
Basal resistance of barley to adapted and non-adapted ... · Basal resistance of barley to adapted...
138
Basal resistance of barley to adapted and non-adapted forms of Blumeria graminis Reza Aghnoum
-
Upload
vuongkhanh -
Category
Documents
-
view
229 -
download
3
Transcript of Basal resistance of barley to adapted and non-adapted ... · Basal resistance of barley to adapted...

Basal resistance of barley to adapted and
non-adapted forms of Blumeria graminis
Reza Aghnoum

II
Thesis committee
Thesis supervisors
Prof. Dr. Richard G.F. Visser
Professor of Plant Breeding
Wageningen University
Dr.ir. Rients E. Niks
Assistant professor, Laboratory of Plant Breeding
Wageningen University
Other members
Prof. Dr. R.F. Hoekstra, Wageningen University
Prof. Dr. F. Govers, Wageningen University
Prof. Dr. ir. C. Pieterse, Utrecht University
Dr.ir. G.H.J. Kema, Plant Research International, Wageningen
This research was conducted under the auspices of the Graduate school of
Experimental Plant Sciences.

III
Basal resistance of barley to adapted and
non-adapted forms of Blumeria graminis
Reza Aghnoum
Thesis
Submitted in partial fulfillment of the requirements for the degree of doctor
at Wageningen University
by the authority of the Rector Magnificus
Prof. Dr. M.J. Kropff,
in the presence of the
Thesis Committee appointed by the Doctorate Board
to be defended in public on Tuesday 16 June 2009
at 4 PM in the Aula.

IV
Reza Aghnoum
Basal resistance of barley to adapted and non-adapted forms of Blumeria graminis
132 pages.
Thesis, Wageningen University, Wageningen, NL (2009)
With references, with summaries in Dutch and English
ISBN 978-90-8585-419-7

V
Contents
Chapter 1
General introduction
1
Chapter 2
Which candidate genes are responsible for natural variation in
basal resistance of barley to barley powdery mildew?
15
Chapter 3
Transgressive segregation for extreme low and high level of
basal resistance to powdery mildew in barley
47
Chapter 4
New perspectives to identify genes that determine non-host
resistance of barley to non-adapted Blumeria graminis forms
61
Chapter 5
Rust-induced powdery mildew resistance in barley
79
Chapter 6
General discussion
101
Abbreviations 111
Glossary 113
Summary in English 115
Summary in Dutch (Samenvatting) 119
Acknowledgments 123
About the author 127
List of publications 129
Education certificate 131

VI
This thesis is dedicated with deep gratitude to my dear wife Fatemeh and our lovely
children, Mahrokh and Pouria

General Introduction

Chapter 1
2
Introduction Powdery mildews, a large group of Ascomycota fungal species, are obligate biotrophs
that colonize a wide range of plant species. These fungi consist of 16 genera and
about 650 species (Inuma et al., 2007). Nearly 10,000 species of angiosperms have
been reported as host plants for powdery mildews (Braun, 2002). The host range of
these fungi is restricted to angiosperms, and does not include ferns and
gymnosperms (Takamatsu, 2004). They cause economic loss on a wide range of
important agricultural crops, including cereals. Powdery mildew of the grass family is
caused exclusively by members of the species Blumeria graminis. On the basis of
host range, B. graminis is divided into formae speciales (f.sp., plural ff.spp.), each of
which can infect mainly species of one genus of gramineous plants (Wyand & Brown,
2003). Powdery mildew of barley caused by Blumeria gramins (DC.) Speer f.sp.
hordei Em. Marchal (syn. Erysiphe graminis DC. f.sp. hordei Em. Marchal) is one of
the most important diseases of barley in Europe and many major barley cultivation
areas.
Barley- Blumeria interaction as a model system Due to several reasons, powdery mildew of barley has been studied extensively and it
can be considered as an experimental model system for the study of host-pathogen
interactions.
1- The development of the fungus is restricted to the host epidermal surface, so the
successive developmental stages of germlings including germination, appressorial
germ tube growth and haustorium formation can be easily monitored by light
microscopy. The synchronized development of the fungus on the host facilitates
meaningful experiments where physiological and molecular responses of the barley
host can be correlated perfectly with the development of the fungus, using bioimaging
analyses and transcript profiling (Collinge et al., 2008).
2- The mode of infection by mildew exclusively involves the epidermis. Epidermal
cells can be transiently transformed relatively easily and transient-induced gene
silencing (TIGS) and transient over-expression in single epidermal cells can be
applied, allowing reverse genetics to identify genes that influence the barley-Blumeria
interaction (Douchkov et al., 2005).
3-There is a great availability of valuable genetic stock. More than 85 race-specific
resistance genes for powdery mildew have been identified in barley (Jørgensen,
1994). Many near-isogenic lines (see Glossary) have been developed by back-
crossing different resistance gene donors into common genetic background, resulting
in, e.g., the Pallas differential set (Kølster et al., 1984). Such sets of near-isogenic

General introduction
3
lines form excellent experimental material for comparative studies of defense
responses (Thordal-Christensen et al., 2000). The incompatible phenotype of different
R genes/barley mildew interactions varies from single cell to multi-cell hypersensitive
reaction (HR) or are based on formation of papillae with a subsequent HR. So far six
major resistance genes of barley to powdery mildew (Mla1, Mla6, Mla7, Mla10, Mla12
and Mla13) have been cloned and characterized (Chełkowski et al., 2003, Stein &
Graner, 2004). Furthermore, some genes have been cloned that are required for R-
gene-mediated resistance viz. Rar1 (required for Mla12 resistance-1) and Sgt1
(suppressor of the G2 allele of skp1) (Kitagawa et al., 1999; Azevedo et al., 2002).
4- Barley is the first crop in which an alternative, durable (see Glossary) type of
resistance, mlo resistance, has been found. This resistance is mediated by the
recessive mlo allele of the Mlo (Mildew resistance locus-o) locus. mlo resistance is
effective against all known isolates of the barley powdery mildew fungus (Jørgensen,
1992;Büschges et al., 1997), and has remained so despite extensive cultivation of
mlo-cultivars in Europe since 1979 (Lyngkjær et al., 2000). mlo resistance is
associated with formation of cell wall appositions (papillae) at the site of attempted
fungal penetration. Cloning of the Mlo wild type allele and Ror2, a gene that is
required for mlo-mediated resistance, provided insights into papilla-based penetration
resistance (Büschges et al., 1997; Collins et al., 2003). Non-host resistance of barley
against non-adapted powdery mildews, the basal host resistance and mlo-mediated
resistance are based on similar mechanisms (Trujillo et al., 2004; Humphry et al.,
2006; Schweizer, 2007). Only recently, more than 60 years after the first report of mlo
in barley, effective homologues have been found in tomato, pea and Arabidopsis
thaliana (Collins et al., 2003; Fondevilla et al., 2006; Bai et al., 2008). In Arabidopsis
thaliana these genes are called penetration mutant genes (PEN), and they have been
shown to be required for both non-host resistance and a host resistance that is similar
to the mlo resistance in barley to B. graminis (Consonni et al., 2006).
5. Barley is a diploid autogamous crop, in which DH production is technically
feasible. This is an advantage over an allogamous cereal crop like rye (heterozygous)
and allopolyploid crops like wheat and oat that possess in principle two or three
homologous copies of gene of interest, one on each genome (Elliott et al., 2002).
Barley-powdery mildew: Compatible interaction The powdery mildew pathogen disperses by asexually formed windborne conidia.
Approximately 2 hours after contact with a leaf surface, the conidium germinates with
a primary germ tube (PGT) (Figure 1). This germ tube penetrates through the leaf
cuticle and may have a function in conidial adhesion, water uptake, recognition of leaf

Chapter 1
4
surface structures and perception and transduction of leaf derived signals (Heitefuss,
2001; Eichmann & Hückelhoven, 2008). Within about 4 h, a secondary germ tube,
that is called the appressorial germ tube (AGT), will be formed. AGT differentiates an
appressorium by 8-12 h, from which the fungus tries to penetrate the host cell wall by
a penetration peg. Attacked epidermal cells react to the fungal penetration peg by
papilla deposition. If fungal penetration succeeds, a haustorium will be formed in the
attacked cell. This is an organ by which the fungus obtains its nutrition from the plant
cells and probably organizes reprogramming of the plant cell’s gene expression. After
successful formation of the first haustorium (called establishment) an elongated
secondary hypha (ESH) will start to grow from the primary appressorium. From these
ESH, secondary appressoria are formed that will penetrate epidermal cells to
establish secondary haustoria. Three to four days after formation of the first
haustorium, the pathogen develops conidiophores, on which chains of conidia are
formed. Therefore this development stage is also called conidiation. Released mature
conidia can initiate a new infection cycle (Figure 1).
Penetration resistance in barley-powdery mildew int eraction Penetration thorough the epidermal cell walls is a critical stage during the infection
process of powdery mildews. Epidermal cells actively resist penetration by powdery
mildews in association with papilla formation at the site of attempted fungal
penetration, and hence may prevent haustorium formation (Figure 1). Papillae consist
of a variety of compounds, including callose, phenolic compounds, H2O2, proteins and
glycoproteins with hydrolytic properties (Zeyen et al., 2002). Even in a susceptible
plant some germlings fail to penetrate in association with papilla formation. The
speed of papilla deposition and their chemical composition play an important role in
the success or failure of haustorium formation at that site (Heitefuss, 2001). Upon
failure of attempted penetration from the first appressorial lobe, the appressorium
may form one or two subsequent lobes to try again (Carver, 1986).
In interaction of barley with the powdery mildew pathogen, three types of
resistance, viz. basal host resistance, non-host resistance and mlo-mediated
resistance operate on the basis of similar mechanisms, viz. penetration resistance in
association with papilla formation.
Basal host resistance Even in susceptible plants a proportion of germlings stop at penetration stage in
association with papilla formation due to basal resistance. This resistance is defined
as the plant’s ability to reduce the disease severity caused by an adapted pathogen

General introduction
5
despite a compatible interaction phenotype. Histological studies on apparently
susceptible barley accessions showed that resistance to primary penetration varies
greatly (e.g. Carver, 1986; Asher & Thomas, 1987). Penetration resistance to
powdery mildew in barley also depends on the type of the epidermal cells. Higher
frequencies of successful penetration occurs on the short cells which directly are in
contact with the stomata (epidermal cells type A) and short cells not directly adjacent
Figure 1. Schematic representation of different outcome of interaction of barley with the powdery mildew
pathogen. hai/dai = hours/days after inoculation, PGT=Primary germ tube AGT=Appressorial germ tube,
C=Conidia, HAU=Haustorium, ESH= Elongated secondary hyphae, Pa=Papilla
to stomata (type B) than on the long epidermal cells covering the vascular tissue (Lin
& Edwards, 1974; Koga et al., 1990). Infection rate in both short and long epidermal
cells also depends on the leaf age. On older leaves, a lower proportion of germlings
succeed to establish than on younger leaves (Lin & Edwards, 1974; Carver, 1986).
Basal resistance is a quantitatively inherited trait, conferred by many, mostly
additively acting genes, each with small effect, so it is also called quantitative
resistance. Advances in molecular marker technology and QTL analysis have led to
identification of many QTLs in different genetic backgrounds of barley, but the genes
underlying the resistance QTLs are still elusive. Using transient-induced gene
silencing and transient over-expression in single epidermal cells, many candidate
genes have been identified that play a role in host, non-host and/or mlo-mediated
resistance (e.g. Douchkov et al., 2005; Zimmermann et al., 2006) but still it is
questionable whether the allelic variation in those candidate genes also determines
natural variation in level of basal resistance between plant accessions.
:::
ESH
Hypersensitive response
R genes, Mla1, Mla6 etc.
Papillae defense
mlo-mediated, Non-host
and basal host resistance
5-8 dai
AGT C
Compatible interaction
HAU
72hai
12hai
:::
PGT PA
C

Chapter 1
6
Non-host resistance The grass powdery mildew pathogen, Blumeria graminis, is classified into several
formae speciales each colonizing an individual genus of the grass family. Blumeria
graminis f.sp. hordei (Bgh), the causal agent of powdery mildew disease on barley,
for example, successfully colonizes cultivated barley Hordeum vulgare and its wild
ancestor H. spontaneum but is probably not able to colonize other related species in
the genus Hordeum nor other species in the Triticeae, such as wheat (Eshed & Wahl,
1970; Menzies & MacNeill, 1989). Conversely, cultivated barley cannot be colonized
by powdery mildew isolates collected on other Hordeum species and other Triticeae.
This resistance is known as non-host resistance, and is mainly associated with papilla
formation at sites of attempted fungal penetration. A proportion of challenged
epidermal cells show, however, a hypersensitive reaction. Although a similar
mechanism of defense is involved in basal resistance of barley to the adapted
pathogen, Bgh, at macroscopic and microscopic level a clear discrimination can be
made between basal host resistance and non-host resistance, since in the latter, the
resistance is complete. Since there is not any barley accession which is susceptible
to the non-adapted mildew forms, like the wheat powdery mildew pathogen (Bgt), it is
not known which genes determine the host status of barley to the non-adapted Bgt.
mlo-mediated resistance Resistance mediated by the recessive loss-of-function mutant at the Mlo locus is
race-non-specific, since it is effective against all known isolates of Bgh (Büschges et
al., 1997). Several resistance alleles of mlo have been derived from mutagenic
treatment of susceptible wildtype Mlo barley cultivars, but the mlo resistance allele
has also been identified from non-mutagenized landraces of barley from Ethiopia
(Jørgensen, 1992). In mlo resistant plants, resistance at the penetration stage in
association with papilla formation prevents the fungus to form haustoria in all kinds of
epidermal cells, except the subsidiary cells of stomata. Papilla formation occurs also
in Mlo susceptible plants when challenged by the penetration peg of the germinated
powdery mildew conidium, but the penetration efficiency of Bgh is considerably higher
in Mlo susceptible plants than in mlo resistant plants (Freialdenhoven et al., 1996).
On the other hand, papilla based responses occur earlier and the size of papillae are
generally larger at the failed penetration sites in mlo resistant lines than at the sites of
failed penetration in Mlo susceptible plants (Jørgensen, 1992).
mlo-mediated resistance shows some level of pathogen specificity, since mlo alleles
are not effective against other fungal pathogens including barley leaf rust (Puccinia
hordei), stripe rust (Puccinia striiformis), stem rust (Puccinia graminis), scald

General introduction
7
(Rhynchosporium secalis), and the take all fungal disease (Gaeumannomyces
graminis) (Jørgensen, 1977; Collins et al., 2007). However mlo seems to play a role
in non-host resistance of barley to the non-adapted mildew pathogen, Bgt
(Peterhänsel et al., 1997; Trujillo et al., 2004; Humphry et al., 2006). Mlo gene
homologs have now been identified in a wide range of plants, including wheat, rice,
tomato, pea, grapevine and the model plant Arabidopsis thaliana, some of which
have been demonstrated to contribute to penetration resistance to powdery mildew
(Elliott et al., 2002; Consonni et al., 2006; Fondevilla et al., 2006; Bai et al., 2008;
Winterhagen et al., 2008).
mlo and non-host resistance share similar molecular co mponents mlo and non-host resistance share many analogous features (Humphry et al., 2006).
1- Both types of resistance are associated with strong penetration resistance based
on papilla formation at the penetration sites. 2- Both mlo and non-host resistance are
broad spectrum and durable. 3- They do not depend on Salicylic acid (SA), Jasmonic
acid (JA) or Ethylene-mediated defense signaling pathways but both depend on actin
cytoskeleton components. 4- Both types of resistance require PEN genes
(Arabidopsis) or Ror genes (barley). 5- Both depend on the function of the SNAP34
protein.
Cloning of the Mlo wild type allele, identification of Ror genes and characterization
of Arabidopsis mutants with atenuated non-host interaction with Bgh contributed a lot
to our understanding of the molecular basis of penetration resistance to powdery
mildews (see Figure 2; Freialdenhoven et al., 1996; Büschges et al., 1997; Collins et
al., 2003; Consonni et al., 2006). Two genes, Ror1 and Ror2 (required for mlo-
specified resistance) are required for the function of mlo in barley. Mutation either in
Ror1 or Ror2 decrease powdery mildew resistance in mlo genotypes (Freialdenhoven
et al., 1996). Peterhänsel et al., (1997) showed that mlo and Ror genes are active
when barley is challenged with Bgt, a non-adapted powdery mildew. Trujillo et al.
(2004) indicated that mutants of Ror1 and Ror2 showed higher frequencies of fungal
penetration in interaction with Bgt. Screening of mutagenized Arabidopsis populations
for increased frequencies of penetration by the non-adapted powdery mildew fungus
Bgh, resulted in identification of several penetration (pen) mutants allowing higher
frequencies of Bgh entry into epidermal cells of which three, designated pen1, pen2,
and pen3, have been characterized and the corresponding genes were identified
(Collins et al., 2003; Lipka et al., 2005; Stein et al., 2006). Cloning of PEN1 showed
that it is a homologue of barley Ror2 and that this gene encodes SYP121, a syntaxin
which is known to contribute to vesicle fusion events through the formation of SNARE

Chapter 1
8
complexes with a corresponding VAMP (vesicle associated membrane protein) and a
SNAP25 homologue (Collins et al., 2003). PEN2 encodes a glycoside hydrolase that
is required for the enzymatic activation of small molecules and is involved in the
production of an antifungal compound (Lipka et al., 2005). PEN2 protein was found to
be localized to peroxisomes, organelles that accumulate at the sites of attempted
powdery mildew penetration (Koh et al., 2005; Lipka et al., 2005). PEN3 encodes an
ATP-binding cassette (ABC) transporter protein (Stein et al., 2006) possibly involved
in exports of a potentially fungi-toxic compound that is processed by PEN2 in
peroxisomes into the apoplast (Figure 2).
Figure 2. Schematic overview of molecular components involved in penetration resistance to powdery mildew. A
fungal appressorium trying to breach the host cell wall by a penetration peg is stopped in a cell wall apposition
(papilla). PEN1/Ror2 syntaxin and HvSNAP34 protein contribute to SNARE complex formation and secretion of
antimicrobial compounds at sites of attempted fungal penetration. PEN3 is involved in export of potentially toxic
compounds that were released by PEN2 enzyme activity in peroxisomes (reprinted by permission from Collins et
al., 2007; http://www.publish.csiro.au/nid/40/paper/AR06065.htm).
The possible functional link between mlo-mediated and non-host resistance was
recently supported by identification of some candidate genes that emerged from
transient overexpression or transient-gene silencing studies. These candidate genes
were found to be required for mlo-mediated resistance against Bgh as well as for
non-host resistance to Bgt (Douchkov et al., 2005; Miklis et al., 2007; Schweizer,
2007). Furthermore, overexpression of Mlo in epidermal cells of barley resulted in
some degree of susceptibility to the non-adapted Bgt pathogen suggesting that Mlo
may negatively regulate non-host resistance as well (Elliott et al., 2002).

General introduction
9
Suppression and induction of penetration resistance Adapted pathogens have the ability to overcome preformed defense barriers and
suppress induced defense responses, but non-adapted pathogens do not have such
ability (Thordal-Christensen, 2003). Different fungal pathogens employ different
suppression strategies to infect and colonize their hosts (Staples & Mayer, 2003).
Biotrophic and haustorium forming fungal pathogens, like powdery mildews and rusts,
first should suppress penetration resistance to be able to successfully penetrate
through epidermal or mesophyll cell walls. Successful penetration and haustorium
formation of virulent isolates of Blumeria graminis induces susceptibility to haustorium
formation by subsequent challenge of either adapted or non-adapted powdery mildew
pathogens. On the other hand, failed attempted penetration induces resistance to
haustorium formation by subsequent challenge inoculation with a compatible isolate
(e.g. Lyngkjær & Carver, 1999; Lyngkjær et al., 2001; Kunoh, 2002; Olesen et al.,
2003). This induction of susceptibility and resistance at cell level is often called
accessibility and inaccessibility. Studies on non-host interactions with cowpea rust
fungus and plantain powdery mildew fungus suggests that the adhesion between the
plant cell wall and the plasma membrane plays a role in cell wall-associated defense
responses and powdery mildews and rust fungi employ different strategies to
successfully induce accessibility of plant cells (Mellersh & Heath, 2001; Heath, 2002).
Scope and outline of the thesis In this thesis we investigated the genetic variation in barley for resistance to the
adapted and non-adapted powdery mildew pathogens and we intend to identify the
genes that are responsible for natural variation in host status and non-host resistance
level. We aimed also to determine whether the basal resistance of barley to powdery
mildew is being suppressed by prior attack by a different pathogenic haustorium
forming fungus, Puccinia hordei that differs from the mildew fungus in its strategy to
induce accessibility.
In Chapter 2, a multi-population QTL analysis was conducted for powdery mildew
resistance of barley at seedling and at adult plant stage. Twenty-three EST/cDNA-
clone sequences were mapped that are likely to play a role in the barley-Blumeria
interaction based on transcript profiling, gene-silencing or over-expression data, and
mlo, Ror1 and Ror2. To compare the map position of QTLs over different mapping
populations, we constructed an improved high-density integrated linkage map of
barley, based on the marker data of seven mapping populations of barley.
In Chapter 3, two experimental barley lines were developed with extremely high and
low level of basal resistance to powdery mildew by convergent crossing between the

Chapter 1
10
most resistant and the most susceptible lines from four mapping populations of
barley.
In Chapter 4, a large collection of barley germplasm was screened and some rare
barley accessions were identified with rudimentary susceptibility to the non-adapted
wheat powdery mildew, Bgt. Those accessions were intercrossed in two cycles, and
resulted in two lines, called SusBgt SC and SusBgt DC, with exceptional susceptibility
to Bgt, that were tested with seven non-adapted forms of Blumeria graminis, to
investigate the specificity of the non-host resistance of barley.
In Chapter 5, the rust-induced powdery mildew resistance at the cellular level was
characterized. We also identified differentially expressed genes that may explain the
strong papilla-based penetration resistance in double inoculated barley plants.
In Chapter 6, the results obtained in the previous Chapters, the importance of the
experimental barley lines that were developed in this study for the identification of
genes that determine basal resistance of barley to adapted and non-adapted
Blumeria graminis forms is being discussed.
References
Asher MJC, Thomas CE. 1987. The inheritance of mechanisms of partial resistance to Erysiphe
graminis in spring barley. Plant Pathology 36: 66-72.
Azevedo C, Sadanandom A, Kitagawa K, Freialdenhoven A, Shirasu K, Schulze-Lefert P. 2002.
The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance.
Science 295: 2073-2076.
Bai Y, Pavan S, Zheng Z, Zappel NF, Reinstädler A, Lotti C, De Giovanni C, Ricciardi L,
Lindhout P, Visser R, Theres K, Panstruga R. 200 8. Naturally occurring broad-spectrum
powdery mildew resistance in a Central American tomato accession is caused by loss of Mlo
function. Molecular Plant-Microbe Interactions 21: 30-39.
Braun U, Cook RTA, Inman AJ, Shin HD. 2002. The taxonomy of powdery mildew fungi. In:
Bélanger RR, Bushnell WR, Dik AJ, Carver TLW, eds. The powdery mildews: A comprehensive
treatise. St. Paul/Minnesota, USA: APS Press, 13–55.
Büschges R, Hollricher K, Panstruga R, Simons G, Wo lter M, Frijters A, vanDaelen R, vander
Lee T, Diergaarde P, Groenendijk J, et al. 1997. The barley Mlo gene: a novel control element
of plant pathogen resistance. Cell 88: 695–705.
Carver TLW. 1986. Histology of infection by Erysiphe graminis f. sp. hordei in spring barley lines with
various levels of partial resistance. Plant Pathology 35: 232–240.
Chelkowski j, Tyrka M, Sobkiewicz A. 2003 . Resistance genes in barley (Hordeum vulgare L.) and
their identification with molecular markers. Journal of Applied Genetics 44: 291-309.
Collinge DB, Jensen MK, Lyngkjaer MF, Rung J. 2008. How can we exploit functional genomics
approaches for understanding the nature of plant defences? Barley as a case study. European
Journal of Plant Pathology 121: 257-266
Collins NC, Niks RE, Schulze-Lefert P, 2007. Resistance to cereal rusts at the plant cell wall - What
can we learn from other host-pathogen systems? Australian Journal of Agricultural Research
58: 476-489.

General introduction
11
Collins NC, Thordal-Christensen H, Lipka V, Bau S, Kombrink E, Qiu JL, Hückelhoven R, Stein,
M, Freialdenhoven, A, Somerville SC, Schulze-Lefert P. 2003. SNARE-protein-mediated
disease resistance at the plant cell wall. Nature 425: 973-977.
Consonni C, Humphry ME, Hartmann HA, Livaja M, Durn er J, Westphal L, Vogel J, Lipka V,
Kemmerling B, Schulze-Lefert P, et al. 2006. Conserved requirement for a plant host cell
protein in powdery mildew pathogenesis. Nature Genetic 38: 716–720.
Douchkov D, Nowara D, Zierold U, Schweizer P. 2005. A high-throughput gene-silencing system for
the functional assessment of defense-related genes in barley epidermal cells. Molecular Plant-
Microbe Interactions 18: 755-761.
Eichmann R, Hückelhoven R. 2008. Accommodation of powdery mildew fungi in intact plant cells.
Journal of Plant Physiology 165: 5–18.
Elliott C, Zhou FS, Spielmeyer W, Panstruga R, Schu lze-Lefert P. 2002. Functional conservation of
wheat and rice Mlo orthologs in defense modulation to the powdery mildew fungus. Molecular
Plant-Microbe Interactions. 15: 1069-1077.
Eshed N, Wahl I. 1970. Host ranges and interrelations of Erysiphe graminis hordei, E. graminis tritici,
and E. graminis avenae. Phytopathology 60: 628-634.
Fondevilla S, Carver TLW, Moreno MT, Rubiales D. 20 06. Macroscopic and histological
characterisation of genes er1 and er2 for powdery mildew resistance in pea. European Journal
of Plant Pathology 115: 309–321.
Freialdenhoven A, Peterhänsel C, Kurth J, Kreuzaler F, Schulze-Lefert P. 1996. Identification of
genes required for the function of non-race specific mlo resistance to powdery mildew in barley.
Plant Cell 8: 5-14.
Heath MC. 2002. Cellular interactions between biotrophic fungal pathogens and host or nonhost
plants. Canadian Journal of Plant Pathology 24: 259–264.
Heitefuss R. 2001. Defence reactions of plants to fungal pathogens: principles and perspectives,
using powdery mildew on cereals as an example. Naturwissenschaften 88: 273-283.
Humphry M, Consonni C, Panstruga R. 2006. mlo-based powdery mildew immunity: silver bullet or
simply non-host resistance? Molecular Plant Pathology 7: 605–610.
Inuma T, Khodaparast SA, and Takamatsu S. 2007. Multilocus phylogenetic analyses within
Blumeria graminis, a powdery mildew fungus of cereals. Molecular Phylogenetic and Evolution
44: 741-51
Jørgensen H. 1994. Genetics of powdery mildew resistance in barley. Critical Review in Plant
Sciences 13: 97-119.
Jørgensen JH. 1977. Spectrum of resistance conferred by mlo powdery mildew resistance genes in
barley. Euphytica 26: 55-62.
Jørgensen JH. 1992. Discovery, characterization and exploitation of Mlo powdery mildew resistance
in barley. Euphytica 63: 141–152.
Kitagawa K, Skowyra D, Elledge SJ, Harper J, Hieter P. 1999. SGT1 encodes an essential
component of the yeast kinetochore assembly pathway and a novel subunit of the SCF ubiquitin
ligase complex. Molecular Cell 4: 21-33.
Koga H, Bushnell WR, Zeyen RJ. 1990. Specificity of cell type and timing of events associated with
papilla formation and the hypersensitive reaction in leaves of Hordeum vulgare attacked by
Erysiphe graminis f. sp. hordei. Canadian Journal of Botany. 68: 2344–2352.

Chapter 1
12
Koh S, Andre A, Edwards H, Ehrhardt D, Somerville S . 2005. Arabidopsis thaliana subcellular
responses to compatible Erysiphe cichoracearum infections. The Plant Journal 44: 516–529.
Kølster P, Munk L, Stølen O, Løhde J, 1986. Near-isogenic barley lines with genes for resistance to
powdery mildew. Crop Science 26: 903–907.
Kunoh H. 2002. Localized induction of accessibility and inaccessibility by powdery mildew. In:
Bélanger RR, Bushnell WR, Dik AJ, Carver TLW, eds. The powdery mildews: A comprehensive
treatise. St. Paul/Minnesota, USA: APS Press, 126–133.
Lin MR, Edwards HH. 1974. Primary penetration process in powdery mildewed barley related to host
cell age, cell type, and occurrence of basic staining material. New Phytologist 73: 131-137.
Lipka V, Dittgen J, Bednarek P, Bhat R, Wiermer M, Stein M, Landtag J, Brandt W, Rosahl S,
Scheel D, et al. 2005. Pre- and postinvasion defences both contribute to nonhost resistance in
Arabidopsis. Science 310: 1180-1183.
Lyngkjær MF, Carver TLW, Zeyen RJ. 2001. Virulent Blumeria graminis infection induces
penetration susceptibility and suppresses race-specific hypersensitive resistance against
avirulent attack in Mla1-barley. Physiological and Molecular Plant Pathology 59: 243-256.
Lyngkjær MF, Carver TLW. 1999. Induced accessibility and inaccessibility to Blumeria graminis f.sp.
hordei in barley epidermal cells attacked by a compatible isolate. Physiological and Molecular
Plant Pathology 55: 151–162.
Lyngkjær MF, Newton AC, Atzema JL, Baker SJ. 2000. The barley mlo gene: an important powdery
mildew resistance source. Agronomie 20: 745–756.
Mellersh DG, Heath MC. 2001. Plasma membrane-cell wall adhesion is required for expression of
plant defense responses during fungal penetration. Plant Cell 13: 413–424.
Menzies JG, MacNeill BH. 1989. Infection of species of the Gramineae by Erysiphe graminis f.sp.
hordei and Erysiphe graminis f.sp. tritici. Canadian Plant Disease Survey 69: 105-108.
Miklis M, Consonni C, Bhat RA, Lipka V, Schulze-Lef ert P, Panstruga R. 2007. Barley mlo
modulates actin-dependent and actin-independent antifungal defense pathways at the cell
periphery. Plant Physiology 144: 1132–1143.
Olesen KL, Carver TLW, Lyngkjær MF . 2003. Fungal suppression of resistance against
inappropriate Blumeria graminis formae speciales in barley, oat and wheat. Physiological and
Molecular Plant Pathology 62: 37-50.
Peterhänsel C, Freialdenhoven A, Kurth J, Kolsch R, Schulze-Lefert P. 1997. Interaction analyses
of genes required for resistance responses to powdery mildew in barley reveal distinct pathways
leading to leaf cell death. Plant Cell 9: 1397–1409.
Schweizer P. 2007. Nonhost resistance of plants to powdery mildew-New opportunities to unravel the
mystery. Physiological and Molecular Plant Pathology 70: 3–7.
Staples RC, Mayer AM. 2003. Suppression of host resistance by fungal plant pathogens. Israel
journal of plant sciences 51: 173-184.
Stein M, Dittgen J, Sanchez-Rodriguez C, Hou BH, Mo lina A, Schulze-Lefert P, Lipka V,
Somerville S. 2006. Arabidopsis PEN3/PDR8, an ATP cassette transporter, contributes to
nonhost resistance to inappropriate pathogens that enter by direct penetration. Plant Cell 18: 1-
16. Stein N, Graner A. 2004. Map-based gene isolation in cereal genomes. In: Gupta PK, Varshney RK,
eds. Cereal Genomics. Dordrecht, the Netherlands: Kluwer Academic Publishers, 331–360.
Takamatsu S .2004. Phylogeny and evolution of the powdery mildew fungi (Erysiphales, Ascomycota)
inferred from nuclear ribosomal DNA sequences. Mycoscience 45: 147-157.

General introduction
13
Thordal-Christensen H, Gregersen PL, Collinge DB. 2 000. The barley/Blumeria (syn. Erysiphe)
graminis interaction: a case study. In: Slusarenko AJ, Fraser RSS, van Loon LC, eds.
Mechanisms of Resistance to Plant Diseases. Dordrecht, the Netherlands: Kluwer Academic
Publishers, 77–100.
Thordal-Christensen H. 2003. Fresh insights into processes of non-host resistance. Current Opinion
in Plant Biology 6: 351-357.
Trujillo M, Troeger M, Niks RE, Kogel KH, Hückelhov en R. 2004. Mechanistic and genetic overlap
of barley host and non-host resistance to Blumeria graminis. Molecular Plant Pathology 5: 389-
396.
Winterhagen P, Howard SF, Qiu W, Kovacs LG. 2008. Transcriptional up-regulation of grapevine
MLO genes in response to powdery mildew infection. American Journal of Enology and
Viticulture. 59: 159-168.
Wyand RA. Brow, JKM. 2003. Genetic and forma specialis diversity in Blumeria graminis of cereals
and its implications for host-pathogen co-evolution. Molecular Plant Pathology 4: 187-198.
Zeyen RJ, Carver TLW, Lyngkjær MF. 2002. The formation and role of papillae. In: Bélanger RR,
Bushnell WR, Dik AJ, Carver TLW, eds. The powdery mildews: A comprehensive treatise. St.
Paul/Minnesota, USA: APS Press, 107-125.
Zimmermann G, Baumlein H, Mock HP, Himmelbach A, Sc hweizer P. 2006. The multigene family
encoding germin-like proteins of barley. Regulation and function in basal host resistance. Plant
Physiology 142: 181-192.

Chapter 1
14

Which candidate genes are responsible for natural
variation in basal resistance of barley to barley
powdery mildew?
R. Aghnoum, T.C. Marcel, A. Johrde, N. Pecchioni, P. Schweizer and R.E. Niks
Submitted

Chapter 2
16
Abstract
Basal resistance of barley against powdery mildew is a quantitatively inherited trait
that is based on non-hypersensitive mechanisms of defense. A functional genomic
approach indicated many candidate genes to be involved in defense against fungal
haustorium formation. It is not known for which of those candidate genes allelic
variation may contribute to the natural variation for powdery mildew resistance, since
many of them may be highly conserved within the barley species. Twenty-three
EST/cDNA-clone sequences that are likely to play a role in the barley-Blumeria
interaction based on transcript profiling, gene-silencing or over-expression data, and
mlo, Ror1 and Ror2, were mapped and are considered candidate genes to contribute
to basal resistance. We mapped QTLs for powdery mildew resistance in six mapping
populations of barley at seedling and/or at adult plant stage and developed an
improved high-density genetic integrated map containing 6,990 markers for
comparing QTLs and candidate gene positions over mapping populations. We
mapped 12 QTLs at seedling stage and 13 QTLs at adult plant stage of which four
were in common between the two developmental stages. Six of the candidate genes
showed coincidence in their map positions with the QTLs identified for basal
resistance to powdery mildew. This co-localization justifies to give priority to those six
candidate genes to verify them as being responsible for the phenotypic effects of the
QTLs for basal resistance.
Introduction The barley-powdery mildew pathosystem is one of the best characterized and highly
specialized systems of host-parasite interaction. More than 85 race-specific
resistance genes to powdery mildew have been identified in barley (Jørgensen, 1994;
Chelkowski et al., 2003). They act through a hypersensitivity response (HR), which is
only elicited by particular avirulent pathogen isolates according to the gene-for-gene
concept (Collins et al., 2002). Upon deployment of resistance genes in breeding
programs, virtually all of these genes became ineffective (Czembor, 2000). Six major
resistance genes of barley to powdery mildew (Mla1, Mla6, Mla7, Mla10, Mla12, and
Mla13) have been cloned and characterized (Chelkowski et al., 2003; Stein & Graner,
2004).
Another type of resistance is known as basal (partial) resistance and is conferred by
many, mostly additively acting genes, each with a small effect. Basal resistance has
been used to indicate a low-level penetration resistance expressed in 'susceptible'
plants, which is not due to hypersensitivity (e.g. Collins et al., 2003). Since partial
resistance is defined as a resistance characterized by a susceptible infection type

Which candidate genes are….
17
and a reduced disease severity (Parlevliet, 1975) we presume in this manuscript that
partial and basal resistance in this pathosystem are synonymous terms. Basal
resistance is considered as an important and potentially durable type of resistance.
Nevertheless, it is the least investigated type of resistance. In barley-powdery mildew
interaction, basal resistance is associated with formation of cell wall appositions, so-
called papillae, at the fungal penetration sites and this reinforcement of cell walls
prevents haustorium formation in the attacked epidermal cells. Failure of haustorium
formation is also called “pre-haustorial” resistance and is in general not associated
with a hypersensitive reaction of the challenged plant cells (Niks & Rubiales, 2002).
Even the most susceptible barley genotypes prevent a substantial proportion of
mildew infection units from forming their first haustorium, implying that all genotypes
have at least some degree of basal resistance. Genotypic variation in level of basal
resistance of barley to powdery mildew has been reported by several researchers
(Asher & Thomas, 1984; Jones & Davies, 1985; Carver, 1986; Knudsen et al., 1986;
Mastebroek & Balkema-Boomstra, 1991). Histological studies on partially resistant
genotypes showed that resistance to primary penetration by non-hypersensitive
mechanisms affect a varying proportion (26-75%, dependent on barley accession) of
attempted penetrations to fail (Asher & Thomas, 1987). Carver (1986) reported that in
spring barley lines with various levels of basal resistance, a range from 48 to 90% of
infection units failed at penetration stage.
Development of molecular marker techniques has enabled researchers to dissect
quantitatively inherited traits, like basal resistance, by QTL analysis and to study the
effects of individual QTLs, including their interactions. In contrast to single and
dominant major genes, the genetic and molecular basis of quantitative disease
resistance is less well understood, because of the polygenic inheritance, the relatively
small effect of individual genes and significant genotype × environment interactions
that make such traits experimentally more difficult to assess. However, research is
underway to isolate the genes underlying QTLs for basal resistance of barley to
barley leaf rust Puccinia hordei following map-based cloning approaches (Marcel,
2007b). Such an approach would be feasible to identify the genes underlying the
powdery mildew QTLs.
The recessive allele of the Mlo-locus of barley mediates a race non-specific
resistance to the barley powdery mildew. mlo resistance and basal resistance share
the same mechanism of defense, i.e. both are “papilla-based” and “pre-haustorial”
(Trujillo et al., 2004; Schweizer, 2007). Mutational approaches revealed that two
genes (Ror1 and Ror2) are required for full expression of mlo resistance
(Freialdenhoven et al., 1996). These two genes have also been implicated in basal

Chapter 2
18
penetration resistance of barley expressed in susceptible (Mlo) genotypes (Collins et
al., 2003). So, mlo, Ror1 and Ror2 can be considered as candidate genes to be
involved in basal resistance of barley to powdery mildew.
Candidate gene analysis is based on the hypothesis that genes with known function
could correspond to loci controlling traits of interest (Pflieger et al., 2001). This
approach has been widely applied for genetic dissections of complex and quantitative
traits (Zhu & Zhao, 2007). For plant geneticists, a candidate gene is any gene
putatively involved in trait variation, based on its biological function, its map position
and/or its expression pattern (Pflieger et al., 2001). Ideally, the list of candidate genes
to explain a particular phenotype can be shortened by following a prioritarization
approach, which consist in combining information on the function, position and
expression of the genes involved (Zhu & Zhao, 2007). However, finally the candidate
genes should be validated through physiological analyses or genetic transformation.
To identify genes that influence barley-Blumeria interactions in host and non-host
combinations, a high-throughput RNAi system for transient-induced gene silencing
(TIGS) as well as a transient over-expression in single epidermal cells of barley has
been developed (Schweizer et al., 1999; Douchkov et al., 2005; Ihlow et al., 2008).
Using this system, the function of 930 barley candidate genes including 693 up-
regulated genes and 101 resistance-gene analogues expressed in barley epidermis
have been tested for their effect on non-host resistance of barley to wheat powdery
mildew and on the level of basal host resistance. The role of 43 genes in non-host
resistance, basal resistance and/or mlo-mediated resistance of barley has been
confirmed by using this method (Dong et al., 2006; Zimmermann et al., 2006; Johrde
& Schweizer, 2008). Identification of candidate genes whose expression is correlated
with haustorium formation as shown after overexpression or silencing does not
necessarily mean that they are also responsible for natural variation in levels of basal
resistance. Such candidate genes may simply be conserved within the plant
population.The study of the inheritance of natural variation can be considered as an
alternative approach to elucidate the functional role of candidate genes in a given
phenotype (Mitchell-Olds & Schmitt, 2006).
In the present study we mapped genes that determine natural variation in level of
basal resistance in six barley mapping populations and compared their map positions
with those of 23 markers derived from gene sequences that were found to be
pathogen-induced or that produced a resistance phenotype upon RNAi or over-
expression during the interaction of barley with powdery mildew and also mlo, Ror1
and Ror2 genes. We combined the expression and function of candidate genes with

Which candidate genes are….
19
their map position in order to prioritize them. We also determined whether QTLs
tended to map to major gene loci of resistance to powdery mildew.
Materials and Methods
Marker data sets Segregating marker data from seven individual mapping populations (Table 1) were
compiled to prepare a high-density genetic integrated map of barley. The populations
consisted of four doubled haploids (DH) populations, i.e. Igri × Franka (IgFr, Graner et
al., 1991), Steptoe × Morex (StMo, Kleinhofs et al., 1993), the Oregon Wolfe Barleys
(OWBs, Costa et al., 2001) and Nure × Tremois (NuTr, Francia et al., 2004), and of
three recombinant inbred lines (RIL) populations, i.e. L94 × Vada (LnVa, Qi et al.,
1998), Vada × SusPtrit (VaSu, Jafary et al., 2006) and Cebada Capa × SusPtrit
(CCSu, Jafary et al., 2008).
We reused the data sets compiled by Marcel et al., (2007a) after excluding the
AFLP markers mapped on the OWBs. We added the DArT (diversity arrays
technology) and TDM (transcript-derived marker) marker data sets released by Wenzl
et al., (2006) and Potokina et al., (2008) [StMo], respectively, the SSR and EST
marker data sets released by Varshney et al., (2007) and Stein et al., (2007) [IgFr,
StMo, OWBs], respectively, and new marker data posted on the Oregon State
University (OSU) Barley Project web site (http://www.barleyworld.org/) [OWBs]. The
marker data set of NuTr consisted of 542 markers, mostly DArT markers (72%), and
was obtained from Dr. Nicola Pecchioni (Università di Modena e Reggio Emilia, Italy).
Segregating data from most of the BIN markers defined by Kleinhofs & Graner (2001)
[StMo] were obtained from Dr. Andris Kleinhofs (Washington State University). Three
new marker data sets were also included in the present integrated map and consist of
168 “PERO” markers mapped in LnVa and VaSu by peroxidase-profiling (to be
published elsewhere by our laboratory), 26 new EST-SSR markers mapped in NuTr
or CCSu by A/T labelling (Marcel et al., 2007a), and the 23 candidate genes
described in the present study (Table 4).
Construction of the barley integrated map The integrated map was constructed essentially as described in Marcel et al., (2007a)
using the software JoinMap 4TM and MapChart (Voorrips, 2002). All the bridge
markers (i.e. markers common to two or more populations), the 210 BIN markers
defined by Marcel et al., (2007a) and the additional BIN markers provided by Dr. A.
Kleinhofs were used to prepare the framework map. The order of the 210 BIN
markers was initially used as fixed order to run the regression mapping algorithm.

Chapter 2
20
After each mapping, the obtained marker order was compared with the marker order
on the integrated map of Marcel et al., (2007a). Most discrepancies between the two
marker orders were investigated by graphical genotyping in order to reject or to
validate the newly calculated map. In cases where the newly calculated map was
rejected, the fixed order was modified and the mapping repeated until the
discrepancy was corrected. Consequently, in cases where the new map was
validated, it could only represent an improvement from the previous integrated map of
Marcel et al., (2007a). Then, the marker order on the constructed framework map was
used as fixed order to run the maximum likelihood mapping algorithm on the 49
linkage groups (i.e. seven barley chromosomes for seven mapping populations);
default values were increased to five map optimization rounds with 3,000 chains.
Three maps were generated for each linkage group and aligned to compare the order
and distances between markers. From each alignment/linkage group, the map
showing the most consistent marker order and the shortest total length was
integrated into the framework map following the “neighbors” approach. The obtained
integrated map was divided into 217 BINs of about 5 cM each. The BIN-defining
markers of Kleinhofs & Graner (2001) were maintained in their role while the BIN-
defining markers of Marcel et al., (2007a) were sometimes modified to obtain a more
homogeneous distribution of the BINs over the map.
Plant and pathogen materials Six out of seven (IgFr excluded) mapping populations were evaluated for their level of
basal resistance to barley powdery mildew at seedling stage under controlled
conditions and five of them (IgFr and OWBs excluded) at adult plant stage under field
conditions. An isolate of Blumeria graminis f.sp. hordei collected in Wageningen 2004
(Wag.04) was used for evaluation of resistance levels at seedling stage. This isolate
was maintained in a climate cabinet and propagated on young seedlings of the
susceptible barley cultivar ‘Manchuria’.
Table 1. Characteristics of the seven barley mapping populations used to construct the integrated map of
which six were used for QTL analysis.
Name of population Type Lines N umber of markers
L94 × Vada (LnVa) RIL 103 1083
Vada × SusPtrit (VaSu) RIL 152 533
C.Capa × SusPtrit (CCSu) RIL 113 519
OWBs DH 94 882
Steptoe × Morex (StMo) DH 150 3561
Nure × Tremois (NuTr) DH 118 542
Igri × Franka (IgFr) DH 71 737

Which candidate genes are….
21
Table 2 . Comparison of several (integrated) linkage maps published for barley Map length
(cM)
Number
of loci
Average locus number per cM Contributing Populations 1
1H
2H
3H
4H
5H
6H
7H
Rostoks et al.,
2005
1,211 1,237 0.9 1.1 1.0 1.1 0.9 1.0 1.2 LiHs, OWBs, StMo
Wenzl et al., 2006 1,161 2,935 2.8 3.1 2.5 1.9 2.0 2.5 3.0 BaCP, ClSa, DaZh, FoCI, FrSt, IgAt, PaTa,
StMo, TXFr, YeFr
Marcel et al., 2007 1,081 3,258 2.6 3.7 3.3 2.4 2.3 2.9 3.8 LnVa, VaSu, StMo, OWBs, IgFr, CCSu
Stein et al., 2007 1,118 1,055 1.0 1.1 0.9 0.8 0.9 0.8 1.2 StMo, OWBs, IgFr
Potokina et al.,
2008
1,010 1,596 1.3 1.8 1.9 1.3 1.4 1.5 1.7 StMo
This report 1,093 6,990 6.4 7.9 6.9 4.5 5.7 5.9 7.3 LnVa, VaSu, StMo, OWBs, IgFr, CCSu,
NuTr
1 LiHs (Lina x Hordeum spontaneum HS92); OWBs (Oregon Wolfe Barleys); StMo (Steptoe x Morex); BaCP (Barque-73 x CPI71284-48); ClSa (Clipper x Sahara);
DaZh (Dayton x Zhepi2); FoCI (Foster x CI4196); FrSt (Fredickson x Stander); IgAt (Igri x Atlas68); PaTa (Patty x Tallon); TXFr (TX9425 X Franklin); YeFr (Yerong x
Franklin); LnVa (L94 x Vada); VaSu (Vada x SusPtrit); IgFr (Igri x Franka); CCSu (Cabada Capa x SusPtrit); NuTr (Nure x Tremois)

Chapter 2
22
Assessment of resistance level at seedling stage Evaluation of powdery mildew resistance at seedling stage was performed on
detached leaves (Schwarzbach, 2004). Barley plants were grown in spore-free
greenhouse compartments. Twelve days after sowing, when the first leaf was fully
expanded, a segment of about 4 cm long was cut from the middle part of the first
leaves, and both cut ends were immersed in 0.6% water agar medium containing 125
ppm benzimidazole in a square (12 × 12 cm) polystyrene petri dish, the adaxial side
of the leaf facing upwards. Each petri dish accommodated about 16 leaf segments.
One leaf segment was taken per RIL/DH line per experiment. Both parents of each
mapping population and the susceptible cultivar ‘Manchuria’ were added to each petri
dish as reference. To use freshly produced conidia for inoculation, young seedlings of
the susceptible cultivar ‘Manchuria’ were densely inoculated with powdery mildew 7
days prior to inoculation of the experiment. A settling tower was used for inoculations
(Schwarzbach, 2004). Per inoculation, four petri dishes containing the leaf segments
were put inside the tower and conidia from heavily infected leaves were blown into
the tower using compressed air and were allowed to settle for 10 minutes. The
density of inoculum was monitored by haemocytometer and was adjusted to 10-15
conidia per mm2. Following inoculation, the closed petri dishes were incubated in a
growth chamber at 20 oC with fluorescent light irradiance (102.8 micromol/m2/s)
supplied for 12 h per day. Four days after incubation, the accessions were scored by
counting the number of young powdery mildew colonies per 2 cm2 of leaf segment,
using a magnifying glass (10x). The experiment was repeated three times and the
average infection frequency used for QTL analysis. Because L94 is a mlo-barley and
fully resistant to powdery mildew, in our inoculation experiment we only included non-
mlo carrying RILs (51 accessions) of the LnVa population (Shtaya et al., 2006).
Histology of infection by powdery mildew To determine the mechanism of resistance at the cellular level, one DH line or RIL
from each mapping population showing high levels of basal resistance at seedling
stage together with the parents were selected for microscopic observation. The
susceptible barley cultivar ‘Manchuria’ was included as reference. From each
accession two seedlings were grown in boxes (37 × 39 cm). Twelve days after
sowing, first formed leaves were fixed in a horizontal position, with the adaxial side
up. The inoculation was performed using the settling tower with freshly produced
spores collected from young susceptible barley plants. The inoculum density was
adjusted to 10-15 conidia per mm2. In each inoculation, one seedling from each
accession was sampled for histological analysis and the other one was kept for

Which candidate genes are….
23
checking the efficiency of inoculation. The experiment was performed in two
replications. Forty eight hours after inoculation (hai) a leaf segment from the
inoculated leaves was cut and transferred to a solution of 1 mg.ml−1 3,3'-
Diaminobenzidine (DAB) for 8 h and, subsequently, to a solution of acetic acid-
ethanol (3/1 v/v) to be cleared overnight. The leaf segments were stained with
0.005% Trypan Blue as described by Anker & Niks (2001). To make the preparations,
the leaf segments were embedded adaxial side up in glycerol. Leaf segments were
examined at 400x magnification using an Axiophot microscope (Zeiss, Germany). In
all microscopic observations only interstomatal (type B epidermal cells, following the
terminology proposed by Koga et al., 1990) were examined. Those germlings that
were stopped by papillae at penetration stage were considered as failed primary
penetration (FPP) and germlings that formed a haustorium and elongated secondary
hyphae (ESH) were considered as successful primary penetration (SPP) (Asher &
Thomas, 1983). Those epidermal cells that were challenged by more than one
conidium were not considered.
Phenotypic assessment at adult plant stage Evaluation of basal resistance at adult plant stage was carried out at field conditions
under naturally occurring powdery mildew infections. The parents and lines of
different mapping populations were sown in three replications according to a
randomized complete block design in spring of 2006 and 2007. Each genotype was
sown in three-row plots of one meter long, row distance 25 cm. To minimize the effect
of interplot interference (Parlevliet & Van Ommeren, 1984; Lipps & Madden, 1992),
three rows of oat were sown between two neighbouring genotypes. The plots were
treated following standard agricultural practices. Mildew severity was recorded two or
three times with 8-10 days interval according to the 0-9 scale (0= no infection and 9=
fully covered by mildew) (Saari & Prescott, 1975). The area under the disease-
progress curve (AUDPC) was calculated (Jeger & Viljanen-Rollinson, 2001) and used
as quantitative trait for QTL analysis. For the CCSu population, in the 2006 field test,
disease severity was recorded only once and directly used for QTL analysis. For
NuTr, the data were collected from 2 × 3 meter yield trial plots of 88 spring lines
during the spring of 2005 and 2006, without alternating oat plots. The possible effects
of interplot interference were minimized by sampling only the middle rows of the plots.
QTL analysis The quantitative resistance data were combined with the already available biparental
marker map data and analyzed with the software package MapQTL version 5 (Van

Chapter 2
24
Ooijen, 2004). First an interval mapping (IM) analysis was performed. Markers with
the highest log-likelihood ratios (LOD value) were selected as the initial set of
possible co-factors for multiple-QTL mapping (MQM) analysis (Jansen, 1993; Jansen
& Stam, 1994). The LOD threshold for detecting QTLs was calculated by a
permutation test with 10000 iterations at the significance level α = 0.05. A threshold
LOD value above or equal to the LOD value suggested by the permutation test was
used to declare the presence of a QTL. Detected QTLs were transferred from the
individual maps to the barley integrated map. QTLs were named as ‘Rbgq’
(quantitative genes for Resistance to Blumeria graminis) followed by the QTL number
as suggested by Shtaya et al., (2006). A common name is given to QTLs for which
the LOD-1 interval was overlapping.
Mapping candidate genes and their co-localization w ith QTLs Thirty four EST-derived sequences were provided by Dr. Patrick Schweizer (The
Leibniz Institute of Plant Genetics and Crop Plant Research, IPK, Germany) and 5
sequences by Dr. Ralph Hückelhoven (Technical University of Munich, Germany).
These ESTs represent candidate genes implicated in basal resistance, since they
promoted or decreased haustorium formation by Blumeria graminis upon RNAi
silencing or over-expression. We tried to convert those sequences to polymorphic
PCR markers to map them on one of the barley mapping populations used to
construct the integrated map. Primer pairs were designed using DNASTAR software
package based on the full-coding-sequence available or using databases of genomic
or cDNA sequences (CR-EST: The IPK Crop EST Database; The National Centre for
Biotechnology Information (NCBI), http://www.ncbi.nlm.nih.gov/ and The TIGR Gene
Index at DFCI). EST sequences were converted to Cleaved Amplified Polymorphic
Sequence (CAPS), Derived Cleaved Amplified Polymorphic Sequences (dCAPS) or
Sequence Characterized Amplified Region (SCAR) markers to be mapped in one of
the four mapping populations: CCSu, OWBs, StMo and LnVa.
The candidate gene based markers were placed on the integrated map of barley to
be compared with the position of QTLs for resistance to powdery mildew. We
considered mlo and the two genes that are required for the mlo-mediated resistance,
Ror1 and Ror2, as candidate genes to compare their map positions with QTLs. mlo
gene was already mapped directly in the LnVa population and was present in our
integrated map. We located the Ror genes on the map based on the position of
reported co-segregating markers (Collins et al., 2001; Collins et al., 2003). To
determine the global significance of co-localizations between QTLs and candidate
genes, a chi-square test was performed. The null hypothesis of independent

Which candidate genes are….
25
distribution of 5 cM BINs containing a QTL peak marker and BINs containing a
candidate gene was tested.
Comparing the map position of QTLs with known R gen es The Mla6 and MlLa genes were already located on the integrated map based on the
marker data of IgFr and LnVa populations. Other R genes (Mlra, Mlk, Mlnn, Mlga,
Mlg, Mlj, mlt, Mlf) were located on the map based on the reported position of co-
segregating markers (Schonfeld et al., 1996; Kurth et al., 2001; Jahoor et al., 2004).
Results
The new barley integrated map The new barley integrated map contains 6,990 morphological/molecular markers
covering a total genetic distance of 1,093 cM, which is similar to previously reported
barley integrated genetic maps (Table 2). The framework map contained 94 (for
chromosome 1H), 149 (2H), 131 (3H), 85 (4H), 126 (5H), 116 (6H) and 122 (7H) (=
823 in total) bridge markers and covers a genetic distance of 1,051 cM. The most
represented types of molecular markers on this integrated map are RFLP (20%),
AFLP (20%), SSR (9%), DArT (19%) and TDM (23%) (= 91% of all markers). RFLP
and AFLP marker techniques are not used so commonly anymore but the integration
of such markers with more modern types of markers allow to integrate past and
present data collected on many different mapping populations as well as to select
markers or candidate genes linked to a specific trait in any of these populations. More
than 43% of the markers on the present barley integrated map target ESTs or gene
sequences (gene-targeted markers, GTM) and can be used for candidate gene
identification (Rostoks et al., 2005; Marcel et al., 2007a) or to perform synteny
analyses (Rostoks et al., 2005; Stein et al., 2007). The StMo map contributed more
than half of the total number of markers (3,561 markers) on the integrated map, most
of these markers coming from the recently released array-based marker data sets
(Wenzl et al., 2006; Potokina et al., 2008). The very low percentage of errors and
high quality of these marker data greatly increase the reliability of our integrated map.
The present integrated linkage map was aligned with the one of Marcel et al., (2007a)
and with the TDM-based map of Potokina et al., (2008) in order to compare their
locus order. The marker order between the present integrated map and the TDM-
based map was highly consistent, with only a few and very localized inversions
pinpointing small inaccuracies inherent to map integration projects (Jackson et al.,
2008). The marker order was also consistent with the integrated map of Marcel et al.,
(2007a) with the exception of reordering between several AFLP markers mapped at

Chapter 2
26
the telomeres of chromosomes 5HL and 6HL in the CCSu population. The inspection
of graphical genotypes mostly agreed with the marker order on the present integrated
map. The constructed barley integrated map has been posted on the GrainGenes
website (Barley, Integrated 2008, Marcel" at http://wheat.pw.usda.gov/).
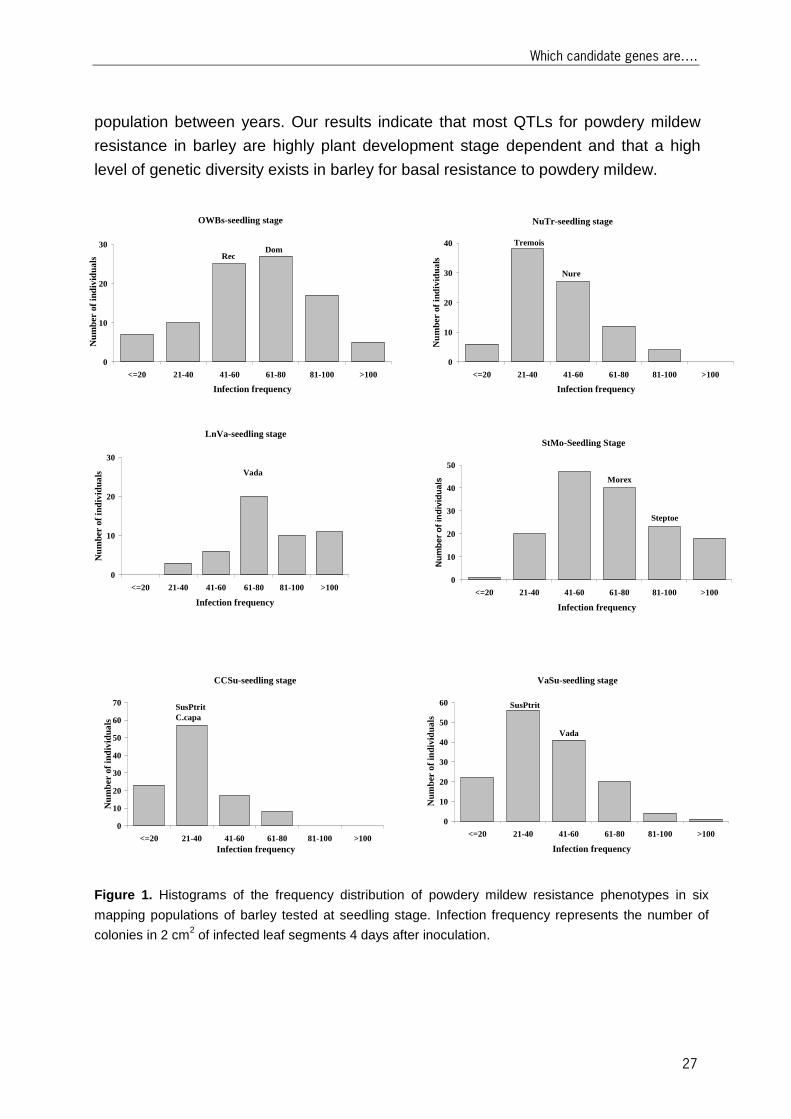
QTLs mapped at seedling stage The infection frequency of powdery mildew in detached seedling leaf tests exhibited
continuous variation in all six mapping populations indicating quantitative inheritance.
In all tested mapping populations, transgressive segregation towards resistance and
towards susceptibility was observed (Figure 1), indicating that the resistance and
susceptibility alleles were contributed by both parents.
The number, genomic position on the barley integrated map, and effect of QTLs
associated with basal resistance to powdery mildew at seedling stage are
summarized in Table 3 and Figure 2. In total, 12 QTLs were mapped that affect the
infection frequency at seedling stage in six mapping populations. The mapped QTLs
were found on all chromosomes. One QTL on chromosome 5H (Rbgq15) was in
common between VaSu and LnVa populations. The phenotypic variation explained by
the QTLs ranged from 6.2-24.8%. The LOD scores of QTLs were generally low
(≤6.1). Our results indicate a high level of diversity in QTLs to be involved in powdery
mildew resistance at seedling stage in barley.
QTLs mapped at adult plant stage The RIL and DH populations showed a continuous distribution for AUDPC or disease
severity data collected at field experiments. Transgressive segregation was common
in four out of five (StMo, no transgression) populations tested (Figure 3). In total, 13
QTLs for adult plant resistance, including QTLs mapped in different populations at the
same location, were detected and these were distributed over all barley
chromosomes (Table 3 and Figure 2). Three out of the 13 QTLs (Rbgq7, Rbgq8 and
Rbgq9) were mapped in more than one population. Four QTLs were detected in both
developmental stages (Rbgq2a, Rbgq7, Rbgq9, and Rbgq13) of which two were
detected in the same population at seedling and adult plant stages. The phenotypic
variation explained by the QTLs ranged from 7.3% to 52.1%, indicating great
differences in effect size of QTLs conferring resistance at adult plant stage. Four
QTLs (Rbgq12, Rbgq4, Rbgq7 and Rbgq8) were detected and confirmed in both
years. The most consistent QTLs were also the ones with the highest LOD scores
(Table 3).The inconsistency of the other QTLs may be due to a high level of
environment dependency or due to different race composition of the pathogen
Which candidate genes are….
27
population between years. Our results indicate that most QTLs for powdery mildew
resistance in barley are highly plant development stage dependent and that a high
level of genetic diversity exists in barley for basal resistance to powdery mildew.
Figure 1. Histograms of the frequency distribution of powdery mildew resistance phenotypes in six
mapping populations of barley tested at seedling stage. Infection frequency represents the number of
colonies in 2 cm2 of infected leaf segments 4 days after inoculation.
OWBs-seedling stage
0
10
20
30
<=20 21-40 41-60 61-80 81-100 >100
Infection frequency
Num
ber
of in
divi
dual
s RecDom
NuTr-seedling stage
0
10
20
30
40
<=20 21-40 41-60 61-80 81-100 >100
Infection frequency
Num
ber
of in
divi
dual
s
Nure
Tremois
CCSu-seedling stage
0
10
20
30
40
50
60
70
<=20 21-40 41-60 61-80 81-100 >100Infection frequency
Num
ber
of in
divi
dual
s
SusPtritC.capa
VaSu-seedling stage
0
10
20
30
40
50
60
<=20 21-40 41-60 61-80 81-100 >100
Infection frequency
Num
ber
of in
divi
dual
s
SusPtrit
Vada
LnVa-seedling stage
0
10
20
30
<=20 21-40 41-60 61-80 81-100 >100
Infection frequency
Num
ber
of in
divi
dual
s Vada
StMo-Seedling Stage
0
10
20
30
40
50
<=20 21-40 41-60 61-80 81-100 >100
Infection frequency
Num
ber
of in
divi
dual
s
Morex
Steptoe

Chapter 2
28
Table 3. Characteristics of QTLs mapped in six barley populations against powdery mildew (Blumeria graminis f.sp. hordei) QTL
Name
Growth
stage
Population Chrom. Peak marker
(PM)
Position
of PM
LOD % exp. a Add. b Donor c
Rbgq4 Field2005 NuTr 1H bPb-4657 23.8 7.2 28.4 10.05 Tremois
Rbgq8 Field2005 NuTr 2H bPb-2230 54.3 6.0 22.1 9.10 Tremois
Rbgq12 Field2005 NuTr 4H E39M61-181 71.6 6.3 31.2 10.66 Tremois
Rbgq4 Field2006 NuTr 1H bPb-4657 23.8 9.6 37.3 6.35 Tremois
Rbgq7 Field2006 LnVa 2H E41M32-83 32.3 7.3 52.1 17.08 Vada
Rbgq21 Field2006 VaSu 7H P15M47-184 40.9 3.3 8.6 0.58 SusPtrit
Rbgq8 Field2006 LnVa 2H E33M54-187 57.0 3.3 27.5 11.86 Vada
Rbgq18 Field2006 CCSu 6H E33M55-333 58.5 2.8 10.4 0.37 Cebada Capa
Rbgq8 Field2006 NuTr 2H bPb-6881 59.2 6.0 19.1 4.66 Tremois
Rbgq8 Field2006 VaSu 2H E33M54-182 61.1 7.7 20.0 0.85 Vada
Rbgq12 Field2006 NuTr 4H E39M61-181 71.6 4.9 37.1 6.34 Tremois
Rbgq9 Field2006 LnVa 2H E38M54-294 147.6 4.7 34.7 13.03 Vada
Rbgq9 Field2006 VaSu 2H P17M54-152 148.4 3.6 9.9 0.60 Vada
Rbgq7 Field2007 LnVa 2H E41M32-83 32.3 3.1 16.1 3.40 Vada
Rbgq7 Field2007 StMo 2H MWG858 37.8 8.7 17.2 2.33 Steptoe
Rbgq5 Field2007 StMo 1H ABG053 43.1 5.7 10.9 1.89 Morex
Rbgq8 Field2007 CCSu 2H E38M54-169 54.3 3.4 14.3 4.00 Cebada Capa
Rbgq8 Field2007 VaSu 2H E38M54-169 54.3 13.3 31.1 4.08 Vada

Which candidate genes are….
29
Table 3 continued
QTL
Name
Growth
stage Population Chrom.
Peak marker
(PM)
Position of
PM
LOD % exp. a Add. b Donor c
Rbgq2a Field2007 LnVa 3H E33M61-131 63.8 4.3 24.9 4.27 L94
Rbgq11 Field2007 StMo 3H CDO113B 106.7 3.6 7.3 1.49 Morex
Rbgq13 Field2007 VaSu 4H E41M40-79 138.3 3.9 7.9 2.07 SusPtrit
Rbgq16 Field2007 StMo 5H CDO484 161.1 4.2 7.6 1.55 Morex
Rbgq17 Field2007 VaSu 5H E33M61-144 178.0 4.8 9.5 2.22 SusPtrit
Rbgq20 Seedling OWBs 7H ABG704 7.4 3.5 18.3 11.12 Rec
Rbgq10 Seedling StMo 3H MWG571C 16.4 4.7 15.5 9.90 Morex
Rbgq7 Seedling CCSu 2H E37M32-288 25.5 3.8 14.4 6.24 SusPtrit
Rbgq14 Seedling OWBs 5H Bmac0047e 44.9 4.9 19.9 11.40 Dom
Rbgq2a Seedling NuTr 3H bPb-0312 47.7 6.1 24.8 9.36 Tremois
Rbgq22 Seedling CCSu 7H E38M61-128 48.7 4.2 20.0 8.68 SusPtrit
Rbgq6 Seedling VaSu 1H E38M54-441 61.5 3.4 8.0 6.07 SusPtrit
Rbgq2b Seedling LnVa 3H E35M48-410a 69.5 2.8 19.8 9.98 L94
Rbgq19 Seedling NuTr 6H scssr05599 96.9 5.6 22.4 8.76 Nure
Rbgq15 Seedling VaSu 5H E42M48-282 115.4 2.7 6.2 5.18 SusPtrit
Rbgq15 Seedling LnVa 5H E33M58-306 117.6 3.1 21.5 10.71 Vada
Rbgq13 Seedling VaSu 4H E35M58-56 144.0 4.2 9.8 6.53 SusPtrit
Rbgq9 Seedling VaSu 2H E42M48-405 146.4 5.0 11.5 6.97 Vada
a The proportion of the phenotypic variance explained by the QT b Effects of the resistant allele in reduction of the number of colonies per 2 cm2; c Donor of the resistance allele

Chapter 2
30
*BCD1434
*ABA305
*E42M51-113GBM1447*MWG938Mlra
*cMWG645
*MWG837Mla6Mlk*GBM1007
*ABA004
*GBR1154
*BCD098
*ABG053Mlnn
*Ica1
*E35M61-92
*JS074GBM1412Ror1GBM1487*Pcr2GBM1336GBM1216GBM1480*bPb-1193*Glb1GBM1153*GBM1108WBE223*DAK123BMlgaGBM1481*GBR0643*PSR330*Bmac0144aGBM5162*cMWG649B
*cMWG706A
*ABC257
*BCD193
*GBR1418
*ABG702
*ABC261WBE221GBM1308*GBR0828GBM1358GBM1407*GBR1390GBM1314
Rbgq4(Trem
ois)_7.2R
bgq6(SusPtrit)_3.4
Rbgq4(Trem
ois)_9.6
Rbgq5(M
orex)_5.7
1H*MWG844A
*GBR0613
*ABG703B
*E37M33-160
*MWG878
*GBS0535
GBM5023GBM5018*ABG318*E33M61-135GBM5140*ABG358*GBR1550WBE226*PoxGBM5095*E38M54-169GBM1446*Bgq60GBM1158GBM1172GBM1203GBM1366GBM1129*GBM5230WBE219*ABC468WBE231*GBMS160GBM1513*ABC451*MWG865GBM1468GBM1408*Bmag0125WBE229*MWG503GBM1328GBM1440*MWG882*GBM1208*ksuD22GBM1469*E33M54-307
*GBS0379
GBM1437*ABC252GBM1200
*GBM1047
*ABC165A
WBE220*E35M61-358
*MWG844C
MlLa*MWG2200
GBM5080
Rbgq7(SusP
trit)_3.8R
bgq8(C.C
apa)_3.4R
bgq9(Vada)_5.0
Rbgq7(V
ada)_3.1R
bgq8(Vada)_13.3
Rbgq7(Steptoe)_8.7
Rbgq8(T
remois)_6.0
Rbgq7(V
ada)_7.3R
bgq8(Vada)_3.3
Rbgq9(V
ada)_4.7
Rbgq8(T
remois)_6.0
Rbgq9(V
ada)_3.6
Rbgq8(V
ada)_7.7
2H*BCD907
WBE212*ABG070
*JS195F
*E35M48-250
*E42M55-233
*ABG321
GBM1430*MWG798B
*MWG584GBM1284
*BCD153
*E37M32-471
*ABG396
*GBM1372
*MWG571BGBM1163
*ABC176GBM5020GBM1343*ABG377WBE232Contig7079_atContig7080_atGBM1090GBM5047*MWG555B*ABG453*ABR320*CDO345*bPt-7238
*CDO113B
*GBS0510
*His4B
GBM1226*MWG847
*ABG004
*WG110
*ABC161GBM1118
*MWG902WBE211*ABC174WBE225GBM1288WBE218*Adh10
*ABC166
*GBR1425
*ABC172GBM1420
Rbgq10(M
orex)_4.7R
bgq2a(Trem
ois)_6.1R
bgq2b(L94)_2.8
Rbgq2a(L
94)_4.3R
bgq11(Morex)_3.6
3H0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
105
110
115
120
125
130
135
140
145
150
155
160
165
170
175
180
Figure 2. Localization of candidate gene markers and QTLs for resistance to powdery mildew on an integrated
map of barley. The candidate genes, including mlo, Ror1 and Ror2 genes, are in bold red. The locations of QTLs
are indicated by vertical bars at the right side of each linkage group. Solid bars represent QTLs identified at
seedling stage under controlled conditions, and shaded bars represent QTLs identified at adult plant stage under
field conditions. The length of a bar approximately represents the LOD-1 support interval and the length of the
exceeding line the LOD-2 support interval of the QTL based on rMQM results. The ruler indicates the distance in
cM. (Continued on next page)

Which candidate genes are….
31
*bPt-7602
*bPt-7852
*MWG634
*E35M54-548
*JS103.3
*HVM40
GBM1252*Ole1*LoxA
*BCD402B
*HVKNOX3
*BCD808B
Mlg*ABC303*ABG484GBM1364*PBI25
*bBE54A
*Bmac0310GBM5210*BCD453B
*GBM1350
*ABG319A
*E39M61-367
GBM1249*KFP221
WBE216*bPb-1325mlo*ABG397
*GBM1448
WBE214*ABG319C
*MWG2037BGBM1324GBM1147WBE104*E42M58-312GBM1453*GBM1018
Rbgq12(T
remois)_6.3
Rbgq13(SusP
trit)_4.2
Rbgq12(T
remois)_4.9
Rbgq13(SusP
trit)_3.9
4H*bPb-2330
*DAK133
*ABR313
*MWG920-1A
*bPb-0091
*scssr07106*E45M55-400GBM5028*ABG705WBE227*CDO669BContig5975_atContig5974_s_atWBE100*ABG395GBM1478*E32M62-321GBM1222GBM1325GBM1508*Ltp1*GBMS196GBM1293*WG530*E35M55-181*GBS0042*ABC302GBM1506
*E38M55-128
*BCD926
*MWG522
*ABG473Mlj*GBM1227GBR0180*E42M51-335*MWG2230GBM1295GBM1141*GBS0531WBE228GBM5008*MWG514BGBM5222GBM5212GBM1384GBM5214*GBS0138*WG908GBM1166Ror2*E42M55-75
*ABG496GBM1490
*GBS0800
*ABG391
*ABG390
*DsT-33
*ABG463
*ABC309GBM1474*MWG851B
Rbgq14(D
om)_4.9
Rbgq15(SusP
trit)_2.7R
bgq16(Morex)_4.2
Rbgq15(V
ada)_3.1R
bgq17(SusPtrit)_4.8
5H*ABG062
*ABG466
*ABG378
*E42M54-160
*E33M61-401
*cMWG652A
*E33M55-63
GBM1270*DD1.1C
*ABG387B*E35M61-269GBM5086GBR0504*GBM1053*iLdh1GBM1267WBE215GBM5012*GBS0468*ABG474
*GBMS180
*ABC170B
*GBS0767
*Nar7
*GBR0621
*MWG934
*Tef1
*scssr00103
*Bmac0040a
*P16M51-215GBM1154GBM5064*GBM1274GBM1275GBM1276*MWG2196*cMWG684A
Rbgq19(N
ure)_5.6
Rbgq18(C
.Capa)_2.8
6H*E37M50-401
*ABG704
mlt*bPt-3623
*ABG320
WBE224*P14M61-73
*ABC151AWBE217*GBM1326GBM5060*ABG380
*bPt-6944
*ksuA1A
*GBM1464
GBM5226*Brz
*ABC255
*HVCMA
*E33M54-261*ABG701GBM1237*ABC455WBE101Contig2163_atContig2170_atMlf*Amy2GBM1303WBE210GBM1472GBM1297GBM1396GBM1428*GBMS141*RZ242GBM5225*bPb-9908*ABC310B*bPb-9104*ABC305*GBR1547GBM1362WBE230*ABG461A
*GBR1326
*GBM1456
*GBR1567
*bPb-1858
Rbgq20(R
ec)_3.5R
bgq22(SusPtrit)_4.2
Rbgq21(SusP
trit)_3.3
7H0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
105
110
115
120
125
130
135
140
145
150
155
160
165
170
175
180

Chapter 2
32
NuTr-Field 2006
0
10
20
30
40
50
60
70
80
<=50 51-70 71-90 91-110 >110
AUDPC
Num
ber
of in
divi
dual
s
Tremois
StMo-Field 2007
0
10
20
30
40
50
60
70
80
90
<=20 21-30 31-40 41-50 >50
AUDPC
Num
ber
of in
divi
dual
s
Steptoe
Morex
VaSu-Field 2007
0
10
20
30
40
50
60
70
80
90
<=20 21-30 31-40 41-50 >50AUDPC
Num
ber
of in
divi
dual
s VadaSusPtrit
LnVa-Field 2006
0
10
20
30
<=20 21-40 41-60 61-80 81-100 >100AUDPC
Num
ber
of in
divi
dual
s
Vada
CCSu-Field2007
0
10
20
30
40
50
60
70
<=20 21-40 41-60 61-80 81-100 >100
AUDPC
Num
ber
of in
divi
dual
s
SusPtritC.Capa
Figure 3. Histograms of the frequency distribution of powdery mildew resistance phenotypes in 5
mapping populations of barley tested at adult plant stage in field conditions.

Which candidate genes are….
33
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Dom Rec
OW
B 3
4( R
)
Nur
e
Tre
moi
s
NuT
r 12
2( R
)
Step
toe
Mor
ex
StM
o 39
( R)
L94
Vad
a
LnV
a 26
( R
)
SusP
trit
VaS
u 11
1( R
)
CC
apa
CC
Su 9
2( R
)
Man
chur
ia
Inte
ract
ion
site
s
SPP-HR
SPP+HR
FPP
Figure 4. Defense reaction of the parents and most resistant line of different mapping populations to Bgh at 48
hai. Data of each group represent the mean of two leaf segments; in each leaf 100 infection sites have been
evaluated. Gray-dotted column (FPP) represent failed primary penetration where the fungus was stopped at
penetration stage; Black column (SPP+HR), successful primary penetration resulting in a haustorium but showed
HR; White column (SPP-HR), successful primary penetration with haustorium and without HR. The lines with the
(R) represent the most resistant line from each mapping population.
Histology of resistance at seedling stage The reaction to Bgh at 48 hai are presented for the parental lines and for the most
resistant lines of each of the six studied mapping populations in Figure 4. At
macroscopic level, all parental lines, except L94 that carries the mlo gene, showed a
compatible infection type (infection type 4). This implies that they do not carry an
effective race-specific resistance gene (R gene) to the isolate Wag.04 that was used.
The proportion of failed infection units that were stopped at penetration stage (pre-
haustorial resistance) in parental lines ranged from 20% (Rec) to 63% (SusPtrit) of
attempted penetration sites. Always a higher proportion of infection units were
stopped at penetration stage in the most resistant lines rather than in the parents of
the mapping populations. Apart from the mlo-caused pre-haustorial resistance in L94,
the highest proportion of papilla-based resistance (83 %) was in line CCSu (RIL 92).
The proportion of penetrated infection units that were stopped in association with a
hypersensitivity reaction of the challenged epidermal cells ranged from 1% (Rec) to
11% (Vada) of the infection units. These results indicate that the QTLs that were
mapped at the seedling stage should determine papilla-based, non-hypersensitive,
basal or partial resistance.

Chapter 2
34
Candidate gene mapping Twenty three out of 39 EST-derived sequences representing candidate genes were
mapped in at least one of the available mapping populations. Eighteen candidate
genes were converted to CAPS markers. For one candidate gene, a SNP was used
to design a dCAPS marker using the dCAPS Finder online software
(http://helix.wustl.edu/dcaps/dcaps.html). Two sequences were mapped as SCAR
markers and two other sequences were previously mapped as RFLP markers. One
sequence(ID: HO02M14) was polymorphic in the CCSu population but could not be
assigned to a map position with confidence. Fiftheen candidate gene sequences
remained for which no polymorphism was detected or for which it was not possible to
obtain a clear PCR amplification product in any of the parental combinations (ID:
HO14H18, HO14K08, HO16C19, HO26F22, HO13M13, HZ40O15, HI05K17,
HO15M05, HO13K06, HW03O11, HU02G09, AJ344223, AJ518932, AJ518933,
AM265370). The position of the mlo gene was determined directly in the LnVa map
on chromosome 4H (Qi et al., 1998; Shtaya et al., 2006). Based on Collins et al.,
(2001), Ror1 is located within a 0.1 cM distance of ABG452 marker on chromosome
1H. Ror2 was located on chromosome 5H based on its flanking markers as published
by Collins et al., (2003).
Co-localizations between candidate genes and QTLs The Chi-square test applied did not indicate a significant global co-localization
between QTLs and candidate genes (Chi-square value, X2=0.79). Nevertheless, six
out of 26 mapped candidate genes showed coincidence in their map positions with
the QTLs identified for basal resistance to powdery mildew (Figure 2, Table 4). We
defined coincidence of a QTL and a candidate gene, when the candidate gene
mapped within the LOD-1 interval of the QTL in our integrated map. In this paper we
only discuss those candidate genes which are within 5 cM from QTL peak markers
because they are the most relevant ones.
Despite similarities in resistance mechanism between basal resistance and mlo
mediated resistance, none of the detected QTLs showed coincidence with the map
positions of mlo and Ror2 genes. The peak marker of Rbgq6, a QTL mapped in VaSu
population at seedling stage, is located within a 0.1 cM distance of ABG452, a marker
closely linked to Ror1 on chromosome 1H. Shtaya et al., (2006) detected a QTL in
LnVa population in the same region where Ror2 is present.

Which candidate genes are….
35
Co-localizations between QTLs and race-specific res istance genes In most cases, the map positions of powdery mildew resistance QTLs did not coincide
with those of the race-specific resistance genes (R genes). Only five out of 21 QTLs
mapped in the same region where an R gene has been reported. We defined
coincidence of a QTL and an R gene, when the putative map position of the R gene
in our integrated map was located within the LOD-1 support interval of a QTL. Since
the R-genes were mapped mostly in mapping populations that did not contribute to
our integrated map, their map position in our integrated map is more indirectly
obtained, and therefore the cM distance to our QTL peak marker is less precise than
the map positions of the candidate gene-derived markers. A QTL mapped in LnVa
and VaSu populations (Rbgq9) showed coincidence with the known map position of
the MlLa gene, a gene derived from “Hordeum laevigatum” and located on the long
arm of chromosome 2H. This gene causes an intermediate type of hypersensitive
reaction to avirulent isolates (Giese et al., 1993; Marcel et al., 2007b). In both
populations, the resistance QTL allele was contributed by Vada, a cultivar that carries
the MlLa gene. Since the powdery mildew isolate used for evaluation of the
populations at seedling stage was virulent to the MlLa carrying genotypes, it is
plausible that the explained phenotypic variation in this region is due to a QTL and
not to MlLa itself. One QTL detected on chromosome 7H in the OWBs (Rbgq20)
showed coincidence with the map position of mlt. A QTL mapped on chromosome 1H
in NuTr population (Rbgq4) showed coincidence with Mla and Mlk loci. Backes et al.,
(2003) also reported co-localization of QTLs mapped in the cross Vada × 1B-87 on
chromosomes 1H and 7H with the Mla and Mlf loci, respectively. A QTL mapped in
the StMo population (Rbgq5) on chromosome 1H showed coincidence with the map
position of Mlnn gene. Finally, a QTL mapped in the NuTr population (Rbgq12) on
chromosome 4H showed coincidence with the map position of Mlg. But at the end, in
the case of co-localizations between QTLs and R genes, only fine-mapping of the
concerned loci can tell whether the phenotypic variation is due to the R gene or not.

Chapter 2
36
1Regulation in barley leaves inoculated with Blumeria graminis ff.ssp.;
2RNAi effect, the effect of transient-induced gene silencing (TIGS) of test sequence on resistance phenotype
in the single-cell system; OEX effect, the effect of single-cell transient over-expression of test sequence on resistance phenotype in the single-cell system; 3Regulation by Bgh or
stem-rust inoculation in experiments BB2, BB4, BB7 or BB49 of Plant Expression Database (PlexDB) .Corresponding probe-set IDs: HV02H04, Contig12036_at; L37358, HI02E21u_s_at; HY07A14, Contig15618_at; AJ534447, Contig11241_at.;
4 Not significant;
5 Not analyzed
Table 4 . Characterization and map position of ESTs or cDNA-clone representing candidate genes for powdery mildew resistance in barley
Clone
ID/Acc.
Regul. 1 OEX or
RNAi
effect 2
Function References Name of
marker
Position
Chrom./cM
Co-localization
with QTL(s)
HY05J20 up yes Bax-inhibitor 1 Hückelhoven et al., 2003 GBR0504 6H/ 51.6
HO08H12 up yes HvGER4d Zimmermann et al., 2006 WBE104 4H/ 135.8 Rbgq13
HW05P16 up yes HvGER5a Zimmermann et al., 2006 GBR0180 5H/ 97.4
HO06J21 up n.a. WIR1a Schweizer et al., 1999 WBE101 7H/ 83.1
HO09K14 up n.a. WIR1b Zierold et al., 2005 WBE100 5H/ 35.5
DQ647621 up yes HvGER3a Zimmermann et al., 2006 WBE210 7H/ 88.6
HO02P09 up n.a. unknown Zierold et al., 2005 WBE216 4H/ 108.5
HO07K06 up n.a. unknown Zierold et al., 2005 WBE218 3H/ 149.0
HO11O12 up n.a. unknown Zierold et al., 2005 WBE224 7H/ 23.0
HO13M20 up n.a. unknown Zierold et al., 2005 WBE212 3H/ 11.8 Rbgq10
HO14K22 up yes Receptor protein kinase-like Zierold et al., 2005 WBE211 3H/ 142.7
HO14P02 up n.a. Receptor protein kinase-like Zierold et al., 2005 WBE220 2H/ 136.2
HO12E03 up n.a. unknown Zierold et al., 2005 WBE217 7H/ 33.0
HO07H15 up n.a. 4-Coumarate coenzyme A ligase Zierold et al., 2005 WBE215 6H/ 59.8 Rbgq18
HO40A01 up n.a. Hydrolase Zierold et al., 2005 WBE214 4H/ 126.1
HO12F09 up yes SNAP-34 Douchkov et al., 2005; Jensen et al., 2007 WBE219 2H/ 63.6 Rbgq8
HO06D23 up yes P-type ATPase Zierold et al., 2005 WBE223 1H/ 79.8
HvPrx40 up yes Peroxidase Johrde et al., 2008 WBE232 3H/ 81.2
L37358 up 3 n.a.5 Lipoxygenase 2 Bachmann et al., 2002
http://www.plexdb.org/index.php
WBE228 5H/ 117.0 Rbgq15
Hv07A14 up 3 n.a.5 ACC synthase http://www.plexdb.org/index.php WBE229 2H/ 83.9
AJ536667 up yes WRKY1 transcription factor Eckey et al., 2004 WBE230 7H/ 123.3
AJ534447 up 3 n.a.5 Apoplastic invertase http://www.plexdb.org/index.php WBE231 2H/ 69.0

Which candidate genes are….
37
Discussion
Diversity of QTLs for powdery mildew resistance in barley Most mapping studies on quantitative, basal resistance, of barley to powdery mildew
have been based on individual populations. Here we studied the genetic variation of
basal resistance using multiple populations to assess also the diversity of the genes
involved in the genetic variation. The QTLs conferring basal resistance were
distributed over all seven chromosomes of barley without obvious clustering. In
different mapping populations different sets of QTLs were involved in basal
resistance. These indicated a great diversity for QTLs (genes) for this trait in barley. A
similar great diversity has been reported in genes conferring basal resistance in some
other barley-pathogen interactions, like barley-Puccinia hordei (Qi et al., 2000),
barley-Rhyncosporium secalis (Zhan et al., 2008), barley-Fusarium (Fusarium head
blight) (Sato et al., 2008) and barley-spot blotch (Bilgic et al., 2005). Qi et al., (2000)
reported, on the basis of two tested mapping populations, that many loci for basal
resistance to leaf rust (Puccinia hordei) should exist on the barley genome. Marcel et
al., (2007a) reported 19 QTLs contributing to the basal resistance of barley to leaf
rust in five mapping populations. They showed that in each mapping population of
barley different sets of QTLs segregate for basal resistance and only few QTLs were
found to be in common in more than two different barley cultivars. QTL analysis for
Fusarium head blight (FHB) resistance in different barley mapping populations
indicate that quantitative resistance of barley to FHB is conditioned by many loci
(Mesfin et al., 2003). QTLs for powdery mildew resistance have been reported in
different mapping populations of barley (e.g. Backes et al., 1995; Backes et al., 1996;
Spaner et al., 1998; Falak et al., 1999; Backes et al., 2003; Von Korff et al., 2005;
Shtaya et al., 2006). We compared the position of the peak marker of the presently
found QTLs with the peak markers of already reported QTLs using our high-density
integrated map of barley. In four cases (Rbgq2a, Rbgq4, Rbgq7 and Rbgq8), the
location of peak marker of QTLs are only about 0-2 cM away from previously reported
QTLs for mildew resistance, and in two cases (Rbgq2b and Rbgq14) the peak
markers are 4-6 cM away. One of these QTLs (Rbgq4) already has been reported in
two different populations of barley, Blenheim × E224, Vada × 1B-87 (Thomas et al.,
1995; Backes et al., 2003). This suggests that some QTLs found in our study might
be the same as already reported QTLs in other genetic backgrounds.
Growth-stage specificity of QTLs for basal resistan ce Growth-stage dependent and independent QTLs both have been reported in many
host-pathogen interactions, including powdery mildew and leaf rust on barley,

Chapter 2
38
septoria blotch on wheat, ascochyta blight on pea, and downy mildew on lettuce (e.g.
Qi et al., 1999; Backes et al., 2003; Eriksen et al., 2003; Prioul et al., 2004; Marcel et
al., 2007a; Zhang et al., 2009). In our study, only 4 out of a total of 21 QTLs were
effective at both growth stages (Rbgq2a, Rbgq7, Rbgq9, and Rbgq13), suggesting
that QTLs for powdery mildew resistance are mainly growth-stage dependent. Similar
results have been reported by Backes et al. (2003). They found that only one out of
five QTLs that mapped in a cross between the barley cultivar ”Vada” and a wild barley
(H. vulgare ssp. spontaneum line ”1B-87”) was growth-stage independent. This is
also in agreement with the report of Mastebroek & Balkema-Boomstra (1991) in which
it is concluded that the phenotypic expression of basal resistance of barley to
powdery mildew is growth-stage dependent. Our histological observations confirm
that the QTLs mapped in the present work contribute mainly to a prehaustorial non-
hypersensitive basal resistance, rather than to an incomplete hypersensitive
resistance.
Co-localization of candidate genes and R genes with QTL regions In field experiments under natural disease infection, R-gene resistance can behave
as QTL resistance, since 1) the R-genes may have an intermediate effect (like MlLa),
and 2) the natural population of pathogen may consist of a mixture of virulent and
avirulent races to a certain R-gene, resulting in a lower epidemic development on
genotypes carrying the R-gene than in genotypes carrying the susceptibility allele
(Parlevliet, 1983; Parlevliet & van Ommeren, 1985). Furthermore, QTLs that co-
localize with R-genes for resistance may be allelic variants of R-genes or residual
effect of defeated R-genes (Kelly & Vallejo, 2005).
The molecular basis of quantitative (basal) resistance, like other genetically
complex traits, is poorly understood. Candidate gene analysis can be considered as
an important approach toward understanding the molecular basis of quantitative
resistance, partly thanks to the availability of huge numbers of ESTs in databases. In
the present study, we found a co-localization of QTLs for resistance with genes that
have been suggested by reverse genetics to be involved in barley-powdery mildew
interaction. Six out of 26 mapped candidate genes showed coincidence with the
position of QTLs for powdery mildew resistance (Figure 2). Transient-induced gene
silencing (TIGS) experiments showed that a t-SNARE protein, HvSNAP34, is involved
in three types of durable, non-race specific resistances, viz. mlo-mediated, non-host
and basal resistance (Douchkov et al., 2005). Map co-localization of HvSNAP34 with
a QTL (Rbgq8) for powdery mildew resistance provides further evidence of the role of
this gene in basal resistance. An other QTL (Rbgq7) co-localized with the known map

Which candidate genes are….
39
position of Ror1 on chromosome 1H. None of the QTLs co-localized with mlo and
Ror2 genes. We conclude that, despite the similarity in resistance mechanism, there
is little evidence that basal resistance is determined by allelic variants of the Mlo or
Ror2 genes.
Map co-localization between resistance gene analogs (RGA) (see Glossary) and
defense-related (DR) genes with QTLs for seedling and/or adult plant resistance have
been reported in many other host-pathogen systems (Faris et al.1999; Geffroy et al.,
2000; Wang et al., 2001; Prioul-Gervais et al., 2007; Marcel et al., 2007a). Hu et al.,
(2008) established a candidate gene strategy that integrates linkage map, expression
profile and functional complementation analyses with the purpose of cloning QTLs
that are responsible for quantitative resistance of rice to bacterial blight and fungal
blast. Using more than one mapping population, however, provides an impression on
the diversity of the genes responsible for basal resistance. With a larger number of
mapping populations and QTLs detected, it is possible to test a larger sample of
QTLs for their statistical coincidence with the candidate genes. The use of an
integrated map to compare data obtained in different populations also increases the
chance of discovering relevant co-localizations. Here we prioritized the candidate
genes by accumulating evidence of position, biological function and functional proof
by TIGS. However, map co-localization of candidate genes with the QTLs involved in
the trait variation is not a sufficient proof of causal relationship, and actual
involvement of the candidate genes in the trait variation should be validated by
complementation tests through genetic transformation and/or physiological analyses
of genes at the mRNA and/or the protein level and by ultra-fine-mapping of both the
candidatee gene and the QTL (Pflieger et al., 2001). The integrated candidate gene
strategy has proven to be a fast and efficient approach for isolation of genes with
small effect (Hu et al., 2008). Information gained from this approach may help
breeders to use more efficiently the valuable sources of diversity to improve the level
of basal resistance in crop plants.
Acknowledgements
Reza Aghnoum was supported by the Agricultural Research and Education Organization (AREO) and the Ministry of Science Research and Technology of Iran. The excellent technical assistance of Anton Vels in greenhouse and field experiments is greatly appreciated. We are indebted to Dr. Ralph Hückelhoven (Technical University of Munich, Germany) for sharing five sequences of candidate genes; to Christine Boyd and Dr. Andris Kleinhofs from the Washington State University for making available the segregation data of many BIN markers; to Prof. Fred van Eeuwijk for allowing us to collect powdery mildew data from his field experiments of the NuTr population; to Dr. Rajeev K. Varshney from ICRISAT for providing primer aliquots and markers information on the new set of EST-SSR markers. Those EST-SSR markers have been developed at IPK in Germany. Marc-Antoine

Chapter 2
40
Bonvoisin from ESITPA in France is acknowledged for his hard work on mapping this new set of EST-SSR markers. We gratefully acknowledge the critical reading of the manuscript by Prof. Richard Visser.
References
Anker CC, Niks RE. 2001. Prehaustorial resistance to the wheat leaf rust fungus, Puccinia triticina, in
Triticum monococcum (s.s.). Euphytica 117: 209–215.
Asher MJC, Thomas CE. 1983. The expression of partial resistance to Erysiphe graminis in spring
barley. Plant Pathology 32: 79-89.
Asher MJC, Thomas CE. 1984. Components of partial resistance to Erysiphe graminis in spring
barley. Plant Pathology 33: 123–130.
Asher MJC, Thomas CE. 1987. The inheritance of mechanisms of partial resistance to Erysiphe
graminis in spring barley. Plant Pathology 36: 66-72.
Bachmann A, Hause B, Maucher H, GarbeE, Vörös K, We ichert H, Wasternack C, Feussner I.
2002. Jasmonate-induced lipid peroxidation in barley leaves initiated by distinct 13-LOX forms of
chloroplasts. Biological Chemistry 383: 1645-1657.
Backes G, Graner A, Foroughi-Wehr B, Fischbeck G, W enzel G, Jahoor A. 1995. Localization of
quantitative trait loci (QTLs) for agronomic important characters by the use of a RFLP map in
barley (Hordeum vulgare L.). Theoretical and Applied Genetics 90: 294– 302.
Backes G, Madsen LH, Jaiser H, Stougaard J, Herz M, Mohler V, Jahoor A. 2003. Localization of
genes for resistance against Blumeria graminis f.sp hordei and Puccinia graminis in a cross
between a barley cultivar and a wild barley (Hordeum vulgare ssp spontaneum) line. Theoretical
and Applied Genetics 106: 353-362.
Backes G, Schwarz G, Wenzel G, Jahoor A. 1996. Comparison between QTL analyses on powdery
mildew resistance in barley based on detached primary leaves and on field data. Plant Breeding
115: 419–421.
Bilgic H, Steffenson BJ, Hayes PM. 2005. Comprehensive genetic analyses reveal differential
expression of spot blotch resistance in four populations of barley. Theoretical and Applied
Genetics 111: 1238–1250.
Carver TLW. 1986. Histology of infection by Erysiphe graminis f. sp. hordei in spring barley lines with
various levels of partial resistance. Plant Pathology 35: 232–240.
Chelkowski j, Tyrka M, Sobkiewicz A. 2003 . Resistance genes in barley (Hordeum vulgare L.) and
their identification with molecular markers. Journal of Applied Genetics 44: 291-309.
Collins NC, Lahaye T, Peterhansel C, Freialdenhoven A, Corbitt M, Schulze-Lefert P. 2001.
Sequence haplotypes revealed by sequence-tagged site fine mapping of the Ror1 gene in the
centromeric region of barley chromosome 1H. Plant Physiology 125: 1236–1247.
Collins NC, Sadanandom A, Schulze-Lefert P. 2002. Genes and molecular mechanisms controlling
powdery mildew resistance in barley. In: Bélanger RR, Bushnell WR, Dik AJ, Carver TLW, eds.
The powdery mildews: A comprehensive treatise. St. Paul/Minnesota, USA: APS Press, 134-145.
Collins NC, Thordal-Christensen H, Lipka V, Bau S, Kombrink E, Qiu JL, Hückelhoven R, Stein
M, Freialdenhoven A, Somerville SC et al. 2003. SNARE-protein-mediated disease resistance
at the plant cell wall. Nature 425: 973-977.

Which candidate genes are….
41
Costa JM, Corey A, Hayes PM, Jobet C, Kleinhofs A, Kopisch-Obusch A, Kramer SF, Kudrna D,
Li M, Riera-Lizarazu O et al. 2001. Molecular mapping of the Oregon Wolfe Barleys: a
phenotypically polymorphic doubled-haploid population. Theoretical and Applied Genetics 103:
415–424.
Czembor JH. 2000. Resistance to powdery mildew in barley landraces from morocco. Journal of Plant
Pathology 82: 187-200.
Dong WB, Nowara D, Schweizer P. 2006. Protein polyubiquitination plays a role in basal host
resistance of barley. Plant Cell 18: 3321-3331.
Douchkov D, Nowara D, Zierold U, Schweizer P. 2005. A high-throughput gene-silencing system for
the functional assessment of defense-related genes in barley epidermal cells. Molecular Plant-
Microbe Interactions 18: 755-761.
Eckey C, Korell M, Leib K, Biedenkopf D, Jansen C, Langen G, Kogel KH. 2004 . Identification of
powdery mildew-induced barley genes by cDNA-AFLP:functional assessment of an early
expressed MAP kinase. Plant Molecular Biology 55: 1-15.
Eriksen L, Borum F, Jahoor A. 2003. Inheritance and location of resistance to Mycosphaerella
graminicola causing septoria tritici blotch and plant height in the wheat (Triticum aestivum L.)
genome with DNA markers. Theoretical and Applied Genetics 107: 515– 527.
Falak I, Falk DE, Tinker NA, Mather DE. 1999. Resistance to powdery mildew in a doubled haploid
barley population and association with marker loci. Euphytica 107: 185–192.
Faris JD, Li W, Gill BS, Liu D, Chen P. 1999. Candidate gene analysis of quantitative disease
resistance in wheat. Theoretical and Applied Genetics 98: 219–225.
Francia E, Rizza F, Cattivelli L, Stanca AM, Galiba G, Tóth B, Hayes PM, Skinner JS, Pecchioni
N. 2004. Two loci on chromosome 5H determine low-temperature tolerance in a ‘Nure’ (winter) x
‘Tremois’ (spring) barley map. Theoretical and Applied Genetics 108: 670–680.
Freialdenhoven A, Peterhänsel C, Kurth J, Kreuzaler F, Schulze-Lefert P. 1996. Identification of
genes required for the function of non-race specific mlo resistance to powdery mildew in barley.
Plant Cell 8: 5-14.
Geffroy V, Sevignac M, De Oliveira JCF, Fouilloux G , Skroch P, Thoquet P, Gepts P, Langin T,
Dron M. 2000. Inheritance of partial resistance against Colletotrichum lindemuthianum in
Phaseolus vulgaris and co-localization of quantitative trait loci with genes involved in specific
resistance. Molecular Plant-Microbe Interactions 13: 287-296.
Giese H, Holm-Jensen AG, Jensen HP, Jensen J. 1993. Localization of the Laevigatum powdery
mildew resistance to barley chromosome 2 by the use of RFLP markers. Theoretical and Applied
Genetics 85: 897–900.
Graner A, Jahoor A, Schondelmaier J, Siedler H, Pil len K, Fischbeck G, Wenzel G, Herrmann
RG. 1991. Construction of an RFLP map of barley. Theoretical and Applied Genetics 83: 250–
256.
Heun M. 1992. Mapping quantitative powdery mildew resistance of barley using a restriction fragment-
length polymorphism map. Genome 35: 1019–1025.
Hu KM, Qiu DY, Shen XL, Li XH, Wang SP. 2008. Isolation and manipulation of quantitative trait loci
for disease resistance in rice using a candidate gene approach. Molecular Plant 1: 786-793.
Hückelhoven R, Dechert C, Kogel KH. 2003. Overexpression of barley BAX inhibitor 1 induces
breakdown of mlo-mediated penetration resistance to Blumeria graminis. Proceedings of the
National Academy of Sciences of the United States of America 100: 5555-5560.

Chapter 2
42
Ihlow A, Schweizer P, Seiffert U. 2008. A high-throughput screening system for barley/powdery
mildew interactions based on automated analysis of light micrographs. BMC Plant Biology. doi:
10.1186/1471-2229-8-6 Jackson BN, Schnable PS, Aluru S. 2008. Consensus genetic maps as median orders from
inconsistent sources. IEEE/ACM Transactions on Computational Biology and Bioinformatics 5:
161–171.
Jafary H, Szabo LJ, Niks RE. 2006. Innate nonhost immunity in barley to different heterologous rust
fungi is controlled by sets of resistance genes with different and overlapping specificities.
Molecular Plant–Microbe Interactions 19: 1270–1279.
Jafary H, Albertazzi G, Marcel TC, Niks RE. 2008. High diversity of genes for nonhost resistance of
barley to heterologous rust fungi. Genetics 178: 2327–2339.
Jahoor A, Eriksen L, Backes G. 2004. QTLs and genes for disease resistance in barley and wheat.
In: Gupta PK, Varshney RK, eds. Cereal Genomics. The Netherlands: Kluwer Academic
Publishers, 199–251.
Jansen RC, Stam P. 1994. High resolution of quantitative traits into multiple loci via interval mapping.
Genetics 136: 1447-1455.
Jeger J, Viljanen-Rollinson SLH. 2001. The use of area under the disease-progress curve (AUDPC)
to assess quantitative disease resistance in crop cultivars. Theoretical and Applied Genetics 102:
32–40.
Jensen MK, Rung JH, Gregersen PL, Gjetting T, Fugls ang AT, Hansen M, Joehnk N, Lyngkjaer
MF, Collinge DB. 2007. The HvNAC6 transcription factor: a positive regulator of penetration
resistance in barley and Arabidopsis. Plant Molecular Biology 65:137-150.
Johrde A, Schweizer P. 2008. A class III peroxidase specifically expressed in pathogen-attacked
barley epidermis contributes to basal resistance. Molecular Plant Pathology 9: 687-696.
Jones IT, Davies IJER. 1985. Partial resistance to Erysiphe graminis f. sp. hordei in old European
barley varieties. Euphytica 34: 499-507.
Jørgensen H. 1994. Genetics of powdery mildew resistance in barley. Critical Review in Plant
Sciences 13: 97-119.
Kelly JD, Vallejo VA. 2005. QTL analysis of multigenic disease resistance in plant breeding. In Tuzun
S, Bent E, eds. Multigenic and induced systemic resistance in plants. USA: Springer, 21–48.
Kleinhofs A, Graner A. 2001. An integrated map of the barley genome. In: Philips RL, Vasil IK, eds.
DNA-Based Markers in Plants. Dordrecht, The Netherlands: Kluwer Academic Publishers, 187–
199.
Kleinhofs A, Kilian A, Saghai MA, Biyashev RM, Haye s P, Chen FQ, Lapitan N, Fenwick A, Blake
TK, Kanazin V et al. 1993. A molecular, isozyme and morphological map of the barley (Hordeum
vulgare) genome. Theoretical and Applied Genetics 86: 705–712
Knudsen JCN, Dalsgaard HH, Jorgensen JH. 1986. Field assessment of partial resistance to
powdery mildew in spring barley. Euphytica 35: 233–243.
Koga H, Bushnell WR, Zeyen RJ. 1990. Specificity of cell type and timing of events associated with
papilla formation and the hypersensitive reaction in leaves of Hordeum vulgare attacked by
Erysiphe graminis f. sp. hordei. Canadian Journal of Botany 68: 2344–2352.
Kurth J, Kolsch R, Simons V, Schulze-Lefert P. 200 1. A high resolution genetic map and a
diagnostic RFLP marker for the Mlg resistance locus to powdery mildew in barley. Theoretical
and Applied Genetics 102: 53–60.

Which candidate genes are….
43
Lipps PE, Madden LV. 1992. Effects of plot size and border width on assessment of powdery mildew
of winter wheat. Plant Disease 76: 299–303.
Marcel TC, Varshney RK, Barbieri M, Jafary H, de Ko ck MJD, Graner A, Niks RE. 2007a. A high-
density consensus map of barley to compare the distribution of QTLs for partial resistance to
Puccinia hordei and of defence gene homologues. Theoretical and Applied Genetics 114: 487-
500.
Marcel TC, Aghnoum R, Durand J, Varshney RK, Niks R E. 2007b. Dissection of the barley 2L1.0
region carrying the ‘Laevigatum’ quantitative resistance gene to leaf rust using near isogenic lines
(NIL) and subNIL. Molecular Plant–Microbe Interactions 20: 1604–1615.
Mastebroek HD, Balkema-Boomstra AG. 1991. Identification of growth stage dependent expression
of partial resistance of barley to powdery mildew. Euphytica 58: 113-118.
Mesfin A, Smith KP, Dill-Macky R, Evans CK, Waugh R , Gustus CD, Muehlbauer GJ. 2003.
Quantitative trait loci for Fusarium head blight resistance in barley detected in a two-rowed by six-
rowed population. Crop Science 43: 307–318.
Mitchell-Olds T, Schmitt J. 2006. Genetic mechanisms and evolutionary significance of natural
variation in Arabidopsis. Nature, 441: 947-952.
Niks RE, Rubiales D. 2002. Potentially durable resistance mechanisms in plants to specialized fungal
pathogens. Euphytica 124: 201–216.
Parlevliet JE, van Ommeren A. 1984. Interplot interference and the assessment of barley cultivars for
partial resistance to leaf rust, Puccinia hordei. Euphytica 33: 685–697.
Parlevliet JE, van Ommeren A. 1985. Race-specific effects in major genic and polygenic resistance
of barley to leaf rust in the field: identification and distinction. Euphytica 34: 689–695.
Parlevliet JE. 1975. Partial resistance of barley to leaf rust, Puccinia hordei. I. Effect of cultivar and
development stage on latent period. Euphytica 24: 21–27.
Parlevliet JE. 1983. Can horizontal resistance be recognized in the presence of vertical resistance in
plants exposed to a mixture of pathogen races?. Phytopathology 73: 379.
Pflieger S, Lefebvre V, Caranta C, Blattes A, Goffi net B, Palloix A. 1999. Disease resistance gene
analogues as candidates for QTLs involved in pepper-pathogen interactions. Genome 42: 1100–
1110.
Pflieger S, Lefebvre V, Causse M. 2001. The candidate gene approach in plant genetics: a review.
Molecular Breeding 7: 275-291.
Potokina E, Druka A, Luo Z, Wise R, Waugh R, Kearse y M. 2008. Gene expression quantitative trait
locus analysis of 16,000 barley genes reveals a complex pattern of genome-wide transcriptional
regulation. The Plant Journal 53: 90–101.
Prioul S, Frankewitz A, Deniot G, Morin G, Baranger A. 2004. Mapping of quantitative trait loci for
partial resistance to Mycosphaerella pinodes in pea (Pisum sativum L.), at the seedling and adult
plant stages. Theoretical and Applied Genetics 108:1322–1334.
Prioul-Gervais S, Deniot G, Receveur EM, Frankewitz A, Fourmann M, Rameau C, Pilet-Nayel
ML, Baranger A. 2007. Candidate genes for quantitative resistance to Mycosphaerella pinodes
in pea (Pisum sativum L.). Theoretical and Applied Genetics 114: 971–984.
Qi X, Fufa F, Sijtsma D, Niks RE, Lindhout P, Stam P. 2000. The evidence for abundance of QTLs
for partial resistance to Puccinia hordei on the barley genome. Molecular Breeding 6: 1–9.
Qi X, Jiang G, Chen W, Niks RE, Stam P, Lindhout P. 1999. Isolate-specific QTLs for partial
resistance to Puccinia hordei in barley. Theoretical and Applied Genetics 99: 877–884.

Chapter 2
44
Qi X, Stam P, Lindhout P. 1998. Use of locus-specific AFLP markers to construct a high-density
molecular map in barley. Theoretical and Applied Genetics 96: 376–384.
Read BJ, Raman H, McMichael G, Chalmers KJ, Able tt GA, Platz GJ, Raman R, Genger RK,
Boyd WJR, Li CD et al. 2003. Mapping and QTL analysis of the barley population Sloop ×
Halycon. Australian Journal of Agricultural Research 54: 1145–1153.
Rostoks N, Mudie S, Cardle L, Russell J, Ramsay L, Booth A, Svensson JT, Wanamaker EM,
Hedley PE, Liu H et al. 2005. Genome-wide SNP discovery and linkage analysis in barley based
on genes responsive to abiotic stress. Molecular Genetics and Genomics 274: 515–527.
Saari EE, Prescott JM. 1975. A scale for appraising the foliar intensity of wheat disease. Plant
Disease Reporter 59: 377–380.
Sato K, Hori K, Takeda K. 2008. Detection of Fusarium head blight resistance QTLs using five
populations of top-cross progeny derived from two-row × two-row crosses in barley. Molecular
Breeding 22: 517–526.
Schonfeld M, Ragni A, Fischbeck G, Jahoor A. 1996. RFLP mapping of three new loci for
resistance genes to powdery mildew (Erysiphe graminis f sp hordei) in barley. Theoretical and
Applied Genetics 93: 48-56.
Schweizer P, Pokorny J, Abderhalden O, Dudler R. 19 99. A transient assay system for the
functional assessment of defense-related genes in wheat. Molecular Plant-Microbe Interactions
12: 647-654.
Schweizer P. 2007. Nonhost resistance of plants to powdery mildew-New opportunities to unravel the
mystery. Physiological and Molecular Plant Pathology 70: 3–7.
Shtaya MJY, Marcel TC, Sillero JC, Niks RE, Rubiale s D. 2006. Identification of QTLs for powdery
mildew and scald resistance in barley. Euphytica 151: 421-429.
Spaner D, Shugar LP, Choo TM, Falak I, Briggs KG, Legge WG, Falk DE, Ullrich SE, Tinker
NA, Steffenson BJ et al.1998. Mapping of disease resistance loci in barley on the basis of
visual assessment of naturally occurring symptoms. Crop Science 38: 843-850.
Stein N, Graner A. 2004. Map-based gene isolation in cereal genomes. In: Gupta PK, Varshney RK,
eds. Cereal Genomics. Dordrecht, the Netherlands: Kluwer Academic Publishers, 331–360.
Stein N, Prasad M, Scholz U, Thiel T, Zhang H, Wolf M, Kota R, Varshney RK, Perovic D, Grosse
I et al. 2007. A 1,000-loci transcript map of the barley genome: new anchoring points for
integrative grass genomics. Theoretical and Applied Genetics 114: 823–839.
Thomas WTB, Powell W, Waugh R, Chalmers KJ, Barua U M, Jack P, Lea V, Forster BP,
Swanston JS, Ellis RP et al. 1995. Detection of quantitative trait loci for agronomic, yield, grain
and disease characters in spring barley (Hordeum vulgare L.). Theoretical and Applied Genetics
91: 1037–1047.
Trujillo M, Troeger M, Niks RE, Kogel KH, Hückelh oven R. 2004 . Mechanistic and genetic overlap
of barley host and non-host resistance to Blumeria graminis. Molecular Plant Pathology 5: 389-
396.
Van Ooijen JW. 2004. MapQTL® version 5, software for the mapping of quantitative trait loci in
experimental populations. Wageningen, the Netherlands: Kyazma BV.
Varshney RK, Marcel TC, Ramsay L, Russell J, Röder M, Stein N, Waugh R, Langridge P, Niks
RE, Graner A. 2007. A high density barley microsatellite consensus map with 775 SSR loci.
Theoretical and Applied Genetics 114: 1091–1103.

Which candidate genes are….
45
Von Korff M, Wang H, Leon J, Pillen K. 2005. AB-QTL analysis in spring barley.I. Detection of
resistance genes against powdery mildew, leaf rust and scald introgressed from wild barley.
Theoretical and Applied Genetics 110: 583- 590.
Voorrips RE. 2002. MapChart: software for the presentation of linkage maps and QTLs. The Journal
of Heredity 93: 77–78.
Wang Z, Taramino G, Yang D, Liu G, Tingey SV, Mia o GH, Wang GL. 2001. Rice ESTs with
disease-resistance gene- or defense-response gene-like sequences mapped to regions
containing major resistance genes or QTLs. Molecular Genetics and Genomics 265: 302–310.
Wenzl P, Li H, Carling J, Zhou M, Raman H, Paul E, Hearnden P, Maier C, Xia L, Caig V et al.
2006. A high-density consensus map of barley linking DArT markers to SSR, RFLP and STS loci
and agricultural traits. BMC Genomics. doi:10.1186/1471-2164-7-206.
Zhan J, Fitt BDL, Pinnschmidt HO, Oxley SJP, Newton AC. 2008. Resistance, epidemiology and
sustainable management of Rhynchosporium secalis populations on barley. Plant Pathology 57:
1–14.
Zhang NW, Lindhout P, Niks RE, Jeuken MJW. 2009. Genetic dissection of Lactuca saligna nonhost
resistance to downy mildew at various lettuce developmental stages. Plant Pathology, in press.
Zhu M, Zhao S. 2007. Candidate Gene Identification Approach: Progress and Challenges.
International Journal of Biological Sciences 3: 420-427.
Zierold U, Scholz U, Schweizer P. 2005. Transcriptome analysis of mlo-mediated resistance in the
epidermis of barley. Molecular Plant Pathology 6: 139-152.
Zimmermann G, Baumlein H, Mock HP, Himmelbach A, Sc hweizer P. 2006. The multigene family
encoding germin-like proteins of barley. Regulation and function in basal host resistance. Plant
Physiology 142: 181-192.

Chapter 2
46

Chapter 3
54
Transgressive segregation for extreme low and high
level of basal resistance to powdery mildew in barley
R. Aghnoum, R. Tadayonirad and R.E. Niks

Chapter 3
48
Abstract
Basal resistance of barley to powdery mildew is a quantitatively inherited trait that
limits the growth and sporulation of barley powdery mildew pathogen by a non-
hypersensitive mechanism of defense. Two experimental barley lines were developed
with extremely high (ErBgh) and low (EsBgh) level of basal resistance to powdery
mildew by convergent crossing followed by phenotypic selection between the most
resistant and the most susceptible lines from four mapping populations of barley. We
tried to verify the alleles of the seven identified QTLs in the four mapping populations
from which the most resistant and the most susceptible individuals had been derived.
Only four out of seven QTLs were confirmed to be present in the selected resistant
lines. Surprisingly, none of the expected QTLs could be detected in the resistant line
ErBgh. Our results showed that phenotypic selection in convergent crossing is very
effective and resulted in very contrasting phenotypes for basal resistance. Histological
observations showed that in ErBgh almost 90% of infection units failed to penetrate
the epidermal cells in association with papilla formation. Established infection units
may fail in formation of a substantial proportion of the secondary and subsequent
haustoria. The extremely resistant and susceptible lines developed here are valuable
material to be used in further experiments to characterize the molecular basis of basal
resistance to powdery mildew.
Introduction R-gene triggered resistance is considered easy to select for, because of the large
phenotypic effect. Most R-gene resistance appears to be associated with HR cell
death (Watanabe & Lam, 2005). This resistance is only able to protect the plants
against those races of the pathogen that possess an avirulence (Avr) gene
corresponding to the R-gene. Pathogens like Blumeria graminis, the cereal powdery
mildew pathogen, that possess a mixed (sexual and asexual) reproduction system
and a high potential for gene flow due to the long-range wind dispersal of spores, are
easily able to break down R-gene mediated resistance. Use of quantitative resistance
has been proposed as a breeding strategy for durable protection against such a
pathogens (McDonald & Linde, 2002). The minor gene, quantitative, resistance is
often considered much more difficult to select for, due to the small contribution of
each gene to the total level of resistance. Marker assisted selection has been
proposed as a strategy to accumulate such resistance genes, in order to achieve high
levels of this durable, non-race specific resistance (Castro et al., 2003, Friedt &
Ordon, 2007). Quantitative resistance of barley to powdery mildew is based on
impaired haustorium formation in the attacked epidermal cells, and is not associated

Transgressive segregation for ….
49
with a hypersensitive reaction of challenged cells.This resistance also called basal
resistance (see Glossary). Such a quantitative pre-haustorial resistance also occurs in
various other mildew and rust fungal pathosystems (e.g. Carver, 1986; Niks, 1986;
Asher & Thomas, 1987; Sillero & Rubiales, 2002). It has been reported frequently that
among the progenies of a cross between two plant genotypes with intermediate levels
of resistance or even susceptibility, descendants can be found with a higher or lower
level of resistance compared to that of both parents (e.g. Jones, 1983; Wallwork &
Johnson, 1984; Roderick & Jones, 1991). This phenomenon is known as
transgressive segregation. Transgressive segregation implies that both parents carry
a similar gene dose for the trait of interest, but those genes are located at different
loci, and alleles for resistance from both parents can end up together in some of the
progeny plants.
Four barley mapping populations that had not been developed with the aim to study
quantitative resistance to powdery mildew demonstrated that there is a great genetic
diversity of minor genes for powdery mildew resistance, with very few of such genes
in common between the populations (Chapter 2). Such a high diversity leads to
substantial transgressive segregation for resistance levels. We presume that further
accumulation of minor genes by convergent crossing and rounds of selection should
result in even higher levels of resistance to be achieved.
The objective of the present study was to investigate genetic plasticity of a limited,
medium resistant barley gene pool for obtaining extremely susceptible and extremely
resistant progeny by phenotypic selection for low and high resistance levels,
respectively. On the basis of the knowledge of the contributing QTLs (see Chapter 2)
we intended to test whether the resulting extremely susceptible and resistant progeny
would indeed carry the expected QTL alleles for susceptibility or resistance.
Material and Methods
Crossing and selection program In a previous study (Chapter2) we mapped QTLs for powdery mildew resistance at
seedling stage in four mapping populations of barley, viz. one recombinant inbred line
(RIL) population, L94 × Vada (LnVa) and three doubled haploid (DH) populations,
Nure × Tremois (NuTr), Steptoe × Morex (StMo) and Oregon Wolfe Barleys (OWBs).
Per mapping population, one line was selected with the lowest infection frequency on
the primary leaf and the lowest haustorium formation rate of challenged epidermal
cells. These homozygous most resistant lines were crossed pairwise (F1), and their
most resistant descendants (in F2) were intercrossed again, forming a Double Cross
(DC), to develop, by phenotypic selection, a descendant (in DC-S1) with the highest

Chapter 3
50
level of quantitative (basal) resistance (Figure 1). In parallel, per mapping population,
one line was selected with the highest infection frequency on the primary leaf and the
highest haustorium formation rate of challenged epidermal cells, representing the
most susceptible line of that mapping population. These lines with very low level of
basal resistance were intercrossed in two rounds to form a Double Cross (DC) to
accumulate QTL alleles for susceptibility. For development of DH barley lines with
extremely high level of basal resistance, the DC-S1 seeds were allowed to self-
pollinate and the most resistant DC-S2 plant was selected. A bulk of microspores of
six plants (DC-S3) derived from the self-pollination of this DC-S2 plant was used for
DHs production. For development of a DH barley line with extremely low level of basal
resistance, three DC-S1 plants were used to make DH lines. From each DH plant
three seedlings (12 days-old) were inoculated with powdery mildew and based on the
infection frequency, four days after inoculation, two DH lines, one with the highest and
one with the lowest infection frequency, were selected. The susceptible barley cv.
Manchuria was used as reference.
Evaluation of basal resistance at seedling stage For evaluation of the mapping populations, selection of the most resistant and the
most susceptible progenies and DHs, the following method was used. Barley plants
were sown in boxes (37 × 39 cm) and were grown in spore free greenhouse
compartments at 18-20 °C with 70% relative humidity and a photoperiod of 16 h. After
12 days, the fully expanded primary leaf was fixed in a horizontal position, with the
adaxial side up. A settling tower was used for inoculation. Freshly produced conidia
on young seedlings of the susceptible cultivar ‘Manchuria’ were blown into the tower.
The density of inoculum was monitored by haemocytometer and was adjusted to 10-
15 conidia per mm2. The inoculated plants were incubated in a growth chamber at 18-
20 oC with fluorescent light irradiance (102.8 micromol/m2/s) supplied for 12 h per
days. Four days after incubation, the barley accessions were scored by counting the
number of young powdery mildew colonies per 2 cm2 of leaf area using a frame with a
0.5 × 4 cm window and a magnifying glass (10x).
Histological assessment of infection Samples were collected from the middle part of inoculated leaves at 48 hai (hours
after inoculation) and transferred to a solution of acetic acid-ethanol (3/1 v/v) to be
cleared overnight. Then the leaf segments were stained with 0.1 mg ml-1 Trypan Blue
in alcoholic lactophenol as described by Peterhänsel et al. (1997). To make the
prepreations, the leaf segments were embedded adaxial side up in glycerol and

Transgressive segregation for ….
51
examined at 400× magnification using a transmitted white light Axiophot microscope
(Zeiss, Germany). Only stomatal and interstomatal cells (type A and type B of
epidermal cells following the terminology of Koga et al., 1990) were examined. Long
epidermal cells over-lying vascular bundles (type C) and subsidiary cells were not
considered. Only those epidermal cells that were attacked by a single germinated
conidium were evaluated.
Verification of QTLs via putative flanking markers One DH line was selected from each double cross DH population. For the resistant
selection (DC-R) there were 132 DHs, for the susceptible selection (DC-S) 129 DHs.
From DC-R and DC-S the most resistant and the most susceptible DH lines,
respectively, were selected based on the phenotypic reaction of DHs to powdery
mildew. The inoculation with powdery mildew and evaluation was performed in the
same way as described earlier. The selected lines were genotyped for flanking
markers of the seven identified QTLs (Table1).
Results
Development of barley lines with extremely low and high level of basal
resistance to powdery mildew Based on both macroscopic and histological data, DC-S1 families showed
transgressive segregation both towards resistance and susceptibility (data not
shown). One DH line was selected from each double cross (DC-R and DC-S)
combination out of 132 and 129 total DH lines produced, respectively. The extremely
resistant and the extremely susceptible barley lines to Bgh were called ErBgh and
EsBgh, respectively. ErBgh and EsBgh showed a remarkable difference in infection
frequency of powdery mildew at 6 dai (Figures 2 and 3). Histological evaluations
revealed that in the most resistant lines of the mapping populations always a higher
proportion of germlings was stopped in association with papilla formation at the sites
of attempted penetration than in the most susceptible lines (Figure 4). The ErBgh line
allowed only about 12 % and the EsBgh line 67% of the mildew germlings to form a
primary haustorium (Figure 4). In ErBgh a considerably lower proportion of secondary
penetration attempts along the elongated secondary hyphae (ESH) resulted in a
haustorium than in EsBgh at 48 hai (19% and 60% respectively). Interestingly, in
ErBgh, most of the successful secondary penetrations were observed in the same cell
as where the first haustorium was formed (Figure 5). On ErBgh no substantial
hypersensitivity reaction was observed.

Chapter 3
52
Our results demonstrated that just by visual selection QTLs from different sources
can be combined to reach an extreme level of basal resistance.
QTL verification We tried to verify the alleles of the seven identified QTLs in the four mapping
populations from which the most resistant and the most susceptible individuals had
been derived. It seemed reasonable to presume that line ErBgh would combine for all
or most of the QTLs the resistance allele, EsBgh for all or most of the QTLs the
susceptibility allele. We checked the genotype of the most resistant and the most
susceptible genotypes of each mapping population that were used for making the first
round of crossings (Table 1). Only four QTLs (Rbgq14, Rbgq2a, Rbgq19 and
Rbgq10) were confirmed to have the resistance allele in those four selected resistant
lines (Table 1). Surprisingly, none of the QTLs for which we can have a conclusion
(Rbgq14, Rbgq2a, Rphq19 and Rphq10) could be detected in the resistant line
ErBgh. Only three QTLs (Rbgq2a, Rbgq19 and Rbgq15) were confirmed to have the
susceptibility allele in the selected susceptible lines. Surprisingly, none of the QTLs
for which we can have a conclusion (Rbgq2a, Rbgq19 and Rbgq15) could be
detected in the susceptible line EsBgh.
The verification of the QTL alleles present in ErBgh and EsBgh was hampered by at
least two technical points.
1-Lack of multiple allelic variations makes a QTL peak or flanking marker-allele not
unique for selection if this allele also occurs in at least one of the parents of the other
mapping populations. This is especially true for markers like AFLP, where each
marker has only two variants (“0” or “1”).
2 -We searched on the latest integrated barley linkage map (Chapter 2) for multiple
allelic SSR markers close to the target QTLs, to increase the chance that the allele
that was indicative for a particular QTL allele, did not occur in any of the other
ancestral lines. However, we found such SSR markers often at some distance from
the peak marker of the QTL. Therefore, recombination between the marker locus and
the QTL might occur during the crossing procedure. In case of such a distance,
absence of the indicative marker allele does not prove absence of the QTL allele.

Transgressive segregation for ….
53
Mapping QTLs for powdery mildew resistance at seedling stage in four mapping populations
Selection of the most resistant line of each population
Two cycles of recurrent selection to accumulate QTLs for resistance
Figure 1. A schematic representation of a crossing programs to develop a barley line with extreme high level of basal resistance to powdery mildew. A parallel crossing program followed to develop a line with extreme low level of resistance.
R1 × R2 R3 × R4
F1 Selfing F1 F2 Selection F2 F2 R1 × R2 × F2 R3 × R4
DC
DC-S1 DC-S2 Selection DC-S3 Making DH lines Selection of the extremely resistant line
Figure 2. Phenotypeof the extremely resistant (ErBgh, left) and the extremely susceptible (EsBgh) barley lines six days after inoculation with Bgh with the same inoculom density and incubation conditions.

Chapter 3
54
Table1: Characteristics of QTLs mapped at seedling stage for powdery mildew resistance and
the genotype of the most resistance and the susceptible lines used for crossing Population QTLs Peak marker LOD Exp% a Donor b R parent c S parent d
OWBs
Rbgq20
Rbgq14
ABG704
Bmac0047e
3.5
4.9
18.3
19.9
Rec
Dom
-
+
+
+
NuTr
Rbgq2a
Rbgq19
bPb-0312
scssr05599
6.1
5.55
24.8
22.4
Tremois
Nure
+
+
-
-
LnVa
Rbgq2b
Rbgq15
E35M48-410a
E33M58-306
2.8
3.1
19.8
21.5
L94
Vada
-
-
+
-
StMo Rbgq10
MWG571C 4.7
15.5
Morex
+ +
a The proportion of the phenotypic variance explained by the QTL b Donor of the resistance allele
c The genotype of the most resistant (R ) line of each population regarding the expected QTL; + indicates that the
line contained a molecular marker that suggested the expected resistance allele of the QTL, - that the line
contained a molecular marker that suggested the unexpected susceptibility allele. d The genotype of the most susceptible (S) line of each population regarding the expected QTL; + indicates that
the line contained a molecular marker that suggested the expected susceptibility allele of the QTL, - that the line
contained a molecular marker that suggested the unexpected resistance allele.

Transgressive segregation for ….
55
0
10
20
30
40
50
60
70
80
90
StMo-
r
LnVa-r
OWBs-r
NuTr-r
StMo-
s
LnVa-s
OWBs-s
NuTr-s
ErBgh
EsBgh
Man
churia
Infe
ctio
n fr
eque
ncy
Figure 3. Infection frequency [colonies per 2 cm2] of the most resistant (r) and
the most susceptible (s) genotypes of different mapping populations. LnVa (L94
× Vada), StMo (Steptoe × Morex), OWBs (Oregon Wolf Barleys) and NuTr
(Nure × Tremois). ErBgh represents the extremely resistant and EsBgh the
extremely susceptible DH barley line to Bgh derived from the double crosses.
Data were collected at 4 dai.
0
10
20
30
40
50
60
70
80
StMo-
r
LnVa-r
OWBs-r
NuTr-r
StMo-
s
LnVa-s
OWBs-s
NuTr-s
ErBgh
EsBgh
Man
churia
Pen
etra
tion
eff
icie
ncy
%
Figure 4. Penetration efficiency (% successfully penetrating germlings) of Bgh
on the most resistant (r) and the most susceptible (s) genotypes of different
mapping populations. ErBgh represents the extremely resistant and EsBgh the
extremely susceptible DH barley line to Bgh derived from the double crosses.

Chapter 3
56
Discussion
Accumulation of genes with minor effect by phenotyp ic selection Our data support the proposed strategy that genotypes with much higher (and lower)
level of resistance than the parents can be selected from progenies derived from
convergent crosses between several parents with intermediate levels of resistance,
due to high transgression of the genetic components from both parents (Wallwork &
Johnson, 1984). This high success of accumulation of quantitative genes for basal
resistance can be ascribed to the great diversity of such genes in the barley
germplasm, as shown in Chapter 2, involving different loci in each parental pair. Such
an accumulation was, for the same reason, also easily achieved by Parlevliet &
Kuiper (1985), in barley to barley leaf rust (Puccinia hordei). Also to that pathogen
loci for basal resistance occur all over the barley genome (Qi et al., 2000; Marcel et
al., 2007), making transgression for basal resistance a very common phenomenon.
Efficiency of phenotypic selection to achieve a hig h level of basal
resistance Our results showed that phenotypic selection in convergent crossing is very effective
and resulted in very contrasting phenotypes for basal resistance. MAS has some
problems, like the lack of unique markers close to the peak marker of the QTLs of
Figure 5. Develpoment of Bgh (48 hai) on ErBgh. A, a successful primary penetration resulted in a haustorium and a failed secondary attempted penetration (SAP). B, a closer view of failed SAP showed in fig. A. C, a successful primary penetration resulted in formation of a secondary haustorium in the same cell. The scale bar represents 100 µm.

Transgressive segregation for ….
57
interest, especially if more than two parents will be used in a convergent crossing
programme. With strong enough linkage disequilibrium (see Glossary) between
markers and QTLs (Dekkers & Hospital, 2002), MAS should in theory be easier,
faster, cheaper, or more efficient than classical (phenotypic) selection but in practice
the MAS is not always as efficient as expected (Hospital, 2008). It should be pointed
out here that the efficiency of MAS also depends on the type of the trait and to the
population. Phenotypic selection is more effective than MAS in some cases or at least
both are of the same value (Romagosa et al., 1999; Flint-Garcia et al., 2003). Flint-
Garcia et al. (2003) compared the efficiency of phenotypic selection versus Marker-
assisted selection for rind penetrometer resistance (RPR) and European corn borer
(ECB). They reported that MAS for high and low RPR was effective in the three
populations studied. Phenotypic selection for both high and low RPR was more
effective than MAS in two of the populations. However, in a third population, MAS for
high RPR using QTL effects from the same population was more effective than
phenotypic selection. MAS is probably not needed for a rather simple trait like
quantitative powdery mildew resistance. Selection for powdery mildew resistance is
quite straightforward, plenty of genes in the ancestors are available that can be
accumulated and non-genetic variation is relatively low if inoculum is applied
homogeneously over the plants to be compared. MAS may be more useful to
introgress or accumulate QTLs for such traits like drought and salt tolerance that are
difficult to select for phenotypically. However, also in such cases unique marker
alleles are required if a convergent crossing scheme is used.
It is remarkable that our initially selected most resistant and most susceptible lines
from the mapping populations did not contain (nearly) all the expected QTL alleles
(Table 1). One possible explanation may be that the effect of QTLs depends strongly
on the genetic background. Marcel et al. (2008) showed that some QTLs for basal
resistance to P. hordei had a particularly high effect if combined with other QTLs, that
alone had hardly any effect. Such interactions would make phenotypic selection
probably more successful than MAS, in case the partner QTLs of such interactions
are still unknown.
High level of basal penetration resistance in ErBgh line Basal resistance of barley to powdery mildew is mainly based on impairment of
haustorium formation. The observation that established infection units in ErBgh may
fail in formation of a substantial proportion of the secondary and subsequent
haustoria (Figures 5A and B) is unexpected. In other barley and oat genotypes
(Carver & Carr, 1978; Gustafsson & Claesson, 1988), lower secondary haustorium
Failed SAP

Chapter 3
58
formation is reported but it is not mentioned whether this is due to failure of
attempted secondary penetration or to slow growth resulting in fewer penetration
attempts. The percentage of successful primary penetration in ErBgh is nearly as low
(12%) as the lowest (10%) reported ever (Carver, 1986), excluding non-host and mlo
resistance. It seems plausable that further rounds of convergent crossing and
selection, involving parents with high quantitative resistance might easily increase
resistance levels even further. It should be further investigated whether the failed
secondary and attempted penetration in ErBgh is due to papilla formation or that
other mechanisms are involved.
The present observation that in ErBgh at 48 hai the second and next haustoria are
often formed in the same cell as the first haustorium (Figure 5C) is intriguing. A
similar phenomenon was observed in SusBgt lines that show a high level of
susceptibility to the non-adapted wheat powdery mildew, B. graminis f.sp.tritici
(Chapter 4). This is a further indication that basal host resistance has similarities with
non-host resistance.
We consider the extremely susceptible and resistant lines developed here as
valuable material to be used in further experiments to characterize the molecular
basis of basal resistance to powdery mildew. Additional work on this material could
also lead to a critical comparison of phenotypic versus marker assisted selection for a
quantitative trait like basal resistance.
Acknowledgements
The excellent technical assistance of Anton Vels in greenhouse experiments is greatly appreciated. Dr.
TC. Marcel is acknowledged for his help in QTL verification using SSR markers.
Reference
Asher MJC, Thomas CE. 1987. The inheritance of mechanisms of partial resistance to Erysiphe
graminis in spring barley. Plant Pathology 36: 66-72.
Carver TLW, Carr AJH. 1978. Effects of host resistance on the development of haustoria and colonies
of oat mildew. Annals of Appliled Biology 88: 171–178.
Carver TLW. 1986. Histology of infection by Erysiphe graminis f. sp. hordei in spring barley lines with
various levels of partial resistance. Plant Pathology 35: 232–240.
Castro AJ, Chen X, Hayes PM, Johnson M. 2003. Pyramiding quantitative trait locus (QTL) alleles
determining resistance to barley stripe rust: Effects on resistance at the seedling stage. Crop
Science 43: 651-659.
Dekkers JCM, Hospital F.2002. The use of molecular genetics in the improvement of agricultural
populations. Nature Reviews Genetics 3: 22–32.
Flint-Garcia SA, Darrah L, McMullen M. 2003. Phenotypic versus marker-assisted selection for stalk
strength and second-generation European corn borer resistance in maize. Theoretical and
Applied Genetics 107: 1331–1336.

Transgressive segregation for ….
59
Friedt W, Ordon F. 2007. Molecular Markers for Gene Pyramiding and Disease Resistance Breeding
in Barley. In:Varshney RK, Tuberosa R, eds, Genomics-Assisted Crop Improvement:Vol. 2:
Genomics Applications in Crops, 81–101.
Gustafsson M, Claesson L. 1988. Resistance to powdery mildew in wild species of barley. Hereditas
108: 231–237.
Hospital F. 2008. Challenges for effective marker-assisted selection in plants. Genetica
10.1007/s10709-008-9307-1
Jones IT. 1983. Transgressive segregation for enhanced level of adult plant resistance to mildew in
the oat cross Mostyn x Maldwyn. Euphytica 32: 499-503.
Koga H, Bushnell WR, Zeyen RJ. 1990. Specificity of cell type and timing of events associated with
papilla formation and the hypersensitive reaction in leaves of Hordeum vulgare attacked by
Erysiphe graminis f. sp. hordei. Canadian Journal of Botany 68: 2344–2352.
Marcel TC, Gorguet B, Ta MT, Kohutova Z, Vels A, Ni ks RE. 2008. Isolate specificity of quantitative
trait loci for partial resistance of barley to Puccinia hordei confirmed in mapping populations and
near-isogenic lines. New Phytologist 177: 743–755.
Marcel TC, Varshney RK, Barbieri M, Jafary H, de Ko ck MJD, Graner A, Niks RE. 2007. A high-
density consensus map of barley to compare the distribution of QTLs for partial resistance to
Puccinia hordei and of defence gene homologues. Theoretical and Applied Genetics 114: 487–
500.
McDonald BA, Linde C. 2002. The population genetics of plant pathogens and breeding strategies for
durable resistance. Euphytica 124: 163-180.
Niks RE. 1986. Failure of haustorial development as a factor in slow growth and development of
Puccinia hordei in partially resistant barley seedlings. Physiological and Molecular Plant
Pathology 28: 309–322.
Parlevliet JE, Kuiper HJ. 1985. Accumulating polygenes for partial resistance to barley leaf rust,
Puccinia hordei. 1. Selection for increased latent period. Euphytica 37: 7–13.
Peterhänsel C, Freialdenhoven A, Kurth J, Kolsch R, Schulze-Lefert P. 1997. Interaction analyses
of genes required for resistance responses to powdery mildew in barley reveal distinct pathways
leading to leaf cell death. Plant Cell 9: 1397–1409.
Qi X, Fufa F, Sijtsma D, Niks RE, Lindhout P, Stam P. 2000. The evidence for abundance of QTLs
for partial resistance to Puccinia hordei on the barley genome. Molecular Breeding 6: 1–9.
Roderick HW, Jones IT. 1991. The evaluation of adult plant resistance to powdery mildew (Erysiphe
graminis f.sp. avenae) in transgressive lines of oats. Euphytica 53: 143-149.
Romagosa I, Han F, Ullrich SE, Hayes PM, Wesenberg DM. 1999. Verification of yield QTL through
realized molecular marker-assisted selection responses in a barley cross. Molecular Breeding 5:
143–152.
Sillero JC, Rubiales D. 2002. Histological characterization of the resistance of faba bean to faba bean
rust. Phytopathology 92: 294–299.
Wallwork H, Johnson R. 1984. Transgressive segregation for resistance to yellow rust in wheat.
Euphytica 33: 123–132.
Watanabe N, Lam E. 2005. The Hypersensitive Response in Plant Disease Resistance. In: S, Tuzun
E, Bent eds, Multigenic and Induced Systemic Resistance in Plants. Springer. 83-111.

Chapter 3
54

New perspectives to identify genes that…
54
New perspectives to identify genes that determine
non-host resistance of barley to non-adapted
Blumeria graminis forms
R. Aghnoum and R.E. Niks
Submitted

Chapter 4
62
Abstract The genetic basis of non-host resistance of barley to non-adapted formae speciales of
Blumeria graminis is not known, since there is no barley line that is susceptible to, for
example, the wheat powdery mildew pathogen, Blumeria graminis f.sp. tritici (Bgt).
We identified barley accessions with rudimentary susceptibility to the non-adapted
Bgt. Those accessions were intercrossed in two cycles and selected two lines, called
SusBgt SC and SusBgt DC, with exceptional susceptibility to Bgt at seedling stage.The
quantitative variation among barley accessions and in the progenies after convergent
crossing suggests a polygenic basis for this non-host resistance. Both lines show a
remarkable level of haustorium formation and colony development by the non-
adapted target mildew. The SusBgt lines and their ancestor lines also allowed
haustorium formation and conidiation by four out of seven tested non-adapted
Blumeria forms. Component analysis of the infection process suggested that non-host
resistance factors are specific to the form and developmental stage of Blumeria.
Resistance to establishment (first haustorium), colonization (subsequent haustoria)
and conidiation are not clearly associated.
The lines developed will serve to elucidate the genetic basis of non-host resistance
in barley to Bgt, and they are useful tools in gene expression and complementation
studies on non-host resistance.
Introduction The majority of plant species are immune to the majority of potentially pathogenic
microbial invaders. This is the most common and broadest spectrum resistance,
termed non-host resistance. It is defined as resistance shown by all members of a
plant species against all members of a given pathogen species (Heath, 2000;
Thordal-Christensen, 2003). Non-host resistance relies on a variety of preformed and
inducible mechanisms of defense (Thordal-Christensen, 2003; Mysore & Ryu, 2004).
Pathogens that are able to overcome the preformed defense mechanisms encounter
extracellular surface receptors that recognize general elicitors, the so-called
pathogen-associated molecular patterns (PAMP) (see Glossary) (Nürnberger et al.,
2004; Chisholm et al., 2006). The pathogen should suppress the PAMP-triggered
immunity (PTI) in order to establish a compatible interaction and successfully colonize
the plant (Chisholm et al., 2006). Those pathogen species that are not able to
suppress the PTI are often called non-adapted, inappropriate or heterologous
pathogens. It is presumed that would-be pathogens deliver effectors (see Glossary) in
the attacked plant cells that may suppress PTI if there is sufficient correspondence
between effectors and their respective operative targets (Block et al., 2008; van der

New perspectives to identify genes that…
63
Hoorn & Kamoun, 2008). The degree of correspondence between effectors and
targets may determine the host status of a plant to a would-be pathogen (Niks &
Marcel, 2009). Two interesting questions are whether different stages of development
of a pathogen in a plant may require different sets of effectors and whether different
strains of a pathogen species need to address the same operative targets in a
particular plant species.
Powdery mildew in cereals is caused by the biotrophic Ascomycete fungus Blumeria
graminis (Bg) formerly known as Erysiphe graminis. Bg has a high level of biological
specialization for its cereal and grass hosts. Barley is considered a strict non-host
plant species to the wheat forma specialis, Blumeria graminis f.sp. tritici (Bgt), and
also to the forms of Blumeria graminis that are pathogenic to oats and rye (Hardison,
1944; Eshed & Wahl, 1970; Menzies & MacNeill, 1989). At the macroscopic level, no
barley accession has been identified that allowed Bgt colonies to develop (Menzies &
MacNeill, 1989; Olesen et al., 2003; Atienza et al., 2004). At the microscopic level,
however, some barley accessions have been reported to permit some development of
Bgt (Tosa & Shishiyama, 1984; Trujillo et al., 2004). Penetration of the plant
epidermal cell walls and subsequent haustorium formation, hereafter called
establishment, is a critical stage in the infection process. In compatible interactions of
barley with the adapted powdery mildew pathogen, Blumeria graminis f.sp. hordei
(Bgh), the fungus can successfully penetrate through epidermal cell walls and form
haustoria to obtain nutrients from the host cells. In non-host interactions of barley with
non-adapted formae speciales such as Bgt, establishment typically fails in association
with formation of cell wall appositions (papillae), in some cases supplemented with
the hypersensitive cell death reaction (HR) of single attacked cells. In the interaction
of barley with the adapted powdery mildew, Bgh, a proportion of attempted germlings
are also stopped by such defense mechanisms (Tosa & Shishiyama, 1984;
Hückelhoven et al., 2001; Trujillo et al., 2004).
It is unknown which genetic factors determine that barley is a host to Bgh but a non-
host to non-adapted formae speciales of Bg. Since there is no barley line that is
susceptible to, for example, Bgt, inheritance studies are not possible. Some candidate
genes have been implicated to play a role in resistance of barley to the wheat forma
specialis (Bgt) by transient overexpression or transient-induced gene silencing (TIGS)
in single epidermal cells (Miklis et al., 2007; Douchkov et al., 2005; Eichmann et al.,
2004; Schweizer, 2007), but it is not known whether allelic variation between these
genes explains the host status difference between wheat and barley to Bgt. Barley is
indeed a model plant in which to study the molecular basis of basal and non-host
resistance to powdery mildews (Hückelhoven, 2007; Schweizer, 2007; Collinge et al.,

Chapter 4
64
2008). We set out to identify the genes that are responsible for natural variation in
host status and non-host resistance level.
In the present paper, we identified barley accessions with rudimentary susceptibility
to the non-adapted wheat powdery mildew (Bgt) and crossed them to develop
experimental barley lines with exceptional susceptibility to Bgt. We quantified
components of infection development of Bgh, Bgt and several other non-adapted
powdery mildew formae speciales and species on these experimental lines to
determine the specificity of resistance factors.
Materials and Methods
Plant and Pathogen Materials A collection of 439 accessions of barley was evaluated at the seedling stage to the
non-adapted wheat powdery mildew fungus, Bgt. The collection consisted of 136
accessions of Hordeum spontaneum and 303 accessions of Hordeum vulgare: viz.,
227 accessions (landraces and wild H. spontaneum barley from the ICARDA barley
germplasm collection) provided by Dr. Rajeev K. Varshney, 54 accessions received
from the Centre for Genetic Resources (CGN), Wageningen, the Netherlands and 22
parental lines of available mapping populations of barley at the Laboratory of Plant
Breeding, Wageningen University. The seeds of barley cv. Turkey 290 were kindly
provided by Dr. H.E. Bockelman (National Small Grains Collection, USDA, USA). An
isolate of Bgt (Swiss field isolate FAL92315), kindly provided by Dr. Patrick Schweizer
(IPK, Germany), was propagated on the susceptible wheat cv. Vivant and used for
inoculation. The lines with some level of susceptibility to Bgt were intercrossed in two
cycles and resulted in two lines with exceptional susceptibility to Bgt (see Results
section). These lines were named SusBgt SC and SusBgt DC. SusBgt lines and the
parental lines were tested to several non-adapted powdery mildew formae speciales
(ff. spp.) and species. The isolates will be referred to according to their source host.
The following non-adapted ff. spp. of Bg were tested: Blumeria graminis f.sp. avenae
(Bga), the powdery mildew of oat (Avena sativa); Blumeria graminis f.sp. secalis
(Bgs), the powdery mildew of rye (Secale cereale); and four isolates collected from
wild grasses, Hordeum murinum, Blumeria graminis f.sp. hordei-murini (Bghm);
Agropyron repens, Blumeria graminis f.sp. agropyri (Bgar); Bromus mollis, Blumeria
graminis f.sp. bromi (Bgb); and Dactylis glomerata, Blumeria graminis f.sp. dactylidis
(Bgd). All of these isolates were collected in the Wageningen region and maintained
in isolation on their respective host plants. We also tested two non-adapted species,
viz. Oidium neolycopersici, the powdery mildew of tomato, and Sphaerotheca
fuliginea, the powdery mildew of cucumber.

New perspectives to identify genes that…
65
Inoculation
Three seedlings of each barley accession were grown in boxes 37 × 39 cm in size.
The susceptible wheat cv. Vivant was sown in each box to monitor the effectiveness
of inoculation. Turkey 290, SusPmur and SusPtrit which have been reported to allow
relatively high haustorium formation by Bgt (Tosa & Shishiyama, 1984; Trujillo et al.,
2004) were included as references. The fully expanded primary leaves of 12-day-old
seedlings were stapled horizontally, adaxial side up. To make a dense inoculation,
freshly produced conidia of a Bgt isolate formed on the susceptible wheat cv. Vivant
were blown onto the leaves to reach a density of approximately 50 conidia per mm2.
The inoculated plants were incubated in a growth chamber at 18-20 °C with 70%
relative humidity and a photoperiod of 16 h (200 micromol/m2/s).
Macroscopic and microscopic evaluation of barley- Bgt interaction First, macroscopic evaluations of all tested accessions were carried out using a
magnifying glass (10x) eight days after inoculation (dai). Those accessions that
macroscopically showed some degree of susceptibility were selected. These
accessions were sown and inoculated again for quantification of haustorium formation
and conidiation under the microscope. At two time points, 72 hours after inoculation
(hai) and 8 dai, a leaf segment approximately 3 cm long was collected from the
middle part of the infected primary leaves. Leaf segments were transferred to a
solution of acetic acid-ethanol (3:1 v:v) to be cleared for at least 3 hours. Then, the
leaf segments were stained in Coomassie Brilliant Blue according to the method of
Wolf & Fric (1981). The experiment was carried out in two consecutive replications.
Per barley accession, 200 observed germlings were scored for the result of the first
penetration attempt to establish a haustorium. Because different types of host
epidermal cells show different degrees of resistance to penetration (Lin & Edwards,
1974; Koga et al., 1990) and penetration attempts by different germlings on the same
cell may induce resistance or susceptibility (Carver et al., 1999; Olesen et al., 2003),
we considered only cells that were attacked by a single conidium and only epidermal
cells adjacent to stomata and interstomatal epidermal cells (type A and type B of
epidermal cells following the terminology of Koga et al., 1990).
Evaluation of susceptibility of SusBgt lines SusBgt lines and the parental lines were tested to non-adapted powdery mildew
isolates in order to quantify the degrees of haustorium formation, hypersensitive and
non-hypersensitive mechanism of defense and conidiation. At 72 hai, leaf samples
were cut and placed in a solution of 1 mg ml−1 3,3'-Diaminobenzidine (DAB)-HCL, pH

Chapter 4
66
3.8, which allows whole-cell accumulation of H2O2, for 8 h and subsequently
transferred to a solution of acetic acid-ethanol (3:1 v:v) to be cleared overnight. Then
the leaf segments were stained with 0.1 mg ml-1 Trypan Blue in alcoholic lactophenol
as described by Peterhänsel et al., (1997). H2O2 accumulation appears as a reddish-
brown coloration of epidermal cells and indicates the degree of HR (Thordal-
Christensen et al., 1997).
Results
Genotypic variation in interaction of barley with t he wheat powdery
mildew pathogen The majority of the 439 barley accessions were (nearly) immune, showing no
macroscopic symptoms or signs of infection whatsoever with the non-adapted
pathogen (Bgt) even 10 days after inoculation. However, a few barley accessions
were identified that showed a low degree of susceptibility to Bgt. On these accessions
very small lesions, or even in a few cases tiny colonies, were visible by 10x
magnifying glass or by naked eye, indicating some haustorium formation by the non-
adapted pathogen. Six such accessions were examined under the microscope to
quantify the amount of haustorium formation in comparison with lines Turkey 290,
SusPtrit and SusPmur and with the immune barley cv. Vada. The cellular reactions of
these selected lines to Bgt is shown in Figure 1. On barley cv. Vada, all germlings
were stopped at the penetration stage in association with the deposition of cell wall
appositions (papillae), and the fungus could not develop further to establish any
haustoria. On the other accessions, the large majority of germlings were also stopped
at penetration stage, but 12 to 25 % of the germlings succeeded in penetrating
through the epidermal cell wall and established a haustorium and elongated
secondary hyphae (ESH). Accessions with similar rates of establishment by Bgt, such
as Hsp17 and SusPtrit, could differ greatly in the rate of conidium formation (Figure
1). The results confirm earlier reports (Tosa & Shishiyama, 1984; Trujillo et. al., 2004)
that there are differences among barley accessions in the degree to which they allow
the non-adapted wheat powdery mildew pathogen to establish haustoria. Such rare
barley accessions with some level of susceptibility to this non-adapted mildew were
used as parental lines to accumulate genes for susceptibility to Bgt.

New perspectives to identify genes that…
67
0
5
10
15
20
25
30
35
Hiproly
Trisuli B
azar 9
Chame 2
Bimtak
othi 7
HSP14
HSP17
Turkey
290
SusPtri
t
SusPmur
Vada
Inte
ract
ion
site
s[%
]
Figure 1. Penetration efficiency and conidiation of Blumeria graminis f.sp. tritici (Bgt) on different barley
accessions that showed some level of susceptibility to Bgt. The immune barley cv. Vada was used as a reference.
White columns represent the successful primary penetration with a haustorium, collected from two leaf segments
(each from independent inoculation experiments) fixed at 72 hai. In each leaf segment, 100 attempted
penetrations were recorded. Gray columns represent the percentage of all successful primary penetrations that
resulted in formation of conidia at 8 dai. Data represent the means of two leaf segments collected from two
independent inoculation experiments.
Development of SusBgt lines, susceptible to the non -adapted wheat
powdery mildew Based on macroscopic and microscopic observations, including the degree of
conidiation, four accessions of barley, viz. Hiproly (CGN00588; Egypt), Trisuli Bazar 9
(CGN00931; Nepal), Chame 2 (CGN01042; Nepal) and SusPtrit (Atienza et al., 2004)
were selected as the most promising lines for crossing to accumulate genes for
susceptibility to Bgt. We made pairwise crosses between these four lines (Trisuli
Bazar 9 × Hiproly) and (SusPtrit × Chame 2) (Figure 2). The second generation
offspring (F2) derived from each of the two crossing combinations were evaluated first
macroscopically for susceptibility to the Bgt isolate, and then a few F2 plants from
each crossing combination that seemed to be more susceptible than the parents were
tested microscopically for penetration efficiency and conidiation. A double cross (DC)
was made between a selected individual F4 plant derived from the cross Trisuli Bazar
9 × Hiproly and a selected individual F3 plant derived from the cross SusPtrit ×
Chame 2. Four DC-S1 plants coming from the same single DC plant were used for
making doubled haploid (DH) lines by microspore culture. We had expected additional
transgression in the DC (because it had four susceptible ancestors) compared to the
single cross (SC) with only two ancestors. However, during the procedure, the
progeny from the cross SusPtrit × Chame 2 seemed so susceptible that we produced
also a DH from that lineage. Two selected F4 individuals derived from one selected F3
plant from the single cross (SC) SusPtrit × Chame 2 were also used for making DH

Chapter 4
68
lines. Progeny from selfed DH plants were first evaluated macroscopically, and those
lines that showed a clear susceptibility were analyzed microscopically. Since the
selected DC and SC lines looked similarly susceptible, we decided to test them both
extensively. The most Bgt-susceptible lines derived from the single cross and double
cross were named SusBgt SC and SusBgt DC.
Susceptibility of SusBgt lines to the adapted ( Bgh) and non-adapted
wheat powdery mildew pathogen ( Bgt) SusBgt lines and all the parental lines showed a compatible interaction (infection type
4 according to the scale of Mains & Dietz (1930)) with the barley powdery mildew.
Interestingly, SusPtrit and both SusBgt lines arrested at least 50% of the infection
units of the adapted pathogen, Bgh, from developing haustoria. This indicates a rather
high level of basal resistance to Bgh (Figure 3).
SusBgt lines allowed the non-adapted Bgt to form many colonies that were clearly
visible by the naked eye at 8 dai and even earlier (Figure 4A). In the barley
accessions, a proportion (depending on genotype, 12 to 27%) of challenged
epidermal cells without detectable haustoria responded by HR (Figure 5A).
Figure 2. Two cycles of convergent crossing in barley to accumulate genes for susceptibility
to the wheat powdery mildew (Blumeria graminis f.sp. tritici).
Crossing Selfing Selection of the Selfing Selfing Double cross
most susceptible (selection) (selection)
F2 plants
P1 × P2 F1 F2 F3 F4
F4 P1 × P2 × F3 P3 × P4
P3 × P4 F1 F2 F3
DC-S1
Making DH lines
Making DH lines
P1 = SusPtrit Selection o f the most
susceptible DH line
P2 = Chame 2 SusBgt SC
Selection of the most
P3 = Trisuli Bazar 9 susceptible DH line
SusBgt DC
P4 = Hiproly

New perspectives to identify genes that…
69
There were great differences between the accessions in the rate of cells in which a
haustorium was formed. Vada was fully resistant to penetration by Bgt. Both SusBgt
SC and SusBgt DC lines showed a higher level of haustorium formation by Bgt than the
parental lines (Figure 5A). In SusBgt SC and SusBgt DC, 51% and 59%, respectively, of
germlings had succeeded in penetration and haustorium formation, while in Chame 2,
the most susceptible parental line, only 35% of germlings had successfully formed a
first haustorium. In SusBgt DC, which showed the highest frequency of successfully
penetrated cells, a larger proportion of penetrated cells with a haustorium (59%) were
stopped by single-cell HR than in SusBgt SC (42%) and the parental lines. The highest
frequency of successfully penetrated epidermal cells without HR was detected in
SusBgt SC (29%).
0%10%20%30%40%50%60%70%80%90%
100%
Hipro
ly
Trisuli
Bazar
9
Chame 2
SusPtri
t
SusBgt
SC
SusBgt
DC
Inte
ract
ion
site
s
Figure 3. Defense reactions of SusBgt lines and their parental lines to the barley powdery mildew (Bgh) at 48 hai.
White columns represent failed primary penetration. Black columns represent the successful primary penetration
with a haustorium and with hypersensitivity (HR). Cross-hatched columns represent the successful primary
penetration with a haustorium but without HR. Data represent the means of four leaf segments collected from two
independent inoculation experiments. In each leaf segment, 100 infection sites were recorded.
Figure 4. Macroscopic phenotypes of interaction of SusBgt lines with Blumeria graminis f.sp. tritici (Bgt) and B.
graminis f.sp. hordei-murini (Bghm). (A) Development of colonies of Bgt on SusBgt SC and SusBgt DC at 8 dai. (B)
Hypersensitive reaction of SusBgt SC to Bghm at 8 dai. Vada is immune to both ff.spp.

Chapter 4
70
The majority of those germlings that had formed a primary haustorium were stopped
at the secondary penetration attempts. In both SusBgt SC and SusBgt DC, a
considerable proportion of successfully penetrated germlings succeeded in
establishment of a secondary haustorium (44 and 42%, respectively, Figure 5D).
Interestingly, the secondary and subsequent haustoria were almost always detected
in the same cell in which the primary haustorium had been established, typically
resulting in two to five haustoria in one epidermal cell (Figure 6E). Subsequent
penetration attempts in epidermal cells located away from the first infected cell usually
failed (Figure 6F). At 8 dai, lines differed in the proportion of established germlings
that had proceeded to the formation of at least some conidiophores. This conidiation
was most conspicuous (34% of those germlings that had formed a haustorium) in
SusBgt SC, and much less so in SusBgt DC (6%) (Figure 5G). In the parental line
Trisuli Bazar 9, a similar proportion of established germlings reached conidiation as in
SusBgt SC (Figure 5G), but the rate of establishment was lower than in SusBgt SC
(Figure 5A). In summary, the results showed that SusBgt lines show an exceptional
susceptibility to the non-adapted Bgt, and the amenability to formation of haustoria is
not necessarily associated with amenability to conidiation. Both lines are valuable to
study the inheritance of non-host resistance in barley to powdery mildews.
The level of susceptibility of SusBgt lines to othe r non-adapted powdery
mildew pathogens To determine whether the SusBgt lines are susceptible to other non-adapted powdery
mildew pathogens, the four parental lines, SusBgt lines and the reference barley cv.
Vada were inoculated with one isolate each of other non-adapted formae speciales
and species of powdery mildew pathogens. SusBgt lines and their parental lines showed relatively high haustorium formation
and conidiation by the powdery mildew fungus of Hordeum murinum, Bghm (Figure
5B, 5E and 7B). At the macroscopic level, SusBgt SC showed a remarkably strong HR
reaction at 8 dai (Figure 4B). All tested accessions allowed at least some haustorium
formation and conidiation by the rye powdery mildew pathogen (Bgs) (Figure 5C and
5F), but the SusBgt lines were not more susceptible to this mildew form than the other
lines were. SusBgt SC and SusPtrit that allowed relatively high establishment by Bgt
(>50 %) allowed considerably less establishment by Bgs (< 26 %). Surprisingly, Vada,
which did not allow any haustorium formation by Bgt, allowed relatively high
haustorium formation by Bgs (31 %). Relatively few germlings of Bgs that had formed
a first haustorium proceeded to form a secondary haustorium (< 22%) and conidia
(<14%) compared to Bgt (< 45 and < 35%, respectively) (compare Figure 5D with

New perspectives to identify genes that…
71
Figure 5F, and note the difference in the scale of the ordinates). The tested barley
lines were all nearly immune to the mildew fungi of oat, Bromus mollis and Dactylis
glomerata. At most, 10% of the germlings formed a haustorium, and none of the
germlings formed conidia. Both SusBgt lines allowed some haustorium formation (35-
51%) by the Agropyron powdery mildew (Bgar), but the rate of established germlings
reaching conidiation was considerably higher in SusBgt SC than in SusBgt DC (80%
and 21%, respectively) (data not shown). As with the Bgt, during infections by the
other non-adapted mildew strains, except Bghm (Figure 7A), second and subsequent
haustoria were almost always formed in the same cell as the first haustorium, and
failed penetration attempts along the spreading hyphae were commonly observed.
The two additional non-adapted mildew species, O. neolycopersici and Sphaerotheca
fuliginea, were not observed to form any haustoria on any of the barley accessions.
Discussion Although we have not yet started to perform formal inheritance studies, several
interesting conclusions on the genetic basis of the basal resistance of barley to non-
adapted Bg mildews have emerged.
1-The resistance has a quantitative inheritance, as suggested by the quantitative
differences between accessions and by the transgressive segregation when
combining the parental lines. Our results suggest that crossing and selection among
some barley accessions with some level of susceptibility resulted in the accumulation
of susceptibility factors (or depletion of resistance factors) to the non-adapted wheat
powdery mildew, Bgt. These lines showed an unprecedented level of haustorium
formation and colony development by this non-adapted target mildew. We conclude
that non-host resistance of barley to Bgt is based on several minor genes, each with a
small effect.
2-Genes for basal resistance have pathogen developmental stage-specific effects. If
quantitative trait loci (QTLs) indeed represent operative targets for pathogen-delivered
effectors to silence aspects of the defense (Jafary et al., 2006; Niks & Marcel, 2009),
silencing of the defense at the first penetration stage may require different effector-
target interactions than ESH development or conidiation. Also, it seems that making
the first haustorium (= silencing the defense of the first plant cell) does not imply that
the defenses of the next cells are silenced easily as well. Arabidopsis penetration
mutants (pen) allow higher frequencies of penetration by the non-adapted powdery
mildew pathogen (Bgh), but other genes (Sag101, Pad4) compromise subsequent
stages in the infection process (Lipka et al., 2005). Consonni et al. (2006) reported

Chapter 4
72
Bgt
0%10%20%30%40%50%60%70%80%90%
100%
Hiproly
Tris
uli Baz
ar 9
Cham
e 2
SusPtri
tSusB
gt SC
SusBgt
DC
Vada
Vivant
Inte
ract
ion
site
s
A Bghm
0%10%20%30%40%50%60%70%80%90%
100%
Hiproly
Tris
uli B
azar
9
Chame 2
SusPtri
tSusB
gt SC
SusBgt
DC
Vada
Inte
ract
ion
site
s
B
Bgs
0%10%20%30%40%50%60%70%80%90%
100%
Hipro
ly
Trisuli B
azar
9
Chame 2
SusP
trit
SusB
gt SC
SusBgt
DC
Vada
Rogo
Inte
ract
ion
site
s
C
Bgt
0
10
20
30
40
50
Hipro
ly
Trisuli B
azar 9
Chame 2
SusPtri
t
SusBgt
SC
SusBgt
DC
Inte
ract
ion
site
s [%
]
D
Bghm
05
10152025303540
Hiproly
Tris
uli Baz
ar 9
Chame 2
SusP
trit
SusBgt
SC SusB
gt D
C
Vada
Inte
ract
ion
site
s [%
]
E
Bgs
0
5
10
15
20
25
30
Hiproly
Trisuli B
azar 9
Chame 2
SusPtri
t
SusBgt S
C
SusBgt D
CVad
a
Inte
ract
ion
site
s [%
]
F
Bgt
0
10
20
30
40
Hiproly
Tris
uli Baz
ar 9
Chame 2
SusPtri
t
SusBgt
SC SusB
gt DC
Vada
Inte
ract
ion
site
s [%
]
G
Bghm
0102030405060708090
100
Hipro
ly Tris
uli Baz
ar 9
Chame 2
SusP
trit
SusBgt
SC SusB
gt DC
Vada
Inte
ract
ion
site
s [%
]
H
Bgs
0
5
10
15
20
25
30
Hiproly
Trisuli B
azar 9
Chame 2
SusPtri
t
SusBgt S
C
SusBgt D
CVada
Inte
ract
ion
site
s [%
]
I
Figure 5 . Interaction of SusBgt lines and the parental lines with three non-adapted Bg forms. Three upper figures (A-C): defense reactions at 72 hai. White-dotted
columns (FPP+HR) represent failed primary penetration with HR. White columns (FPP-HR) represent failed primary penetration without HR. Black columns (SPP+HR)
represent the successful primary penetration with a haustorium and with HR. Cross-hatched columns (SPP-HR) represent the successful primary penetration with a
haustorium but without HR. Data represent the means of four leaf segments collected from two independent inoculation experiments. In each leaf segment, 100 infection
sites were recorded. Vivant is a wheat cultivar, and Rogo is a rye cultivar (Each is susceptible to its own mildew forms). Three middle figures (D-F): development of
secondary haustoria in the SPP infection units of A-C. Black-dotted columns represent the percentage of infection units that developed at least one secondary haustorium
at 72 hai. Three lower figures (G-I): development of conidia. The gray columns represent the percentage of established infection units that formed at least one
conidiophore at 8 dai. Data represent the means of four or two leaf segments collected from two independent inoculation experiments.
SPP-HR
SPP+HR
FPP-HR
FPP+HR

New perspectives to identify genes that…
73
Figure 6. Interaction of SusBgt lines with non-adapted Bgt mildew fungus. (A) A germling that stopped at the penetration stage in an epidermal cell, which shows an H2O2 reaction in the whole cell, indicating HR. (B) A failed germling that stopped at the penetration stage in association with papilla formation. (C) A successfully penetrating germling that elicited HR. (D) A successfully established germling that formed elongated secondary hyphae (ESH). (E) Multiple haustorium formation (arrows) in an epidermal cell where the primary haustorium was formed. (F) Failed secondary penetrations (arrows) along an ESH. (G) An established colony of Bgt at 8 dai on SusBgt DC that did not result in conidiation. (H) Abundant conidiation by an established colony of Bgt at 8 dai on SusBgt SC. The scale bar represents 100 µm.

Chapter 4
74
Figure 7. Haustorium formation and conidiation of non-adapted Bghm and Bgar mildews on SusBgt
lines at 8 dai. (A) A successfully established germling of Bghm on SusBgt SC that resulted in formation
of a hyphal network and several secondary haustoria (arrows). (B) Abundant conidiation by an
established colony of Bghm at 8 dai on SusBgt SC. (C) An established colony of Bgar at 8 dai on
SusBgt DC that did not result in conidiation. (D) Abundant conidiation by an established colony of Bgar
at 8 dai on SusBgt SC. The scale bar represents 100 µm.
that the Arabidopsis Atmlo2 pen1 double mutant allows more cell entry by
Golovinomyces cichoracearum, but no significant increase in conidiophore
production. For those later stages of development, apparently EDS1, SAG101 and
PAD4 need to be silenced.
3-Genes for basal resistance are mildew forma specialis-specific. SusBgt lines and
SusPtrit are remarkably accessible to Bgt and allow relatively high frequencies of
fungal establishment. Yet, they show a relatively high level of basal resistance to the
adapted powdery mildew pathogen, Bgh. Vada had a high level of resistance to Bgt,
but was not equally resistant to the rye mildew Bgs. The factors in SusBgt DC that
hampered conidiation by Bgt compared to SusBgt SC (Figure 5G) seem to also be
effective against Bgar (Figure 7C and D), but not against Bghm (Figure 5H and 7B) or
Bgs (Figure 5I). Selection for high susceptibility to Bgt has not affected the level of
resistance to Bga and other non-adapted mildews.
Our results in the present study show many parallels with the barley–rust interaction
(Atienza et al., 2004; Jafary et al., 2006; Marcel et al., 2007; Jafary et al., 2008). In

New perspectives to identify genes that…
75
both pathosystems, accumulation of genes for susceptibility to non-adapted pathogen
forms and species appears feasible. Non-host resistance to rust species and to non-
adapted formae speciales of powdery mildew seems similar to quantitative, basal host
resistance, and the resistance mechanism in both pathosystems is mainly based on
non-hypersensitive mechanisms.
Powdery mildew pathogens should suppress PAMP-triggered immunity (PTI) to
establish basic compatibility. This suppression of PTI requires compatibility between
the effectors and the operative targets in the plant. Effectors (O’Connell & Panstruga,
2006) and their operative targets in the plant (van der Hoorn & Kamoun, 2008) both
seem to be under strong diversifying selection. For each plant species, a different
bouquet of effectors may be required to suppress PTI (Almeida et al., 2009; Niks &
Marcel, 2009), implying that each mildew form probably has a different set of
effectors, each tuned to the operative targets in their host plant species. Depending
on the fungal development stage, different aspects of defense should be
reprogrammed, again requiring different effectors. By common evolutionary ancestry,
however, some compatible variants of operative target motifs may occur in some non-
host plant accessions. Non-adapted mildews cannot be expected to suppress PTI of
barley effectively, but, in the SusBgt lines, we may have accumulated variants of the
targets that can be suppressed relatively easily by Bgt.
New evidence has emerged on the involvement of some candidate genes in the
basal host and non-host resistance of barley against mildew pathogens (Douchkov et
al., 2005; Miklis et al., 2007). An example of such a candidate gene is HvSNAP34,
which plays a role in basal host and non-host resistance of barley to Bg. Transient-
induced gene silencing (TIGS) of this candidate gene strongly increased the rate of
successful haustorium formation of both adapted (Bgh) and non-adapted powdery
mildew fungi (Bgt) (Douchkov et al., 2005). HvSNAP34 could be one of the operative
targets of effectors to enhance successful establishment of haustoria by invading
mildew fungi. We are now developing mapping populations of SusBgt SC × Vada and
SusBgt DC × Vada to map the QTLs (genes) that determine the level of basal
resistance to the various mildew forms used in this study, similar to what was done for
rusts in Jafary et al. (2006). This will enable us to identify the targets involved in non-
host immunity in barley against mildew pathogens. Finally, the existence of relatively
high levels of susceptibility of SusBgt lines to more than one forma specialis of Bg,
may facilitate the cross-hybridization between different ff. spp. of Bg, as done on the
shared host Triticum urartu for crossing f.sp. tritici with f.sp. agropyri by Tosa (1989).
Such pathogen and plant inheritance studies can contribute considerably to our

Chapter 4
76
understanding of the evolutionary and genetic basis of the host-pathogen interactions
in Gramineous powdery mildew pathosystems. Acknowledgements
Anton Vels is gratefully acknowledged for his excellent technical assistance in greenhouse experiments. We thank Dr. Patrick Schweizer (Leibniz-Institute of Plant Genetics and Crop Plant Research (IPK), Germany) for providing the wheat isolate of mildew. Bert Essenstam from the Unifarm of Wageningen University is acknowledged for providing the cucumber powdery mildew and multiplication of wheat powdery mildew. We thank Dr. Harold E. Bockelman (National Small Grains Collection, USDA, USA) for providing the seeds of barley cv. Turkey 290, the Centre for Genetic Resources, the Netherlands (CGN) for proving the seeds of 54 accessions of barley, and Dr. Rajeev K. Varshney from IPK, Germany, for providing the seeds of 227 accessions of barley (ICARDA collection) used in this study. ICARDA (Syria) is acknowledged for its indirect contribution by preparing the seeds. We thank Dr. Viktor Korzun and Clemens Springmann (PLANTA Angewandte Pflanzengenetik und Biotechnologie, Germany) for making the DHs. We thank Prof. Richard G.F. Visser for critically reading the manuscript and for his continuous support.
References
Atienza S, Jafary H, Niks RE. 2004. Accumulation of genes for susceptibility to rust fungi for which barley is nearly a nonhost results in two barley lines with extreme multiple susceptibility. Planta 220: 71-79.
Block A, Li G, Fu Z, Alfano JR. 2008. Phytopathogen type III effector weaponry and their plant targets. Current Opinion in Plant Biology 11: 396-403.
Carver TLW, Lyngkjær MF, Neyron L, Strudwicke CC. 1 999. Induction of cellular accessibility and inaccessibility and suppression and potentiation of cell death in oat attacked by Blumeria graminis f.sp. avenae. Physiological and Molecular Plant Pathology 55: 183–196.
Chisholm ST, Coaker G, Day B, Staskawicz BJ. 2006. Host-microbe interactions: shaping the evolution of the plant immune response. Cell 124: 803-814.
Collinge DB, Jensen MK, Lyngkjaer MF, Rung J. 2008. How can we exploit functional genomics approaches for understanding the nature of plant defences? Barley as a case study. European Journal of Plant Pathology 121: 257-266.
Consonni C, Humphry ME, Hartmann HA, Livaja M, Durn er J, Westphal L, Vogel J, Lipka V, Kemmerling B, Schulze-Lefert P, et al. 2006. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nature Genetic 38: 716–720.
Douchkov D, Nowara D, Zierold U, Schweizer P. 2005. A high-throughput gene-silencing system for the functional assessment of defense-related genes in barley epidermal cells. Molecular Plant-Microbe Interactions 18: 755-761.
Eichmann R, Schultheiss H, Kogel KH, Hückelhoven R. 2004. The barley apoptosis suppressor homologue bax inhibitor-1 compromises nonhost penetration resistance of barley to the inappropriate pathogen Blumeria graminis f.sp tritici. Molecular Plant-Microbe Interactions 17: 484–490.
Eshed N, Wahl I. 1970. Host ranges and interrelations of Erysiphe graminis hordei, E. graminis tritici, and E. graminis avenae. Phytopathology 60: 628-634.
Hardison JR. 1944. Specialization of pathogenicity in Erysiphe graminis on wild and cultivated grasses. Phytopathology 34: 1–20.
Heath MC. 2000. Nonhost resistance and nonspecific plant defences. Current Opinion in Plant Biology 3: 315-319.
Hückelhoven R. 2007. Cell wall-associated mechanisms of disease resistance and susceptibility. Annual Review of Phytopathology 45: 101–127.
Hückelhoven R, Dechert C, Kogel KH. 2001. Non-host resistance of barley is associated with hydrogen peroxide burst at sites of attempted penetration by wheat powdery mildew fungus. Molecular Plant Pathology 2: 199-205.

New perspectives to identify genes that…
77
Jafary H, Albertazzi G, Marcel TC, Niks RE. 2008. High diversity of genes for nonhost resistance of barley to heterologous rust fungi. Genetics 178: 2327–2339.
Jafary H, Szabo LJ, Niks RE. 2006. Innate nonhost immunity in barley to different heterologous rust fungi is controlled by sets of resistance genes with different and overlapping specificities. Molecular Plant–Microbe Interactions 19: 1270–1279.
Koga H, Bushnell WR, Zeyen RJ. 1990. Specificity of cell type and timing of events associated with papilla formation and the hypersensitive reaction in leaves of Hordeum vulgare attacked by Erysiphe graminis f.sp. hordei. Canadian Journal of Botany. 68: 2344–2352.
Lin MR, Edwards HH. 1974. Primary penetration process in powdery mildewed barley related to host cell age, cell type, and occurrence of basic staining material. New Phytologist 73: 131-137.
Lipka V, Dittgen J, Bednarek P, Bhat R, Wiermer M, Stein M, Landtag J, Brandt W, Rosahl S, Scheel D, et al. 2005. Pre- and postinvasion defences both contribute to nonhost resistance in Arabidopsis. Science 310: 1180-1183.
Mains EB, Dietz SM. 1930. Physiologic forms of barley mildew, Erysiphe graminis hordei Marchal. Phytopatlogy 20: 229-239.
Marcel TC, Varshney RK, Barbieri M, Jafary H, de Ko ck MJD, Graner A, Niks RE. 2007. A high-density consensus map of barley to compare the distribution of QTLs for partial resistance to Puccinia hordei and of defence gene homologues. Theoretical and Applied Genetics 114: 487–500.
Menzies JG, MacNeill BH. 1989. Infection of species of the Gramineae by Erysiphe graminis f.sp. hordei and Erysiphe graminis f.sp. tritici. Canadian Plant Disease Survey 69: 105-108.
Miklis M, Consonni C, Bhat RA, Lipka V, Schulze-Lef ert P, Panstruga R. 2007. Barley mlo modulates actin-dependent and actin-independent antifungal defense pathways at the cell periphery. Plant Physiology 144: 1132–1143.
Mysore KS, Ryu CM. 2004. Nonhost resistance: how much do we know? Trends in Plant Science 9: 97-104.
Niks RE, Marcel TC. 2009. Non-host and basal resistance: how to explain specificity? New Phytologist 182: 817–828.
Nurnberger T, Brunner F, Kemmerling B. Piater L. 20 04. Innate immunity in plants and animals: striking similarities and obvious differences. Immunological Reviews 198: 249-266.
O'Connell RJ, Panstruga R. 2006. Tête à tête inside the plant cell: establishing compatibility between plants and biotrophic fungi and oomycetes. New Phytolologist 171: 699-718.
Olesen KL, Carver TLW, Lyngkjær MF. 2003. Fungal suppression of resistance against inappropriate Blumeria graminis formae speciales in barley, oat and wheat. Physiological and Molecular Plant Pathology 62: 37-50.
Peterhänsel C, Freialdenhoven A, Kurth J, Kolsch R, Schulze-Lefert P. 1997. Interaction analyses of genes required for resistance responses to powdery mildew in barley reveal distinct pathways leading to leaf cell death. Plant Cell 9: 1397–1409.
Schweizer P. 2007. Nonhost resistance of plants to powdery mildew-New opportunities to unravel the mystery. Physiological and Molecular Plant Pathology 70: 3–7.
Thordal-Christensen H, Zhang Z, Wei Y, Collinge DB. 1997. Subcellular localization of H2O2 in plants: H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant Journal 11: 1187–1194.
Thordal-Christensen H. 2003. Fresh insights into processes of non-host resistance. Current Opinion in Plant Biology 6: 351-357.
Tosa Y, Shishiyama J. 1984. Defense reactions of barley cultivars to an inappropriate forma specialis of the powdery mildew fungus of gramineous plants. Canadian Journal of Botany 62: 2114–2117.
Tosa Y. 1989. Genetic analysis of the avirulence of wheatgrass powdery mildew fungus on commons wheat. Genome 32: 913-917.
Trujillo M, Troeger M, Niks RE, Kogel KH, Hückelh oven R. 2004 . Mechanistic and genetic overlap of barley host and non-host resistance to Blumeria graminis. Molecular Plant Pathology 5: 389-396.
van der Hoorn RAL, Kamoun S. 2008. From guard to decoy: a new model for perception of plant pathogen effectors. Plant Cell 20: 2009-2017.
Wolf G, Fric F. 1981. A rapid method for staining Erysiphe graminis f.sp. hordei in and on whole barley leaves with a protein-specific dye. Phytopathology 71: 596-598.

New perspectives to identify genes that…
73

Rust)induced powdery mildew resistance
79
Rust-induced powdery mildew resistance in barley
R. Aghnoum, R.G.F. Visser, P. Schweizer and R.E. Niks

Chapter 5
80
Abstract Different pathogens employ different strategies to suppress their host defense
mechanisms. We observed that a compatible interaction of barley-Puccinia hordei
induced increased papilla-based resistance to a challenge infection by a compatible
powdery mildew isolate despite the different strategies of suppression of defense by
the two organisms. The level of rust-induced mildew resistance varies among barley
accessions and is not determined by the virulent/avirulent spectra of the challenger
isolate. Our histological data suggest that the inducer effect is due to rust-plant
communication during stoma penetration by the rust fungus, or to the presence of the
rust hyphae in the apoplast, or to some degree of pre-haustorial resistance mounted
by the mesophyll cells to the invading rust fungus. Macroarray gene expression
analysis showed that several genes related to metabolism and photosynthesis were
highly down regulated and a few genes involved in plant defense including a
pathogenesis related protein (PR-1), and Cysteine synthase were highly up regulated
in the double inoculated treatment. Both our histological and gene expression
analysis suggest that the rust “primes” the basal mildew resistance genes prior to the
challenge mildew infection.
Introduction Infection by a pathogen or parasite may condition plants to activate or suppress
defense reactions against subsequent infection attempts by other pathogen strains or
species. Induction of plant defense systems as a result of prior exposure to
pathogens (Ryals et al., 1996), non-pathogenic root-colonizing bacteria (van Loon et
al., 1998), feeding by herbivores (Kessler & Baldwin, 2002) or treatment with
chemicals (Oostendorp et al., 2001) is termed induced resistance. Three types of
induced resistance have been described, viz. systemic acquired resistance (SAR)
(see Glossary) (Durrant & Dong, 2004), induced systemic resistance (ISR) (see
Glossary) (van Loon et al., 1998) and induced localized resistance (Kunoh, 2002).
Suppression of plant defense systems against a non-adapted or incompatible race of
a pathogen as a result of prior exposure to a compatible pathogen is called induced
susceptibility.
Induced localized resistance and induced susceptibility have been reported in many
plant-pathogen combinations (Table 1). In cereals induction of localized susceptibility
or resistance has been reported as a result of a prior attack by both necrotrophic and
biotrophic fungi (e.g. Jørgensen et al., 1998; Kunoh et al., 1988; Lyngkjær & Carver,
1999; Niks, 1989). In cereals, most research on localized induction of resistance and
susceptibility has concentrated on interaction of compatible and incompatible isolates

Rust)induced powdery mildew resistance
81
of the powdery mildew pathogen, Blumeria graminis (hereafter Bg). Failed
establishment (see Glossary) by an adapted or non-adapted powdery mildew
pathogen induces penetration resistance to subsequent challenge inoculation with a
compatible isolate (e.g. Hwang & Heitefuss, 1982; Cho & Smedegaard-Petersen,
1986; Carver et al., 1999; Lyngkjær & Carver, 1999; Kunoh, 2002). Conversely,
successful penetration and haustorium formation by an adapted, virulent isolate of Bg
induces susceptibility at cell level to haustorium formation by subsequent challenge
with adapted virulent or avirulent isolates and also non-adapted formae speciales and
species (e.g. Tsuchiya & Hirata, 1973; Carver et al., 1999; Lyngkjær et al., 2001;
Olesen et al., 2003). “Susceptibility” at cell level is often called accessibility, because
the avirulent or non-adapted challenger pathogen typically will not complete its life
cycle, due to defense mechanisms that back-up the penetration resistance. For
similar reasons “resistance” at cell level is often called inaccessibility. Most of the
reports indicate that this induction of accessibillity and inaccessibility is localized and
limited to the challenged cell and its adjacent cells.
Induced resistance to rust pathogens as a result of prior inoculation with the same
or different rust pathogens has been reported by several workers (e.g. Kochman &
Brown, 1975; Sebesta et al., 1996; Calonnec, et al. 1996; Pei et al. 2003). In the
barley leaf rust (Puccinia hordei) system, failed haustorium formation by an non-
adapted rust fungus, however, does not induce reduced haustorium formation by a
compatible challenger rust (Niks, 1989) but a compatible interaction suppresses the
defense system of the same cell at challenge with a non-adapted rust species.
Powdery mildew and rust fungi both have biotrophic lifestyles and require living
plant tissues to survive and reproduce. Successful establishment of specialized
feeding structures of the pathogen, the so-called haustoria, is a prerequisite for a
compatible interaction. In barley, non-host resistance against non-adapted powdery
mildews, the quantitatively inherited basal resistance and mlo-mediated resistance to
the adapted powdery mildew all act on the basis of papilla formation at the point of
attempted ingress into the cell, which results in impaired haustorium formation. Papilla
formation is involved in induced inaccessibility in the barley-powdery mildew (Sahashi
& Shishiyama, 1986). A similar papilla formation is involved in basal and non-host
resistance against rust fungi (Niks, 1986, 1989).
Mellersh & Heath (2001) reported evidence that mildews and rust fungi follow
different strategies to successfully suppress plant defense, and hence induce
accessibility of plant cells. Rust fungi should decrease adhesion between the cell wall
and the plasma membrane at the cell wall penetration point, but for successful
infection by mildew no such decreased adhesion is required. We set out to determine

Chapter 5
82
whether a compatible rust interaction would induce increased accessibility to mildew,
despite the different strategies of suppression of defense by the two organisms. We
found, however, in exploring experiments, that compatible infections by Puccinia
hordei induced increased papilla based resistance rather than decreased papilla
based resistance to a challenge infection by a compatible powdery mildew isolate. In
this paper we characterize this rust-induced mildew resistance at the cellular level.
We also identified differentially expressed genes that can explain the strong papilla-
based penetration resistance in double inoculated barley plants.
Materials and Methods
Plant material Three barley near isogonics lines (Pallas, Pallas-Mla3, Pallas-MlLa), the mildew
susceptible barley cv. ‘Manchuria’, two barley lines that possess high levels of basal
resistance to leaf rust (Vada and 17-5-16), and three near-isogenic barley lines with
different genotypes of MlLa and Rphq2, the latter being a QTL for basal resistance to
leaf rust (Marcel et al., 2007) (Vada rphq2 mlLa, Vada rphq2 MlLa, and Vada Rphq2
mlLa), were used to study the effect of pre-inoculation with leaf rust on powdery
mildew development. Three seeds of each genotype were sown in trays of 37×39 cm
in two rows on the right and left side of each tray. Plants were grown in a greenhouse
compartment where the temperature was set at 18-20°C with 16 hours of light and a
relative humidity of about 70%.
Pathogen material The isolate 1.2.1 of Puccinia hordei was used as the ‘inducer’. This isolate is able to
establish a compatible interaction with all tested barley lines. Two isolates of Blumeria
graminis f.sp. hordei, isolate C15 and Wag.04, were used as ‘challenger’. Isolate C15
is avirulent to Pallas-MlLa but virulent to Pallas-Mla3. Wag.04 isolate is the reverse,
virulent to Pallas-MlLa but avirulent to Pallas-Mla3. Leaf rust isolate 1.2.1 was
maintained on line L94, and the powdery mildew isolates on Manchuria. An isolate of
Puccinia recondita, the rye leaf rust pathogen, freshly propagated on a susceptible
rye genotype and an isolate of Puccinia antirrhini, the snapdragon rust, collected from
naturally infected common snapdragon (Antirrhinum majus) in Wageningen were
used in this study.
Inducer inoculation Ten days after sowing, the fully expanded first seedling-leaves were positioned
horizontally, adaxial side up. In the preliminary experiments, prior to the inoculation

Rust)induced powdery mildew resistance
83
with rust, the middle part of each leaf was covered with a 3 cm strip of aluminum foil
to keep that middle section free of rust infection, in order to determine whether the
effect of inducer inoculation on the challenger is localized or systemic. In each tray
only the plants of one row were inoculated by the inducer and the plants of the other
row were covered to keep them free of rust. For each box, 6 mg of fresh
urediospores of P. hordei isolate 1.2.1 were diluted ten times with Lycopodium
powder and dusted over the plants in a settling tower. This inoculum dose results in
about 350 urediospores per cm2. Inoculated plants were incubated overnight in a
dark chamber with 100% humidity and then transferred to a greenhouse
compartment with the above mentioned conditions.
Challenger inoculation Five days after the inoculation with the inducer, plants were subjected to inoculation
by the challenger mildew. A settling tower was used for inoculation. Freshly produced
conidia of powdery mildew were blown into the tower over the horizontally fixed
adaxial sides of the first leaves. The density of inoculum was adjusted to 100-150
conidia per cm2. The inoculated plants were incubated in a growth chamber at 18-20 oC with fluorescent light irradiance (100 micromol/m2/s) supplied for 16 h per day.
Histological assessment of challenger infection Forty eight hour after inoculation (hai) of the challenger, a leaf segment of about 3 cm
long was collected from the basal side of the rust-mildew inoculated portion and the
whole rust-free middle portion. From the control plants a leaf segment of about 3 cm
long was collected from the middle part of the leaves. From each genotype, two
seedlings were sampled and one seedling was kept to monitor the efficiency of
inoculation and infection type of the powdery mildew at macroscopic level. Infection
types (IT) of powdery mildew were scored based on the 0–4 scale proposed by
Mains & Dietz (1930) on those plants that were only inoculated with the challenger.
The leaf segments were submerged in a solution of acetic acid-ethanol (3/1 v/v) at
least for 3 hours to be cleared. They were stained by boiling for 8 min in a solution of
alcoholic lactophenol (96% ethanol-lactophenol 1/1 v/v) containing 0.1 mg ml-1
Trypan Blue and then cleared in a Chloral Hydrate solution (2.5 mg ml-1) overnight
(Peterhänsel et al., 1997). Those infection units that were stopped at penetration
stage in association with papilla deposition were considered failed primary
penetration (FPP) and infection units that succeeded to penetrate the epidermal cells
and established a haustorium and an elongated secondary hypha (ESH) were

New perspectives to identify genes that…
122
1 Induced resistance (IR) and induced susceptibility (IS) have been used for the effect of inducer on challenger development on entire leaves, whole plants or at cell level 2 ISR, Induced systemic resistance; 3nhR, non-host resistance; 4Bga, Bgh and Bgt stand for Blumeria graminis f.sp. avenea, hordei and tritici respectively
Table 1: A summary of published literature on pathogen-induced susceptibility and resistance
Crop Inducer Challenger Plant-inducer interaction
Plant-challenger interaction
Outcome of interaction 1 Reference(s)
Bean Uromyces phaseoli Uromyces phaseoli S S IR Yarwood 1954 Melon Sphaerotheca fuliginea Bgh4, Bgt4 S nhR3 IS Oku & Ouchi 1976
Rice Pyricularia oryzae Bipolaris sorokiniana
Pyricularia oryzae Pyricularia oryzae
R nhR
S S
IR IR
Manandhar et al., 1998
Oat Puccinia triticina P. graminis tritici
P. coronata avenae P. graminis avenae
nhR nhR
S S
IR IR
Kochman & Brown 1975
Oat Oat
Bga4 Bga Bga
Bga Bga Bgh,Bgt
S R S
S S nhR
IS IR IS
Carver et al., 1999; Lyngkjær & Carver 2000 Olesen et al., 2003
Wheat Puccinia striiformis P. striiformis R S IR
Calonnec et al., 1996
Wheat Septoria tritici Septoria tritici R S IR Halperin et al., 1996
Wheat Bgt Bgh
Bgh,Bga Bgt
S R
nhR S
IS IR
Olesen et al., 2003 Schweizer et al., 1989
Barley F. oxysporum f.sp. radicis-lycopersici Bgh nhR S ISR2 Nelson 2005 Barley Cladosporium macrocarpum Bgh Saprophyte S IR Gregersen & Smedegaard, 1989
Barley Puccinia hordei P. recondita f.sp. recondita(Prr)
Prr P. hordei
S nhR
nhR S
IS No IR
Niks, 1989
Barley Bipolaris maydis Septoria nodorum
Drechslera teres Drechslera teres
nhR nhR
S S
IR IR
Jørgensen et al., 1998
Barley Bgh Sph. fuliginea S nhR IS Oku & Ouchi, 1976 Barley Bgh Erysiphe pisi S nhR IS Kunoh et al., 1988; Kunoh, 2002 Barley Bgh Various PM fungi S nhR IS Tsuchiya & Hirata, 1973 Barley Bgh Bgt,Bga S nhR IS Ouchi et al., 1976a; Olesen et al., 2003 Barley Erysiphe pisi Bgh nhR S IR Kunoh et al., 1988; Kunoh, 2002
Barley Bgt Bgh nhR S IR Ouchi et al., 1976a; Ouchi et al., 1976b; Gregersen and Smedegaard 1989
Barley Bgh Bgh R S IR Cho and Smedegaard-Petersen, 1986; Lyngkjær &
Carver, 2000; Lyngkjær et al., 2001
Barley
Bgh Bgh S S /R IS Chin et al., 1984; Lyngkjær et al., 2001; Lyngkjær &
Carver, 2000
84

Rust)induced powdery mildew resistance
85
considered as successful primary penetration (SPP) (Asher & Thomas, 1983).
Because different types of epidermal cells show different reactions to powdery
mildew infection (Lin & Edwards, 1974) only stomatal and interstomatal cells (type A
and type B epidermal cells following the terminology of Koga et al., 1990) were
examined, and long epidermal cells over-lying vascular tissues (type C) were not
considered. In each leaf segment 150 powdery mildew germlings were scored. Only
those epidermal cells attacked by a single mildew fungal germling were considered.
Preparations were examined at a magnification of 400× using a Zeiss Axiophot
microscope, using transmitted white light.
Transcriptome analysis of rust-induced mildew resis tance In order to study defense-related gene expression in barley that was double
inoculated with rust and powdery mildew, a macroarray expression analysis was
carried out. Barley line 17-5-16 (Parlevliet & Kuiper, 1985) which showed in our
histological experiments a high level of rust-induced penetration resistance to
powdery mildew was selected for this gene expression study. Treatments were as
follows: treatment 1, inducer (P. hordei, isolate 1.2.1.) + challenger (Bgh, isolate
C15); treatment 2, only inducer; treatment 3, only challenger; treatment 4, mock, no
inoculation, only Lycopodium powder applied as for the other treatments. About 60
plants were grown in each box (37 × 39 cm). For each type of treatment one box was
used. Ten days after sowing, first formed leaves were fixed in a horizontal position,
with the abaxial side up. The inoculation procedure, the density of inducer and
challenger and the incubation conditions were as in the previous experiments.
Twelve hai, 15 leaves were sampled per treatment. Leaf samples were shock-frozen
in liquid nitrogen immediately and stored at –80 oC. The experiment was carried out
in three independent inoculation experiments in a randomized block design.
The barley PGRC2 cDNA array We used the barley PGRC2 cDNA macroarray, which is an extended version of the
PGRC1 array described by Schweizer (2008). These cDNA arrays have been
developed at IPK (Leibniz Institute of Plant Genetics and Crop Plant Research,
Gatersleben, Germany) and contain approximately 13,000 and 10,000 unigenes
respectively, spotted on two membranes.
RNA isolation Total RNA was extracted from whole-leaf material sampled at 12 hai. About 10 leaf
samples were ground in liquid nitrogen and about 100 mg of the homogenized tissue

Chapter 5
86
samples was used for RNA extraction with TRIZOL® Reagent according to the
manufacturer’s protocol with some modification of the procedure, viz. the aqueous
phase was extracted twice with chloroform.
Synthesis of 33P-labelled cDNA and hybridization procedures mRNA was extracted from 37.5 µg of the total RNA using Dynal magnetic beads
according to the manufacturer’s protocol. 33P-labelled second-strand cDNA probes
were prepared as described by Sreenivasulu et al. (2002). cDNA arrays were first
pre-hybridized and then hybridized with cDNA probes as described by Sreenivasulu
et al. (2002). The hybridization period was prolonged to 18 h. as suggested by
Zierold et al. (2005).
Spot detection and data normalization The hybridized membranes were scanned by the phosphorimager (Fuji BAS3000
reader). Spots were detected and spot intensities were quantified by using the
ArrayVision 8.0 software package (Imaging Research Inc., St. Catharines, Ontario,
Canada). Signal intensities were normalized after background subtraction as
described by Sreenivasulu et al. (2002). Only unigenes with a signal/background
ratio of more than 2.0-fold in at least two hybridizations were considered.
Differentially regulated genes were identified by calculating the Q value (false
discovery rate) by using the EDGE (Extraction of Differential Gene Expression)
software (Leek et al. 2006). Transcripts that showed a Q-value of less than 0.2
(FDR<20%) and a p-value of less than 0.05 between sample pairs were selected for
further analysis.
Results
Effect of prior infection with leaf rust on powdery mildew development Induction of localized inaccessibility to powdery m ildew by inducer
The leaf rust fungus (inducer) was able to establish a compatible interaction with all
tested barley genotypes. At five days after inoculation (dai) with the inducer, when
the plants were subjected to the attack by challenger, the leaf rust had formed pale
green flecks and at 8 dai mature rust pustules had formed.
On the control plants that were only inoculated with challenger mildew, depending
on the barley genotype, 23 to 41% of the infection units were stopped at penetration
stage in association with papilla formation. When the challenger isolate was avirulent
to the accession that carry corresponding R gene, the successfully penetrated host
cells died in response to the attack. The inducer inoculation caused a significant

Rust)induced powdery mildew resistance
87
reduction in successful haustorium formation by the challenger mildew in all tested
barley accessions, but the level of induced resistance differed between the
accessions (Figure 2). The strongest effect of induced resistance was observed in
the interaction of Pallas-Mla3 with the Wag.04 mildew isolate. On Pallas-Mla3 plants
pre-conditioned by the inducer, only 9% of germlings succeeded to penetrate and
form a haustorium, versus 58 % on the control plants. In the very mildew susceptible
accession Manchuria, the lowest effect of induced resistance was observed. Data
obtained from the middle parts of the leaves that were covered during inducer
inoculation, indicated that this induction of penetration resistance hardly extends to
tissue away from the rust infections, and therefore is hardly or not systemic (Figures
1 and 2A).
Figure 1. Rust-induced mildew resistance phenotype on barley line Pallas inoculated with leaf rust and powdery
mildew (wag.04 isolate). Colonies of powdery mildew are visible on the right part of the leaf that only was
inoculated with powdery mildew; on the left side, that was inoculated with leaf rust 5 days before mildew
inoculation, hardly any colony is visible.
Induced inaccessibly is not determined by the virul ent/avirulent spectra
of the challenger isolate Because of the different levels of induced resistance on the three Pallas near-
isogenic lines (Figure 2A), an obvious question to be answered was whether the
virulence/avirulence of the challenger isolate (powdery mildew) to the Pallas near-
isogenic lines that carrying the corresponding R gene, can influence the level of
induced resistance. Similar results were obtained when we used another powdery
mildew isolate (C15) that reacted differentially with the two near-isogenic lines
(Pallas-Mla3 and Pallas-MlLa) in comparison with Wag.04 isolate (Figure 2B). This
indicates that induced resistance is not determined by the virulent/avirulent spectra of
the challenger isolate.
We tested Vada recombinants with different genotypes regarding Rphq2 and MlLa.
Rphq2 is an allele for higher basal resistance to P. hordei, rphq2 for lower basal
resistance to this rust fungus. MlLa is an allele for race-specific incomplete

Chapter 5
88
hypersensitive resistance to powdery mildew, mlLa is the susceptibility allele. The
Rphq2 and MlLa loci are linked, and recombinants between these loci have been
developed in Vada background (Marcel et al., 2007). The four combinantions of
alleles on these two loci showed similar and rather low levels of induced resistance
(Figure 3). This is preliminary evidence that the level of basal resistance to P. hordei
does not determine the level of induced resistance to powdery mildew, and that
mildew R-gene MlLa is not a determining factor either. We also tested barley
experimental line 17-5-16, a barley line that possesses a very high level of basal
resistance to leaf rust (Parlevliet & Kuiper, 1985). In this line the inducer caused a
high level of penetration resistance to the mildew challenger (Figure 3).
0
10
20
30
40
50
60
70
80
90
100
Pallas Pallas-Mla3 Pallas-MlLa Manchuria
Pen
etra
tion
eff
icie
ncy
[%]
aa
b
b
a
a
b
aa b
a a
A
0
10
20
30
40
50
60
70
80
90
100
Pallas Pallas-Mla3 Pallas-MlLa Manchuria
Pen
etra
tion
eff
icie
ncy
[%]
a
a
a
b
a
b
a
b
B
Figure 2. Penetration efficiency of powdery mildew germlings on different barley lines at 48 hai. A, Wag.04 isolate, avirulent to Pallas-Mla3 but virulent to Pallas-MlLa. B, C15 isolate, avirulent to Pallas-MlLa but virulent to Pallas-Mla3. Gray-dotted columns represent the penetration efficiency of powdery mildew when barley leaves were inoculated with the inducer (Puccinia hordei isolate 1.2.1) five days before inoculation with the challenger. White columns represent penetration efficiency in a 3 cm strip of leaves that was covered during inoculation with the inducer and were inoculated only with the challenger. Gray columns represent penetration efficiency on control plants that were inoculated only by powdery mildew at the same time as inoculation of other treatments with the challenger. In Figure 2A data of each column are based on an average of 600 mildew infection units collected from 4 leaf segments (two leaves in two experiments per accession/treatment combination). In Figure 2B data of each column are based on an average of 300 mildew infection units collected from 2 leaf segments each from an independent inoculation experiment. Means followed by the same letter within a genotype are not significantly different (p= 0.05).

Rust)induced powdery mildew resistance
89
Stoma penetration and intracellular hyphae may cont ribute to induced
resistance We determined which aspect of rust infection may contribute to induced resistance.
We applied a non-adapted rust, viz. the snapdragon rust fungus P. antirrhini. We
checked the barley leaves inoculated with this rust at microscopic level. This rust
produces only germ tubes on the barley leaf surface, and hardly or not penetrates
through stomata. Although the snapdragon rust was applied at about two times
higher inoculum dose than the leaf rust, we observed that it did not induce resistance
against powdery mildew (Figure 4). This indicates that induced resistance is not due
to communication between the rust germ tube and the barley epidermal cells. Next
we applied the non-adapted rust P. recondita as inducer. P. recondita is able to find
the stomata and penetrate through them, but hardly forms any haustorium in barley
(Niks, 1989). We found that application of P. recondita reduced the haustorium
formation by the mildew fungus from 47% in the control to 25% (Figure 4). There is a
tendency that haustorium formation is not required for the induction of resistance to
the mildew fungus, more replications are required to test whether this difference is
significant. Application of the inducer inoculum P. hordei and challenger mildew on
opposite leaf sides resulted in a similar level of induced penetration resistance to
subsequent attack by powdery mildew as when both inoculations were applied to the
adaxial leaf surface (Figure 4). This induction was not observed when P. recondita
was applied on the opposite side of the leaf. This suggests that the rust intracellular
hyphae and stoma penetration both may contribute to induced resistance.
Transcript profiling Out of approximately 13,000 unigenes spotted on the array, 6,053 unigenes showed
hybridization signals at least 2.0 fold above the local background and they were
selected for further statistical analysis. Only those unigenes that showed a Q-value
less than 0.2 between sample pairs and P value < 0.05 were considered differentially
regulated genes. In total, 285 gene/treatment combinations showed differential
expression compared with the mock inoculation (Figure 5). A considerable set (67%)
of differentially regulated genes in LR+PM was also differentially regulated in LR.
Higher numbers of genes were found to be differentially regulated in LR+PM
treatment (128) than in single inoculation treatments. On the other hand, direct
comparison of gene expression between LR (Leaf Rust) and LR+PM (Leaf Rust +
Powdery mildew) inoculations did not reveal any transcripts that were significantly

Chapter 5
90
Figure 3. Penetration efficiency of powdery mildew germlings (C15 isolate) on different near isogenic barley
lines on the background of Vada and a line with high level of basal resistant to leaf rust (17-5-16). Gray-
dotted columns represent the penetration efficiency of powdery mildew when barley leaves were inoculated
with the inducer (Puccinia hordei isolate 1.2.1) five days before inoculation with the challenger. Gray
columns represent penetration efficiency on control plants that were inoculated only by powdery mildew at
the same time as inoculation of other treatments with the challenger. Data of each column are based on an
average of 300 infection units collected from 2 leaf segments (one leaf in two independent inoculation
experiments per treatment). C15 isolate is avirulent on genotypes carrying the MlLa allele.
Figure 4. Penetration efficiency of powdery mildew germlings (Wag.04 isolate) on barley cv. Pallas
inoculated with different inducer rusts. The inducers were inoculated five days before inoculation with the
challenger. Data of each column are based on an average of 600 infection units collected from 4 leaf
segments (two leaves in two experiments per treatment). * The mean difference is significant at 0.05 level
(one sided test).
Basal resistance
to P. hordei high low low high very high
Incomplete HR
to Bgh yes no yes no no
0
10
20
30
40
50
60
70
(Rphq2 MlLa) (rphq2 mlLa) (rphq2 MlLa) (Rphq2 mlLa) 17-5-16
Pen
etra
tion
eff
icie
ncy
[%]
0
10
20
30
40
50
60
Only mildew P. hordei P. hordeidownside
P. recondita P. recondita downside
Snapdragonrust
Pen
etra
tion
eff
icie
ncy
[%] *
*

Rust)induced powdery mildew resistance
91
different in their abundance, indicating the existence of quantitative rather than
qualitative differences between induced but not challenged and induced-challenged
leaves. Such differential expression patterns may have escaped detection due to the
limited number of samples used for the analysis. To assess the quantitative
difference in normalized signal intensities (Int) of differentially regulated genes
between double inoculation treatment (LR+PM) and single mildew inoculation (PM) in
a more robust manner, a differential index (DI; Zierold et al., 2005) was calculated for
each gene as follows:
DI = (IntLR+PM − IntPM)/ (IntLR+PM + IntPM)
DI > 0 indicates genes that on average show higher transcript abundance in LR+PM
compared to PM. DI < 0 indicates genes that on average show higher transcript
abundance in PM than in LR+PM. The genes with the greatest difference in
expression level between LR+PM and PM showing a DI > 0.33 or DI < -0.33, which
corresponds to a two-fold signal difference on average, are listed in Table 2. The
results indicate that there are great differences in transcript abundance among the
genes with different functions. The majority of the differentially regulated genes found
to be involved in metabolism and photosynthesis and a few genes in disease
resistance (Table 2 and Figure 6). Up-regulation of a pathogenesis related protein
(PR-1), a marker for Systemic Acquired Resistance (SAR) and Cysteine synthase
which is involved in balancing the production of ROS (reactive oxygen species) was
observed in double inoculated treatment and in single rust treatment (Table 2, Figure
6).
Figure 5. Venn diagram representation of overlapping and non-overlapping sets of differentially
regulated genes (versus mock) between different treatments in barley line 17-5-16.
14 23
30
1
55 3
40
Powdery mildew (48) Leaf rust (109)
Leaf rust + powdery mildew (128)

Chapter 5
92
Table 2. Differentially regulated genes in double inoculated ( leaf rust + powdery mildew)
barley plants versus powdery mildew inoculated plants Clone ID Blast X description 1 Regulation DI 2
HD01N05 Pathogenesis-related protein up 0.97
HO16H10 PR-1, a pathogenesis related protein up 0.97
HO06C23 Protease inhibitor up 0.93
HW03J02 Hypothetical protein up 0.66
HO08E02 Cysteine synthase up 0.66
HO05I23 Ribulose-1,5-bisphosphate carboxylase down -0.33
HO03F06 (TPR)-containing protein down -0.34
HO05F14 Ribulose-1,5-bisphosphate carboxylase down -0.34
HB27P08 Phosphoethanolamine methyltransferase down -0.34
HO28H23 Fructose-bisphosphate aldolase down -0.34
HE01K15 Hypothetical protein down -0.36
GCW002K24 Unknown down -0.37
HO19N07 Chlorophyll a/b binding protein down -0.37
HO04E03 Ribulose-1,5-bisphosphate carboxylase down -0.38
HO02G01 Ribulose-1,5-bisphosphate carboxylase down -0.40
HZ48I22 Photosystem I antenna protein down -0.41
HM11H23 Photosystem I antenna protein down -0.42
HO01J24 Ribulose-1,5-bisphosphate carboxylase down -0.45
HB21N18 Chlorophyll a/b binding protein down -0.48
HO01D09 Chlorophyll a-b binding protein down -0.49
HE01E12 Chlorophyll a/b binding protein 2 down -0.50
HZ53A20 Metallothioneine type2 down -0.50
HO36J03 Carbonic anhydrase down -0.51
HA06N06 Fructose-bisphosphate aldolase down -0.51
HO38I13 Carbonic anhydrase down -0.52
HO02J18 Putative GTP-binding protein down -0.53
HG01D09 Carbonic anhydrase down -0.53
HA12L17 Phosphoglycerate kinase down -0.56
HV04H14 Putative thiamine biosynthesis protein down -0.57
HO28I13 Carbonic anhydrase down -0.57
HDP10O13 Photosystem II 10 kDa polypeptide down -0.76
HG01P09 Unknown down -0.80
HV03C20 Metallothionein-like protein type 3 down -0.80
HD08N06 Metallothionein-like protein down -0.81 1Description of the most significant Blast X hit (the lowest E value). 2Differential index of transcript abundance between double inoculated and single mildew inoculated plants. Only genes that showed at least 2-fold difference in transcript abundance were presented.

Rust)induced powdery mildew resistance
93
Discussion Our histological data indicated that pre-inoculation of barley with leaf rust increases
the efficiency of the barley defense system to arrest the establishment of the
challenger mildew fungus. This induction of resistance is localized rather than
systemic. The lack of induction by the germinating snapdragon rust suggests that the
effect is not caused by the rust germ tubes growing over the epidermis. The inducing
effect by the rye leaf rust in barley suggests that rust haustoria are not essential
either. We presume that the inducer effect is due to rust-plant communication during
stoma penetration by the rust fungus, or to the presence of the rust hyphae in the
apoplast, or to some degree of pre-haustorial resistance mounted by the mesophyll
cells to the invading rust fungus.
The level of induced mildew resistance varies among barley accessions. Although
great differences in level were found among Pallas near-isogenic lines (Figure 2),
suggesting a role of the Ml genes in these lines, the virulence or avirulence of the
challenging mildew apparently did not matter (Figure 2A versus 2B). The high level of
induced resistance observed in Pallas-Mla3 and Pallas-MlLa compared to Pallas may
be due to some other genetic difference between these NILs either linked to the Ml
gene, or elsewhere on the genome. Also we did not find sufficient evidence that level
of basal resistance to P. hordei determines the level of induced resistance. It is
intriguing that the experimental line 17-5-16 that shows a high level of basal
resistance to P. hordei, shows also a high level of induced resistance. However, this
may be a co-incidental association, since the two Vada NILs with a decreased level
of basal resistance (rphq2), did not induce a convincingly lower level of induced
resistance than the two lines carrying the alleles Rphq2 for higher level of basal
resistance (Figure 3). The control treatment inoculated only with the PM suggests that the lower the basal
resistance to powdery mildew (Manchuria, Figure 2), the lower the induction of
penetration resistance by the rust. This possible association should be verified in
larger sets of barley accessions, for which we might use the material characterized in
Chapter 2 and 3, showing a wide range of levels of such basal resistance. If indeed
the level of rust induced resistance depends on the level of basal resistance to
mildew, an interesting hypothesis would be that the rust “primes” the basal mildew
resistance genes prior to the challenge mildew infection. Such a “priming” of defense
genes (ISR) has been reported to occur in plants that were colonized by
Pseudomonas fluorescence (Verhagen et al., 2004).

Chapter 5
94
0
1
2
3
4
PR-1
Pathogen
esis-
relat
ed p.
Protea
se in
h ibitor
Cystein
e synth
ase
Hypo th
etical
p rote
in
Log
sign
al in
tens
ity
Mildew LR LR+PM
A
0
1
2
3
4
Pho t. II 1
0 kDa
Carbon
i c anhydra
se
Phosp
hoglyc
erate
kinas
e
Chloro
phyll a/b
b. p
.
Ribulose-1
,5-b. c
.
Log
sign
al in
tens
ity
B
Figure 6 Examples of highly up-regulated and down-regulated host genes in PM, LR and LR+PM treatments. A,
five genes with the high DI and B, five genes with the low DI. Data represent mean of signal intensities of three
inoculation experiments.
Macroarray transcriptome analysis To get insight into the molecular basis of the rust-induced mildew resistance
phenomenon, we compared the global gene expression profile of double inoculated
barley plants with that of single inoculated ones.
We found a considerable level of overlap in differentially regulated genes in double
inoculated plants with that of single powdery mildew or rust inoculated ones (25%
and 67% respectively) although these two pathogens are adapted to live in different

Rust)induced powdery mildew resistance
95
host tissues. P. Schweizer (IPK, Germany, unpublished data) found a considerable
overlap in transcriptional response between entire leaf samples challenged with
powdery mildew or rust, suggesting that rust and powdery mildew affect the barley
transcriptome in a similar way. However, since powdery mildew interacts exclusively
with the epidermis of the host, a considerable proportion of powdery mildew induced
transcripts that are epidermis-specific may escape from detection in whole-leaf
samples due to dilution of the signal by the extract of the mesophyll (Zierold et al.,
2005). Accordingly, this experiment was initially set up to compare gene expression
using peeled epidermis. However, this turned out to be technically not feasible
because rust pustules damaged the epidermis and thereby, prevented efficient
peeling by 6 dai. Therefore only whole leaf samples were collected.
By using DI as criterion for differential gene regulation we identified 46 genes
belonging to different functional categories, predominantly activated in the double
inoculated plants versus single powdery mildew inoculated plants. In double
inoculated and also single rust inoculated plants, high up-regulation of PR-1 protein,
a marker for induced resistance was observed. Pathogenesis-related proteins (PRs)
are usually defined as host-specific proteins that are induced in several plant species
in response to attack by pathogens or related situations and have antimicrobial
activity (van Loon et al., 2006). Transient silencing of PR-1 expression decreased the
penetration resistance to the powdery mildew fungus in barley, indicating a role for
PR-1 in basal resistance to mildew (Schultheiss et al., 2003). We observed high up-
regulation of Cysteine synthase (Table 2) in the double inoculated treatment but not
in single rust or powdery mildew inoculated treatments. Fofana et al., (2007) reported
that in interaction of wheat with the wheat leaf rust pathogen (Puccinia triticina) the
expression of Cysteine synthase is up-regulated in incompatible (Lr gene) but down-
regulated in compatible interactions. Cysteine synthase is involved in reducing the
concentration of reactive oxygen species (ROS), signaling molecules that are
involved in various processes including pathogen defense (Apel & Hirt, 2004).
However, to confirm that PRs and ROS are indeed involved in rust-induced mildew
resistance, further investigations are necessary.
Biotrophic fungal pathogens including rust and powdery mildews were reported to
decrease the rate of photosynthesis in their host plants (e.g. Scholes & Farrar, 1986;
Scholes et al., 1994; Scholes & Rolfe, 1996). Our results showed that many genes
that were strongly down-regulated in double inoculated plants are involved in
photosynthesis (Table 2). Similar down-regulation of genes involved in
photosynthesis reported in wheat attacked by leaf rust pathogen including the 1,5-
Ribulose bisphosphate carboxylase which is the main source of energy production

Chapter 5
96
for the plant cell, generating ATP and reductive potential (NADPH) through
photosynthesis (Fofana et al., 2007). The role of this high down-regulation of genes
involved in photosynthesis in rust-mildew induced resistance is unknown, but it is
tempting to speculate that the strong suppression of photosynthesis in the mesophyll
might prevent proper powdery mildew development in the epidermis due to the lack
of energy.
This study presents the detection and histological and molecular characterization of
rust induced mildew resistance. Important questions that need to be addressed in
subsequent studies are the genes that determine the level of induced resistance, and
the mechanisms by which the rust infections apparently reprogramme the epidermal
cells to become more resistant to the mildew pathogen. An entirely different type of
question is, whether this interaction may have epidemiological consequences in case
both pathogens occur side-by-side in the field.
Acknowledgement
Reza Tadayonirad is acknowledged for his great help during the inoculations and samplings. The
macroarray analysis was done at the Leibniz Institute of Plant Genetics and Crop Plant Research
(IPK). The excellent technical assistance of Ines Walde is acknowledged. We thank Dr. Axel
Himmelbach from IPK and Dr. Chris Maliepaard form Plant Breeding for thier help in data analysis.
References
Apel K, Hirt H, 2004. Reactive oxygen species: metabolism, oxidative stress, and signal transduction,
Annual Review of Plant Biology 55: 373–399.
Asher MJC, Thomas CE. 1983. The expression of partial resistance to Erysiphe graminis in spring
barley. Plant Pathology 32: 79-89.
Calonnec A, Goyeau H, de Vallavielle-Pope C . 1996. Effects of induced resistance on infection
efficiency and sporulation of Puccinia striiformis on seedlings in varietal mixtures and on field
epidemics in pure stands. European Journal of Plant Pathology 102: 733–741.
Carver TLW, Lyngkjær MF, Neyron L, Strudwicke CC. 1 999. Induction of cellular accessibility and
inaccessibility and suppression and potentiation of cell death in oat attacked by Blumeria
graminis f.sp avenae. Physiological and Molecular Plant Pathology 55: 183–196.
Chin KM, Wolfe MS, Minchin PN. 1984. Host-mediated interaction between pathogen genotypes.
Plant Pathology 33: 161-171.
Cho BH, Smedegaard-Petersen V. 1986. Induction of resistance to Erysiphe graminis f.sp. hordei in
near-isogenic barley lines. Phytopathology 76: 301–305.
Durrant WE, Dong X. 2004. Systemic acquired resistance. Annual Review of Phytopathology 42:
185-209.
Fofana B, Banks TW, McCallum B, Strelkov SE, Clouti er S.2007. Temporal Gene Expression
Profiling of the Wheat Leaf Rust Pathosystem Using cDNA Microarray Reveals Differences in
Compatible and Incompatible Defence Pathways. International Journal of Plant Genomics.
doi:10.1155/2007/17542

Rust)induced powdery mildew resistance
97
Gregersen PL, Smedegaard V. 1989. Induction of resistance in barley against Erysiphe graminis
f.sp. hordei after preinoculation with the saprophytic fungus, Cladosporium macrocarpum.
Journal of Phytopathology 124: 128–136.
Halperin T, Schuster S, PniniCohen S, Zilberstein A , Eyal Z. 1996 . The suppression of pycnidial
production on wheat seedlings following sequential inoculation by isolates of Septoria tritici.
Phytopathology 86: 728–732.
Hammerschmidt R, Kuc J. 1982. Lignification as a mechanism for induced systemic resistance in
cucumber. Physiological Plant Pathology 17: 61–71.
Heath MC. 2002. Cellular interactions between biotrophic fungal pathogens and host and nonhost
plants. Canadian Journal of Plant Pathology 24: 259–264.
Holub EB, Cooper A. 2004. Matrix, reinvention in plants: how genetics is unveiling secrets of non-
host disease resistance. Trends in Plant Science 9: 211-214.
Hwang BK, Heitefuss R. 1982. Induced resistance of spring barley to Erysiphe graminis f.sp hordei.
Phytopathol. Z. 103: 41–47.
Jørgensen HJL, Lübeck PS, Thordal-Christensen H, de Neergaard E, Smedegaard-Petersen V.
1998. Mechanisms of induced resistance in barley against Drechslera teres. Phytopathology 88:
698-707.
Kessler A,. Baldwin IT. 2002. Plant responses to insect herbivory: the emerging molecular analysis.
Annual Review Plant Biology 53: 299–328.
Kochman J, Brown J F. 1975. Studies on the mechanism of cross protection in cereal rusts.
Physiological Plant Pathology 6: 19–27.
Koga H, Bushnell WR, Zeyen RJ. 1990. Specificity of cell type and timing of events associated with
papilla formation and the hypersensitive reaction in leaves of Hordeum vulgare attacked by
Erysiphe graminis f. sp. hordei. Canadian Journal of Botany 68: 2344–2352.
Kogel KH, Langen G. 2005. Induced disease resistance and gene expression in cereals. Cell
Microbiology 7: 1555–1564.
Kovats K, Binder A, Hohl HL. 1991. Cytology of induced systemic resistance of tomato to
Colletotrichum lagenarium. Planta 183: 484–490.
Kunoh H, Katsuragawa N, Yamaoka N, Hayashimoto A. 1 988. Induced accessibility and enhanced
inaccessibility at the cellular level in barley coleoptiles. III. Timing and localization of enhanced
inaccessibility in a single coleoptile cell and its transfer to an adjacent cell. Physiological and
Molecular Plant Pathology 33: 81–93.
Kunoh H. 2002. Localized induction of accessibility and inaccessibility by powdery mildew. In:
Bélanger RR, Bushnell WR, Dik AJ, Carver TLW, eds. The powdery mildews: A comprehensive
treatise. St. Paul/Minnesota, USA: APS Press, 126–133.
Lin MR, Edwards HH. 1974. Primary penetration process in powdery mildewed barley related to host
cell age, cell type, and occurrence of basic staining material. New Phytologist 73: 131-137.
Lyngkjær MF, Carver TLW, Zeyen RJ. 2001. Virulent Blumeria graminis infection induces
penetration susceptibility and suppresses race-specific hypersensitive resistance against
avirulent attack in Mla1-barley. Physiological and Molecular Plant Pathology 59: 243-256.
Lyngkjær MF, Carver TLW. 1999. Induced accessibility and inaccessibility to Blumeria graminis f.sp.
hordei in barley epidermal cells attacked by a compatible isolate. Physiological and Molecular
Plant Pathology 55: 151–162.

Chapter 5
98
Lyngkjær MF, Carver TLW. 2000 . Conditioning of cellular defence responses to powdery mildew in
cereal leaves by prior attack. Molecular Plant Pathology 1: 41-50.
Mains EB, Dietz SM. 1930. Physiologic forms of barley mildew, Erysiphe graminis hordei Marchal.
Phytopathology 20: 229-39.
Manandhar HK, Jørgensen HJL, Mathur SB, Smedegaard- Petersen V. 1998. Suppression of rice
blast by preinoculation with avirulent Pyricularia oryzae and the nonrice pathogen Bipolaris
sorokiniana. Phytopathology 88: 735-739.
Marcel TC, Aghnoum R, Durand J, Varshney RK, Niks R E. 2007. Dissection of the barley 2L1.0
region carrying the ‘Laevigatum’ quantitative resistance gene to leaf rust using near isogenic
lines (NIL) and subNIL. Molecular Plant–Microbe Interactions 20: 1604–1615.
Mellersh DG, Heath MC. 2001. Plasma membrane – cell wall adhesion is required for expression of
plant defense responses during fungal penetration. Plant Cell 13: 413–424.
Nelson HE. 2005. Fusarium oxysporum f.sp. radicis-lycopersici can induce systemic resistance in
barley against powdery mildew. Journal of Phytopathology 153: 366-370.
Niks RE. 1986. Failure of haustorial development as a factor in slow growth and development of
Puccinia hordei in partially resistant barley seedlings. Physiological and Molecular Plant
Pathology 28: 309–322.
Niks RE. 1989. Induced accessibility and inaccessibility of barley cells in seedling leaves inoculated
with two leaf rust species. Journal of Phytopathology 124: 296–308.
Oku H, Ouchi S. 1976. Host plant accessibility to pathogens. Review of Plant Protection Research 9:
58–71.
Olesen KL, Carver TLW, Lyngkjær MF . 2003. Fungal suppression of resistance against
inappropriate Blumeria graminis formae speciales in barley, oat and wheat. Physiological and
Molecular Plant Pathology 62: 37-50.
Oostendorp M, Kunz W, Dietrich B, Staub T. 2001. Induced disease resistance in plants by
chemicals. European Journal of Plant Pathology 107: 19–28.
Ouchi S, Hibino C, Oku H. 1976a. Effect of earlier inoculation on the establishment of a subsequent
fungus as demonstrated in powdery mildew of barley by a triple inoculation procedure.
Physiological and Molecular Plant Pathology 9: 25–32.
Ouchi S, Oku H, Hibino C. 1976b. Localization of induced resistance and susceptibility in barley
leaves inoculated with the powdery mildew fungus. Phytopathology 66: 901-905.
Parlevliet JE, Kuiper HJ. 1985. Accumulating polygenes for partial resistance to barley leaf rust,
Puccinia hordei. 1. Selection for increased latent period. Euphytica 37: 7–13.
Pei MH, Ruiz C, Hunter T, Bayon C. 2003. Rust resistance in Salix induced by inoculations with
avirulent and virulent isolates of Melampsora larici-epitea. Forest Pathology 33: 383–94.
Peterhänsel C, Freialdenhoven A, Kurth J, Kolsch R, Schulze-Lefert P. 1997. Interaction analyses
of genes required for resistance responses to powdery mildew in barley reveal distinct pathways
leading to leaf cell death. Plant Cell 9: 1397–1409.
Ryals JA, Neuenschwander UH, Willits MG, Molina A , Steiner HY, Hunt MD. 1996. Systemic
acquired resistance, Plant Cell 8: 1809–1819.
Sahashi N, Shishiyama J. 1986. Increased papilla formation, a major factor of induced resistance in
the barley-Erysiphe graminis f.sp. hordei system. Canadian Journal of Botany 64: 2178-2181.
Scholes JD, Farrar JF.1986. Increased rates of photosynthesis in localised regions of a barley leaf
infected with brown rust. New Phytologist 104: 601–612.

Rust)induced powdery mildew resistance
99
Scholes JD, Lee PJ, Horton P, Lewis DH. 1994. Invertase: understanding changes in the
photosynthetic and carbohydrate metabolism of barley leaves infected with powdery mildew. New
Phytologist 126: 213-222.
Scholes JD, Rolfe SA. 1996. Photosynthesis in localised regions of oat leaves infected with crown
rust (Puccinia coronata): quantitative imaging of chlorophyll fluorescence . Planta 199: 573-582.
Schultheiss H, Dechert C, Király L, Fodor J, Michel K, Kogel K-H, Huckelhoven R. 2003.
Functional assessment of the patgogenesis-related protein PR-1b in barley. Plant Science 165:
1275–1280.
Schweizer P, Hunziker, Mosinger E. 1989. cDNA cloning, in vitro transcription and partia1 sequence
analysis of mRNAs from winter wheat (Triticum aestivum L.) with induced resistance to Erysiphe
graminins f.sp. tritici. Plant Mo1ecular Biology 12: 643-654.
Sebesta J, Epstein AH, Formanova M. 1996. Induction to resistance to crown rust in oat. Journal of
phytopathology 144: 455-458.
Sreenivasulu N, Altschmied L, Panitz R, Hahnel U, M ichalek W, Weschke W, Wobus U. 2002.
Identification of genes specifically expressed in maternal and filial tissues of barley caryopses: a
cDNA array analysis. Molecular Genetics and Genomics 266: 758–767.
Sticher L, Mauch-Mari B, Metraux JP. 1997. Systemic acquired resistance. Annual Review of
Phytopathology 35: 235–270.
Ton J, Pieterse CMJ, Van Loon LC. 2006. The relationship between basal and induced resistance in
Arabidopsis. In: Tuzun S, Bent E, eds. Multigenic and induced systemic resistance in plants.
New York, USA: Springer, 197–224.
Trujillo M, Troeger M, Niks RE, Kogel KH, Hückelh oven R. 2004. Mechanistic and genetic overlap
of barley host and non-host resistance to Blumeria graminis. Molecular Plant Pathology 5: 389-
396.
Tsuchiya K, Hirata K. 1973. Growth of various powdery mildew fungi on the barley leaves infected
preliminarily with the barley powdery mildew fungus. Annals of the Phytopathological Society of
Japan 39: 396–403.
Vallad GE, Goodman RM. 2004. Systemic acquired resistance and induced systemic resistance in
conventional agriculture. Crop Science 44: 1920-1934.
Vallelian BL, Mosinger E, Metraux JP, Schweizer P. 1998. Structure, expression and localization of
a germin-like protein in barley Hordeum vulgare L. that is insolubilized in stressed leaves. Plant
Molecular Biology 37: 297–308.
Van Loon LC, Bakker PAHM, Pieterse CMJ. 1998. Systemic resistance induced by rhizosphere
bacteria. Annual Review of Phytopathology 36: 453–483.
van Loon, LC, Rep M, Pieterse CMJ. 2006. Significance of inducible defense related proteins in
infected plants. Annual Review of Phytopathology 44: 1-28.
Verhagen BWM, Glazebrook J, Zhu T, Chang H, Van Loo n LC, Pieterse CMJ. 2004 . The
transcriptome of rhizobacteria-induced systemic resistance in Arabidopsis. Molecular Plant-
Microbe Interaction 17: 895-908.
Yarwood CE. 1954. Mechanism of acquired immunity to a plant rust. Proceedings of the National
Academy of Sciences of the United States of America 40: 374-377.
Zierold U, Scholz U, Schweizer P. 2005. Transcriptome analysis of mlo-mediated resistance in the
epidermis of barley. Molecular Plant Pathology 6: 139-152.

Chapter 6
100

General discussion
101
General discussion

Chapter 6
102
Introduction In the barley-Blumeria interaction attacked plant epidermal cells defend themselves
against fungal penetration by localized responses leading to papilla formation and
preventing the fungus from haustorium formation. This mechanism of defense
conveys a race non-specific form of resistance and is common to basal host
resistance and mlo-mediated resistance to adapted powdery mildew (Bgh) and non-
host resistance to non-adapted forms of Bg. Identification of molecular components
underlying mlo resistance in barley and non-host resistance in Arabidopsis revealed
that these two types of resistance share many similarites (Humphry et al., 2006).
However, it is not known which genes make some barley accessions less susceptible
to the adapted form of Bg than others, and which genes determine the host status of
barley to non-adapted Bg forms. We undertook an integrated approach combining
QTL mapping and candidate genes analysis in order to identify genes that are
involved in basal resistance to adapted Bgh. Two experimental barley lines were
developed with exceptional susceptibility to a non-adapted form of Bg, the wheat
powdery mildew fungus. In this Chapter we discuss the implications of our findings for
understanding the genetics and mechanisms of basal resistance of barley to adapted
and non-adapted forms of powdery mildew.
Abundance and diversity of genes for basal resistan ce to powdery
mildew Basal resistance of barley to powdery mildew is a quantitatively inherited trait that
limits the growth and sporulation of powdery mildew colonies by a non-hypersensitive
mechanism of defense. This resistance is valuable for the breeders since it is
assumed to be race non-specific and durable. However, due to the polygenic
inheritance, interaction with environmental factors and subjectivity of disease
assessment, this type of resistance has always been considered difficult to breed for.
New molecular tools provided the opportunity to dissect this type of resistance and
understand its molecular basis, and might lead to more efficient breeding methods.
QTL mapping has broaden our understanding of the number and location of the
genes that are involved in basal resistance in different genetic backgrounds of barley
(Chapter 2). Most mapping studies on basal resistance of barley to powdery mildew
have been based on single biparental mapping populations, without verfication of the
mechanism of resistance at the cellular level. QTL mapping in biparental populations
reveals only a slice of the genetic architecture for a trait because only alleles that
differ between the two parental lines will segregate (Holland, 2007). In this thesis, the
genetics and mechanism of basal resistance of barley to powdery mildew have been

General discussion
103
studied using six mapping populations, involving 10 parents, to assess the diversity of
the genes involved in the genetic variation and the mechanism underlying the
resistance conferred by those QTLs. In different mapping populations different sets of
QTLs were responsible for the level of basal resistance and only a few QTLs were
found to be in common between populations. Even when a same parent participated
in two mapping populations (Vada and SusPtrit, see Chapter 2), we found that QTLs
discovered to be contributed by a particular parent in one mapping population, almost
never were detected in a mapping population of that same parent with a different
barley accession. These findings indicate a tremendous wealth of minor genes for
basal resistance to mildew. In addition, the results suggested that QTLs for powdery
mildew resistance in barley are mainly growth-stage dependent. This high diversity of
the genes for the basal resistance and their growth-stage dependency explain why
this type of resistance is race non-specific and probably durable, since multiple genes
with partial and inconsistent effects are involved, pathogen variants that overcome
basal resistance gain only a marginal advantage (Poland, 2008).
The results showing diversity of QTLs for basal resistance to powdery mildew within
the mapping populations of barley suggest that at each development stage there are
sufficient numbers of QTLs available that can be used for plant selection and
breeding.
Co-localization of QTLs and candidate genes Gene expression studies, motif directed profiling and high throughput gene silencing,
transient transformation and large scale mutation assays have been very productive
to indicate candidate genes for traits of interest, including disease resistance. In
Chapter 2, we prioritized many candidate genes that showed an interaction
phenotype with powdery mildew after RNAi silencing or overexpression studies, by
acquiring evidence of their mapping position. Six out of 26 mapped candidate genes
co-localized with the QTLs for powdery mildew resistance. This co-localization
suggests that those candidate genes are likely to contribute to the variation observed
in the mapping population in which the QTL at that locus had been detected. The
information obtained from the co-localization and the knowledge of the sequences of
those candidate genes can speed up the isolation of the genes at those QTLs.
However, for final proof that the QTLs that co-localize with the candidate genes are
indeed caused by allelic variation in those candidate genes, a mapping at much
higher genetic resolution is required. HvSNAP34 is one of the prioritized candidate
genes. This gene encodes a t-SNARE protein that is involved in mlo-mediated, non-
host and host basal resistance (Douchkov et al., 2005). This is an interesting gene to

Chapter 6
104
be cloned to compare its allelic variation between Dom and Rec, that had different
QTL alleles at that locus. Both alleles could be used in transient expression assays to
investigate whether one confers more resistance than the other.
Accumulation of genes for basal resistance The multi-population QTL analysis showed that basal resistance of barley to powdery
mildew is conferred by many genes, most of which are growth stage dependent. QTL
analysis followed by Marker Assisted Selection (MAS) has been suggested to be an
efficient strategy in developing new lines with levels of resistance superior to that of
their parents. This process is known as QTL pyramiding. For pyramiding, after the
discovery of interesting QTLs in the mapping population, QTLs can be combined
through crossing of the genotypes carrying the QTL alleles to be followed by MAS
(Ashikari & Matsuoka, 2006). However in practical breeding, accumulation of genes
using MAS has to be repeated in each crossing cycle since the combined resistance
genes will segregate in the progeny. For traits that are difficult to measure and show a
low heritability, the MAS strategy should be considered as a potential alternative for
crop improvement. However, there are not always marker alleles close to the desired
QTL allele to monitor reliably the introgression of that QTL in new progeny. Other
parents may carry the same marker allele, or the marker allele may be too far away
from the target QTL allele. In that case haplotypes around the target QTL allele
should be determined, which is a laborious effort. Another aspect that may reduce the
efficiency of MAS is the possible occurrence of QTL interactions. That may lead to
disappointing phenotypic performance conferred by designed QTL combinations. We
showed here that accumulation of unknown QTLs just by visual selection for higher
level of basal resistance among the progenies is feasible and easy.
Non-host immunity of barley to non-adapted powdery mildews Complexity of the mechanism of non-host resistance Many hypotheses about the mechanisms of non-host resistance have been
suggested, but two of them are currently in the focus of interest.
1-The non-adapted pathogens produces effectors to suppress PAMP-triggered
immunity in the plant, but these effectors correspond insufficiently with the operative
targets.
2- Plants contain multiple “R genes” acting together to recognize the pathogen
avirulence (Avr) gene products (functionally these are presumed to be effectors).
Since the multiple R-gene combination requires several simultaneous Avr � avr
mutations, it is impossible for the pathogen to break this defense.

General discussion
105
Both of these two models can explain the durability of non-host resistance (Jones &
Dangl, 2006). Genetic studies of interaction of Arabidopsis with non-adapted powdery
mildew forms have shown that both pre- and post-penetration defense mechanisms
are involved in non-host resistance (Lipka et al., 2005). Our histological studies
revealed that resistance to penetration in association with papilla formation is the
main mechanism involved in non-host resistance to non-adapted forms of Bg.
However, a considerable proportion of successfully penetrated germlings (30 to 60%
depending on the barley-non-adapted mildew form combination) were stopped by a
hypersensitive reaction. One forma specialis, from Hordeum murinum, induced
extensive hypersensitivity upon haustorium formation on some tested lines. However,
Christopher-Kozjan & Heath, (2003) presented evidence that hypersensitivity in non-
host combinations not necessarily imply the activity of R/Avr interaction. The
combination of non-hypersensitive and hypersensitive defense mechanisms involved
suggest that non-adapted powdery mildew forms should evolve strategies to
overcome different defense mechanisms in different developmental stages to acuire a
plant species as suitable host.
Genetics of non-host resistance to non-adapted powd ery mildews Most of the genes that have been implicated in non-host resistance have been
discovered by mutagenesis or gene expression studies (Niks & Marcel, 2009; Table
1). Only a few studies reported genes that play a role in natural variation for non-host
resistance (Shafiei et al., 2007; Rodrigues et al., 2004; Jafary et al., 2006; Jafary et
al., 2008; Zhang et al., 2009). Barley is a full non-host to the non-adapted Bgt. We
tested here a large collection of barley accessions against Bgt, and only a few
accessions were found to possess susceptibility to Bgt at seedling stage, and at most
at a very low level. Crossings among some barley accessions with some level of
susceptibility to Bgt resulted in the accumulation of susceptibility factors in two barley
lines called SusBgt SC and SusBgt DC. The accumulation of susceptibility genes in
barley to Bgt by successive crosses suggest that the resistance has a quantitative
inheritance. However we intend to follow a mapping approach to determine the
genetics of non-host resistance in barley against non-adapted forms of Bg.
Furthermore both SusBgt lines are useful tools in gene expression and
complementation studies on non-host resistance. The two lines show surprisingly a
differential level of susceptibility in the conidiation stage of non-adapted powdery
mildews. Therefore, mapping populations based on both lines may lead to the
identification of QTLs that are effective in particular stages of fungal development.

Chapter 6
106
Enhancement of basal resistance to powdery mildew Our results showed that basal resistance of barley against powdery mildew is
enhanced by pre-inoculation with leaf rust. We hypothesized that this rust-induced
mildew resistance might be due to rust-plant communication during stoma
penetration, or to the presence of the rust hyphae in the apoplast, or to some degree
of pre-haustorial resistance mounted by the mesophyll cells to the invading rust
fungus.
Using a barley cDNA macroarray, we compared the genome-wide transcriptome
changes upon double inoculation (LR+PM) with single inoculation leaf rust (LR) and
powdery mildew (PM) treatments. The majority of genes up-regulated by LR are also
up-regulated in the LR+PM, and hence, not silenced by PM. Analysis of quantitative
transcriptome abundance of differentially regulated genes in double inoculation
treatment versus single inoculations (PM and LR) revealed that most of the down-
regulated genes are involved in plant metabolism and photosynthesis. This suggests
that the rust-induced powdery mildew resistance state might be due to a lack of
energy in the plant cells. In double inoculated and also single rust inoculated plants, a
high up-regulation of PR-1 protein, a Pathogenesis-related protein (PR) was
observed. Accumulation of PRs is a hallmark of pathogen-induced systemic acquired
resistance (van Loon et al., 2006). Accumulation of PR proteins after infection by
pathogens has been reported to be associated with increased effectiveness of plant
defense responses upon subsequent pathogen infection, a state that has been called
‘’Priming’’. Primed plants are capable of activating defense responses either faster,
stronger, or both upon attack by pathogens, insects or in response to abiotic stress
(Conrath et al., 2006). Both our histological and expression studies suggest that pre-
inoculation of barley with leaf rust prime the basal defense mechanism of plants
against subsequent challenge inoculation with powdery mildew.
Concluding remarks
In this thesis we found that the barley genome is a rich source of QTLs (genes)
conferring basal resistance to powdery mildew. The great diversity of such genes in
the barley germplasm making transgression for basal resistance a very common
phenomenon. Identification of the genes underlying the powdery mildew seems
feasible considering the prioritarized candidate genes that were found to co-localize
with resistance QTLs. Information gained from the prioritization of candidate genes
can help in the isolation of the genes that underlie resistance QTLs. These candidate
genes have been shown already to affect the level of basal resistance, and now they
have been mapped to the QTL positions found in our mapping studies. In qualitative

General discussion
107
Table 1. Genes with known sequences that were implicated to play a role in basal host and/or non-
host resistance of barley to powdery mildews ( adapted from Niks & Marcel, 2009)
Gene name Description BHR a NHRb References *
Genes involved in transport and secretion systems
ROR2 Plasma membrane-anchored
syntaxin
Bgh Bgt 1,2,3,8,10
SNAP34
ADF3
Synaptosome-associated protein
Actin depolymerizing factor 3
Bgh
Bgh
Bgt
Bgt
11,3,10
11
Genes involved in the positive or negative regulati on of basal defenses
BI-1 BAX inhibitor 1 Bgh Bgt 3,9,11
CDPKs Calcium-dependent protein
kinases
Bgh Bgt 8
MLO Seven-transmembrane domain
protein
Bgh 2,3,4,7,10,11
Genes involved in redox signaling
GLPs Germin-like proteins Bgh 2,3
RBOHs NADPH oxidase Bgh 3
PRXs: Prx07/Prx40 Class III peroxidase proteins Bgh Bgt 6
Transcription factors and signaling cascades
NACs: NAC6 NAC transcription factors Bgh 5
WRKYs: WRKY1/2 Group II WRKYs transcription
factors
Bgh 12
a, basal host resistance
b, non-host resistance
* references:
1: Collins et al. (2003)
2: Collins et al. (2007)
3: Hückelhoven (2007)
4: Jarosch et al. (1999)
5 : Jensen et al., (2007)
6: Johrde & Schweizer (2008)
7: Kumar et al. (2001)
8: Lipka et al. (2008)
9: Nürnberger & Lipka (2005)
10: O’Connell & Panstruga (2006)
11: Schweizer (2007)
12: Shen et al.(2007)
genes this would be sufficient proof to consider these sequences as the “responsible
gene”. In QTLs it is, however, too early to consider these candidates as “confirmed”.
We propose that either complementation tests with allelic forms or ultra-fine physical
QTL mapping are carried out to confirm the candidate genes that emerge from our
study as the most likely. The developed barley lines with exceptional susceptibility to
Bgt will serve to determine the genetic basis of non-host resistance in barley to wheat
powdery mildew by both forward and reverse genetic approaches. This material will
also be useful to quantify subsequent lines of basal defense that the mildew fungus
encounters during its development, such as primary haustorium and secondary
haustorium formation and conidiation. Finally, the rust-induced mildew resistance

Chapter 6
108
phenomenon that was reported here for the first time can be further investigated to
improve our knowledge about the molecular mechanism of penetration resistance at
cell level and the mechanisms that plants deploy to protect themselves to different
biotic stress agents.
References
Ashikari M, Matsuoka M. 2006. Identification, isolation and pyramiding of quantitative trait loci for rice
breeding. Trends Plant Science 11: 344–350.
Christopher-Kozjan R, Heath MC. 2003. Cytological and pharmacological evidence that biotrophic
fungi trigger different cell death execution processes in host and nonhost cells during the
hypersensitive response. Physiological and Molecular Plant Patholology 62: 265-275.
Collins NC, Niks RE, Schulze-Lefert P, 2007. Resistance to cereal rusts at the plant cell wall-What
can we learn from other host-pathogen systems? Australian Journal of Agricultural Research 58:
476-489.
Collins NC, Thordal-Christensen H, Lipka V, Bau S, Kombrink E, Qiu J-L, Hückelhoven R, Stein
M, Freialdenhoven A, Somerville S et al. 2003. SNARE-protein-mediated disease resistance at
the plant cell wall. Nature 425: 973-977.
Conrath U, Beckers GJM, FlorsV, Garcia-Agustin P, J akab G, Mauch F, Newman MA, Pieterse
CMJ, Poinssot B, Pozo MJ, Pugin A, Schaffrath U, To n J, Wendehenne D, Zimmerli L,
Mauch-Mani B. 2006. Priming: getting ready for battle. Molecular Plant-Microbe Interaction 19:
1062-1071. Douchkov D, Nowara D, Zierold U, Schweizer P. 2005. A high-throughput gene-silencing system for
the functional assessment of defense-related genes in barley epidermal cells. Molecular Plant-
Microbe Interactions 18: 755-761.
Holland JB. 2007. Genetic architecture of complex traits in plants. Current Opinion in Plant Biology
10: 156–161.
Hückelhoven R. 2007. Cell wall-associated mechanisms of disease resistance and susceptibility.
Annual Review of Phytopathology 45: 101-127.
Humphry M, Consonni C, Panstruga R. 2006. mlo-based powdery mildew immunity: silver bullet or
simply non-host resistance? Molecular Plant Pathology 7: 605–610.
Jafary H, Albertazzi G, Marcel TC, Niks RE. 2008. High diversity of genes for nonhost resistance of
barley to heterologous rust fungi. Genetics 178: 2327–2339.
Jafary H, Szabo LJ, Niks RE. 2006. Innate nonhost immunity in barley to different heterologous rust
fungi is controlled by sets of resistance genes with different and overlapping specificities.
Molecular Plant–Microbe Interactions 19: 1270–1279.
Jarosch B, Kogel KH, Schaffrath U. 1999. The ambivalence of the barley Mlo locus: Mutations
conferring resistance against powdery mildew (Blumeria graminis f.sp. hordei) enhance
susceptibility to the rice blast fungus Magnaporthe grisea. Molecular Plant-Microbe Interactions
12: 508–514.
Jensen MK, Rung JH, Gregersen PL, Gjetting T, Fugls ang AT, Hansen M, Joehnk N, Lyngkjaer
MF, Collinge DB. 2007. The Hv NAC6 transcription factor: a positive regulator of penetration
resistance in barley and Arabidopsis. Plant Molecular Biology 65: 137-150.
Johrde A, Schweizer P. 2008. A class III peroxidase specifically expressed in pathogen-attacked
barley epidermis contributes to basal resistance. Molecular Plant Pathology 9: 687- 696.

General discussion
109
Jones JDG, Dangl JL. 2006. The plant immune system. Nature 444: 323–329.
Kumar J, Hückelhoven R, Beckhove U, Nagarajan S, Ko gel KH. 2001. A compromised Mlo
pathway affects the response of barley to the necrotrophic fungus Bipolaris sorokiniana
(Teleomorph: Cochliobolus sativus) and its toxins. Phytopathology 91: 127–133.
Lipka U, Fuchs R, Lipka V. 2008. Arabidopsis non-host resistance to powdery mildews. Current
Opinion in Plant Biology 11: 1-8.
Lipka V, Dittgen J, Bednarek P, Bhat R, Wiermer M, Stein M,Landtag J, Brandt W, Rosahl S,
Scheel D, et al. 2005. Pre- and post invasion defences both contribute to nonhost resistance in
Arabidopsis. Science 310: 1180-1183.
Niks RE, Marcel TC. 2009. Non-host and basal resistance: how to explain specificity? New Phytologist
182: 817–828
Nürnberger T, Lipka V. 2005. Non-host resistance in plants: new insights into an old phenomenon.
Molecular Plant Pathology 6: 335-345.
O'Connell RJ, Panstruga R. 2006. Tête à tête inside the plant cell: establishing compatibility between
plants and biotrophic fungi and oomycetes. New Phytolologist 171: 699-718.
Poland JA, Balint-Kurti2 PJ, Randall J, Wisser RCP, Nelson RJ.2008 . Shades of gray: the world of
quantitative disease resistance. Trends in Plant Science doi:10.1016/j.tplants.2008.10.006
Rodrigues P, Garrood JM, Shen QH, Smith PH, Boyd LA . 2004. The genetics of non-host disease
resistance in wheat to barley yellow rust. Theoretical and Applied Genetics 109: 425-432.
Schweizer P. 2007. Nonhost resistance of plants to powdery mildew - New opportunities to unravel
the mystery. Physiological and Molecular Plant Patholology 70: 3-7.
Shafiei R, Hang C, Kang J-G, Loake GJ. 2007. Identification of loci controlling non-host disease
resistance in Arabidopsis against the leaf rust pathogen Puccinia triticina. Molecular Plant
Pathology 8: 773-784.
Shen, Q-H, Saijo Y, Mauch S, Biskup C, Bieri S, Kel ler B, Seki H, Ülker B, Somssich IE, Schulze-
Lefert P. 2007. Nuclear activity of MLA immune receptors links isolate-specific and basal
disease-resistance responses. Science 315: 1098-1103.
van Loon, LC, Rep M, Pieterse CMJ. 2006. Significance of inducible defense related proteins in
infected plants. Annual Review of Phytopathology 44: 1-28.
Zhang NW, Lindhout P, Niks RE, Jeuken MJW. 2009. Genetic dissection of Lactuca saligna nonhost
resistance to downy mildew at various lettuce developmental stages. Plant Pathology, in press

Chapter 6
110

Abbreviations
111
Abbreviations
AUDPC Area Under the Disease-Progress Curve
Bg Blumeria graminis
Bga Blumeria graminis f.sp. avenea
Bgar Blumeria graminis f.sp. agropyri
Bgb Blumeria graminis f.sp. bromi
Bgd Blumeria graminis f.sp. dactylidis
Bgh Blumeria graminis f.sp. hordei
Bghm Blumeria graminis f.sp hordei-murini
Bgt Blumeria graminis f.sp. tritici
CAPS Cleaved Amplified Polymorphic Sequences
dai day after inoculation
DArT Diversity Arrays Technology
DC Double Cross
dCAPS derived Cleaved Amplified Polymorphic Sequences
DHs Doubled Haploids
DI Differential Index
ESH Elongated Secondary Hypha
EST Expressed Sequence Tag
f.sp. forma specialis
ff.spp. formae speciales
FPP Failed Primary Penetration
FPP+HR Failed Primary Penetration with HR
FPP-HR Failed Primary Penetration without HR
hai hour after inoculation
HR Hypersensitive Response
IR Induced Resistance
IS Induced Susceptibility
ISR Induced Systemic Resistance
LOD Logarithm of odds
LR Leaf Rust

Abbreviations
112
MAS Marker-Assisted Selection
Mla Mildew resistance locus a
Mlo Mildew resistance locus o
nhR non-host resistance
NILs Near Isogenic Lines
PAMP Pathogen-Associated Molecular Patterns
PEN Penetration (mutant genes)
PM Powdery mildew
PRs Pathogenesis-Related proteins
PTI PAMP-Triggered Immunity
QTL Quantitative Trait Locus
Rbgq quantitative genes for Resistance to Blumeria graminis
RILs Recombinant Inbred Lines
RNAi RNA interference
Ror Required for mlo-specified resistance
Rphq2 An allele for quantitative resistance to Puccinia hordei
SAP Secondary Attempted Penetration
SAR Systemic Acquired Resistance
SC Single Cross
SCAR Sequence Characterized Amplified Region
SPP Successful Primary Penetration
SPP+HR Successful Primary Penetration with a haustorium and with HR
SPP-HR Successful Primary Penetration with a haustorium but without HR
TDM Transcript-Derived Marker
TIGS Transient-Induced Gene Silencing

Glossary
113
Glossary Avirulent: carrying an avirulence (Avr) gene that hampers development and
reproduction of pathogen in a host plant accession that carries a corresponding
resistance gene.
Basal defense/resistance:
qualitative: defense that plant species mount against non-adapted microbial
intruders.
quantitative: defense that inhibits pathogen spread after successful infection and
onset of disease.
Basic compatibility: state that results from the capacity of the microbe to deal
effectively with the defense that plant species mount against non-adapted microbial
intruders.
Challenger: in induced-resistance terminology, it refers to an adapted or non-
adapted pathogen which is inoculated to a plant subsequently after inoculation with
inducer pathogen (see there).
Colonization: the development stage after establishment of the infection (see
there). In mildews the stage of development of secondary and subsequent haustoria.
Compatible interaction: a host–pathogen interaction that results in production of
the pathogen without elicitation of gross defense responses (the host is susceptible).
Durable resistance: resistance that has remained effective for a long period in
which it has been applied at large scale in an environment conducive to the
pathogen.
Effector: secreted pathogen protein that manipulates host cell functions to suppress
defense and promote infection. As a secondary effect, an effector may elicit in
particular plant genotypes a hypersensitive response, and be an avirulence factor.
Establishment: the development of a primary haustorium in attacked plant cells by
a pathogenic fungus, especially rust and powdery mildew fungi.
Haustorium: specialized organ formed by most biotrophic (pathogenic) fungi and
oomycetes in a living plant cell for nutrient absorption and molecular communication
with the plant cell.
Host specificity: the property of a pathogen species to be able to establish basic
compatibility with only one or a few plant species.
Host status: status of a plant species being susceptible to a particular pathogen
species. If not, the plant is a marginal host or a non-host.
Incompatible interaction: a host–pathogen interaction that does not result in
disease (the host is resistant).

Glossary
114
Induced Systemic Resistance (ISR): the phenomenon that plants acquire an
enhanced defensive capacity against subsequent pathogen attack as a result of root
colonization by selected strains of non-pathogenic bacteria.
Inducer: In induced-resistance terminology, it refers to a pathogen, non-pathogenic
microorganism or other factors that induce resistance or susceptibility to a
subsequent challenger.
Linkage disequilibrium (LD). The condition in which the frequency of a particular
haplotype for two loci is significantly different from that expected under random
mating.
Near isogenic lines (NILs): Inbred lines that differ at only a small genomic region.
Non-adapted pathogen: pathogen species that cannot infect a particular (non-host)
plant species (syn. heterologous, inappropriate).
Non-host resistance: resistance occurring in all genotypes of a plant species to all
genotypes of a potential pathogen species.
Operative target: host target that, when manipulated by a pathogen effector, results
in enhanced pathogen fitness.
PAMP: pathogen-associated molecular pattern; a pathogen-specific molecular
sequence or structure, often indispensable for the microbial lifestyle that, elicits an
innate immune response on receptor-mediated perception.
PAMP-triggered immunity (PTI): first line of active plant defense that relies on the
recognition of pathogen-associated (or microbe-associated) molecular patterns
(PAMPs/MAMPs) by pattern-recognition receptors (PRRs).
Pattern-recognition receptors: plant proteins that identify PAMP (see above)
molecules, such as flagellin or chitin components, that are associated with microbial
pathogens.
Quantitative trait locus (QTL): a locus with an effect on a quantitative trait (i.e. a
trait showing continuous variation) that can only be mapped by the application of
QTL mapping software.
Recombinant inbred line (RIL): an inbred line produced from an initial cross
followed by continuous inbreeding; populations of RILs are often used for QTL
mapping studies.
Resistance gene analogs (RGA): a sequence identified in whole genome analyses
based on their structural homology (presence of NBS and/or LRR domains) with
previously identified R-genes.
Systemic Acquired Resistance (SAR): the phenomenon that plants acquire an
enhanced defensive capacity against subsequent pathogen attack as a result of pre-
inoculation with pathogens that causes necrotic lesions.

Summary
115
Summary In the barley-Blumeria interaction, resistance at penetration stage in association with
papilla formation is a commonly occurring mechanism. This mechanism of defense
reduces the infection severity by adapted powdery mildew pathogen (basal
resistance to Blumeria graminis f.sp. hordei, Bgh) and fully protects the plant against
non-adapted powdery mildew pathogens (non-host resistance to non-adapted forms
of B. graminis). In this thesis we followed an integrated approach based on QTL
mapping and candidate gene analysis. It was our objective to determine which of the
known candidate genes have a map position that coincides with one of the QTLs
detected in our mapping study. Such candidate genes might determine the natural
variation of basal resistance against powdery mildew in barley. We also investigated
whether the genotypic variation in level of resistance of barley to the non-adapted
wheat powdery mildew, B. graminis f.sp. tritici (Bgt) could be used to develop an
experimental line to determine the inheritance of non-host resistance of barley to this
pathogen. Finally we aimed to determine whether the basal resistance of barley to
powdery mildew is being suppressed by prior attack by a different pathogenic
haustorium forming fungus, Puccinia hordei.
Chapter 1 presents an introduction about basal resistance of barley to powdery
mildew and briefly lists evidence on molecular associations between basal, mlo-
mediated and non-host resistance. In Chapter 2 , we performed QTL analysis for
powdery mildew resistance at seedling and at adult plant stage in six mapping
populations of barley. In that analysis quantitative resistance of barley to powdery
mildew was found to be based on a large number of genes, with only few detected in
more than one mapping population. These QTLs for powdery mildew resistance in
barley are mainly plant growth stage dependent. From gene-expression, gene
silencing and transient transformation assays 39 genes emerged that increased or
decreased basal resistance of barley to the powdery mildew fungus. They are
considered candidate genes that might be responsible for natural variation in level of
basal resistance. By looking for polymorphism in those genes, we could determine
the map position of 23 of these candidate genes. Mapping positions of three more
genes, mlo, Ror1 and Ror2, were already available, and these were also considered
as candidate genes in our study.To compare the map position of QTLs in different
mapping populations with the map positions of the 26 candidate genes, we
constructed an improved high-density integrated linkage map of barley. This
improved map was based on the marker data of seven mapping populations of
barley. More than 43% of the markers on our improved integrated map target ESTs
or gene sequences (gene-targeted markers, GTM) and can be used for candidate

Sammary
116
gene identification. Six out of the 26 candidate genes co-localized with QTLs for
basal powdery mildew resistance. They are interesting targets for further studies,
since their allelic forms may be responsible for part of the differences in basal
resistance between the parents in our mapping study. Chapter 3 reports the
development of two experimental barley lines with extremely high and low level of
basal resistance to barley powdery mildew. These lines were obtained by convergent
crossing between the most resistant and the most susceptible lines, respectively,
from four mapping populations of barley studied in Chapter 2. We consider the
extremely susceptible and resistant lines developed here as valuable material to be
used in further experiments to characterize the molecular basis of basal resistance to
powdery mildew. The results suggest that phenotypic selection is sufficiently efficient
to achieve high levels of basal resistance. We report some difficulties in the
application of marker assisted selection that might make that approach unnecessary
and less efficient to achieve a similar high level of basal resistance. In Chapter 4 the
development of two more experimental lines is described. These lines have, at the
seedling stage an unprecedented level of susceptibility to the non-adapted wheat
powdery mildew. A large collection of barley germplasm was screened and some
rare barley accessions were identified with rudimentary susceptibility to the wheat
powdery mildew, Bgt. Those accessions were intercrossed in two cycles, and
resulted in the two exceptional research lines, called SusBgt SC and SusBgt DC. The
quantitative variation among barley accessions and in the progenies after convergent
crossing suggest a polygenic basis of this non-host resistance. Component analysis
of the infection process suggested that non-host resistance factors are Blumeria-form
specific. The developed lines will serve to elucidate the genetic basis of non-host
resistance in barley to wheat powdery mildew, and are useful tools in gene
expression and complementation studies on non-host resistance. In Chapter 5 we
reported that a compatible interaction of barley-Puccinia hordei induces increased
papilla based resistance to a challenge infection by a compatible powdery mildew
isolate. These pathogens differ in strategies to suppress plant cell defense. We
showed that the level of rust-induced mildew resistance varies among barley
accessions and is not determined by the virulent/avirulent spectra of the challenger
isolate. Macroarray gene expression analysis showed that several genes involved in
metabolism and photosynthesis were highly down-regulated and a few genes
involved in plant defense including a pathogenesis related protein (PR-1) and
cysteine synthase were highly up-regulated in the double inoculated treatment. Our
histological and gene expression analyses are compatible with the hypothesis that
the rust “primes” the basal mildew resistance genes prior to the challenge mildew

Summary
117
infection. In Chapter 6 , the results obtained in the previous Chapters are being
discussed. The perspectives of the experimental barley lines that were developed in
this study for the identification of genes that determine basal resistance of barley to
adapted and non-adapted Blumeria graminis forms are highlighted.

Sammary
118

Samenvatting
119
Samenvatting
In de gerst-meeldauw (Blumeria) interactie is resistentie tegen celwandpenetratie
door de vorming van papillen een algemeen voorkomend verschijnsel. Dit
resistentiemechanisme reduceert de aantastingsgraad van gerst door
gerstmeeldauw (basisresistentie tegen Blumeria graminis f.sp. hordei, Bgh), en
beschermt de plant volledig tegen niet op gerst gespecialiseerde vormen van
meeldauw (niet-waardresistentie). In dit proefschrift volgden we een geïntegreerde
benadering door de kartering van QTLs te combineren met gegevens uit een
kandidaatgenbenadering. Doel was om te bepalen welke van de bekende
kandidaatgenen karteren op een van de door ons in kaart gebrachte QTLs.
Dergelijke kandidaatgenen kunnen verantwoordelijk zijn voor de gevonden
verschillen in basisresistentie van gerst tegen de gerstmeeldauw. We hebben ook
onderzocht of de genetische variatie in resistentieniveau tegen de
tarwemeeldauwschimmel (B. graminis f.sp. tritici) zou kunnen worden benut om een
experimentele gerstlijn te ontwikkelen die zou kunnen dienen om de overerving te
bepalen van de niet-waard resistentie van gerst tegen dat pathogeen. Tenslotte
bepaalden we of de basisresistentie van gerst tegen meeldauw zou worden
onderdrukt na eerdere infectie door een ander op gerst gespecialiseerd
haustoriumvormend pathogeen, nl. de dwergroestschimmel Puccinia hordei.
Hoofdstuk 1 introduceert het onderwerp, basisresistentie van gerst tegen
meeldauw, en geeft een korte beschrijving van de argumenten die suggereren dat
basisresistentie, mlo-resistentie en niet-waard resistentie op moleculair niveau
geassocieerd zijn. In Hoofdstuk 2 beschrijven we een QTL analyse die we
uitvoerden om de genetische loci te vinden die bijdragen aan variatie in
basisresistentie van gerst tegen meeldauw in het zaailingstadium en in het
volwassenplantstadium. We deden deze analyse in zes karteringspopulaties van
gerst. Die studie leidde tot de conclusie dat deze kwantitatieve resistentie gebaseerd
is op een groot aantal genen. Slechts enkele daarvan werden gevonden in meer dan
een populatie. De meeste van deze QTLs voor meeldauwresistentie zijn
plantontwikkelingsstadium-specifiek. Uit genexpressie-, genexpressie
onderdrukkings- en transformatie-experimenten kwamen 39 genen naar voren die
bijdroegen aan de kans dat een meeldauwschimmel met succes een haustorium in
een plantencel vormde. Deze genen kunnen worden beschouwd als kandidaatgenen
die verantwoordelijk zijn voor natuurlijke verschillen in niveau van basisresistentie.
We zochten sequentieverschillen in deze kandidaatgenen, zodat we hun kaartpositie
konden bepalen. Dit lukte voor 23 van deze kandidaatgenen. Kaartposities van drie
andere bij meeldauwresistentie betrokken genen, mlo, Ror1 en Ror2, waren al

Samenvatting
120
bekend, en ook deze werden voor onze studie beschouwd als kandidaatgenen voor
basisresistentie. Vervolgens vergeleken we de kaartpositie van deze 26
kandidaatgenen met die van de gevonden QTLs voor basisresistentie. Daarvoor
ontwikkelden we een verbeterde en zeer dichte, geïntegreerde koppelingskaart van
gerst. Deze verbeterde kaart was gebaseerd op zeven karteringspopulaties van
gerst. Meer dan 43% van de merkers die op deze kaart zijn samengebracht
vertegenwoordigen gensequenties, en zijn van nut om in de toekomst
kandidaatgenen te identificeren. Zes van de 26 kandidaatgenen hadden een
kaartpositie die samenviel met die van een QTL voor basisresistentie tegen
meeldauw. Deze zes kandidaatgenen zijn interessant voor vervolgstudies, omdat
variatie in deze genen wel eens verantwoordelijk zouden kunnen zijn voor een deel
van de verschillen in basisresistentie die gevonden werden in de karteringsstudies.
Hoofdstuk 3 beschrijft de ontwikkeling van twee experimentele gerstlijnen met een
extreem hoge en een extreem lage basisresistentie tegen gerstmeeldauw. Deze
lijnen werden verkregen door toepassing van convergente kruisingen tussen
respectievelijk de meest resistentie en tussen de meest vatbare lijnen in de
karteringspopulaties die beschreven zijn in Hoofdstuk 2. We beschouwen de extreem
vatbare en extreem resistente lijnen als waardevol materiaal voor verder onderzoek
naar de moleculaire basis van basisresistentie tegen meeldauw. De resultaten wijzen
erop dat fenotypische selectie voldoende effectief is om zeer hoge niveaus van
basisresistentie te verkrijgen. We geven enkele argumenten dat merkergestuurde
selectie wel eens onnodig en minder efficiënt zou kunnen zijn om een dergelijk hoog
niveau van resistentie te bereiken. In Hoofdstuk 4 beschrijven we de ontwikkeling
van twee andere experimentele gerstlijnen. Deze lijnen hebben, in het
zaailingstadium, een niet eerder gemeld hoog niveau van vatbaarheid voor de op
tarwe gespecialiseerde tarwemeeldauwschimmel. In een grote collectie gerstlijnen
werd gezocht naar zeldzame gerstlijnen met een rudimentaire vatbaarheid voor dit
tarwepathogeen. Die gerstlijnen werden in twee cycli onderling gekruist, wat
resulteerde in twee buitengewone onderzoekslijnen, SusBgt SC en SusBgt DC. De
kwantitatieve variatie tussen gerstlijnen en binnen de hier ontwikkelde
nakomelingschappen wijst op een kwantitatieve overerving van deze niet-waard
resistentie. Componentenanalyse van het infectieproces suggereerde dat factoren
die de niet-waard resistentie van gerst tegen meeldauwvormen bepalen Blumeria-
vorm-specifiek zijn. De ontwikkelde lijnen zullen dienen om de overerving van niet-
waard resistentie in gerst tegen de tarwemeeldauw te bepalen. De lijnen zijn voorts
een nuttig instrument in genexpressie- en complementatiestudies naar niet-waard
resistentie. In Hoofdstuk 5 vermelden we dat een compatibele interactie tussen

Samenvatting
121
gerst en Puccinia hordei (de veroorzaker van dwergroest) de basisresistentie tegen
een daarna aangebrachte meeldauw verhoogt. Deze pathogenen verschillen in
manier waarop ze de afweer van plantencellen onderdrukken. We toonden aan dat
het niveau van deze roest-geïnduceerde resistentie verschilt tussen gerstlijnen, en
niet afhangt van het virulentiespectrum van het gebruikte meeldauwisolaat. Een
macroarray genexpressie-experiment liet zien dat in de dubbel-geïnoculeerde
behandeling verscheidene genen die een rol spelen in plant metabolisme en
fotosynthese, verminderd tot expressie kwamen. Dit, terwijl enkele genen die een rol
spelen in de afweer van planten versterkt tot expressie kwamen. Onder de
laatstgenoemde zijn pathogenese gerelateerde eiwitten (PR-1) en cysteine synthase
het vermelden waard. Onze histologische en genexpressiebevindingen zouden erop
kunnen wijzen dat de eerdere infectie door de roest tot gevolg heeft dat genen voor
basisresistentie tegen meeldauw worden “geprimed”, d.w.z. klaargemaakt om snel
en sterk te reageren zodra een meeldauwschimmel de cel aanvalt. In Hoofdstuk 6
bespreken we de in de voorgaande hoofdstukken behaalde resultaten. Met name
wijzen we op de perspectieven die de verkregen experimentele lijnen bieden voor de
identificatie van genen die verantwoordelijk zijn voor basisresistentie van gerst tegen
aangepaste en niet-aangepaste vormen van Blumeria graminis.

New perspectives to identify genes that…
122

Acknowledgments
123
Acknowledgments
In the name of God, most gracious, most merciful. Looking back, I feel so fortunate of
having had the opportunity to do my PhD at the Laboratory of Plant Breeding of
Wageningen University. I did my PhD at a very friendly atmosphere and received
continuous technical and emotional support during the last four years. It is a pleasure
to convey my gratitude to all people who helped me in achieving this momentous
task.
First of all, I would like to express my deep sense of gratitude to my supervisor Dr.
Rients Niks, for his patient guidance, encouragement and excellent advice
throughout this study. Dear Rients, you were always ready to help. Your energetic
personality and your enthusiasm for seeing new results have been stimulating me to
improve my scientific background and also to enjoy my research outputs. You were
so quick in commenting and correcting my written materials and your positive words
gave me new confidence in myself and my ability. I will never forget your efforts to
find a good living place for my family before our arrival and how happily you informed
me that you could find a beautiful family house, a case of too-nice-to-be-true! You
and your respectful wife, Marlien was very kind and supportive to me and my family.
Thank you for all your help and support.
I acknowledge my gratitude to Prof. Richard Visser for his valuable advices
throughout my study. Dear Richard, thank you for stimulating discussions and for
your comments and suggestions for the editing of my thesis.
My sincere thanks to Thierry Marcel for his fruitful collaboration and his kind
assistance in the research as well as for his valuable friendship. Dear Thierry, I have
really enjoyed working in the same group with you and have learned a lot from you. I
count you and your respectful wife, Hoa, among my best family friends.
Many thanks go to Hossein Jafary and his respectful wife Shirin. Dear Hossein, I
and my family will never forget your kind help and your warm and generous
hospitality during the first days of our stay in the Netherlands. You kindly helped me
to find my way in the Lab and easily shared your experience. Thank you for your
support, help and friendship.
My cordial thanks to Anton Vels for his kind assistance in setting up the field and
the greenhouse trials. Dear Anton, I was so pleased to hear that you are recovering
from your illness. You are indeed a great colleague.
I am thankful to Annie Marchal, Letty Dijker and Mariame Gada at the secretary of
Plant Breeding for dealing with administration and being always kind and helpful.
I had the great fortune of sharing ideas with every member of resistance group, but
more important, I have been honored with their friendship. It is a pleasure to mention

Acknowledgments
124
Sjaak van Heusden, Ben Vosman, Yuling Bai, Marieke Jeuken, Ningwen Zhang,
Stefano Pavan, Luigi Faino, Zheng Zheng, Erik den Boer and Freddy Yeo.
Furthermore, I would like to give special thanks to our Lab technicians, Petra vanden
Berg, Fien Meijer-Dekens, Koen Pelgrom and Martijn van Kaauwen for creating such
a nice atmosphere in the Lab.
I am thankful to my friendly officemates Paul Dijkhuis, Awang Maharijaya, Firdaus
Syarifin and Heungryul Lee. Extra thanks go to Awang for his kind help to design the
cover of my thesis. I am grateful to Nadeem Khan, Arnold Kuzniar, Anitha Kumari,
Sameer Joshi, Estelle Verzaux, Anoma Lokossou, Xingfeng Huang, Miqia Wang and
Shital Dixit for their friendship. Dear Shital thank you also for your kind acceptance to
be one of my paranimphs.
My special thanks also go to Dr. Patrick Schweizer from IPK, Germany for his
cooperation during the last two years and his great hospitality during my stay at IPK.
I had a great opportunity to meet and establish friendship with many Iranian
scientists in Wageningen. It is a pleasure to mention Dr. Mahmoud Otreshy, Rahim
Mehrabi, Ramin Roohparvar, Mostafa Karbasioun, Benyamin Houshyani, Farhad
Nazarian and their families. Dear Rahim and Farhad we shared various thoughts
during our tea breaks and sport activities in the late afternoon together with Hossein.
Thank you for your friendship. Furthermore I am grateful to Mahdi Arzanlou, Ali
Haghnazari, Reza Talebi, Sara Dezhsetan, Reza Tadayonirad, Leila Samiei, Nasim
Mansouri, Kambiz Baghalian and Alireza Babaei for their friendship. Dear Alireza, we
shared many funny moments during our trips together, I value our friendship very
much.
It is a pleasure to convey my gratitude to Alireza Seifi. Dear Alireza, thank you for
your encouragement and your kind help in the Lab as well as for creating such a
great friendship.
My special thanks go to Hedayat Bagheri, Mohammad Balali, Afshin Mehraban,
Mahmoud Tabib Ghaffary and their families. Dear Hedayat and Mohammad, we have
many good memories from our family trips and gatherings. I have found both of you
trustworthy and honest friends. Dear Afshin and Mahmoud thank you for being
always supportive to me and my family. You are very good friends. Furthermore, I
want to thank Mr. and Mrs. Raei for their support and their kindness to me and my
family.
I would like to take this opportunity to thank my colleagues at Agricultural Research
and Education Organization (AREO) and the Ministry of Science Research and
Technology (MSRT) of my home country, IRAN, for financial support and also dealing

Acknowledgments
125
with a lot of official tasks during the last four years. Many thanks go in particular to
the Staff Training Office of AREO and the scientific counselor of MSRT in Paris.
I would like to express my special thanks to my brother-in-law Reza Tajy and my
sister-in-law Maryam for their support and kindness.
My PhD study could never be fulfilled without the full support from my family. I
would like to express my profound gratitude to my beloved parents for their
continuous emotional support and prayers during my study abroad. You made a long
journey to visit us, indeed those were our best days in the Netherlands. To my lovely
daughter, Mahrokh and my lovely son, Pouria, I feel the joy and happiness as watch
you grow everyday. I am really proud of you! Finally I would like to express my
deepest sense of gratitude to my wife, Fatemeh for her moral support and patience
during my study. Dear Fatemeh, thank you for your generous love and persistent
confidence in me. I love you so much.
Reza Aghnoum
Wageningen, June 2009

Acknowledgments
123

About the author
127
About the author Reza Aghnoum was born on June 22, 1972 in
Tehran, Iran but he grew up in a small city in the
north east of Iran, where his parents had originally
come from. After finishing his high- school, Reza
started his academic studies in Plant Protection at
Ferdowsi University of Mashhad in 1991. He
followed a Master degree at the same university
in Plant Pathology in 1995.
Upon earning his Master degree in 1997, he was employed by the Agricultural
Research and Education Organization (AREO) as a cereal pathologist-breeder at
Mashhad Agricultural Research Center, where he worked for 7 years with a group of
cereal breeders aiming to breed high yielding and disease resistant wheat and barley
cultivars. In 2001 he passed the national examination and received a scholarship to
do a PhD abroad. In February 2005 he started his PhD program at the Laboratory of
Plant Breeding of Wageningen University under the supervision of Dr. Rients E. Niks
and worked on basal resistance of barley to powdery mildew as his PhD thesis. Reza
will join the scientific community of his home country, Iran, after finishing his PhD
studies in the Netherlands.
Reza Aghnoum
Khosaran Agricultural and Natural Resources Research Center
P.O.Box: 91735-488
Mashhad,
IRAN
Email: [email protected]

Acknowledgments
128

List of publications
129
List of publications
Marcel TC, Aghnoum R, Durand J, Varshney RK, Niks RE. 2007. Dissection of the
barley 2L1.0 region carrying the ‘Laevigatum’ quantitative resistance gene to leaf rust
using near isogenic lines (NIL) and subNIL. Molecular Plant–Microbe Interactions 20:
1604-1615.
Aghnoum R , Marcel TC, Johrde A, Pecchioni N, Schweizer P, Ni ks RE. Which
candidate genes are responsible for natural variation in basal resistance of barley to
barley powdery mildew? Submitted.
Aghnoum R , Niks RE. New perspectives to identify genes that determine non-host
resistance of barley to non-adapted Blumeria graminis forms. Submitted.
Abstracts
Aghnoum R, Marcel TC, Jafary H, Niks RE. Genetic diversity for quantitative
resistance of barley against powdery mildew. Plant & Animal Genomes XV
Conference. San Diego, California, USA. January 13-17, 2007.
Aghnoum R, Niks RE. Development of barley lines with extreme low and high level
of basal resistance to powdery mildew.The 10th International Barley Genetics
Symposium. Alexandria, Egypt, April 5-10, 2008.

Acknowledgments
130

Education certificate
131
Education certificate Education Statement of the Graduate School Experimental Plant Sciences Issued to: Reza Aghnoum Date: 16 June 2009 Group: Plant Breeding, Wageningen University and Re search Centre 1) Start-up phase date ► First presentation of your project Genetics of non-hypersensitive resistance of barley to powdery mildew Nov 1, 2005 ► Writing,or rewriting a project proposal Polygenic, non-hypersensitive resistance of barley to barley powdery mildew Feb-Aug 2005 ► Writing a review or book chapter ► MSc courses Genetic Analysis, Tools and Concepts (GEN-30306) Nov-Dec 2005 Breeding for Resistance (PBR 30306) Mar-Apr 2005 ► Laboratory use of isotopes Subtotal Start-up Phase 13.5 credits* 2) Scientific Exposure date ► EPS PhD student days EPS PhD student day 2005 in Radboud University Nijmegen Jun 2, 2005 SDV PhD student day 2006 in Paris Jun 9, 2006 EPS PhD student day 2006 in wageningen Sep 19, 2006 EPS PhD student day 2007 in wageningen Sep 13, 2007
1st Joint Retreat of the PhD Students in EPS
Oct 2-3, 2008
► EPS theme symposia EPS Theme 2 'Interactions between Plants and Biotic Agents, Utrecht University Jun 23, 2005 ► NWO Lunteren days and other National Platforms EPS - ALW Lunteren Apr 3-4, 2006 EPS - Lunteren meeting Apr 6-7, 2009 ► Seminars (series), workshops and symposia Molecular markers in yellow rust and powdery mildew: Dr. Xia-Yu Duan Nov 21, 2007 Plant Breeding Research day 2008, wageningen university, Jun 17, 2008 DNA demethylation in Arabidopsis:Prof.Jian-Kang Zhu Nov 3, 2008 Seminar: Prof. Nürnberger:Receptors in plant immunity Dec 17, 2008 ► Seminar plus ► International symposia and congresses Plant and Animal Genome XV Conference,USA Jan 13-17, 2007 Bioexploit meeting, Wageningen Mar 31-Apr 1, 2009 ► Presentations Poster: Plant and Animal Genome XV Conference,USA Jan 13-17, 2007 Poster:Plant Breeding Research day 2007, wageningen university, Sep 27, 2007 Poster:The 10th International Barley Genetics Symposium, Alexandria, Egypt Apr 5-10, 2008 Poster:Work shop "Natural Variation in plants", Aug 26-29, 2008 Oral presentation:Summerschool "On the Evolution of Plant Pathogen Interactions Jun 20, 2008
Poster:1st Joint Retreat of the PhD Students in EPS Oral presentation:EPS-Lunteren meeting
Oct 2-3, 2008 Apr 7, 2009
► IAB interview Sep 14, 2007 ► Excursions Excursion to Barenburg grass breeding company, Wolfheze, Oct 2,2007 Excursion to Bejo seeds and Royal van Zanten companies, Jun 11, 2008 Excursion to Syngenta company in Einkhuzen Sep 25, 2008
Subtotal Scientific Exposure 10.4 credits* 3) In-Depth Studies date ► EPS courses or other PhD courses Summerschool "Signalling in Plant Development and Plant Defense" Jun 21-23, 2006 Summerschool "On the Evolution of Plant Pathogen Interactions Jun 18-20, 2008 EPS PhD Work shop "Natural Variation in plants", Aug 26-29, 2008 ► Journal club Litterature discussion in Department of Plant Breeding 2005-2008 ► Individual research training Two weeks Lab training on gene expression analysis, IPK Germany Jan 25-Feb 06, 2009
Subtotal In-Depth Studies 9.0 credits* 4) Personal development date ► Skill training courses Working with EndNote 8, Wageningen UR Library Mar 8, 2005 Techniques for writing and presenting a scientific paper Oct 16-19, 2007 ► Organisation of PhD students day, course or confere nce ► Membership of Board, Committee or PhD council
Subtotal Personal Development 1.4 credits* TOTAL NUMBER OF CREDIT POINTS* 34.3
Herewith the Graduate School declares that the PhD candidate has complied with the educational requirements set by the Educational Committee of EPS which comprises of a minimum total of 30 credits * A credit represents a normative study load of 28 hours of study

Acknowledgments
132
This research has been carried out at the Laboratory of Plant Breeding of
Wageningen University and finantially was supported by the Agricultural Research
and Education Organization (AREO) and the Ministry of Science Research and
Technology of I. R. of Iran.
Thesis layout: by the author
Cover design: by Awang Maharijaya and the author
Front page: schematically represents the complexity of the papilla-based defense
Back page: abundant conidiation by the non-adapted wheat powdery mildew on the
barley line SusBgt SC
Printed by: Wöhrmann Print Service, Postbus 92, 7200 AB, Zutphen, the
Netherlands


















