
aureus Staphylococci on Staphylococcus aureus Growth and ...

University of Calgary
PRISM: University of Calgary's Digital Repository
Graduate Studies The Vault: Electronic Theses and Dissertations
2017
Bacteriocins of bovine non-aureus staphylococci
Carson, Domonique
Carson, D. (2017). Bacteriocins of bovine non-aureus staphylococci (Unpublished master's
thesis). University of Calgary, Calgary, AB. doi:10.11575/PRISM/25092
http://hdl.handle.net/11023/4124
master thesis
University of Calgary graduate students retain copyright ownership and moral rights for their
thesis. You may use this material in any way that is permitted by the Copyright Act or through
licensing that has been assigned to the document. For uses that are not allowable under
copyright legislation or licensing, you are required to seek permission.
Downloaded from PRISM: https://prism.ucalgary.ca

UNIVERSITY OF CALGARY
Bacteriocins of bovine non-aureus staphylococci
by
Domonique Carson
A THESIS
SUBMITTED TO THE FACULTY OF GRADUATE STUDIES
IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE
DEGREE OF MASTER OF SCIENCE
GRADUATE PROGRAM IN VETERINARY MEDICAL SCIENCES
CALGARY, ALBERTA
SEPTEMBER, 2017
© Domonique Carson 2017

ii
Abstract
The non-aureus staphylococci (NAS) species are among the most prevalent isolated from bovine
milk and have been reported to inhibit major mastitis pathogens, likely by producing
bacteriocins. This thesis is comprised of two sections, focusing on in vitro inhibition assays and
in silico identification of bacteriocin gene clusters and bacteriocin resistance genes in NAS and
Staphylococcus aureus, using isolates obtained from the Canadian Bovine Mastitis and Milk
Quality Research Network. The first part determined the inhibitory capability of 441 bovine
NAS isolates (comprising 25 species) against bovine S. aureus and human methicillin-resistant S.
aureus (MRSA) and determined the presence of bacteriocin biosynthetic gene clusters in NAS
whole genomes. Overall, 40 isolates from 9 species (S. capitis, S. chromogenes, S. epidermidis,
S. pasteuri, S. saprophyticus, S. sciuri, S. simulans, S. warneri, and S. xylosus) inhibited growth
of S. aureus in vitro; of which, 23 isolates (from S. capitis, S. chromogenes, S. epidermidis, S.
pasteuri, S. simulans, and S. xylosus) also inhibited MRSA. 105 putative bacteriocin gene
clusters encompassing 6 different subclasses (lanthipeptides, sactipeptides, lasso peptides, class
IIa, class IIc, and class IId) in 95 whole genomes from 16 species were identified. The second
part of the thesis determined the susceptibility of 139 bovine S. aureus isolates to a bacteriocin
producing S. chromogenes isolate and identified and described the distribution of genes
potentially associated with susceptibility and resistance in S. aureus whole genomes. Overall, 90
S. aureus isolates (65%) were resistant to inhibition by the S. chromogenes isolate. We identified
77 genes that were associated with an isolate being resistant. We also identified 76 genes that
were associated with an isolate being susceptible to the S. chromogenes. Bacteriocin
susceptibility and resistance seems to be linked to a large number of genes, the majority of which

iii
are annotated as hypothetical proteins and will need further assessment to determine their role in
S. aureus susceptibility. Overall, bacteriocins may be a potential source of novel antimicrobials
and this thesis represents the foundation to explore novel NAS bacteriocins.

iv
Acknowledgements
Firstly, I would like to thank my supervisors Herman Barkema and Jeroen De Buck for
all of their patience and support during this project. Thank you to Herman for the opportunity to
attend so many conferences and speak about my research. Thank you to Jeroen for all your
direction with my projects. I also would like to thank my committee members for their support
and Dr. John Kastelic for his edits of my manuscript and for his scientific writing courses.
I would also like to thank Uliana Kanevets and Aaron Lucko for their help in the lab.
Thank you to Matthew Workentine for all his help with bioinformatics.
Thank you to the original CNS crew, Larissa Condas and Diego Nobrega for everything.
Larissa, thank you for showing me the CNS ropes, I came into this project with little lab
experience and your expertise was essential for my success. Your endless encouragement and
honest life talks during your time here were also so appreciated- “Everything is AWESOME”.
Diego, thank you for all your help, from whole genome sequencing to talking through absolutely
everything with me, I could not have done it without you. I am also so grateful for the rest of the
CNS crew that has joined us. Thank you Ali Naqvi for helping me with statistics and thank you
Ana Paula Monteiro Alves for being my conference buddy and letting me be the absolute
introvert that I am. Lastly, thank you to Sohail Naushad, without you I would have not been able
to do this. Thank you for the countless conversations about genes, bioinformatics, and how to
interpret my results. Thank you for the coffee dates and the pep talks and for always being on my
team. Thank you for all of your help with organizing genomes, creating trees, running jobs, and
giving me feedback whenever I ask.

v
Thank you to all my fellow graduate students. Thank you to Emily Morabito for
beginning this program at the same time as me and becoming such a huge support as we figured
this all out together. Thank you to Caroline Corbett for being a rock in all of this. You were
always there to talk and were always able to provide valuable feedback on my research. Thank
you for being such a good friend to me. Also thank you Casey Jacobs for always being there for
me (usually with a beer and chocolate ready). Our daily dog walks became so special and
without them life would have been much more difficult.
I would also like to thank Dr. Keliesha Roth and Amy Stanley for being the best friends a
girl could ask for in this life. Keliesha, thank you for always making me feel like I could do this.
Thank you for letting me come work at your house, for taking such good care of me after my
surgery this spring, for always taking the dogs for me whenever they needed somewhere to go,
and for bringing Took into my life. You are one of the most amazing people I’ve ever met and a
huge reason that I am the person I am today. Amy, thank you for being the most understanding
and amazing human. You supported me through this every way a person possibly could… you
ran away to the mountains with me when I needed that, you exercised the dogs for me when I
couldn’t, you provided hours of deep conversations about life, and you have always just wanted
what is best for me. Thank you to you both.
Thank you to Reid Anderson. You came into my life on that airplane at the exact moment
I needed you to. You have been my strength in times of weakness, my never ending support
system, my inspiration to keep working towards my goals, and (most importantly) the best dog
dad I could ever ask for. You mean more to me than you’ll ever know.
Lastly, thank you to my family for the endless support and understanding. Thank you for
answering all of my phone calls and always giving me words of encouragement. It has been hard

vi
to be so far apart but it never felt too far. Thank you for instilling in me a passion for learning
and for sticking things out.

vii
Dedication
To my family,
in particular my parents
and my chosen family
who have always supported me
and to my dogs
who have kept me sane.

viii
Table of Contents
Abstract .......................................................................................................................................... ii
Acknowledgements ...................................................................................................................... iv
Dedication .................................................................................................................................... vii
Table of Contents ....................................................................................................................... viii
List of Tables ................................................................................................................................ xi
List of Figures and Illustrations ................................................................................................ xii
List of Symbols, Abbreviations and Nomenclature ................................................................ xiv
Preface .......................................................................................................................................... xv
Chapter One: General Introduction ........................................................................................... 1
1.1 Mastitis in the Canadian dairy industry ........................................................................... 1
1.2 Non-aureus staphylococci ................................................................................................... 2
1.3 Bacteriocins ......................................................................................................................... 4
1.4 Classification of bacteriocins ............................................................................................. 5
1.4.1 Lanthipeptides ................................................................................................................ 7
1.4.2 Sactipeptides ................................................................................................................ 12
1.4.3 Lasso Peptides .............................................................................................................. 14
1.4.4 Class IIa Bacteriocins .................................................................................................. 15
1.4.5 Class IIb Bacteriocins .................................................................................................. 16
1.4.6 Class IIc Bacteriocins .................................................................................................. 17
1.4.7 Class IId Bacteriocins .................................................................................................. 18
1.5 Immunity genes and cross immunity .............................................................................. 19
1.6 Bacteriocin discovery and purification ........................................................................... 20
1.7 In silico screening .............................................................................................................. 23
1.8 Applications of bacteriocins ............................................................................................. 26
1.9 Bacteriocin resistance ....................................................................................................... 31
1.10 Thesis outline ................................................................................................................... 34
1.10.1 Bacteriocins of non-aureus staphylococci isolated from bovine milk ....................... 34

ix
1.10.2 Identifying putative bacteriocin resistance genes in Staphylococcus aureus whole
genomes ................................................................................................................................ 35
Chapter Two: Bacteriocins of non-aureus staphylococci isolated from bovine milk ........... 36
2.1 Abstract .............................................................................................................................. 36
2.2 Introduction ....................................................................................................................... 38
2.3 Materials and methods ..................................................................................................... 40
2.3.1 Isolates ......................................................................................................................... 40
2.3.2 Phenotypic testing ........................................................................................................ 41
2.3.3 Effect of proteinase K on inhibition ............................................................................. 41
2.3.4 Whole genome sequencing, assembly, and annotation ................................................ 42
2.3.5 Screening of genomes for bacteriocin clusters ............................................................ 43
2.3.6 BLAST ......................................................................................................................... 44
2.3.7 Genome comparison .................................................................................................... 44
2.3.8 Precursor gene alignments ........................................................................................... 45
2.4 Results ................................................................................................................................ 45
2.4.1 Phenotypic testing ........................................................................................................ 45
2.4.2 Effect of proteinase K on inhibition ............................................................................. 46
2.4.3 Screening of genomes for bacteriocin clusters ............................................................ 46
2.5 Discussion .......................................................................................................................... 51
2.6 Conclusions ........................................................................................................................ 58
Chapter Three: Identifying putative bacteriocin resistance genes in Staphylococcus aureus
whole genomes ............................................................................................................................. 79
3.1 Abstract .............................................................................................................................. 79
3.2 Introduction ....................................................................................................................... 81
3.3 Materials and methods ..................................................................................................... 83
3.3.1 Isolates ......................................................................................................................... 83
3.3.2 Phenotypic testing ........................................................................................................ 84
3.3.3 Whole genome sequencing, assembly, and annotation ................................................ 84
3.3.4 Screening of genomes for bacteriocin clusters ............................................................ 86
3.3.5 Screening of genomes for immunity related genes ...................................................... 86

x
3.4 Results ................................................................................................................................ 87
3.4.1 Phenotypic test ............................................................................................................. 87
3.4.2 Screening of genomes for bacteriocin clusters ............................................................ 87
3.4.3 Screening of genomes for putative resistance related genes ........................................ 88
3.5 Discussion .......................................................................................................................... 90
3.6 Conclusions ........................................................................................................................ 95
Chapter Four: Summarizing discussion ................................................................................. 102
4.1 NAS phenotypic testing .................................................................................................. 102
4.2 Bacteriocin clusters in NAS ............................................................................................ 105
4.3 Staphylococcus aureus susceptibility ............................................................................. 107
4.4 Potential immunity genes ............................................................................................... 107
4.5 Conclusions and future research ................................................................................... 108

xi
List of Tables
Table 2-1 Bacteriocin gene clusters identified in bovine non-aureus staphylococci genomes
and inhibitory phenotypes tested against Staphylococcus aureus and MRSA. .................... 60
Table 3-1 Bacteriocin gene clusters identified in bovine Staphylococcus aureus and in vitro
susceptibility to a lanthipeptide-producing S. chromogenes. ................................................ 96
Table 3-2 The number and percentage of S. aureus isolates containing a putative bacteriocin
resistance gene associated with an isolate being resistant to a lanthipeptide encoding S.
chromogenes. ........................................................................................................................ 98
Table 3-3 The number of and percentage of S. aureus isolates containing genes associated
with an isolate being susceptible to a lanthipeptide encoding S. chromogenes. ................... 99

xii
List of Figures and Illustrations
Figure 2-1 Distribution of bacteriocin biosynthetic gene clusters in species of non-aureus
staphylococci isolated from milk of Canadian dairy cows displayed on the phylogenetic
tree from Naushad et al (2017). ............................................................................................ 63
Figure 2-2 Biosynthetic gene clusters and LanA alignments of type A lanthipeptides
identified in non-aureus staphylococci isolated from milk of Canadian dairy cows. .......... 64
Figure 2-3 Phylogenetic tree of Staphylococcus epidermidis isolates from bovine milk
indicating growth inhibition against Staphylococcus aureus and genomically identified
bacteriocin clusters. ............................................................................................................... 66
Figure 2-4 Biosynthetic gene clusters and LanA alignments of type B lanthipeptides with a
single LanM identified in non-aureus staphylococci isolated from milk of Canadian
dairy cows. ............................................................................................................................ 68
Figure 2-5 Biosynthetic gene clusters and LanA alignments of type B lanthipeptides with
dual lanM enzymes identified in non-aureus staphylococci isolated from milk of
Canadian dairy cows. ............................................................................................................ 70
Figure 2-6 Biosynthetic gene clusters and alignments of precursor peptides from
sactipeptides identified in non-aureus staphylococci isolated from milk of Canadian
dairy cows. ............................................................................................................................ 72

xiii
Figure 2-7 Biosynthetic gene clusters and alignments of precursor peptides from the lasso
peptide identified in non-aureus staphylococci isolated from milk of Canadian dairy
cows. ..................................................................................................................................... 73
Figure 2-8 Biosynthetic gene clusters of Class II double glycine leader peptide bacteriocins
identified in non-aureus staphylococci isolated from milk of Canadian dairy cows. .......... 74
Figure 2-9 Biosynthetic gene clusters and precursor alignments of Class IIc circular
bacteriocins identified in non-aureus staphylococci isolated from milk of Canadian
dairy cows. ............................................................................................................................ 75
Figure 2-10 Biosynthetic gene clusters and precursor alignments of Class IId lactococcin-like
bacteriocins identified in non-aureus staphylococci isolated from milk of Canadian
dairy cows. ............................................................................................................................ 77
Figure 3-1 Maximum Likelihood phylogenetic tree of Staphylococcus aureus isolates with
isolates resistant to a bacteriocin producing S. chromogenes indicated with a black dot. .... 97
Figure 3-2 LanA alignments and phylogenetic tree of type A lanthipeptides identified in S.
aureus isolated from milk of Canadian dairy cows. ........................................................... 100

xiv
List of Symbols, Abbreviations and Nomenclature
Symbol Definition aa Amino acid ABC transporter ATP binding cassette transporter BMSCC Bulk milk somatic cell count C-terminal Carboxyl-terminus of protein C-domain Carboxyl-terminus of signal peptide CBMQRN Canadian Bovine Mastitis and Milk Quality Network CNS Coagulase negative staphylococci CM Clinical mastitis GRAS Generally regarded as safe IMI Intramammary infection kDa Molecular mass unit, kilo Dalton LAB Lactic acid bacteria MIC Minimal inhibition concentration MDR Multi drug resistant NAS Non-aureus staphylococci OD600 Optical density measured at a wavelength of 600nm NSR Nisin resistance protein PBS Phosphate buffered saline PCR Polymerase chain reaction pH Potential Hydrogen SCC Somatic cell count TMD Transmembrane domain WGS Whole genome sequencing

xv
Preface
This dissertation consists of two manuscripts – the first manuscript is in press, planned
for issue 17 of Applied and Environmental Microbiology in September, 2017. For both
manuscripts, the first author was involved with study concept and design, acquisition of isolates
and data, laboratory analysis, analysis and interpretation of data, drafting of the manuscript, and
critical revision. This was done under the guidance of the senior author, supervisor and co-
supervisor. All authors provided critical reviews of the manuscripts and contributed intellectual
content. Both manuscripts were reproduced in their entirety as chapters in this dissertation.
Manuscript I) Domonique Carson, Herman W Barkema, Sohail Naushad, and Jeroen De Buck.
Bacteriocins of non-aureus staphylococci isolated from bovine milk. Accepted for publication in
Applied and Environmental Microbiology, 83 (17).
Manuscript II) Domonique Carson, Sohail Naushad, Herman W Barkema, and Jeroen De Buck.
Identifiying putative bacteriocin immunity genes in S. aureus whole genomes.

1
Chapter One: General Introduction
1.1 Mastitis in the Canadian dairy industry
Mastitis, inflammation of the mammary gland, is predominantly caused by microbial
infection in the mammary gland and costs Canadian dairy producers an estimated $400 million
per year due to direct losses in milk yield and treatment costs, and indirect losses, such as
discarded milk during treatment and future milk yield reduction (Rollin, Dhuyvetter, and
Overton 2015; Bradley 2002). Mastitis causing pathogens can be divided into major and minor
pathogens, as well as into environmental or contagious, depending on their transmission patterns
(Olde Riekerink et al. 2008). The most common contagious major pathogens are Staphylococcus
aureus, Streptococcus uberis, Streptococcus dysgalactiae, and Streptococcus agalactiae,
whereas as Escherichia coli, and Klebsiella spp. are the most frequently isolated environmental
bacteria causing mastitis. Intramammary infection (IMI) of these pathogens can result in clinical
mastitis (CM), characterized by decrease in milk production and quality or by udder
abnormalities. Cows can also have subclinical mastitis (SCM), presenting with inflammation of
the udder determined by an increase in somatic cell count (SCC). Staphylococcus aureus and E.
coli are the most frequently isolated bacteria from CM on dairy farms in developed countries.
Staphylococcus aureus often leads to persistent IMIs that can influence culling decisions (Olde
Riekerink et al. 2006; Bradley 2002).
Traditionally, research and control programs have focused on decreasing the prevalence
of mastitis caused by major contagious pathogens, and producers and herd veterinarians have
been quite successful in this regard. Improvements have been made in hygiene, housing

2
management, milking management, and with timely treatment of infections has led to a
distribution shift of mastitis causing pathogens (Piepers et al. 2007). Non-aureus staphylococci
(NAS), generally considered minor mastitis pathogens, are the most commonly isolated bacteria
from the udder (Piepers et al. 2007; Pitkälä et al. 2004; Sampimon et al. 2009).
Even with the significant improvement being made on farm, mastitis is still a very
important disease for both the producer and consumer as it affects milk production, milk quality,
animal welfare, and can have public health concerns. Mastitis is the leading cause of antibiotic
usage for lactating dairy cows, for both the treatment and prevention of the disease (Saini et al.
2012; Oliveira and Ruegg 2014). In addition to treating any apparent CM in the lactating herd,
blanket dry cow therapy contributes heavily to the amount of overall of antibiotic use on farm
and high levels of antibiotic use are associated with higher levels of antibiotic resistant bacteria.
Consequently, consumer demands are driving the industry towards less antibiotic use, as evident
by the ever-increasing national production level of organic milk (Canadian dairy information
center, http://www.dairyinfo.gc.ca/pdf/org-bio_can_e.pdf). Although, the organic farm
regulations may result in producers being less likely to treat sick animals with antibiotics as
treated cows may be prohibited from returning to the milking herd, which may lead to welfare
implications (Barkema et al. 2015). Thus, in addition to judicious use of antibiotics, the need for
safe and effective alternatives is undeniable.
1.2 Non-aureus staphylococci
Staphylococci belong to the phylum Firmicutes, order Bacillales, class Bacilli, family
Staphylococcaceae, genus Staphylococcus. They are a group of Gram-positive and catalase-

3
positive bacteria that are round and form grape like structures. The heterogeneous genus
comprises over 50 species that are teat skin opportunists and generally considered minor udder
pathogens (White et al. 1989). NAS were originally differentiated from S. aureus based on the
ability of S. aureus to coagulate plasma (coagulase-positive), and therefore were named
coagulase-negative staphylococci (CNS). Originally, it was thought that only S. aureus was
pathogenic. At this point, several members of the group (e.g. S. agnetis) are now identified as
coagulase-variable (Dos Santos et al. 2016); it is, therefore, more appropriate that the group be
termed non-aureus staphylococci in order to encompass both the coagulase-negative and
coagulase-variable members.
Twenty-five NAS species have been isolated from cows, with S. chromogenes being the
most frequently isolated species from both milk and skin (Vanderhaeghen et al. 2015). While
NAS can be the cause of clinical mastitis, infection commonly results in subclinical mastitis,
raising the milk SCC of the infected quarter (Taponen et al. 2006). Therefore, a high prevalence
of NAS IMI can contribute to an increase in bulk milk SCC (BMSCC) in herds with a low
BMSCC (Schukken et al. 2009). Consequently, controlling NAS could allow producers to
further lower BMSCC. On the other hand, it has been reported that NAS IMI can result in
increased milk production when compared with uninfected cows (Schukken et al. 2009).
Interestingly, several studies have found that NAS have a protective effect against IMI by
major mastitis pathogens (De Vliegher et al. 2004; Matthews, Harmon, and Smith 1990). The
first challenge study using S. chromogenes reported a protective effect when 53% of S.
chromogenes-infected quarters were protected from a S. aureus challenge (Matthews, Harmon,
and Smith 1990). Later, De Vliegher et al. (2004) reported that two of 10 S. chromogenes
isolates were able to inhibit the in vitro growth of all S. aureus, S. dysgalactiae, and S. uberis,

4
but none of the tested E. coli isolates. On the other hand, field studies have reported no protective
effects from staphylococcal infections against environmental pathogens (Hogan et al. 1988) and
that NAS IMI was neither a risk factor or a protective against mastitis caused by S. aureus or S.
uberis (Zadoks et al. 2001). The inconsistent results regarding if NAS are protective or not
against IMI from major pathogens are likely due to undifferentiated NAS species and genotypes,
which may have different pathogenicity and effects in the udder.
When looking at the effects of NAS on udder health at a species level, certain species
have been shown to inhibit the growth of mastitis pathogens in vitro. NAS strains from Brazilian
bovine mastitis cases, including Staphylococcus epidermidis, Staphylococcus simulans,
Staphylococcus saprophyticus , Staphylococcus hominis , and Staphylococcus arlettae, inhibited
growth of indicator species Corynebacterium fimi (Nascimento et al. 2005). The isolated
antimicrobial substances were considered to be bacteriocins due to sensitivity to proteolytic
enzymes (Nascimento et al. 2005). Recently, Braem et al. (2014) identified NAS strains (from 6
species) that inhibited S. aureus, S. uberis, and S. dysgalactiae and an inhibitory substance (from
an inhibiting S. chromogenes) was isolated and identified to be a nukacin-like bacteriocin
(Braem et al. 2014). Thus, it is possible that the variation that was seen when examining NAS
inhibition was due to different species and isolates being able to produce bacteriocins, which are
antimicrobial peptides.
1.3 Bacteriocins
Bacteriocins are ribosomally synthesized and (generally) post-translationally modified
peptides (RiPPs) that are produced by bacteria to kill other bacteria, thus bacteriocin production

5
gives the producing strain an advantage in certain ecological niches, allowing them to compete
for common resources (Parada et al. 2007). The bacteriocins generally exert their bacteriocidal
activity against a narrow spectrum of bacteria. The first antimicrobial peptide compounds
identified, termed colicins, were produced by E. coli and now, bacteriocins have been found in
all major lineages of bacteria (Riley and Wertz 2002). The lactic acid bacteria (LAB) are
abundant producers of bacteriocins and LAB have been used for centuries to ferment foods
(Riley and Wertz 2002). Bacteriocins have the potential to be used as antimicrobial peptides in
the food and health industries. Nisin, a bacteriocin produced by Lactococcus lactis, was
discovered in 1947 (Mattick, Hirsch, and Berridge 1947) and is now the most widely used
bacteriocin.
Bacteriocins produced by Gram-positive bacteria are usually smaller than 8kDa and they are
usually pore forming. The producer contains specific proteins, which confer host immunity
against the bacteriocin (Cotter, Ross, and Hill 2013). Bacteriocins are generally classified into
two main groups, the modified peptides and the unmodified peptides (Cotter, Ross, and Hill
2013).
1.4 Classification of bacteriocins
A variety of classification systems for bacteriocins exist, based on chemical structure,
molecular mass, enzymatic susceptibility, mode of action, genetic mechanisms, thermo stability,
producing strains, spectrum of activity, or presence of post translationally modified residues
(Klaenhammer 1993). This assortment of classification schemes can result in bacteriocins being

6
assigned to multiple different classes (if they were discovered by different groups at the same
time), according to the classification scheme each group choses.
Historically, bacteriocins were first grouped in four main classes (Klaenhammer 1993).
Class I were the lantibiotics (subdivided into two groups), class II were the non-modified
peptides (subdivided into three groups), class III were the large and heat labile bacteriocins, and
class IV were the bacteriocins proposed to form large complexes with macromolecules
(Klaenhammer 1993). In 2005, Cotter, Hill, and Ross (2005) proposed a classification system
that recognized just two classes of bacteriocins: class I, the post translationally modified
lantibiotics and class II, the non-modified bacteriocins. The class II bacteriocins were broken
into four categories: IIa, small heat stable peptides, IIb, the two component bacteriocins, IIc, the
circular bacteriocins, and IId, the other class II bacteriocins, including sec-dependent and
leaderless bacteriocins (Cotter, Hill, and Ross 2005). Subsequently, Heng and Tagg (2006)
suggested that the class IIc bacteriocins, the circular bacteriocins, should be grouped as their own
class, class IV bacteriocins, and that the large bacteriolysins (which are non-bacteriocin lytic
proteins that were not included in the Cotter classification scheme) be classified as class III
bacteriocins.
In 2013, Arnison et al. (2013) proposed universal nomenclature for RiPPs, which has
been generally accepted (Cotter, Ross, and Hill 2013). This scheme includes all modified
bacteriocins grouped as class I, and the unmodified or circular bacteriocins grouped as subclasses
of class II bacteriocins (Arnison et al. 2013). Interestingly, a study in 2010 classified 107
bacteriocins according to amino acid structure, resulting in 12 different bacteriocin groups, each
representing a distinct branch on the phylogenetic tree and containing a conserved motif (Zouhir
et al. 2010). This study represented a unique classification scheme, although one that has not

7
been adapted by the bacteriocin researchers community. In 2016, Alvarez-Sieiro et al. (2016)
proposed a scheme modified from Arnison et al. (2013), where circular bacteriocins are included
in class I modified bacteriocins and the bacteriolysins are included as class III bacteriocins. In
this thesis, to remain consistent with the proposed universal nomenclature, we will follow the
two-class classification scheme recommended by Arnison et al. (2013).
1.4.1 Lanthipeptides
Traditionally termed lantibiotics (lanthionine-containing antibiotic), lanthipeptides are
characterized by the presence of several uncommon amino acids, including meso-lanthionine and
3-methyllanthionine, that are a result of post-translational modifications (Bierbaum et al. 1996).
The name lanthipeptides was changed to encompass the non-antibiotic peptides (type C and D
lanthipeptides) from the same biosynthetic origin (Goto et al. 2010), although the majority of
lanthipeptides are either type A or type B, which exert antimicrobial activity (Bierbaum and Sahl
2009). Lanthipeptides are the most common RiPPs in available genomes, with more than 95
lanthipeptides from Gram-positive bacteria having been isolated and described (Dischinger, Basi
Chipalu, and Bierbaum 2014). Genome mining has identified many more potential compounds
waiting to be characterized (Knerr and van der Donk 2012), many of which are closely related
and likely have common peptide ancestors. The most well known lanthipeptide, nisin, which was
isolated from L. lactis in 1947 (Mattick, Hirsch, and Berridge 1947), has been used in the food
preservation industry for over 50 years.
Lanthipeptides are the bacteriocins most frequently isolated from Staphylococcus species,
thus, there are many well-characterized “Staphylococcin” examples. The first to be discovered
and described were pep5 (Sahl et al. 1985) and epidermin (Allgaier et al. 1986) from S.

8
epidermidis and gallidermin from S. gallinarum (Schnell et al. 1989). Multiple additional type-A
lanthipeptides have since been identified. Staphylococcus cohnii produces staphyloccocin T,
which has an identical sequence to gallidermin and is active against a broad spectrum of Gram-
positive bacteria (Furmanek et al. 1999). Staphylococcus hyicus produces hyicin 3862 (Fagundes
et al. 2011), a bacteriocin likely related to epidermin and Bsa (a bacteriocin produced by S.
aureus) (Daly et al. 2010) and represents the first bacteriocin to be identified in S. hyicus. The
genetic organization of hyicin 3862 was recently elucidated and showed 91% identity with Bsa
but was found to have a broader spectrum of activity than Bsa (Fagundes et al. 2017). Along
with the bacteriocin pep5, S. epidermidis produces other lanthipeptides that are closely related to
pep5 including epilancin K7 (van de Kamp et al. 1995), epicidin 280 (Heidrich et al. 1998), and
epilancin 15X (Ekkelenkamp et al. 2005). Staphylococcus warneri produces a type-AII
lanthipeptide, nukacin ISK-1 (Sashihara et al. 2000), which is a member of the lacticin 481 group
(Bierbaum and Sahl 2009). Nukacin-like bacteriocins have also been identified in S. simulans
(Ceotto et al. 2010), S. hominis (Wilaipun et al. 2008), and S. chromogenes (Braem et al. 2014).
Lastly, S. aureus produces a two-component lanthipeptide, C55 (Maduwe, Sahl, and Tagg 1999).
1.4.1.1 Genetic Organization
Genes related to lanthipeptide synthesis are generically named with the locus symbol lan,
where each characterized bacteriocin has its individual naming system, for example nis for nisin.
All lanthipeptides have a precursor peptide, LanA, which contains both a leader peptide and a
core peptide, of which the former is cleaved off to yield the active peptide. The leader peptide is
thought to play roles in posttranslational modification, immunity, and export (Arnison et al.
2013). The mature lanthipeptide results from posttranslational modifications to the core peptide

9
by one or more enzymes, also encoded on the operon. There is a small subclass of lanthipeptides
that require two LanA peptides, A1 and A2, for complete antimicrobial activity. These low
identity peptides each require their own posttranslational modifications by separate modification
enzymes to be able to work synergistically together outside of the cell (Bierbaum and Sahl
2009).
The modification enzymes, which are responsible for creating thioether cross-links (by
dehydrating the serine and threonine residues followed by addition of a thiol of a cysteine
residue) and for cleaving off the leader peptide either during or before export, are the basis for
dividing lanthipeptides into their groups (Knerr and van der Donk 2012). Type A lanthipeptides
contain lanB and lanC genes, where the dehydration of the serine and threonine residues to
dehydroalanine (Dha) and dehydrobutyrine (Dhb) residues in the pre-peptide are completed by a
dehydratase, LanB, and the thioether crosslinks are formed on the dehydrated amino acids by a
cyclase, generically called LanC. For type B lanthipeptides, a bifuncational enzyme, LanM,
carries out the dehydration and cyclisation. The C terminal of the LanM synthetase shares
homology with the LanC cyclases of the type A lanthipeptides (Blin et al. 2014; Asaduzzaman
and Sonomoto 2009).
Lanthipetides also contain LanP serine proteases, which cleave the leader peptide from
the core peptide, and a LanT, which is responsible for exporting the mature peptide. Immunity
genes are also located on the operon to provide the producer with self-immunity. In
lanthipeptides, immunity is conveyed with lanI, or the lanFE(G) immunity cluster. The LanI
protein is thought to function by interception or target shielding (Stein et al. 2005) or by
sequestering the bacteriocin on the bacterial cell wall membrane, as is the case with NisI from
nisin. In contrast, the LanFEG proteins work by removing the peptide via the dedicated ABC

10
transporter (Stein et al. 2005). Additional genes found on the gene clusters are lanR and lanK
which are both involved in lanthipeptide regulation by a quorum sensing system (Chatterjee et al.
2005). Presence of the active lanthipeptide leads to a signalling cascade initiated by the LanK
histidine kinase, followed by activation of the LanR response regulator to activate biosynthesis
of the bacteriocin (Dischinger, Basi Chipalu, and Bierbaum 2014).
1.4.1.2 Mode of Action
Type A lanthipeptides, like nisin, which are linear positively charged peptides, have dual
modes of action where they inhibit cell wall biosynthesis and form pores that mainly act upon
Gram-positive bacteria (Bierbaum and Sahl 2009). In the specific case of nisin, which has been
studied extensively, the cell wall precursor lipid II is the target and upon binding to lipid II the
nisin inhibits peptidoglycan synthesis. Additionally, upon binding, nisin is able to insert itself
into the membrane to form stable pores consisting of eight nisin and four lipid II molecules
(Hasper, de Kruijff, and Breukink 2004). This dual mode of action makes nisin very potent and
makes resistance harder to acquire (Bastos, Coelho, and Santos 2015). Other lanthipeptides, like
epidermin and gallidermin that are too short to span the lipid bilayer to form pores, also bind to
lipid II, yet are still able to kill target bacteria, indicating that they have other lipid II mediated
mechanisms (Bastos, Coelho, and Santos 2015). The proposed mode of action is that the
bacteriocin sequesters the lipid II away from its functional location, thus blocking cell wall
synthesis (Hasper et al. 2006). Targets have yet to be identified for pep5 and epilancin K7
(Draper et al. 2015). Type AII lanthipeptides (lacticin 481 and nukacin ISK-1 groups) consist of
an N-terminal linear region and a C-terminal globular region (Chatterjee et al. 2005), and it is
thought that their structures may prevent them from forming pores (Islam, Nagao, et al. 2012).

11
For certain bacteriocins in this group the positive lysine residues in the linear region are essential
for binding to the cell membrane, although not all members of this group have positively charged
residues (Islam, Nagao, et al. 2012), indicating additional areas of the bacteriocin that facilitate
antimicrobial activity. One member of this group, lacticin 481 binds lipid II leading to the
inhibition of the transglycosylation step, therefore stopping cell wall synthesis (Knerr et al.
2012). Nukacin ISK-1 contains a conserved region similar to the lipid II binding region of
mersacidin (a type B lanthipeptide), suggesting a similar mode of action. Preliminary studies
seem to indicate that nukacin ISK-1 indeed has a bacteriostatic mode of action caused by binding
to lipid II with its conserved A ring region to inhibit cell wall synthesis (Islam, Nishie, et al.
2012). This has been recently confirmed along with the illumination of other important residues
in the peptide (Elsayed et al. 2017). Therefore, type AII lanthipeptides are not pore formers but
interact with lipid II on the cell wall surface and inhibit cell wall biosynthesis, resulting in
termination of cell growth. Lacticin 3147, a two peptide lanthipeptide, has a proposed three step
mechanism where the a-peptide binds to lipid II inducing a conformational change in the a-
peptide which the b-peptide recognizes and binds to, resulting in insertion into the membrane,
therefore forming a pore (Srinivas et al. 2012). The type B lanthipeptides (e.g. mersacidin) are
globular peptides with no charge, or are negatively charged (Islam, Nagao, et al. 2012). These
lanthipeptides act by inhibiting cell wall biosynthesis and are also not pore formers. Mersacidin
binds to lipid II and interferes with the transglycosylation step of peptidoglycan synthesis (Brötz
et al. 1997). Mersacidin, as the name indicates, is active against methicillin-resistant S. aureus
(MRSA) (Kruszewska et al. 2004) and is also able to inhibit the growth of vancomycin-resistant
Enterococcus faecium strains even though mersacidin and vancomycin have the same target.

12
This is because mersacidin binds to a different site on the lipid II (Brötz et al. 1997; van Heel,
Montalban-Lopez, and Kuipers 2011).
1.4.2 Sactipeptides
Sactipeptides, formerly known as sactibiotics, are a small class of RIPPs that are
characterized by the presence of at least one thioether bond between the cysteine sulphur and the
a-carbon of an acceptor amino acid. They are all relatively hydrophobic with a 3D hairpin like
structure (Arnison et al. 2013). Sactipeptides originally had only been characterized in Bacillus
species, though recent genome approaches have identified them in Clostridium species (Murphy
et al. 2011; Haft, Basu, and Mitchell 2010). Subtilosin-A, produced by Bacillus subtilis 168, is
the best characterized sactipeptide (Babasaki et al. 1985) and has been reported to have
antimicrobial activity against both Gram-positive and Gram-negative pathogens (Shelburne et al.
2007). One member of the sactipeptide family, Thuricin CD, is effective against Clostridium
difficile, yet it has a narrow spectrum of activity thus not impacting the host commensal flora
(Rea et al. 2010). A sactipeptide was recently identified in S. hyicus, named hyicin 4244 (Duarte
et al. 2017), making it the first sactipeptide characterized from Staphylococcus.
1.4.2.1 Genetic Organization
The biosynthetic gene clusters of sactipeptides typically include a precursor peptide,
along with one radical SAM enzyme per precursor, a putative protease, and two potential export
and immunity proteins (Fluhe and Marahiel 2013). The nomenclature for these clusters is based
off the thuricin CD gene cluster (Rea et al. 2010), where “A” is the precursor peptide, “C” and
“D” refer to the radical-SAM protein(s), “F” and “G” indicate the ABC-transporters, and “P” is

13
used to identify any yet unidentified proteases in the cluster (Arnison et al. 2013). The SAM
enzyme is responsible for the sulphur to a-carbon crosslinks, formed from linking the sulphur of
a cysteine residue to the a-carbon of an acceptor amino acid (Fluhe and Marahiel 2013), and has
been used as the target for genome mining for novel sactipeptides (Murphy et al. 2011).
1.4.2.2 Mode of Action
The precise modes of action of the characterized sactipeptides are not well understood,
although they seem to be able to interact with and disrupt target cell walls (Thennarasu et al.
2005; Wang et al. 2014). It has been demonstrated that thurincin H does not cause cell membrane
permeability or cell wall lysis, although it does decrease cell viability (Wang et al. 2014).
Subtilosin A seems to have variable modes of action, depending on the target organism. In a
study looking at subtilosin A’s effect on Gardnerella vaginalis, the bacteriocin resulted in an
immediate depletion of the cells pH and triggered an efflux of ATP, suggesting that subtilosin A
forms pores, leading to cell death (Sutyak Noll, Sinko, and Chikindas 2011). When investigating
subtilosin A’s effects on Listeria monocytogenes, there was no efflux of ATP and only minor
effects on pH and transmembrane potential, which the authors suggest indicates subtilosin A
interacts with the cell membrane, causing intracellular damage leading to cell death (Kuijk, Noll,
and Chikindas 2012). This mode of action was previously suggested, as evidence showed
subtilosin A adopts a partially buried orientation in the lipid bilayer, inducing conformational
changes and leading to membrane permeability (Thennarasu et al. 2005).

14
1.4.3 Lasso Peptides
Lasso peptides get their name from their structure, which resembles a lariat with its
threaded configuration. Based on their structure and experimental reports, these peptides are
highly resistant to proteases and denaturing agents, which make them a topic of much interest
(Arnison et al. 2013), although not all lasso peptides seem to follow this pattern (Hegemann et al.
2015). Actinobacteria, and occasionally Proteobacteria, most commonly produce lasso peptides
(Arnison et al. 2013), although putative gene clusters have been identified in genomes from other
phyla (Hegemann et al. 2015). As of 2015, 38 lasso peptides had been discovered, largely due to
genome mining (Hegemann et al. 2015). A few of these lasso peptides have a narrow spectrum
of antimicrobial activity (Iwatsuki et al. 2006; Knappe et al. 2008; Salomón and Farías 1992).
Lasso peptides require only two posttranslational modifications, cleavage of the leader
peptide and formation of a disulphide bridge(s). The presence or absence of disulphide bridges in
the structure determine if the peptide belongs to class one, two, or three (Maksimov and Link
2014), where class I lasso peptides contain two disulphide bridges, class II contain none, and
class III contain just one. To the best of our knowledge, no lasso peptides have been identified in
Staphylococcus.
1.4.3.1 Genetic Organization
The recently adapted universal nomenclature for gene organization of lasso peptides
follows an “ABCD” structure (Arnison et al. 2013). Studies report that the ABC genes are
necessary for lasso peptide production (Maksimov and Link 2014), where A is the structural
gene, B encodes for the ATP dependent protease, and C encodes for the enzyme responsible for
isopeptide bond formation (Pan and Link 2011). Some clusters contain an ABC transporter,

15
encoded by the D gene, which transports the peptide out of the cell and is also responsible for
producer immunity (Pan and Link 2011). For clusters where the D gene is absent, the peptide is
still excreted from the cell and there is no host cell death, indicating the producer may use an
existing ABC transporter or that the peptide is able to diffuse out of the cell membrane (Knappe
et al. 2008). Newer clusters have been reported that contain highly conserved genes adjacent to
the ABC genes, such as isopeptidases (Hegemann et al. 2015), which may be indicative of the
evolutionary nature of the lasso peptide as clusters with these genes branch together in clades
(Maksimov and Link 2013).
1.4.3.2 Mode of Action
In general, lasso peptides are enzyme inhibitors or receptor antagonists, with a narrow
spectrum of activity against closely related bacteria (Arnison et al. 2013). The most well
characterized lasso peptide, microcin J25 produced by E. coli AY25, exerts its antimicrobial
effects by entering the target cell via the iron siderophore receptor (Destoumieux-Garzón et al.
2005) and inhibiting RNA polymerase (Delgado et al. 2001). Likewise, capistruin, produced by
Burkholderia thailandensis E264, was experimentally shown to have the same mode of action as
microcin J25 and the authors propose that all structurally similar lasso peptides potentially have
the same target (Kuznedelov et al. 2011). Lassomycin is a protease inhibitor that specifically
inhibits Mycobacterium tuberculosis (Gavrish et al. 2014).
1.4.4 Class IIa Bacteriocins
Class IIa bacteriocins refer to pediocin-like bacteriocins with a broad inhibitory spectrum
including potent anti-listerial activity (Kjos et al. 2011). These bacteriocins contain an N

16
terminal consensus sequence (YGNGVxCxxxxCxVxWxxA, where x is any amino acid) (Cotter,
Hill, and Ross 2005). These bacteriocins normally contain two distinct regions separated by a
flexible hinge (Kjos et al. 2011). Pediocin PA-1, produced by Pediococcus acidilactici UL5, is
the model bacteriocin for this group.
1.4.4.1 Genetic organization
The pediocin PA-1 gene cluster contains four genes, ABCD, where A is the structural
gene, B encodes the immunity protein, C is the ABC transporter and D encodes for an accessory
protein (Alvarez-Sieiro et al. 2016).
1.4.4.2 Mode of Action
Class IIa bacteriocins form pores in target cells, and unlike other class II bacteriocins,
these mechanisms have been elucidated. The target for these bacteriocins is the proteins of the
sugar transporter mannose phosphotransferase system (Man-PTS) (Kjos et al. 2011). However, it
is unknown if the bacteriocins use the Man-PTS the same way lanthipeptides use lipid II as a
docking molecule or if the bacteriocin interacts with the Man-PTS gate causing it to permanently
open, for which the latter model is more likely (Kjos et al. 2011)
1.4.5 Class IIb Bacteriocins
Class IIb bacteriocins require the presence of two distinct peptides that work
synergistically to provide maximum antimicrobial activity. Lactococcin G, produced by L. lactis
(Nissen-Meyer et al. 1992), was the first class IIb bacteriocin to be isolated and is subsequently
the best characterized at this point. As of 2015, this class contained 15 additional bacteriocins

17
(Kjos et al. 2014). All peptides in this class require two different peptides (located on the same
operon) each produced in equal amounts to obtain peak antimicrobial activity (Kjos et al. 2014).
1.4.5.1 Genetic organization
These bacteriocins require at least five different genes, on one or two operons (Alvarez-
Sieiro et al. 2016). Generally, there are two structural genes, an ABC transporter, an immunity
protein gene and a gene encoding an accessory protein.
1.4.5.2 Mode of Action
Lactococcin G, produced by L. lactis, was initially reported to interact with a receptor on
the cell membrane of the target bacteria to induce cell leakage (Rogne et al. 2008). Recently, it
was determined the target is likely bacA, a membrane protein involved in peptidoglycan
synthesis (Kjos et al. 2014), which was the first time a target has been identified for class IIb
bacteriocins.
1.4.6 Class IIc Bacteriocins
Class IIc bacteriocins are circular bacteriocins, characterized by an amide bond between
the N and C termini (Maqueda et al. 2008) and as of 2011 have only been identified in Gram-
positive bacteria (van Belkum, Martin-Visscher, and Vederas 2011). Their head to tail
cyclization attribute to their reported resistance to proteases and pH and heat treatment (van
Belkum, Martin-Visscher, and Vederas 2011). The first and most well characterized circular
bacteriocin is Enterocin AS-48, isolated from Enterococcus (Martínez-Bueno et al. 1994; Samyn
et al. 1994). At least nine bacteriocins from this group have been isolated, purified, and

18
characterized (Arnison et al. 2013). The first report of a circular bacteriocin in Staphylococcus is
aureocyclicin 4185, produced by S. aureus (Potter, Ceotto, Coelho, Guimaraes, et al. 2014).
1.4.6.1 Genetic organization
The genetic organization for many circular bacteriocins has been well-described (van
Belkum, Martin-Visscher, and Vederas 2011), although a universal nomenclature has not been
adopted for this group yet, except for the use of A for the precursor peptide and the
recommendation to use B for the putative membrane protein (Arnison et al. 2013). The enterocin
AS-48 gene cluster contains ten genes, termed A, B, C, C1, D, D1, E, F, G, and H, which function
as production, modification, transport, and immunity genes (Maqueda et al. 2008).
1.4.6.2 Mode of Action
In general, circular bacteriocins exert their antimicrobial activity by targeting cell
membranes and forming pores (Arnison et al. 2013). Enterocin AS-48 interacts with the cell
membrane, inserting itself in a voltage-independent manner causing loss of membrane potential
and cell death (Maqueda et al. 2008). Carnocyclin A, on the other hand, is able to form pores in a
voltage-dependent manner (Gong et al. 2009).
1.4.7 Class IId Bacteriocins
Class IId bacteriocins are a heterogenous group of bacteriocins that are single linear
peptides. The most well characterized bacteriocin from this group is lactococcin 972, produced
by L. lactis subsp. lactis IPLA 972 (Martı́nez et al. 1999).

19
1.4.7.1 Genetic Organization
Along with the 91aa structural gene, LclA, the complete lactococcin 972 cluster contains
a transporter, LclB, and an immunity protein (Martı́nez et al. 1999). The genetic structure of
lactococcin A is similar, with a structural gene, an immunity gene, an ABC transporter, and an
accessory gene (Stoddard et al. 1992).
1.4.7.2 Mode of Action
Latococcin 972 blocks the incorporation of lipid II of closely related bacteria.
Lactococcin A, like Class IIa bacteriocins, targets the Man-PTS proteins, although unlike the
Class IIa bacteriocins, it is very specific and targets only the lactococcal Man-PTS system
(Alvarez-Sieiro et al. 2016).
1.5 Immunity genes and cross immunity
Bacteriocin producers encode for specific immunity proteins to provide protection from
the lethal activities of their products. The peptide structural and modification/transport genes are
present on the same operon as the immunity genes, meaning that the producing strain is sensitive
to the bacteriocin product when in a non-producing state (Eijsink 1998).
Lanthipeptides have the most complex immunity proteins, termed LanI and LanFEG.
These two proteins are thought to function independently and have different mechanisms of
protection (Stein et al. 2005). LanFEG have been experimentally determined to be involved in
exporting bacteriocin out of the cell and fall into the ABC-2 subfamily of drug resistance
exporters (Stein et al. 2005). On the other hand, LanI appears to sequester lanthipeptides on the

20
surface, thus preventing pore formation. One interesting note from a study done examining the
lanthipeptide subtilin and its immunity proteins was that SpaI interacts specifically with subtilin,
and not with the structurally similar nisin (Stein et al. 2005). Similarly, NisI only confers
protection against nisin, and not against subtilin (Stein et al. 2003). This indicates that cross
immunity due to bacteriocin cluster immunity proteins can be rare, even between closely related
lanthipeptides. For certain bacteriocins, like nisin, lanI and lanFEG are both needed for optimal
immunity, however certain bacteriocin clusters only contain one self protection mechanism, for
example the pep5 cluster contains only lanI whereas the epidermin cluster only contains the
lanFEG transporter (Stein et al. 2003).
There are additional genes related to immunity that have been identified, although not
well described. Abi proteins, or CAAX immunity proteins, are putative membrane bound
metalloproteases and have been shown to be involved with self-immunity for Class IIb
bacteriocins (Kjos et al. 2011). They can show extensive cross immunity, which could mean that
the immunity proteins give immunity by a common, shared mechanism (Kjos et al. 2010). These
mechanisms could be degradation of the bacteriocin, or by modifying the bacteriocins receptor
(Kjos et al. 2010). Class IIa bacteriocins, as well as lactococcin A and B, which target the Man-
PTS system studies have shown that their immunity proteins bind and lock the bacteriocin onto
the receptor target to prevent pore formation (Diep et al. 2007).
1.6 Bacteriocin discovery and purification
Traditionally, discovery of bacteriocins starts with a large screen of bacterial isolates to
assess their inhibitory capability, and thus potential bacteriocin production, against indicator

21
bacteria in vitro. A commonly used method of detection is the ‘spot on lawn’ assay (Fleming,
Etchells, and Costilow 1975), where drops of the producer broth are spotted onto agar and
incubated overnight to allow colonies to develop. The top of the agar is overlayed with soft agar
(0.5% agar) inoculated with the indictor species, and incubated overnight, followed by
examination and measuring of the zones of inhibition around the producer colonies (Fleming,
Etchells, and Costilow 1975). De Vliegher et al. (2004) utilized a cross-streaking method, where
the potential producer was inoculated as a center streak down a blood agar plate and incubated
overnight. The indicator species was spread over the entirety of the agar after flipping the agar
over, with the producer center streak ending up on the bottom of the plate. The zones of
inhibition were measured after incubation for 24hr, perpendicular to the center streak (De
Vliegher et al. 2004). Well diffusion assays are also used to assess inhibition (Schillinger and
Lücke 1989). For this assay, agar plates are overlayed with soft agar inoculated with the
indicator species, followed by the drilling of wells into the agar once set. Cell free supernatant of
the potential producer is added to each well, and following overnight incubation, zones of
inhibition are assessed (Schillinger and Lücke 1989). Identified potential producers are then
grown up in conditions conducive for bacteriocin production. Staphylococcus aureus isolates
were shown to have peak bacteriocin production during the late-log or early stationary growth
phase of cultures grown in brain heart infusion (BHI) medium at 37°C (Nascimento et al. 2004).
Bacteriocin production during the late-log or early stationary phase can be 4.6 to 7.5 fold higher
than production in the early exponential growth phase (Sedgley, Clewell, and Flannagan 2009).
The bacteriocin is extracellularly secreted into the medium during growth, normally in small
quantities, so a large amount of medium is recommended to be able to isolate enough quantity of
bacteriocin. Optimal isolation and purification protocols depend on the bacteriocin, as they have

22
different properties that can be taken advantage of for separation from the culture medium.
Therefore, there is not one technique that is suitable for all classes of bacteriocin (Kaškonienė et
al. 2017). Common isolation techniques take advantage of their charge and hydrophobic natures
(Parada et al. 2007). Ammonium sulphate precipitation is the most commonly used method to
reduce the volume and concentrate the bacteriocin from the producer medium (Kaškonienė et al.
2017), although chloroform extractions have proven to be less expensive, less time consuming,
and result in a higher bacteriocin yield (Burianek and Yousef 2000). Upon obtaining an active
partially purified bacteriocin (at this point there are still proteins and peptides from the growth
medium present), Reverse-Phase High Pressure Liquid Chromatography (RP-HPLC) is most
commonly performed to purify the bacteriocin (Kaškonienė et al. 2017). Other techniques, such
as ultrafiltration or filter assisted size exclusion protein fractionation, can be used for purification
(Kaškonienė et al. 2017). Upon obtaining a purified bacteriocin, SDS-PAGE can be used to
determine an approximate molar mass, and the bacteriocin can be sent to sequencing to obtain
the amino acid sequence, or it can be detected using matrix-assisted laser desorption ionisation
time-of-flight mass spectrometry (MALDI-TOF MS) (Zhu et al. 2016). This lab-based
bacteriocin discovery approach is time consuming, as well as labour intensive, requiring
qualified technicians to use the equipment needed (Kaškonienė et al. 2017). There also is the risk
of “re-discovery” of a bacteriocin, as was the case with a bacteriocin discovered in an S.
simulans isolate, initially named simulancin 3299, although after purification and identification it
was discovered to be identical to known bacteriocin nukacin ISK-1 (Ceotto et al. 2010). There is
also a risk of missing identification of some bacteriocins that are not expressed under normal
laboratory conditions.

23
1.7 In silico screening
In silico screening, or genome mining, is an approach that is being used more frequently
for the discovery of new bacteriocins. This methodology is able to examine whole genome
sequences of bacteria and identify bacteriocin biosynthetic gene clusters, independent of
laboratory phenotype analysis.
One of the initial genome screenings for bacteriocins was done using the lanM protein
(from type B lanthipeptides) as the driver sequence. Because the associated genes are well
conserved in specific classes of bacteriocins, this approach could yield identification of novel
type B lanthipeptides. The study resulted in 89 LanM homologues identified, 61 of which were
in bacteria not associated with lanthipeptide production. For identified genomes, with both a
LanM homologue and available whole genomes, BAGEL was used to further analyze the
genome for potential bacteriocin clusters. One bacteriocin containing isolate that was able to
inhibit pathogens was selected for further laboratory analysis and bacteriocin characterization as
a proof of concept. Consequently, lichenicidin was isolated and characterized, providing
testimony that in silico screening is a valuable tool that can result in the identification of novel
bacteriocins (Begley et al. 2009).
LanT, a lanthipeptide associated transporter, was used as the driver sequence to identify
novel lanthipeptides in available sequences on NCBI, taking the top 72 hits for further analysis
(Singh and Sareen 2014). This approach led to the identification of 54 strains containing LanT
homologues, strains that were not previously associated with lanthipeptide production. Overall,
the study identified 8 novel two-component lanthipeptides for further characterization (Singh and
Sareen 2014). Likewise, the radical SAM enzyme present in sactipeptide clusters was used as the

24
driver sequence in genome mining for novel sactipeptides, yielding putative sactipeptides in
phyla not typically associated with bacteriocin production (Murphy et al. 2011). Using a similar
associated gene homology approach with lasso peptides, Burkholderia thailandensis E264 was
identified to contain putative lasso peptide associated genes. Capistriun, a novel lasso peptide
was subsequently isolated (Knappe et al. 2008).
A unique precursor-centric genome mining approach was created to search for small
areas of conserved regions in the structural gene of lasso peptides (Maksimov, Pelczer, and Link
2012). Using this approach, out of 3000 prokaryotic genomes mined, 78 were identified to be
putative producers. To validate this approach, one putative producer was selected and the lasso
peptide was expressed in E. coli, leading to the production of a novel lasso peptide, astexin-1
(Maksimov, Pelczer, and Link 2012). In total, genome mining has resulted in the considerable
increase in identified members of the lasso peptide family, inflating to 38 members as of 2015
(Hegemann et al. 2015).
Currently, approaches have been automated that combine direct mining for the structural
gene along with indirect mining for associated genes in order to comprehensively search the
genomes. BAGEL3 is one such available software, which mines for bacteriocins in single or
multiple DNA sequences such as (un)finished genomes, scaffold files but also meta-genomics
data (van Heel et al. 2013). BAGEL3 is also a source for databases of structural genes in each
class of bacteriocin, although these databases were last updated in 2013. antiSMASH (antibiotics
and Secondary Metabolite Analysis SHell) is another tool for genome mining that identifies all
secondary metabolite biosynthetic gene clusters, not just bacteriocins (Weber et al. 2015).
Novel bacteriocins were recently identified in anaerobic bacteria using a combination of
antiSMASH, and BAGEL and bactibase databases (Letzel, Pidot, and Hertweck 2014). Out of

25
221 anaerobe genomes from 18 different phyla, they identified 25% of genomes (from 8
different phyla) were able to encode for bacteriocins. This study determined 43 out of 81
identified clusters were novel and described 23 clusters that had not been identified in anaerobes
before, although had similarities to previously identified bacteriocins from other phyla (Letzel,
Pidot, and Hertweck 2014). In another study, 34 genomes from 34 different species of LAB that
have not been identified as bacteriocin producers were mined through BAGEL3, resulting in the
identification of 20 of the species containing bacteriocin gene clusters (Singh et al. 2015).
Azevedo et al. (2015) screened 224 ruminal bacteria strains and 5 ruminal archaea to determine
the distribution and diversity of ruminal bacteria. This study identified 46 bacteriocin gene
clusters in 33 strains of bacteria. Whole and partial genome sequences were uploaded into
Bagel3 and antiSMASH software for the detection of bacteriocin gene clusters. Before that study
only 9 bacteriocins had been fully or partially characterized from ruminal bacteria (Azevedo et
al. 2015). In another study of substantial size, 382 genome isolates from the gastrointestinal tract,
available as a subset of the Human Microbiome Project, were mined for bacteriocin clusters
using BAGEL3 (Walsh et al. 2015). In total, 74 clusters from 59 isolates were detected, and the
majority of the species containing isolates were from species not previously associated with
bacteriocin production (Walsh et al. 2015). It is apparent that genome mining is an incredibly
useful tool to identify bacteriocin gene clusters.
Although, caution has to be taken to draw conclusions from genome mining, as precursor
peptides may be modified in different ways than anticipated from examining the modification
genes and may result in a mature peptide belonging to a different class than once thought
(Arnison et al. 2013). Additionally, proximity on a contig may not mean there is a
target/substrate relationship. Additionally, in silico screening may not ultimately result in

26
identification and purification of bacteriocin, though there have been successes (Begley et al.
2009; Dischinger et al. 2009; Knappe et al. 2008). There is also the potential to miss completely
novel clusters, as neither the precursor genes nor associated genes are known or lack sufficient
homology to be identified.
It is nevertheless a good starting point to identify isolates for future characterization in
the laboratory and to identify the distribution of bacteriocin associated gene clusters in large
groups of related bacteria, or bacteria from unique environmental niches. In general, due to
extensive genome mining projects, bacteriocins are now known to be more prevalent and present
in more phyla of bacteria than what was once thought (Arnison et al. 2013).
1.8 Applications of bacteriocins
Nisin is currently the most widely studied and used bacteriocin. In 1969, the Joint Food
and Agriculture Organization/World Health Organization approved nisin for use as a food
additive (Shin et al. 2016). In 1988, nisin was given a generally regarded as safe (GRAS) status
for use in cheeses by the Food and Drug Administration in the United States (Cotter, Hill, and
Ross 2005). Presently, it is licensed in over 50 countries as a food preservative (Shin et al. 2016).
The two main identified areas of research for bacteriocin utilization are in the food preservation
industry and in the medical and veterinary fields (Pieterse and Todorov 2010). This thesis will
focus on the potential applications of bacteriocins in the medical and veterinary fields.
Bacteriocins have been identified as attractive alternatives to antibiotics (Cotter, Ross,
and Hill 2013). Although it is generally accepted that bacteriocins produced by Gram-positive
bacteria possess less potential to be used in a clinical setting for treatment of Gram-negative

27
pathogens because these bacteriocins normally only have activity against other Gram-positive
pathogens. However, nisin and epidermin have both demonstrated activity against Gram-
negative pathogens in vitro (Kuwano et al. 2005; Lacroix et al. 2001). Subtilosin A also has
antimicrobial effects against both Gram-positive and Gram-negative pathogens (Shelburne et al.
2007), so there is potential for other bacteriocins to be identified to be useful against Gram-
negative pathogens. Additionally, applying heat stress increased the effectiveness of subtilosin A
against Gram-negative bacteria (Shelburne et al. 2007), which was similar to what was reported
for nisin and Gram-negative bacteria (Boziaris and Adams 2001).
The prospective applications in the human medical field range from topical treatments of
skin infections to treatments of ulcers (Pieterse and Todorov 2010). One reason that bacteriocins
could be so useful is that medicine could exploit their narrow spectrum of activity. While there
are many bacteriocins that have broad spectrums of activity, which is appealing while dealing
with an infection of unknown etiology, there are also many bacteriocins with a narrow spectrum
of activity. These bacteriocins are of value because they can target a specific pathogen while
leaving the commensal microbiota untouched. An example of this is Thuricin CD, a sactipeptide
produced by Bacillus thuringiensis DPC 6431, which has promising results as a therapeutic
against C. difficile infections, while showing no significant adverse effects on the normal colon
microbiota (Rea et al. 2010). Another promising study showed that nisin exerted anti-biofilm
effects against saliva derived biofilms without causing cytotoxic effects to the human oral cells
(Shin et al. 2015). Yet another benefit to using narrow spectrum bacteriocins in place of
antibiotics is that antibiotics could be used less frequently, thus reducing the selection pressure
for resistance, and therefore maintaining the usefulness of that antibiotic for future need (Riley
and Wertz 2002).

28
Nevertheless, there are some complications that must be addressed to put these
bacteriocins to clinical use. Bacteria that are able to encapsulate themselves, such as Klebsiella
species, are resistant to subtilosin A by way of preventing the bacteriocin by getting to the cell
wall surface, either by exclusion or by binding on the capsule surface (Shelburne et al. 2007).
With nisin for example, it has low solubility and low activity at high pH, and it has a tendency to
interact with blood components (Dischinger, Basi Chipalu, and Bierbaum 2014). Additionally,
because lanthipeptides lack stability against intestinal enzymes, optimal methods of delivery
would have to be investigated. Studies have suggested the use of a pill (Arthur, Cavera, and
Chikindas 2014) to encapsulate and protect the bacteriocin from the proteolytic enzymes in the
stomach. Another proposed mechanism to avoid degradation by digestive enzymes is to colonize
the gastrointestinal tract with a strain that produce the bacteriocin. In a study assessing L.
monocytogenes inhibition by a bacteriocin produced by Lactobacillus salivarius UCC118, mice
were fed either a control, a bacteriocin producing strain, or a bacteriocin knockout strain then
orally infected with luciferase-tagged Listeria (Corr et al. 2007). Thirty minutes post-challenge,
there was no fluorescence in the bacteriocin-positive infected mice, indicating anti-listerial
activity from the probiotic strain (Corr et al. 2007).
In terms of applications for bacteriocins in the dairy industry, bacteriocins have been
studied for use in preventing mastitis, although to date, only nisin and lacticin 3147 have been
explored extensively for use. Nisin was first observed to considerably reduce the amount of S.
aureus and E. coli recovered from the bovine teat skin after 1 minute of exposure to the nisin
preparation when compared with conventional iodine and chlorohexidine treatments (Sears et al.
1992). Additionally, the product posed little risk of skin irritation (Sears et al. 1992). Immucell
Corporation developed two nisin based products, Wipe-Out® and Mast Out® for use in the

29
industry, although they do not appear to be available anymore (Pieterse and Todorov 2010).
Though, according to Immucell Corp’s website, they are in the construction stage of a new
pharmaceutical facility for the production of nisin for use in a product to treat SCM
(http://immucell.com/products/purified_nisin/). Lacticin 3147 has been investigated to replace
antibiotics in dry cow therapy by use in teat sealants. Initial studies indicated lacticin 3147
inhibited mastitis pathogen S. dysgalatiae in vitro and resulted in no irritation upon inoculation
into the teat (Ryan et al. 1998). In an experimental trial on 68 uninfected quarters lacticin 3147
was tested on its ability to inhibit a challenge of S. dyscalactiae (issued 3 days post bacteriocin
infusion) for 8 days (Ryan et al. 1999). The results indicated that only 6% of the quarters
inoculated with both teat sealant and lacticin 3147 contracted clinical mastitis or shed S.
dysgalactiae in the 8 days post challenge, compared to 66% of the control quarters (Ryan et al.
1999). In a subsequent study, the usefulness of lacticin 3147 against S. aureus was assessed
(Twomey et al. 2000). In this study, one quarter of each lactating cow was infused with lacticin
3147 and teat sealant, where the other two quarters were left as untreated controls. Two hours
after inoculation, the quarters were challenged with S. aureus and 18 h later the recovery count
of S. aureus was assessed. The quarters with lacticin 3147 and teat sealant showed less S. aureus
recovery (Twomey et al. 2000). Lacticin 3147 was also assessed for mastitis prevention in the
lactating herd in the use of a teat dip (Klostermann et al. 2010). Here, teats were coated with S.
aureus, S. dysgalactiae, or S. uberis and then dipped with a teat dip containing lacticin 3147. The
dip was able to eliminate 80% of the S. aureus, 97% of S. dysgalactiae and 90% of S. uberis
after a 10-minute contact time (Klostermann et al. 2010).
Many other bacteriocins have demonstrated activity against mastitis causing pathogens.
Five characterized bacteriocins from B. thuringiensis were tested for inhibition against 50 bovine

30
subclinical mastitis S. aureus isolates, and all of the S. aureus isolates were sensitive to at least
one of the bacteriocins (Barboza-Corona et al. 2009). More recently, a study reported that yet
unidentified bacteriocins from B. thuringiensis were able to inhibit 60% of pathogens isolated
from dairy goat mastitis cases in vitro, many of which were multi-drug resistant isolates
(Gutiérrez-Chávez et al. 2016). Nukacin 3299, isolated from S. simulans, showed inhibitory
activity against Staphylococcus strains from mastitis origins (Ceotto et al. 2010). Staphylococcus
aureus (165 strains) and S. agalactiae (74 strains) from bovine mastitis were assessed for
susceptibility to three bacteriocins produced by S. aureus and four bacteriocins produced by S.
epidermidis. Epidermin (an S. epidermidis bacteriocin) showed the most widespread inhibition of
mastitis pathogens (>85%), followed by using a combination of S. aureus bacteriocins (Coelho et
al. 2007).
The delivery of these inhibiting bacteriocins will need to be studied more, although it
appears that teat dips, wipes, and teat sealants containing the bacteriocin are the most promising.
The benefits of combining bacteriocins with teat sealants mean that the bacteriocin is localized in
the teat, allowing for antimicrobial activity against pathogens that manage to enter the teat canal
after dry off. These potential products could allow dairy farmers to reduce their antibiotic use by
implementing selective dry cow therapy, only treating cows with infected quarters with
antibiotics and teat sealants at dry off, and treating uninfected cows with the teat sealant plus
bacteriocin product. Studies longer than 8 days will need to be done in order to ensure the cow is
provided with protection for the entire duration of the dry period. Teat wipes and dips could
provide effective control against Gram-positive mastitis in the lactating herd, potentially limiting
antibiotic use by decreasing the cases of CM. Thus, there is great potential for bacteriocins to be

31
used for treatment and control of mastitis. Future studies need to be done on the best ways to use
these bacteriocins, as well as on the best delivery system.
1.9 Bacteriocin resistance
An important consideration when moving forward with investigating bacteriocins for
potential use in health and food industries is to assess the risk of target bacteria to develop
resistance (Cotter, Ross, and Hill 2013). As with antibiotic resistance, the specific mechanisms
of resistance can be classified into two groups, acquired resistance (developed by a formerly
susceptible strain) and innate resistance (intrinsic). Only a few bacteriocins have been studied
with respect to the development of resistance: nisin, lacticin 3147, pediocin-like bacteriocins,
and lysostaphin (Bastos, Coelho, and Santos 2015). Little resistance to nisin has been reported
among food spoilage bacteria in the field; therefore, the evidence of bacteriocin resistance is
limited to laboratory experiments and lanthipeptide resistance has indeed been induced in
laboratory settings (Draper et al. 2015). The mechanisms which target bacteria confer resistance
to lanthipeptides are often innate and provide protection against cationic antimicrobial peptides
in general, not specifically lanthipeptides (Draper et al. 2015).
One very specific mechanism of innate resistance is through immune mimicry, which
occurs when non-lanthipeptide producing strains are immune to lanthipeptides as a result of
possessing immunity gene functional homologues (Draper et al. 2009). The expression of these
orphan immunity clusters confers protection against the associated bacteriocin (Draper et al.
2015). Additionally, some bacteria may encode for enzymes, which specifically degrade
bacteriocins. Nisinase, which degrades nisin, is an example and confers protection against nisin

32
and subtilin (Bastos, Coelho, and Santos 2015). Nisin resistance protein NSR protease is another
anti-nisin enzyme that has been identified in non-producer L. lactis strains (Sun et al. 2009).
Transcriptome analysis revealed 92 genes directly or indirectly involved in the
acquisition of nisin resistance in L. lactis (Kramer et al. 2006), indicating that resistance can
potentially be incredibly complex. The authors suggest four ways that L. lactis was able to
decrease susceptibility to nisin. The first and main mechanism is the prevention of nisin from
interacting with the cell membrane, and therefore lipid II. It likely does this by increasing the
thickness of the cell wall, becoming more densely packed, and by becoming less negatively
charged, effectively repelling nisin. Secondly, expression of genes changes in order to change the
local pH at the outside of the cytoplasmic membrane, which may either sequester the nisin or
promote its degradation. Thirdly, the phospholipids can become less saturated and more loosely
packed by changing expression of genes that are involved in elongation and saturation of said
phospholipids. This may hamper nisins ability to insert itself and form pores. Lastly, ABC
transporters may be able to remove nisin from the cytoplasmic membrane, keeping it from
binding to lipid II (Kramer et al. 2006). It is notable that the structure of lipid II does not change
as a mechanism of resistance, as where nisin binds is synthesized early and necessary for
function (Kramer et al. 2006).
In terms of acquired resistance, the frequency of spontaneous mutations that result in
nisin resistance can vary from <10-9 to 10-2 in L. monocytogenes and from 10-8 to 10-2 in other
Gram-positive organisms (Bastos, Coelho, and Santos 2015). Nisin resistance seems to have
little effect on bacterial fitness, and the resistance phenotype appears to be stable in the
population (Bastos, Coelho, and Santos 2015). For lacticin 3147, the frequency of resistance in
L. lactis appears to be low, with resistant isolates only able to resist low levels of the bacteriocin

33
after one round of pressure. Although, higher levels of resistance to lacticin 3147 can be selected
for and the resistance phenotype also appears to be stable (Bastos, Coelho, and Santos 2015).
Moderate to high levels of resistance have been reported in L. monocytogenes and E.
faecalis for Class IIa bacteriocins, where the resistant strains can decrease expression of the
Man-PTS system, which is the bacteriocins target (Guinane et al. 2006). Additionally, sensitivity
of cells to Class IIa bacteriocin depends on the sequence of the Man-PTS proteins, as only a
specific clade in the Man-PTS proteins’ phylogeny is susceptible (Kjos, Nes, and Diep 2009).
The authors’ approach of examining receptor sequences (in this case the Man-PTS protein) in
relation to susceptibility can potentially be used for all bacteriocins once a target receptor is
determined in order to quantify the inhibitory ability of the bacteriocin on a large group of target
organisms.
Because of the resistance observed in in vitro studies, it is not unlikely that bacteriocin
resistance will arise with the application of bacteriocins in clinical settings (Bastos, Coelho, and
Santos 2015). Although, a study examining the risk of bacteriocin E-760 resistance developing in
Campylobacter in a chicken model only found low levels of resistance (Hoang, Stern, and Lin
2011). Resistance risk may depend on the bacteriocins mode of action; specifically, bacteriocins
with dual modes of action may be less likely to induce resistance. Therefore, the lack of apparent
resistance towards nisin may be due to its multiple modes of action with a number of distinct
targets (Shin et al. 2015). More studies need to be done on the specific risk of resistance
developing in food model systems. Once resistance mechanisms are determined, ways can be
created to mitigate the resistance in clinical applications. For example, An ABC pump confers
resistance of Campylobacter to E-760, therefore it is postulated that use of the bacteriocin with
efflux pump inhibitors may increase the susceptibility of Campylobacter to E-760 for use in

34
production systems (Hoang, Stern, and Lin 2011). More studies need to be done on the risk of
resistance development in clinical settings for Staphylococci bacteriocins. To do this,
information needs to be obtained on specific bacteriocins, their structure, modes of action, and
spectrums of activity. Identification of bacteriocins is the first step in this process. With the NAS
being such ubiquitous members of the udder microbiome, paired with evidence of NAS
inhibition on major mastitis pathogens due to bacteriocin production, large-scale investigations
of bacteriocin production in NAS has merit.
1.10 Thesis outline
The overall aim of this thesis research was to assess the species-specific inhibitory
capability of bovine NAS, to identify bacteriocin genes potentially relating to inhibition, and in
an attempt to understand the mechanisms of susceptibility, identify genes related to susceptibility
in a large group of bovine S. aureus to an S. chromogenes bacteriocin. The study consisted of
two parts with distinct (albeit complementary) aims and hypotheses.
1.10.1 Bacteriocins of non-aureus staphylococci isolated from bovine milk
Aim: To determine the inhibitory capability of 441 bovine NAS isolates from 26 different
species against a bovine S. aureus and a human methicillin-resistant S. aureus (MRSA). The
second aim was to identify and describe the organization of bacteriocin biosynthetic gene
clusters in the corresponding 441 whole genome sequences.
Hypothesis: Phenotypic inhibition will be widespread across isolates and genotype will be
able to predict phenotypic inhibition in most cases.

35
The research to address this aim and hypothesis is described in Chapter 2.
1.10.2 Identifying putative bacteriocin resistance genes in Staphylococcus aureus whole genomes
Aim: To determine the sensitivity of 139 bovine S. aureus isolates to a bacteriocin
producing S. chromogenes isolate and to identify and describe putative bacteriocin resistance
genes in S. aureus whole genome sequences.
Hypothesis: The presence of specific bacteriocin resistance genes in S. aureus isolates will
correspond to the in vitro phenotype.
The research to address this aim and hypothesis is described in Chapter 3.

36
Chapter Two: Bacteriocins of non-aureus staphylococci isolated from bovine milk
2.1 Abstract
Non-aureus staphylococci (NAS), the bacteria most commonly isolated from the bovine
udder, potentially protect the udder against infection by major mastitis pathogens due to
bacteriocin production. In this study, we determined the inhibitory capability of 441 bovine NAS
isolates (comprising 25 species) against bovine S. aureus. Furthermore, inhibiting isolates were
tested against a human methicillin-resistant S. aureus (MRSA) isolate using a cross-streaking
method. We determined the presence of bacteriocin clusters in NAS whole genomes using
genome mining tools, BLAST, and comparison of genomes of closely related inhibiting and non-
inhibiting isolates and determined the genetic organization of any identified bacteriocin
biosynthetic gene clusters. Forty isolates from 9 species (S. capitis, S. chromogenes, S.
epidermidis, S. pasteuri, S. saprophyticus, S. sciuri, S. simulans, S. warneri, and S. xylosus)
inhibited growth of S. aureus in vitro; 23 of which, from S. capitis, S. chromogenes, S.
epidermidis, S. pasteuri, S. simulans, and S. xylosus, also inhibited MRSA. 105 putative
bacteriocin gene clusters in 95 whole genomes from 16 species were identified. Twenty-five
novel bacteriocin precursors were described. Additionally, 7 NAS species were identified with
clusters for the first time and sactipeptides and lasso peptides, which have never been reported in
Staphylococcus species, were identified. In conclusion, NAS from bovine mammary glands are a
source of potential bacteriocins, with >21% being possible producers, representing potential for
future characterization and clinical applications.

37
Importance
Mastitis (particularly infections caused by Staphylococcus aureus) cost Canadian dairy
producers $400 million/year and is the leading cause of antibiotic use on dairy farms. With
increasing antibiotic resistance and regulations regarding use, there is impetus to explore
bacteriocins (bacterially produced antimicrobial peptides) for treatment and prevention of
bacterial infections. We examined the ability of 441 non-aureus staphylococci (NAS) bacteria
from Canadian bovine milk samples to inhibit growth of S. aureus in the laboratory. Overall, 9%
inhibited growth of S. aureus and 58% of those also inhibited methicillin-resistant S. aureus
(MRSA). In NAS whole-genome sequences, we identified 21% of NAS as having bacteriocin
genes. Our study provides the first comprehensive genomic report of NAS bacteriocins from
bovine milk and represents the foundation to further explore these antimicrobial peptides for
clinical use.

38
2.2 Introduction
Non-aureus staphylococci (NAS), a heterogeneous group of approximately 50 species that
can be considered teat skin opportunists and minor pathogens, are the bacteria most commonly
isolated from the bovine udder (Piepers et al. 2007; Pitkälä et al. 2004; Sampimon et al. 2009;
White et al. 1989). Several studies reported that NAS may confer protection against
intramammary infection (IMI) by major mastitis pathogens (De Vliegher et al. 2004; Matthews,
Harmon, and Smith 1990). In a challenge study, 53% of Staphylococcus chromogenes-infected
quarters were protected from S. aureus challenge (Matthews, Harmon, and Smith 1990). In
another study, S. chromogenes isolates inhibited in vitro growth of all tested Staphylococcus
aureus, Streptococcus dysgalactiae, and Streptococcus uberis isolates, but none of the Gram-
negative isolates (De Vliegher et al. 2004). Similarly, NAS strains from Brazilian bovine mastitis
cases, including Staphylococcus epidermidis, Staphylococcus simulans, Staphylococcus
saprophyticus , Staphylococcus hominis , and Staphylococcus arlettae, inhibited growth of
Corynebacterium fimi (Nascimento et al. 2005). The isolated antimicrobial substances were
considered to be bacteriocins due to sensitivity to proteolytic enzymes (Nascimento et al. 2005).
Recently, Braem et al. (2014) identified NAS strains (from 6 species) that inhibited S. aureus, S.
uberis, and S. dysgalactiae and an inhibitory substance (from an inhibiting S. chromogenes) was
isolated and identified to be a nukacin-like bacteriocin.
Bacteriocins are ribosomally synthesized and post-translationally modified peptides
(RiPPs) produced by Gram-positive bacteria that mainly inhibit growth of similar bacterial
species, or occasionally a broad spectrum of bacteria (Cotter, Hill, and Ross 2005). As
regulations surrounding antibiotic usage get stricter, bacteriocins represent potential alternatives

39
to antibiotics (Cotter, Ross, and Hill 2013). Nisin, produced by Lactococcus lactis, was available
for use in the dairy industry in the form of Wipe Out®, with germicidal activity against S. aureus
and S. agalactiae (Sears et al. 1992). Lacticin 3147, another bacteriocin produced by L. lactis,
was tested for use in a teat sealant and reduced the number of S. aureus recovered from the
bovine mammary gland 18 h post challenge when compared to infusion with teat sealant alone
(Crispie et al. 2005).
Traditionally, identification of novel bacteriocins used culture-based approaches that
involved screening numerous isolates for antimicrobial activity, followed by lengthy biochemical
characterization. However, due to growing accessibility of genome sequence data, in silico
screening (genome mining) is a promising approach to identify novel biosynthetic gene clusters.
Although bacteriocin precursor genes are often small and lack homology, making in silico
identification challenging, bacteriocin-associated genes present on the same operon are highly
conserved. Therefore, the associated modification genes are used in classification schemes, with
2 classes: Class 1, post-translationally modified bacteriocins, including lanthipeptides,
sactipeptides, and lasso peptides; and Class 2, non-modified or cyclic peptides (Cotter, Hill, and
Ross 2005). By screening genomes for bacteriocin-associated genes, new lanthipeptides (Marsh
et al. 2010; Begley et al. 2009) and new Class IIa bacteriocin gene clusters (Kjos et al. 2011)
have been identified. In addition, BLAST-based approaches have been used to identify
bacteriocins in cyanobacteria (Wang, Fewer, and Sivonen 2011) and to identify lanthipeptide
clusters by using a transport gene for screening (Singh and Sareen 2014). Current software-based
approaches (e.g. BAGEL3 and antiSMASH) combine direct mining for the structural gene with
indirect mining for bacteriocin-associated genes. Using this approach, novel bacteriocins were
identified in ruminal bacteria (Azevedo et al. 2015), anaerobic bacteria (Letzel, Pidot, and

40
Hertweck 2014), lactic acid bacteria (Singh et al. 2015), and human gut microbiota (Walsh et al.
2015). A large in silico screen has apparently not been done on NAS whole genomes, which
could be the foundation for future investigations into alternatives for antimicrobials in the dairy
industry.
The first objective was to determine the inhibitory capability of 441 bovine NAS isolates
from 25 species against a bovine S. aureus and a human methicillin-resistant S. aureus (MRSA).
The second objective was to identify and describe the organization of bacteriocin biosynthetic
gene clusters in the corresponding 441 whole genome sequences.
2.3 Materials and methods
2.3.1 Isolates
NAS isolates and a S. aureus isolate from a clinical case of bovine mastitis were collected
in the National Cohort of Dairy Farms (NCDF) conducted across Canada during 2007 and 2008,
as described by Reyher et al. (2011). The Canadian Bovine Mastitis and Milk Quality Research
Network (CBMQRN) at the University of Montreal stored the samples before sending them to
the University of Calgary. Overall, 441 NAS isolates were selected from the stock of 5507
isolates over the 25 NAS species identified previously (Naushad et al. 2016; Condas et al. 2017
). These isolates originated from 87 herds across Canada from Nova Scotia, Prince Edward
Island, New Brunswick (representing Atlantic Canada), Québec and Ontario (representing
Central Canada), and Alberta (representing Western Canada) (Reyher et al. 2011). Isolates
included 68 NAS isolates from clinical mastitis cases, 26 multi-drug resistant (MDR) isolates,
maximum 1 isolate per cow of any uncommon species (defined as <20 unique isolates at cow

41
level), and maximum 1 randomly selected isolate per cow for all other species until 441 were
chosen (Naushad et al. 2016). The multi-drug resistant MRSA clinical strain H176 from human
origin was obtained from Dr. K. Zhang’s laboratory at the University of Calgary.
2.3.2 Phenotypic testing
All 441 NAS isolates were tested for antimicrobial activity against a S. aureus isolate
(derived from clinical mastitis). Only the NAS isolates that inhibited this S. aureus were tested
against the MRSA. Testing was done using a cross-streaking method, modified from a previous
report (De Vliegher et al. 2004). Each isolate was plated on 5% defimbrinated sheep blood agar
plates (BD Diagnostics, Mississauga, ON, Canada) and incubated overnight at 37°C. A single
colony was diluted in PBS to a McFarland 0.5 standard and was used to inoculate a centre streak
(5 mm) on a 5% sheep blood agar plate and subsequently incubated at 37°C for 24 h. On day 2,
the agar was loosened from the plate with sterile metal tweezers and flipped onto the lid of the
plate so the NAS centre streak was face down. Then, 100 µL of a 10-3 dilution in PBS of a
McFarland 0.5 standard of a single colony from an overnight culture on 5% sheep blood agar of
S. aureus was spread over the entire agar surface and incubated at 37° for 24 h. On day 3, plates
were examined for bacterial growth and any inhibition (total or partial) of pathogen growth was
recorded. All experiments included a negative control (PBS was used to make the center streak
on day 1).
2.3.3 Effect of proteinase K on inhibition
Strains that were both inhibitors of S. aureus and potential bacteriocin producers were
tested to confirm the peptide nature of the inhibitory product using proteinase K (20 mg ml -1;

42
Sigma Aldrich) and an agar well diffusion assay. Concentrated cell free supernatant was obtained
from BHI cultures of the 21 inhibiting and potentially producing NAS isolates by performing a
modified version of a chloroform extraction described previously (Burianek and Yousef 2000).
Briefly, 40ml of BHI broth was inoculated with 0.1% of an overnight culture of NAS and
incubated at 37° for approximately 20hr. Cells were removed by centrifugation at 4500g at 4° for
15 min. 20ml of chloroform (Sigma-Aldrich) was mixed with the cell free supernatant of each
NAS and stirred vigorously for 20 min, followed by centrifugation at 4500 g at 4° for 15 min.
The cloudy interfacial precipitate was collected and dried overnight. The dried product was
dissolved in 1ml phosphate buffer solution (PBS) using a magnetic spinner at 4° for 24h.
Proteinase K was added to each concentrated cell free supernatant. Samples with and
without proteinase K were incubated at 37° for 1 hr. Residual inhibition was tested using an agar
well diffusion assay (Schillinger and Lücke 1989).
2.3.4 Whole genome sequencing, assembly, and annotation
Sequencing, assembly, and annotation for NAS and the clinical mastitis S. aureus isolate
were performed as described (Naushad et al. 2016). Briefly, genomic DNA was extracted with
DNeasy Blood and Tissue Kit (Qiagen, Toronto, ON, Canada), according to the corresponding
protocol for Gram-positive bacteria. Sequencing of these samples was performed using the
Illumina MiSeq platform (Illumina, San Diego, CA, USA); DNA libraries for sequencing were
prepared using a Nextera XT DNA library preparation kit (Illumina, San Diego, CA, USA). All
sequencing steps, including cluster generation, paired-end sequencing (2 × 250 bp), and primary
data analysis for quality control, were performed on the instrument. Genome assembly was
automated using the Snakemake workflow engine (Köster and Rahmann 2012). Raw read pairs

43
were screened for adapters and quality trimmed using Cutadapt 1.8.3 (Martin 2011) as
implemented in Trim Galore! 0.4.0 (with default parameters). Genomes were assembled using
Spades 3.6.0 (Nurk et al. 2013) using the built-in error correction and default parameters. To
assess coverage, reads were mapped back to the assembled genome using BWA 0.7.12-r1039 (Li
and Durbin 2009). Contigs > 200 bp were annotated with Prokka 1.11 (Seemann 2014) using the
provided Staphylococcus database. Assembly quality was evaluated with Quast 3.0 (Gurevich et
al. 2013). Contigs, as well as the annotated protein sequences, were used for custom blast
searches using SequenceServer (Priyam et al. 2015). Data were previously submitted to NCBI
under BioProject ID PRJNA342349.
2.3.5 Screening of genomes for bacteriocin clusters
Identification of biosynthetic gene clusters related to secondary metabolite production and
analysis of sequences of interest was done using antiSMASH 3 (Weber et al. 2015). Each gene in
identified clusters was further examined using the BlastP web server on NCBI
(http://www.ncbi.nlm.nih.gov/BLAST), and the presence of conserved domains from the
Conserved Domain Database (Marchler-Bauer et al. 2017) was noted in each coding region and
compared to previously identified known conserved domains in bacteriocin-associated genes
(Azevedo et al. 2015). Putative gene clusters were classified according to Cotter et al. (Cotter,
Ross, and Hill 2013). Additionally, any identified structural genes in the NAS genomes were
used to BLAST back against our NAS genomes, to potentially identify any clusters not detected
by antiSMASH.

44
2.3.6 BLAST
The 441 NAS whole genomes were assessed by BLAST for any bacteriocin structural
genes contained in Classes I, II, and III databases from BAGEL
(http://bagel.molgenrug.nl/index.php/bacteriocin-database). Any genomic regions with identified
bacteriocin-associated genes after the BLAST search were visualized using Geneious version
8.1.6 (Kearse et al. 2012) to determine if the bacteriocin gene cluster was complete by assessing
if the structural gene and known essential associated genes were present using the BlastP web
server on NCBI (http://www.ncbi.nlm.nih.gov/BLAST).
2.3.7 Genome comparison
To further identify potential bacteriocin-associated genes, genomes of the inhibiting NAS
were compared to closely related genomes of non-inhibiting NAS of the same species, according
to the phylogenetic trees of each species. For this purpose, the phylogenetic trees for each
species of NAS were constructed using methods as described (Naushad et al. 2016). Briefly,
trees were rooted using Macrococcus caseolyticus and created based on the core genome of the
individual NAS species. The core set were identified using the UCLUST algorithm (Edgar 2010)
and protein families with at least 30% sequence identity and 50% sequence length were
considered core. However, protein families present in ≥ 95% of the input genomes were
considered core and protein families containing potential paralogous sequences (duplicated
sequence in same genome) were excluded. Each protein family was individually aligned using
MAFFT 7 (Katoh and Standley 2013). Aligned amino acid positions which contained gaps in
more than 50% of genomes, were excluded from further analysis. Remaining amino acid
positions were concatenated to create a combined dataset. A maximum-likelihood tree based on

45
this alignment was constructed using FastTree 2.1 (Price, Dehal, and Arkin 2010) using the
Whelan and Goldman substitution model (Whelan and Goldman 2001).
Comparisons were done by identifying shared genes, present in both closely related
inhibiting and non-inhibiting isolates using Spine, a web based application that identifies
common sequences in the input genomes (Ozer, Allen, and Hauser 2014). Sequences unique to
inhibiting isolates were then determined using AGEnt, by subtracting the output of shared
sequences acquired from Spine from the genome of an inhibiting isolate (Ozer, Allen, and
Hauser 2014). Sequences unique to inhibiting isolates were then visualized using Geneious
version 8.1.6 (Kearse et al. 2012) and genes and conserved domains were determined using the
BLASTn web server on NCBI (http://www.ncbi.nlm.nih.gov/BLAST) and Conserved Domain
Database (Marchler-Bauer et al. 2017) to establish any additional bacteriocin-associated
sequences.
2.3.8 Precursor gene alignments
Protein alignments of precursor peptides were generated using MUSCLE (Edgar 2004).
Sequence alignments were viewed and edited with Jalview alignment editor (Waterhouse et al.
2009).
2.4 Results
2.4.1 Phenotypic testing
Out of 441 NAS isolates, 40 isolates (9.1%) from 9 species (S. capitis, S. chromogenes, S.
epidermidis, S. pasteuri, S. saprophyticus, S. sciuri, S. simulans, S. warneri, and S. xylosus)

46
inhibited growth of the bovine clinical mastitis S. aureus isolate (Table 1). Of the 40 inhibiting
isolates, 23 (57.5%) from S. capitis, S. chromogenes, S. epidermidis, S. pasteuri, S. simulans, and
S. xylosus also inhibited growth of the MRSA isolate.
2.4.2 Effect of proteinase K on inhibition
Out of 21 inhibitors that were potential producers, four isolates showed inhibition in the
well diffusion assay, and for all of which inhibition was eliminated with the addition of
proteinase K.
2.4.3 Screening of genomes for bacteriocin clusters
In the 441 NAS genomes, 184 putative bacteriocin gene clusters belonging to 143 isolates
were identified. A total of 105 clusters from 95 isolates belonging to 16 species were determined
to be viable clusters, whereas the others were eliminated due to absence of either a structural
gene or essential bacteriocin-associated genes (Table 1). Overall, 21.5% of NAS potentially
produced bacteriocins. Ten of the 441 genomes encoded 2 clusters of different classes/types,
whereas the remaining 85 potential producers contained 1 cluster. Of the 40 inhibitors, 21 were
putative producers, whereas no viable bacteriocin gene clusters were identified in 19 inhibitors.
Class II bacteriocins were most frequently identified, with 69 clusters in 68 isolates from S.
equorum, S. gallinarum, S. haemolyticus, S. hyicus, S. saprophyticus, S. sciuri, S. simulans, S.
succinus, S. warneri, and S. xylosus (Fig 1). Nine of the Class II producers were also inhibitors.
For Class I bacteriocins, lanthipeptides were the most frequently identified type, with 29 clusters
in 29 isolates from S. capitis, S. chromogenes, S. cohnii, S. epidermidis, S. equorum, S.
gallinarum, S. sciuri, S. simulans, S. succinus, and S. vitulinus (Fig 1). Fifteen of the 29

47
lanthipeptide producers were also inhibitors. Three sactipeptide clusters were identified from 3 S.
capitis isolates, 2 of which were inhibitors. Lastly, 4 lasso peptide clusters were identified in 2
non-inhibiting S. fleuretti genomes, a non-inhibiting S. sciuri genome, and an inhibiting S. sciuri
genome. Putative bacteriocin-associated genes were distributed throughout the phylogeny of
NAS (Naushad et al. 2016) (Fig 1), although no isolates from S. agnetis (n=13), S. arlettae
(n=15), S. auricularis (n=2), S. caprae (n=1), S. devriesei (n=8), S. hominis (n=11), S. kloosii
(n=1), S. nepalensis (n=2), and S. pasteuri (n=6) contained putative bacteriocin gene clusters.
There was no obvious clustering based on phylogeny or class of bacteriocin.
2.4.3.1 Class I bacteriocins
Lanthipeptide clusters. Twenty-nine lanthipeptide gene clusters were detected in NAS genomes
(Table 1). Fifteen clusters were classified as type 1 (Fig 2); 12 of those clusters, all from S.
capitis, had an identical LanA structural peptide (Fig 2). Six of the 12 S. capitis producers were
also inhibitors. The 44 aa precursor had 59% identity with nisin (a Lactococcus lactis
bacteriocin) and contained the conserved domain, pfam02052. Two of the remaining type 1
clusters came from 2 inhibiting isolates of S. epidermidis, containing an identical 52 aa precursor
peptide, including the epidermin-conserved domain (TIGR03731) and had 96% identity with
epidermin (accession number P08136; Fig 2). The cluster in S. epidermidis 1778 additionally
contained the LanFEG immunity system and a LanT protein (Fig 2). These 2 isolates were both
the only inhibitors and only producers identified in S. epidermidis, suggesting that the
bacteriocin was responsible for in vitro inhibition (Fig 3). The final type 1 lanthipeptide cluster
identified was detected in non-inhibitor S. equorum 1644. This cluster harboured a 57 aa
precursor that contained the type 1 lanthipeptide conserved domain pfam08130 and had 53%

48
identity with Pep5 and 48% identity with epicidin 280 (Fig 2). This precursor was not identified
in any of our other 441 isolates. Four of the 5 clusters contained a LanR regulator, whereas 3 of
the 5 contained a LanP protease.
Fourteen of the 29 lanthipeptide clusters contained the type 2 LanM modification system.
Ten of those clusters contained single LanM proteins (Fig 4). Of those 10 clusters, 2 inhibitors
from S. chromogenes contained 61 aa LanA1 and 82 aa A2 precursors, which contained the
mersacidin conserved domain (pfam16934); the A1 precursor had 20% identity with the Cyl-L
component of cytolysin produced by Enterococcus faecalis and the A2 precursor had 26%
identity with the Cyl-S component of cytolysin (Fig 3), whereas A1 and A2 had 40% identity
with each other (Fig 4). The S. chromogenes 1348 cluster additionally contained a LanT
transporter (Fig 4). Staphylococcus cohnii 5 contained a cluster with a 39 aa hypothetical protein
that had 19% identity to LanA from mersacidin, although no bacteriocin-associated conserved
domains were identified during BLASTp analysis. Staphylococcus cohnii 1067 also had no
identified structural peptide, although a 41 aa hypothetical protein adjacent to the LanM had 21%
identity with the S. cohnii 5 potential LanA and 25% with LanA from S. simulans 1336 identified
in this study. Staphylococcus simulans 1336, a non-inhibitor, contained a 2-component
lanthipeptide, where the 68 aa A1 had 56% identity with the Cyl-L component of cytolysin and
the 68aa A2 had 57% identity with Cyl-L. Furthermore, A1 and A2 have 80% identity with one
another and contained the 2-component Enterococcus faecalis cytolysin (EFC) conserved
domain (pfam16934). The next 3 lanthipeptide clusters were from 3 inhibiting S. simulans
isolates and all contained identical clusters with LanA peptides (conserved domain pfam04604)
that were identical to NukA, the structural peptide for Nukacin ISK-1(accession number
Q9KWM4). Another cluster, in S. simulans 3061, contained a 105 aa hypothetical protein which

49
may function as a LanA (Fig 3) and had 15% identity with a potential LanA in S. cohnii 5.
Lastly, S. vitulinus 730 contained 2 LanA peptides, which had 53% identity with each other.
Furthermore, A1 had 35% identity with Cyl-L and A2 had 32% identity with both Cyl-S and
mersacidin. Three clusters contained an NADPH-dependent FMN reductase (not normally
associated with lanthipeptide clusters).
The remaining 4 type lanthipeptide clusters were part of a different category of type 2
lanthipeptides that contain 2 structural peptides, plus dual lanM proteins, where each lanM is
responsible for modifying a distinct precursor peptide (Fig 5). Staphylococcus succinus 6028
contained a 73 aa A1 precursor and a 68 aa A2 precursor. Staphylococcus succinus 6028A1 had
34% identity with Lacticin 3147A1, whereas the A2 precursor had 32% identity with Lacticin
3147A2. In inhibitor S. sciuri 225, the 63 aa A1 precursor had 28% identity to Lacticin 3147A1
and the 68 aa A2 precursor had 36% identity with Lacticin 3147A2 and 40% identity with
Lichenicidin A2. Lastly, S. gallinarum 2094 and 1388 contained identical novel two-peptide type
2 lanthipeptide biosynthetic clusters with 83aa A1 precursors (conserved domain pfam14867)
that had 34% identity with Lichenicidin A1 and 68aa A2 precursors that had 43% identity with
Lichenicidin A2. The 2 precursors, A1 and A2, had 21.9% identity with each other. Three of the
4 clusters contained a lanP protease.
Sactipeptide clusters. A total of 26 potential Subtilosin-A like clusters were identified, although
23 were excluded due to absence of the critical AlbF gene (Azevedo et al. 2015), leaving 3 S.
capitis isolates that contained viable clusters (Table 1). In S. capitis 1319 (non-inhibitor), 2487
(inhibitor), and 3379 (inhibitor), the structural peptides were all identical and had 63% identity
with the Subtilosin-A precursor, SboA (accession number O07623; Fig 6).

50
Lasso Peptide clusters. Three non-inhibiting NAS isolates, 2 S. fleuretti and 1 S. sciuri, along
with an inhibiting S. sciuri, contained identical clusters encoding for a lasso peptide (Fig 7). The
40 aa structural peptide, ‘A’, had no identified conserved domains, but had 31% identity with a
previously characterized lasso peptide lariatin, produced by Rhodococcus sp. K01-B0171
(Iwatsuki et al. 2006).
2.4.3.2 Class II bacteriocins
Double-glycine leader. Three S. equorum and 1 S. saprophyticus isolate (all non-inhibitors)
encoded for bacteriocin clusters that contained 2 precursor peptides that were annotated as
bacteriocin class II with double-glycine leader peptides (Fig 8). Although the precursor peptides
in all 4 clusters were identical, additional associated proteins varied. The S. saprophyticus and 2
of the S. sciuri clusters contained a SecA protein, possibly related to secretion.
Class IIc circular bacteriocin clusters. Nineteen isolates, 2 of which were inhibitors (across
various clades of NAS phylogeny; Fig 1) contained circular bacteriocin gene clusters encoding 4
distinct bacteriocins (Fig 9). Of the 19 identified clusters, 16 were in S. gallinarum isolates, and
all 16-precursors were identical, although the clusters contained varying bacteriocin-associated
genes (Fig 9). The S. gallinarum precursor showed 25% identity with both gassericin and
enterocin AS-48 (Fig 9). All but 2 of the clusters contained a signal peptidase. Staphylococcus
haemolyticus 109 contained a 97 aa structural peptide with 25% identity with both circularin and
enterocin AS-48. The cluster identified in inhibiting S. simulans 1355 contained a structural
peptide (conserved domain TIGR03651) with 23% identity with enterocin AS-48. The final

51
cluster, identified in inhibiting S. xylosus 4938 contained a precursor with 32% identity with
circularin A.
Class IId clusters. Forty-seven NAS genomes contained complete lactococcin-like clusters (Fig
10). Of these 47 producers, 8 were inhibitors. Twenty-seven lactococcin-like clusters were
identified in 27 S. simulans isolates. Eleven isolates contained a 94 aa structural precursor,
whereas the remaining 15 contained a 105 aa structural precursor, with 88% identity with each
other (Fig 10).
Fourteen isolates contained clusters with the same organization of genes, although the
precursor gene (conserved domain pfam09683) varied slightly among isolates (Fig 10). All 8 S.
xylosus isolates in this group contained an identical 93 aa structural gene with 62% identify with
the structural gene identified in the 2 S. equorum clusters. The S. hyicus cluster’s structural
peptide had 52% identity with the S. xylosus’s structural peptide. Lastly, 3 S. sciuri isolates
structural peptides had 43% identity with the S. xylosus precursor.
An additional S. sciuri isolate, with an identical structural peptide to the other S. sciuri
isolates mentioned above, was identified, although it lacked the ABC transporter in the cluster.
The remaining 5 clusters were identified in S. warneri. The 95 aa putative precursor had 60%
with the 94 aa precursor from S. simulans (Fig 10).
2.5 Discussion
In this study 95 isolates (22%) of 441 NAS encoded for 105 bacteriocin biosynthetic gene
clusters, making them potential bacteriocin producers. Bacteriocin gene clusters were detected,

52
apparently for the first time, in S. equorum, S. haemolyticus, S. succinus, S. capitis, S. cohnii, S.
vitulinus, and S. fleuretti, although some species had demonstrated antimicrobial activity in
previous studies (Braem et al. 2014). According to the phylogenetic tree, bacteriocin gene
clusters are spread throughout the NAS phylogeny, although lasso peptides were only present in
2 species from the same clade and sactipeptides were only identified in S. capitis. In contrast,
only 40 (9%) of 441 NAS were inhibitors when antimicrobial activity was tested in vitro against
a bovine clinical mastitis S. aureus isolate. We identified a higher percentage of NAS with
antimicrobial activity than a previous study that reported only 6.4% of NAS from bovine mastitis
cases were inhibitory (Nascimento et al. 2005). The NAS isolated from the teat apex skin of
dairy cows may have potential for greater bacteriocin production, as 13% of those isolates had
antimicrobial properties in one study(Braem et al. 2014).
Our study identified 29 lanthipeptide clusters from 29 NAS isolates. Both Class I and II
lanthipeptides were identified in NAS genomes. Lanthipeptides are characterized by the presence
of several uncommon amino acids, including meso-lanthionine and 3-methyl-lanthionine, the
result of post-translational modifications (Bierbaum et al. 1996). For Class I lanthipeptides,
dehydration of the serine and threonine residues to dehydroalanine (Dha) and dehydrobutyrine
(Dhb) residues in the pre-peptide, LanA, are completed by a dehydratase (LanB). Thereafter,
thioether crosslinks are formed on the dehydrated amino acids by a cyclase (generically termed
LanC), whereas each specific bacteriocin has its individual naming system, for example nisB and
nisC for nisin. More than 50 lanthipeptides from Gram-positive bacteria have been isolated and
described (Asaduzzaman and Sonomoto 2009), with genome mining identifying many more
potential compounds (Knerr and van der Donk 2012). Previously uncommon genes, e.g. FMN
reductase and N-acetyltransferases, were identified in some clusters in our study, which have

53
been reported in multiple lanthipeptide clusters identified by Singh and Sareen (2014). As we
learn more about post-translational modifications, these genes could help identify novel clusters
in additional genomes. Class I lanthipeptides were identified in S. capitis, S. epidermidis, and S.
equorum. The S. epidermidis strains harbouring the bacteriocin biosynthetic gene cluster also
inhibited S. aureus. The structural gene had 96% similarity with epidermin, the most frequently
produced lanthipeptide by NAS (Bastos et al. 2009) and reported to inhibit human MRSA
(Nascimento et al. 2006) and S. aureus isolated from bovine mastitis (Coelho et al. 2007). Only 6
of 12 S. capitis lanthipeptide producers were inhibitors; therefore, if it is in fact the bacteriocin
identified that is responsible for the inhibition then the remaining 6 non-inhibitors could be
producers of a yet unidentified inhibitory substance, or perhaps the bacteriocin cluster is not
being expressed in the 6 non-inhibitors. This study apparently represents the first time
lanthipeptides have been identified in S. capitis. The S. equorum producer was also a non-
inhibitor, but harbours a gene that likely encodes for a novel bacteriocin (similar to pep5 and
epidicin 280). Class I lanthipeptides pre-peptides typically have a FNLD conserved region
approximately at positions -15 to -20 and a proline at -2 from the cleavage site, which have roles
in modifications of the peptide (Lubelski et al. 2008). Only the LanA identified in the S.
epidermidis strains had the FNLD conserved region and the proline at -2. The LanA identified in
S. capitis had a FDLD motif with the proline at -2, similar to gallidermin and lanthipeptides
recently identified in ruminal bacteria (Azevedo et al. 2015). However, the S. equorum LanA had
a FDLE motif, similar to pep5 and epicidin 280, which had the closest identity when aligned.
One of the most common lanthipeptides identified in NAS to date has been nukacin-like
bacteriocin. In our study, nukacin was discovered in 3 inhibiting S. simulans isolates. The LanA
precursor was identical to Nukacin ISK-1 produced by S. warneri (Sashihara et al. 2000),

54
Nukacin 3299 produced from S. simulans (Ceotto et al. 2010), and Nukacin L217 produced by S.
chromogenes (Braem et al. 2014). That both Nukacin 3299 and Nukacin L217 producer strains
were also isolated from bovine milk and the potential nukacin producer in our study was able to
inhibit S. aureus suggests a role for this bacteriocin in NAS colonization and pathogen inhibition
in the udder environment.
Class II lanthipeptides are characterized by a bifuncational enzyme, LanM, responsible for
both dehydration and cyclisation, whereas the C terminal shares homology with LanC cyclases
of the Class I lanthipeptides (Asaduzzaman and Sonomoto 2009). Ten of 14 clusters identified
contained a single LanM, with 4 containing 2 LanA precursors. The 2 LanA precursors in each
cluster had a high sequence identity (80% for the S. simulans 1336 cluster), suggesting that the
same single LanM enzyme could modify both precursors. For dual precursors with low sequence
identity (e.g. in lichenicidin and haloduracin), there are multiple lanM enzymes to modify each
unique precursor (Dischinger et al. 2009) (McClerren et al. 2006). These represent a distinct
group of type II lanthipeptides, of which 3 were identified in our study. Herein, A1 and A2
precursors had 26, 35, and 22% identity with one another for clusters in S. succinus, S. sciuri,
and S. gallinarum, respectively. However, only the S. sciuri isolate was an inhibitor. Unlike class
II lanthipeptides produced by ruminal bacteria (Azevedo et al. 2015), eight type II clusters in this
study harboured the LanP protease.
Sactipeptides are a group of class I bacteriocins that contain a sulphur to α-carbon linkage,
catalyzed by a recombinant S-adenosylmethionine (rSAM) protein (Fluhe and Marahiel 2013;
Arnison et al. 2013). They were originally only isolated from Bacillus species, although genome
mining has now identified putative gene clusters in the genera Clostridium, Blautia, Kandleria,
Lachnobacterium, Peptostreptococcus, Roseburia, and Ruminococcus (Azevedo et al. 2015;

55
Walsh et al. 2015; Murphy et al. 2011). Hyicin 4244, produced by S. hyicus, was recently
characterized and represents the first sactipeptide in Staphylococcus (Duarte et al. 2017). One
sactipeptide cluster was identified in 3 isolates in our study, although the bacteriocin-associated
genes varied slightly between clusters. Our analysis identified a histidine kinase (HK) and
response regulator (RR) in each cluster, indicating that bacteriocins may be subjected to a 2-
component regulatory system, previously only reported in sactipeptides clusters from ruminal
bacteria (Azevedo et al. 2015). Two of the 3 clusters containing isolates were also inhibitors,
although the S. capitis 3379 isolate additionally contained a lanthipeptide, which could be
responsible for inhibition. The inhibiting S. capitis 2784 did not contain any additional clusters to
our knowledge, although, subtilosin-A, which this novel bacteriocin is most related to,
moderately inhibited S. aureus in vitro (Shelburne et al. 2007). Thus, the non-inhibiting S. capitis
producer was potentially not expressing its bacteriocin gene or the inhibition seen in the other
two isolates was due to another product.
Lasso peptides are an emerging group of RiPPs that do not undergo extensive
modification, although they are folded so that the C terminal is threaded through a ring formed
by a single isopeptide bond, yielding their signature lariat-like form (Maksimov and Link 2014).
To the best of our knowledge, no lasso peptides have been identified in Staphylococcus species
(Hegemann et al. 2015). In this study, an identical, novel cluster was identified in 3 non-
inhibiting isolates, 2 from S. fleuretti and 1 from S. sciuri, along with 1 inhibiting S. sciuri. The
identified clusters contained all 4 of the essential enzymes (‘ABCD’) for peptide production
(Letzel, Pidot, and Hertweck 2014), a transglutaminase-like protein that is likely the ‘B’ gene
which acts as a protease to cleave the leader sequence, a protein with an asparagine synthase
conserved domain, which is likely the ‘C’ gene responsible for isopeptide bond formation, and 2

56
units of an ABC-type dipeptide/oligopeptide/nickel transport system functioning as the ‘D’ gene,
as well as a 53 aa putative protein.
Although most identified putative bacteriocin clusters in this study were class II, there are
only 12 unique precursors in those clusters, compared to the 15 unique lanthipeptide precursors
identified. To date, the majority of Class II bacteriocins in Staphylococcus species have been
identified in S. aureus (Bastos et al. 2009). The first circular bacteriocin in Staphylococcus
species, discovered in 2014, was aureocyclicin 4185 from S. aureus 4185 (Potter, Ceotto,
Coelho, Guimarães, et al. 2014). Herein, we identified putative novel circular bacteriocin gene
clusters from S. gallinarum, S. haemolyticus, S, simulans, and S. xylosus. These bacteriocin
precursors had limited similarity with previously characterized circular bacteriocins, but all
contained the conserved domain associated with the circularin A/uberolysin family. Although the
S. xylosus producer was not the only producer in the species (lactococcin-like clusters were also
identified in other isolates), it was the only inhibitor in the species, suggesting that this
bacteriocin is responsible for activity against S. aureus, although further investigation after
isolation and purification of the peptide is needed. The S. simulans isolate was also an inhibitor
in vitro, but producers from S. gallinarum and S. haemolyticus were non-inhibitors, suggesting
these bacteriocins either have a different spectrum of activity or were not activated.
Class IId bacteriocins are described as linear non-pediocin-like. Lactococcin 972 belongs
to this group and is produced by Lactococcus lactis subsp. lactis IPLA 972 (Martı́nez et al.
1999). The gene cluster, along with the LclA precursor, encodes for a transporter, LclB, and an
immunity gene (Letzel, Pidot, and Hertweck 2014). The 7 class IId lactococcin-like precursors
all had varying degrees of similarity with one another, but all contained the lactococcin 972
conserved domain. Generally, lactococcin-like bacteriocins have a narrow spectrum of activity

57
against Lactococcus species, due to the nature of their binding to receptors (Kjos, Nes, and Diep
2009), indicating it was unlikely that these producers would be inhibitory against S. aureus.
Nonetheless, 8 of 47 lactococcin-like producers were inhibitors. Of these, 8 inhibitors, 4 also
contained a lanthipeptide cluster, whereas 1 also contained a circular bacteriocin cluster, which
could have been the bacteriocins responsible for S. aureus inhibition. Remaining inhibitors could
harbour an unidentified novel bacteriocin responsible for inhibition, or could be producing a non-
bacteriocin inhibitory substance. All of these novel bacteriocins will need further assessments for
spectrum of activity and biochemical characterization to complete identification.
When comparing phenotype and genotype, 95 NAS contained bacteriocin biosynthetic
gene clusters, whereas only 40 of the NAS had inhibition towards S. aureus. This could be due to
several factors, including bacteriocins produced were not effective against inhibiting S. aureus or
the condition tested in vitro did not lead to sufficient levels of bacteriocin production
(Nascimento et al. 2004). This highlights a substantial benefit of genome mining, as variability
of in vitro inhibition testing is negated, enabling clusters that may be silent or repressed in vitro
to be identified. Of the 40 inhibitors, 21 were identified as potential producers and the peptide
nature of the inhibitory product was verified in four of these isolates by elimination of the
inhibition the addition of with proteinase K. Further investigation into the remaining isolates
should be carried out to optimize the extraction conditions and confirm the proteinaceous nature
of the inhibitory compound. For the remaining 19 inhibiting isolates bacteriocin gene clusters
were not detected, perhaps due to inhibition from production of other inhibiting substances, e.g.
low molecular weight antibiotics, lytic enzymes, or metabolic by-products (Leroy and De Vuyst
2004). It could also be due to the nature of the detection software, as identification of clusters
using antiSMASH is based on similarity to previously described genes, with potential to miss

58
completely novel clusters. However, as knowledge increases regarding bacteriocin-associated
genes and structural precursors, detection methods will improve and more bacteriocins will be
described, allowing for even greater detection. In order to conduct the most comprehensive
analysis of bacteriocins currently available, our analysis methods were ordered and combined to
maximize detection. By first using antiSMASH and BLAST searches using the BAGEL
databases, we identified the bulk of the genomes containing bacteriocin gene clusters. We then
used any precursor genes in clusters identified by antiSMASH for further BLAST searches in our
whole genomes, which led to identification of additional lasso peptide clusters not detected by
antiSMASH. Lastly, using our phenotypic results and comparing genomes of inhibitors to
closely related non-inhibitors in the same species and using BLAST searches to analyze unique
sequences in the inhibitor led to identification of bacteriocin-associated genes in 6 of 19
inhibitors not initially identified as producers. However, no complete clusters were identified in
these isolates, due to the absence of peptide precursor genes in close proximity upon
visualization in Geneious.
2.6 Conclusions
In conclusion, all clusters identified, excluding the nukacin identified in S. simulans and
the epidermin variant identified in S. epidermidis, were novel bacteriocin clusters, having less
than 70% identity with previously described bacteriocins. The combination of genome mining
tools, such as antiSMASH along with BLAST searches, makes discovery of novel bacteriocins
quicker and more comprehensive than conventional approaches. The identified putative
producers should be further studied to characterize the bacteriocins described here in order to

59
elucidate structures, modes of action, and spectrum of activity. The NAS isolated from mammary
origin are a rich source of bacteriocins, with >21% being potential producers, thereby
representing a promising source for future research and potential clinical application.
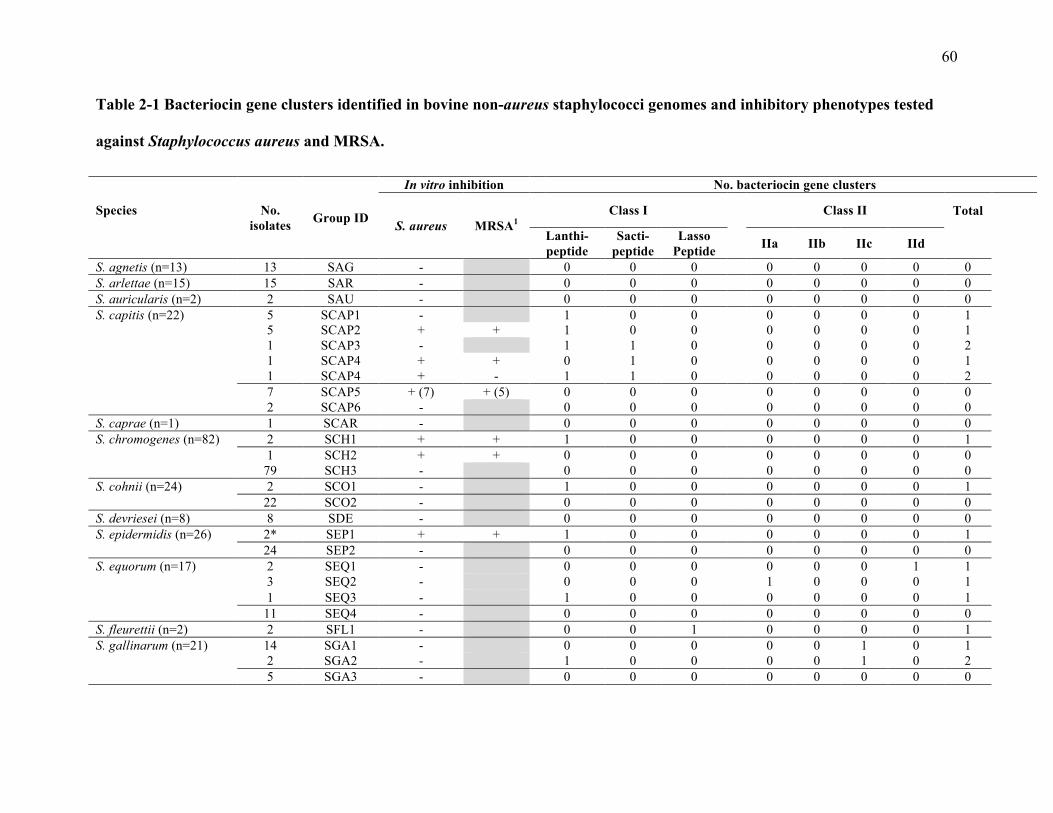
60
Table 2-1 Bacteriocin gene clusters identified in bovine non-aureus staphylococci genomes and inhibitory phenotypes tested
against Staphylococcus aureus and MRSA.
Species No. isolates Group ID
In vitro inhibition No. bacteriocin gene clusters
S. aureus MRSA1 Class I
Class II Total
Lanthi-peptide
Sacti-peptide
Lasso Peptide
IIa IIb IIc IId
S. agnetis (n=13) 13 SAG - 0 0 0 0 0 0 0 0 S. arlettae (n=15) 15 SAR - 0 0 0 0 0 0 0 0 S. auricularis (n=2) 2 SAU - 0 0 0 0 0 0 0 0 S. capitis (n=22) 5 SCAP1 - 1 0 0 0 0 0 0 1
5 SCAP2 + + 1 0 0 0 0 0 0 1 1 SCAP3 - 1 1 0 0 0 0 0 2 1 SCAP4 + + 0 1 0 0 0 0 0 1 1 SCAP4 + - 1 1 0 0 0 0 0 2 7 SCAP5 + (7) + (5) 0 0 0 0 0 0 0 0 2 SCAP6 - 0 0 0 0 0 0 0 0
S. caprae (n=1) 1 SCAR - 0 0 0 0 0 0 0 0 S. chromogenes (n=82) 2 SCH1 + + 1 0 0 0 0 0 0 1
1 SCH2 + + 0 0 0 0 0 0 0 0 79 SCH3 - 0 0 0 0 0 0 0 0
S. cohnii (n=24) 2 SCO1 - 1 0 0 0 0 0 0 1 22 SCO2 - 0 0 0 0 0 0 0 0
S. devriesei (n=8) 8 SDE - 0 0 0 0 0 0 0 0 S. epidermidis (n=26) 2* SEP1 + + 1 0 0 0 0 0 0 1
24 SEP2 - 0 0 0 0 0 0 0 0 S. equorum (n=17) 2 SEQ1 - 0 0 0 0 0 0 1 1
3 SEQ2 - 0 0 0 1 0 0 0 1 1 SEQ3 - 1 0 0 0 0 0 0 1
11 SEQ4 - 0 0 0 0 0 0 0 0 S. fleurettii (n=2) 2 SFL1 - 0 0 1 0 0 0 0 1 S. gallinarum (n=21) 14 SGA1 - 0 0 0 0 0 1 0 1
2 SGA2 - 1 0 0 0 0 1 0 2 5 SGA3 - 0 0 0 0 0 0 0 0

61
S. haemolyticus (n=29) 1 SHA1 - 0 0 0 0 0 1 0 1 28 SHA2 - 0 0 0 0 0 0 0 0
S. hominis (n=11) 11 SHO - 0 0 0 0 0 0 0 0
S. hyicus (n=3) 1 SHY1 - 0 0 0 0 0 0 1 1 2 SHY2 - 0 0 0 0 0 0 0 0
S. kloosii (n=1) 1 SKL - 0 0 0 0 0 0 0 0 S. nepalensis (n=2) 2 SNE - 0 0 0 0 0 0 0 0 S. pasteuri (n=6) 1 SPA1 + + 0 0 0 0 0 0 0 0
5 SPA2 - 0 0 0 0 0 0 0 0 S. saprophyticus (n=16) 1 SSA1 - 0 0 0 1 0 0 0 1
1 SSA2 + - 0 0 0 0 0 0 0 0 14 SSA2 - 0 0 0 0 0 0 0 0
S. sciuri (n=30) 3 SSC1 - 0 0 0 0 0 0 1 1 1 SSC2 + - 1 0 0 0 0 0 0 1 1 SSC3 + - 0 0 0 0 0 0 1 1 1 SSC4 + - 0 0 1 0 0 0 0 1 1 SSC4 - 0 0 1 0 0 0 0 1 6 SSC5 + (6) - 0 0 0 0 0 0 0 0
17 SSC6 - 0 0 0 0 0 0 0 0 S. simulans (n=42) 19 SSI1 - 0 0 0 0 0 0 1 1
1 SSI2 - 1 0 0 0 0 0 1 2 1* SSI3 + - 1 0 0 0 0 0 1 2 1* SSI4 + - 0 0 0 0 0 1 1 2 3 SSI5 + + 1 0 0 0 0 0 1 2 2 SSI6 + + 0 0 0 0 0 0 1 1 1 SSI7 + - 0 0 0 0 0 0 0 0
14 SSI8 - 0 0 0 0 0 0 0 0 S. succinus (n=15) 1 SSU1 - 1 0 0 0 0 0 0 1
14 SSU2 - 0 0 0 0 0 0 0 0 S. vitulinus (n=6) 1 SVI1 - 1 0 0 0 0 0 0 1
5 SVI2 - 0 0 0 0 0 0 0 0 S. warneri (n=19) 5 SWA1 - 0 0 0 0 0 0 1 1
1 SWA2 + - 0 0 0 0 0 0 0 0 13 SWA3 - 0 0 0 0 0 0 0 0
S. xylosus (n=28) 8 SXY1 - 0 0 0 0 0 0 1 1 1* SXY2 + + 0 0 0 0 0 1 0 1 19 SXY3 - 0 0 0 0 0 0 0 0

62
Grey shading: not tested; 1MRSA = methicillin-resistant Staphylococcus aureus; aAn asterisk in the No. isolates column indicates
inhibitory activity was achieved with the chloroform-extracted product and inhibition was suppressed with the addition of proteinase
K.

63
Figure 2-1 Distribution of bacteriocin biosynthetic gene clusters in species of non-aureus
staphylococci isolated from milk of Canadian dairy cows displayed on the phylogenetic tree
from Naushad et al (2017).
Bacteriocin types are indicated according to the following; L, lanthipeptide; S, sactipeptide; Ls,
lasso peptide; II, Class II double glycine leader peptides; C, circular bacteriocins; Lt,
lactococcin-like.
Macrococcus caseolyticus ATCC 13548 Macrococcus caseolyticus JCSC5402
Staphylococcus sciuri (n = 29)
Staphylococcus fleuretti (n = 2) Staphylococcus vitulinus (n = 6)
Staphylococcus chromogenes (n = 83)
Staphylococcus hyicus (n = 3)
Staphylococcus agnetis (n = 13)
Staphylococcus simulans (n = 42)
Staphylococcus aureus (n = 9)
Staphylococcus hominis (n = 11)
Staphylococcus devriesei (n = 8)
Staphylococcus haemolyticus (n= 29)
Staphylococcus pasteuri (n = 6)
Staphylococcus warneri (n = 19)
Staphylococcus epidermidis (n = 26)
Staphylococcus caprae SCR4023 4023
Staphylococcus capitis (n = 22)
Staphylococcus auricularis (n = 2) Staphylococcus kloosii SKL4696 4696
Staphylococcus arlettae (n = 15)
Staphylococcus gallinarum (n = 21)
Staphylococcus succinus (n = 15)
Staphylococcus nepalensis (n = 2)
Staphylococcus cohnii (n = 24)
Staphylococcus equorum (n = 17)
Staphylococcus saprophyticus (n = 16)
Staphylococcus xylosus (n = 28)
100
100
100
10099
100
100
100
10030
100
100
100
100
100
100
100
100
100
100
100
100
100
100
100
86
82
100
100
100
100
100
100
100
100
100
100
100
100
100
75
100
99
100
100
100
100
100
100
100
0.5
L
L
L
L
L
L
L
L
L
Ls
Ls
S
Lt
Lt
Lt
Lt
II
II
C
C
C
C
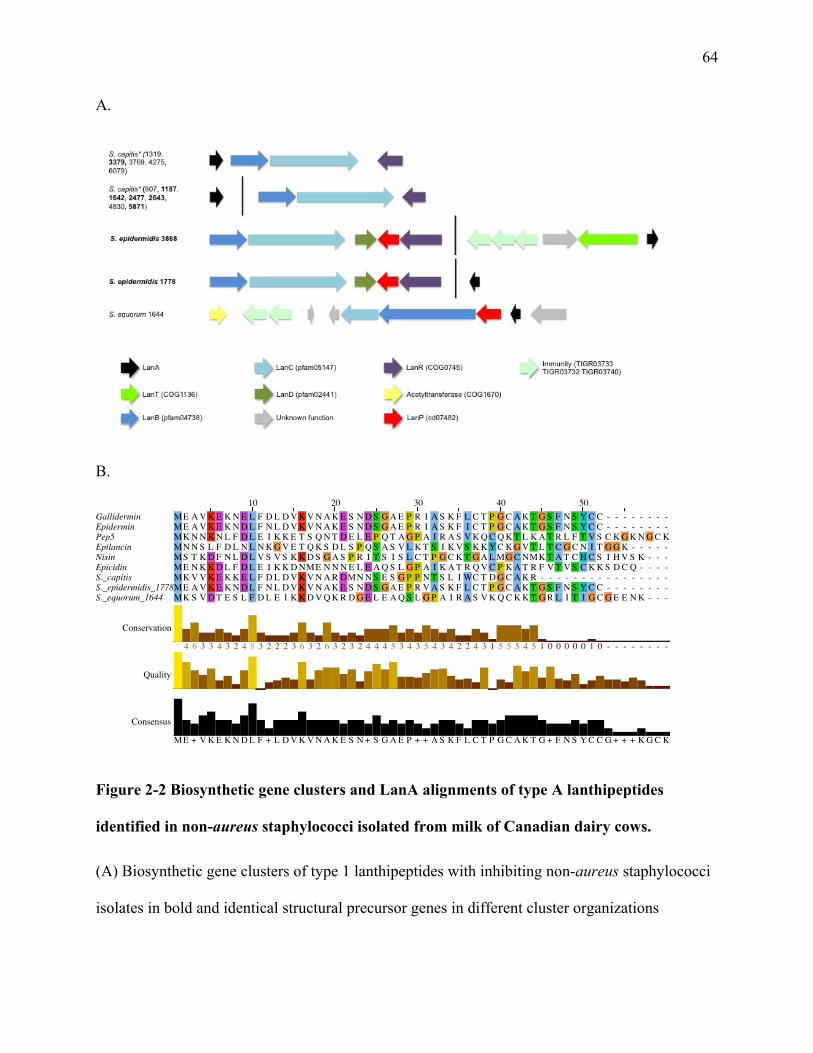
64
A.
B.
Figure 2-2 Biosynthetic gene clusters and LanA alignments of type A lanthipeptides
identified in non-aureus staphylococci isolated from milk of Canadian dairy cows.
(A) Biosynthetic gene clusters of type 1 lanthipeptides with inhibiting non-aureus staphylococci
isolates in bold and identical structural precursor genes in different cluster organizations
10 20 30 40 50GalliderminEpiderminPep5EpilancinNisinEpicidinS._capitisS._epidermidis_1778/3868S._equorum_1644
ME A V K E K N E L F D L D V K V N A K E S N D S G A E P R I A S K F L C T P G C A K T G S F N S Y C C - - - - - - - -ME A V K E K N D L F N L D V K V N A K E S N D S G A E P R I A S K F I C T P G C A K T G S F N S Y C C - - - - - - - -MK N N K N L F D L E I K K E T S Q N T D E L E P Q T A G P A I R A S V K Q C Q K T L K A T R L F T V S C K G K N G C KMN N S L F D L N L N K G V E T Q K S D L S P Q S A S V L K T S I K V S K K Y C K G V T L T C G C N I T G G K - - - - -MS T K D F N L D L V S V S K K D S G A S P R I T S I S L C T P G C K T G A L MG C NMK T A T C H C S I H V S K - - -ME N K K D L F D L E I K K D NME N N N E L E A Q S L G P A I K A T R Q V C P K A T R F V T V S C K K S D C Q - - - -MK V V K E K K E L F D L D V K V N A R DMN N S E S G P P N T S L I WC T D G C A K R - - - - - - - - - - - - - - - -ME A V K E K N D L F N L D V K V N A K E S N D S G A E P R V A S K F L C T P G C A K T G S F N S Y C C - - - - - - - -MK S V D T E S L F D L E I K K D V Q K R D G E L E A Q S L G P A I R A S V K Q C K K T G R L I T I G C G E E N K - - -
Conservation
Quality
Consensus
* 4 6 3 3 4 3 2 4 8 3 2 2 2 3 6 3 2 6 3 2 3 2 4 4 4 5 3 4 3 5 4 3 4 2 2 4 3 1 5 5 5 4 5 1 0 0 0 0 0 1 0 - - - - - - - -
ME + V K E K N D L F + L D V K V N A K E S N + S G A E P + + A S K F L C T P G C A K T G + F N S Y C C G + + + K G C K

65
indicated by *. (B) Multiple sequence alignments of LanA genes identified in type 1
lanthipeptide clusters and known bacteriocins nisin (accession number P13068), gallidermin
(accession number P21838), epidermin (accession number P08136), epilancin K7 (accession
number Q57312), pep5 (accession number P19578), and epicidin 280 (accession number
O54220).

66
Figure 2-3 Phylogenetic tree of Staphylococcus epidermidis isolates from bovine milk
indicating growth inhibition against Staphylococcus aureus and genomically identified
bacteriocin clusters.
L L

67
The percentage of trees in which the associated isolates clustered together is shown next to the
branches, with branch lengths measured in numbers of substitutions per site. Phenotypically
inhibiting isolates are surrounded with a box. Bacteriocin gene clusters are identified by the
following; L, lanthipeptide.

68
A.
B.
Figure 2-4 Biosynthetic gene clusters and LanA alignments of type B lanthipeptides with a
single LanM identified in non-aureus staphylococci isolated from milk of Canadian dairy
cows.
10 20 30 40 50 60 70 80 90 100 110CinnamycinCylL-LCylL-SMersacidinNukacin_ISK-1S._chromogenes_803/1348AS._chromogenes_803/1348BS._cohnii_1067S._cohnii_5S._simulans_1336AS._simulans_1336BS._simulans_1390/1346/1897S._simulans_3061S._vitulinus_730AS._vitulinus_730B
- - - - - - - - - - - - - - - - - - - - - - - - - - - MT A S I L Q Q S V V D A D F R A A L L E N P A A F - - - - G A S A A A L P T P V E A Q D Q - - A S L D F WT K D I A A T E A F A C R Q S C S - - F G P F T F V C D G N T K- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ME N L S V V P S F E E L S V E E ME A I Q G - - S G D V Q A E T T P V C A V A - - - - - - A T A A A S S A A C GWV G G G I F T G V T V V V S L K H C - - - - -- - - - - - - - - - - - - - - - - - - - ML N K E N Q E N Y Y S N K L E L V G P S F E E L S L E E ME A I Q G - - S G D V Q A E T T P A C - - - - - - - - - - - - - - - - - - - - - F T I G L G V G - - A L F S A K F C - - - - -- - - - - - - - - - - - - - MS Q E A I I R S WK D P F S R E N S T Q N P A G N P F S E L K E A QMD K L V G - - A G DME A A C T F T L P G G - - - - - - - - - - - - - - - - - - - - G G - - - - - - V C T L T S E C I C - - -- - - - - - - - - - - - - - - - - - - - - - - - - - ME N S K VMK D I E V A N L L E E V Q E D E L N E V L G - - - A K K K S G V I P T V S H D - - - C HMN S - - - - - - - - - - - - - - - - - - - - F Q F V F T C C S - - - -- - - - - - - - - - - - - - - - - - - - - - - - - - - MS E K A F T E E I V G K S L H S L T Q E E MH S F Y G D F D D N A E V R A S P V A S A V V K E T V K Q G A K A S S A K C A G V I S L S G I G G - A A S S A N D C L G - - -- - - - - - - - - - - - - - - - - - - - - - - - - - - - - MK F N V E E I V G K S I Q T L S E E D L K D F Y N - Q N D D V E V R A T P L T T A P - - - - - - - - - - - - - - - - V S F L A S Y - - - - - L A S E T I D C G S D K -- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - MD E I L K L Q K L N Q A N H T S E DMN S L I D Y R T G R I F T - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - WS S L S N H C - - - - -- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - ME D V L E L Q K L A S D E - - - - - - - - A D V S E K G Y T P T T V T T - - - V G - - - - - - - - - - - - - - - - - - - - - - - L S T I S N G C - - - - -- - - - - - - - - - - - - - - - - - - - - - - - - - - - - MF N K E L K E V V P S F D S L T T E E ME A L I G - - E G D I Q A E T T P A C A A A - - - - - - - - - A A S S G A C A A G A G G F I V G A T A I F S V K Q C - - - - -- - - - - - - - - - - - - - - - - - - - - - - - - - - - - MF N K E L K E V V P S F D S L T T E E ME A L I G - - E G D I Q A E T T P V C A A A - - - - - - - - - A A S S G T C A S F I T G A A S G - - V A F S I K E C - - - - -- - - - - - - - - - - - - - - - - - - - - - - - - - ME N S K VMK D I E V A N L L E E V Q E D E L N E V L G - - - A K K K S G V I P T V S H D - - - C HMN S - - - - - - - - - - - - - - - - - - - - F Q F V F T C C S - - - -
MK Q K I Q L L F S P A I I F L L I Y V I S F T S S Y I T K S Y F I F ML F V Y I F L T I F I H E L G H Y I L - - A R I K K G I L I KM I V G P - - - F Q F S R D A Y I S S N K E WA Y V G G L T - - - I I Y F L N D E K I MY L- - - - - - - - - - - - - - - - - - - - - - - - - - - - - ML N E T N N L V G L S F T E L N Q E E MD F I S G - A E G T V E P Q A T P T I A S P - - - F T P Y A V E I S I G A T V S A V S G - - - - - - L V S Y T K D C L - - - -- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - MK N E N I I G K S F ME L E Q E E MD F I S G - A E G T V E P Q A T P T I S S A - - - V C V R V S L G V S A A G A S F V V S Y - - - - - V A S A T S D C D - - - -
Conservation
Quality
Consensus
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 0 0 0 0 0 2 2 2 2 1 2 6 1 3 3 1 3 2 7 0 0 0 1 0 0 - - 0 3 2 3 2 2 0 0 0 0 1 0 1 0 1 - - - - - - - - - - - - - - - - - - - - - - - - - - - - 5 5 3 2 4 2 1 5 - - - - -
MK Q K I Q L L F S P A I I + + + + + + I + + + + + ME NMK N + + L + E V G P S F E E L T E E E ME A + I G + A E G D V E A E + T P T C A A A + K E + HMN S A A A S S A A C A S F + G G Y G + G + T V A + S T K D C + G + + +

69
(A) Biosynthetic gene clusters of type B lanthipeptides with inhibiting NAS isolates in bold and
identical precursors in various cluster organizations within the same species indicated by *. (B)
Multiple sequence alignments of LanA genes identified in type B lanthipeptide clusters and
known bacteriocins cinnamycin (accession number P29827), cytolysin-L (accession number
H7C7B0), cytolysin-S (accession number H7C7B5), mersacidin (accession number P43683), and
nukacin ISK-1 (accession number Q9KWM4).

70
A.
B-1.
B-2.
Figure 2-5 Biosynthetic gene clusters and LanA alignments of type B lanthipeptides with
dual lanM enzymes identified in non-aureus staphylococci isolated from milk of Canadian
dairy cows.
10 20 30 40 50 60 70 80Lacticin_3174A1Lichenicidin_A1Staphylococci_C55A1*S._gallinarum_2094/1388A1S._sciuri_225A1S._succinus_6028A1
- - - - - - - - - - - - - MN K N E I E T Q - - - - P V T WL E E V S D Q N F - - - - - - - - - - - D E D V F G A C S T N T F S L S D YWG N N G AWC T L T H E C MAWC KMS K K E M I L S WK N P MY R T E S S Y H - - - P A G N I L K E L Q E E E Q H S I A G - - - - - - G T I T L S T C A I - - - - L S K P L G N N G Y L C T V T K E C MP S C N- - - - - - - - - MK S S F L E K D I E E Q - - - - - V T WF E E V S E Q E F - - - - - - - - - - - D D D I F G A C S T N T F S L S D YWG N K G NWC T A T H E C MS WC K- MR K N L E - - - R N P Y L R N E N Q E S V D L P L D N P I R E L K E D E L L K L N G A K S R F K P G A E I A V S T I G C Y A G S V Y L G N D G F MC T T T V E C Q N Q C K- - - - - - - - - - - - - MN K Q E I E L F E - - S A G N F A K E L E N G S L E N I F G G D S - - - Q P R E F T S D N D G K Y - K S I T W - - E C S I C P - T H T C F G - C -- - - MS D F F K Q R K N L T S N T A E K L - - - - D E A I L E E V E D Q S S MG G F - - - - - - - N T WN L T P T S T V G I A A S V G L G N K G K V C T Y T V E C V N N C S
Conservation
Quality
Consensus
- - - - - - - - - - - - - 7 5 4 6 6 5 6 2 2 - - - - 0 3 7 4 6 7 * 9 4 7 4 6 4 - - - - - - - - - - - 3 3 1 4 8 7 5 5 7 4 1 1 1 - 4 * 3 4 7 3 2 5 9 3 6 * 7 1 * 4 6 * 5 6 1 * 1
M+ + K + + + + + + + N P M+ + N E I E E Q + D L P A + N + L E E + + E Q E + + + I F G + + S R F K D T D E F + A C S T + T + + L S + Y + G N + G + WC T + T H E C MNWC K
10 20 30 40 50 60 70 80Lacticin_3174A2Lichenicidin_A2Staphylococci_C55A2S._gallinarum_2094/1388A2S._sciuri_225A2S._succinus_6028A2
- - MK E K NMK K - N D T I E L Q L G K Y L E D DM I E L A E G D E S H G G T T - P - - - - - - - A T P A I S I L S A Y - - - I S T N T C P T T K C T - - - R A C - - -- - MK N S A A R E A F K G A N H P A GMV S E E E L K A L V G G N D V N P E T T - P - - - A T T S S WT C I T A G V T - - - - V S A S L C P T T K C T - - - S R C - - -Q S DMK N E L G K F L E E N E L E L G K F S E S DML E I T D - D E V Y A A G T - P - - - - L A L L G G A A T G V I G Y - - - I S N Q T C P T T A C T - - - R A C - - -- - MK K E D Y K G - - - - - - - - I G L V D E K E L K KMA G A G E V T P R T T I P C G A A V I T A G G V I G G G V K Y T L Q D S A N S C P T G Q C T - - - S K - - - -- - MT N - - - - - - - - - - - - - I Q K V T I K E L E N I S K G S A S S P R V T - P - - - - T T T V V P A S L A V - - - - - - - - - - - C P T T K C A S V V K A C P G K- - MS K D Q E K Q L N E I - - - - T G L I S E D E L E E S L L - E D T F G G T H - P - - - T L S Y I G G A L G G V S A V T A V T A Q S P C P T S A C S - - - K S C - - -
Conservation
Quality
Consensus
- - 5 4 5 1 0 0 0 0 - - - - - - - - 6 6 6 6 3 5 4 9 9 4 3 5 5 2 - 4 4 6 3 7 3 7 6 - * - - - - 1 0 0 5 5 6 8 5 5 7 7 0 1 - - - - 0 2 0 1 0 * * * 7 4 * 7 - - - 5 4 2 - - -
Q S MK K + + + K K + N E + + E L + + G K V S E + E L + E + A G G D E V + P + T T I P C G A A + T T A G G A I + G V + A Y T + + I S A + T C P T T K C T S V V + A C P G K

71
(A) Biosynthetic gene clusters of identified dual precursor type B lanthipeptides with inhibiting
NAS isolates in bold. (B-1) Multiple sequence alignments of LanA1 genes identified in type B
lanthipeptide clusters and known bacteriocins lacticin 3147 A1 (accession number O87236),
lichenicidin A1 (accession number P86475), and staphylococcin C55 A1 (accession number
Q9S4D3). (B-2) Multiple sequence alignments of LanA2 genes identified in type 2 lanthipeptide
clusters and known bacteriocins lacticin 3147 A2 (accession number O87237), lichenicidin A2
(accession number P86476), and staphylococcin C55 A2 (accession number Q9S4D2).
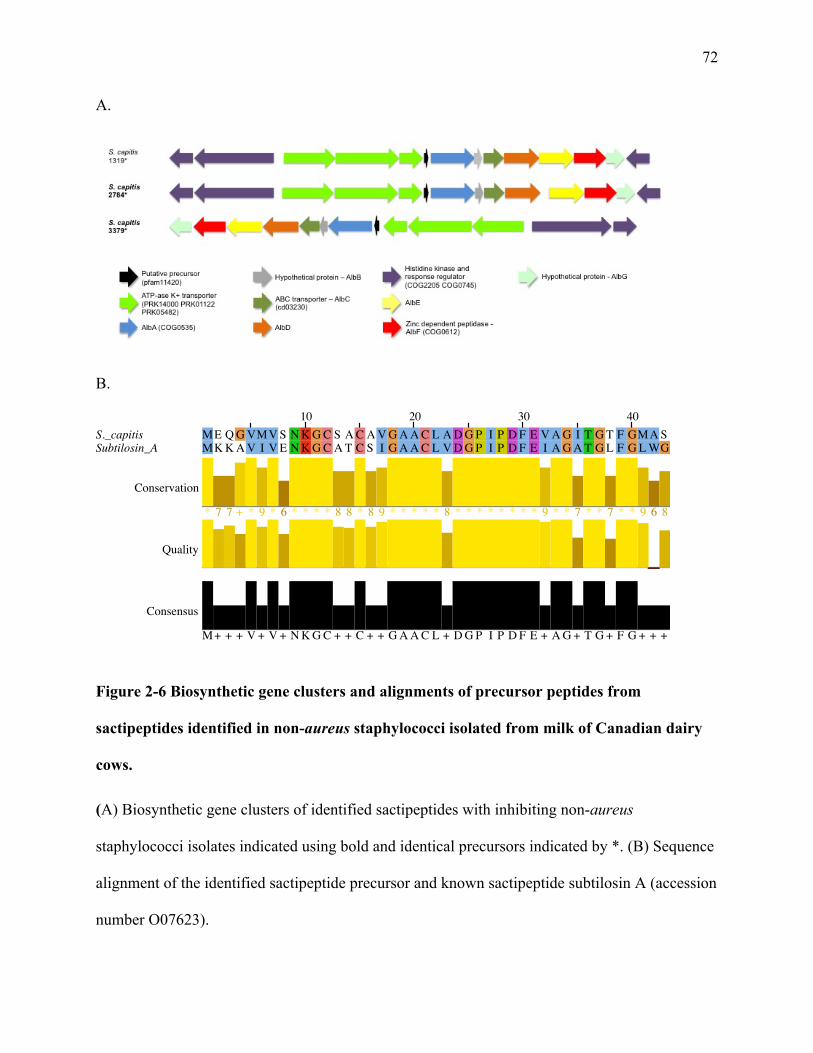
72
A.
B.
Figure 2-6 Biosynthetic gene clusters and alignments of precursor peptides from
sactipeptides identified in non-aureus staphylococci isolated from milk of Canadian dairy
cows.
(A) Biosynthetic gene clusters of identified sactipeptides with inhibiting non-aureus
staphylococci isolates indicated using bold and identical precursors indicated by *. (B) Sequence
alignment of the identified sactipeptide precursor and known sactipeptide subtilosin A (accession
number O07623).
10 20 30 40S._capitisSubtilosin_A
ME Q G VMV S N K G C S A C A V G A A C L A D G P I P D F E V A G I T G T F GMA SMK K A V I V E N K G C A T C S I G A A C L V D G P I P D F E I A G A T G L F G L WG
Conservation
Quality
Consensus
* 7 7 + * 9 * 6 * * * * 8 8 * 8 9 * * * * * 8 * * * * * * * * 9 * * 7 * * 7 * * 9 6 8
M+ + + V + V + N K G C + + C + + G A A C L + D G P I P D F E + A G + T G + F G + + +

73
A.
B.
Figure 2-7 Biosynthetic gene clusters and alignments of precursor peptides from the lasso
peptide identified in non-aureus staphylococci isolated from milk of Canadian dairy cows.
(A) Biosynthetic gene cluster of the identified lasso peptide. Identical precursors indicated with
an * (B) Sequence alignments of the identified lasso peptide precursor and known lasso peptides
lariatin (accession number H7C8I3) and microcin J25 (accession number Q9X2V7).
10 20 30 40 50LariatinMicrocin_J25S._fleuretti/S._sciuri
- - - - - - - - - - MT S Q P S K K T Y N A P S L V Q R G K F A R - - T T A G S Q L V Y R E WV G H S N V I K P G PM I K H F H F N K L S S G K K N N V P S P A K G V I Q I K K S A S Q L T K G G A G H V P E Y F V G I G T P I S F Y G- - - - - - - - - - - - - - - MK K T Y T S P S L V L L G N A K D L I K G S H S S R I P E N A V G I S - - - F Y K G
Conservation
Quality
Consensus
- - - - - - - - - - - - - - - 6 6 5 7 6 7 8 4 8 9 9 7 5 5 6 5 5 5 - - 7 5 8 4 8 6 4 9 5 8 5 6 * * 5 8 - - - 3 5 4 7
M I K H F H F N K L + + + + + + K K T Y + A P S L V Q + G K + A + + + T + + G S + + V P E + + V G I S + + I + + + G

74
Figure 2-8 Biosynthetic gene clusters of Class II double glycine leader peptide bacteriocins
identified in non-aureus staphylococci isolated from milk of Canadian dairy cows.
Identical precursors are identified by *.

75
A.
B.
Figure 2-9 Biosynthetic gene clusters and precursor alignments of Class IIc circular
bacteriocins identified in non-aureus staphylococci isolated from milk of Canadian dairy
cows.
10 20 30 40 50 60 70 80 90 100 110Carnocyclin_ACircularin_AEnterocin_AS-48GassericinUberolysinS._gallinarumS._haemolyticus_109S._simulans_1355
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - ML Y E L V A - - - - - - - Y G I - - A Q G T A E K V V S L I N A G L T V G S I I S I L G G V - - - - - T V G L S G V F T A V K A - - - - - - - - - - - A I A K Q G I K K A I Q L- - - - - - - - - - - - - - - - - - - - - - - - - - - MS L L A L V A G T - - - - - - L G V - - S Q S I A T T V V S I V L T G - - - S T L I S I I L G I T - - - - A I L S G G V D A I L E I GWS A F V A T V K K I V A E R G K A A A I AW- - - - MV K E N K F S K I F I L MA L S F L G L A L F S A S L Q F L P I A HMA K E F G I - - P A A V A G T V L N V V E A G GWV T T I V S I L T - - - - - - - A V G S G G L S L L A A A G R E S I K A Y L K K E I K K K G K R A V I AW
MV T K Y G R N L G L N K V E L F A I - - - WA V L V V A L L L T T A N I YW I A D Q F G I H L A T G T A R K L L D AMA S G A S L G T A F A A I L G V T L P AWA L A A A G A L G A T A A - - - - - - - - - - - - - - - - - - - - - - - -- - - - - - - - - - - - - - - - - - - - - - - - - - - MD I L L E L A G Y - - - - - - T G I - - A S G T A K K V V D A I D K G A A A F V I I S I I S - - - - - - - T V I S A G A L G A V S A S A D F I I L T V K N Y I S R N L K A Q A V I W
MF L R L K N S K E E I K I AMF I T I S S L A V L L I G P L F N A A A A - - - - - - MG L - - N A A T A T S L Y Y A L N F V GWG A T V A S I V A S F - - - - - G L A S I G A Y T I WA A - - - - - - - - - - - - V K R MA L R S F I KW- - - - MN A T K T I N S I I F L T A L V AMS S I A F V L F S T V S P N - - L V S ML S I - - D S G T A L V I V N R V L D T V N L A T I V A S V G T L T - - - - G A G A I G V G L L Q T A KW - - - - - - - - - L A L K F G K K R A Q AW- - - - - MS K V Q D S N V S F L L T L S A S L L V V T V L MF T L P N A P F I A Q Q L G L - - S T A A S L G L A Q A L K T V G N V A T A L T I I G T F T - - - - G V G T I G S S L A A V I - - - - - - - - - L Q K V K K E G A K K A A A F
Conservation
Quality
Consensus
- - - - - - - - - - - - - - - - - - - - - - - - - - - 1 1 5 4 4 4 5 4 6 0 - - - - - - 6 8 9 - - 4 6 8 6 8 2 3 9 5 4 3 8 2 2 7 1 0 1 5 6 7 6 7 5 9 5 - - - - - - - 8 7 7 5 5 * 5 2 6 7 4 2 7 - - - - - - - - - - - - 1 0 2 0 1 1 0 0 1 0 0 1
M+ + + M+ + + K + + + K I + F L + + L S A L A + L + + + L L L T L A + + + + I A + Q L G I H L A + G T A + K V V + A V + + G G + V A T I I S I I G G + T L P AW+ V G S I G + + L A + A A GW+ + I + A T V K K + + K K + G K K + A I AW

76
(A) Biosynthetic gene clusters of identified Class IIc bacteriocins with inhibiting NAS isolates
indicated in bold and identical precursors in S. gallinarum are indicated by *. (B) Alignment of
precursor peptides from identified class IIc bacteriocins and known bacteriocins carnocyclin A
(accession number B2MVM5), and circularin A (accession number BAC164), enterocin AS-48
(accession number Q47765), gassericin (accession number O24790), and uberolysin (accession
number A5H1G9).
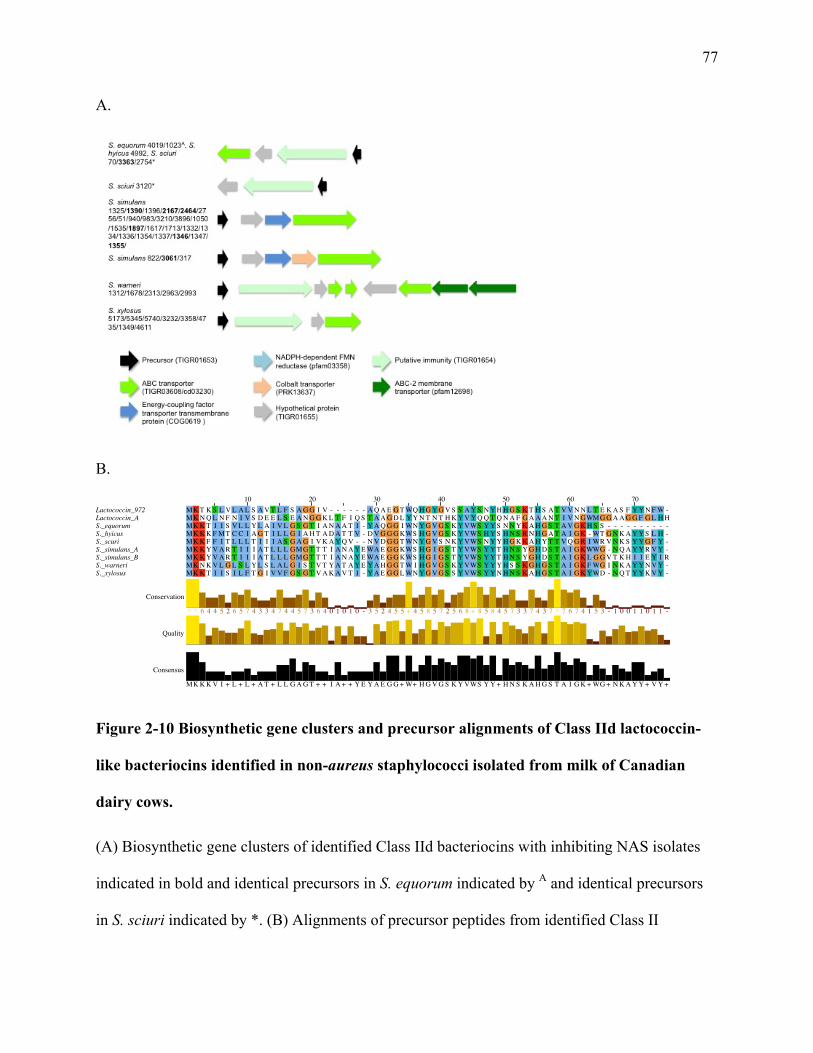
77
A.
B.
Figure 2-10 Biosynthetic gene clusters and precursor alignments of Class IId lactococcin-
like bacteriocins identified in non-aureus staphylococci isolated from milk of Canadian
dairy cows.
(A) Biosynthetic gene clusters of identified Class IId bacteriocins with inhibiting NAS isolates
indicated in bold and identical precursors in S. equorum indicated by A and identical precursors
in S. sciuri indicated by *. (B) Alignments of precursor peptides from identified Class II
10 20 30 40 50 60 70Lactococcin_972Lactococcin_AS._equorumS._hyicusS._scuriS._simulans_AS._simulans_BS._warneriS._xylosus
MK T K S L V L A L S A V T L F S A G G I V - - - - - - A Q A E G T WQ H G Y G V S S A Y S N Y H H G S K T H S A T V V N N L T E K A S F Y Y N F W -MK N Q L N F N I V S D E E L S E A N G G K L T F I Q S T A A G D L Y Y N T N T H K Y V Y Q Q T Q N A F G A A A N T I V N GWMG G A A G G F G L H HMK K T I I S V L L Y L A I V L G S G T I A N A A T I - Y A Q G G I WN Y G V G S K Y VWS Y Y S N N Y K A H G S T A V G K H S S - - - - - - - - - -MK K K F MT C C I A G T I L L G I A H T A D A T T V - D V G G G KWS H G V G S K Y VWS H Y S H N S R N H G A T A I G K - WT G N K A Y Y S L H -MK K F F I T L L L T I I I A S G A G I V K A Y Q V - - N V D G G T WN Y G V S N K Y VWS N Y Y H G K K A H Y T T V Q G R I WR V N K S Y Y G F Y -MK K Y V A R T I I I A T L L L GMG T T T I A N A Y E WA E G G KWS H G I G S T Y VWS Y Y T H N S Y G H D S T A I G KWWG - N Q A Y Y R V Y -MK K Y V A R T I I I A T L L L GMG T T T I A N A Y E WA E G G KWS H G I G S T Y VWS Y Y T H N S Y G H D S T A I G K L G G V T K H I I E Y I RMK N K V L G L S L Y L S L A L G I S T V T Y A T A Y E Y A H G G T W I H G V G S K Y VWS Y Y Y H S S K G H G S T A I G K F WG I N K A Y Y N V Y -MK K T I I S I L F T G I V V F G S G T V A K A V T I - Y A E G G L WN Y G V G S S Y VWS Y Y N H N S K A H G S T A I G K YWD - N Q T Y Y K V Y -
Conservation
Quality
Consensus
* * 6 4 4 5 2 6 5 7 4 3 3 4 7 4 4 5 7 3 6 4 0 1 0 1 0 - 3 5 2 4 5 5 + 4 5 8 5 7 2 5 6 8 + 8 5 8 4 5 7 3 3 7 4 3 7 * 7 6 7 4 1 5 3 - 1 0 0 1 1 0 1 1 -
MK K K V I + L + L + A T + L L G A G T + + I A + + Y E Y A E G G + W+ H G V G S K Y VWS Y Y + H N S K A H G S T A I G K + WG + N K A Y Y + V Y +

78
bacteriocins and known bacteriocins Lactococcin 972 (accession number O86283) and Lactoccin
A (accession number P0A313).

79
Chapter Three: Identifying putative bacteriocin resistance genes in Staphylococcus aureus whole genomes
3.1 Abstract
Mastitis is the most costly disease in dairy cattle and is the largest contributor to antibiotic
use on farm. Staphylococcus aureus is the most detrimental mastitis causing pathogens. In this
study, we determined the susceptibility of 139 bovine S. aureus isolates to a S. chromogenes
isolate that was previously identified to inhibit the growth of S. aureus and to encode a
lanthipeptide bacteriocin. We also determined the presence of bacteriocin gene clusters in the S.
aureus whole genomes to examine the association with susceptibility to the growth inhibition
caused by the S. chromogenes isolate. At the pan genome level, we also identified genes
potentially related to susceptibility and resistance by comparing the presence of putative
immunity related genes or genes encoding hypothetical proteins between susceptible and
resistant S. aureus phenotypes. The distributions of additional putative bacteriocin resistance
genes, which were obtained through the NCBI database and through examining accessory
genomes of resistant isolates, were also determined for each susceptibility group. Forty-nine
(35%) S. aureus isolates were susceptible to the S. chromogenes, whereas 90 (65%) isolates were
resistant in the in vitro inhibition assay. Seventy-four putative bacteriocin gene clusters in 73 S.
aureus whole genomes were identified. The presence of having a bacteriocin cluster was
associated with an isolate being susceptible to the S. chromogenes. The pan genome of S. aureus
isolates contained 6559 genes. Genes that were present in over 90% of all isolates, as well as
genes contained in less than 40% of isolates, were disregarded. Thence, genes were selected for
further analysis on the basis of them either encoding for hypothetical proteins or for proteins that

80
are known to be potentially involved in bacteriocin resistance. This resulted in 273 genes to be
assessed for the distribution between the susceptible and resistant groups. Seventy-seven genes
had a positive association with an isolate being resistant to the S. chromogenes, while 76 genes
were associated with the S. aureus isolate being susceptible. In conclusion, susceptibility seems
to be linked to a large number of genes, the majority of which are annotated as hypothetical
proteins and will need further assessment to determine their role in S. aureus susceptibility.
Key-words: Staphylococcus aureus, non-aureus staphylococci, coagulase-negative
staphylococci, bacteriocin

81
3.2 Introduction
Staphylococcus aureus is most harmful mastitis pathogen on dairy farms, causing clinical
mastitis (CM) cases, increased somatic cell count (SCC), and large economic losses due to
decreased milk yield and increased culling. Antibiotic therapy is frequently inadequate against
chronic intramammary infections (IMI) caused by S. aureus and with increased consumer
interest and stricter antibiotic use policies on the horizon due to the mounting antibiotic
resistance there is a need to investigate alternatives to antibiotic use for the treatment of bacterial
infections on farm. Conceivable alternatives are bacteriocins, which have been shown to inhibit
mastitis pathogens in vitro (Braem et al. 2014; Brito, Somkuti, and Renye 2011; Ceotto‐Vigoder
et al. 2016; Chaimanee et al. 2009; Coelho et al. 2007; Kim et al. 2010; De Vliegher et al. 2004;
Leon-Galvan et al. 2015; Nascimento et al. 2005) and have been investigated for use in a teat dip
(Klostermann et al. 2010) and teat sealants (Crispie et al. 2004).
Bacteriocins are ribosomally synthesized and post-translationally modified peptides
(RiPPs) that generally exert bacteriocidal action across a small spectrum of closely related
bacteria (Cotter, Ross, and Hill 2013). Bacteriocins from Gram-positive bacteria are classified
into two main groups, the lanthipeptides, containing post translationally modified peptides, and
class II bacteriocins, containing the circular and the non-modified peptides (Cotter, Ross, and
Hill 2013). Bacteriocins have been garnering attention as potential alternatives to antibiotics due
to their ability to inhibit the growth of drug resistant bacteria (Nascimento et al. 2006;
Kruszewska et al. 2004) and to not cause significant effects on host natural microbiota (Rea et al.
2010). Bacteriocin producers encode specific proteins in the bacteriocin biosynthetic gene cluster
to provide self-protection. In lanthipeptides, immunity is conveyed with proteins designated as

82
LanI, or the LanFE(G) immunity clusters. LanI is thought to function by interception, target
shielding (Stein et al. 2005) or by sequestering the bacteriocin on the bacterial cell wall
membrane, as is the case with NisI from nisin (Stein et al. 2003). In contrast, the lanFEG cluster
works by removing the peptide via a dedicated ABC transporter (Stein et al. 2003). Immunity for
class IIa and IIc bacteriocins is obtained when the immunity protein binds directly to the
bacteriocin receptor, inhibiting pore formation (Diep, D 2007). A variety of other families of
genes have been identified as potential immunity proteins, although with no known mechanisms
such as Abi genes, which encode CAAX metalloproteases (Kjos et al. 2010).
Cross immunity between closely related lanthipeptides was initially thought to be rare
(Draper et al. 2009); the nisin immunity proteins, NisI and NisFEG, conferred no protection to
subtilin, a closely related lanthipeptide (Stein et al. 2003), and vice versa (Stein et al. 2005), but
cross-immunity has been reported (Draper et al. 2009). Immunity to the two-peptide
lanthipeptide lacticin 3147 has been demonstrated in isolates that encode the closely related
bacteriocin staphylococcin C55, as well as in pathogens that contain functional immunity
homologues (Draper et al. 2009). The fact that pathogens can contain orphan immunity
homologues has implications for clinical use, as a natural reservoir of resistant organisms
containing immunity homologues could be selected for under conditions of antimicrobial
pressure and render the bacteriocin therapeutic unusable. There is also the potential to transfer
these genes to other organisms, including mastitis pathogens. Although there are many suspected
immunity genes, an investigation into the occurrence of these genes in a large group of S. aureus
isolates has not been explored to date. This information could potentially be extrapolated to
predict the usefulness of a specific bacteriocin as a preventative or therapeutic against S. aureus
infections.

83
The first objective of this study was to determine the susceptibility of 139 bovine S.
aureus isolates against a previously identified lanthipeptide producing bovine S. chromogenes in
vitro (Carson et al. 2017). The second objective was to identify any bacteriocin biosynthetic gene
clusters using genome-mining software in the corresponding 139 S. aureus whole genomes in an
attempt to explain the resistance phenotype observed in vitro. The third objective was to identify
putative bacteriocin resistance genes that are associated with resistant isolates by determining the
distribution of genes of interest from the pan genome in both the susceptible and resistant groups
and examining the associations.
3.3 Materials and methods
3.3.1 Isolates
Staphylococcus aureus and NAS isolates were collected in the National Cohort of Dairy
Farms (NCDF) study conducted across Canada during 2007 and 2008, as described by Reyher et
al. (2011). The Canadian Bovine Mastitis and Milk Quality Research Network (CBMQRN) at
the University of Montreal stored the samples before sending them to the University of Calgary.
A total of 146 S. aureus isolates were selected. The isolates originated from 125 lactating cows
from 52 herds across Canada from Nova Scotia, Prince Edward Island, New Brunswick
(representing Atlantic Canada), Québec and Ontario (representing Central Canada), and Alberta
(representing Western Canada) (Reyher et al. 2011). Two random selections for isolates were
made, one for the clinical cases (51 out of 387 isolates) and one for IMI (95 out of 2915 isolates).

84
3.3.2 Phenotypic testing
An S. chromogenes isolate, which was previously identified to inhibit S. aureus in vitro
and encode a lanthipeptide bacteriocin gene cluster (Carson et al. 2017) was tested for inhibition
against 139 S. aureus isolates. Testing was done using a modified cross-streaking method (De
Vliegher et al. 2004). The S. chromogenes isolate was plated on 5% defimbrinated sheep blood
agar plates (BD Diagnostics, Mississauga, ON, Canada) and incubated overnight at 37°C. A
single colony was diluted in PBS to a McFarland 0.5 standard and was used to inoculate a centre
streak (5 mm) on a 5% sheep blood agar plate and subsequently incubated at 37°C for 24 h. On
day 2, the agar was loosened from the plate with sterile metal tweezers and flipped onto the lid of
the plate so the NAS centre streak was face down. 100µL of a 10-3 dilution in PBS of a
McFarland 0.5 standard of a single colony from an overnight culture on 5% sheep blood agar of
each S. aureus isolate was spread over the entire agar surface and incubated at 37° for 24 h. On
day 3, plates were examined for bacterial growth and any inhibition (total or partial) of the
pathogen growth was recorded. All experiments included a negative control whereby only PBS
was used to make the center streak on day 1.
3.3.3 Whole genome sequencing, assembly, and annotation
Sequencing, assembly, and annotation for NAS and S. aureus isolates were performed as
described previously (Naushad et al. 2016). Briefly, genomic DNA was extracted with DNeasy
Blood and Tissue Kit (Qiagen, Toronto, ON, Canada), according to the corresponding protocol
for Gram-positive bacteria. Sequencing of these samples was performed using the Illumina
MiSeq platform (Illumina, San Diego, CA, USA); DNA libraries for sequencing were prepared
using a Nextera XT DNA library preparation kit (Illumina, San Diego, CA, US). All sequencing

85
steps, including cluster generation, paired-end sequencing (2 × 250 bp), and primary data
analysis for quality control, were performed on the instrument. Genome assembly was automated
using the Snakemake workflow engine (Köster and Rahmann 2012). Raw read pairs were
screened for adapters and quality trimmed using cutadapt 1.8.3 (Martin 2011) as implemented in
Trim Galore! 0.4.0 with default parameters. Genomes were assembled using Spades 3.6.0 (Nurk
et al. 2013) using the built-in error correction and default parameters. To assess coverage, reads
were mapped back to the assembled genome using BWA 0.7.12-r1039 (Li and Durbin 2009).
Contigs larger than 200 bp were annotated with Prokka 1.11 (Seemann 2014) using the provided
Staphylococcus database. Assembly quality was evaluated with Quast 3.0 (Gurevich et al. 2013).
Contigs, as well as the annotated protein sequences, were used for custom blast searches using
SequenceServer (Priyam et al. 2015). The data for NAS were previously submitted to NCBI
under BioProject ID PRJNA342349. The process to submit S. aureus data has been initiated.
A phylogenetic tree for the S. aureus was constructed using methods as described
previously (Naushad et al. 2016). Briefly, a genome-based phylogenetic tree of 136 S. aureus
isolates was constructed using the published pipeline PhyloPhlAn (Segata et al. 2013). The
PhyloPhlAn approach is based on the use of 400 ubiquitous and phylogenetically informative
proteins. Orthologs of these proteins in S. aureus genomes were detected using USEARCH
v5.2.32 (Edgar 2010). Multiple sequence alignments of these proteins were generated using
MUSCLE v3.8.31 (Edgar 2004) and a Maximum-Likelihood tree was constructed and formatted
using MEGA 6.0 (Tamura et al. 2013).

86
3.3.4 Screening of genomes for bacteriocin clusters
Identification of biosynthetic gene clusters related to secondary metabolite production and
analysis of sequences of interest were done using antiSMASH 3 (Weber et al. 2015). Putative
gene clusters were classified according to Cotter et al. (Cotter, Ross, and Hill 2013).
3.3.5 Screening of genomes for immunity related genes
3.3.5.1 Pan genome
The pan genome of all S. aureus isolates was computed using BPGA (Bacterial Pan
Genome Analysis tool) (Chaudhari et al., 2016). Genes were discarded that were present in over
90% of isolates, as they were likely a part of the core genome and would not have enough
differences between the resistant and resistant groups for differentiation. Genes were also
discarded that were present in less than 40% of genomes, as there likely would not be enough
representation in each group to determine differences. Genes were then further selected for
analysis based on annotation. Only genes that would reasonably be considered to be involved in
immunity, such as those encoding any ABC transporters, proteases, lipoproteins, hypothetical
proteins, bacteriocin precursors, and any genes annotated as bacteriocin immunity proteins, were
selected. The presence of these genes in susceptible and resistant groups was determined using
an in-house python BLAST script. Two x two tables were created to examine the association
between an isolate being susceptible or resistant and having a specific gene or not having that
specific gene. The significance of that association was examined using Fisher’s exact test
(Fisher’s exact test of independence). Data were analyzed using Microsoft Excel, with P < 0.05
considered statistically significant.

87
3.3.5.2 BLAST
Two groups of 10 genomes, each containing 5 randomly selected susceptible and 5
resistant S. aureus isolates, were compared using Spine, a web based application that identifies
both common and accessory sequences in the input genomes (Ozer, Allen, and Hauser 2014).
Sequences identified in the accessory genome of the resistant isolate group were explored for
genes potentially related to immunity based on their annotations. A list of genes was compiled
and the distribution of putative bacteriocin resistance genes was determined in all S. aureus
genomes in both the resistant and susceptible groups by conducting BLASTp searches against a
local S. aureus BLAST database, using command line BLAST+ 2.5.0 (Camacho et al. 2009).
Additionally, an immunity gene database based on literature was compiled to BLAST on
the local S. aureus database to determine the distribution of genes in resistant and susceptible
genomes. Data were analyzed as stated above.
3.4 Results
3.4.1 Phenotypic test
Out of 139 S. aureus isolates, 49 (35%) had growth inhibition from the S. chromogenes
(Table 1) and were deemed susceptible. The remaining 90 isolates were not susceptible and
therefore were deemed resistant.
3.4.2 Screening of genomes for bacteriocin clusters
Seventy-four putative bacteriocin gene clusters belonging to 73 isolates were identified
(Table 1). Overall, 53% of S. aureus potentially produces bacteriocins. Of the 90 resistant S.

88
aureus, 36 (40%) contained putative clusters. Of the 49 susceptible S. aureus, 37 (76%)
contained putative clusters. Lanthipeptides were the main bacteriocin class identified in the
genomes (73 clusters contained lanthipeptides). A class IId lactococcin-like bacteriocin cluster
was also identified. Two distinct lanthipeptide gallidermin-like precursors were identified, which
had 79% identity with one another (Fig 3-1). The precursors, although annotated as gallidermin
precursors, had only 55 and 53% identity with gallidermin (Fig 3-1).
3.4.3 Screening of genomes for putative resistance related genes
The pan genome of the S. aureus isolates consisted of 6559 genes. 847 genes were present
in less than 90% of all isolates but greater than or equal to 40%. Out of this, 273 were selected
based on their potential for being involved in resistance, or were hypothetical proteins. Based on
the p-values of the fisher’s exact test, 156 had significant associations to be potentially related to
susceptibility (124 of which encoded for hypothetical proteins, with no known function).
Of the 156 genes with significant associations, 73 were associated with an isolate being
resistant. An association with a resistant isolate was defined as having a significant p-value from
the fisher’s exact test and by the gene being present in over 50% of the resistant isolates and in
less than 50% of the susceptible isolates. The resulting list consisted of genes encoding three
ABC superfamily ATP binding cassette transporters, a cytosol aminopeptidase, 61 hypothetical
proteins, a lipoprotein, three oligopeptide ABC superfamily ATP binding cassette transporters, a
putative ABC-type transporter, a putative staphylococcal lipoprotein, a putative staphylococcal
tandem lipoprotein, and a S1B family serine protease SplE (Table 3-2).
Seventy-three genes with significant associations were present in greater than 50% of
susceptible isolates and less than 50% of resistant isolates and were thus attributed to an isolate

89
being susceptible. This list contained six ABC superfamily ATP binding cassette transporters,
an ABC-2 family transporter protein, 56 hypothetical proteins, two gallidermin precursors, a
nisin biosynthesis NisC protein, four oligopeptide ABC superfamily ATP binding cassette
transporters, a putative ABC-type transporter, a serine protease SplE_1, and a thermophilic
serine protease precursor (Table 3-3).
Putative bacteriocin resistance genes identified in the accessory genomes of resistant
isolates using the SPINE approach yielded 10 potential genes of interest, three ABC superfamily
ATP binding cassette transporters, an antimicrobial peptide ABC transporter ATPase, a CAAX
amino terminal protease self-immunity, a gallidermin precursor, a macrolide export
ATO_binding/permease protein, two oligopeptide ABC superfamily ATP binding cassette
transporter, and an P-ATPase superfamily P-type ATPase potassium (K+) transporter subunit A.
One oligopeptide ABC superfamily ATP binding cassette transporter was associated with an
isolate being resistant, with 62% of the resistant isolates encoding for that protein, while only
27% of the susceptible isolates did (Table 3-2). Additionally, two ATP transporters were
associated with isolates being non-susceptible, with 54% and 50% presence in resistant isolates,
compared to 22% for both in susceptible isolates (Table 3-2). Presence of the gallidermin
precursor was associated with an isolate being susceptible (Table 3-3).
From the BLAST searches based on literature, no additional genes were identified that
were associated with an isolate being resistant. However, two self-immunity proteins present on
the epidermin biosynthetic gene cluster, EpiE and EpiF, were associated with an isolate being
susceptible (Table 3-3).

90
3.5 Discussion
In this study, 90 isolates (65%) of 139 S. aureus were not susceptible to growth inhibition
by an S. chromogenes isolate that harbours genes encoding a lanthipeptide. A total of 74
bacteriocin biosynthetic gene clusters were identified with antiSMASH and of the 90 resistant
genomes, 36 (40%) were identified to contain 37 clusters, while of the 49 (76%) susceptible
isolates contained 37 putative bacteriocin clusters. Genomes of the susceptible and resistant S.
aureus were examined and 73 genes were identified that were associated with an isolate being
resistant to the S. chromogenes.
Only two classes of bacteriocins were identified in the S. aureus genomes, a lanthipeptide
and a class IId lactococcin-like bacteriocin gene cluster. The presence of a bacteriocin gene
cluster was associated with an isolate being susceptible to the S. chromogenes bacteriocin. The
lanthipeptide clusters had two distinct precursors that were frequently identified in the same
cluster. They were annotated as gallidermin precursors, although they only showed 55 and 53%
identity with gallidermin. In the phylogenetic tree created with other lanthipeptide precursors, it
appears the identified precursors and gallidermin share a common ancestor but the S. aureus
precursors are probable novel lanthipeptides. Further characterization will need to be done to
conclude firstly, if they are viable clusters followed by determining their spectrum of activity,
structures, and modes of action. From the BLAST data looking at the distribution of potential
immunity related proteins in susceptible and resistant isolates, it indicates that the gallidermin
precursor (as it was annotated) is associated with an isolate being susceptible to the S.
chromogenes bacteriocin producing strain. One conceivable mechanism for this is that the
lanthipeptide cluster the S. aureus contains can provide cross immunity to the S. chromogenes

91
bacteriocin, but because resistance is dependent on expression of immunity proteins (Eijsink et
al. 1998), the S. aureus likely was not producing bacteriocin at the time when it was streaked on
the plate, therefore the immunity proteins on the lanthipeptide operon may not have been
produced, thus leaving the S. aureus susceptible to the S. chromogenes. Although, this would
mean that by containing this gallidermin precursor, and thus a lanthipeptide operon, other
bacteriocin immunity genes are excluded, or that the operon affects other bacteriocin immunity
genes, leaving it susceptible.
One method of bacteriocin resistance is immune mimicry, where the target organism
contains an orphan immunity homologue, which normally confers self-immunity to the
bacteriocin producer (Draper et al. 2009). Therefore, investigating the target genomes for
immunity homologues would be a straightforward way to predict susceptibility to a specific
bacteriocin. Unfortunately, the bacteriocin biosynthetic gene cluster of the producing S.
chromogenes did not contain any recognizable immunity proteins or protein clusters (Carson et
al. 2017) to use as a driver sequence to BLAST in the S. aureus genomes. It is possible that the S.
chromogenes contains immunity genes elsewhere on the genome to provide self-protection,
although further investigation into that isolate would need to be conducted for confirmation.
Investigating the gene differences between susceptible and resistant isolates by creating a
pan genome and describing the distribution in each group, as well as by conducting BLAST
searches from accessory genomes of resistant isolates and from literature, we identified 76 genes
where its presence in a genome is associate with that isolate being resistant to a S. chromogenes
isolate. The majority of these genes encoded for hypothetical proteins, meaning that no known
function has been described. Therefore, there is a need for characterization of these hypothetical
proteins to determine their specific functions and how they are potentially involved in immunity

92
to the S. chromogenes isolate. Fifty-six hypothetical proteins were associated with the isolate
containing them to be susceptible to the S. chromogenes isolate. Characterization and
investigation into the roles of these proteins should also be assessed to have a better
understanding of how these proteins contribute to the susceptibility phenotype observed in vitro.
Homology to known genes can be assessed, as well as presence of conserved domains to predict
functions of the hypothetical proteins. Many different strategies can be utilized simultaneously
for resistance to a bacteriocin, as proven with nisin resistance (Kramer et al. 2006), so it is no
surprise that there are so many potential proteins associated with susceptibility. Although, the
high number of hypothetical proteins that seem to be involved in susceptibility is quite large and
it seems unlikely that they are all involved in immunity. The methodology used may result in a
high level of false discovery.
The LanI immunity proteins in lanthipeptide clusters are lipoproteins, which confer the
producer self-immunity by, in the case of nisin, sequestering nisin and thus rendering it inactive
(Stein et al. 2003). Only the structures for NisI (Hacker et al. 2015) and SpaI (for subtilin)
(Christ et al. 2012) have been elucidated. Even though nisin and subtilin are sequentially and
structurally similar, the two immunity lipoproteins do not show cross-resistance for the two
bacteriocins (Hacker et al. 2015). Therefore, the lipoproteins that confer immunity to the
producer need to be quite specific for that bacteriocin, although, it is plausible that other
lipoproteins present in non-producing strains may be able to provide protection against certain
bacteriocins. We identified three lipoproteins that are associated with an isolate being resistant to
the S. chromogenes strain. The next step would be to do multiple sequence alignments with
known lipoproteins involved in immunity to assess any relatedness and also to compare the

93
structures of each lipoprotein to assess for any similar regions that could be indicative of being
related to immunity.
An S1B family serine protease was found to be associated with an isolate being not
susceptible to the S. chromogenes. This family of protease is typically involved in intracellular
protein turnover (Di Cera 2009). These proteases are also found associated with bacteriocin gene
clusters for cleaving the leader peptide off of the core peptide before exporting from the cell. In
terms of bacteriocin resistance, it in feasible that the protease is able to degrade the bacteriocin
before it has a chance to interact with the cell membrane. Studies to indicate where the protease
localizes in the cell could provide some evidence of potential interactions with bacteriocins, as
the nisin resistance protein (NSR), which is able to proteolytically inactivate nisin, localizes on
the cell membrane where it interacts with nisin to provide resistance to the non-nisin producing
strain (Sun et al. 2009). Nisin resistance protein genes (nsr) have been identified across many
species, including Staphylococcus epidermidis, in an operon that potentially provides nisin
immunity (Draper et al. 2015), although it was not identified in any S. aureus genomes in this
study. A serine protease SplE_1 and thermophilic serine protease precursors were both identified
in the pan genome of the S. aureus isolates and were determined to be associated with an isolate
being susceptible to inhibition from the S. chromogenes. The thermophilic serine protease
precursor was identified with the lanthipeptide clusters in antiSMASH (data not shown),
therefore, the possible mechanisms for how it is related to an isolates sensitivity to the S.
chromogenes is likely the same one mentioned previously for the gallidermin precursors.
ATP transporters are also known to be involved in bacteriocin immunity, as they make up
the LanFEG components of lanthipeptide immunity clusters (Stein et al. 2003). They are also
involved in nisin resistance for non-nisin producing strains, by way of removing nisin from the

94
cytoplasmic membrane to prevent it from binding to lipid II to form pores (Kramer et al.
2006). After hypothetical proteins, the ABC transporters were the most identified proteins to be
associated with susceptibility to the S. chromogenes isolate. Although, they were involved in
both isolates being resistant and isolates being susceptible. We could postulate that the proteins
associated with susceptible isolates are likely the FEG components of the lanthipeptide clusters
identified with antiSMASH and it would make sense that they showed the same association to
the susceptible isolates that the gallidermin precursor did.
Studies have demonstrated Abi proteins, which are metalloproteases also called CAAX
proteases, are involved in bacteriocin immunity (Kjos et al. 2010). In this study, one gene that
was annotated as a CAAX amino terminal protease self-immunity proteins were identified in the
S. aureus pan genome and assessed for differences in proportions in the susceptible and resistant
groups, although no associations were significant. Potentially, this protein is not involved in
resistance to the specific S. chromogenes strain tested, but could provide resistance for the S.
aureus strains to a different bacteriocin. There is also potential for the Abi proteins to belong to a
bacteriocin biosynthetic gene cluster (Kjos et al. 2010), and therefore provide the producer with
self-immunity, although no clusters were identified in antiSMASH that contained these proteins.
Future analysis visualizing the areas of the S. aureus genome containing these Abi proteins
should be conducted to rule out the presence of a novel bacteriocin that antiSMASH overlooked.
Further directions could involve transcriptome analyses to determine how the levels of
expression of certain genes change between a susceptible and resistant isolate, as was done with
nisin (Kramer et al. 2006). There is also the potential to focus on characterizing the bacteriocin
produced by S. chromogenes in vitro. Once a mode of action is determined, assessing
mechanisms of resistance and therefore predicting susceptibility to the bacteriocin will be more

95
straightforward. Identifying a target for the bacteriocin can lead to sequence analyses of the
receptors in the S. aureus genomes to potentially link differences in sequences to susceptibility,
as was done with the man PTS system and Class IIa bacteriocins (Kjos, Nes, and Diep 2009).
Additionally, the whole pan genome could be analysed for protein associations between
susceptibility groups, as there are possibilities for proteins to be involved in susceptibility that
are not necessarily recognizable to be classified as putative bacteriocin resistance genes.
3.6 Conclusions
In conclusion, 73 genes were identified that were associated with S. aureus isolates being
resistant to a bacteriocin producing S. chromogenes strain. The majority of identified genes were
coding for hypothetical proteins, but ABC transporters, lipoproteins, and a serine protease were
also detected. The identified putative bacteriocin resistance genes should be further studied to
elucidate their potential role in bacteriocin immunity.

96
Table 3-1 Bacteriocin gene clusters identified in bovine Staphylococcus aureus and in
vitro susceptibility to a lanthipeptide-producing S. chromogenes.
In vitro susceptibility
No. isolates
Group ID
No. bacteriocin gene clusters
Class I
Class II Total
Lanthi-peptide
Sacti-peptide
Lasso Peptide
IIa IIb IIc IId
Susceptible (n=49)
37 AUR1 1 0 0 0 0 0 0 1
12 AUR2 0 0 0 0 0 0 0 0
Resistant (n=90)
35 AUR3 1 0 0 0 0 0 0 1
1 AUR4 1 0 0 0 0 0 1 2
54 AUR4 0 0 0 0 0 0 0 0
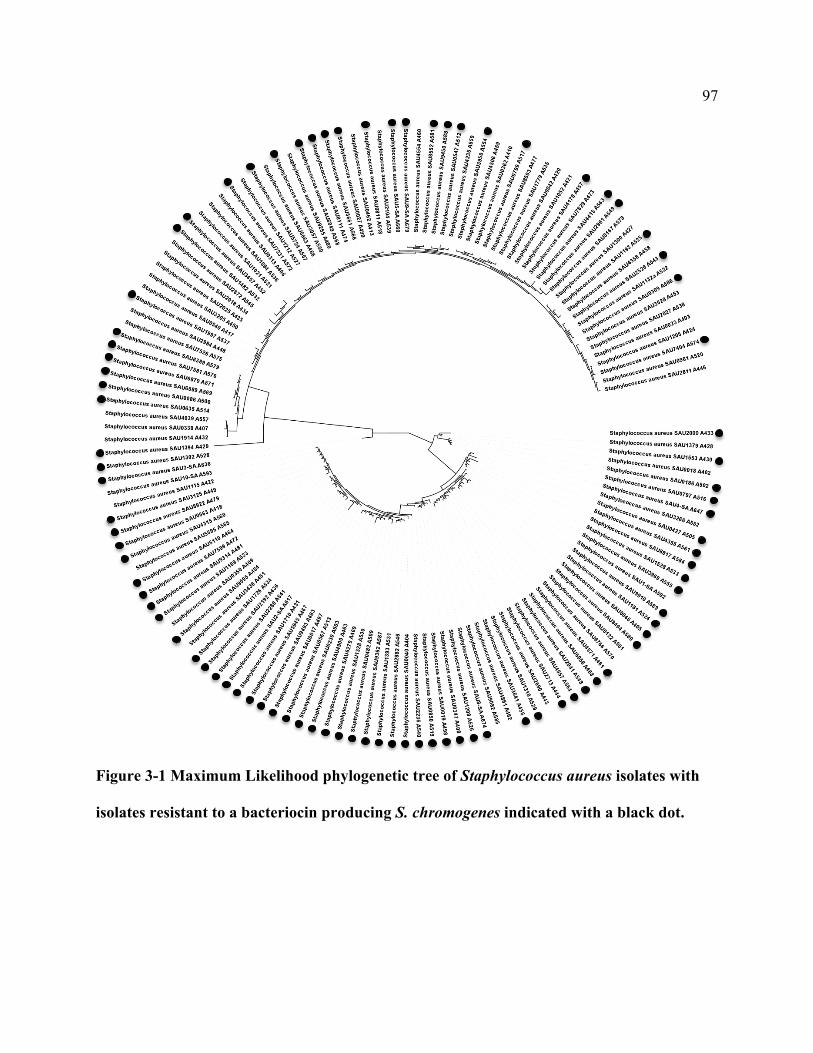
97
Figure 3-1 Maximum Likelihood phylogenetic tree of Staphylococcus aureus isolates with
isolates resistant to a bacteriocin producing S. chromogenes indicated with a black dot.
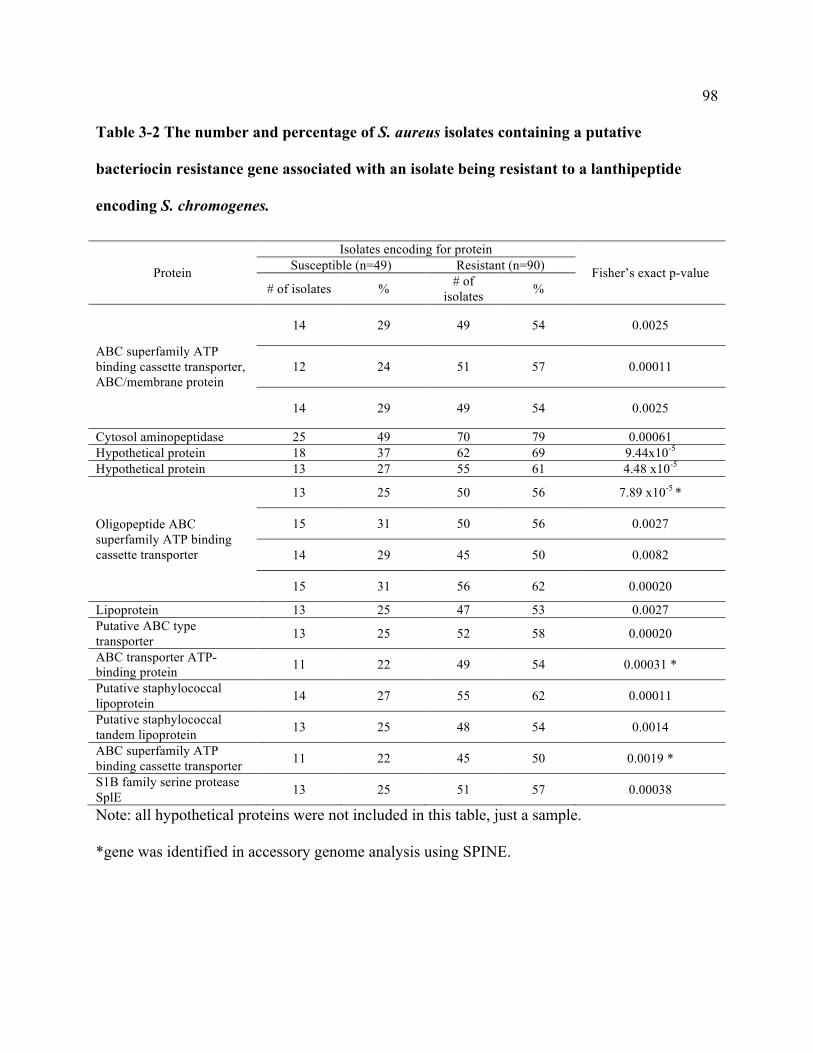
98
Table 3-2 The number and percentage of S. aureus isolates containing a putative
bacteriocin resistance gene associated with an isolate being resistant to a lanthipeptide
encoding S. chromogenes.
Protein
Isolates encoding for protein
Fisher’s exact p-value Susceptible (n=49) Resistant (n=90)
# of isolates % # of isolates %
ABC superfamily ATP binding cassette transporter, ABC/membrane protein
14 29 49 54 0.0025
12 24 51 57 0.00011
14 29 49 54 0.0025
Cytosol aminopeptidase 25 49 70 79 0.00061 Hypothetical protein 18 37 62 69 9.44x10-5 Hypothetical protein 13 27 55 61 4.48 x10-5
Oligopeptide ABC superfamily ATP binding cassette transporter
13 25 50 56 7.89 x10-5 *
15 31 50 56 0.0027
14 29 45 50 0.0082
15 31 56 62 0.00020
Lipoprotein 13 25 47 53 0.0027 Putative ABC type transporter 13 25 52 58 0.00020
ABC transporter ATP-binding protein 11 22 49 54 0.00031 *
Putative staphylococcal lipoprotein 14 27 55 62 0.00011
Putative staphylococcal tandem lipoprotein 13 25 48 54 0.0014
ABC superfamily ATP binding cassette transporter 11 22 45 50 0.0019 *
S1B family serine protease SplE 13 25 51 57 0.00038
Note: all hypothetical proteins were not included in this table, just a sample.
*gene was identified in accessory genome analysis using SPINE.
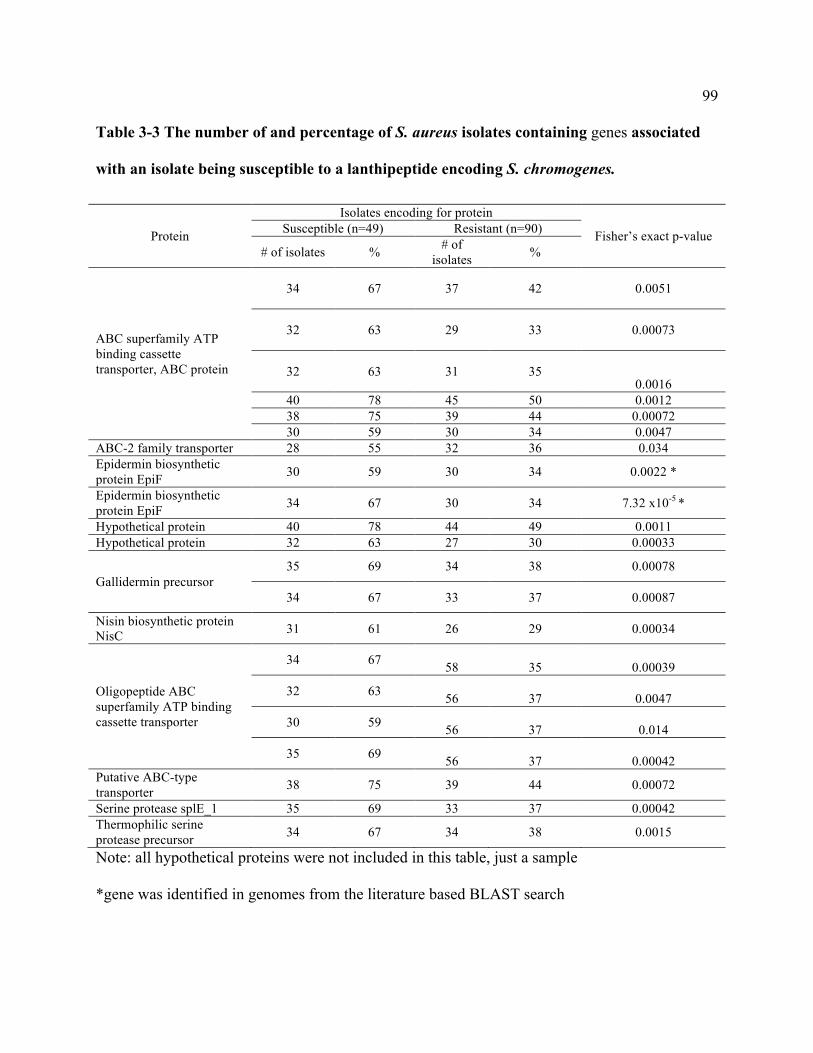
99
Table 3-3 The number of and percentage of S. aureus isolates containing genes associated
with an isolate being susceptible to a lanthipeptide encoding S. chromogenes.
Protein
Isolates encoding for protein
Fisher’s exact p-value Susceptible (n=49) Resistant (n=90)
# of isolates % # of isolates %
ABC superfamily ATP binding cassette transporter, ABC protein
34 67 37 42 0.0051
32 63 29 33 0.00073
32 63 31 35 0.0016
40 78 45 50 0.0012 38 75 39 44 0.00072 30 59 30 34 0.0047
ABC-2 family transporter 28 55 32 36 0.034 Epidermin biosynthetic protein EpiF 30 59 30 34 0.0022 *
Epidermin biosynthetic protein EpiF 34 67 30 34 7.32 x10-5 *
Hypothetical protein 40 78 44 49 0.0011 Hypothetical protein 32 63 27 30 0.00033
Gallidermin precursor 35 69 34 38 0.00078
34 67 33 37 0.00087
Nisin biosynthetic protein NisC 31 61 26 29 0.00034
Oligopeptide ABC superfamily ATP binding cassette transporter
34 67 58 35 0.00039
32 63 56 37 0.0047
30 59 56 37 0.014
35 69 56 37 0.00042 Putative ABC-type transporter 38 75 39 44 0.00072
Serine protease splE_1 35 69 33 37 0.00042 Thermophilic serine protease precursor 34 67 34 38 0.0015
Note: all hypothetical proteins were not included in this table, just a sample
*gene was identified in genomes from the literature based BLAST search

100
A.
B.
Figure 3-2 LanA alignments and phylogenetic tree of type A lanthipeptides identified in S.
aureus isolated from milk of Canadian dairy cows.
10 20 30 40 50 60 70
NisinS._aureus_aS._aureus_bS._capitisEpiderminS._epidermidis_1778/3868GalliderminEpilancin_K7S._equorum_1644Pep5Epicidin_280
- - - - - MS TKD FN L D L - VSV - - SKKDSGASPR I TS I S L - - C TPGC - K T - - - - - - - - - - GA L MGCNMK TA TCHCS I HVSK- - - - - - MENV L D L DVQVKAKND TSDSAGDER I TS F I G - - C TPGCGK T - - - - - - - - - - GS F - - - - - - - - NS FCC - - - - -- - - - - - MEKV L D L DVQVKGNNN TNDSAGDER I TSH F L - - CS FGCGK T - - - - - - - - - - GS F - - - - - - - - NS FCC - - - - -MKVVKEKKE L FD L DVKVNAR - DMNNSESGPPN TS L I W- - C TDGCAKR - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -MEAVKEKND L FN L DVKVNAK - ESNDSGAEPR I ASK F I - - C TPGCAK T - - - - - - - - - - GS F - - - - - - - - NSYCC - - - - -MEAVKEKND L FN L DVKVNAK - ESNDSGAEPRVASK F L - - C TPGCAK T - - - - - - - - - - GS F - - - - - - - - NSYCC - - - - -MEAVKEKNE L FD L DVKVNAK - ESNDSGAEPR I ASK F L - - C TPGCAK T - - - - - - - - - - GS F - - - - - - - - NSYCC - - - - -- - - - - MNNS L FD L N L NKGV - - E TQKSD L SPQSAS - V L K TS I KVSKKY - - - - - - - - - - CKG - - - - - V T L TCGCN I TGGK- MKSVD TES L FD L E I KKDV - - QKRDGE L EAQS L GPA I RASVKQCKK T - - - - - - - - - - GR L - - - - - - - - I T I GCGEENK- - - MKNNKN L FD L E I KKE T - - SQN TDE L EPQTAGPA I RASVKQCQK T L KA TR L F TVSCKG - - - - - - - - KNGCK - - - - -- - - MENKKD L FD L E I KKDN - - MENNNE L EAQS L GPA I KA TRQVCPKA - - - - - - - - - - TR F - - V TVSCKKSDCQ - - - - -
Conservation
Quality
Consensus
- - - - - 0 4 5 4 4 8 8 * 7 9 1 5 3 6 - - 4 3 5 5 5 4 5 4 3 5 5 6 8 0 4 5 - - 7 3 2 5 7 0 * 3 - - - - - - - - - - 2 0 1 - - - - - - - - 0 1 0 2 0 - - - - -
MEAVKEKND L FD L DVKVNAKNE +NDS + +EPR I AS + F L +AC TPGCAK T L KA TR L F TVSGS FMG+ + + + T +NSYCC I + + +K
65.13 Pep5
169.88 Nisin
49.39 S._aureus_a
53.61 S._aureus_b
64.06
14.00Epidermin
14.00S._epidermidis_1778/3868
5.61
15.89 Gallidermin
56.94
13.26
145.74 S._capitis
33.99
27.90
141.57 Epilancin_K7
25.16
127.18 S._equorum_1644
21.32
116.73 Epicidin_280
65.13

101
(A) Multiple sequence alignments of LanA identified in type A lanthipeptide clusters and
known bacteriocins nisin (accession number P13068), gallidermin (accession number P21838),
epidermin (accession number P08136), epilancin K7 (accession number Q57312), pep5
(accession number P19578), epicidin 280 (accession number O54220), and lanthipeptide
precursors identified in Chapter 2. (B) Neighbour joining tree of LanA.

102
Chapter Four: Summarizing discussion
The foremost goal of this thesis was to gain knowledge on the ability of NAS to inhibit S.
aureus. NAS are the most frequently isolated bacteria from the bovine mammary gland, yet are
considered minor pathogens and there are conflicting reports in the literature about the effects of
NAS on udder health. Some studies report that NAS increase SCC (Schukken et al. 2009) and
reduce milk yield, whereas others report an increase in milk yield (Piepers et al. 2010) and a
protective effect against IMI by major pathogens (Matthews, Harmon, and Smith 1990; Braem et
al. 2014). The discrepancy in study results could be due to considering NAS as a group instead
of at the species level. The majority of reports of NAS inhibition seem to be due to S.
chromogenes (De Vliegher et al. 2004; Braem et al. 2014). The apparent protective effect seems
to be due, at least in part, to the production of bacteriocins, which are small antimicrobial
peptides, although a large-scale investigation on bacteriocin production by NAS had not been
done. Therefore, the overall goal of this thesis was to determine the inhibitory capability of NAS
against bovine S. aureus and to determine the distribution and describe any bacteriocin
biosynthetic gene clusters in 441 NAS whole genomes. Additionally, the thesis investigated the
susceptibility of 139 S. aureus isolates to a bacteriocin producing S. chromogenes strain and
aimed to identify any bacteriocin biosynthetic gene clusters and any potential genes related to
susceptibility in the S. aureus whole genomes.
4.1 NAS phenotypic testing

103
The modified cross-streaking method was chosen for this portion of the experiment
due to the ease of assessing inhibition with this methodology, though there are some limitations.
The expression of certain bacteriocins may be dependent on the culture conditions (Espeche et
al. 2014), so different NAS strains that potentially produce different bacteriocins may not all
express that bacteriocin in the conditions that were used in this thesis. Additionally, there is
potential to miss inhibition by only using one indicator species, as different bacteriocins have
different spectrums of activity. The variation in concentration of bacteriocin being produced and
subsequently diffused through the agar is another limitation, which could lead to false negatives
when assessing inhibition. Inhibition can be a result of inhibiting substances other than
bacteriocins, e.g. low molecular weight antibiotics, lytic enzymes, or metabolic by-products
(Leroy and De Vuyst 2004). In order to confirm inhibition is due to a bacteriocin, the bacteriocin
can be isolated, purified, and subsequently tested in an inhibition assay. Although, an often
quicker and less expensive option is to assess cessation of inhibition with the addition of
proteolytic enzyme, which gives a good indication that inhibition is due to a substance that is
protein-like in nature. Though, the cross-streaking method we used for phenotypic assessment is
not well suitable for testing the potential bacteriocin producing strain with proteinase K;
therefore, the well-diffusion assay was employed to test this. Although, because bacteriocins are
produced in broth culture in such low concentrations, the cell-free supernatant of the producing
strain was not concentrated enough to elicit an inhibitory effect in the well diffusion assay.
Consequently, chloroform extractions were carried out on the 21 potential producer/inhibitors in
order to concentrate the bacteriocin from the cell-free supernatant to assess for protease
sensitivity. To this end, we only obtained five NAS chloroform extracted products that were able
to inhibit the S. aureus in the well diffusion assay. For each of the five that we were able to

104
obtain inhibition for, we were able to eliminate inhibition with the addition of proteinase K to
the bacteriocin product, thus indicating that the inhibitory compound is protein-like in nature and
therefore, likely due to bacteriocins. For the remaining 16 isolates that inhibited S. aureus in the
cross streaking method yet did not inhibit in the well diffusion assay via chloroform extractions,
there could be a number of different reasons. The chloroform extraction technique may not have
been the optimal extraction procedure for all of the bacteriocins potentially being produced, as
the properties of the bacteriocins likely differ. It is thus possible that another extraction protocol
could result in a higher yield of bacteriocin from the cell free supernatant. Additionally, the
growing conditions (e.g. temperature) of the producer in broth could have affected the inhibition,
as chloroform extracted products of a nukacin-like bacteriocin were compared and products that
were initially incubated at 30°C had stronger inhibitory effects than those grown at 37°C (Braem
et al. 2014). As well, some bacteria may not produce similar amounts of bacteriocin when grown
in broth, compared to when grown on solid medium. A variety of temperatures while incubating
the producer in broth medium can be explored in the future to optimize bacteriocin extraction.
Another method of assessing proteinase K sensitivity could be done directly on the agar, to
eliminate the need to perform any extractions on cell free supernatant.
In conclusion, the cross-streaking method to initially assess phenotype was sufficient at
identifying 40 NAS that inhibited the growth of S. aureus in vitro. Using this method, combined
with the well diffusion assay to assess for proteolytic enzyme effects on inhibition was adequate
at determining the species-specific effects on NAS on S. aureus and was able to confirm the
protein-like nature of the inhibitory product for 5 NAS potential bacteriocin producers.

105
4.2 Bacteriocin clusters in NAS
Various NAS species have been reported to produce bacteriocins. Staphylococcus
epidermidis produces pep5 (Sahl et al. 1985), epidermin (Allgaier et al. 1986), epilancin K7 (van
de Kamp et al. 1995), epicidin 280 (Heidrich et al. 1998), and epilancin 15X (Ekkelenkamp et al.
2005). Staphylococcus gallinarum produced gallidermin (Schnell et al. 1989). Staphylococcus
cohnii produces staphyloccocin T (Furmanek et al. 1999). Staphylococcus hyicus produces the
lanthipeptide hyicin 3862 (Fagundes et al. 2011) and the sactipeptide, hyicin 4244 (Duarte et al.
2017). Staphylococcus warneri produces a type-AII lanthipeptide, nukacin ISK-1 (Sashihara et
al. 2000). Nukacin-like bacteriocins have also been identified in S. simulans (Ceotto et al. 2010),
S. hominis (Wilaipun et al. 2008), and S. chromogenes (Braem et al. 2014). Lastly, S. aureus
produces Bsa (Daly et al. 2010), the circular bacteriocin aureocyclicin 4185 (Potter, Ceotto,
Coelho, Guimaraes, et al. 2014), and a two-component lanthipeptide, C55 (Maduwe, Sahl, and
Tagg 1999). In this study, we were able to identify 105 putative bacteriocin clusters in 95 NAS
isolates. We identified potential new additions to the pep5 and epidermin groups of
lanthipeptides, along with novel lanthipeptides not closely related to previously described
lanthipeptides. Staphylococcus epidermidis 1778 and 3868 had the strongest inhibition of any
isolate out of the 441 tested, and the identified lanthipeptide varied in one aa from epidermin.
Thus, it is likely a variant of epidermin, and its spectrum of activity should be explored further as
epidermin has shown promise for use in clinical settings (Fontana, de Bastos, and Brandelli
2006; Nascimento et al. 2006).
There are some limitations associated with in silico screening that need to be stated.
Genome mining does not provide any information about biological activity, as evident when

106
comparing our phenotypic test with our genomic screen. We identified isolates with potential
clusters that were not active against S. aureus in vitro, although there is the possibility that the
isolate potentially produces a bacteriocin that is not active against S. aureus or the clusters
identified may not be functional in producing active bacteriocins. One of the initial problems
regarding identifying bacteriocins using software was that bacteriocin-finding tools only looked
for precursor genes in the genomes and as a result clusters were often missed due to the fact that
precursor genes are so small and are frequently overlooked in annotation (Wang, Fewer, and
Sivonen 2011). Current approaches combine mining for the structural gene with mining for the
homologous bacteriocin-associated genes present on the operon, which has increased the
identification of clusters. However, completely novel bacteriocin classes will likely be missed
because they lack homology to known biosynthetic genes and/or structural genes. In this study
however, as discussed in the discussion of Chapter 2, we utilized multiple tools for discovery, in
order to have the most comprehensive analysis of the available genomes. Lastly, although it is
possible to deduce which classes the bacteriocins we identified belong to, the placement into
classes based on bioinformatics alone are not always correct (Arnison et al. 2013), thus further
characterization in the laboratory is necessary for confirmation. Similarity in sequence to a
known precursor also does not guarantee that the two belong to the same class, additionally,
proximity between precursors and biosynthetic genes do not guarantee a target/substrate
relationship (Arnison et al. 2013). For these reasons, biochemical characterization in the
laboratory is necessary to completely characterize the discovered bacteriocins. That being said,
genome mining is a valuable resource to identify potential clusters to be characterized in vitro.
Henceforth, genome mining and laboratory analysis should be paired together in a cyclic
process to maximize bacteriocin discovery. Genome mining can identify isolates to pursue with

107
laboratory analysis, leading to elucidating the bacteriocins biology, which will provide
further information to enhance genome-mining algorithms, leading to the identification of more
bacteriocins.
4.3 Staphylococcus aureus susceptibility
The S. chromogenes isolate for this experiment was chosen based on initial pilot
experiment, wherein the susceptibility of nine S. aureus isolates to the S. chromogenes was
variable (data not shown). The same cross-streaking method utilized in Chapter 2 was used for
this experiment. The limitations of this method discussed above (the differential expression of
bacteriocins in different environments or the spectrum of activity variation) do not apply here, as
we know that the S. chromogenes strain is able to inhibit S. aureus in vitro. We also know that
the S. chromogenes isolate is able to produce a protein-like inhibitory product (as evident from
the positive results of the proteinase K experiment in Chapter 2) that is likely a bacteriocin.
Therefore, the variability in inhibition that we see in the S. aureus is due to something the
specific S. aureus isolate inherently contains, making it resistant to the S. chromogenes.
4.4 Potential immunity genes
Utilizing access to the S. aureus whole genome sequences and determining phenotypic
data for inhibition, assessing putative bacteriocin resistance genes present in the S. aureus was
the next logical step. By comparing the distribution of genes of interest (ones that have been
previously established as being involved in immunity or annotated as hypothetical proteins)

108
present in susceptible and resistant isolates, potential genes could be determined that
contribute to susceptibility. By using the pan genome to select genes for investigation, we were
able to determine 77 genes that were associated with an isolate not being susceptible to the S.
chromogenes and we also were able to determine 76 genes that were associated with an isolate
being susceptible to the S. chromogenes. The majority of these genes encoded for hypothetical
proteins, which will require functional determination to evaluate their role in susceptibility. One
future method that could potentially yield a smaller pool of genes would be to use a different
NAS strain for the phenotypic assessment, one that is less variable in inhibition. The large
amount of variation seen in the susceptibility of S. aureus to the S. chromogenes bacteriocin is no
doubt likely due to multiple resistance mechanisms, whereas with fewer resistant isolates to a
different NAS could potentially mean there are a smaller number of resistance mechanism at
work. Therefore, less variation may result in a better chance at determining what susceptible
strains are lacking that resistant strains contain. Nonetheless, these results show that S. aureus
isolates from the same ecological niche show variable susceptibility to the bacteriocin encoding
S. chromogenes strain and there are a large number of proteins in play that determine
susceptibilities.
4.5 Conclusions and future research
The current studies contribute towards understanding bacteriocin production among
Staphylococcus strains isolated from bovine milk. The results indicate that in silico screening for
bacteriocins is a promising and time effective method to identify potential novel bacteriocins.
This knowledge will be the foundation for future in vitro and in vivo investigations. The results

109
also indicate that susceptibility in S. aureus to an S. chromogenes lanthipeptide can be varied
and there are many genes that are potentially associated with susceptibility.
Chapter one, a review of NAS inhibition and bacteriocins was provided. Bacteriocins
produced by select NAS are likely the reason for the inhibition on mastitis pathogens reported by
previous studies. Indeed, bacteriocins have been recently isolated and characterized from NAS
following in vitro inhibition studies (Braem et al. 2014; Ceotto et al. 2010).
Overall, 40 isolates from S. capitis, S. chromogenes, S. epidermidis, S. pasteuri, S.
saprophyticus, S. sciuri, S. simulans, S. warneri, and S. xylosus inhibited growth of a bovine
clinical mastitis S. aureus isolate. Twenty-three of the 40 inhibiting isolates from S. capitis, S.
chromogenes, S. epidermidis, S. pasteuri, S. simulans, and S. xylosus also inhibited growth of the
MRSA isolate. Ninety-five isolates belonging to 16 species, S. capitis, S. chromogenes, S. cohnii,
S. epidermidis, S, equorum, S. fleurettii, S. gallinarum, S. haemolyticus, S, hyicus, S.
saprophyticus, S. sciuri, S. simulans, S. succinus, S. vitulinus, S. warneri, and S. xylosus, were
identified to contain putative bacteriocin biosynthetic gene clusters, and thus potentially produce
bacteriocins. Only 21 of the inhibiting NAS were also potential producers, as we were not able to
identify bacteriocin clusters in the remaining 19 genomes from inhibitors, although this is not to
definitively say that they do not contain them. It should be noted that although species
identification is important with NAS for a multitude of reasons, bacteriocin production appears
to be strain specific, and not species-related.
The susceptibility of a group of 139 S. aureus to a single S. chromogenes bacteriocin was
examined in Chapter 3, as well as the possible association between susceptibility and putative
bacteriocin resistance genes. Forty-nine S. aureus isolates were inhibited by the S. chromogenes,
while the remaining 90 isolates were not susceptible. After assessing the S. aureus pan genome,

110
as well as creating a BLAST database for genes present in the resistant isolates accessory
genomes and using putative bacteriocin resistance genes based on the literature, 77 proteins were
found that were associated with the resistant S. aureus isolates. The majority of the identified
proteins were hypothetical proteins, which will require future studies for elucidating of their role
in susceptibility.
Described below are suggestions for future research mainly regarding the next steps to
take in the laboratory working towards potential future applications in the dairy industry. One of
the initial next steps could focus on optimizing the growing conditions and purification of
bacteriocins from potential NAS and S. aureus producers, as mentioned previously. Generally,
following a large-scale screen of isolates for inhibitory activity, isolation and characterization of
the bacteriocin from the isolate that exhibited the strongest inhibiting potential in vitro (Braem et
al. 2014) or from those isolates in which a bacteriocin has not been previously described (Ceotto
et al. 2010) is common practice. If we were to pursue the strongest inhibitors from this thesis, the
two inhibiting S. epidermidis isolates exhibited the strongest inhibition of the S. aureus. As for
pursuing bacteriocin isolation from species that have not been identified as having clusters, S.
equorum, S. haemolyticus, S. succinus, S. capitis, S. cohnii, S. vitulinus, and S. fleuretti all were
identified with having clusters in this study and none of which were previously reported to,
although some species had demonstrated antimicrobial activity in previous studies (Braem et al.
2014). An advantage of this study is the availability of the genomic data for presence of
bacteriocin biosynthetic gene clusters. This will eliminate the chance of “re-discovery” of
bacteriocins, as has happened previously with selecting isolates based on inhibition experiments
alone (Ceotto et al. 2010; Braem et al. 2014). With the exception of the nukacin bacteriocin
identified in 3 inhibiting S. chromogenes isolates, all the bacteriocins identified from the in silico

111
screen of NAS and S. aureus are novel. The S. epidermidis isolates that were the strongest
inhibitors contained a bacteriocin cluster, likely encoding for an epidermin variant. Additionally,
because this study represents the first time lasso peptides have been identified in Staphylococcus
genomes the second time sactipeptides have been noted (Duarte et al. 2017), characterization of
those bacteriocins would be noteworthy.
Following purification of a bacteriocin, it can be characterized by determining structure,
size, class, stability, and spectrum of activity. A wide range of pathogens should be used for
spectrum of activity experiments in order to assess the potential usefulness because along with
the benefits of identifying bacteriocins active against mastitis pathogens, identifying bacteriocins
with inhibition against food pathogens, such as Listeria, could be beneficial and offer new
bacteriocins for use in the food preservation industry. Once the bacteriocin is characterized,
experiments can take place to determine the bacteriocins mode of action and cellular target(s).
Certainly, as mentioned previously, assessing the risk of resistance would have to be
thoroughly examined and deemed low, or at least manageable, for there to be any merit of
developing bacteriocins for applications in the industry. The resistance distribution should be
first assessed. The target receptor sequences, if known, can be taken from a wide range of target
organisms and a comparison of receptor sequence and isolate susceptibility can take place, much
like the experiment looking at the man-PTS sequence and Class IIa bacteriocins (Kjos, Nes, and
Diep 2009) to determine the range of isolate susceptibility. Additionally, transcriptome analyses
can be done in order to determine factors related to acquire resistance by determining which
genes up and down-regulate in resistant isolates (Kramer et al. 2006). The rate of acquiring
resistance in vitro should also be examined. Once mechanisms of resistance have been
documented, steps can be taken to decrease the amount of resistance when administering

112
bacteriocins for clinical use. Some recommendations to avoid resistance include using
multiple bacteriocins with different modes of action (Cotter, Ross, and Hill 2013), which can
also increase spectrum of activity (Coelho et al. 2007). Using bacteriocins in conjunction with
antibiotics could also reduce the risk of resistance developing, as bacteriocins and antibiotics
have different modes of action. Activity can also be increased for certain bacteriocins when
combined with other antimicrobials or membrane-active substances (Cotter, Ross, and Hill
2013), which would reduce the selection pressure for resistance. Bioengineering the bacteriocins
is also a possibility for decreasing some resistance, examples of which include increasing the
bacteriocins resistance to proteases (Rink et al. 2010), or to enable them to bind to receptors
which carry gene modifications that made the target bacteria previously resistant (Cotter, Ross,
and Hill 2013).
Once novel bacteriocins have been fully described and risk of resistance has been
assessed, studies can continue to assess the bacteriocins potential for use in the dairy industry. As
indicated previously, the most likely use of NAS bacteriocins in the dairy industry would be in
teat wipes or teat sealants for the prevention of mastitis from Gram-positive pathogens and to
reduce the use of antibiotics. When the Dutch dairy industry stopped blanket dry cow therapy,
antibiotic usage decreased by 56% from 2007 to 2013 (Speksnijder et al. 2015). Although, with
the decrease in antibiotic use at dry off, the incidence rate of CM was 1.7 times higher in quarters
dried off without antibiotics compared to quarters dried off with antibiotics (Scherpenzeel et al.
2014). The pathogen responsible for these CM cases was S. uberis (Scherpenzeel et al. 2014),
and it should be noted that S. uberis is a Gram-positive pathogen that has been shown to be
inhibited by bacteriocins in vitro. Thus, the use of bacteriocins with a teat sealant at dry off could
potentially prevent these CM mastitis cases and mitigate the increase of CM observed. Using just

113
teat sealant without any antibiotics could conceivably push any bacteria in the teat duct up
into the canal during application; having bacteriocin in the teat sealant therefore would reduce
new infections that could occur as a result of this mechanism of infection (Crispie et al. 2005).
Consequently, more studies need to be done on the persistence of bacteriocin in a teat sealant to
determine how long protection could be provided to the cow to be able to establish the usefulness
of bacteriocins in teat sealants at dry off.
There is also the possibility to use bacteriocins to treat mastitis cases in the lactating herd.
Occasionally, intramammary antibiotic treatment for mastitis is ineffective due to the formation
of biofilms in the teat canal, which antibiotics are not able to access (Pieterse and Todorov
2010). Lysostaphin, which is a bacteriolysin, or class III bacteriocin according to some
classification schemes (Heng and Tagg 2006) produced by S. simulans, shows antimicrobial
activity against staphylococci (Bastos, Coutinho, and Coelho 2010) and was recently
demonstrated to disrupt S. aureus biofilms and result in bacterial cell death (Ceotto‐Vigoder et
al. 2016). Nisin application also resulted in S. aureus cell death in pre-formed biofilms (Ceotto‐
Vigoder et al. 2016). The authors also suggest that using a combination of bacteriocins for
prevention and treatment of S. aureus biofilms will decrease the risk of developing resistance, as
the two bacteriocins have different modes of action (Ceotto‐Vigoder et al. 2016). A recent study
from Argentina identified an S. chromogenes isolated from a non-infected quarter that was able
to prevent the formation of biofilms, but had no antimicrobial activity (Isaac et al. 2017). The
bioactive component of the S. chromogenes cell free supernatant was inhibited with the addition
of proteinase K, indicating that it is protein-like in nature (Isaac et al. 2017), and therefore, the
anti-biofilm effects could be due to a bacteriocin. These results indicate that bacteriocins can
potentially be used to treat persistent S. aureus infections with a biofilm component, potentially

114
resulting in less antibiotic use and less economic losses due to IMI. Another benefit of using
bacteriocins to treat lactating cows is the potential reduction of withdrawal times because the
digestive enzymes present in milk would degrade bacteriocins more quickly than antibiotics,
therefore reducing the cost association with lost milk after treatment (Pieterse and Todorov
2010). The persistence of the bacteriocin in the mammary gland would have to be assessed, as
well as its activity and stability at physiological pH.
Also, with mounting consumer interest in probiotic use, one could appreciate the use of
probiotics to battle mastitis and because NAS have shown to inhibit mastitis pathogens in the
udder, it raises the question if they could be used as such. However, it remains to be determined
if the producing strain would be able to produce enough bacteriocin in milk to be effective, and
what the effect the colonization of a bacteriocin producing NAS strain would have on the udder
and milk quality. Lactococcus lactis 19.3 produces a bacteriocin with a wide inhibitory spectrum,
including against L. monocytogenes and was able to grow and produce similar amounts of
bacteriocin in MRS medium, cow's milk and soya milk, respectively (Zamfir et al. 2016).
Another study infused live L. lactis 3147 (which produces lacticin 3147) into the udder, which
had previously showed killing effects of Gram-positive mastitis pathogens (Klostermann et al.
2008), and the authors concluded the inhibition likely came from stimulating the host’s
mammary immune system and the stimulation may be a result of the bacteriocin production
(Crispie et al. 2008). If NAS strains behaved the same in bovine milk, there is potential for
bacteriocin production in the milk of the udder and could explain the inhibition of mastitis
pathogens observed in the field (Matthews, Harmon, and Smith 1990; Piepers et al. 2010), and
why teats colonized by S. chromogenes produce milk with low SCC (De Vliegher et al. 2003).
Staphylococcus chromogenes is the most frequently isolated NAS from the mammary gland

115
(Condas et al. 2017 ) and it has been identified as a producer of bacteriocins in the literature
(Braem et al. 2014; De Vliegher et al. 2004). Although, S. capitis, S. gallinarum, S. hyicus, S.
agnetis, and S. simulans are the most frequently isolated NAS species from quarters with high
SCC (Condas et al. 2017), so using isolates from those species has the potential to increase the
SCC which would not be desirable to the dairy producer. Although, the increase of SCC with
IMI from NAS is relatively small (Condas et al. 2017) and the strain selected for potential
probiotic use would have to be assessed for how it persists in the udder and how it specifically
impacts the udder health and milk quality. Because enzymes in the milk can potentially degrade
purified bacteriocins, administering bacteriocin-producing bacteria that can persist may be a
more effective approach for treatment or prevention of infection. Although, to be used in food
production the strain would have to have a GRAS (generally regarded as safe) status.
In conclusion, this study indicates that NAS are potentially prolific producers of
bacteriocins, some of which have the ability to inhibit S. aureus and MRSA in vitro. This study
also highlights the larger number of proteins that are potentially involved in bacteriocin
resistance in S. aureus. The substantial amount of information generated from the in silico
analyses warrants further investigation.

116
References
Allgaier, Hermann, Gunther Jung, Rolf G. Werner, Ursula Schneider, and Hans Zahner. 1986.
'Epidermin: sequencing of a heterodet tetracyclic 21-peptide amide antibiotic', Eur J
Biochem, 160: 9-22.
Alvarez-Sieiro, Patricia, Manuel Montalbán-López, Dongdong Mu, and Oscar P. Kuipers. 2016.
'Bacteriocins of lactic acid bacteria: extending the family', Appl Microbiol Biotechnol,
100: 2939-51.
Arnison, Paul G., Mervyn J. Bibb, Gabriele Bierbaum, Albert A. Bowers, Tim S. Bugni,
Grzegorz Bulaj, Julio A. Camarero, Dominic J. Campopiano, Gregory L. Challis, Jon
Clardy, Paul D. Cotter, David J. Craik, Michael Dawson, Elke Dittmann, Stefano
Donadio, Pieter C. Dorrestein, Karl-Dieter Entian, Michael A. Fischbach, John S.
Garavelli, Ulf Göransson, Christian W. Gruber, Daniel H. Haft, Thomas K. Hemscheidt,
Christian Hertweck, Colin Hill, Alexander R. Horswill, Marcel Jaspars, Wendy L. Kelly,
Judith P. Klinman, Oscar P. Kuipers, A. James Link, Wen Liu, Mohamed A. Marahiel,
Douglas A. Mitchell, Gert N. Moll, Bradley S. Moore, Rolf Müller, Satish K. Nair, Ingolf
F. Nes, Gillian E. Norris, Baldomero M. Olivera, Hiroyasu Onaka, Mark L. Patchett,
Joern Piel, Martin J. T. Reaney, Sylvie Rebuffat, R. Paul Ross, Hans-Georg Sahl, Eric W.
Schmidt, Michael E. Selsted, Konstantin Severinov, Ben Shen, Kaarina Sivonen, Leif
Smith, Torsten Stein, Roderich D. Süssmuth, John R. Tagg, Gong-Li Tang, Andrew W.
Truman, John C. Vederas, Christopher T. Walsh, Jonathan D. Walton, Silke C. Wenzel,
Joanne M. Willey, and Wilfred A. van der Donk. 2013. 'Ribosomally synthesized and

117
post-translationally modified peptide natural products: overview and
recommendations for a universal nomenclature', Nat Prod Rep, 30: 108-60.
Arthur, T. D., V. L. Cavera, and M. L. Chikindas. 2014. 'On bacteriocin delivery systems and
potential applications', Future Microbiol, 9: 235-48.
Asaduzzaman, Sikder M., and Kenji Sonomoto. 2009. 'Lantibiotics: Diverse activities and unique
modes of action', J Biosci Bioeng, 107: 475-87.
Azevedo, Analice C., Cláudia B. P. Bento, Jeronimo C. Ruiz, Marisa V. Queiroz, and Hilário C.
Mantovani. 2015. 'Distribution and genetic diversity of bacteriocin gene clusters in rumen
microbial enomes', Appl Environ Microbiol, 81: 7290-304.
Babasaki, Katsuhiko, Toshifumi Takao, Yasutsugu Shimonishi, and Kiyoshi Kurahashi. 1985.
'Subtilosin A, a new antibiotic peptide produced by Bacillus subtilis 168: isolation,
structural analysis, and biogenesis', J Biochem, 98: 585-603.
Barboza-Corona, José Eleazar, Norma de la Fuente-Salcido, Nayeli Alva-Murillo, Alejandra
Ochoa-Zarzosa, and Joel E. López-Meza. 2009. 'Activity of bacteriocins synthesized by
Bacillus thuringiensis against Staphylococcus aureus isolates associated to bovine
mastitis', Vet Microbiol, 138: 179-83.
Barkema, H. W., M. A. G. von Keyserlingk, J. P. Kastelic, T. J. G. M. Lam, C. Luby, J. P. Roy,
S. J. LeBlanc, G. P. Keefe, and D. F. Kelton. 2015. 'Invited review: Changes in the dairy
industry affecting dairy cattle health and welfare', J Dairy Sci, 98: 7426-45.
Bastos, M. C. F., H. Ceotto, M. L. V. Coelho, and J. S. Nascimento. 2009. 'Staphylococcal
antimicrobial peptides: relevant properties and potential biotechnological applications',
Curr Pharm Biotechnol, 10: 38-61.

118
Bastos, M. D. D., M. L. V. Coelho, and O. C. D. Santos. 2015. 'Resistance to bacteriocins
produced by Gram-positive bacteria', Microbiology, 161: 683-700.
Bastos, Maria do Carmo de Freire, Bruna Gonçalves Coutinho, and Marcus Lívio Varella
Coelho. 2010. 'Lysostaphin: A Staphylococcal bacteriolysin with potential clinical
applications', Pharmaceuticals, 3: 1139-61.
Begley, Máire, Paul D. Cotter, Colin Hill, and R. Paul Ross. 2009. 'Identification of a novel two-
peptide lantibiotic, lichenicidin, following rational genome mining for LanM proteins',
Appl and Environ Microbiol, 75: 5451-60.
Bierbaum, G., F. Götz, A. Peschel, T. Kupke, M. van de Kamp, and H. G. Sahl. 1996. 'The
biosynthesis of the lantibiotics epidermin, gallidermin, Pep5 and epilancin K7', Antonie
van Leeuwenhoek, 69: 119-27.
Bierbaum, G., and H. G. Sahl. 2009. 'Lantibiotics: Mode of action, biosynthesis and
bioengineering', Curr Pharm Biotechnol, 10: 2-18.
Blin, Kai, Daniyal Kazempour, Wolfgang Wohlleben, and Tilmann Weber. 2014. 'Improved
lanthipeptide detection and prediction for antiSMASH', PloS one, 9: e89420.
Boziaris, I. S., and M. R. Adams. 2001. 'Temperature shock, injury and transient sensitivity to
nisin in Gram negatives', J Appl Microbiol, 91: 715-24.
Bradley, A. J. 2002. 'Bovine mastitis: An evolving disease', Vet J, 164: 116-28.
Braem, G., B. Stijlemans, W. Haken, S. Vliegher, L. Vuyst, and F. Leroy. 2014. 'Antibacterial
activities of coagulase�negative staphylococci from bovine teat apex skin and their
inhibitory effect on mastitis�related pathogens', J Appl Microbiol, 116: 1084-93.
Brito, M. A. V. P., G. A. Somkuti, and J. A. Renye. 2011. 'Production of antilisterial bacteriocins
by staphylococci isolated from bovine milk', J Dairy Sci, 94: 1194-200.

119
Brötz, H., G. Bierbaum, P. E. Reynolds, and H. G. Sahl. 1997. 'The lantibiotic mersacidin
inhibits peptidoglycan biosynthesis at the level of transglycosylation', Eur J Biochem,
246: 193-99.
Burianek, L. L., and A. E. Yousef. 2000. 'Solvent extraction of bacteriocins from liquid cultures',
Lett Appl Microbiol, 31: 193-97.
Camacho, Christiam, George Coulouris, Vahram Avagyan, Ning Ma, Jason Papadopoulos,
Kevin Bealer, and Thomas L. Madden. 2009. 'BLAST+: architecture and applications',
BMC bioinformatics, 10: 421-21.
Carson, Domonique, H. W. Barkema, Sohail Naushad, and Jeroen De Buck. 2017. 'Bacteriocins
of non-aureus staphylococci isolated from bovine milk', Appl and Environ Microbiol, 83:
In Press.
Ceotto, Hilana, Helge Holo, Karlla Fernanda Silva da Costa, Janaína Santos Nascimento, Zhian
Salehian, Ingolf F. Nes, and Maria do Carmo de Freire Bastos. 2010. 'Nukacin 3299, a
lantibiotic produced by Staphylococcus simulans 3299 identical to nukacin ISK-1', Vet
Microbiol, 146: 124-31.
Ceotto�Vigoder, H., S. L. S. Marques, I. N. S. Santos, M. D. B. Alves, E. S. Barrias, A. Potter, D.
S. Alviano, and M. C. F. Bastos. 2016. 'Nisin and lysostaphin activity against preformed
biofilm of Staphylococcus aureus involved in bovine mastitis', J Appl Microbiol, 121:
101-14.
Chaimanee, Veeranan, Chotipa Sakulsingharoj, Somkid Deejing, Pramot Seetakoses, and
Piyanuch Niamsup. 2009. 'Screening and characterisation of bacteriocin-producing
bacteria capable of inhibiting the growth of bovine mastitis', Maejo International Journal
of Science and Technology, 3: 43-52.

120
Chatterjee, Champak, Moushumi Paul, Lili Xie, and Wilfred A. van der Donk. 2005.
'Biosynthesis and mode of action of lantibiotics', Chemical Reviews, 105: 633-84.
Christ, Nina A., Sophie Bochmann, Daniel Gottstein, Elke Duchardt-Ferner, Ute A. Hellmich,
Stefanie Düsterhus, Peter Kötter, Peter Güntert, Karl-Dieter Entian, and Jens Wöhnert.
2012. 'The first structure of a lantibiotic immunity protein, SpaI from Bacillus subtilis,
reveals a novel fold', J Biol Chem, 287: 35286-98.
Coelho, Marcus Lívio Varella, Janaína Santos Nascimento, Patrícia Carlin Fagundes, Danielle
Jannuzzi Madureira, Selma Soares Oliveira, Maria Aparecida Vasconcelos Paiva Brito,
and Maria do Carmo Freire Bastos. 2007. 'Activity of staphylococcal bacteriocins against
Staphylococcus aureus and Streptococcus agalactiae involved in bovine mastitis', Res
Microbiol, 158: 625-30.
Condas, Larissa A. Z., Jeroen De Buck, Diego B. Nobrega, Domonique A. Carson, Jean-Philippe
Roy, Greg P. Keefe, Trevor J. DeVries, John R. Middleton, Simon Dufour, and Herman
W. Barkema. 2017. 'Distribution of non-aureus staphylococci species in udder quarters
with low and high somatic cell count, and clinical mastitis', J Dairy Sci, 100: 5613-27.
Condas, Larissa, Jeroen De Buck, Diego Nobrega, Domonique Carson, Sohail Naushad, John
Kastelic, S. De Vliegher, R. N. Zadoks, John Middleton, S. Dufour, and H. W. Barkema.
2017 'Prevalence of non-aureus staphylococci isolated from milk samples in Canadian
dairy herds', J Dairy Sci: Forthcoming.
Corr, Sinéad C., Yin Li, Christian U. Riedel, Paul W. O'Toole, Colin Hill, and G. M. Gahan
Cormac. 2007. 'Bacteriocin production as a mechanism for the antiinfective activity of
Lactobacillus salivarius UCC118', Proceedings of the National Academy of Sciences of
the United States of America, 104: 7617-21.

121
Cotter, Paul D., Colin Hill, and R. Paul Ross. 2005. 'Bacteriocins: developing innate
immunity for food', Nat Rev Micro, 3: 777-88.
Cotter, Paul D., R. Paul Ross, and Colin Hill. 2013. 'Bacteriocins-a viable alternative to
antibiotics?', Nat Rev Microbiol, 11: 95-105.
Crispie, Fiona, Mercedes Alonso-Gómez, Collette O'Loughlin, Katja Klostermann, James Flynn,
Seán Arkins, William Meaney, R. Paul Ross, and Colin Hill. 2008. 'Intramammary
infusion of a live culture for treatment of bovine mastitis: effect of live lactococci on the
mammary immune response', J Dairy Res, 75: 374-84.
Crispie, Fiona, James Flynn, R. Paul Ross, Colin Hill, and William J. Meaney. 2004. 'Update on
the development of a novel dry cow therapy using a bismuth-based intramammary teat
seal in combination with the bacteriocin lacticin 3147', Ir Vet J, 57: 652-56.
Crispie, Fiona, Denis Twomey, James Flynn, Colin Hill, Paul Ross, and William Meaney. 2005.
'The lantibiotic lacticin 3147 produced in a milk-based medium improves the efficacy of
a bismuth-based teat seal in cattle deliberately infected with Staphylococcus aureus', J
Dairy Res, 72: 159-67.
Daly, Karen M., Mathew Upton, Stephanie K. Sandiford, Lorraine A. Draper, Philip A.
Wescombe, Ralph W. Jack, Paula M. O'Connor, Angela Rossney, Friedrich Götz, Colin
Hill, Paul D. Cotter, R. Paul Ross, and John R. Tagg. 2010. 'Production of the Bsa
lantibiotic by community-acquired Staphylococcus aureus strains', J Bacteriol, 192:
1131-42.
De Vliegher, S., H. Laevens, L. A. Devriese, G. Opsomer, J. L. M. Leroy, H. W. Barkema, and
A. de Kruif. 2003. 'Prepartum teat apex colonization with Staphylococcus chromogenes

122
in dairy heifers is associated with low somatic cell count in early lactation',
Veterinary Microbiology, 92: 245-52.
De Vliegher, S., G. Opsomer, A. Vanrolleghem, L. A. Devriese, O. C. Sampimon, J. Sol, H. W.
Barkema, F. Haesebrouck, and A. de Kruif. 2004. 'In vitro growth inhibition of major
mastitis pathogens by Staphylococcus chromogenes originating from teat apices of dairy
heifers', Vet Microbiol, 101: 215-21.
Delgado, Mónica A., María R. Rintoul, Ricardo N. Farías, and Raúl A. Salomón. 2001.
'Escherichia coli RNA polymerase is the target of the cyclopeptide antibiotic microcin
J25', J Bacteriol, 183: 4543-50.
Destoumieux-Garzón, Delphine, Sophie Duquesne, Jean Peduzzi, Christophe Goulard, Michel
Desmadril, Lucienne Letellier, Sylvie Rebuffat, and Pascale Boulanger. 2005. 'The iron–
siderophore transporter FhuA is the receptor for the antimicrobial peptide microcin J25:
role of the microcin Val(11)–Pro(16) β-hairpin region in the recognition mechanism',
Biochem J, 389: 869-76.
Di Cera, Enrico. 2009. 'Serine Proteases', IUBMB Life, 61: 510-15.
Diep, Dzung B., Morten Skaugen, Zhian Salehian, Helge Holo, and Ingolf F. Nes. 2007.
'Common mechanisms of target cell recognition and immunity for class II bacteriocins',
Proceedings of the National Academy of Sciences of the United States of America, 104:
2384-89.
Dischinger, Jasmin, Shradha Basi Chipalu, and Gabriele Bierbaum. 2014. 'Lantibiotics:
promising candidates for future applications in health care', Int J Med Microbiol, 304: 51.

123
Dischinger, Jasmin, Michaele Josten, Christiane Szekat, Hans-Georg Sahl, and Gabriele
Bierbaum. 2009. 'Production of the novel two-peptide lantibiotic Lichenicidin by Bacillus
licheniformis DSM 13', PloS one, 4: e6788.
Dos Santos, Danielle Cabral, Carla Christine Lange, Pedro Avellar-Costa, Katia Regina Netto
Dos Santos, Maria Aparecida Vasconcelos Paiva Brito, and Marcia Giambiagi-deMarval.
2016. 'Staphylococcus chromogenes, a coagulase-negative staphylococcus species that
can clot plasma', J Clin Microbiol, 54: 1372.
Draper, Lorraine A., Paul D. Cotter, Colin Hill, and R. Paul Ross. 2015. 'Lantibiotic resistance',
Microbiol Mol Biol Rev, 79: 171-91.
Draper, Lorraine A., Karen Grainger, Lucy H. Deegan, Paul D. Cotter, Colin Hill, and R. Paul
Ross. 2009. 'Cross-immunity and immune mimicry as mechanisms of resistance to the
lantibiotic lacticin 3147', Mol Microbiol, 71: 1043-54.
Duarte, Andreza Freitas de Souza, Hilana Ceotto-Vigoder, Emile Santos Barrias, Thaïs Cristina
Baeta Soares Souto-Padrón, Ingolf Figved Nes, and Maria do Carmo de Freire Bastos.
2017. 'Hyicin 4244, the first sactibiotic described in staphylococci, exhibits an anti-
staphylococcal biofilm activity', Int J Antimicrob Agents, In Press.
Edgar, Robert C. 2004. 'MUSCLE: a multiple sequence alignment method with reduced time and
space complexity', BMC bioinformatics, 5: 113-13.
———. 2010. 'Search and clustering orders of magnitude faster than BLAST', Bioinformatics,
26: 2460-61.
Eijsink, Vincent G. H., Marianne Skeie, P. Hans Middelhoven, May Bente Brurberg, and Ingolf
F. Nes. 1998. 'Comparative studies of class IIa bacteriocins of lactic acid bacteria', Appl
and Environ Microbiol, 64: 3275-81.

124
Ekkelenkamp, Miquel B., Micha Hanssen, Shang-Te Danny Hsu, Ad de Jong, Dana
Milatovic, Jan Verhoef, and Nico A. J. van Nuland. 2005. 'Isolation and structural
characterization of epilancin 15X, a novel lantibiotic from a clinical strain of
Staphylococcus epidermidis', FEBS Lett, 579: 1917-22.
Elsayed, Khaled M., Mohammad R. Islam, Mahin Abdullah Al, Jun-ichi Nagao, Takeshi Zendo,
and Kenji Sonomoto. 2017. 'LiaRS reporter assay: A simple tool to identify lipid II
binding moieties in lantibiotic nukacin ISK-1', J Biosci Bioeng, 123: 398-401.
Espeche, M. C., M. S. J. Tomas, B. Wiese, E. Bru, and M. E. F. Nader-Macias. 2014.
'Physicochemical factors differentially affect the biomass and bacteriocin production by
bovine Enterococcus mundtii CRL1656', J Dairy Sci, 97: 789-97.
Fagundes, Patrícia Carlin, Hilana Ceotto, Amina Potter, Maria Aparecida Vasconcelos de Paiva
Brito, Dag Brede, Ingolf F. Nes, and Maria do Carmo de Freire Bastos. 2011. 'Hyicin
3682, a bioactive peptide produced by Staphylococcus hyicus 3682 with potential
applications for food preservation', Res Microbiol, 162: 1052-59.
Fagundes, Patrícia Carlin, Ilana Nascimento de Sousa Santos, Márcia Silva Francisco, Rodolpho
Mattos Albano, and Maria do Carmo de Freire Bastos. 2017. 'Genetic and biochemical
characterization of hyicin 3682, the first bacteriocin reported for Staphylococcus hyicus',
Microbiol Res.
Fleming, H. P., J. L. Etchells, and R. N. Costilow. 1975. 'Microbial Inhibition by an Isolate of
Pediococcus from cucumber brines', Appl Microbiol, 30: 1040-42.
Fluhe, Leif, and Mohamed A. Marahiel. 2013. 'Radical S-adenosylmethionine enzyme catalyzed
thioether bond formation in sactipeptide biosynthesis', Curr Opin Chem Biol, 17: 605-12.

125
Fontana, Mariana Buss Cezar, Maria do Carmo Freire de Bastos, and Adriano Brandelli.
2006. 'Bacteriocins pep5 and epidermin inhibit Staphylococcus epidermidis adhesion to
catheters', Curr Microbiol, 52: 350-53.
Furmanek, B., T. Kaczorowski, R. Bugalski, K. Bielawski, J. Bogdanowicz, Podhajska, and J. A.
1999. 'Identification, characterization and purification of the lantibiotic staphylococcin T,
a natural gallidermin variant', J Appl Microbiol, 87: 856-66.
Gavrish, Ekaterina, Clarissa S. Sit, Shugeng Cao, Olga Kandror, Amy Spoering, Aaron Peoples,
Losee Ling, Ashley Fetterman, Dallas Hughes, Anthony Bissell, Heather Torrey, Tatos
Akopian, Andreas Mueller, Slava Epstein, Alfred Goldberg, Jon Clardy, and Kim Lewis.
2014. 'Lassomycin, a ribosomally synthesized cyclic peptide, kills Mycobacterium
tuberculosis by targeting the ATP-dependent protease ClpC1P1P2', Chem Biol, 21: 509-
18.
Gong, Xiandi, Leah A. Martin-Visscher, Drew Nahirney, John C. Vederas, and Marek Duszyk.
2009. 'The circular bacteriocin, carnocyclin A, forms anion-selective channels in lipid
bilayers', Biochim Biophys Acta, 1788: 1797-803.
Goto, Yuki, Bo Li, Jan Claesen, Yanxiang Shi, Mervyn J. Bibb, and Wilfred A. van der Donk.
2010. 'Discovery of unique lanthionine synthetases reveals new mechanistic and
evolutionary insights', PLoS Biol, 8: e1000339.
Guinane, Caitriona M., Paul D. Cotter, Colin Hill, and R. Paul Ross. 2006. 'Spontaneous
resistance in Lactococcus lactis IL1403 to the lantibiotic lacticin 3147', FEMS Microbiol
Lett, 260: 77-83.
Gurevich, Alexey, Vladislav Saveliev, Nikolay Vyahhi, and Glenn Tesler. 2013. 'QUAST:
Quality assessment tool for genome assemblies', Bioinformatics, 29: 1072-75.

126
Gutiérrez-Chávez, A. J., E. A. Martínez-Ortega, M. Valencia-Posadas, M. F. León-Galván,
N. M. de la Fuente-Salcido, D. K. Bideshi, and J. E. Barboza-Corona. 2016. 'Potential use
of Bacillus thuringiensis bacteriocins to control antibiotic-resistant bacteria associated
with mastitis in dairy goats', Folia Microbiol, 61: 11-19.
Hacker, Carolin, Nina A. Christ, Elke Duchardt-Ferner, Sophie Korn, Christoph Göbl, Lucija
Berninger, Stefanie Düsterhus, Ute A. Hellmich, Tobias Madl, Peter Kötter, Karl-Dieter
Entian, and Jens Wöhnert. 2015. 'The solution structure of the lantibiotic immunity
protein NisI and its interactions with nisin', J Biol Chem, 290: 28869-86.
Haft, Daniel H., Malay Kumar Basu, and Douglas A. Mitchell. 2010. 'Expansion of ribosomally
produced natural products: a nitrile hydratase- and Nif11-related precursor family', BMC
Biol, 8: 70-70.
Hasper, Hester E., Naomi E. Kramer, James L. Smith, J. D. Hillman, Cherian Zachariah, Oscar
P. Kuipers, Ben de Kruijff, and Eefjan Breukink. 2006. 'An alternative bactericidal
mechanism of action for lantibiotic peptides that target lipid II', Science, 313: 1636-37.
Hasper, Hester Emilie, Ben de Kruijff, and Eefjan Breukink. 2004. 'Assembly and stability of
nisin−lipid II pores', Biochem, 43: 11567-75.
Hegemann, Julian D., Marcel Zimmermann, Xiulan Xie, and Mohamed A. Marahiel. 2015.
'Lasso peptides: an intriguing class of bacterial natural products', Acc Chem Res, 48:
1909-19.
Heidrich, Christoph, Ulrike Pag, Michaele Josten, Jörg Metzger, Ralph W. Jack, Gabriele
Bierbaum, Günther Jung, and Hans-Georg Sahl. 1998. 'Isolation, characterization, and
heterologous expression of the novel lantibiotic epicidin 280 and analysis of its
biosynthetic gene cluster', Appl and Environ Microbiol, 64: 3140-46.

127
Heng, Nicholas C. K., and John R. Tagg. 2006. 'What's in a name? Class distinction for
bacteriocins', Nat Rev Micro, 4.
Hoang, K. V., N. J. Stern, and J. Lin. 2011. 'Development and stability of bacteriocin resistance
in Campylobacter spp: Bacteriocin resistance in Campylobacter', J Appl Microbiol, 111:
1544-50.
Hogan, J. S., K. L. Smith, D. A. Todhunter, and P. S. Schoenberger. 1988. 'Rate of
environmental mastitis in quarters infected with Corynebacterium bovis and
Staphylococcus species', J Dairy Sci, 71: 2520-25.
Isaac, Paula, Luciana Paola Bohl, María Laura Breser, María Soledad Orellano, Agustín Conesa,
Marcela Alejandra Ferrero, and Carina Porporatto. 2017. 'Commensal coagulase-negative
Staphylococcus from the udder of healthy cows inhibits biofilm formation of mastitis-
related pathogens', Vet Microbiol, 207: 259-66.
Islam, Mohammad R., Jun-Ichi Nagao, Takeshi Zendo, and Kenji Sonomoto. 2012.
'Antimicrobial mechanism of lantibiotics', Biochem Soc Trans, 40: 1528.
Islam, Mohammad R., Mami Nishie, Jun-ichi Nagao, Takeshi Zendo, Sandro Keller, Jiro
Nakayama, Daisuke Kohda, Hans-Georg Sahl, and Kenji Sonomoto. 2012. 'Ring A of
nukacin ISK-1: a lipid II-binding motif for type-A(II) lantibiotic', J Am Chem Soc, 134:
3687.
Iwatsuki, Masato, Hiroshi Tomoda, Ryuji Uchida, Hiroaki Gouda, Shuichi Hirono, and Satoshi
Ōmura. 2006. 'Lariatins, antimycobacterial peptides produced by Rhodococcus sp.
K01−B0171, have a lasso structure', J Am Chem Soc, 128: 7486-91.
Kaškonienė, Vilma, Mantas Stankevičius, Kristina Bimbiraitė-Survilienė, Gintarė Naujokaitytė,
Loreta Šernienė, Kristina Mulkytė, Mindaugas Malakauskas, and Audrius Maruška.

128
2017. 'Current state of purification, isolation and analysis of bacteriocins produced by
lactic acid bacteria', Appl Microbiol Biotechnol, 101: 1323-35.
Katoh, Kazutaka, and Daron M. Standley. 2013. 'MAFFT Multiple Sequence Alignment
Software Version 7: Improvements in performance and usability', Mol Biol Evol, 30: 772-
80.
Kearse, Matthew, Richard Moir, Amy Wilson, Steven Stones-Havas, Matthew Cheung, Shane
Sturrock, Simon Buxton, Alex Cooper, Sidney Markowitz, Chris Duran, Tobias Thierer,
Bruce Ashton, Peter Meintjes, and Alexei Drummond. 2012. 'Geneious Basic: An
integrated and extendable desktop software platform for the organization and analysis of
sequence data', Bioinformatics, 28: 1647-49.
Kim, S. Y., S. Shin, H. C. Koo, J. H. Youn, H. D. Paik, and Y. H. Park. 2010. 'In vitro
antimicrobial effect and in vivo preventive and therapeutic effects of partially purified
lantibiotic lacticin NK34 against infection by Staphylococcus species isolated from
bovine mastitis', J Dairy Sci, 93: 3610-15.
Kjos, Morten, Juan Borrero, Mona Opsata, Dagim J. Birri, Helge Holo, Luis M. Cintas, Lars
Snipen, Pablo E. Hernández, Ingolf F. Nes, and Dzung B. Diep. 2011. 'Target
recognition, resistance, immunity and genome mining of class II bacteriocins from Gram-
positive bacteria', Microbiology, 157: 3256-67.
Kjos, Morten, Ingolf F. Nes, and Dzung B. Diep. 2009. 'Class II one-peptide bacteriocins target a
phylogenetically defined subgroup of mannose phosphotransferase systems on sensitive
cells', Microbiology, 155: 2949.
Kjos, Morten, Camilla Oppegård, Dzung B. Diep, Ingolf F. Nes, Jan-Willem Veening, Jon
Nissen-Meyer, and Tom Kristensen. 2014. 'Sensitivity to the two-peptide bacteriocin

129
lactococcin G is dependent on UppP, an enzyme involved in cell-wall synthesis', Mol
Microbiol, 92: 1177-87.
Kjos, Morten, Lars Snipen, Zhian Salehian, Ingolf F. Nes, and Dzung B. Diep. 2010. 'The Abi
proteins and their involvement in bacteriocin self-immunity', J Bacteriol, 192: 2068-76.
Klaenhammer, Todd R. 1993. 'Genetics of bacteriocins produced by lactic acid bacteria', FEMS
Microbiol Rev, 12: 39-85.
Klostermann, Katja, Fiona Crispie, James Flynn, R. Paul Ross, Colin Hill, and William Meaney.
2008. 'Intramammary infusion of a live culture of Lactococcus lactis for treatment of
bovine mastitis: comparison with antibiotic treatment in field trials', J Dairy Res, 75: 365-
73.
Klostermann, Katja, Fiona Crispie, Jim Flynn, William J. Meaney, R. Paul Ross, and Colin Hill.
2010. 'Efficacy of a teat dip containing the bacteriocin lacticin 3147 to eliminate Gram-
positive pathogens associated with bovine mastitis', J Dairy Res, 77: 231-38.
Knappe, Thomas A., Uwe Linne, Séverine Zirah, Sylvie Rebuffat, Xiulan Xie, and Mohamed A.
Marahiel. 2008. 'Isolation and structural characterization of capistruin, a lasso peptide
predicted from the genome sequence of Burkholderia thailandensis E264', J Am Chem
Soc, 130: 11446-54.
Knerr, Patrick J., Trent J. Oman, Chantal V. Garcia De Gonzalo, Tania J. Lupoli, Suzanne
Walker, and Wilfred A. van der Donk. 2012. 'Non-proteinogenic amino acids in lacticin
481 analogues result in more potent inhibition of peptidoglycan transglycosylation', ACS
Chem Biol, 7: 1791.
Knerr, Patrick J., and Wilfred A. van der Donk. 2012. 'Discovery, biosynthesis, and engineering
of lantipeptides', Annu Rev Biochem, 81: 479-505.

130
Köster, Johannes, and Sven Rahmann. 2012. 'Snakemake—a scalable bioinformatics
workflow engine', Bioinformatics, 28: 2520-22.
Kramer, Naomi E., A. F. T. van Hijum Sacha, Jan Knol, Jan Kok, and Oscar P. Kuipers. 2006.
'Transcriptome analysis reveals mechanisms by which Lactococcus lactis acquires nisin
resistance', Antimicrob Agents Chemother, 50: 1753-61.
Kruszewska, Danuta, Hans-Georg Sahl, Gabriele Bierbaum, Ulrike Pag, Sean O. Hynes, and Asa
Ljungh. 2004. 'Mersacidin eradicates methicillin-resistant Staphylococcus aureus
(MRSA) in a mouse rhinitis model', J Antimicrob Chemother, 54: 648-53.
Kuijk, van S. J. A., K. S. Noll, and M. L. Chikindas. 2012. 'The species-specific mode of action
of the antimicrobial peptide subtilosin against Listeria monocytogenes Scott A', Lett Appl
Microbiol, 54: 52-58.
Kuwano, Koichi, Noriko Tanaka, Takashi Shimizu, Kohei Nagatoshi, Shinsuke Nou, and Kenji
Sonomoto. 2005. 'Dual antibacterial mechanisms of nisin Z against Gram-positive and
Gram-negative bacteria', J Antimicrob Agents, 26: 396-402.
Kuznedelov, Konstantin, Ekaterina Semenova, Thomas A. Knappe, Damir Mukhamedjarov,
Aashish Srivastava, Sujoy Chatterjee, Richard H. Ebright, Mohamed Marahiel, and
Konstantin Severinov. 2011. 'The antibacterial threaded-lasso peptide capistruin inhibits
bacterial RNA polymerase', J Mol Biol, 412: 842-48.
Lacroix, Christophe, Marilaine Mota-Meira, Marc C. Lavoie, Hélène Morency, and Gisèle
LaPointe. 2001. 'Comparison of the activity spectra against pathogens of bacterial strains
producing a mutacin or a lantibiotic', Can J Microbiol, 47: 322-31.
Leon-Galvan, Ma Fabiola, Jose E. Barboza-Corona, A. Arianna Lechuga-Arana, Mauricio
Valencia-Posadas, Daniel D. Aguayo, Carlos Cedillo-Pelaez, Erika A. Martinez-Ortega,

131
and Abner J. Gutierrez-Chavez. 2015. 'Molecular detection and sensitivity to
antibiotics and bacteriocins of pathogens isolated from bovine mastitis in family dairy
herds of central mexico', BioMed Res Int, 2015: 1-9.
Leroy, Frédéric, and Luc De Vuyst. 2004. 'Lactic acid bacteria as functional starter cultures for
the food fermentation industry', Trends Food Sci Technol, 15: 67-78.
Letzel, Anne-Catrin, Sacha J. Pidot, and Christian Hertweck. 2014. 'Genome mining for
ribosomally synthesized and post-translationally modified peptides (RiPPs) in anaerobic
bacteria', BMC Genomics, 15: 983.
Li, Heng, and Richard Durbin. 2009. 'Fast and accurate short read alignment with Burrows–
Wheeler transform', Bioinformatics, 25: 1754-60.
Lubelski, J., R. Rink, R. Khusainov, G. N. Moll, and O. P. Kuipers. 2008. 'Biosynthesis,
immunity, regulation, mode of action and engineering of the model lantibiotic nisin', Cell
Mol Life Sci, 65: 455-76.
Maduwe, A. D. B. Navaratna, Hans-Georg Sahl, and John R. Tagg. 1999. 'Identification of genes
encoding two-component lantibiotic production in Staphylococcus aureus C55 and other
phage group II S. aureus Ssrains and demonstration of an association with the exfoliative
toxin B gene', Infect Immun, 67: 4268-71.
Maksimov, Mikhail O., and A. James Link. 2013. 'Discovery and characterization of an
isopeptidase that linearizes lasso peptides', J Am Chem Soc, 135: 120-38.
———. 2014. 'Prospecting genomes for lasso peptides', J Ind Microbiol Biotechnol, 41: 333-44.
Maksimov, Mikhail O., István Pelczer, and A. James Link. 2012. 'Precursor-centric genome-
mining approach for lasso peptide discovery', Proceedings of the National Academy of
Sciences of the United States of America, 109: 15223-28.

132
Maqueda, Mercedes, Marina Sánchez-Hidalgo, Matilde Fernández, Manuel Montalbán-
López, Eva Valdivia, and Manuel Martínez-Bueno. 2008. 'Genetic features of circular
bacteriocins produced by Gram-positive bacteria', FEMS Microbiol Rev, 32: 2-22.
Marchler-Bauer, Aron, Yu Bo, Lianyi Han, Jane He, Christopher J. Lanczycki, Shennan Lu,
Farideh Chitsaz, Myra K. Derbyshire, Renata C. Geer, Noreen R. Gonzales, Marc
Gwadz, David I. Hurwitz, Fu Lu, Gabriele H. Marchler, James S. Song, Narmada Thanki,
Zhouxi Wang, Roxanne A. Yamashita, Dachuan Zhang, Chanjuan Zheng, Lewis Y. Geer,
and Stephen H. Bryant. 2017. 'CDD/SPARCLE: functional classification of proteins via
subfamily domain architectures', Nucleic Acids Res, 45: D200-D03.
Marsh, Alan J., Orla O'Sullivan, R. Paul Ross, Paul D. Cotter, and Colin Hill. 2010. 'In silico
analysis highlights the frequency and diversity of type 1 lantibiotic gene clusters in
genome sequenced bacteria', BMC Genomics, 11: 679-700.
Martin, Marcel. 2011. 'Cutadapt removes adapter sequences from high-throughput sequencing
reads', EMBnet .journal, 17: 10-12.
Martı́nez, Beatriz, Marı́a Fernández, Juan E. Suárez, and Ana Rodrı́guez. 1999. 'Synthesis of
lactococcin 972, a bacteriocin produced by Lactococcus lactis IPLA 972, depends on the
expression of a plasmid-encoded bicistronic operon', Microbiology, 145: 3155-61.
Martínez-Bueno, M., M. Maqueda, A. Gálvez, B. Samyn, J. Van Beeumen, J. Coyette, and E.
Valdivia. 1994. 'Determination of the gene sequence and the molecular structure of the
enterococcal peptide antibiotic AS-48', J Bacteriol, 176: 6334-39.
Matthews, K. R., R. J. Harmon, and B. A. Smith. 1990. 'Protective effect of Staphylococcus
chromogenes infection against Staphylococcus aureus infection in the lactating bovine
mammary gland', J Dairy Sci, 73: 3457-62.

133
Mattick, A. T. R., A. Hirsch, and N. J. Berridge. 1947. 'Further observations on an inhibotory
substance (nisin) from lactic streptococci', Lancet, 250: 5-8.
McClerren, Amanda L., Lisa E. Cooper, Chao Quan, Paul M. Thomas, Neil L. Kelleher, and
Wilfred A. van der Donk. 2006. 'Discovery and in vitro biosynthesis of haloduracin, a
two-component lantibiotic', Proc Natl Acad Sci U S A, 103: 17243-48.
Murphy, Kiera, Orla O'Sullivan, Mary C. Rea, Paul D. Cotter, R. Paul Ross, and Colin Hill.
2011. 'Genome mining for radical SAM protein determinants reveals multiple sactibiotic-
like gene clusters', PloS one, 6: e20852.
Nascimento, J. S., H. Ceotto, S. B. Nascimento, M. Giambiagi-deMarval, K. R. N. Santos, and
M. C. F. Bastos. 2006. 'Bacteriocins as alternative agents for control of multiresistant
staphylococcal strains', Lett Appl Microbiol, 42: 215-21.
Nascimento, Janaína Santos, Janaína Abrantes, Marcia Giambiagi-deMarval, and Maria Carmo
Freire Bastos. 2004. 'Growth conditions required for bacteriocin production by strains of
Staphylococcus aureus', World J Microbiol Biotechnol, 20: 941-47.
Nascimento, Janaína Santos, Patricia Carlin Fagundes, Maria Aparecida Vasconcelos Paiva
Brito, Kátia Regina Netto Santos, and Maria Carmo Freire Bastos. 2005. 'Production of
bacteriocins by coagulase-negative staphylococci involved in bovine mastitis', Vet
Microbiol, 106: 61-71.
Naushad, Sohail, Herman Barkema, Christopher Luby, Larissa Condas, Diego Nobrega,
Domonique Carson, and Jeroen De Buck. 2016. 'Comprehensive phylogenetic analysis of
bovine non-aureus staphylococci species based on whole-genome sequencing', Front
Microbiol, 7.

134
Nissen-Meyer, J., H. Holo, L. S. Håvarstein, K. Sletten, and I. F. Nes. 1992. 'A novel
lactococcal bacteriocin whose activity depends on the complementary action of two
peptides', J Bacteriol, 174: 5686-92.
Nurk, Sergey, Anton Bankevich, Dmitry Antipov, Alexey A. Gurevich, Anton Korobeynikov,
Alla Lapidus, Andrey D. Prjibelski, Alexey Pyshkin, Alexander Sirotkin, Yakov Sirotkin,
Ramunas Stepanauskas, Scott R. Clingenpeel, Tanja Woyke, Jeffrey S. McLean, Roger
Lasken, Glenn Tesler, Max A. Alekseyev, and Pavel A. Pevzner. 2013. 'Assembling
single-cell genomes and mini-metagenomes from chimeric MDA products', J Comput
Biol, 20: 714-37.
Olde Riekerink, R. G. M., H. W. Barkema, D. F. Kelton, and D. T. Scholl. 2008. 'Incidence Rate
of Clinical Mastitis on Canadian Dairy Farms', J Dairy Sci, 91: 1366-77.
Olde Riekerink, Richard G. M., Herman W. Barkema, Stefan Veenstra, Doris E. Poole, Randy T.
Dingwell, and Gregory P. Keefe. 2006. 'Prevalence of contagious mastitis pathogens in
bulk tank milk in Prince Edward Island', Can Vet J, 47: 567-72.
Oliveira, L., and P. L. Ruegg. 2014. 'Treatments of clinical mastitis occurring in cows on 51
large dairy herds in Wisconsin', J Dairy Sci, 97: 5426-36.
Ozer, Egon A., Jonathan P. Allen, and Alan R. Hauser. 2014. 'Characterization of the core and
accessory genomes of Pseudomonas aeruginosa using bioinformatic tools Spine and
AGEnt', BMC Genomics, 15: 737.
Pan, Si Jia, and A. James Link. 2011. 'Sequence diversity in the lasso peptide framework:
discovery of functional microcin J25 variants with multiple amino acid substitutions', J
Am Chem Soc, 133: 5016-23.

135
Parada, José Luis, Carolina Ricoy Caron, Adriane Bianchi P. Medeiros, and Carlos Ricardo
Soccol. 2007. 'Bacteriocins from lactic acid bacteria: purification, properties and use as
biopreservatives', Braz Arch Biol Technol, 50: 512-42.
Piepers, S., L. De Meulemeester, A. de Kruif, G. Opsomer, H. W. Barkema, and S. De Vliegher.
2007. 'Prevalence and distribution of mastitis pathogens in subclinically infected dairy
cows in Flanders, Belgium', J Dairy Res, 74: 478-83.
Piepers, S., G. Opsomer, H. W. Barkema, A. de Kruif, and S. De Vliegher. 2010. 'Heifers
infected with coagulase-negative staphylococci in early lactation have fewer cases of
clinical mastitis and higher milk production in their first lactation than noninfected
heifers', J Dairy Sci, 93: 2014-24.
Pieterse, Reneé, and Svetoslav D. Todorov. 2010. 'Bacteriocins: exploring alternatives to
antibiotics in mastitis treatment', Braz J Microbiol, 41: 542-62.
Pitkälä, A., M. Haveri, S. Pyörälä, V. Myllys, and T. Honkanen-Buzalski. 2004. 'Bovine mastitis
in Finland 2001—Prevalence, distribution of bacteria, and antimicrobial resistance', J
Dairy Sci, 87: 2433-41.
Potter, A., H. Ceotto, M. L. V. Coelho, A. J. Guimaraes, and M. D. D. Bastos. 2014. 'The gene
cluster of aureocyclicin 4185: the first cyclic bacteriocin of Staphylococcus aureus',
Microbiology, 160: 917-28.
Potter, Amina, Hilana Ceotto, Marcus Lívio Varella Coelho, Allan J. Guimarães, and Maria do
Carmo de Freire Bastos. 2014. 'The gene cluster of aureocyclicin 4185: the first cyclic
bacteriocin of Staphylococcus aureus', Microbiology, 160: 917-28.
Price, Morgan N., Paramvir S. Dehal, and Adam P. Arkin. 2010. 'FastTree 2 – Approximately
maximum-likelihood trees for large alignments', PloS one, 5: e9490.

136
Priyam, Anurag, Ben J Woodcroft, Vivek Rai, Alekhya Munagala, Ismail Moghul, Filip Ter,
Mark Anthony Gibbins, HongKee Moon, Guy Leonard, Wolfgang Rumpf, and Yannick
Wurm. 2015. 'Sequenceserver: a modern graphical user interface for custom BLAST
databases', bioRxiv.
Rea, Mary C., Clarissa S. Sit, Evelyn Clayton, Paula M. O'Connor, Randy M. Whittal, Jing
Zheng, John C. Vederas, R. Paul Ross, and Colin Hill. 2010. 'Thuricin CD, a
posttranslationally modified bacteriocin with a narrow spectrum of activity against
Clostridium difficile', Proceedings of the National Academy of Sciences of the United
States of America, 107: 9352-57.
Reyher, S. Dufour, H. W. Barkema, L. Des Côteaux, T. J. DeVries, I. R. Dohoo, G. P. Keefe, J.
P. Roy, and D. T. Scholl. 2011. 'The National Cohort of Dairy Farms—A data collection
platform for mastitis research in Canada', J Dairy Sci, 94: 1616-26.
Riley, Margaret A., and John E. Wertz. 2002. 'Bacteriocins: Evolution, ecology, and application',
Annu Rev Microbiol, 56: 117-37.
Rink, R., A. Arkema-Meter, I. Baudoin, E. Post, A. Kuipers, S. A. Nelemans, M. Haas Jimoh
Akanbi, and G. N. Moll. 2010. 'To protect peptide pharmaceuticals against peptidases', J
Pharmacol Toxicol Methods, 61: 210-18.
Rogne, Per, Gunnar Fimland, Jon Nissen-Meyer, and Per Eugen Kristiansen. 2008. 'Three-
dimensional structure of the two peptides that constitute the two-peptide bacteriocin
lactococcin G', Biochim Biophys Acta, 1784: 543-54.
Rollin, E., K. C. Dhuyvetter, and M. W. Overton. 2015. 'The cost of clinical mastitis in the first
30 days of lactation: An economic modeling tool', Prev Vet Med, 122: 257.

137
Ryan, M. P., J. Flynn, C. Hill, R. P. Ross, and W. J. Meaney. 1999. 'The natural food grade
inhibitor, lacticin 3147, reduced the incidence of mastitis after experimental challenge
with Streptococcus dysgalactiae in nonlactating dairy cows', J Dairy Sci, 82: 2108-14.
Ryan, Máire P., William J. Meaney, R. Paul Ross, and Colin Hill. 1998. 'Evaluation of lacticin
3147 and a teat seal containing this bacteriocin for inhibition of mastitis pathogens', Appl
and Environ Microbiol, 64: 2287-90.
Sahl, H. G., M. Grossgarten, W. R. Widger, W. A. Cramer, and H. Brandis. 1985. 'Structural
similarities of the staphylococcin-like peptide Pep-5 to the peptide antibiotic nisin',
Antimicrob Agents Chemother, 27: 836-40.
Saini, V., J. T. McClure, D. Léger, S. Dufour, A. G. Sheldon, D. T. Scholl, and H. W. Barkema.
2012. 'Antimicrobial use on Canadian dairy farms', J Dairy Sci, 95: 1209-21.
Salomón, R. A., and R. N. Farías. 1992. 'Microcin 25, a novel antimicrobial peptide produced by
Escherichia coli', J Bacteriol, 174: 7428-35.
Sampimon, O. C., H. W. Barkema, I. M. G. A. Berends, J. Sol, and T. J. G. M. Lam. 2009.
'Prevalence and herd-level risk factors for intramammary infection with coagulase-
negative staphylococci in Dutch dairy herds', Vet Microbiol, 134: 37-44.
Samyn, Bart, Manuel Martinez-Bueno, Bart Devreese, Mercedes Maqueda, Antonio Gálvez, Eva
Valdivia, Jacques Coyette, and Jozef Van Beeumen. 1994. 'The cyclic structure of the
enterococcal peptide antibiotic AS-48', FEBS Lett, 352: 87-90.
Sashihara, Toshihiro, Sashihara Toshihiro, Kimura Hirokazu, Higuchi Toshimasa, and Adachi
Asaho. 2000. 'A novel lantibiotic, Nukacin ISK-1, of Staphylococcus warneri ISK-1',
Biosci Biotechnol Biochem, 64: 2420.

138
Scherpenzeel, C. G. M., I. E. M. den Uijl, G. van Schaik, R. G. M. Olde Riekerink, J. M.
Keurentjes, and T. J. G. M. Lam. 2014. 'Evaluation of the use of dry cow antibiotics in
low somatic cell count cows', J Dairy Sci, 97: 3606-14.
Schillinger, U., and F. K. Lücke. 1989. 'Antibacterial activity of Lactobacillus sake isolated from
meat', Appl and Environ Microbiol, 55: 1901-06.
Schnell, Norbert, Karl-Dieter Entian, Friedrich Götz, Thomas Hörner, Roland Kellner, and
Günther Jung. 1989. 'Structural gene isolation and prepeptide sequence of gallidermin, a
new lanthionine containing antibiotic', FEMS Microbiol Lett, 58: 263-67.
Schukken, Y. H., R. N. Gonzalez, L. L. Tikofsky, H. F. Schulte, C. G. Santisteban, F. L.
Welcome, G. J. Bennett, M. J. Zurakowski, and R. N. Zadoks. 2009. 'CNS mastitis:
nothing to worry about?', Vet Microbiol, 134: 9-14.
Sears, P. M., B. S. Smith, W. K. Stewart, R. N. Gonzalez, S. D. Rubino, S. A. Gusik, E. S.
Kulisek, S. J. Projan, and P. Blackburn. 1992. 'Evaluation of a nisin-based germicidal
formulation on teat skin of live cows', J Dairy Sci, 75: 3185-90.
Sedgley, Christine M., Don B. Clewell, and Susan E. Flannagan. 2009. 'Plasmid pAMS1-
Encoded, Bacteriocin-Related “Siblicide” in Enterococcus faecalis', J Bacteriol, 191:
3183.
Seemann, Torsten. 2014. 'Prokka: rapid prokaryotic genome annotation', Bioinformatics, 30:
2068–69.
Segata, Nicola, Daniela Börnigen, Xochitl C. Morgan, and Curtis Huttenhower. 2013.
'PhyloPhlAn is a new method for improved phylogenetic and taxonomic placement of
microbes', Nat Commun, 4: 2304.

139
Shelburne, Charles E., Florence Y. An, Vishnu Dholpe, Ayyalusamy Ramamoorthy, Dennis
E. Lopatin, and Marilyn S. Lantz. 2007. 'The spectrum of antimicrobial activity of the
bacteriocin subtilosin A', J Antimicrob Chemother, 59: 297-300.
Shin, J. M., J. W. Gwak, P. Kamarajan, J. C. Fenno, A. H. Rickard, and Y. L. Kapila. 2016.
'Biomedical applications of nisin', J Appl Microbiol, 120: 1449-65.
Shin, Jae M., Islam Ateia, Jefrey R. Paulus, Hongrui Liu, J. Christopher Fenno, Alexander H.
Rickard, and Yvonne L. Kapila. 2015. 'Antimicrobial nisin acts against saliva derived
multi-species biofilms without cytotoxicity to human oral cells', Front Microbiol, 6: 617.
Singh, M., and D. Sareen. 2014. 'Novel LanT associated lantibiotic clusters identified by genome
database mining', PloS one, 9: e91352.
Singh, Neetigyata Pratap, Abhay Tiwari, Ankiti Bansal, Shruti Thakur, Garima Sharma, and
Reema Gabrani. 2015. 'Genome level analysis of bacteriocins of lactic acid bacteria',
Comput Biol Chem, 56: 1-6.
Speksnijder, D. C., D. J. Mevius, C. J. M. Bruschke, and J. A. Wagenaar. 2015. 'Reduction of
Veterinary Antimicrobial Use in the Netherlands. The Dutch Success Model', Zoonoses
Public Health, 62: 79-87.
Srinivas, Suda, D. Cotter Paul, Hill Colin, and R. Paul Ross. 2012. 'Lacticin 3147 - Biosynthesis,
molecular analysis, immunity, bioengineering and applications', Curr Protein Pept Sci,
13: 193-204.
Stein, Torsten, Stefan Heinzmann, Stefanie Düsterhus, Stefan Borchert, and Karl-Dieter Entian.
2005. 'Expression and functional analysis of the subtilin immunity genes spaIFEG in the
subtilin-sensitive host Bacillus subtilis MO1099', J Bacteriol, 187: 822-28.

140
Stein, Torsten, Stefan Heinzmann, Irina Solovieva, and Karl-Dieter Entian. 2003. 'Function
of Lactococcus lactis nisin immunity senes nisI and nisFEG after coordinated expression
in the surrogate host Bacillus subtilis', J Biol Chem, 278: 89-94.
Stoddard, G. W., J. P. Petzel, M. J. van Belkum, J. Kok, and L. L. McKay. 1992. 'Molecular
analyses of the lactococcin A gene cluster from Lactococcus lactis subsp. lactis biovar
diacetylactis WM4', Appl and Environ Microbiol, 58: 1952-61.
Sun, Zhizeng, Jin Zhong, Xiaobo Liang, Jiale Liu, Xiuzhu Chen, and Liandong Huan. 2009.
'Novel mechanism for nisin resistance via proteolytic degradation of nisin by the nisin
resistance protein NSR', Antimicrob Agents Chemother, 53: 1964-73.
Sutyak Noll, Katia, Patrick J. Sinko, and Michael L. Chikindas. 2011. 'Elucidation of the
molecular mechanisms of action of the natural antimicrobial peptide subtilosin against
the bacterial vaginosis-associated pathogen Gardnerella vaginalis', Probiotics Antimicrob
Proteins, 3: 41-47.
Tamura, Koichiro, Glen Stecher, Daniel Peterson, Alan Filipski, and Sudhir Kumar. 2013.
'MEGA6: Molecular evolutionary genetics analysis version 6.0', Mol Biol Evol, 30: 2725-
29.
Taponen, Suvi, Heli Simojoki, Maarit Haveri, Helle D. Larsen, and Satu Pyörälä. 2006. 'Clinical
characteristics and persistence of bovine mastitis caused by different species of
coagulase-negative staphylococci identified with API or AFLP', Veterinary
Microbiology, 115: 199-207.
Thennarasu, Sathiah, Dong-Kuk Lee, Alan Poon, Karen E. Kawulka, John C. Vederas, and
Ayyalusamy Ramamoorthy. 2005. 'Membrane permeabilization, orientation, and
antimicrobial mechanism of subtilosin A', Chem Phys Lipids, 137: 38-51.

141
Twomey, D. P., A. I. Wheelock, J. Flynn, W. J. Meaney, C. Hill, and R. P. Ross. 2000.
'Protection against Staphylococcus aureus mastitis in dairy cows using a bismuth-based
teat seal containing the bacteriocin, lacticin 3147', J Dairy Sci, 83: 1981-88.
van Belkum, Marco J., Leah A. Martin-Visscher, and John C. Vederas. 2011. 'Structure and
genetics of circular bacteriocins', Trends Microbiol, 19: 411-18.
van de Kamp, M., H. W. van den Hooven, R. N. Konings, G. Bierbaum, H. G. Sahl, O. P.
Kuipers, R. J. Siezen, W. M. de Vos, C. W. Hilbers, and F. J. van de Ven. 1995.
'Elucidation of the primary structure of the lantibiotic epilancin K7 from Staphylococcus
epidermidis K7. Cloning and characterisation of the epilancin-K7-encoding gene and
NMR analysis of mature epilancin K7', Eur J Biochem, 230: 587-600.
van Heel, Auke J., Anne de Jong, Manuel Montalbán-López, Jan Kok, and Oscar P. Kuipers.
2013. 'BAGEL3: automated identification of genes encoding bacteriocins and
(non)bactericidal posttranslationally modified peptides', Nucleic Acids Res, 41: W448-
W53.
van Heel, Auke Johan, Manuel Montalban-Lopez, and Oscar P. Kuipers. 2011. 'Evaluating the
feasibility of lantibiotics as an alternative therapy against bacterial infections in humans',
Expert Opin Drug Metab Toxicol, 7: 675-80.
Vanderhaeghen, Wannes, Sofie Piepers, Frédéric Leroy, Els Van Coillie, Freddy Haesebrouck,
and Sarne De Vliegher. 2015. 'Identification, typing, ecology and epidemiology of
coagulase negative staphylococci associated with ruminants', Vet J, 203: 44-51.
Walsh, Calum J., Caitriona M. Guinane, Colin Hill, R. Paul Ross, Paul W. O'Toole, and Paul D.
Cotter. 2015. 'In silico identification of bacteriocin gene clusters in the gastrointestinal

142
tract, based on the Human Microbiome Project's reference genome database', BMC
Microbiol, 15: 183.
Wang, Gaoyan, Guoping Feng, Abigail B. Snyder, David C. Manns, John J. Churey, and Randy
W. Worobo. 2014. 'Bactericidal thurincin H causes unique morphological changes in
Bacillus cereus F4552 without affecting membrane permeability', FEMS Microbiol Lett,
357: 69-76.
Wang, Hao, David P. Fewer, and Kaarina Sivonen. 2011. 'Genome mining demonstrates the
widespread occurrence of gene clusters encoding bacteriocins in cyanobacteria', PloS
one, 6: e22384.
Waterhouse, Andrew M., James B. Procter, David M. A. Martin, Michèle Clamp, and Geoffrey
J. Barton. 2009. 'Jalview Version 2—a multiple sequence alignment editor and analysis
workbench', Bioinformatics, 25: 1189-91.
Weber, T., K. Blin, S. Duddela, D. Krug, H. U. Kim, R. Bruccoleri, S. Y. Lee, M. A. Fischbach,
R. Müller, W. Wohlleben, R. Breitling, E. Takano, and M. H. Medema. 2015.
'antiSMASH 3.0—a comprehensive resource for the genome mining of biosynthetic gene
clusters', Nucleic Acids Res, 43: W237-W43.
Whelan, Simon, and Nick Goldman. 2001. 'A general empirical model of protein evolution
derived from multiple protein families using a maximum-likelihood approach', Mol Biol
Evol, 18: 691-99.
White, D. G., R. J. Harmon, J. E. S. Matos, and B. E. Langlois. 1989. 'Isolation and identification
of coagulase-negative staphylococcus species from bovine body sites and streak canals of
nulliparous heifers', J Dairy Sci, 72: 1886-92.

143
Wilaipun, Pongtep, Takeshi Zendo, Ken-ichi Okuda, Jiro Nakayama, and Kenji Sonomoto.
2008. 'Identification of the nukacin KQU-131, a new type-A(II) lantibiotic produced by
Staphylococcus hominis KQU-131 isolated from thai fermented fish product (Pla-ra)',
Biosci Biotechnol Biochem, 72: 2232-35.
Zadoks, R. N., H. G. Allore, H. W. Barkema, O. C. Sampimon, G. J. Wellenberg, Y. T. Gröhn,
and Y. H. Schukken. 2001. 'Cow- and quarter-level risk factors for Streptococcus uberis
and Staphylococcus aureus mastitis', J Dairy Sci, 84: 2649-63.
Zamfir, Medana, Iulia-Roxana Stefan, Mihaela-Marilena Stancu, and Silvia-Simona Grosu-
Tudor. 2016. 'Production, mode of action and sequencing of the corresponding gene of a
bacteriocin produced by Lactococcus lactis 19.3', Int J Food Sci Technol, 51: 2164-70.
Zhu, Shaozhou, Christopher D. Fage, Julian D. Hegemann, Andreas Mielcarek, Dushan Yan,
Uwe Linne, and Mohamed A. Marahiel. 2016. 'The B1 Protein Guides the Biosynthesis
of a Lasso Peptide', Scientific Reports, 6: 35604.
Zouhir, Abdelmajid, Riadh Hammami, Ismail Fliss, and Jeannette Ben Hamida. 2010. 'A new
structure-based classification of Gram-positive bacteriocins', Protein J, 29: 432-39.


















