Behavioral ecology of young-of-the-year kelp rockfish ...directory.umm.ac.id/Data...
18
Journal of Experimental Marine Biology and Ecology 256 (2001) 33–50 www.elsevier.nl / locate / jembe Behavioral ecology of young-of-the-year kelp rockfish, Sebastes atrovirens Jordan and Gilbert (Pisces: Scorpaenidae) * Peter A. Nelson Institute of Marine Sciences, University of California, Santa Cruz, CA 95064, USA Received 2 June 2000; received in revised form 14 August 2000; accepted 25 September 2000 Abstract Young-of-the-year (YOY) kelp rockfish, Sebastes atrovirens, recruit initially to the canopy of the giant kelp, Macrocystis pyrifera (L.), along the coast of central California. During the summer of 1991, I observed major fluctuations in YOY kelp rockfish abundance in the canopy before the onset of a vertical migration downward to the Macrocystis holdfasts. Within the Macrocystis canopy, YOY kelp rockfish make a series of microhabitat shifts preparatory to their downward migration, forming smaller and more closely held aggregations. Experimental manipulations of Macrocystis architecture indicate that YOY kelp rockfish densities are reduced in areas with only small Macrocystis (few stipes) compared to unmanipulated areas with a range of Macrocystis sizes. Unusually large Macrocystis, created by binding multiple kelps together, did not compensate for a preponderance of small Macrocystis in the surrounding areas. In unmanipulated habitat, YOY kelp rockfish densities were correlated with the size of the Macrocystis (number of stipes). Habitat selection in the canopy and holdfasts appears to be behavioral and is closely tied to the architecture of the alga. The onset of the downward migration appears to be triggered by a combination of ontogenetic and environmental cues. 2001 Elsevier Science B.V. All rights reserved. Keywords: Macrocystis; Microhabitat selection; Recruitment; Rockfish; Sebastes 1. Introduction Recent work on tropical and temperate reef fishes continues to debate the relative importance of recruitment versus post-recruitment processes in structuring these reef fish *Correspondence address: Hawaii Institute of Marine Biology, University of Hawaii at Manoa, Coconut Island, Kaneohe, HI 96744-1346, USA. Tel.: 11-808-236-7428; fax: 11-808-236-7443. E-mail address: [email protected] (P.A. Nelson). 0022-0981 / 01 / $ – see front matter 2001 Elsevier Science B.V. All rights reserved. PII: S0022-0981(00)00305-1
Transcript of Behavioral ecology of young-of-the-year kelp rockfish ...directory.umm.ac.id/Data...

Journal of Experimental Marine Biology and Ecology256 (2001) 33–50
www.elsevier.nl / locate / jembe
Behavioral ecology of young-of-the-year kelp rockfish,Sebastes atrovirens Jordan and Gilbert (Pisces: Scorpaenidae)
*Peter A. NelsonInstitute of Marine Sciences, University of California, Santa Cruz, CA 95064, USA
Received 2 June 2000; received in revised form 14 August 2000; accepted 25 September 2000
Abstract
Young-of-the-year (YOY) kelp rockfish, Sebastes atrovirens, recruit initially to the canopy ofthe giant kelp, Macrocystis pyrifera (L.), along the coast of central California. During the summerof 1991, I observed major fluctuations in YOY kelp rockfish abundance in the canopy before theonset of a vertical migration downward to the Macrocystis holdfasts. Within the Macrocystiscanopy, YOY kelp rockfish make a series of microhabitat shifts preparatory to their downwardmigration, forming smaller and more closely held aggregations. Experimental manipulations ofMacrocystis architecture indicate that YOY kelp rockfish densities are reduced in areas with onlysmall Macrocystis (few stipes) compared to unmanipulated areas with a range of Macrocystissizes. Unusually large Macrocystis, created by binding multiple kelps together, did not compensatefor a preponderance of small Macrocystis in the surrounding areas. In unmanipulated habitat, YOYkelp rockfish densities were correlated with the size of the Macrocystis (number of stipes). Habitatselection in the canopy and holdfasts appears to be behavioral and is closely tied to thearchitecture of the alga. The onset of the downward migration appears to be triggered by acombination of ontogenetic and environmental cues. 2001 Elsevier Science B.V. All rightsreserved.
Keywords: Macrocystis; Microhabitat selection; Recruitment; Rockfish; Sebastes
1. Introduction
Recent work on tropical and temperate reef fishes continues to debate the relativeimportance of recruitment versus post-recruitment processes in structuring these reef fish
*Correspondence address: Hawaii Institute of Marine Biology, University of Hawaii at Manoa, Coconut Island,Kaneohe, HI 96744-1346, USA. Tel.: 11-808-236-7428; fax: 11-808-236-7443.
E-mail address: [email protected] (P.A. Nelson).
0022-0981/01/$ – see front matter 2001 Elsevier Science B.V. All rights reserved.PI I : S0022-0981( 00 )00305-1

34 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
populations (Sale, 1990; Levin, 1994; Eggleston, 1995; Caley et al., 1996; Ault andJohnson, 1998a,b). The debate has emphasized the differences in the ecologies of aprerecruitment, planktonic, larval stage as opposed to those of a postrecruitment,substratum-associated juvenile or adult stage, each stage influenced by a different set ofvariables. Recruitment of a cohort of temperate reef fishes may be subject to anadditional level of complexity where species rely on macroalgae as habitat (Carr, 1989,1994a,b; Holbrook et al., 1990; Anderson, 1994).
Given the vagaries of predation, algal variability, and availability of food, postrecruit-ment processes may obscure variation in the recruitment of some fish species ontemperate reefs (Love et al., 1990). The mechanisms by which recruits select habitat andhow variability of habitat may affect the subsequent distribution of juveniles areunknown. In order to understand the population biology of fishes for which macroalgaeplays an important ecological role, it is clear that we must know something about howthose fishes use the resource. This paper describes the pattern of recruitment for aspecies of rockfish, Sebastes atrovirens, which first appears in near-shore areas in thecanopy of the kelp beds which form large, continuous areas of habitat in the easterntemperate Pacific Ocean.
Spatial and temporal variation in the abundance and distribution of macroalgae havebeen shown to have a strong effect on the recruitment of many temperate reef fishes (e.g.Bodkin, 1988; Carr, 1989, 1991; Holbrook et al., 1990; Levin, 1991, 1993; Anderson,1994). Algal density, architecture and distribution are subject to processes independentof recruiting fish, and the presence of suitable settlement habitat may potentially be
˜strongly affected by El Nino events (Holbrook et al., 1990) and, in some instances, thecommercial harvest of macroalgae (e.g. Miller and Geibel, 1973; Feder et al., 1974;McCleneghan and Houk, 1985; Bodkin, 1988). Clearly, the presence or absence ofmacroalgae, or its sudden disappearance, affects the ecology of algal-associated fishes,but we do not have a good understanding of how variation in the structure ofmacroalgae, including canopy density and health, the size of individual algae (demo-graphics) or algal architecture may affect fish ecology.
Here, I provide results from observational and empirical studies of the relationshipbetween the detailed structure of the giant kelp, Macrocystis pyrifera C.Ag. andyoung-of-the-year (YOY) kelp rockfish, Sebastes atrovirens Jordan and Gilbert. I reporton the recruitment of YOY kelp rockfish to the Macrocystis canopy and fluctuations infish densities in the canopy over the 1991 season (July–October). I describe changes inthe behavior and distribution of YOY aggregations within parts of the algal canopy overtime, and test the hypothesis that sea surface temperature is correlated with YOY S.atrovirens density in the canopy. After the fish had settled to the kelp holdfasts, Imanipulated the number of Macrocystis stipes to determine if the availability ofdifferent-sized Macrocystis affected YOY density at the holdfasts. I tested the hypothesisthat the number of YOY kelp rockfish is positively correlated with the size of the kelpwith which the fish are associated. Finally, I provide evidence for the ontogenetic controlof the vertical migration by YOY kelp rockfish, from the Macrocystis canopy down tothe holdfasts.
Rockfishes (Sebastes spp., family Scorpaenidae) are an important component ofcommercial fisheries (Lenarz, 1986; CDFG, 1998; Love et al., 1998) in the temperate

P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50 35
Pacific. Several near-shore stocks, including species which utilize the kelp beds asjuvenile fishes, along the California coast are in severe decline (Love et al., 1998). Thepossible establishment of no-take marine reserves is thought to be particularly relevantto Sebastes spp. as these fishes are long-lived and slow to reach reproductive maturity,adults show high site fidelity and recruitment into the fishery is variable (Love andWestphal, 1981; Wyllie Echeverria, 1987; Leaman, 1991; Love and Johnson, 1998). Ifmarine reserves are to be successful in providing not only a local refuge but also abenefit to populations (and fisheries) outside of the reserve, dispersal into other areas willbe essential. Pelagic larval and juvenile stages might be expected to provide suchdispersal capabilities in these sedentary fishes, but the efficacy of such a plan is difficultto establish in advance. Additional information on the ecology of juvenile fishes isneeded to anticipate the habitat requirements for juveniles recruiting outside of theirnatal areas and the potential for postsettlement dispersal.
This study provides information on behavior, movement, growth, distribution andtiming of the vertical migration of YOY kelp rockfish. In particular, I provide evidencethat the architectural characteristics of the macroalga into which these juveniles recruitare important in determining the density of juveniles which make the vertical migrationto the reef. Observational data suggest that algal-associated juveniles are unlikely todisperse far from the spot in the algal canopy where they first recruit. My observationsmay provide predictive tools for understanding variability in year-to-year recruitmentstrength in at least one important fish in kelp beds on the central Californian coast.
To understand reef fish population structure, we must have basic information aboutthe processes likely to affect fluctuations in cohort strength. Models of populationecology of necessity divide life history into convenient stages — larval period,settlement, reef-associated, etc. — but what happens within those stages? Whatprocesses account for a strong showing for a year class in the larval stages but a weakone once the cohort is reef-associated? Particularly when postrecruitment ecologies areas multifaceted as in some of the Sebastes spp., we have to understand what ishappening within those discrete stages.
2. Materials and methods
2.1. Numbers of fish in the canopy
YOY S. atrovirens recruit to the surface canopy of Macrocystis in mid to late summer(Love et al., 1990; Carr, 1991; Nelson, 1992). They occupy a sequence of microhabitatswithin the kelp before migrating to holdfasts near the reef bottom (Nelson, 1992). YOYkelp rockfish occupy these holdfasts for several weeks before making an additionalhabitat shift to rocky interstices of the reef (Nelson, 1992). For this reason, postrecruit-ment behavior and algal conditions potentially affect numbers of fish.
In the kelp beds near Otter Point, along the southern shore of Monterey Bay,California (Fig. 1), I swam five 20 m transects distinguished by subsurface buoyintervals and by timed swim, at 3–4-day intervals between 12 July, 1991 and 7 October,1991 (n 5 23). These transects were contiguous, end-to-end, such that considerable

36 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
Fig. 1. Study and collection sites.
movement by these juvenile fish in the canopy in response to the presence of a divercould have compromised the independence of these transects. However, these fishresponded to diver presence by seeking shelter in the kelp, and exhibited little tendencyto disperse away from the transects. Hoelzer (1988) has found that a congener (S.carnatus) with a similar natural history exhibits limited lateral movement within thecanopy environment, with 63.5% of his tagged fish resighted within 2 m (range: 0–10 m)of the Macrocystis from which the fish had been captured. Hoelzer (1988) found nocorrelation between the time since tagging and the distance from capture site.
I recorded the number of aggregations, the number and species of YOY Sebastes spp.,and the microhabitat(s) occupied by the fish within a path approximately 2 m wide 3 2 mdeep (standardized by measured diver limb 1 swim fin lengths; total volume of each
3transect 5 80 m ) immediately after swimming each transect. I defined an aggregation astwo or more YOY Sebastes spp. within 1 m of each other which moved together andoccupied the same microhabitat (defined below) during my observations. From these
3data, I calculated the mean number of YOY kelp rockfish/m . I used linear regression totest the hypothesis that sea surface temperature was correlated with fish density.

P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50 37
Observations were aborted when the horizontal visibility was , 4 m or when largeswells prevented accurate assessment of aggregation size.
The kelp beds of the central California coast consist in part of multiple Macrocystispyrifera, each anchored to the substratum by a holdfast (Feder et al., 1974; Dayton,1985). Multiple stipes, buoyed by the pneumatocysts of the leaf-like blades, rise towardsthe water’s surface. At the surface, the stipes bend, creating a canopy of stipes andblades. I defined the microhabitats occupied by the fishes as follows (Fig. 2): gladeswere the areas where the canopy was thinnest and light penetrated readily, usuallyfarthest from where the stipes rising from the holdfasts met the surface. Fish in theseglades were within the top 30 cm of water, among or just below the stipes and blades ofthe canopy. Where the vertical stipes met the canopy and bent over I termed an elbow.This microhabitat was heavily shaded due to the density and abundance of stipes andblades. Fish associated with the elbows were inside the curve and on the downcurrent
Fig. 2. Macrocystis pyrifera microhabitat sites occupied by YOY kelp rockfish.

38 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
side, from 30 to 100 cm below the surface. Fish among or next to the vertical stipes,1–1.5 m below the surface (to distinguish this microhabitat from the elbows) occupiedthe stipes. A 1-way analysis of variance for unequal variances (Rice and Gaines, 1989)was used to test the hypothesis that mean group size varied among canopy microhabitats.
I recorded the sea surface temperature (SST) at the start of each sample date. I usedlinear regression to test the hypothesis that SST was correlated with fish density.
2.2. Experimental manipulations
Casual observations during the prior season (summer, 1990) suggested that postsettle-ment YOY Sebastes atrovirens (i.e. at the kelp holdfasts) were more numerous at thelarger kelps (with a greater number of stipes). I tested the following hypothesesregarding the distribution of YOY S. atrovirens at the kelp holdfasts. (1) The availabilityof different sized Macrocystis (number of stipes) will affect the mean density of YOY S.atrovirens in an area. Further, I predicted that, where all available Macrocystis are small,YOY kelp rockfish density will be reduced, compared to unmanipulated areas orlocations where ‘superkelps’ are available. (2) The number of YOY kelp rockfish ispositively correlated with the size of the kelp with which the fish are associated. (3)YOY kelp rockfish are actively selecting larger kelp (entire bundles of stipes), ratherthan (a) following individual stipes from the canopy to the holdfasts or (b) selectingMacrocystis at random. To test these hypotheses, I manipulated Macrocystis characteris-tics in the kelp beds off Otter Point (Fig. 1) as described below, and then documenteddistribution patterns of postsettlement fish at the holdfasts. I use the term ‘settlement’here to refer to the ontogenetic vertical migration by juvenile Sebastes spp. fromMacrocystis canopy to Macrocystis holdfast or reef bottom.
I laid out six 20 3 20 m plots (A–F), 10–12 m inshore from the outer edge of the kelpbed, at depths of 10–14 m, and separated from any other plot by at least 10 m. Theseplots were delimited with nylon cord on the substratum (Fig. 3), and plot locations werechosen deliberately rather than randomly to minimize any apparent differences amongthem such as depth, distance from the edge of the kelp bed, reef substrate or Macrocystischaracteristics. The substratum was granitic rock and boulders interspersed with patchesof coarse sand; surfaces were covered by coralline algae and sessile invertebrates. Otherunderstory algae were sparse, although Cystoseira osmundacea was present.
I used a nested experimental design, with two replicates for each of three treatments(described below, six plots in total). I assigned treatments to these plots in such a way asto maximize the amount of unmanipulated habitat between treatment plots to ensurethese treatment plots were independent (Fig. 3). A larger number of plots with thetreatments assigned at random would have been preferred with regard to experimentaldesign, but larger plots, located in a comparable habitat, seemed the best compromisebetween replication and feasibility.
Three treatments were applied to the plots (Fig. 3): Plots B and E (Control plots) wereunmanipulated controls. In Plots A and D (Culled plots), every Macrocystis wastrimmed so that no single alga had more than 15 stipes; those with 15 or fewer stipeswere left undisturbed. Cut stipes were pulled away from the holdfast, and allowed tofloat to the surface. They were pulled into open water or left for the currents and swells

P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50 39
Fig. 3. Schematic of plot treatments and layout. Different sized dots represent different sized Macrocystispyrifera.
to pull from the canopy; no cut stipes remained in the canopy of the experimental plotsby 6 days after the initial culling. Macrocystis within a 5-m wide strip surrounding plotsA and D were similarly culled to minimize possible edge effects. In Plots C and F(SuperKelp plots), at four places approximately equidistant from each other and fromplot borders, 4–5 Macrocystis (number of stipes 5 15 each) within approximately 2 m ofeach other were located. The stipes of these algae were then bound together 1.0–1.2 mfrom the bottom. The result was a thick column of stipes ( . 60 stipes) rising to thecanopy, a ‘superkelp.’ The remaining Macrocystis in the SuperKelp plots were culled as

40 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
in A and D. A 5 m-wide buffer zone around C and F was also culled to minimizepossible edge effects.
The plots were sampled beginning on 13 September, 1991 and concluding on 17October, 1991 (eleven sample dates). Sampling from September 13 to September 30 (sixdates) documented the arrival of the first fish to complete the downward verticalmigration from canopy to holdfast (20 September); samples from October 1 to October17 (five dates) were used for the analyses. By October 1, there were virtually no YOYkelp rockfish in the canopy; the last three canopy transects (September 29th, October 3rd
3and 7th) yielded densities of 0.1360.13, 0.0460.11 and 0.0 fish/m (mean61 SEM).Nearly all surviving YOY had settled out at the holdfasts and fish density calculations atthe bottom should have been minimally affected by late migrators. Large swellsdamaged transect markers in the first three plots (A, B and C), and no data wereobtained from these plots after October 7th.
I and a diving assistant counted the number of YOY kelp rockfish associated withMacrocystis holdfasts along transects in each of the plots (Fig. 3). On each sample date,a plot was sampled by swimming one or more transects chosen randomly from one offive fixed positions along the east side of each plot. The transect was swum, 1 m abovethe bottom, along a bearing perpendicular to the eastern plot boundary until the otherside was reached or ten M. pyrifera had been sampled, whichever came first. Thesampler stopped 2.5 m from each M. pyrifera encountered within a corridor, 1 m inwidth, and recorded the number of fish within the stipes or within 1 m of the holdfast(Fig. 2). Then, the diver counted the number of stipes, and noted the presence or absenceof drift algae entwined about the Macrocystis. Although some transects were swum morethan once over the course of the study, I used data only from the last visit to a particulartransect to assure independence; transects sampled only once during the study wereincluded in the analyses as well. YOY associated with drift algae or associated with anattached Macrocystis entangled with drift algae were recorded separately from thosewith unentangled, attached Macrocystis.
To test for a treatment effect, a nested ANOVA was used. The variability inMacrocystis density among plots and the loss of three plots (A–C) due to large swellslate in the study resulted in unequal sample sizes among cells (range: 34–41 data).Because the design was unbalanced (unequal numbers of Macrocystis sampled acrossplots), I took a random sample from each data set (n 5 34 for each cell). These data were
2not normally distributed (D’Agostino-Pearson K test, P,0.05), and were transformed1 / 2 1 / 2[X 1 (X 1 1) ]. I compared the number of fish observed at Macrocystis holdfasts in
the control plots (B, E) across a range of Macrocystis sizes to test the hypothesis that thenumber of postsettlement YOY kelp rockfish at a holdfast is related to Macrocystis size(by number of stipes) using linear regression.
While YOY kelp rockfish may be exhibiting a behavioral preference for largerMacrocystis, a positive relationship between the number of kelp rockfish and Mac-rocystis size might also be explained by one of two simple presettlement encountereffects: (a) the YOY are following individual stipes down to the holdfasts and fishdistribution is actually a function of the distribution of stipes among different sizedMacrocystis. (b) The YOY are following entire bundles of stipes without regard to thenumber of stipes in those bundles, and the frequency of different sized Macrocystis

P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50 41
determines the distribution of postsettlement kelp rockfish. To test the first hypothesis, Icompared the observed distribution of postsettlement fish in the unmanipulated plots (B,E) across Macrocystis size classes (number of stipes51–5, 6–10, 11–15, 16–20, 21–25,26–30, 31–35, 36–40 and .40) to a random (Poisson) distribution based on thedistribution of all stipes across the same range of Macrocystis size classes. Similarly, Itested the second hypothesis by comparing the observed fish distribution across the arrayof Macrocystis sizes to a random distribution based on the availability of differentMacrocystis sizes. In each instance, I used a G test to compare the two frequencydistributions (Sokal and Rohlf, 1995).
2.3. Life history
YOY kelp rockfish have been reported to move from the canopy to the holdfast atabout 45 mm TL (total length, Anderson, 1983; Carr, 1983). To test the hypothesis thatYOY kelp rockfish do not leave the kelp canopy for the Macrocystis holdfasts beforereaching a critical size, I collected YOY kelp rockfish from both the canopy and theholdfast habitats at a single geographic location and on a single date. I collected from asingle locality and date so that size differences could be attributable to developmentaldifferences between the YOY kelp rockfish found in the two habitats. These collectionswere made on 6 October, 1991 at Big Creek (Fig. 1). I predicted a priori that the meanTL in the canopy would be less than the mean TL at the holdfasts, and compared themeans with a 1-tailed t-test for unequal variances.
3. Results
3.1. Numbers of fish in the canopy
YOY kelp rockfish of the 1991 year class were first observed in the kelp canopy off3Otter Point on July 9th in low numbers (two individuals in 160 m ) during canopy
surveys for the California Department of Fish and Game (CDFG, unpublished data).3YOY kelp rockfish first appeared on my transects (one individual in 240 m of transects;
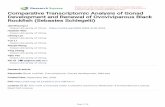
3mean61 SEM50.00860.008 fish/m ) on July 17, reached a maximum mean density of3 32.460.7 fish/m July 26th, and dropped to 0.0760.00 fish/m August 3rd before
3reaching a second, lower peak (1.360.5 fish/m ) on August 16th (Fig. 4). I observedkelp rockfish on the transects from July 17th to October 3rd. No fish were found on thetransects on October 7th, 1991, and casual observations at Otter Point after this daterevealed no kelp rockfish in the canopy (Fig. 4). Sea surface temperature did not explain
2the fluctuations in YOY kelp rockfish density in the canopy (R 50.07; ANOVA:F 51.76, P50.20; Fig. 4).(1, 22)
Between July 8th and September 3rd, YOY kelp rockfish appeared in three distinctmicrohabitats, the glades, the elbows and the upper stipes of the canopy (Fig. 2) beforetheir downward vertical migration to the holdfasts. The fraction of all fish observed onthe canopy transects associated with each of these microhabitats changed over the courseof the season (Fig. 5). One hundred percent of all fish observed through August 12th
42 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
Fig. 4. Seasonal fluctuations in YOY Sebastes atrovirens density in the canopy and sea surface temperature.
were found in the glades. Data collected through the latter half of August and intoSeptember show an increasing proportion of the fish occupying the elbows. By the latterhalf of September, most of the fish were found in the stipes (Fig. 5).
YOY kelp rockfish in each of these microhabitats differed significantly with regard toboth the mean group size and their behavior. In the glades, YOY kelp rockfish formedlarge (mean61 SEM531.562.6 fish), loose and unpolarized aggregations (at a givenmoment in time, most fish were not oriented in the same direction, sensu Schilt and
Fig. 5. Habitat shifts in the Macrocystis pyrifera canopy; YOY kelp rockfish in the canopy move from gladesto elbows to stipes over the course of the season.
P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50 43
Norris, 1997) just below the stipes and fronds of the Macrocystis canopy, often withYOY Sebastes chrysomelas /S. carnatus. (S. chrysomelas and S. carnatus could not bedistinguished from each other in the field at this age.) Within the elbows, aggregations(15.361.6 fish) were composed entirely of YOY kelp rockfish, and individuals werecloser together than in aggregations observed in the glades. In the upper stipes, groupswere small (4.260.2 fish), and individuals were typically observed amongst the stipesand fronds. It is likely that a greater proportion of fish in this microhabitat wereoverlooked than in the glades or elbows. The mean group size (including individual fish)differed significantly among canopy microhabitats (ANOVA for unequal variancesF 51318, df52, 197, P,0.0001).U
3.2. Experimental manipulations
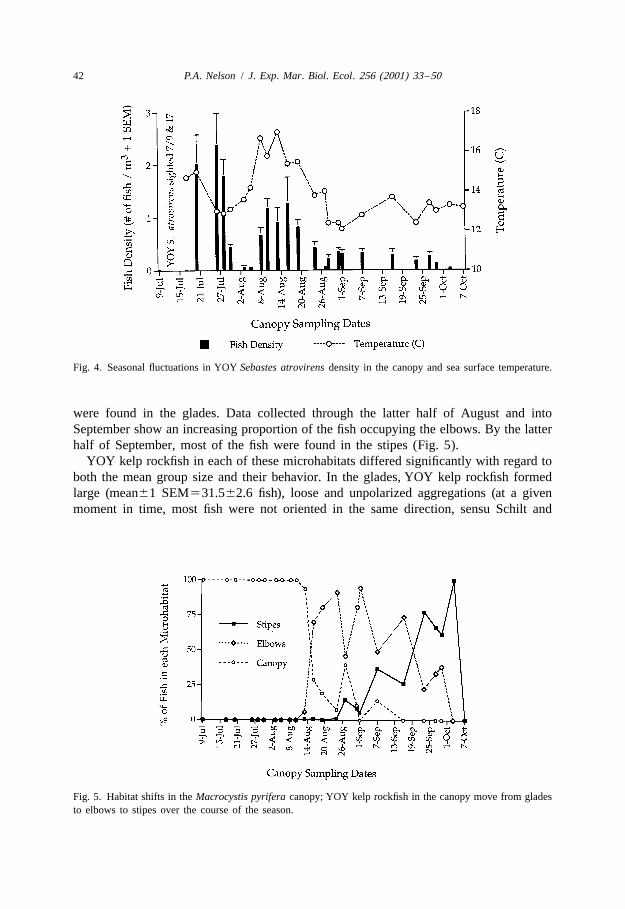
Results from the nested ANOVA indicate that there was a significant treatment effect(Fig. 6, F 510.58, P50.04), and that differences among plots did not contribute(2,198)
significantly to the variance (F 50.61, P50.61). A Student–Newman–Keuls(3,198)
(SNK) multiple comparison test (Underwood, 1997) was used to determine whichtreatment means were different. The Control treatment differed significantly from boththe Culled and the SuperKelp treatments (P,0.05), but the Culled did not differsignificantly from the SuperKelp treatment (P.0.05).
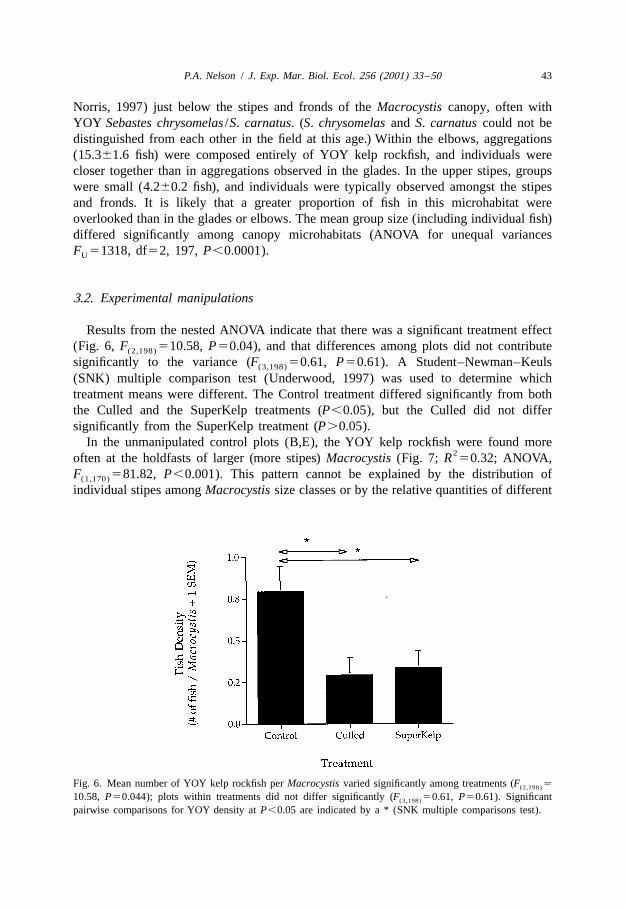
In the unmanipulated control plots (B,E), the YOY kelp rockfish were found more2often at the holdfasts of larger (more stipes) Macrocystis (Fig. 7; R 50.32; ANOVA,
F 581.82, P,0.001). This pattern cannot be explained by the distribution of(1,170)
individual stipes among Macrocystis size classes or by the relative quantities of different
Fig. 6. Mean number of YOY kelp rockfish per Macrocystis varied significantly among treatments (F 5(2,198)
10.58, P50.044); plots within treatments did not differ significantly (F 50.61, P50.61). Significant(3,198)
pairwise comparisons for YOY density at P,0.05 are indicated by a * (SNK multiple comparisons test).
44 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
Fig. 7. Number of YOY Sebastes atrovirens at the holdfasts increased with the size of the Macrocystispyrifera in Control plots (B, E). Line forced through the origin; b ± 0, ANOVA, F 586.25, P,0.001.(1,76)
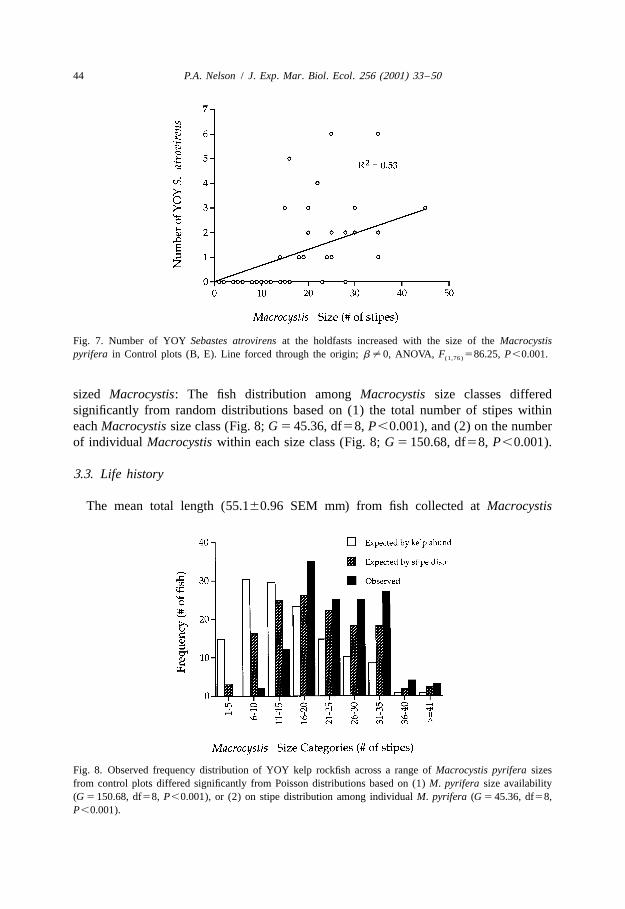
sized Macrocystis: The fish distribution among Macrocystis size classes differedsignificantly from random distributions based on (1) the total number of stipes withineach Macrocystis size class (Fig. 8; G 5 45.36, df58, P,0.001), and (2) on the numberof individual Macrocystis within each size class (Fig. 8; G 5 150.68, df58, P,0.001).
3.3. Life history
The mean total length (55.160.96 SEM mm) from fish collected at Macrocystis
Fig. 8. Observed frequency distribution of YOY kelp rockfish across a range of Macrocystis pyrifera sizesfrom control plots differed significantly from Poisson distributions based on (1) M. pyrifera size availability(G 5 150.68, df58, P,0.001), or (2) on stipe distribution among individual M. pyrifera (G 5 45.36, df58,P,0.001).

P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50 45
pyrifera holdfasts was significantly greater than the mean total length from fish collectedin the canopy (49.660.76 SEM mm) at Big Creek (t 5 4.55, df546, P (1-tailed),
0.001).
4. Discussion
During the time YOY kelp rockfish were present in the kelp canopy, three significantevents were observed: (1) the nearly complete disappearance and subsequent reappear-ance of large number of these juvenile fish in the canopy over a period of 3 weeks (2)the sequential use of three different microhabitats in the canopy, prior to the fish’smovement to the bottom and (3) sequential changes in group size and group structure.
The abrupt, though temporary, disappearance of YOY Sebastes atrovirens from thecanopy appeared to be at least partially attributable to predation by YOY S. paucispinis. Iobserved YOY S. paucispinis pursuing YOY kelp rockfish in the canopy, and increasingnumbers of these fast-growing, piscivorous (Love et al., 1990) rockfish presaged thecrash in YOY kelp rockfish near the end of July (CDFG, unpublished data; Nelson,1992). The disappearance of the YOY S. paucispinis from the canopy coincided with therecruitment of additional YOY kelp rockfish to the canopy.
Unpublished data of the California Department of Fish and Game and my observa-tions from the 1990 and 1991 seasons suggest that kelp rockfish may recruit tonear-shore habitats over a broader time span than many of the other substrate-associatedjuvenile Sebastes species in this area. The timing of recruitment is, in part, related to thetiming of parturition (Love et al., 1990), and adult kelp rockfish may reproduce over abroader time frame than some of its congeners (VenTresca and Lea, unpublished data).While a broad period of parturition by itself would not account for a broad period ofrecruitment (i.e. hydrographic factors are important too), it may mean that there arelarval and juvenile kelp rockfish in the plankton over a long time period during whichrecruits may appear in the kelp canopy.
The shifts between canopy microhabitats described in this study and the accompany-ing changes in aggregation structure appear to have two functions: they should reducethe threat of predation, positioning the YOY in greater proximity to dense cover, and thetight aggregations may confer some of the protective functions attributed to a schoolingformation (Hobson, 1978; Pitcher and Parrish, 1986; Norris and Schilt, 1988). Inaddition, these microhabitat shifts have the potential to enable the YOY kelp rockfish toselect a large Macrocystis at the surface, and remain associated with that alga over thecourse of the fish’s move to the bottom, occupying the holdfast of the same alga whosestipes and elbow it occupied at the surface.
The migration from the surface canopy to the holdfasts appears to be triggered byboth endogenous and environmental cues. Love et al. (1990) describe this movement asontogenetic, citing the observation of Carr, (1983, 1991) that the timing of themovement coincides with increased size. The results presented here from the life historydata on the role of size in the timing of this migration support the suggestion thatontogenetic factors are important. However, it would seem that there should be strongselective pressure to remain in the canopy until a critical size is reached. The larger a

46 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
juvenile fish grows, the greater its top swimming speed (Weihs, 1977), and the fewerpiscivores will possess a gape large enough to eat it. Greater experience in predatoravoidance may also be involved. Before its destruction by storms, the canopy shouldoffer a far greater expanse of cover and substrate for prey than the holdfasts. For thisreason, I suggest that environmental cues are likely to have an important influence on thetiming of this vertical migration. The timing of the downward migration coincidesroughly with the onset of winter storms, raising the possibility that wave action or watermotion may initiate this behavior. Love et al. (1990) have suggested that ‘‘ . . . turbulentconditions may initiate the migration (of juvenile S. entomelas, S. flavidus, S. jordani, S.melanops, S. miniatus, S. ovalis, S. paucispinis and S. pinniger) toward adult depths.’’ Amore likely cue for kelp rockfish, however, is the loss of the kelp canopy which mayresult from high wave action and the deterioration of the algae due to warm water andlow nutrient availability in late summer (Dayton, 1985) inside of Monterey Bay. At BigCreek and at Cypress Point, south and outside of Monterey Bay (Fig. 1), YOY kelprockfish were observed in the canopy in large numbers on October 6th and October 20th,respectively. Within Monterey Bay, at Otter Point, the majority of the YOY kelp rockfishhad settled to the bottom by September 28th and none were found in the canopy onOctober 7th. The Big Creek and Cypress Point sites are much more exposed than is theOtter Point site, but both had large expanses of relatively healthy canopy well after muchof the canopy had been destroyed off Otter Point. YOY Sebastes spp. (including kelprockfish) in the canopy do not appear to be able to survive its removal due tooverwhelming predation pressure in the absence of any refuge (Bodkin, 1988).Therefore, YOY kelp rockfish appear to make the migration first on the basis ofendogenous cues (ontogenetic), and secondarily in response to habitat loss late in theseason. Although there was no correlation between the abundance of YOY kelp rockfishin the canopy and the sea surface temperature, water temperature may be an importantparameter in the migration from surface to bottom. Love et al. (1990) point out that forgrowth, temperature-dependent assimilation efficiency may be a factor in the initialrecruitment to the relatively warm, surface waters prior to a vertical movement to colder,bottom water common in juvenile Sebastes spp.
Once at the holdfasts, YOY kelp rockfish remain associated with Macrocystis for thenext 2 months (Nelson, 1992), before moving into rocky crevices. During this 2-monthperiod, most fish observed at the bottom are associated with holdfasts of the largerMacrocystis. There are four alternative explanations for this non-random distribution:(1) behavioral selection, (2) differential recruitment, (3) differential mortality or (4)differential site fidelity. The pattern may be predetermined, while the fish are still in thecanopy, or the pattern may emerge after the settlement process.
I believe that this pattern is attributable to behavioral selection: the frequencydistributions of stipes and fish across Macrocystis size categories were significantlydifferent — while a simple encounter effect (more fish at the larger Macrocystis becausethe greatest proportion of the stipes in the canopy lead to the holdfasts of the largeralgae) should yield matching distributions (Fig. 8). Thus, fish distribution cannot beexplained by a simple encounter effect, whereby YOY follow individual stipes from thecanopy to the holdfasts, or select Macrocystis without regard to the number of stipes. Inaddition, the YOY kelp rockfish, both in the canopy and at the holdfasts, appeared to be

P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50 47
associating with stipe bundles, rather than individual stipes. Throughout the series ofshifts in microhabitat in the canopy, aggregations of fish changed from relativelydispersed distributions to clumped distributions, occupying an area in close proximity tothe vertical bundles of stipes, rather than associating with a single kelp stipe.
Crowe and Underwood (1998) caution against ascribing an observed nonrandomdistribution among available microhabitats to behavioral selection (e.g. differentialmortality may also produce the observed distribution). Indeed, the density of fish /Macrocystis differed significantly among the three treatments, raising the possibilitiesthat either mortality was higher in the Culled plots or that YOY emigrated from thesetreatment plots in higher numbers than from other plots. However, pretreatment canopyconditions were uniform throughout the experimental area, so I expect that recruitmentinto the Macrocystis canopy was comparable in all plots. Although a considerablesurface canopy remained after the completion of both the culled and the superkelptreatments, it is possible that the process of removing considerable quantities of algalmaterial ‘dislodged’ some proportion of the YOY rockfish in the canopy at the time.
Although comparatively large numbers of YOY S. atrovirens were observed at theSuperKelps (mean50.9460.28 S.E. fish/SuperKelp, max.512; at small Macrocystis,mean50.2760.08 S.E. fish/Macrocystis, max.53), this was not sufficient to compen-sate for the paucity of medium and large kelp (.10 stipes). The availability of theselarger Macrocystis appears to be crucial to maintaining some density of YOY rockfish atthe holdfasts. Events which may affect the age or size distribution of macroalgae, such
˜as kelp harvesting, severe storms or El Nino events could have a direct effect on thestrength of a subsequent YOY S. atrovirens year class. These results should beconsidered when managing kelp harvesting.
5. Conclusions
Observations and transects in the canopy indicate that YOY kelp rockfish exhibit aseries of microhabitat shifts within the Macrocystis canopy, prior to their verticalmigration to the holdfasts. Predation by YOY congeners, particularly S. paucispinis, mayhave the potential to affect the success of a year class of YOY kelp rockfish. The broadtime period over which kelp rockfish arrives in the Macrocystis canopy may beadvantageous in dealing with the variable nature of two unpredictable conditions, thepresence of piscivorous YOY Sebastes spp. and the loss of the Macrocystis canopy tothe first winter storms. It is conceivable that the initial recruitment to the canopy,followed by the migration down to the Macrocystis holdfasts may have evolved to dealwith an ontogenetic change in temperature-related assimilation efficiency and growthrates (Love et al., 1990). The greater abundance of benthos-associated piscivores overwater-column piscivores in the kelp forest may also be incentive to utilize the surfacecanopy as a nursery. Macrocystis pyrifera with more stipes are more often utilized thanare those with fewer stipes, and the loss or absence of the larger Macrocystis has asignificant effect on the overall abundance of the YOY. It seems likely that thispreference for the larger kelp is related to microhabitat selection in the canopy, andprovides for improved habitat at the bottom with greater shelter and prey substrate at the

48 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
larger holdfasts. The timing of the vertical migration appears to be cued first byendogenous cues and, secondarily, by the loss of the surface substrate. This shift inhabitat may be secondarily related to a change in prey availability, though some authorsbelieve that these ontogenetic movements are not mediated by prey availability (Love etal., 1990).
The behavior and ecology of young-of-the-year kelp rockfish is closely tied to thestructure of Macrocystis pyrifera. Postsettlement factors, including predation pressureand the population biology of Macrocystis, have strong effects on the local recruitment
˜success of a given year class. El Nino events and the commercial harvest of Macrocystisare important considerations in the management of this Sebastes species and inunderstanding the demographies of local populations.
Acknowledgements
I am grateful to M. Paddack and N. Crane, and to the California Department of Fishand Game — particularly D. VenTresca, J. Hardwick and J. Houk — for theirconsiderable assistance. Funding was provided by the Friends of the Long MarineLaboratory, Gaines Memorial Award, Institute of Marine Sciences (UCSC), Lerner-GrayFund (AMNH), and the Myers Oceanographic and Marine Biology Trust. I thank K.S.Norris, J. Estes, R. Larson, and L. Montgomery for reviewing early versions of thispaper and A.J. Underwood and an anonymous reviewer for their advice. This paper isdedicated to the late Kenneth S. Norris. [AU]
References
Anderson, T., 1983. Identification and development of nearshore juvenile rockfishes (genus Sebastes) in kelpforests off central California. California State University, M.A. Thesis.
Anderson, T.W., 1994. Role of macroalgal structure in the distribution and abundance of a temperate reef fish.Mar. Ecol. Progr. Ser. 113, 279–290.
Ault, T.R., Johnson, C.R., 1998a. Relationships between habitat and recruitment of three species of damselfish(Pomacentridae) at Heron Reef, Great Barrier Reef. J. Exp. Mar. Biol. Ecol. 223, 145–166.
Ault, T.R., Johnson, C.R., 1998b. Spatially and temporally predictable fish communities on coral reefs. Ecol.Monogr. 68 (1), 25–50.
Bodkin, J.L., 1988. Effects of kelp forest removal on associated fish assemblages in central California. J. Exp.Mar. Biol. Ecol. 117, 227–238.
Caley, M.J., Carr, M.H., Hixon, M.A., Hughes, T.P., Jones, G.P., Menge, B.A., 1996. Recruitment and the localdynamics of open marine populations. Ann. Rev. Ecol. Syst. 27, 477–500.
CDFG, 1998. California Department of Fish and Game, Review of some California fisheries for 1997.CalCOFI Rep 39, 9–18.
Carr, M.H., 1983. Spatial and temporal patterns of recruitment of young-of-the-year rockfishes (genusSebastes) into a central California kelp forest. California State University, M.A. Thesis.
Carr, M.H., 1989. Effects of macroalgal assemblages on the recruitment of temperate zone reef fishes. J. Exp.Biol. Ecol. 126, 59–76.
Carr, M.H., 1991. Habitat selection and recruitment of an assemblage of temperate zone reef fishes. J. Exp.Mar. Biol. Ecol. 146, 113–137.

P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50 49
Carr, M.H., 1994a. Effects of macroalgal dynamics on recruitment of a temperate reef fish. Ecology 75 (5),1320–1333.
Carr, M.H., 1994b. Predicting recruitment of temperate reef fishes in response to changes in macrophytedensity caused by disturbance. In: Stouder, D.J., Fresh, K.L., Feller, R.J. (Eds.). Theory and Application inFish Feeding Ecology, Vol. 18. University of South Carolina Press, Columbia, pp. 255–269.
Crowe, T.P., Underwood, A.J., 1998. Testing behavioural ‘preference’ for suitable microhabitat. J. Exp. Mar.Biol. Ecol. 225, 1–11.
Dayton, P.K., 1985. Ecology of kelp communities. Ann. Rev. Ecol. Syst. 16, 215–245.Eggleston, D.B., 1995. Recruitment in Nassau grouper Epinephelus striatus: postsettlement abundance,
microhabitat features, and ontogenetic habitat shifts. Mar. Ecol. Progr. Ser. 124, 9–22.Feder, H.M., Turner, C.H., Limbaugh, C., 1974. Observations on fishes associated with kelp beds in southern
California. The Resources Agency of California Department of Fish and Game, pp. 4–136.Hobson, E.S., 1978. Aggregating as a defense against predators in aquatic and terrestrial environments. In:
Reese, E.S., Lighter, F.J. (Eds.), Contrasts in Behavior. Wiley, New York, pp. 219–234.Hoelzer, G.A., 1988. Juvenile movement patterns in a territorial scorpaenid fish before and during settlement.
Mar. Ecol. Progr. Ser. 45, 193–195.Holbrook, S.J., Carr, M.H., Schmitt, R.J., Coyer, J.A., 1990. Effect of giant kelp on local abundance of reef
fishes: the importance of ontogenetic resource requirements. Bull. Mar. Sci. 47 (1), 104–114.Leaman, B.M., 1991. Reproductive styles and life history variables relative to exploitation and management of
Sebastes stocks. Environ. Biol. Fish. 30, 253–271.Lenarz, W.H., 1986. A history of California rockfish fisheries. In: Proceedings of the International Rockfish
Symposium. Vol. 87–82, Alaska Sea Grant Report, pp. 35–41.Levin, P.S., 1991. Effects of microhabitat on recruitment variation in a Gulf of Maine reef fish. Mar. Ecol.
Progr. Ser. 75, 183–189.Levin, P.S., 1993. Habitat structure, conspecific presence and spatial variation in the recruitment of a temperate
reef fish. Oecologia 94, 176–185.Levin, P.S., 1994. Fine-scale temporal variation in recruitment of a temperate demersal fish: the importance of
settlement versus post-settlement loss. Oecologia 97, 124–133.Love, M.S., Carr, M.H., Haldorson, L.J., 1990. The ecology of substrate-associated juveniles of the genus
Sebastes. Environ. Biol. Fish. 30 (1–2), 225–243.Love, M.S., Caselle, J.E., Buskirk, W.v., 1998. A severe decline in the commercial passenger fishing vessel
rockfish (Sebastes spp.) catch in the southern California Bight, 1980–1996. CalCOFI Rep. 39, 180–195.Love, M.S., Johnson, K., 1998. Aspects of the life histories of grass rockfish, Sebastes rastrelliger, and brown
rockfish, S. auriculatus, from southern California. Fish. Bull. 87, 100–109.Love, M.S., Westphal, W.V., 1981. Growth, reproduction, and food habits of olive rockfish, Sebastes
serranoides, off central California. Fish. Bull. 79 (3), 533–545.McCleneghan, K., Houk, J.L., 1985. The effects of canopy removal on holdfast growth in Macrocystis pyrifera
(Phaeophyta; Laminariales). Calif. Fish Game 71 (1), 21–27.Miller, D.J., Geibel, J.J., 1973. Summary of blue rockfish and lingcod life histories; a reef ecology study; and
giant kelp, Macrocystis pyrifera, experiments in Monterey Bay, California, California Department of Fishand Game.
Nelson, P.A., 1992. Kelp rockfish and Macrocystis: Behavioral ecology and habitat structure. University ofCalifornia, M.S. Thesis.
Norris, K.S., Schilt, C.R., 1988. Cooperative societies in three-dimensional space: on the origins ofaggregations, flocks, and schools, with special reference to dolphins and fish. Ethol. Sociobiol. 9, 149–179.
Pitcher, T.J., Parrish, J.K., 1986. Functions of shoaling behavior in teleosts. In: Pitcher, T.J. (Ed.), Behavior ofTeleost Fishes. Chapman and Hall, London, pp. 363–439.
Rice, W.R., Gaines, S.D., 1989. One-way analysis of variance with unequal variances. Proc. Nat. Acad. Sci. 86,8183–8184.
Sale, P.F., 1990. Recruitment of marine species: Is the bandwagon rolling in the right direction? Trends Ecol.Evol. 5 (1), 25–27.
Schilt, C.R., Norris, K.S., 1997. Perspectives on sensory integration systems: Problems, opportunities, andpredictions. In: Parrish, J.K., Hamner, W.M. (Eds.), Animal Groups in Three Dimensions. CambridgeUniversity Press, Cambridge, pp. 225–244.

50 P.A. Nelson / J. Exp. Mar. Biol. Ecol. 256 (2001) 33 –50
Sokal, R.R., Rohlf, F.J., 1995. Biometry. W.H. Freeman, New York, p. 887.Underwood, A.J., 1997. Experiments in Ecology. Cambridge University Press, Cambridge, p. 504.Weihs, D., 1977. Effects of size on sustained swimming speeds of aquatic organisms. In: Pedley, T.J. (Ed.),
Scale Effects in Animal Locomotion. Academic Press, New York, pp. 333–338.Wyllie Echeverria, T., 1987. Thirty-four species of California rockfishes: maturity and seasonality of
reproduction. Fish. Bull. 85 (2), 229–250.