![Stability of bacteriophages in burn wound care products · sidered is phage therapy [6–8], i.e. the use of bacterial viruses named bacteriophages (phages for short) to treat bacterial](https://static.fdocuments.in/doc/165x107/5ecd79c66a5b361e1e24365d/stability-of-bacteriophages-in-burn-wound-care-products-sidered-is-phage-therapy.jpg)
Bacterial viruses - International University of Sarajevo
36
Bacterial viruses Mirela Musić Emina Huseinbegović BIO418 Fall 2019
Transcript of Bacterial viruses - International University of Sarajevo

Bacterial viruses
Mirela Musić
Emina Huseinbegović
BIO418 Fall 2019

Introduction
• Bacterial viruses/Bacteriophages/Phages
• Diversity of phages
• Common in most environments where bacteria are found and they are important in regulating their abundance and distribution
Morphology
Genetic characteristics
Simple – icosahedral and filamentous
Complex – tailed with icosahedral head
DNA or RNA
ss or ds
circular or linear

Introduction
• Phages can be classified as: • Virulent → lysis cycle
• Temperate → productive (lytic infection) and reproductive (lysogeny)
• Roles in developments in molecular biology and biotechnology • model systems for plant and animal viruses
• understanding mechanisms in cell
• first genomes to be sequenced
• restriction enzymes
• Therapeutic potential

ssRNA phages
• small, icosahedral viruses of family Leviviridae
• high mutation rate and some of the smallest RNA genomes
• few genes and infect various Gram- bacteria

Virion structure of ssRNA phages
• 180 molecules of major coat protein (CP), one molecule of maturation protein (A) and linear ssRNA genome of 3500 to 4200 nucleotides

Genome of ssRNA phages
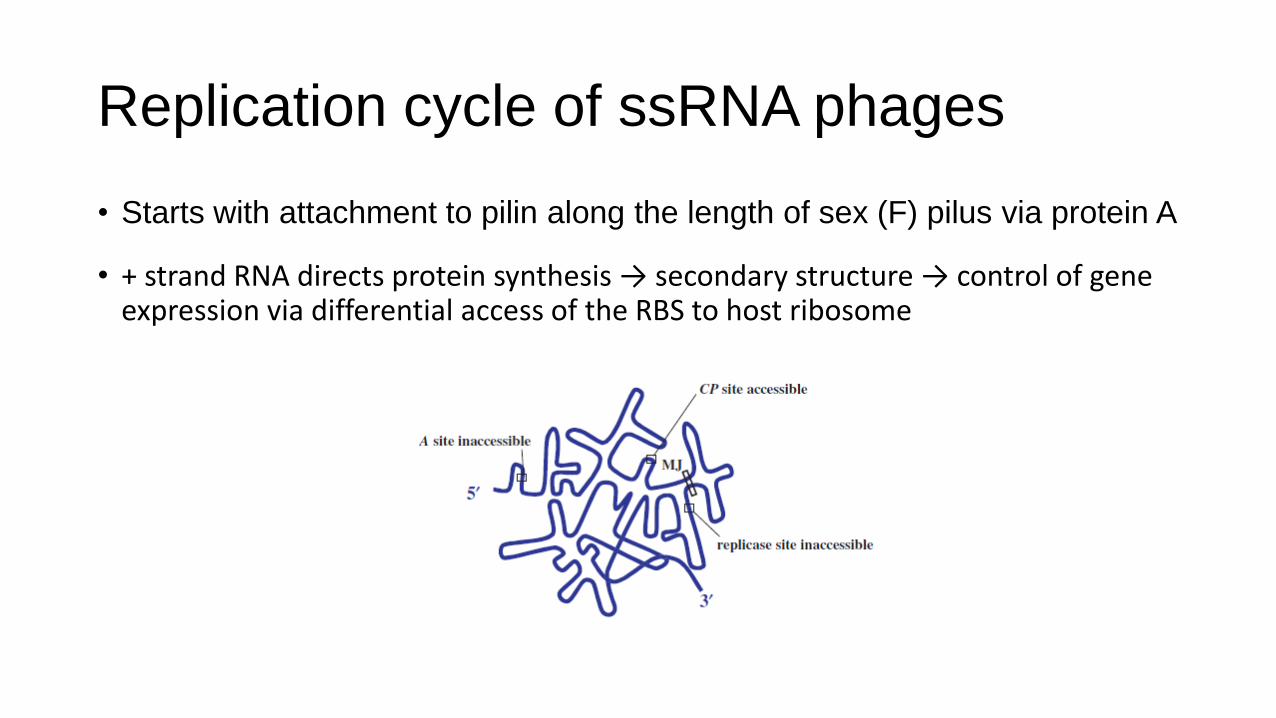
Replication cycle of ssRNA phages
• Starts with attachment to pilin along the length of sex (F) pilus via protein A
• + strand RNA directs protein synthesis → secondary structure → control of gene expression via differential access of the RBS to host ribosome
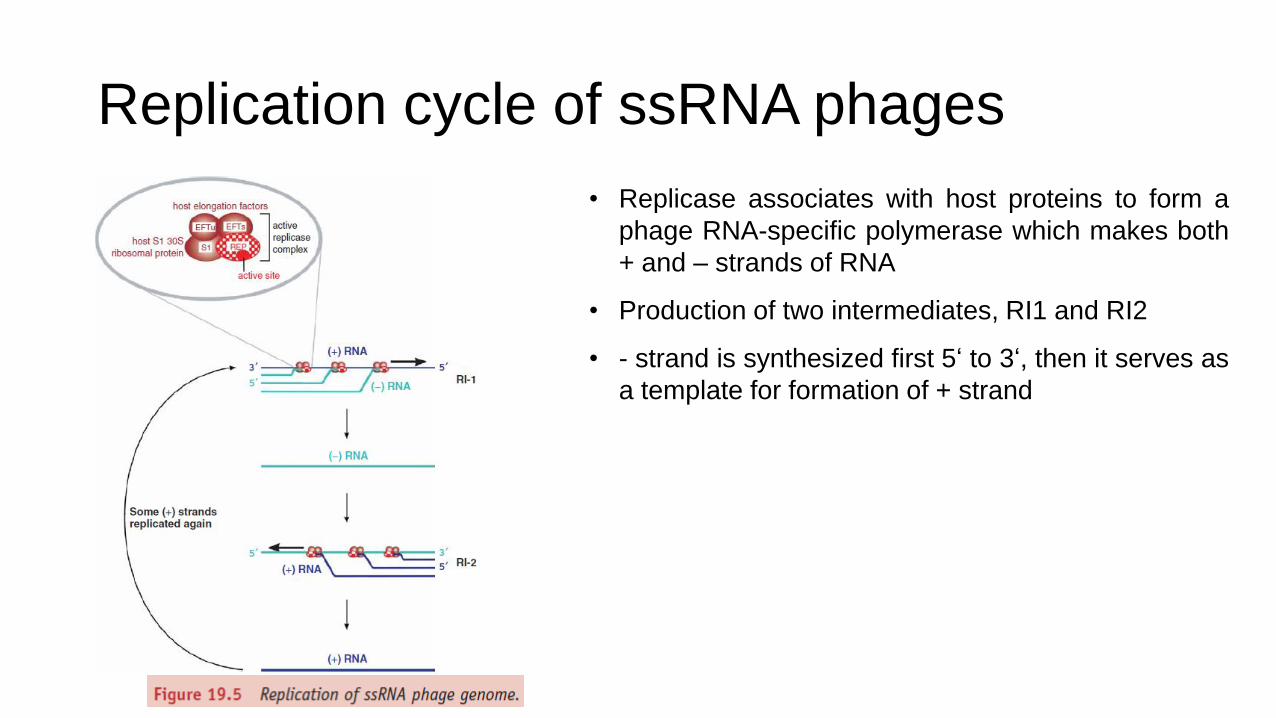
Replication cycle of ssRNA phages
• Replicase associates with host proteins to form a
phage RNA-specific polymerase which makes both
+ and – strands of RNA
• Production of two intermediates, RI1 and RI2
• - strand is synthesized first 5‘ to 3‘, then it serves as
a template for formation of + strand

Replication cycle of ssRNA phages

dsRNA phages
• family Cystoviridae, segmented and packed in a polyhedral inner core with a lipid containing envelope
• φ6 → first to be isolated, genome comprises 3 linear segments
• RNA L (large) → early proteins that assemble to form polymerase complex
• RNA M (medium) → structural proteins for membranes
• RNA S (small) → structural proteins for capsid, membrane assembly, lysis and entry, and non-structural proteins for envelopment of capsid

DNA PHAGES

19.4 Single-stranded DNA phages
• Icosahedral ssDNA phages • ϕX174 (virion, genome, and
replication cycle) • Belong to the family Microviridae
• Such phages provided the first evidence for overlapping genes
• Studies on replication of these phages led to the discovery of rolling circle replication and to the identity of various genes encoding proteins for host DNA replication.
• Filamentous ssDNA phages • Ff phages (virion, and
replication cycle) • Belong to family Inoviridae.
• Unlike many other DNA phages, filamentous phages do not inject their DNA into the host cell; rather entire phage particles are ingested.
• Furthermore, these phages do not lyse infected cells, but progeny are continuously extruded through the cell membrane.
• Despite this, plaque-like zones are formed in a lawn of sensitive bacteria, since infected cells grow more slowly than uninfected cells.
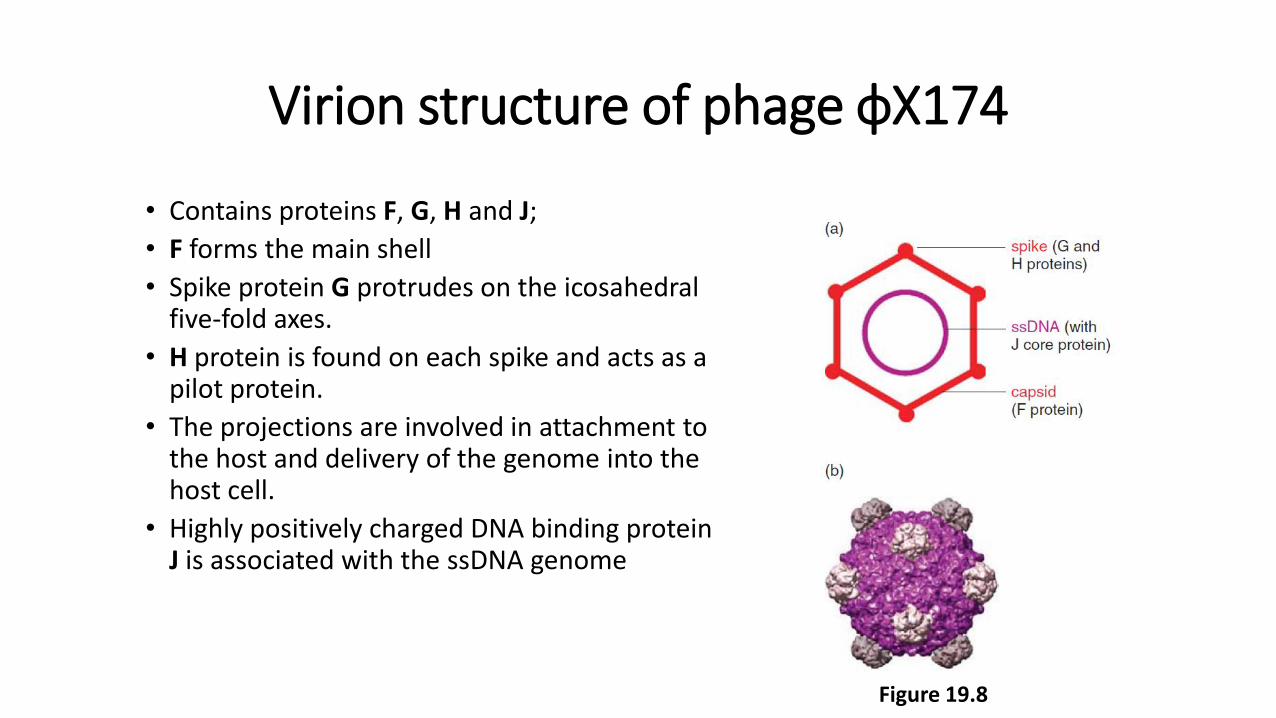
Virion structure of phage ϕX174
• Contains proteins F, G, H and J;
• F forms the main shell
• Spike protein G protrudes on the icosahedral five-fold axes.
• H protein is found on each spike and acts as a pilot protein.
• The projections are involved in attachment to the host and delivery of the genome into the host cell.
• Highly positively charged DNA binding protein J is associated with the ssDNA genome
Figure 19.8

Genome of phage ϕX174
• The genome is a circular ssDNA molecule of 5386 nucleotides, coding for 11 proteins.
• Different proteins are generated through extensive use of overlapping genes translated in alternative reading frames or employing different start codons.
• Gene A contains an internal translation initiation site to encode protein A*;
• B is encoded within A in a different reading frame;
• K is at the end of gene A and extends into gene C and is translated in a different frame to A and C;
• E is totally within D, but in a different reading frame;
• The termination codon for D overlaps initiation codon for J
Figure 19.9
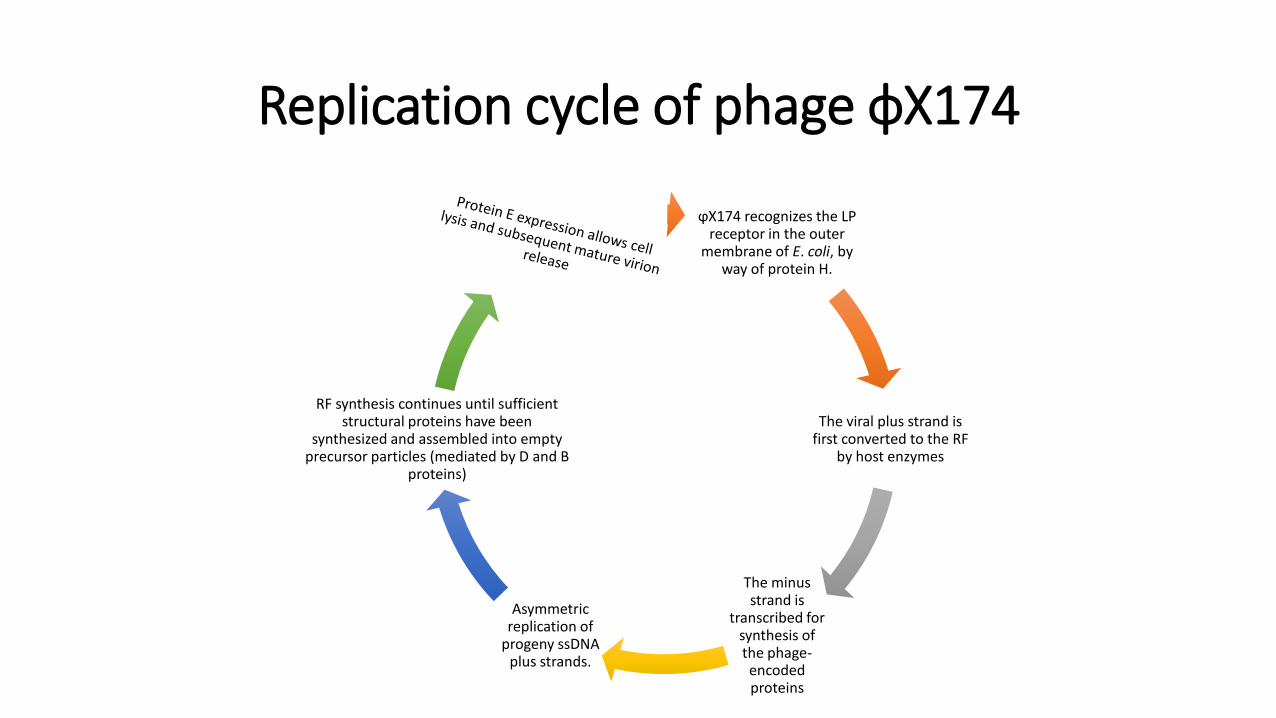
Replication cycle of phage ϕX174
ϕX174 recognizes the LP receptor in the outer
membrane of E. coli, by way of protein H.
The viral plus strand is first converted to the RF
by host enzymes
The minus strand is
transcribed for synthesis of the phage-encoded proteins
Asymmetric replication of
progeny ssDNA plus strands.
RF synthesis continues until sufficient structural proteins have been
synthesized and assembled into empty precursor particles (mediated by D and B
proteins)

Genome of Ff phages
• The genome of the Ff phages is a circular ssDNA molecule of about 6400 nucleotides.
• The genes are tightly packed on the genome, with a number of intergenic (IG) regions.
• The major IG region of 507 nucleotides (between genes IV and II ) contains a number of regulatory regions:
• the origin of replication for (+) and (−) DNA synthesis, • a strong rho-dependent transcription terminator,
• 78- nucleotide hairpin region, which is the packaging signal (PS) for efficient phage assembly.
• The genome encodes 11 proteins. Genes are grouped by function: • genes II, V and X for phage DNA synthesis;
• genes III, VI, VII, VIII and IX for capsid structure; • genes I , IV and XI for assembly
• Transcription is complex, involving several strong promoters.
• The two strong transcription terminators divide the genome into two transcription regions:
• a frequently transcribed region from gene II to VIII and • an infrequently transcribed region from gene III to IV.
• All the genes are transcribed in the same direction.
Figure 19.10

Virion structure of Ff phages
• The capsid is a flexible protein filament of variable length, depending on genome size;
• The filament comprises about 2700 copies of the 50 amino acid major coat protein, arranged in an overlapping array to form a helix.
• The basic positively charged C-terminal domain of pVIII forms the inside wall of the filament and interacts with the DNA.
• The N-terminal negatively charged domain is exposed on the outside.
• The minor coat proteins are present at the ends of the filament.
• At one terminus (where assembly terminates) are two of these proteins:
• gene III protein (pIII) and gene VI protein (pVI) (together form an adsorption complex for recognition ofthe sex pilus)
• At the other terminus (where assembly initiates) are:
• gene VII protein (pVII) and gene IX protein (pIX).
Figure 19.11

Replication cycle of Ff phages
The phage binds first to the tip of
the F pilus through domain N2 of pIII. The pilus retracts
bringing pIII to the periplasm and
domain N1 of pIII can then interact
with TolA
The phage particle enters the cell and
disassembles.
TolA, TolQ and TolR proteins
mediate depolymerization
of the coat proteins
pIII enters the cell and havs a role in the initiation of
replication
Phage DNA is plus strand and DNA
replication occurs immediately upon infection in order to provide the (−)
DNA for transcription.
Replication occurs in three stages
N1= membrane penetration (interacts with TolA) N2= adsorption to receptor
TolA, TolQ, TolR = translocation of the phage DNA to cytoplasm and coat proteins to cytoplasmic membrane

Figure 19.13

Cloning vectors and phage display
• The relatively small size and non-lytic infection cycle make the filamentous phages easy to manipulate and use as cloning vectors.
• Since filament size is governed by size of the genomic DNA, insert DNA of variable length can be cloned.
• Cloned ssDNA can be recovered from the phage particles for use in, for example, DNA sequencing, designing DNA probes and site-directed mutagenesis.
• RF based vector function based on the lacZ gene detection principle.

Phagemids
• Phagemids combine useful features of plasmids and ssDNA phages. • larger inserts can be cloned,
• double-stranded sequences as plasmids,
• amplified and recovered as ssDNA in virions.
• A phagemid typically contains a selectable marker, an origin of plasmid replication, polylinker and site for insertional inactivation and the IG of an ssDNA phage for initiation and termination of DNA replication.

Phage display
• Phage display technology is an in vitro screening technique for identifying ligands for proteins and other macromolecules. At the crux of phage display technology is the ability to express peptide or protein sequences as fusions to the coat proteins of a bacteriophage. Libraries of phage-displayed peptides or proteins are thereby physically linked to their encoding nucleic acid, allowing selection of binding partners for myriad target types by iterative rounds of in vitro panning and amplification, followed by DNA sequencing.
• A notable application of phage display technology has been in the engineering of antibodies
New England Biolabs https://www.youtube.com/watch?v=AqQDZxoCGqE

Double-stranded DNA phages
• T and λ phages have been particularly well characterized.
Phage family
Siphovidirae: T1 and λ
Podoviridae: T3 and T7
Myoviridae: T2, T4, T6

Phage T4
• Elongated icosahedral head with a number of different types of protein, including two accessory proteins: Hoc (highly antigenic outer capsid protein) and Soc (small outer capsid protein) decorating the external surface, a contractile tail, baseplate, tail fibres and pins.
• The T4 genome is linear dsDNA of about 169 kb.
• The genome is AT-rich and contains modified bases in the form of 5-hydroxy-methyl-cytosine.
• The genome comprises about 300 probable genes assigned by the timing of their expression, with three different types of promoter: early (Pe), middle (Pm) and late (Pl).
Figure 19.15

Phage T7 • T7 is a virulent T-odd phage, with icosahedral head and short, non-contractile tail.
• The linear genome (about 39 kbp) is smaller than that of T4. It comprises about 50 genes, which are closely packed, divided into three groups.
Figure 19.17

λ phage • Phage λ comprises an isometric head
containing a dsDNA genome of about 48.5 kb and a long, flexible tail with central tail tip fibre and side tail fibres.
• Many genes encode head and tail components of the phage, but only two (O and P) for replication.
• The genome in the virion is linear, with 12 bases that mediate circularization of the DNA after infection of the host. Circularization of the λ genome protects it from degradation.
• As a temperate phage, λ can either establish lysogeny, being maintained as a prophage integrated into the bacterial chromosome between the biotin and galactose operons, or multiply lytically to produce progeny virions upon infection.
Figure 19.19


Introduction
• Imbalances in the gut microbiota resulting from poor diet, stress, antibiotic use, and other lifestyle and environmental factors are associated with the development of intestinal inflammation and bowel irregularities.
• Bacteriophages (or phages) are among the candidates being explored as potential microbial modifiers in promoting intestinal health. These ubiquitous, bacteria-targeting viruses exhibit a high degree of host specificity, suggesting utility for selectively reducing pathogenic or pro-inflammatory bacteria in the microbial environment.
• While antibiotics can cause or exacerbate microbiota imbalances or dysbiosis, phages offer the opportunity to subtly and selectively modify the gut microbiota. Several bacteriophages are GRAS for human consumption by the U.S. FDA.
• AIM:
• to determine how daily consumption of supplemental E. coli-targeting phages influences the gut microbiota of healthy adults with self-reported gastrointestinal distress;
• Additional effects are also reported in this study.

Materials and methods
• Participants:
• Forty-three healthy adults aged 18–65 with self-reported gastrointestinal issues
• Exclusion criteria included a) a previous diagnosis of gastrointestinal or metabolic
conditions, cancer, liver, or kidney diseases; b) pregnancy or breastfeeding; c) smoking; d) use of antibiotics in the last 2 months; and e) current medication or dietary supplement use that
may impact gut microbiota.
• Participants were asked to a) maintain their regular diet and exercise habits
throughout the study, b) refrain from supplemental prebiotics or probiotics, c) and limit alcohol consumption to 1 drink per day or
no more than 7 drinks per week
• 36 participants completed at least one arm of the study and 32 of those completed the entire study
• The data shown here represent analysis from these 36 participants.
• Study design:
• Randomized, double-blind, placebo-controlled crossover intervention trial, with two 28 day intervention periods and a washout period of at least two weeks between treatments.
• The treatments consisted of 4 supplemental bacteriophage strains at a titer of 106 phages per dose included in PreforPro® commercial capsules prepared by Deerland Enzymes.
• The placebo was rice maltodextrin and coconut triglycerides, which were also the carrier materials used in the treatment capsules.
• One 15 mg capsule per day in test and control group.
• The study was conducted at the Human Performance Clinical Research Laboratory (HPCRL) at Colorado State University.
• Four times participants provided a fresh stool and blood samples.

DNA extraction and sequencing
Fecal DNA was extracted from the swabs using the FastDNA® KIT; sequencing by MiSeq flow cell
Microbiota analysis
Bioinformatic analysis
Short chain fatty acids
• Supernatant analysis by Gas Chromatograph with Flame Ionization Detection
Fecal tryglicerides
(Triglycerides Assay Kit)
Local and systemic inflammation
(sIgA-ELISA; CRP, chemokines and cytokines-Miliplex T cell panel)
Plasma lipids
(HDL, TRIGs, nHDL, tC/HDL, LDL, VLDL by Piccolo Xpress Chemistry Blood Analyzer for lipid panel)
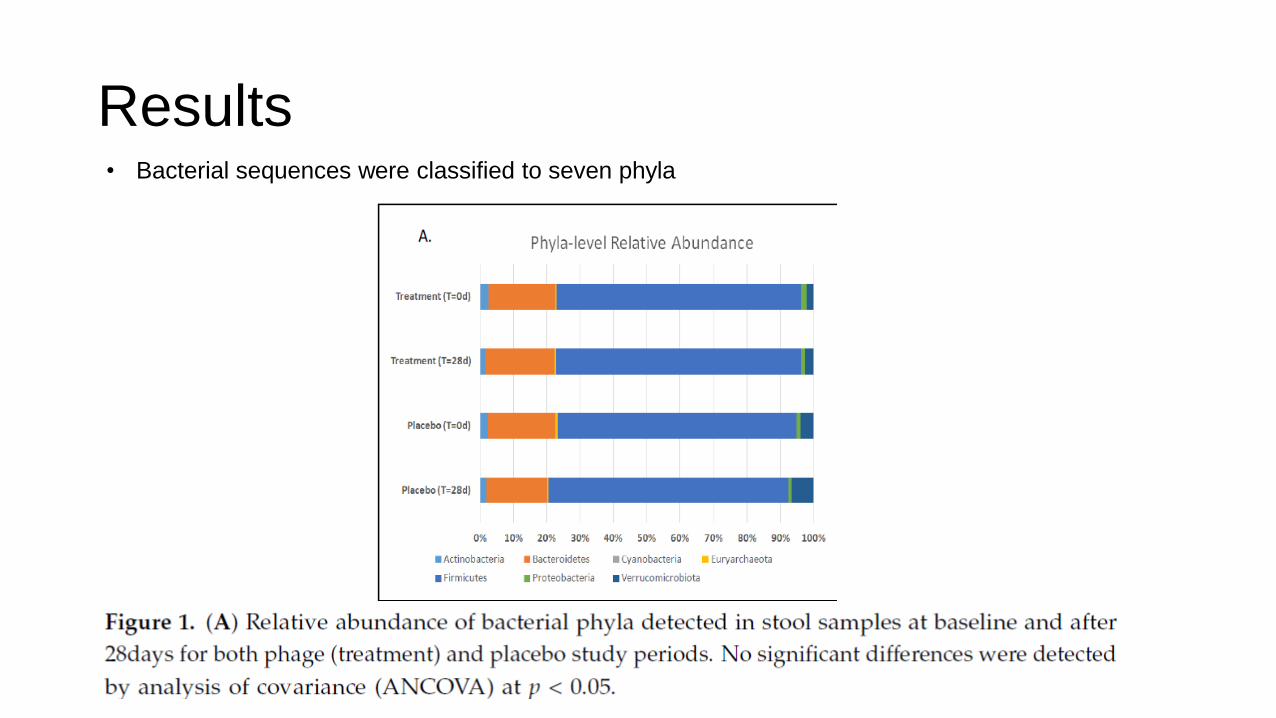
Results • Bacterial sequences were classified to seven phyla

Results
• Identification of ASVs in samples that mapped to E.coli
• E.coli levels were significantly reduced after phage treatment

Results
• Several microbial taxa were significantly correlated with fecal levels of E.coli

Results • Three ASVs representing potentially pathogenic or inflammation-
associated taxa were reduced relative to the placebo
• Two ASVs were increased after phage consumption

Results
• Stool and plasma lipid profiles
• Since the microbiota plays a role in lipid absorption , levels of triglyceride
were measured; however, there was no significant difference across
timepoints or between treatment periods (except: significant period effect
noted for participants starting on the placebo and transitioning to the
treatment group)
• Immunological and Inflammatory markers
• In stool, calprotectin and sIgA were examined (normal range)
• In plasma, C-reactive protein and human T-cell-derived cytokines were
analyzed, only interleukin-4 (Il-4) was significantly reduced

Conclusion
• Bacteriophage consumption caused minimal disruption to the gut microbiota, but did elicit minor changes
• The reduction of E.coli decreased the level of pro-inflammatory bacteria, and increased level of fermentative taxa (shift toward a healthier gut environment)
• This study confirms the safety and tolerability of phage consumption in a human population


















