table 5. Cannabinoid Effects on Behavioural Sensitization Studies in Experimental Animals
Cannabinoid Drug Dose Challenge Dose Treatment Effect Species Ref.
9-THC 5 mg/kg, ip 14 days, twice/day No sensitization Sprague-Dawley rats [167]
9-THC 2, 4, 8 mg/kg, ip
CD: 75 and 150 μg/kg, iv
( 9-THC or WIN 55,212-2)
3 days, twice/day
Washout: 14 days
Oral stereotyped items, less in-tense and shorter than that of typi-
cal drugs of abuse
Sprague-Dawley rats [158]
9-THC 5, 10, 20, 40 mg/kg, ip
CD: 5 mg/kg, ip
5 days, twice/day
Washout: 20 days
Oral stereotyped items, less in-tense and shorter than that of typi-
cal drugs of abuse
Sprague-Dawley rats [159]
9-THC 5, 10, 20, 40 mg/kg, ip
Autoradiography:
CP 55,940
5 days, twice/day
Washout: 21 days
receptor binding
Stimulation of GTP S binding
Sprague-Dawley rats [160]
9-THC 0.5 up to 4 mg/kg, ip
CD : 0.5 or 2 mg/kg, ip
12 days, Washout: 30 days
No sensitized locomotor activity Sprague-Dawley rats [166]
9-THC 10 mg/kg, ip
CD: 10 mg/kg, ip
5, 10, 20, 40 mg/kg, ip
CD: 5 mg/kg, ip
6.5 days
Washout: 14 days
5 days
Washout: 20 days
No sensitization Institute of Cancer Research mice
Sprague-Dawley rats
[165]
CD: Challenge Dose.

360 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
THC, although with the same experimental procedure they observed a clear behavioral sensitization to methampheta-mine. Similarly, in a recent study by Kolb et al. [166], rats showed no evidence of sensitized locomotor activity after repeated treatment either with a constant dose or escalating doses of
9-THC. A failure for
9-THC to induce behavioral
sensitization has also been observed in a study by Wu and French [167]. Interestingly, in this study,
9-THC acutely
produced hypoactivity, which was diminished with repeated exposure to the drug, indicating the development of toler-ance to this effect. To our knowledge, in the relevant litera-ture behavioral sensitization following repeated exposure to synthetic cannabinoid agonists or endocannabinoid modula-tors has not so far been reported.
In summary, cannabinoids do not share with other drugs of abuse the potential to produce clear and robust behavioral sensitization. The fact that the relevant neuronal circuits which are responsible for the phenomenon of sensitization observed with typical drugs of abuse, failed to become sensi-tized after repeated exposure to cannabinoids, may explain why marijuana abuse in the human population usually is not accompanied by obsessive-compulsive drug-seeking behav-ior.
CANNABINOID TOLERANCE
Tolerance can be defined operationally as a right- or downward shift in a dose-effect function, so that higher doses of the drug are required to produce the same effect. Unequivocally this process represents an adaptation to a spe-cific drug effect. Many aspects of tolerance have been hy-pothesized to play important modulatory roles in the devel-opment of physical dependence. However, regular drug ex-posure might well lead to tolerance regardless of weather physical dependence occurs. Interestingly, under specific circumstances it has been shown that tolerance and physical dependence can be dissociated and that they can have dis-tinct molecular mechanisms (e.g., [168]).
Acute or subchronic administration of 9-THC or other
cannabinoid agonists in laboratory animals produces multi-ple neurobiological effects including sedation, catalepsy, motor inhibition, suppression of operant behavior, ataxia, incoordination, analgesia, anticonvulsing activity, memory impairment, antiemetic effect, hypothermia and increased appetite [169]. Cannabinoids also produce a variety of pe-ripheral effects, such as hypotension, reduction of intraocular pressure, immunosuppression, intestinal hypomotility, and changes in adrenal function [170]. Although there is little evidence for the development of sensitization in the cannabi-noid system (see previous section), there is ample evidence indicating that in most of the central and peripheral effects of cannabinoids tolerance develops when the administration prolongs for several days in laboratory animals [171, 172] (Table 6a).
Tolerance has been reported in various species [173], but the degree and the time-course are dependent on the species used, the agonist’s efficacy, the dose, the duration of the treatment, and the experimental procedures used to deter-mine tolerance, as well as the system in which it is assessed [171]. The development of cannabinoid tolerance is re-markably rapid and culminates soon after the start of the
chronic treatment [174-177]. However, although tolerance for certain pharmacological effects of cannabinoids, such as analgesia, motor inhibition, hypothermia, discriminative stimulus effects develops typically within the range of 3–7 days [131, 175, 178-181], other effects, such as the appetite stimulant, the antiemetic, the antihyperalgesic, memory im-pairment and neuroendocrine effects either do not develop tolerance or they require weeks for such a development or larger doses of the compounds [182-188].
Similarly, tolerance to anandamide-induced suppression of spontaneous activity, antinociception, catalepsy, hypo-thermia, and the twitch response in mouse vas deferens has also been reported [180, 189-194], although the magnitude of tolerance tended to be lower than that for
9-tetrahydro-
cannabinol [193, 194].
The development of cannabinoid tolerance is mostly pharmacodynamic in nature (down-regulation/desensiti-zation of cannabinoid receptors), and the contribution of pharmacokinetic properties of cannabinoids (changes in drug absorption, distribution, biotransformation, and excretion), if any, seems minor [195-198]. In fact, a significant reduction in the total number of CB1 receptors, which are predominant in the central nervous system, along with an attenuation of G-protein activation has been repeatedly shown after chronic administration for most cannabinoids [199]. These adaptive responses occur in all brain regions that express cannabinoid receptors, but there is a regional variability both in the rate and the magnitude of this response [171, 199]. Nevertheless, there have also been reports of increased receptor binding [200] or no change in receptor binding in whole brain [201] during the development of tolerance. Furthermore, chronic exposure to
9-THC has also been shown to produce region-
dependent alterations in brain endocannabinoid levels in the rat. Specifically, anandamide concentrations were increased in the limbic forebrain and decreased in striatum, midbrain and diencephalon in
9-THC tolerant rats, whereas 2-
arachidonoylglycerol concentrations increased in the cerebel-lum, brainstem and hippocampus, and decreased in the stria-tum [202, 203]. Tolerance at least to some of the pharmacol-ogical effects of cannabinoids dissipated from 7 to 11 days after the end of the chronic cannabinoid exposure [175]. This effect fits very well with the reported reversal of the adaptive changes in the CB1 receptors even after long-term treatment with
9-THC [204]. Interestingly, the CB1 receptor antago-
nist SR141716A did not always block the development of tolerance to central behavioral actions of cannabinoids, such as motor incoordination [176]. This finding is very unusual, and may be related to the long duration of
9-THC action.
As might be expected, cross-tolerance has been reported between different exogenous CB1 receptor agonists [178, 180] (Table 6b). However, controversial results have been reported on the possible cross-tolerance between anandamide or its metabolically stable analogs and
9-THC. Fride [191]
demonstrated cross-tolerance to the antinociceptive, hypolo-comotor, cataleptic and hypothermic effect between
9-THC
and anandamide in female mice, but not to the decreased defecation. On the contrary, Pertwee et al. [180] did not re-port cross-tolerance between
9-THC and anandamide to
their hypothermic effect. Similarly, Welch [193] did not show cross-tolerance between
9-THC and anandamide to
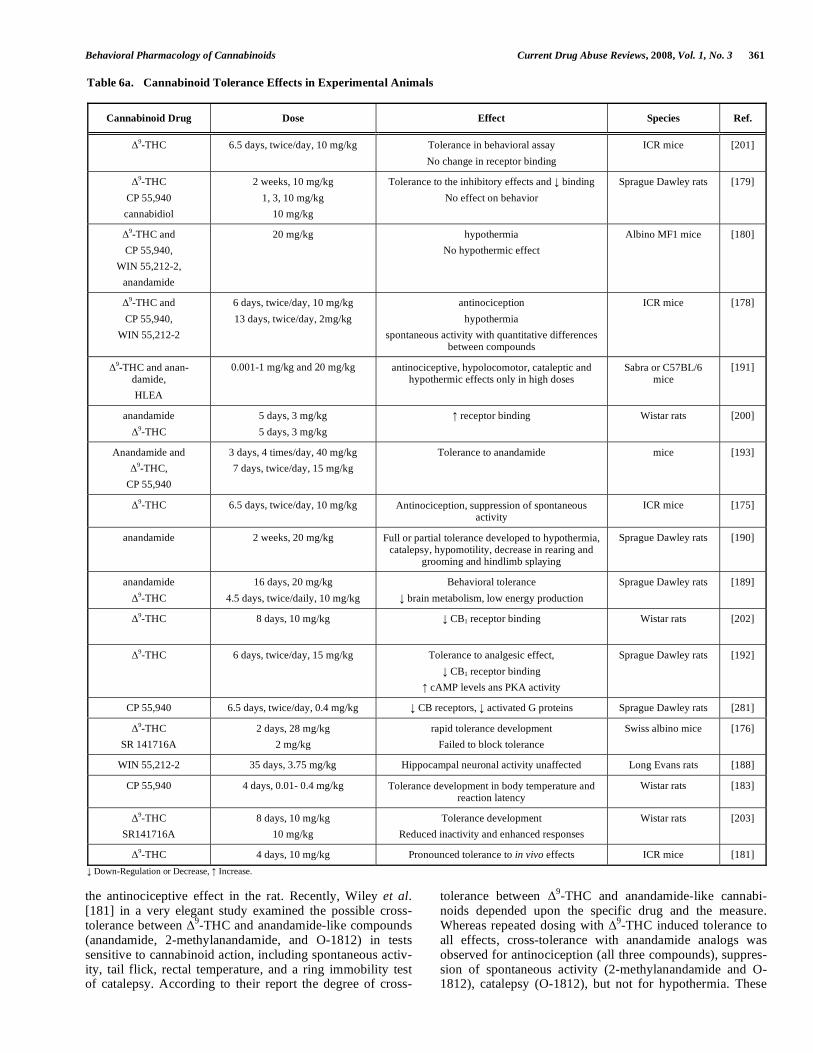
Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 361
the antinociceptive effect in the rat. Recently, Wiley et al. [181] in a very elegant study examined the possible cross-tolerance between
9-THC and anandamide-like compounds
(anandamide, 2-methylanandamide, and O-1812) in tests sensitive to cannabinoid action, including spontaneous activ-ity, tail flick, rectal temperature, and a ring immobility test of catalepsy. According to their report the degree of cross-
tolerance between 9-THC and anandamide-like cannabi-
noids depended upon the specific drug and the measure. Whereas repeated dosing with
9-THC induced tolerance to
all effects, cross-tolerance with anandamide analogs was observed for antinociception (all three compounds), suppres-sion of spontaneous activity (2-methylanandamide and O-1812), catalepsy (O-1812), but not for hypothermia. These
Table 6a. Cannabinoid Tolerance Effects in Experimental Animals
Cannabinoid Drug Dose Effect Species Ref.
9-THC 6.5 days, twice/day, 10 mg/kg Tolerance in behavioral assay
No change in receptor binding
ICR mice [201]
9-THC
CP 55,940
cannabidiol
2 weeks, 10 mg/kg
1, 3, 10 mg/kg
10 mg/kg
Tolerance to the inhibitory effects and binding
No effect on behavior
Sprague Dawley rats [179]
9-THC and
CP 55,940,
WIN 55,212-2,
anandamide
20 mg/kg hypothermia
No hypothermic effect
Albino MF1 mice [180]
9-THC and
CP 55,940,
WIN 55,212-2
6 days, twice/day, 10 mg/kg
13 days, twice/day, 2mg/kg
antinociception
hypothermia
spontaneous activity with quantitative differences between compounds
ICR mice [178]
9-THC and anan-damide,
HLEA
0.001-1 mg/kg and 20 mg/kg antinociceptive, hypolocomotor, cataleptic and hypothermic effects only in high doses
Sabra or C57BL/6 mice
[191]
anandamide 9-THC
5 days, 3 mg/kg
5 days, 3 mg/kg
receptor binding Wistar rats [200]
Anandamide and 9-THC,
CP 55,940
3 days, 4 times/day, 40 mg/kg
7 days, twice/day, 15 mg/kg
Tolerance to anandamide mice [193]
9-THC 6.5 days, twice/day, 10 mg/kg Antinociception, suppression of spontaneous activity
ICR mice [175]
anandamide 2 weeks, 20 mg/kg Full or partial tolerance developed to hypothermia, catalepsy, hypomotility, decrease in rearing and
grooming and hindlimb splaying
Sprague Dawley rats [190]
anandamide 9-THC
16 days, 20 mg/kg
4.5 days, twice/daily, 10 mg/kg
Behavioral tolerance
brain metabolism, low energy production
Sprague Dawley rats [189]
9-THC 8 days, 10 mg/kg CB1 receptor binding
Wistar rats [202]
9-THC 6 days, twice/day, 15 mg/kg Tolerance to analgesic effect,
CB1 receptor binding
cAMP levels ans PKA activity
Sprague Dawley rats [192]
CP 55,940 6.5 days, twice/day, 0.4 mg/kg CB receptors, activated G proteins Sprague Dawley rats [281]
9-THC
SR 141716A
2 days, 28 mg/kg
2 mg/kg
rapid tolerance development
Failed to block tolerance
Swiss albino mice [176]
WIN 55,212-2 35 days, 3.75 mg/kg Hippocampal neuronal activity unaffected Long Evans rats [188]
CP 55,940 4 days, 0.01- 0.4 mg/kg Tolerance development in body temperature and reaction latency
Wistar rats [183]
9-THC
SR141716A
8 days, 10 mg/kg
10 mg/kg
Tolerance development
Reduced inactivity and enhanced responses
Wistar rats [203]
9-THC 4 days, 10 mg/kg Pronounced tolerance to in vivo effects ICR mice [181]
Down-Regulation or Decrease, Increase.
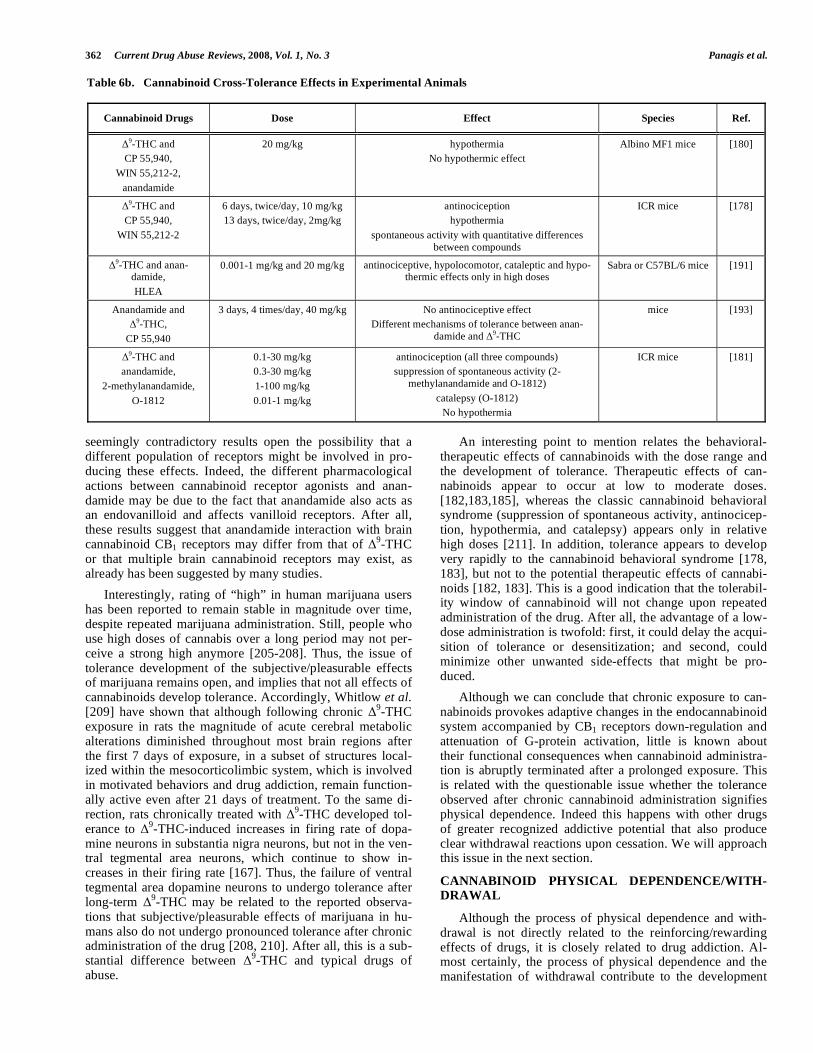
362 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
seemingly contradictory results open the possibility that a different population of receptors might be involved in pro-ducing these effects. Indeed, the different pharmacological actions between cannabinoid receptor agonists and anan-damide may be due to the fact that anandamide also acts as an endovanilloid and affects vanilloid receptors. After all, these results suggest that anandamide interaction with brain cannabinoid CB1 receptors may differ from that of
9-THC
or that multiple brain cannabinoid receptors may exist, as already has been suggested by many studies.
Interestingly, rating of “high” in human marijuana users has been reported to remain stable in magnitude over time, despite repeated marijuana administration. Still, people who use high doses of cannabis over a long period may not per-ceive a strong high anymore [205-208]. Thus, the issue of tolerance development of the subjective/pleasurable effects of marijuana remains open, and implies that not all effects of cannabinoids develop tolerance. Accordingly, Whitlow et al. [209] have shown that although following chronic
9-THC
exposure in rats the magnitude of acute cerebral metabolic alterations diminished throughout most brain regions after the first 7 days of exposure, in a subset of structures local-ized within the mesocorticolimbic system, which is involved in motivated behaviors and drug addiction, remain function-ally active even after 21 days of treatment. To the same di-rection, rats chronically treated with
9-THC developed tol-
erance to 9-THC-induced increases in firing rate of dopa-
mine neurons in substantia nigra neurons, but not in the ven-tral tegmental area neurons, which continue to show in-creases in their firing rate [167]. Thus, the failure of ventral tegmental area dopamine neurons to undergo tolerance after long-term
9-THC may be related to the reported observa-
tions that subjective/pleasurable effects of marijuana in hu-mans also do not undergo pronounced tolerance after chronic administration of the drug [208, 210]. After all, this is a sub-stantial difference between
9-THC and typical drugs of
abuse.
An interesting point to mention relates the behavioral-therapeutic effects of cannabinoids with the dose range and the development of tolerance. Therapeutic effects of can-nabinoids appear to occur at low to moderate doses. [182,183,185], whereas the classic cannabinoid behavioral syndrome (suppression of spontaneous activity, antinocicep-tion, hypothermia, and catalepsy) appears only in relative high doses [211]. In addition, tolerance appears to develop very rapidly to the cannabinoid behavioral syndrome [178, 183], but not to the potential therapeutic effects of cannabi-noids [182, 183]. This is a good indication that the tolerabil-ity window of cannabinoid will not change upon repeated administration of the drug. After all, the advantage of a low-dose administration is twofold: first, it could delay the acqui-sition of tolerance or desensitization; and second, could minimize other unwanted side-effects that might be pro-duced.
Although we can conclude that chronic exposure to can-nabinoids provokes adaptive changes in the endocannabinoid system accompanied by CB1 receptors down-regulation and attenuation of G-protein activation, little is known about their functional consequences when cannabinoid administra-tion is abruptly terminated after a prolonged exposure. This is related with the questionable issue whether the tolerance observed after chronic cannabinoid administration signifies physical dependence. Indeed this happens with other drugs of greater recognized addictive potential that also produce clear withdrawal reactions upon cessation. We will approach this issue in the next section.
CANNABINOID PHYSICAL DEPENDENCE/WITH-DRAWAL
Although the process of physical dependence and with-drawal is not directly related to the reinforcing/rewarding effects of drugs, it is closely related to drug addiction. Al-most certainly, the process of physical dependence and the manifestation of withdrawal contribute to the development
Table 6b. Cannabinoid Cross-Tolerance Effects in Experimental Animals
Cannabinoid Drugs Dose Effect Species Ref.
9-THC and
CP 55,940,
WIN 55,212-2,
anandamide
20 mg/kg hypothermia
No hypothermic effect
Albino MF1 mice [180]
9-THC and
CP 55,940,
WIN 55,212-2
6 days, twice/day, 10 mg/kg
13 days, twice/day, 2mg/kg
antinociception
hypothermia
spontaneous activity with quantitative differences
between compounds
ICR mice [178]
9-THC and anan-damide,
HLEA
0.001-1 mg/kg and 20 mg/kg antinociceptive, hypolocomotor, cataleptic and hypo-thermic effects only in high doses
Sabra or C57BL/6 mice [191]
Anandamide and 9-THC,
CP 55,940
3 days, 4 times/day, 40 mg/kg No antinociceptive effect
Different mechanisms of tolerance between anan-
damide and 9-THC
mice [193]
9-THC and
anandamide,
2-methylanandamide,
O-1812
0.1-30 mg/kg
0.3-30 mg/kg
1-100 mg/kg
0.01-1 mg/kg
antinociception (all three compounds)
suppression of spontaneous activity (2-
methylanandamide and O-1812)
catalepsy (O-1812)
No hypothermia
ICR mice [181]

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 363
and maintenance of drug addiction through negative rein-forcement processes [212-214]. On the other hand, the de-velopment of physical dependence and the manifestation of withdrawal symptoms limit the use of many drugs in the clinic.
Two procedures are commonly used to investigate the development of physical dependence and the occurrence of withdrawal syndrome in laboratory animals. The first is the abstinence withdrawal procedure, in which following re-peated administration of a drug, it is abruptly discontinued and the physical signs that emerge are evaluated (spontane-ous withdrawal). In the second procedure, the precipitated withdrawal, an appropriate receptor antagonist is adminis-tered to animals that have been made dependent upon a drug (the agonist). The rationale of this model is that the antago-nist displaces the agonist from the appropriate receptor, elic-iting molecular changes and withdrawal signs that can easily be evaluated.
In addition to the somatic aspects of drug withdrawal, another relevant symptom, which is closely related to the brain reward circuit, is anhedonia. Anhedonia is a core symptom in both depression and drug withdrawal and can be investigated using the ICSS paradigm, as elegantly has been shown by Markou and colleagues [215-217]. In this case, ICSS thresholds are used operationally as a measure of the animals' "hedonic-anhedonic" state. ICSS thresholds are found to be elevated under withdrawal conditions from many drugs of abuse, including psychostimulants, opiates, alcohol, nicotine, and phencyclidine. This reflects an "anhedonic" state for the animal exhibiting drug withdrawal.
Although experimental animals treated chronically with typical drugs of abuse exhibit withdrawal reactions either following drug cessation or upon the administration of the appropriate receptor antagonist, data on cannabinoids are largely contentious (Table 7). Thus, there is no general agreement whether cannabinoid tolerance can produce physical dependence, as has been demonstrated with typical drugs of abuse. First of all, several studies failed to report spontaneous withdrawal symptoms following chronic
9-
THC administration in experimental animals [72, 73, 177, 218-222]. It has been proposed that the main reason for this is the particular pharmacokinetic properties of
9-THC [171-
173, 223]. Indeed, its high lipophilicity, the long duration and the slow metabolic clearance may prevent the develop-ment of a pronounced withdrawal syndrome, when the drug is abruptly discontinued. This may also explain the general contention that cannabis withdrawal syndrome is seldom in recreational cannabis users, or when appeared it is mild and clinically insignificant (DSM-IV, [224]). Indeed, most scien-tists agree that a reliable cannabis withdrawal syndrome may emerge only following discontinuation of heavy use of can-nabis preparations [2, 225]. To our knowledge, the only study in rats reporting a spontaneous withdrawal reaction after chronic
9-THC treatment has been published by Kay-
makcalan et al. [226]. However, in this study, the authors treated the rats for 5 weeks with extremely high doses that the last two weeks reached 40mg/kg twice daily. In spite of the failure to observe clear spontaneous withdrawal reactions following
9-THC cessation, Beardsley et al. [227] reported
a suppression of operant behavior for food reinforcement in
monkeys. The authors interpreted this specific condition as a manifestation of spontaneous withdrawal, since it returned to normal after re-administration of the drug. In a latter study, they replicated the same behavioral pattern in rats [228]. Fredericks and Benowitz [229] also reported that following discontinuation of
9-THC, monkeys showed an increase in
gross movement, eye contact, and tooth bearing of greater frequency and/or duration than observed before
9-THC.
The authors suggest that this may represent a cannabis absti-nence syndrome. However, these manifestations deviate from the view of the classical withdrawal syndrome ob-served with typical drugs of abuse.
Only three studies have reported the occurrence of spon-taneous cannabinoid withdrawal following chronic treatment with synthetic cannabinoid agonists [230-232]. However, it should be emphasized that these studies used synthetic ago-nists of powerful pharmacological potency for the cannabi-noid receptors (WIN55,212-2 and CP55,940), compared to
9-THC, and with a substantial different pharmacokinetic
profile. Furthermore, the spontaneous withdrawal observed was not very pronounced, but rather mild and particularly modest compared with other studies in which precipitated withdrawal from cannabinoids was manifested. In a more recent study in rats, Moreno et al. [233] observed, following withdrawal of chronic treatment with the CB1 receptor ago-nist HU-210, a decrease in spontaneous motor activity, which was interpreted as an aspect of withdrawal reaction. To the contrary, Oliva et al. [231,232] reported during spon-taneous withdrawal from CP55,940 a marked increase in motor activity and exploratory behavior along with decreases in the number of groomings, rubbings and wet-dog shakes. Abrupt cessation of treatment with CP55,940 does not in-duce the classic somatic signs of withdrawal observed after cannabinoid antagonist-precipitated withdrawal. This may be related to the different experimental protocol (i.e. spontane-ous vs precipitated), the different agonists used or the differ-ent timing of the dosing regimen.
In the absence of a clear spontaneous withdrawal from 9-THC in experimental animals a number of investigators
have tried to demonstrate physical dependence and with-drawal signs by challenging chronically treated animals with a CB1 receptor antagonist, such as SR141716A. In fact, ad-ministration of the CB1 receptor antagonist SR141716A in experimental animals chronically treated with
9-THC pre-
cipitates specific withdrawal reactions. The signs observed are largely somatic (mainly of motor nature), with no rele-vant vegetative changes. The most characteristic withdrawal signs in rodents are wet-dog shakes, head shakes, facial rub-bing, front paw tremor, body tremor, ataxia, eyelid ptosis, piloerection, hypolocomotion, retropulsion, licking, scratch-ing and mastication [94, 109, 116, 203, 218, 234-244]. Simi-lar precipitated withdrawal reactions have also been ob-served following chronic treatments with synthetic cannabi-noid agonists [245-249]. These somatic signs, even if they appear very mild, are mediated through the CB1 receptor, since in CB1 knockout mice receiving chronic treatment with
9-THC administration, the CB1 receptor antagonist
SR141716A did not precipitate any signs of abstinence [94]. However, in most of the above mentioned studies the authors used extreme large doses of cannabinoid agonist and/or an-tagonist. This may have complicated the obtained results.

364 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
Table 7. Cannabinoid Physical Dependence/Withdrawal Effects in Experimental Animals
Cannabinoid Drug Dose Effect Species Ref.
9-THC No spontaneous withdrawal symptoms dogs [220]
9-THC 11, 13, 28 days
10, 20, 40, 80 mg/kg
No spontaneous withdrawal symptoms QS strain mice [219]
9-THC 20 days, 12h access, 0.025-0.3 mg/kg No spontaneous withdrawal symptoms rhesus monkeys [72]
Suspension of cannabis resin 126 days, 0.1 mg/ml of resin No spontaneous withdrawal symptoms Wistar rats [73]
9-THC 5 weeks, up to 40mg/kg Spontaneous withdrawal reaction rats [226]
3 weeks, every 6h, 0.5 mg/kg gross movement, eye contact, tooth baring rhesus monkeys [229]
9-THC 10 days, 0.05 mg/kg No spontaneous withdrawal reactions, but suppression of operant behavior for food
reinforcement
rhesus monkeys [227]
9-THC
Withdrawal by: SR141716A
6.5 days, twice/day
15 mg/kg
5 mg/kg
Disorganized pattern of constantly changing brief sequences of motor behaviour, no evi-
dent autonomic signs
rats [242]
9-THC
Withdrawal by: SR141716A
4 days, 0.5 up to 100 mg/kg No spontaneous withdrawal symptoms Sprague Dawley rats
[218]
HU-210
Withdrawal by: SR141716A
2 weeks, 100 μg/kg
3 mg/kg
Precipitated withdrawal reactions,
elevation in extracellular CRF concentration,
Fos activation in the central nucleus of the
amygdale
rats [248]
Anandamide
SR141716A
Arachidonic acid
Me-F-AN
10-100 mg/kg/day, ip, 4 days
50-200mg/kg/infusion
5-20 mg/kg
No spontaneous withdrawal symptoms Sprague Dawley rats
[251]
9-THC
Withdrawal by: SR141716A
1 week, twice/day,
10 mg/kg
10, 30 mg/kg
profound withdrawal syndrome
paw tremors, head shakes,
normal behavior such as grooming and scratching
ICR mice [238]
9-THC,
Withdrawal by: SR141716A
6.5 days, twice/day, 15 mg/kg
10 mg/kg
No spontaneous withdrawal symptoms,
intense behavioral withdrawal syndrome
Sprague Dawley albino rats
[221]
9-THC
Withdrawal by: SR141716A
6 days, 10-20 mg/kg
10 mg/kg
wet dog shakes, frontpaw tremor, ataxia, hunched posture, tremor, ptosis, piloerection,
decreased locomotor activity and mastication
CD-1 mice [109]
9-THC
anandamide
Withdrawal by: SR141716A
9 days, twice/day, 0.1-0.6 mg/kg-1
0.3, 1, 3 mg/kg-1
1 mg/kg-1
Unique pattern of withdrawal signs: diarrhea, vomiting, excessive salivation, social behav-
ior, restless behavior and trembling
Beagle dogs [250]
CP 55,940
Withdrawal by: SR141716A
6.5 days, twice/day, 0.4 mg/kg
5 mg/kg
Precipitated withdrawal reactions Sprague Dawley rats
[249]
9-THC
Withdrawal by: SR141716A
10, 20 mg/kg No spontaneous withdrawal symptoms CB1 knock-out mice
[94]
9-THC
Withdrawal by: SR141716A
Profound withdrawal syndrome Rats [240]
9-THC
Withdrawal by: SR141716A
6 days, twice/day,
up to 9 mg/kg
No spontaneous withdrawal reactions, but suppression of operant behavior for food
reinforcement
Long Evans Hooded rats
[228]
Anandamide
Withdrawal by: SR 141716A
2 weeks, 20 mg/kg
3 mg/kg
Withdrawal reaction: scratching, licking, biting, eating of feces, ptosis, arched back,
wet-dog and head shakes, myoclonic spasms,
writhing, forepaw fluttering, teeth chattering and piloerection
Sprague Dawley rats
[190]

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 365
For example, Rodriguez de Fonseca et al. [248] observed even in rats chronically treated with vehicle that SR141716A elicits some withdrawal signs (spontaneous withdrawal). Interestingly, in
9-THC-dependent dogs, SR141716A pre-
cipitated a unique pattern of withdrawal signs that included diarrhea, vomiting, excessive salivation, decreases in social behavior, and increases in restless behavior and trembling [250].
Only two studies have examined the possibility that en-docannabinoids, such as anandamide, might produce physi-cal dependence when administered exogenously; their results move in opposite directions. Aceto et al. [251] in a very ele-gant study showed that abrupt discontinuation of anan-damide infusion in rats did not elicit any withdrawal re-sponses; furthermore, neither SR141716A was able to pre-
cipitate withdrawal reactions in those rats. The authors de-cided to administer the drug through continuous infusion (rats were infused i.p. on a continuous basis with anan-damide [25-100 mg/kg/day] for 4 days), in order to address its rapid metabolism. Furthermore, they examined if the pu-tative, metabolically stable anandamide analog 2-Me-F-AN could produce physical dependence (5-20mg infused for 4 days). Even with the use of 2-Me-F-AN they were unable to demonstrate signs of physical dependence. Accordingly, it seems unlikely that the failure of chronic anandamide to produce physical dependence signs is related to its short half-life or metabolism.
On the other hand, rats injected i.p. for two weeks with anadamide (20mg/kg) displayed a withdrawal reaction both upon abrupt cessation and under a challenge with
(Table 7) contd…..
Cannabinoid Drug Dose Effect Species Ref.
9-THC
Withdrawal by: SR141716A
5 days, twice/day, 20 mg/kg
10 mg/kg
basal and calcium-calmodulin-stimulated adenylyl cyclase activities,
active PKA in the cerebellum
CD-1 mice [243]
WIN 55,212-2
Withdrawal by: SR141716A
4 days, once/day, 1-16 mg/kg Mild spontaneous withdrawal Sprague Dawley rats
[230]
HU-210
Withdrawal by: AM281
5 days, twice/day, 100 μg/kg
3 mg/kg
Precipitated withdrawal reactions Long Evans rats [246]
9-THC
Withdrawal by: SR141716A
5 days, 10 mg/kg
10 mg/kg
paw tremors and head shakes Four different mouse strains
[239]
9-THC
Withdrawal by: SR141716A
11 times, every 12h, 20 mg/kg
10 mg/kg
forepaw licking, facial preening, grooming, forepaw tremor, head shakes and weight loss
ddY mice [235]
9-THC
Withdrawal by: SR141716A
4 days, 12.5-100 mg/kg Antagonist precipitated withdrawal signs Sprague-Dawley rats
[236]
9-THC
Withdrawal by: SR141716A
5 days, twice/day, 20 mg/kg
10 mg/kg
No spontaneous withdrawal symptoms mu-, delta-, and kappa- knock-
out mice
[237]
CP 55,940 14 days, 0.5 mg/kg/12h motor activity, exploratory behavior
grooming, rubbing, wet-dog shakes
Swiss albino mice
[231]
HU-210
Withdrawal by: SR141716A
14 days, 40-100 μg/kg
3 mg/kg
Precipitated withdrawal reactions Wistar rats [245]
9-THC
Withdrawal by: SR141716A
8 days, 10 mg/kg
10 mg/kg
inactivity in the open-field test,
responses as tremor, turning and retropul-sion
Wistar rats [203]
CP 55,940 6 days, twice/day, 0.5 mg/kg motor activity, exploratory behavior
grooming, rubbing, wet-dog shakes
Swiss albino mice
[232]
9-THC
Withdrawal by: SR141716A
5 days, 20 mg/kg
10 mg/kg
Tolerance, but no spontaneous withdrawal symptoms
A2A knock-out mice
[116]
HU-210 14 days, 20 μg/kg daily spontaneous motor activity Wistar rats [233]
WIN 55,212-2
Withdrawal by: SR141716A
Precipitated withdrawal reactions rats [247]
Marijuana 9-THC
Withdrawal by: SR141716A
5 days, 50, 100, 200 mg
1, 3, 10 mg/kg
10 mg/kg
paw tremors by SR141716A
blood, brain concentrations of 9-THC
Precipitated withdrawal effect
ICR mice [244]
9-THC
Withdrawal by: SR141716A
5 days, twice/day, 20 mg/kg
10 mg/kg
Precipitated withdrawal reactions CD-1 mice [241]
Increase, Decrease, 2-Me-F-AN [2-methylarachidonyl-(2'-fluoroethyl)-amide].

366 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
SR141716A [190]. The withdrawal signs observed in that study resemble those observed with other cannabinoid ago-nists and include scratching, licking and biting, eating of feces, ptosis, arched back, wet-dog shakes, head shakes, myoclonic spasms, writhing, forepaw fluttering, teeth chat-tering and piloerection. The different findings in these two studies are likely due to the different experimental protocol employed (length of drug exposure, timing of injections, dose range of anandamide and SR141716A used, behavioral method to measure withdrawal). To our knowledge, there is no study in the literature examining if drugs that protect en-docannabinoids from endogenous inactivation may produce physical dependence upon repeated administration.
Although most drugs of abuse upon withdrawal after chronic treatment exhibit a clear "anhedonic" state as re-flected in elevated ICSS thresholds [215-217], this has not been investigated properly with cannabinoids. Gardner and Vorel [43] report that “withdrawal from as little as a single 1.0 mg/kg dose of
9-THC produces significant elevations in
electrical brain stimulation thresholds”. However, in our opinion it seems rather unusual that a single and relatively low dose of
9-THC could produce dependence, in order to
observe withdrawal anhedonia, as typically more robust regimens of administration are employed for other drugs of abuse to study these neurobiological phenomena. Since Gardner and colleagues in their studies adopt a very strict criterion of stable responding (i.e., 0.01 log units for three consecutive days), it is possible that the observed effect of
9-THC in their study could reflect normal baseline variation
over days.
In summary, there is no conclusive evidence for a robust spontaneous withdrawal in experimental animals treated chronically with
9-THC. The analogous precipitated with-
drawal, or the withdrawal observed after chronic treatment with potent synthetic CB1 receptor agonists has been shown using very high doses of
9-THC or the equivalent CB1 re-
ceptor agonists, which exceed the relevant dose range in hu-man cannabis users [252]. Regarding endocannabinoids, the available data are limited and contradictory; this make it difficult to conclude if the endogenous substances have ad-dictive potential, much more since we do not, as yet, know if agents capable to protect endocannabinoids from endoge-nous inactivation produce any physical dependence and/or withdrawal.
NEUROCHEMICAL EFFECTS OF CANNABINOIDS ON DOPAMINE RELEASE IN THE NUCLEUS AC-
CUMBENS IN VIVO
It has been well established that certain typical drugs of abuse, e.g. psychostimulants, opiates, ethanol, and nicotine, increase dopamine release in the nucleus accumbens, a main projection area of the ascending mesotelencephalic dopa-minergic system (also referred to as mesolimbocortical do-paminergic projection), in experimental animals in vivo [253]. This effect has been considered as a neurochemical hallmark of the dependence-producing properties and abuse potential of these classes of drugs, given the intricate in-volvement of the mesolimbocortical dopaminergic projection in processes related to reinforcement of behavior, hedonic homeostasis and incentive salience attribution to intero- and exteroceptive physiological and pharmacological stimuli
[254, 255]. Acute administration of addictive drugs typically increases dopamine release in the nucleus accumbens, espe-cially in the shell subdivision, to a greater extent than in other projection areas, e.g., the striatum and the core of the nucleus accumbens [256, 257]. This action is either main-tained or even enhanced after repeated administration de-pending on the conditions and the context of administration [258-260]. Additional work on the exact site(s), mecha-nism(s) of drug action and related physiological conse-quences has revealed important information on where and how addictive drugs act to mediate their stimulatory effects on dopamine and how these neurochemical changes interface with and shape behavioral and physiological responses [261, 262]. Importantly, drug withdrawal elicited either spontane-ously or after precipitation with specific receptor antagonists seems to result in a consistent reduction of dopamine release in the nucleus accumbens that has been related to dysphoria and emotional distress often exhibited by dependence-producing substances in an abstinence state [263].
The cannabinoid agonist 9-THC increases dopamine
release in the major projection areas of the ascending dopa-minergic system of experimental animals in vivo, as assessed by microdialysis (Table 8), similarly to many, but not all, drugs of abuse. Several methodological parameters, such as the route of administration, the strain of animals used, and the dopaminergic projection area sampled appear to be major determinants of the robustness and reproducibility of this neurochemical effect [90, 223, 264-272]. Tanda et al. [269] in a critical study showed beyond any doubt that both
9-
THC and WIN55,212-2 administered intravenously increase dopamine release selectively in the shell of the nucleus ac-cumbens, through stimulation of CB1 receptors, and con-firmed previous work by Chen et al. [264] that opioid recep-tor stimulation plays an important role in the observed find-ing. Interestingly, chronic administration of
9-THC did not
affect basal release of dopamine in the shell of the nucleus accumbens, but challenge with SR141716A in these animals decreased accumbal dopamine release, representing a neuro-chemical response related to precipitated withdrawal, simi-larly to other drugs of abuse [273]. In direct relevance to behavioral responses that are indicative of the dependence-producing properties of psychotropic compounds, it was re-cently shown that self-administration of WIN55,212-2 is associated with an increase in DA release in the nucleus ac-cumbens [91, 92]. In terms of the effects of endocannabi-noids on dopamine release in the brain, Solinas et al. [155, 274] showed that intravenous, but not intraperitoneal, ad-ministration of anandamide and its longer-lasting analog, methanandamide, increase dopamine release in the nucleus accumbens in a multiphasic manner with a differential in-volvement of the CB1 receptors. It is striking, however, that the same authors did not find an effect of the endocannabi-noid neurotransmission enhancers URB597 and AM404 ad-ministered intraperitonally on accumbal dopamine release, in spite of the fact that these compounds increase endocannabi-noid levels in plasma and brain and potentiate anandamide’s stimulatory action on dopamine release [155, 274]. Thus, the route of administration of endocannabinoids or their modula-tors (see also, [275]) and the mechanism underlying the en-hancement of endocannabinoid neurotransmission seems to be of major importance in determining effects on dopamine release in the nucleus accumbens.

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 367
Taken together, it is safe to conclude that cannabinoid ago-nists and endocannabinoid modulators affect dopamine release in the nucleus accumbens in a very “idiosyncratic”, i.e. related to this particular class of compounds, manner, in which meth-odological variables are critical. Clearly, more work is required to fully understand the pharmacokinetic/pharmacodynamic dimensions of the effects of cannabinoids on dopamine release and the relevance of this phenomenon to their behavioral ac-tions. Specifically, full dose-response curves after different routes of administration in the presence or absence of selective antagonists (or genetic invalidation of cannabinoid targets) and in relation to behavioral or physiological responses need to be obtained, and the site(s) and mechanism(s) of action fully dis-closed. In addition, the actions of endocannabinoids and endo-cannabinoid modulators on this neurochemical parameter need to be studied further, as new selective compounds are devel-oped and the dynamics of endocannabinoid neurotransmission elucidated. To the extent that an enhancement of dopamine release in the nucleus accumbens is directly related to the de-pendence-producing properties of a studied class of com-pounds, it remains of paramount importance to reveal the de-tails of the interaction of cannabinoids with the dopaminergic system in order to assess their addictive potential.
CONCLUDING REMARKS
Different animal models are now available to examine motivational/reinforcing and dependence-producing properties of cannabinoids. These models are similar to those used for other drugs of abuse. Studies using these behavioral models to date suggest that particular experimental conditions are re-quired to reveal cannabinoid motivational/reinforcing proper-ties and abuse potential in experimental animals. Specifically, the majority of the studies using the ICSS paradigm failed to show any reward facilitating effects for
9-THC or other can-
nabinoids, or on the contrary, they present data for anhedonic actions of these compounds, at least in the high dose range. Similarly, most attempts to obtain a robust self-administration of
9-THC or other synthetic cannabinoids, under typical,
drug-free experimental conditions, have been negative and only a limited number of studies report a robust procedure for cannabinoid self-administration. Conflicting results have also been reported in the literature with the conditioned place pref-erence paradigm with various cannabinoids producing either preference or aversion depending on the specific methodologi-cal design. Likewise, cannabinoids do not share with other drugs of abuse the potential to produce clear and robust behav-
Table 8. Neurochemical Effects of Cannabinoids (e.g., Cannabinoid Receptor Agonists, Endocannabinoids, Endocannabinoid
Neurotransmission Enhancers) on Dopamine Release in the Mesotelencephalic Dopaminergic Projection Regions of the
Brain as Assessed by In Vivo Microdialysis
Cannabinoid Species Brain Region Regimen Effect Antagonism Ref.
9-THC Lewis rats N.Accumbens 0.5, 1 mg/kg, IP DA (Naloxone) [264]
9-THC Lewis rats Prefrontal Cortex
(medial)
1, 2 mg/kg, IP DA [265]
9-THC SD rats N.Accumbens 0.5, 1 mg/kg, IP - DA (0.5), DA (1.0) [266]
9-THC Long-Evans rats Striatum
N.Accumbens
1, 10 mg/kg, PO
1 mg/kg, PO
- DA
- DA
[268]
9-THC Lewis rats N.Accumbens 50, 200 μM (local) DA [267]
9-THC SD rats N.Accumbens
(shell, core)
0.15, 0.3 mg/kg, IV DA (shell)
- DA (core)
SR141716A
(Naloxone)
[269]
WIN55,212-2 SD rats N.Accumbens
(shell, core)
0.15, 0.3 mg/kg, IV DA (shell)
- DA (core)
SR141716A
(Naloxone)
[269]
9-THC SD rats N.Accumbens
(shell)
2-12 mg/kg/day, IP
X 8 d
- DA (basal)
DA (SR141716A)
[273]
9-THC Glaxo-Wistar
rats
Striatum 0.5, 2, 5 mg/kg, IV DA SR141716A
(Fluoxetine)
[270]
9-THC SD rats Prefrontal Cortex 1 mg/kg, IV DA SR141716A [271]
9-THC Fischer 344 rats N.Accumbens 0.5, 1 mg/kg, IP - DA (0.5), - DA (1.0) [272]
Anandamide
(meth-)
URB597
SD rats N.Accumbens
(shell)
0.01-10 mg/kg, IV
0.3 mg/kg, IP
DA
- DA
SR141716A
Capsazepine
[274,155]
WIN55,212-2 SD rats N.Accumbens
(shell, core)
12.5 μg/kg, IV, X 3 w
self-administration
DA (shell)
DA (core)
[92]
WIN55,212-2 Lister Hooded,
Long-Evans rats
N.Accumbens
(shell)
12.5 μg/kg, IV,
self-administration
DA (shell)
[91]
Anandamide
AM404
SD rats N.Accumbens
(shell)
10 mg/kg, IP
10 mg/kg, IP
- DA [155]
URB597
OEA, PEA
Wistar rats N.Accumbens
(core)
10 μg/5 μl, ICV DA [275]

368 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
ioral sensitization. Overall, experimental data using the above mentioned paradigms suggest that the experimental conditions required revealing cannabinoid-reinforcing properties are dif-ferent from those used with other classical drugs of abuse.
Although many cannabinoids are found in cannabis sativa preparations, maybe only
9-THC is really abused by humans.
However, cannabis is more than simply 9-THC. Indeed, can-
nabis preparations contain several cannabinoid and non-cannabinoid compounds, which may contribute to the behav-ioral effects of cannabis in humans. After all, however, the pleasurable effects of cannabis preparations in humans appear to be in disagreement with preclinical experimental studies. Nevertheless, most preclinical studies utilize
9-THC or other
synthetic cannabinoid agonists that stimulate cannabinoid re-ceptors. Several other reasons might also explain this differ-ence. There may be significant species differences in functions of the cannabinoid system in animals and humans. Perhaps the recreational use of cannabis by humans reflects more complex sensory and cognitive effects that cannot be experienced by lower animals. This may, also, explain why cannabinoids do not induce the typical pattern of obsessive drug-seeking and compulsive drug-taking behavior, observed in humans ad-dicted to other, more typical drugs of abuse, such as cocaine, heroin, alcohol and nicotine. Pharmacokinetic factors (i.e. factors related to bioavailability of the drug formulation used, lipophilicity, distribution and storage in fat tissue, metabolism and elimination rate) seem to be responsible in part for the subjective effects of cannabinoids. Thus, the possibility still remains, that the abuse of cannabinoids by humans is related to a rewarding action which could be difficult to reveal in the currently used animals models as a consequence of the long duration and slow elimination action of cannabinoid drugs or other unknown factors. Unequivocally, however, preclinical experimental studies utilizing the above mentioned behavioral models have provided a better understanding of the neurobi-ological mechanisms involved in exogenous and endogenous cannabinoids functions and have revealed some commonali-ties between cannabinoids and typical drugs of abuse. On the other hand, studies using the drug discrimination paradigm have shown subjective effects of cannabinoid drugs, that are specific to this group and in general do not generalize to other drugs of abuse. However, we do not know yet the characteris-tics and the significance of those subjective effects. Interest-ingly, similarly with other drugs of abuse, chronic treatment with cannabinoids leads to the development of tolerance to most of their pharmacological effects which probably relates with adaptive changes in the endocannabinoid system accom-panied by CB1 receptors down-regulation and attenuation of G-protein activation. However, there is no conclusive evi-dence for a robust spontaneous withdrawal in experimental animals treated chronically with cannabinoids. Cannabinoid agonists seem to increase dopamine release in the nucleus accumbens under certain experimental conditions, a neuro-chemical marker shared with traditional drugs of abuse. How-ever, critical details of the conditions under which this neuro-chemical effect is effectuated in relation to the physiology of the cannabinoid system remain to be revealed. In the near fu-ture, the explosion of research in the field of cannabinoids and the discovery of optimal pharmacological tools will most cer-tainly also provide insights into the physiological role of the
endogenous cannabinoid system in brain reward and in habit-ual adaptive function.
Key Learning Objectives:
1. Despite the clinical evidence that cannabis derivatives produce clear
subjective motivational responses in humans leading to drug-seeking behavior and in a specific proportion in repeated drug use,
the reinforcing/rewarding attributes of these subjective effects are difficult to define in experimental animals.
2. The majority of the studies using the intracranial self-stimulation
paradigm failed to show any reward facilitating effects for 9-THC or other cannabinoids, or on the contrary, they present data for an-
hedonic actions of these compounds, at least in the high dose range.
3. Most attempts to obtain a robust self-administration of 9-THC or other synthetic cannabinoids, under standard experimental condi-
tions, have been negative. However, several recent studies demon-strate that canabinoids can initiate and sustain active self-
administration in experimental animals, although the range of condi-tions under which this occurs is more limited than with typical drugs
of abuse.
4. There is limited evidence that 9-THC or other cannabinoids can induce the development of conditioned place preference in rodents
under certain experimental conditions.
5. Drug discrimination studies have shown subjective effects of can-nabinoid drugs, that are specific to this group and in general do not
generalize to other drugs of abuse. Thus, the discriminative effects of cannabinoids appear to be neurobiologically distinct involving
most likely other neuronal circuits than those participating in the discriminative effects of classical drugs of abuse.
6. Cannabinoids do not share with other drugs of abuse the potential to
produce clear and robust behavioral sensitization.
7. Chronic treatment with cannabinoids leads to the development of tolerance to most of their pharmacological effects. However, there is
no conclusive evidence for a robust spontaneous withdrawal in ex-perimental animals treated chronically with cannabinoids.
8. Cannabinoid agonists increase dopamine release in the nucleus
accumbens of experimental animals under certain experimental con-ditions.
9. All the above illustrate the differential status of cannabinoids as
atypical drugs of dependence and abuse.
Future Research Questions:
1. The available animal models for measuring the potential reinforc-
ing/rewarding effects of cannabinoids together with the availability of new and more selective compounds that act on the cannabinoid
system provide exciting opportunities to search for drugs that pos-sess the therapeutic potential attributed to cannabis and 9-THC, but
lack the undesirable psychotropic effect of dependence and abuse liability.
2. Careful scrutiny of the commonalities and differences in the rein-
forcing/rewarding properties of cannabinoids and other drugs of abuse should provide a better understanding of the subtle but differ-
ent neurobiological mechanisms underlying their abuse potential.
3. A multidisciplinary approach examining molecular and cellular changes in conjunction with animal models for studying reinforcing
and dependence-producing properties of cannabinoids will undoubt-edly further our current understanding.
4. The neurobiological underpinnings and the physiological conse-
quences of the stimulatory effects of cannabinoids and endocan-nabinoid neurotransmission modifiers on dopamine release in the
nucleus accumbens, in particular in relation to their pertinent moti-vational and dependence-producing actions, remain a subject of ac-
tive research.
5. Detailed research into cannabinoid actions may help to elucidate not only the relevant actions of cannabinoid drugs, but also the role of
the endogenous cannabinoid system in brain reward.

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 369
ACKNOWLEDGEMENTS
This study was supported by the Department of Psychol-ogy of the University of Crete. Styliani Vlachou was sup-ported by a scholarship from PROPONDIS Foundation.
REFERENCES
[1] Green B, Kavanagh D, Young R. Being stoned: a review of self-
reported cannabis effects. Drug Alcohol Rev 2003; 22: 453-60. [2] Budney AJ, Hughes JR, Moore BA, Vandrey R. Review of the
validity and significance of cannabis withdrawal syndrome. Am J Psychiatry 2004; 161: 1967-77.
[3] Chen K, Kandel DB, Davies M. Relationships between frequency and quantity of marijuana use and last year proxydependence
among adolescents and adults in the United States. Drug Alcohol Depend 1997; 46: 53-67.
[4] Swift W, Hall W, Copeland J. One year follow-up of cannabis dependence among long-term users in Sydney, Australia. Drug Al-
cohol Depend 2000; 59: 309-318. [5] Di Marzo V. A brief history of cannabinoid and endocannabinoid
pharmacology as inspired by the work of British scientists. Trends Pharmacol Sci 2006; 27: 134-40.
[6] Gaoni Y, Mechoulam R. Isolation, structure and partial synthesis of an active constituent of hashish. J Am Chem Soc 1964; 86: 1646-7.
[7] Hawks RL. The constituents of cannabis and the disposition and metabolism of cannabinoids. NIDA Res Monogr 1982; 42: 125-37.
[8] Zuardi AW, Crippa JA, Hallak JE, Moreira FA, Guimarães FS. Cannabidiol, a Cannabis sativa constituent, as an antipsychotic
drug. Braz J Med Biol Res 2006; 39: 421-9. [9] Hillard CJ, Harris RA, Bloom AS. Effects of the cannabinoids on
physical properties of brain membranes and phospholipid vesicles: fluorescence studies. J Pharmacol Exp Ther 1985; 232: 579-88.
[10] Matsuda LA, Lolait SJ, Brownstein MJ, Young AC, Bonner TI. Structure of a cannabinoid receptor and functional expression of
the cloned cDNA. Nature 1990; 346: 561-4. [11] Munro S, Thomas KL, Abu-Shaar SM. Molecular characterization
of a peripheral receptor for cannabinoids. Nature 1993; 365: 61-5. [12] Howlett AC, Barth F, Bonner TI, et al. International Union of
Pharmacology: XXVII. Classification of cannabinoid receptors. Pharmacol Rev 2002; 54: 161-202.
[13] Howlett AC. The cannabinoid receptors. Prostaglandin Other Lip-ids Mediat 2002; 68-69: 619-31.
[14] Gong JP, Onaivi ES, Ishiguro H, et al. Cannabinoid CB2 receptors: immunohistochemical localization in rat brain. Brain Res 2006;
1071: 10-23. [15] Van Sickle MD, Duncan M, Kingsley PJ, et al. Identification and
functional characterization of brainstem cannabinoid CB2 recep-tors. Science 2005; 310: 329-32.
[16] Begg M, Pacher P, Bátkai S, et al. Evidence for novel cannabinoid receptors. Pharmacol Ther 2005; 106: 133-45.
[17] Devane WA, Hanus L, Breuer A, et al. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science
1992; 258: 1946-9. [18] Di Marzo V, Fontana A, Cadas H, et al. Formation and inactivation
of endogenous cannabinoid anandamide in central neurons. Nature 1994; 372: 686-91.
[19] Di Marzo V, Melck D, Bisogno T, De Petrocellis L. Endocannabi-noids: Endogenous cannabinoid receptor ligans with neuromodula-
tory function. Trends Neurosci 1998; 21: 521-8. [20] Mechoulam R, Ben-Shabat S, Hanus L, et al. Identification of an
endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem Pharmacol 1995; 50: 83-90.
[21] Hashimotodani Y, Ohno-Shosaku T, Kano M. Endocannabinoids and synaptic function in the CNS. Neuroscientist 2007; 13: 127-37.
[22] Sugiura T, Kobayashi Y, Oka S, Waku K. Biosynthesis and degra-dation of anandamide and 2-arachidonoylglycerol and their possi-
ble physiological significance. Prostaglandines Leukot Essent Fatty Acids 2002; 66: 173-92.
[23] Di Marzo V, Petrosino S. Endocannabinoids and the regulation of their levels in health and disease. Curr Opin Lipidol 2007; 18: 129-
40. [24] Fowler CJ. The cannabinoid system and its pharmacological ma-
nipulation--a review, with emphasis upon the uptake and hydrolysis of anandamide. Fundam Clin Pharmacol 2006; 20: 549-62.
[25] Pertwee RG. The pharmacology of cannabinoid receptors and their
ligands: an overview. Int J Obes 2006; 30: S13-S18. [26] Palmer SL, Thakur GA, Makriyannis A. Cannabinergic ligands.
Chem Phys Lipids 2002; 121: 3-19. [27] Kunos G, Batkai S. Novel physiologic functions of endocannabi-
noids as revealed through the use of mutant mice. Neurochem Res 2001; 26: 1015-21.
[28] Valverde O, Karsak M, Zimmer A. Analysis of the endocannabi-noid system by using CB1 cannabinoid receptor knockout mice.
Handb Exp Pharmacol 2005; 168:117-45. [29] Pertwee RG. Pharmacological actions of cannabinoids. Handb Exp
Pharmacol 2005; 168: 1-51. [30] Makriyannis A, Mechoulam R, Piomelli D. Therapeutic opportuni-
ties through modulation of the endocannabinoid system. Neuro-pharmacology 2005; 48: 1068-71.
[31] Goutopoulos A, Makriyannis A. From cannabis to cannabinergics: new therapeutic opportunities. Pharmacol Ther 2002; 103-17.
[32] Devane W, Dysarz FA, Johnson MR, Melvin LS, Howlett AC. Determination and characterization of a cannabinoid receptor in rat
brain. Mol Pharmacol 1988; 34: 605-13. [33] Houston D, Howlett AC. Differential receptor-G-protein coupling
evoked by dissimilar cannabinoid receptor agonists. Cell Signal 1998; 10: 667-74.
[34] Pertwee RG. Cannabinoid receptor ligands: clinical and neuro-pharmacological considerations, relevant to future drug discovery
and development. Expert Opin Investig Drugs 2000; 1553-71. [35] Ortega-Gutiérrez S. Therapeutic Perspectives of inhibitors of endo-
cannabinoid degradation. Curr Drug Targets CNS Neurol Disord 2005; 4: 697-707.
[36] Cravatt BF, Lichtman AH. Fatty acid amide hydrolase: an emerg-ing therapeutic target in the endocannabinoid system. Curr Opin
Chem Biol 2003; 7: 469-75. [37] Kornetsky C. Brain-stimulation reward: A model for the neuronal
bases for drug induced euphoria. Natl Inst Drug Abuse Res Monogr Ser 1985; 62: 30-50.
[38] Wise RA. Action of drugs of abuse on brain reward systems. Pharmacol Biochem Behav 1980; 13 (Suppl 1): 213-23.
[39] Wise RA. Addictive drugs and brain stimulation reward. 1996; Annu Rev Neurosci 19: 319-40.
[40] Wise RA Drug-activation of brain reward pathways. Drug Alcohol Depend 1998; 51:13-22.
[41] Gardner EL, Paredes W, Smith D, et al. Facilitation of brain stimu-lation reward by delta-9-tetrahydrocannabinol. Psychopharma-
cology 1988; 96: 142-4. [42] Gardner EL, Paredes W, Smith D, Zukin RS. Facilitation of brain
stimulation reward by delta-9-tetrahydrocannabinol is mediated by an endogenous opioid mechanism. Adv Biosci 1989; 75: 671-4.
[43] Gardner EL, Vorel SR. Cannabinoid transmission and reward re-lated events. Neurobiol Dis 1998; 5: 502-33.
[44] Lepore M, Liu X, Savage V, Matalon D, Gardner EL. Genetic differences in 9-tetrahydrocannabinol-induced facilitation of brain
stimulation reward as measured by a rate-frequency curve-shift electrical brain stimulation paradigm in three different rat strains.
Life Sci 1996; 58: 365-72. [45] Stark P, Dews PB. Cannabinoids: Behavioral Effects. J Pharmacol
Exp Ther 1980; 214: 124-30. [46] Kucharski LT, Williams JEG, Kornetsky C. The effects of levonan-
tradol on rewarding brain stimulation thresholds in the rat. Pharma-col Biochem Behav 1983; 19: 149-51.
[47] Vlachou S, Nomikos GG, Stephens D, Panagis G. Lack of evidence for appetitive effects of 9-tetrahydrocannabinol in the intracranial
self-stimulation and conditioned place preference procedures in ro-dents. Behav Pharmacol 2007; 18: 311-19.
[48] Kornetsky C, Esposito RU. Euphorigenic drug: effects on the re-ward pathways of the brain. Fed Proc 1979; 38: 2473-6.
[49] Bauco P, Wise RA. Potentiation of lateral hypothalamic and mid-line mesencephalic brain stimulation reinforcement by nicotine:
examination of repeated treatment. J Pharmacol Exp Ther 1994; 271: 294-301.
[50] Bauco P, Wise RA. Synergistic effects of cocaine with lateral hy-pothalamic brain stimulation reward: lack of tolerance or sensitiza-
tion. J Pharmacol Exp Ther 1997; 283: 1160-7. [51] Lin HQ, Jackson DM, Atrens DM, Christie MJ, McGregor IS.
Serotonergic modulation of 3,4-methylene-dioxymethamphetamine (MDMA)-elicited reduction of response rate but not rewarding

370 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
threshold in accumbal self-stimulation. Brain Res 1997; 744: 351-
7. [52] Arnold JC, Hunt GE, McGregor IS. Effects of the cannabinoid
receptor agonist CP 55,940 and the cannabinoid receptor antagonist SR 141716 on intracranial self-stimulation in Lewis rats. Life Sci
2001; 70: 97-108. [53] Antoniou K, Galanopoulos A, Vlachou S, et al. Behavioral phar-
macological properties of a novel cannabinoid 1’,1’-dithiolane 8-THC analogue, AMG-3. Behav Pharmacol 2005; 16: 499-510.
[54] Vlachou S, Nomikos GG, Panagis G. WIN 55,212-2 decreases the reinforcing actions of cocaine through CB1 cannabinoid receptor
stimulation. Behav Brain Res 2003; 141: 215-22. [55] Vlachou S, Nomikos GG, Panagis G. CB1 cannabinoid receptor
agonists increase intracranial self-stimulation thresholds in the rat. Psychopharmacology 2005; 179: 498-508.
[56] Vlachou S, Nomikos GG, Panagis G. Effects of endocannabinoid neurotransmission modulators on brain stimulation reward. Psy-
chopharmacology 2006; 188: 293-305. [57] Deroche-Gamonet V, Le Moal M, Piazza PV, Soubrié P. SR
141716, a CB1 receptor antagonist, decreases the sensitivity to the reinforcing effects of electrical brain stimulation in rats. Psycho-
pharmacology 2001; 157: 254-9 [58] Bouaboula M, Perrachon S, Milligan L, et al. A selective inverse
agonist for central cannabinoid receptor inhibits mitogen-activated protein kinase activation stimulated by insulin or insulin-like
growth factor 1. Evidence for a new model of receptor/ligand inter-actions. J Biol Chem 1997; 272: 22330-9.
[59] Landsman RS, Burkey TH, Consroe P, Roeske WR, Yamamura HI. SR141716A is an inverse agonist at the human cannabinoid CB1
receptor. Eur J Pharmacol 1997; 334: R1-2. [60] De Vry J, Jentzsch KR. Intrinsic activity estimation of cannabinoid
CB1 receptor ligands in a drug discrimination paradigm. Behav Pharmacol 2003; 14: 471-6.
[61] Christensen R, Kristensen PK, Bartels EM, Bliddal H, Astrup A. Efficacy and safety of the weight-loss drug rimonabant: a meta-
analysis of randomised trials. Lancet 2007; 370: 1706-13. [62] Parker LA, Gillies T. THC-induced place and taste aversion in
Lewis and Sprague-Dawley rats. Behav Neurosci 1995; 109: 71-8. [63] Gardner EL. What we have learned about addiction from animal
models of drug self-administration. Am J Addict 2000; 9: 285-313. [64] Griffiths RR. Common factors in human and infrahuman drug self-
administration, Psychopharmacol Bull 1980; 16: 45–7. [65] Henningfield JE, Cohen C, Heishman SJ. Drug self-administration
methods in abuse liability evaluation. Br J Addict 1991; 86: 1571-7.
[66] Schuster CR, Thompson T. Self administration of and behavioral dependence on drugs. Annu Rev Pharmacol 1969; 9: 483–502.
[67] Young AM, Herling S, Woods JH History of drug exposure as a determinant of drug-self administration. NIDA Res Monogr 1981;
37: 75-88. [68] Wise RA, Bozarth MA Brain substrates for reinforcement and drug
self-administration. Prog Neuropsychopharmacol 1981; 5: 467-74. [69] Yokel RA In: Bozarth MA Ed, Methods of assessing the reinforc-
ing properties of abused drugs. New York, Springer. 1987; 1-33. [70] Carney JM, Uwaydah IM, Balster RL. Evaluation of a suspension
system for intravenous self-administration studies of water-insoluble compounds in the rhesus monkey. Pharmacol Biochem
Behav 1977; 7: 357–64. [71] Corcoran ME, Amit Z. Reluctance of rats to drink hashish suspen-
sions: free choice and forced consumption and the effects of hypo-thalamic stimulation. Psychopharmacologia 1974; 352: 129-47.
[72] Harris RT, Waters W, McLendon D. Evaluation of reinforcing capability of 9-tetrahydrocannabinol in monkeys. Psychopharma-
cologia 1974; 37: 23-9. [73] Leite JL, Carlini EA. Failure to obtain “cannabis directed behavior”
and abstinence syndrome in rats chronically treated with cannabis sativa extracts. Psychopharmacologia 1974; 36: 133-45.
[74] van Ree JM, Slangen J, de Wied D. Intravenous self-administration of drugs in rats. J Pharmacol Exp Ther 1978; 20: 547-57.
[75] Mansbach RS, Nicholson KL, Martin BR, Balster RL. Failure of 9-tetrahydrocannabinol and CP 55,940 to maintain intravenous
self-administration under a fixed-interval schedule in rhesus mon-keys. Behav Pharmacol 1994; 5: 210-25.
[76] Young AM, Katz JL, Woods JH. Behavioral effects of levonantra-dol and nantradol in the rhesus monkey. J Clin Pharmacol 1981b;
21 (Suppl. 8-9): 348S-360S.
[77] Deneau GA, Kaymakcalan S. Physiological and psychological
dependence to synthetic 9-tetrahydrocannabinol (THC) in rhesus monkeys. Pharmacologist 1971; 13: 246-8.
[78] Kaymakcalan S. Tolerance to and dependence on cannabis. Bull Narc 1973; 25: 39-47.
[79] Pickens R, Thompson T, Muchow DC. In: Goldfarb L, Hoffmeister F Eds, Psychic dependence [Bayer Symposium IV]. Berlin Heidel-
berg New York, Springer. 1973; 78-86. [80] Tanda G, Munzar P, Goldberg SR. Self-administration behavior is
maintained by the psychoactive ingredient of marijuana in squirrel monkeys. Nat Neurosci 2000; 3: 1073-74.
[81] Bergman J, Johanson CE. The reinforcing properties of diazepam under several conditions in the rhesus monkey. Psychopharma-
cology 1985; 86: 108–13. [82] Cabeza de Vaca S, Carr KD. Food restriction enhances the central
rewarding effect of abused drugs. J Neurosci 1998; 18: 7502–7510. [83] Campbel UC, Carroll ME. Acquisition of drug-self-administration:
environmental and pharmacological interventions. Exp Clin Psy-chopharmacol 2000; 8: 312-25.
[84] Carr KD. Augmentation of drug reward by chronic food restriction: behavioural evidence and underlying mechanisms. Physiol Behav
2002; 76: 353-64. [85] Carroll ME, France CP, Meisch RA. Food deprivation increases
oral and intravenous drug intake in rats. Science 1979; 205: 319–21.
[86] de la Garza R, Johanson CE. The effects of food deprivation on the self-administration of psychoactive drugs. Drug Alcohol Depend
1987; 19: 17–27. [87] Takahashi RN, Singer G. Self-administration of delta 9-
tetrahydrocannabinol by rats. Pharmacol Biochem Behav 1979; 11: 737–40.
[88] Takahashi RN, Singer G. Effects of body weight levels on cannabis self-injection. Pharmacol Biochem Behav 1980; 13: 877–81.
[89] Fattore L, Cossu G, Martellotta CM, Fratta W. Intravenous self-administration of the cannabinoid CB1 receptor agonist WIN
55,212-2 in rats. Psychopharmacology 2001; 156: 410–16. [90] Fadda P, Scherma M, Spano MS, et al. Cannabinoid self-
administration increases dopamine release in the nucleus accum-bens. Neuroreport 2006; 17: 1629-32.
[91] Fattore L, Vigano D, Fadda P, Rubino T, Fratta W, Parolaro D. Bidirectional regulation of mu-opioid and CB1-cannabinoid recep-
tor in rats self-administering heroin or WIN 55,212-2. Eur J Neuro-sci 2007; 25: 2191-2200.
[92] Lecca D, Cacciapaglia F, Valentini V, Di Chiara G. Monitoring extracellular dopamine in the rat nucleus accumbens shell and core
during acquisition and maintenance of intravenous WIN 55,212-2 self-administration. Psychopharmacology 2006; 188: 63-74.
[93] Martellotta MC, Cossu G, Fattore L, Gessa GL, Fratta W. Self-administration of the cannabinoid receptor agonist WIN 55,212-2
in drug-naive mice. Neuroscience 1998; 85: 327–30. [94] Ledent C, Valverde O, Cossu G, et al. Unresponsiveness to can-
nabinoids and reduced addictive effects of opiates in CB1 knockout mice. Science 1999; 283: 401-4.
[95] Navarro M, Carrera MR, Fratta W, et al. Functional interaction between opioid and cannabinoid receptors in drug self-
administration. J Neurosci 2001; 21: 5344–50. [96] Justinova Z, Tanda G, Redhi GH, Goldberg SR. Self-administration
of 9-tetrahydrocannabinol (THC) by drug naive squirrel monkeys. Psychopharmacology 2003; 169: 135–40.
[97] Justinova Z, Solinas M, Tanda G, Redhi GH, Goldberg SR. The endogenous cannabinoid anandamide and its synthetic analog R(+)-
methanandamide are intravenously self-administered by squirrel monkeys. J Neurosci 2005; 25: 5645-50.
[98] Braida D, Pozzi M, Parolaro D, Sala M. Intracerebral self-administration of the cannabinoid receptor agonist CP 55,940 in the
rat: interaction with the opioid system. Eur J Pharmacol 2001; 413: 227-34.
[99] Braida D, Sala M. 9-tetrahydrocannabinol-induced conditioned place preference and intracerebroventricular self-administration in
rats. Eur J Pharmacol 2004; 506: 63-9. [100] Zangen A, Solinas M, Ikemoto S, Goldberg SR, Wise RA. Two
brain sites for cannabinoid reward. J Neurosci 2006; 26: 4901-7. [101] Bardo MT, Bevins RA. Conditioned place preference: what does it
add to our preclinical understanding of drug reward? Psychophar-macology 2000; 153: 31-43.

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 371
[102] Hoffman DC. The use of place conditioning in studying the neuro-
pharmacology of drug reinforcement. Brain Res Bul 1989; 23: 373-87.
[103] Tzschentke TM. Measuring reward with the conditioned place preference paradigm: a comprehensive review of drug effects, re-
cent progress and new issues. Prog Neurobiol 1998; 56: 613-72. [104] Tzschentke TM. Measuring reward with the conditioned place
preference (CPP) paradigm: update of the last decade. Addict Biol 2007; 12: 227-462.
[105] Sanchis-Segura C, Spanagel R. Behavioural assessment of drug reinforcement and addictive features in rodents: an overview. Ad-
dict Biol 2006; 11: 2-38. [106] Lepore M, Vorel SR, Lowinson J, Gardner EL. Conditioned place
preference induced by 9-tetrahydrocannabinol: comparison with cocaine, morphine and food reward. Life Sci 1995; 56: 2073-80.
[107] Cheer JF, Kendall DA, Marsden CA Cannabinoid receptors and reward in the rat: a conditioned place preference study. Psycho-
pharmacology 2000; 151: 25-30. [108] Elsmore TF, Fletcher GV. Delta-9-tetrahydrocannabinol: aversive
effects in rat at high doses. Science 1972; 175: 911-2. [109] Hutcheson DM, Tzavara ET, Smadja C, et al. Behavioral and bio-
chemical evidence for signs of abstinence in mice chronically treated with 9-tetrahydrocannabinol. Br J Pharmcol 1998; 125:
1567-77. [110] Mallet PE, Beninger RJ. 9 –tetrahydrocannabinol, but not the
endogenous cannabinoid receptor ligand anandamide, produces conditioned place avoidance. Life Sci 1998; 62: 2431-9.
[111] Robinson L, Hinder L, Pertwee RG, Riedel G. Effects of 9-THC and WIN 55,212-2 on place preference in the water maze in rats.
Psychopharmacology 2003; 166: 40-50. [112] Sañudo-Peña MC, Tsou K, Delay ER, Hohman AG, Force M,
Walker JM. Endogenous cannabinoids as an aversive or counter-rewarding system in the rat. Neurosci Lett 1997; 223: 125-8.
[113] Zimmer A, Valjent E, Konig M, et al. Absence of 9-tetrahydrocannabinol dysphoric effects in dynorphin-deficient
mice. J Neurosci 2001; 21: 9499-505. [114] Le Foll B, Wiggins M, Goldberg SR. Nicotine pre-exposure does
not potentiate the locomotor or rewarding effects of -9-tetrahydrocannabinol in rats. Behav Pharmacol 2006; 17: 195-9.
[115] Valjent E, Maldonado R. A behavioural model to reveal place preference to 9-tetrahydrocannabinol in mice. Psychopharma-
cology 2000; 147: 436-8. [116] Soria G, Castañé A, Berrendero F, et al. Adenosine A2A receptors
are involved in physical dependence and place conditioning in-duced by THC. 2004; Eur J Neurosci 20: 2203-13.
[117] McGregor IS, Issakidis CN, Prior G Aversive effects of the syn-thetic cannabinoid CP 55,940 in rats. Pharmacol Biochem Behav
1996; 53: 657-64. [118] Chaperon F, Soubrie P, Puech AJ, Thiebot MH. Involvement of
central cannabinoid (CB1) receptors in the establishment of place conditioning in rats. Psychopharmacology 1998; 135: 324-32.
[119] Braida D, Pozzi M, Cavallini R, Sala M. Conditioned place prefer-ence induced by the cannabinoid agonist CP 55,940: interaction
with the opioid system. Neuroscience 2001; 104: 923-6. [120] Castané A, Maldonado R, Valverde O. Role of different brain
structures in the behavioural expression of WIN 55,212-2 with-drawal in mice. Br J Pharmacol 2004; 142: 1309-17.
[121] Gobbi G, Bambico FR, Mangieri R, et al. Antidepressant-like activity and modulation of brain monoaminergic transmission by
blockade of anandamide hydrolysis. Proc Natl Acad Sci USA 2005; 102: 18620-5.
[122] Bortolato M, Campolongo P, Mangieri RA, et al. Anxiolytic-Like Properties of the anandamide transport inhibitor AM404. Neuro-
psychopharmacology 2006; 31: 2652-9. [123] Colpaert FV. Drug discrimination in neurobiology. Pharmacol
Biochem Behav 1999; 64: 337-45. [124] Stolerman IP, Rasul F, Shine PG. Trends in drug discrimination
research analyzed with a cross-indexed bibliography, 1984-1987. Psychopharmacology 1989; 98: 1-19.
[125] Balster RL. Drug abuse potential evaluation in animals. Br J Addict 1991; 86: 1549-58.
[126] Katz JL, Goldberg SR. Preclinical assessment of abuse liability of drugs. Agents Actions 1988; 23: 18-26.
[127] Balster RL, Prescott WR. Delta 9-tetrahydrocannabinol discrimina-tion in rats as a model for cannabis intoxication. Neurosci Biobe-
hav Rev 1992; 16: 55-62.
[128] Barrett RL, Wiley JL, Balster RL, Martin BR. Pharmacological
specificity of 9-tetrahydrocannabinol discrimination in rats. Psy-chopharmacology 1995; 118: 419-24.
[129] De Vry J, Jentzsch KR. Discriminative stimulus effects of the structurally novel cannabinoid CB1/CB2 receptor partial agonist
BAY 59-3074 in the rat. Eur J Pharmacol 2004; 505: 127-33. [130] Gold LH, Balster RL, Barrett RL, Britt DT, Martin BR. A compari-
son of the discriminative stimulus properties of delta 9-tetrahydrocannabinol and CP 55,940 in rats and rhesus monkeys. J
Pharmacol Exp Ther 1992; 262: 479-86. [131] Wiley JL, Barrett RL, Balster RL, Martin BR Tolerance to the
discriminative stimulus effects of delta 9-tetrahydrocannabinol. Behav Pharmacol 1993; 4: 581-5.
[132] Wiley JL, Barrett RL, Britt DT, Balster RL, Martin BR. Discrimi-native stimulus effects of 9-tetrahydrocannabinol and 9-11-
tetrahydrocannabinol in rats and rhesus monkeys. Neuropharma-cology 1993b; 32: 359-65.
[133] Wiley JL, Balster RL, Martin BR. Discriminative stimulus effects of anandamide in rats. Eur J Pharmacol 1995; 276: 49-54.
[134] Wiley JL, Huffman JW, Balster RL, Martin BR. Pharmacological specificity of the discriminative stimulus effects of 9-
tetrahydrocannabinol in rhesus monkeys. Drug Alcohol Depend 1995b; 40: 81-6.
[135] McMahon LR. Characterization of cannabinoid agonists and appar-ent pA2 analysis of cannabinoid antagonists in rhesus monkeys
discriminating 9-tetrahydrocannabinol. J Pharmacol Exp Ther 2006; 319: 1211-8.
[136] Järbe TUC, Lamb RJ, Lin S, Makriyannis A. (R)-Methanandamide and 9-THC as discriminative stimuli in rats: tests with the can-
nabinoid antagonist SR-141716 and the endogenous ligand anan-damide. Psychopharmacology 2001; 156: 369-80.
[137] Järbe TUC, Liu Q, Makriyannis A. Antagonism of discriminative stimulus effects of 9-THC and (R)-methanandamide in rats. Psy-
chopharmacology 2006; 184: 36-45. [138] Mansbach RS, Rovetti CC, Lowe JA, Winston EN. Effects of the
cannabinoid CB1 receptor antagonist SR141716A on the behavior of pigeons and rats. Psychopharmacology 1996; 124: 315-22.
[139] Pério A, Rinaldi-Carmona M, Maruani J, Barth F, Le Fur G, Sou-brie P. Central mediation of the cannabinoid cue: activity of a se-
lective CB1 antagonist, SR 141716A. Behav Pharmacol 1996; 7: 65-71.
[140] Kallman MJ, Kaempf GL, Harris LS. Discriminative stimulus properties of intraventricular delta-9-tetrahydrocannabinol. Fed
Proc 1984; 43: 952. [141] Brown RG, Weissman A. Discriminative stimulus properties of 9-
tetrahydrocannabinol. Mechanistic studies. J Clin Pharmacol 1981; 21: 227S-34S.
[142] Järbe TUC, Lamb RJ, Makriyannis A, Lin S, Goutopoulos A. 9-THC training dose as a determinant for (R)-methanandamide gen-
eralization in rats. Psychopharmacology 1998; 140: 519-22. [143] Järbe TUC, Lamb RJ. Effects of lithium dose (UCS) on the acquisi-
tion and extinction of a discriminated morphine aversion: tests with morphine and 9-THC. Behav Pharmacol 1999; 10: 349-58.
[144] Mokler DJ, Nelson BD, Harris LS, Rosecrans, JA. The role of benzodiazepine receptors in the discriminative stimulus properties
of delta-9-THC. Life Sci 1986; 38: 1581–89. [145] Wiley JL, Martin BR. Effects of SR141716A on diazepam substitu-
tion for 9-tetrahydrocannabinol in rat drug discrimination. Phar-macol Biochem Behav 1999; 64: 519-22.
[146] Solinas M, Zangen A, Thiriet N, Goldberg SR. -Endorphin eleva-tions in the ventral tegmental area regulate the discriminative ef-
fects of delta-9-tetrahydrocannabinol. Eur J Neurosci 2004; 19:3183–92.
[147] Solinas M, Goldberg SR. Involvement of mu-, delta- and kappa-opioid receptor subtypes in the discriminative-stimulus effects of
delta-9-tetrahydrocannabinol (THC) in rats. Psychopharmacology 2005; 179: 804-12.
[148] Solinas M, Scherma M, Tanda G, Wertheim CE, Fratta W, Gold-berg SR. Nicotinic facilitation of 9-tetrahydrocannabinol dis-
crimination involves endogenous anandamide. J Pharmacol Exp Ther 2007; 321: 1127-34.
[149] Burkey RT, Nation JR. (R)-methanandamide, but not anandamide, substitutes for 9-THC in a drug-discrimination procedure. Exp
Clin Psychopharmacol 1997; 5: 195-202. [150] Wiley JL, Golden KM, Ryan WJ, Balster RL, Razdan RK, Martin
BR. Evaluation of cannabimimetic discriminative stimulus effects

372 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
of anandamide and methylated fluoroanandamide in rhesus mon-
keys. Pharmacol Biochem Behav 1997; 58: 1139-43. [151] Wiley JL, Ryan WJ, Razdan RK, Martin BR. Evaluation of can-
nabimimetic effects of structural analogs of anandamide in rats. Eur J Pharmacol 1998; 355: 113-8.
[152] Wiley JL, Lowe JA, Balster RL, Martin BR. Antagonism of the discriminative stimulus effects of delta 9-tetrahydrocannabinol in
rats and rhesus monkeys. J Pharmacol Exp Ther 1995; 275: 1-6. [153] Alici T, Appel JB. Increasing the selectivity of the discriminative
stimulus effects of 9-tetrahydrocannabinol: complete substitution with methanandamide. Pharmacol Biochem Behav 2004; 79: 431-
7. [154] Wiley JL, LaVecchia KL, Karp NE, et al. A comparison of the
discriminative stimulus effects of delta(9)-tetrahydrocannabinol and O-1812, a potent and metabolically stable anandamide analog,
in rats. Exp Clin Psychopharmacol 2004; 12: 173-9. [155] Solinas M, Tanda G, Justinova Z, et al. The endogenous cannabi-
noid anandamide produces 9-tetrahydrocannabinol-like discrimi-native and neurochemical effects that are enhanced by inhibition of
fatty acid amide hydrolase but not by inhibition of anandamide transport. J Pharmacol Exp Ther 2007b; 321: 370-80.
[156] Robinson TE, Berridge KC. The neural basis of drug craving: an incentive sensitization theory of addiction. Brain Res Rev 1993; 18:
247-91. [157] Steward J, Badiani A. Tolerance and sensitization to the behavioral
effects of drugs. Behav Pharmacol 1993; 4: 289-312. [158] Cadoni C, Pisanu A, Solinas M, Acquas E, Di Chiara G. Behav-
ioural sensitization after repeated exposure to 9-tetrahydrocannabinol and cross-sensitization with morphine. Psy-
chopharmacology 2001; 158: 259-66. [159] Rubino T, Vigano D, Massi P, Parolaro D. The psychoactive ingre-
dient of marijuana induces behavioural sensitization. Eur J Neuro-sci 2001; 14: 884-6.
[160] Rubino T, Vigano D, Massi P, Parolaro D. Cellular mechanisms of 9-tetrahydrocannabinol behavioural sensitization. Eur J Neurosci
2003;17: 325-30. [161] Cadoni C, Di Chiara G. Reciprocal changes in dopamine respon-
siveness in the nucleus accumbens shell and core and in the dorsal caudate-putamen in rats sensitized to morphine. Neuroscience
1999; 90: 447-55. [162] Leith NJ, Kuczenski R. Two dissociable components of behavioral
sensitization following repeated amphetamine administration. Psy-chopharmacology 1982; 76: 310-15.
[163] Panagis G, Nisell M, Nomikos GG, Chergui K, Svensson TH. Nicotine injections into the ventral tegmental area increase locomo-
tion and Fos-like immunoreactivity in the nucleus accumbens of the rat. Brain Res 1996; 730: 133-42.
[164] Shuster L, Webster GW, Yu G. Increased running response to morphine in morphine-pretreated mice. J Pharmacol Exp Ther
1975; 192: 64-7. [165] Varvel SA, Martin BR, Lichtman AH. Lack of behavioral sensitiza-
tion after repeated exposure to THC in mice and comparison to methamphetamine. Psychopharmacology 2007; 193: 517-9.
[166] Kolb B, Gorny G, Limebeer CL, Parker LA. Chronic treatment with 9-tetrahydrocannabinol alters the structure of neurons in the
nucleus accumbens shell and medial prefrontal cortex of rats. Syn-apse 2006; 60: 429-36.
[167] Wu X, French ED. Effects of chronic 9-tetrahydrocannabinol on rat midbrain dopamine neurons: an electrophysiological assess-
ment. Neuropharmacology 2000; 39: 391-8. [168] Bohn LM, Gainetdinov RR, Lin FT, Lefkowitz RJ, Caron MG. μ-
opioid receptor desensitization by -arrestin-2 determines morphine tolerance but not dependence. Nature 2000; 408: 720-3.
[169] Chaperon F, Thiébot MH. Behavioral effects of cannabinoid agents in animals. Crit Rev Neurobiol 1999; 13: 243-81.
[170] Rodriguez de Fonseca F, Del Arco I, Bermudez-Silva FJ, Bilbao A, Cippitelli A, Navarro M. The endocannabinoid system: physiology
and pharmacology. Alcohol Alcohol 2005; 40: 2-14. [171] González S, Cebeira M, Fernández-Ruiz J. Cannabinoid tolerance
and dependence: A review of studies in laboratory animals. Phar-macol Biochem Behav 2005; 81: 300-18.
[172] Lichtman AH, Martin BR. Marijuana withdrawal syndrome in the animal model. J Clin Pharmacol 2002; 42: 20S–27S.
[173] Maldonado R. Study of cannabinoid dependence in animals. Phar-macol Ther 2002; 95: 153–64.
[174] Abood ME, Martin BR. Neurobiology of marijuana abuse. Trends
Pharmacol Sci 1992; 13: 201–6. [175] Bass CE, Martin BR. Time course for the induction and mainte-
nance of tolerance to 9-tetrahydrocannabinol in mice. Drug Alco-hol Depend 2000; 60: 113–9.
[176] da Silva GE, Morato GS, Takahashi RN. Rapid tolerance to 9-tetrahydrocannabinol and cross-tolerance between ethanol and 9-
tetrahydrocannabinol in mice. Eur J Pharmacol 2001; 431: 201-7. [177] McMillan DE, Dewey WL, Harris, LS. Characteristics of tetrahy-
drocannabinol tolerance. Ann NY Acad Sci 1971; 191: 83–99. [178] Fan F, Compton DR, Ward S, Melvin L, Martin BR. Development
of cross-tolerance between 9-tetrahydrocannabinol, CP 55,940 and WIN 55,212. J Pharmacol Exp Ther 1994; 271: 1383–90.
[179] Oviedo A, Glowa J, Herkenham M. Chronic cannabinoid admini-stration alters cannabinoid receptor binding in rat brain: a quantita-
tive autoradiographic study. Brain Res 1993; 616: 293–302. [180] Pertwee RG, Stevenson LA, Griffin G. Cross-tolerance between
9-tetrahydrocannabinol and the cannabimimetic agents, CP 55,940, WIN 55,212-2 and anandamide. Br J Pharmacol 1993; 110:
1483–90. [181] Wiley JL, Smith FL, Razdan RK, Dewey WL. Task specificity of
cross-tolerance between 9-tetrahydrocannabinol and anandamide analogs in mice. Eur J Pharmacol 2005; 510: 59-68.
[182] De Vry J, Denzer D, Reissmueller E, et al. 3-[2-cyano-3-(trifluoromethyl)phenoxy]phenyl-4,4,4-trifluoro-1-butanesulfonate
(BAY 59-3074): A novel cannabinoid CB1/CB2 receptor partial agonist with antihyperalgesic and antiallodynic effects. J Pharma-
col Exp Ther 2004; 310: 620-32. [183] De Vry J, Jentzsch KR, Kuhl E, Eckel G. Behavioral effects of
cannabinoids show differential sensitivity to cannabinoid receptor blockade and tolerance development. Behav Pharmacol 2004b; 15:
1-12. [184] Deadwyler SA, Heyser CJ, Hampson RE, Complete adaptation to
the memory disruptive effects of 9-tetrahydrocannabinol follow-ing 35 days of exposure. Neurosci Res Commun 1995; 17: 9–18.
[185] Felder CC, Glass M. Cannabinoid receptors and their endogenous agonists. Annu Rev Pharmacol Toxicol 1998; 38: 179-200.
[186] Ferraro DP, Grilly DM. Lack of tolerance to 9-tetrahydrocanna-binol in chimpanzees. Science 1973; 179: 490–2.
[187] González S, Manzanares J, Berrendero F, et al. Identification of endocannabinoids and cannabinoid CB1 receptor mRNA in the pi-
tuitary gland. Neuroendocrinology 1999; 70: 137–45. [188] Hampson RE, Simeral JD, Kelly EJ, Deadwyler SA. Tolerance to
the memory disruptive effects of cannabinoids involves adaptation by hippocampal neurons. Hippocampus 2003; 13: 543–56.
[189] Costa N, Colleoni M. Changes in rat brain energetic metabolism after exposure to anandamide or 9-tetrahydrocannabinol. Eur J
Pharmacol 2000; 395: 1-7. [190] Costa B, Giagnoni G, Colleoni M. Precipitated and spontaneous
withdrawal in rats tolerant to anandamide. Psychopharmacology 2000; 149: 121–8.
[191] Fride E. Anandamides: tolerance and cross-tolerance to 9-tetrahydrocannabinol. Brain Res 1995; 697: 83–90.
[192] Rubino T, Vigano D, Costa B, Colleoni M, Parolaro D. Loss of cannabinoid-stimulated guanosine 5 -O-(3-[35S]Thiotriphosphate)
binding without receptor down-regulation in brain regions of anan-damide-tolerant rats. J Neurochem 2000; 75: 2478-84.
[193] Welch SP. Characterization of anandamide-induced tolerance: comparison to 9-THC-induced interactions with dynorphinergic
systems. Drug Alcohol Depend 1997; 45: 39–45. [194] Welch SP, Dunlow LD, Patrick GS, Razdan RK. Characterization
of anandamide-and fluoroanandamide-induced antinociception and cross-tolerance to 9-THC after intrathecal administration to mice:
blockade of 9-THC-induced antinociception. J Pharmacol Exp Ther 1995; 273: 1235–44.
[195] Costa B, Parolaro D, Colleoni M. Chronic cannabinoid, CP-55,940, administration alters biotransformation in the rat. Eur J Pharmacol
1996; 313: 17–24. [196] Dewey WL, McMillan DE, Harris LS, Turk RF. Distribution of
radioactivity in brain of tolerant and nontolerant pigeons treated with 3H- 9-tetrahydrocannabinol. Biochem Pharmacol 1973; 22:
399–405. [197] Magour S, Copher H, Fahndrich C. Is tolerance to delta-9-THC
cellular or metabolic? The subcellular distribution of delta-9-tetrahydrocannabinol and its metabolites in brains of tolerant and
non-tolerant rats. Psychopharmacology 1977; 51: 141–5.

Behavioral Pharmacology of Cannabinoids Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 373
[198] Siemens A, Kalant H. Metabolism of 1-tetrahydrocannabinol by
rats tolerant to cannabis. Can J Physiol Pharmacol 1974; 52: 1154–66.
[199] Martin BR, Sim-Selley LJ, Selley DE. Signaling pathways in-volved in the development of cannabinoid tolerance. Trends Phar-
macol Sci 2004; 25: 325–30. [200] Romero J, García-Gil L, Fernández-Ruiz J, Cebeira M, Ramos JA.
Changes in rat brain cannabinoid binding sites after acute or chronic exposure to their endogenous agonist, anandamide, or to
9-tetrahydrocannabinol. Pharmacol Biochem Behav 1995; 51: 731–7.
[201] Abood ME, Sauss C, Fan F, Tilton CL, Martin BR. Development of behavioral tolerance to 9-tetrahydrocannabinol without altera-
tion of cannabinoid receptor binding or mRNA levels in whole brain. Pharmacol Biochem Behav 1993; 46: 575–9.
[202] Di Marzo V, Berrendero F, Bisogno T, et al. Enhancement of anandamide formation in the limbic forebrain and reduction of en-
docannabinoid contents in the striatum of 9-tetrahydrocannabinol-tolerant rats. J Neurochem 2000; 74: 1627–35.
[203] González S, Di Marzo V, Fernández-Ruiz J, et al. Behavioral and molecular changes elicited by acute administration of SR141716 to
9-tetrahydrocannabinol-tolerant rats: an experimental model of cannabinoid abstinence. Drug Alcohol Depend 2004; 74: 159-174.
[204] Westlake TM, Howlett AC, Ali SF, Paule MG, Scallet AC, Slikker JrW. Chronic exposure to 9-tetrahydrocannabinol fails to irre-
versibly alter brain cannabinoid receptor. Brain Res 1991; 544: 145–9.
[205] Hart CL, Ward AS, Haney M, Comer SD, Foltin RW, Fischman MW. Comparison of smoked marijuana and oral 9-
tetrahydrocannabinol in humans. Psychopharmacology 2002; 164: 407-15.
[206] Iversen LL. The science of marijuana. Oxford, Oxford University Press. 2000.
[207] Kirk JM, de Wit H. Responses to oral 9-tetrahydrocannabinol in frequent and infrequent marijuana users. Pharmacol Biochem Be-
hav 1999; 63: 137-42. [208] Perez-Reyes M, White WR, McDonald SA, Hicks RE, Jeffcoat AR,
Cook CE. The pharmacologic effects of daily marijuana smoking in humans. Pharmacol Biochem Behav 1991; 40: 691-4
[209] Whitlow CT, Freedland CS, Porrino LJ. Functional consequences of the repeated administration of 9-tetrahydrocannabinol in the
rat. Drug Alcohol Depend 2003; 71: 169-77. [210] Haney M, Ward AS, Comer SD, Foltin RW, Fischman MW. Absti-
nence symptoms following smoked marijuana in humans. Psycho-pharmacology 1999; 141: 395-404.
[211] Martin BR, Compton DR, Thomas BF, et al. Behavioral, biochemi-cal, and molecular modeling evaluations of cannabinoid analogs.
Pharmacol Biochem Behav 1991; 40: 471-8 [212] Koob GF, Ahmed SH, Boutrel B, et al. Neurobiological mecha-
nisms in the transition from drug use to drug dependence. Neurosci Biobehav Rev 2004; 27: 739-49.
[213] Le Moal M, Koob GF. Drug addiction: pathways to the disease and pathophysiological perspectives. Eur Neuropsychopharmacol 2007;
17: 377-93. [214] Lyvers M. Drug addiction as a physical disease: the role of physical
dependence and other chronic drug-induced neurophysiological changes in compulsive drug self-administration. Exp Clin Psycho-
pharmacol 1998; 6: 107-25. [215] Barr AM, Markou A, Phillips AG. A 'crash' course on psy-
chostimulant withdrawal as a model of depression. Trends Pharma-col Sci 2002; 23: 475-82.
[216] Markou A, Koob GF. Postcocaine anhedonia. An animal model of cocaine withdrawal. Neuropsychopharmacology 1991; 4:17-26.
[217] Paterson NE, Markou A. Animal models and treatments for addic-tion and depression co-morbidity. Neurotox Res 2007;11: 1-32.
[218] Aceto MD, Scates SM, Lowe JA, Martin BR. Dependence on 9-tetrahydrocannabinol: studies on precipitated and abrupt with-
drawal, J Pharmacol Exp Ther 1996; 278: 1290–95. [219] Chesher GB, Jackson DM. The effect of withdrawal from cannabis
on pentylentetrazol convulsive threshold in mice. Psychopharma-cologia 1974; 40: 129–35.
[220] Dewey WL, Jenkins J, O'Rourke T, Harris LS. The effects of chronic administration of trans- 9-tetrahydrocannabinol on behav-
ior and the cardiovascular system of dogs. Arch Int Pharmacodyn Ther 1972; 198: 118–31.
[221] Diana M, Melis M, Muntoni AL, Gessa GL. Mesolimbic dopa-
minergic decline after cannabinoid withdrawal. Proc Natl Acad Sci USA 1998; 95: 10269–73.
[222] McMillan DE, Harris LS, Frankenheim JM, Kennedy JS. L- 9-tetrahydrocannabinol in pigeons: tolerance to the behavioral ef-
fects. Science 1970; 169: 501–3. [223] Tanda G, Goldberg SR. Cannabinoids: reward, dependence, and
underlying neurochemical mechanisms-a review of recent preclini-cal data. Psychopharmacology 2003; 169: 115-34.
[224] Smith NT. A review of the published literature into cannabis with-drawal symptoms in human users. Addiction 2002; 97: 621-32.
[225] Budney AJ, Moore BA, Vandrey RG, Hughes JR. The time course and significance of cannabis withdrawal. J Abnorm Psychol 2003;
112:393-402. [226] Kaymakcalan S, Ayhan Ih, Tulunay FC. Naloxone-induced or
postwithdrawal abstinence signs in 9-tetrahydrocannabinol-tolerant rats. Psychopharmacology 1977; 55: 243-9.
[227] Beardsley PM, Balster RL, Harris LS. Dependence on tetrahydro-cannabinol in rhesus monkeys. J Pharmacol Exp Ther 1986; 239:
311–19. [228] Beardsley PM, Martin BR. Effects of the cannabinoid CB1 receptor
antagonist, SR141716A, after 9-tetrahydrocannabinol withdrawal. Eur J Pharmacol 2000; 387: 47–53.
[229] Fredericks AB, Benowitz NL. An abstinence syndrome following chronic administration of delta-9-terahydrocannabinol in rhesus
monkeys. Psychopharmacology 1980; 71: 201-2. [230] Aceto MD, Scates SM, Martin BR. Spontaneous and precipitated
withdrawal with a synthetic cannabinoid, WIN55,212-2. Eur J Pharmacol 2001; 416: 75-81.
[231] Oliva JM, Ortiz S, Palomo T, Manzanares J. Behavioural and gene transcription alterations induced by spontaneous cannabinoid with-
drawal in mice. J Neurochem 2003; 85: 94–104. [232] Oliva JM, Ortiz S, Palomo T, Manzanares J. Spontaneous cannabi-
noid withdrawal produces a differential time-related responsiveness in cannabinoid CB1 receptor gene expression in the mouse brain. J
Psychopharmacol 2004; 18: 59-65. [233] Moreno M, Lopez-Moreno JA, Rodriguez de Fonseca F, Navaro
M. Behaviural effects of quinpirole following withdrawal of chronic treatment with the CB1 agonist, HU-210, in rats. Behav
Pharmacol 2005; 16: 441-6. [234] Aceto MD, Scates SM, Lowe JA, Martin BR. Cannabinoid precipi-
tated withdrawal by the selective cannabinoid receptor antagonist, SR 141716A. Eur J Pharmacol 1995; 282: R1-2.
[235] Anggadiredja K, Yamaguchi T, Tanaka H, et al. Prostaglandin E2 attenuates SR141716A-precipitated withdrawal in tetrahydrocan-
nabinol-dependent mice. Brain Res 2003; 966: 47-53. [236] Breivogel CS, Scates SM, Beletskaya IO, Lowery OB, Aceto MD,
Martin BR. The effects of 9-tetrahydrocannabinol physical de-pendence on brain cannabinoid receptors. Eur J Pharmacol 2003;
459: 139-150. [237] Castané A, Robledo P, Matifas A, Kieffer BL, Maldonado R. Can-
nabinoid withdrawal syndrome is reduced in double mu and delta opioid receptor knockout mice. Eur J Pharmacol 2003; 17: 155-9.
[238] Cook SA, Lowe JA, Martin BR. CB1 receptor antagonist precipi-tates withdrawal in mice exposed to 9-tetrahydrocannabinol. J
Pharmacol Exp Ther 1998; 285: 1150-6. [239] Litchtman AH, Sheikh HH, Loh SM, Martin BR. Opioid and can-
nabinoid modulation of precipitated withdrawal in 9-tetrahydrocanabinol and morphine-dependent mice. J Pharmacol
Exp Ther 2001; 298: 1007–14. [240] Sañudo-Peña MC, Force M, Tsou K, McLemore G, Roberts L,
Walker JM. Dopaminergic system does not play a major role in the precipitated cannabinoid withdrawal syndrome. Acta Pharmacol
Sin 1999; 20: 1121–4. [241] Tourino C, Maldonado R, Valverde O. MDMA attenuates THC
withdrawal syndrome in mice. Psychopharmacology 2007; 193: 75-84.
[242] Tsou K, Patrick SL, Walker JM. Physical withdrawal in rats toler-ant to 9-tetrahydrocannabinol precipitated by a cannabinoid recep-
tor antagonist. Eur J Pharmacol 1995; 280: R13-R15. [243] Tzavara ET, Valjent E, Firmo C, et al. Cannabinoid withdrawal is
dependent upon PKA activation in the cerebellum. Eur J Neurosci 2000; 12: 1038–46.
[244] Wilson DM, Varvel SA, Harloe JP, Martin BR, Lichtman AH. SR 141716A (Rimonabant) precipitates withdrawal in marijuana-
dependent mice. Pharmacol Biochem Behav 2006; 85: 105-13.

374 Current Drug Abuse Reviews, 2008, Vol. 1, No. 3 Panagis et al.
[245] Caberlotto L, Rimondini R, Hansson A, Eriksson S, Heilig M.
Corticotropin-releasing hormone (CRH) mRNA expression in rat central amygdala in cannabinoid tolerance and withdrawal: evi-
dence for an allostatic shift? Neuropsychopharmacology 2004; 29: 15-22.
[246] Cui SS, Bowen RC, Gu GB, Hannesson DK, Yu PH, Zhang X. Prevention of cannabinoid withdrawal syndrome by lithium: in-
volvement of oxytocinergic neuronal activation. J Neurosci 2001; 21: 9867-76.
[247] Moranta D, Esteban S, García-Sevilla JA. Acute, chronic and with-drawal effects of the cannabinoid receptor agonist WIN55212-2 on
the sequential activation of MAPK/Raf-MEK-ERK signaling in the rat cerebral frontal cortex: short-term regulation by intrinsic and
extrinsic pathways. J Neurosci Res 2007; 85: 656-67. [248] Rodriguez de Fonseca F, Carrera MRA, Navarro M, Koob GF,
Weiss F. Activation of corticotropin-releasing factor in the limbic system during cannabinoid withdrawal. Science 1997; 276: 2050–
4. [249] Rubino T, Patrini G, Massi P, et al. Cannabinoid-precipitated with-
drawal: a time-course study of the behavioral aspect and its correla-tion with cannabinoid receptors and G protein expression. J Phar-
macol Exp Ther 1998; 285: 813-9. [250] Lichtman AH, Wiley JL, LaVecchia KL, et al. Effects of SR
141716A after acute or chronic cannabinoid administration in dogs. Eur J Pharmacol 1998; 357: 139-48.
[251] Aceto MD, Scates SM, Razdan RK, Martin BR. Anandamide, an endogenous cannabinoid, has a very low physical dependence po-
tential. J Pharmacol Exp Ther 1998; 287: 598-605. [252] Justinova Z, Goldberg SR, Heishman SJ, Tanda G. Self-
administration of cannabinoids by experimental animals and human marijuana smokers. Pharmacol Biochem Behav 2005; 81: 285-99.
[253] Di Chiara G, Imperato A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic sys-
tem of freely moving rats. Proc Natl Acad Sci USA 1988; 85: 5274-8.
[254] Berridge KC. The debate over dopamine's role in reward: the case for incentive salience. Psychopharmacology 2007; 191: 391-431.
[255] Pierce RC, Kumaresan V. The mesolimbic dopamine system: the final common pathway for the reinforcing effect of drugs of abuse?
Neurosci Biobehav Rev 2006; 30: 215-38. [256] Di Chiara. Nucleus accumbens shell and core dopamine: differen-
tial role in behavior and addiction. Behav Brain Res 2002; 137: 75-114.
[257] Pontieri FE, Tanda G, Di Chiara G. Intravenous cocaine, morphine, and amphetamine preferentially increase extracellular dopamine in
the "shell" as compared with the "core" of the rat nucleus accum-bens. Proc Natl Acad Sci USA 1995; 92:12304-8.
[258] Cadoni C, Solinas M, Di Chiara G. Psychostimulant sensitization: differential changes in accumbal shell and core dopamine. Eur J
Pharmacol 2000; 388: 69-76. [259] Fiorino DF, Phillips AG. Facilitation of sexual behavior and en-
hanced dopamine efflux in the nucleus accumbens of male rats af-ter D-amphetamine-induced behavioral sensitization. J Neurosci
1999; 19: 456-63. [260] Lecca D, Valentini V, Cacciapaglia F, Acquas E, Di Chiara G.
Reciprocal effects of response contingent and noncontingent intra-venous heroin on in vivo nucleus accumbens shell versus core do-
pamine in the rat: a repeated sampling microdialysis study. Psy-chopharmacology 2007; 194: 103-16.
[261] Di Chiara G, Bassareo V. Reward system and addiction: what dopamine does and doesn't do. Curr Opin Pharmacol 2007; 7: 69-
76. [262] Kauer JA, Malenka RC. Synaptic plasticity and addiction. Nat Rev
Neurosci 2007; 8: 844-58. [263] Weiss F, Koob GF. Drug addiction: functional neurotoxicity of the
brain reward systems. Neurotox Res 2001; 3: 145-56.
[264] Chen JP, Paredes W, Li J, Smith D, Lowinson J, Gardner EL. Delta
9-tetrahydrocannabinol produces naloxone-blockable enhancement of presynaptic basal dopamine efflux in nucleus accumbens of con-
scious, freely-moving rats as measured by intracerebral microdialy-sis. Psychopharmacology 1990; 102: 156-62.
[265] Chen J, Paredes W, Lowinson JH, Gardner EL. Delta 9-tetrahydrocannabinol enhances presynaptic dopamine efflux in me-
dial prefrontal cortex. Eur J Pharmacol 1990b; 190: 259-62. [266] Chen JP, Paredes W, Lowinson JH, Gardner EL. Strain-specific
facilitation of dopamine efflux by delta 9-tetrahydrocannabinol in the nucleus accumbens of rat: an in vivo microdialysis study. Neu-
rosci Lett 1991; 129: 136-40. [267] Chen J, Marmur R, Pulles A, Paredes W, Gardner EL. Ventral
tegmental microinjection of delta-9-tetrahydrocannabinol enhances ventral tegmental somatodendritic dopamine levels but not fore-
brain dopamine levels: evidence for local neural action by mari-juana's psychoactive ingredient. Brain Res 1993; 621: 65-70.
[268] Castaneda E, Moss DE, Oddie SD, Whishaw IQ. THC does not affect striatal dopamine release: microdialysis in freely moving
rats. Pharmacol Biochem Behav 1991; 40: 587-91 [269] Tanda G, Pontieri FE, Di Chiara G. Cannabinoid and heroin activa-
tion of mesolimbic dopamine transmission by a common mu1 opioid receptor mechanism. Science 1997; 276: 2048-50.
[270] Malone DT, Taylor DA. Modulation by fluoxetine of striatal do-pamine release following Delta9-tetrahydrocannabinol: a mi-
crodialysis study in conscious rats. Br J Pharmacol 1999; 128: 21-6.
[271] Pistis M, Ferraro L, Pira L, et al. Delta(9)-tetrahydrocannabinol decreases extracellular GABA and increases extracellular gluta-
mate and dopamine levels in the rat prefrontal cortex: an in vivo microdialysis study. Brain Res 2002; 948:155-8.
[272] Gardner EL. Addictive potential of cannabinoids: the underlying neurobiology. Chem Phys Lipids 2002; 121: 267-90.
[273] Tanda G, Loddo P, Di Chiara G. Dependence of mesolimbic do-pamine transmission on delta9-tetrahydrocannabinol. Eur J Phar-
macol 1999; 376: 23-6. [274] Solinas M, Justinova Z, Goldberg SR, Tanda G. Anandamide ad-
ministration alone and after inhibition of fatty acid amide hydrolase (FAAH) increases dopamine levels in the nucleus accumbens shell
in rats. J Neurochem 2006; 98: 408-19. [275] Murillo-Rodriguez E, Vazquez E, Millan-Aldaco D, Palomero-
Rivero M, Drucker-Colin R. Effects of the fatty acid amide hydro-lase inhibitor URB597 on the sleep-wake cycle, c-Fos expression
and dopamine levels of the rat. Eur J Pharmacol 2007; 562: 82-91. [276] Xi ZX, Spiller K, Pak AC, et al. Cannabinoid CB1 receptor an-
tagonists attenuate cocaine’s rewarding effects: experiments with self-administration and brain stimulation reward in rats. Neuropsy-
chopharmacology 2008; 33(7): 1735-45. [277] Drews E, Schneider M, Koch M. Effects of the cannabinoid recep-
tor agonist WIN 55,212-2 on operant behavior and locomotor activ-ity in rats. Pharmacol Biochem Behav 2005; 80: 145-150.
[278] Mendizabal V, Zimmer A, Maldonado R. Involvement of kappa/dynorphin system in WIN 55,212-2 self-administration in
mice. Neuropsychopharmachology 2006; 31: 1957-66. [279] J rbe TU, Lamb RJ, Liu Q, Makriyannis A. Discriminative stimu-
lus functions of AM-1346, a CB1R selective anandamide analog in rats trained with Delta-9-THC or (R)-Methanadamide (AM356).
Psychopharmacology 2006 ; 188 : 315-23. [280] McMahon LR. Discriminative stimulus effects of the cannabinoid
CB1 antagonist SR 141716A in rhesus monkeys pretreated with Delta9-tetrahydrocannabinol. Psychopharmacology 2006; 188:
306-14. [281] Rubino T, Vigano D, Massi P, Parolaro D. Changes in the cannabi-
noid receptor binding, G protein coupling and cyclic cAMP cas-cade in the CNS of rats tolerant to and dependent on the synthetic
cannabinoid compound CP55,940. J Neurochem 2000; 75: 2080-6.
Received: October 22, 2007 Revised: January 25, 2008 Accepted: February 20, 2008


















