
b -Adrenergic Receptor Agonists Modulate Human Airway Smooth Muscle Cell Migration … ·...
7
b 2 -Adrenergic Receptor Agonists Modulate Human Airway Smooth Muscle Cell Migration via Vasodilator-Stimulated Phosphoprotein Elena A. Goncharova 1 , Dmitry A. Goncharov 1 , Hengjiang Zhao 1 , Raymond B. Penn 4 , Vera P. Krymskaya 1–3 *, and Reynold A. Panettieri, Jr. 1 * 1 Airways Biology Initiative, Division of Pulmonary, Allergy, and Critical Care, Department of Medicine, 2 Cardiovascular Institute, and 3 Abramson Cancer Center, University of Pennsylvania, Philadelphia, Pennsylvania; and 4 Department of Medicine, University of Maryland, Baltimore, Maryland Severe asthma manifests as airway remodeling and irreversible airway obstruction, in part because of the proliferation and migra- tion of human airway smooth muscle (HASM) cells. We previously reported that cyclic adenosine monophosphate–mobilizing agents, including b 2 -adrenergic receptor (b 2 AR) agonists, which are main- stay of asthma therapy, and prostaglandin E2 (PGE2), inhibit the migration of HASM cells, although the mechanism for this migration remains unknown. Vasodilator-stimulated phosphoprotein (VASP), an anticapping protein, modulates the formation of actin stress fibers during cell motility, and is negatively regulated by protein kinase A (PKA)–specific inhibitory phosphorylation at serine 157 (Ser157). Here, we show that treatment with b 2 AR agonists and PGE2 induces the PKA-dependent phosphorylation of VASP and inhibits the migration of HASM cells. The stable expression of PKA inhibitory peptide and the small interfering (si) RNA-induced deple- tion of VASP abolish the inhibitory effects of albuterol and PGE2 on the migration of HASM cells. Importantly, prolonged treatment with albuterol prevents the agonist-induced phosphorylation of VASP at Ser157, and reverses the inhibitory effects of albuterol and formo- terol, but not PGE2, on the basal and PDGF-induced migration of HASM cells. Collectively, our data demonstrate that b 2 AR agonists selectively inhibit the migration of HASM cells via a b 2 AR/PKA/ VASP signaling pathway, and that prolonged treatment with albu- terol abolishes the inhibitory effect of b-agonists on the phos- phorylation of VASP and migration of HASM cells because of b 2 AR desensitization. Keywords: airway hyperresponsiveness; b 2 -adrenergic receptor desen- sitization; protein kinase A; albuterol; formoterol Human airway smooth muscle (HASM) hyperplasia and remod- eling are characteristic features of airways that contribute to their irreversible obstruction in patients with severe asthma. Ac- cumulating evidence suggests that cell migration contributes to airway smooth muscle hyperplasia and remodeling (1, 2). Re- cently, the migration of airway smooth muscle (ASM) cells to- ward the epithelium and lumen of the airway was reported as a potential contributor to the increased smooth muscle mass in patients with asthma (3, 4). Further, cell-culture studies show that mitogens and inflammatory mediators involved in asthma pathogenesis and pharmacological agents in current used for the treatment of asthma modulate the migration of ASM cells (1, 5, 6). Taken together, these studies suggest that the migration of ASM cells contributes to ASM cell hyperplasia under asthma- related conditions (1). Although the role of HASM hyperplasia in asthma is well established, the molecular mechanisms regu- lating the migration of HASM cells under asthma-related con- ditions are poorly understood (1). Evidence demonstrates that cyclic adenosine monophosphate (cAMP)–inducing agents, including the long-acting b 2 -adrenergic receptor (b 2 AR) agonist formoterol, inhibit the agonist-dependent migration of ASM cells (5, 7). We previously reported that the cAMP-inducing agents salmeterol, cilomolast, and prostaglandin E2 (PGE2) attenuate the migration of HASM cells (6) and induce actin depolimerization in HASM cells (8). However, the mecha- nisms by which b 2 AR agonists modulate the actin cytoskeleton and inhibit cell migration remain unknown. Cell migration requires actin cytoskeleton remodeling, in- cluding the disassembly of actin stress fibers and the formation of actin branching at the lamellipodia protrusion at the leading edge of migrating ASM cells. Vasodilator-stimulated phospho- protein (VASP) belongs to a conserved family of actin- regulatory proteins that regulate actin stress fiber assembly during cell migration (9). VASP facilitates the assembly of the actin filaments by serving as an anticapping protein (9). Agents that elevate cellular concentrations of cAMP and cyclic guano- sine monophosphate promote the inhibitory phosphorylation of VASP and attenuate the anticapping activity of VASP (9). The activity of VASP is negatively regulated by the inhibitory phos- phorylation of serine 157 (Ser157) by protein kinase A (PKA) (Received in original form June 24, 2011 and in final form July 21, 2011) *These authors contributed equally to this work. This work was supported by GSK (R.A.P.), Sepracor (R.A.P.), and National Insti- tutes of Health grant HL58506 (R.B.P.). Correspondence and requests for reprints should be addressed to Reynold A. Panettieri, Jr., M.D., Airways Biology Initiative, Division of Pulmonary, Allergy, and Critical Care, Department of Medicine, University of Pennsylvania, 125 South 31st St., TRL Suite 1200, Philadelphia, PA 19104. E-mail: [email protected] This article has an online supplement, which is accessible from this issue’s table of contents at www.atsjournals.org Am J Respir Cell Mol Biol Vol 46, Iss. 1, pp 48–54, Jan 2012 Originally Published in Press as DOI: 10.1165/rcmb.2011-0217OC on July 28, 2011 Internet address: www.atsjournals.org CLINICAL RELEVANCE Understanding the cellular and molecular mechanisms of human airway smooth muscle (HASM) remodeling is critically important, not only for advancing our knowledge about disease pathobiology but also in terms of indentifying novel molecular targets for therapeutic intervention. This study identifies a mechanism for the inhibitory effects of b 2 -adrenergic re- ceptor (b 2 AR) agonists on the migration of HASM cells. We found that the b 2 AR agonist–induced activation of protein kinase A modulates the phosphorylation of vasodilator- stimulated phosphoprotein, which inhibits the migration of HASM cells. Furthermore, we found that the prolonged treatment of HASM cells with b2AR agonists desensitizes the airways to further treatment with the b2AR agonist. Our data suggest that the prolonged use of b2AR agonists in asthma as a single therapy may promote the development of tolerance to b2AR agonists and the migration of airway smooth muscle cells.
Transcript of b -Adrenergic Receptor Agonists Modulate Human Airway Smooth Muscle Cell Migration … ·...

b2-Adrenergic Receptor Agonists Modulate HumanAirway Smooth Muscle Cell Migration viaVasodilator-Stimulated Phosphoprotein
Elena A. Goncharova1, Dmitry A. Goncharov1, Hengjiang Zhao1, Raymond B. Penn4,Vera P. Krymskaya1–3*, and Reynold A. Panettieri, Jr.1*
1Airways Biology Initiative, Division of Pulmonary, Allergy, and Critical Care, Department of Medicine, 2Cardiovascular Institute, and 3Abramson
Cancer Center, University of Pennsylvania, Philadelphia, Pennsylvania; and 4Department of Medicine, University of Maryland, Baltimore, Maryland
Severe asthma manifests as airway remodeling and irreversibleairway obstruction, in part because of the proliferation and migra-tion of human airway smooth muscle (HASM) cells. We previouslyreported that cyclic adenosine monophosphate–mobilizing agents,including b2-adrenergic receptor (b2AR) agonists, which are main-stay of asthma therapy, and prostaglandin E2 (PGE2), inhibit themigrationofHASMcells, although themechanism for thismigrationremains unknown. Vasodilator-stimulated phosphoprotein (VASP),an anticapping protein, modulates the formation of actin stressfibers during cell motility, and is negatively regulated by proteinkinase A (PKA)–specific inhibitory phosphorylation at serine 157(Ser157). Here, we show that treatment with b2AR agonists andPGE2 induces the PKA-dependent phosphorylation of VASP andinhibits the migration of HASM cells. The stable expression of PKAinhibitory peptide and the small interfering (si) RNA-induced deple-tion of VASP abolish the inhibitory effects of albuterol and PGE2 onthemigrationofHASMcells. Importantly, prolonged treatmentwithalbuterol prevents the agonist-induced phosphorylation of VASP atSer157, and reverses the inhibitory effects of albuterol and formo-terol, but not PGE2, on the basal and PDGF-induced migration ofHASMcells. Collectively, our data demonstrate thatb2AR agonistsselectively inhibit the migration of HASM cells via a b2AR/PKA/VASP signaling pathway, and that prolonged treatmentwith albu-terol abolishes the inhibitory effect of b-agonists on the phos-phorylation of VASP and migration of HASM cells because ofb2AR desensitization.
Keywords: airway hyperresponsiveness; b2-adrenergic receptor desen-
sitization; protein kinase A; albuterol; formoterol
Human airway smooth muscle (HASM) hyperplasia and remod-eling are characteristic features of airways that contribute totheir irreversible obstruction in patients with severe asthma. Ac-cumulating evidence suggests that cell migration contributes toairway smooth muscle hyperplasia and remodeling (1, 2). Re-cently, the migration of airway smooth muscle (ASM) cells to-ward the epithelium and lumen of the airway was reported asa potential contributor to the increased smooth muscle mass inpatients with asthma (3, 4). Further, cell-culture studies showthat mitogens and inflammatory mediators involved in asthma
pathogenesis and pharmacological agents in current used for thetreatment of asthma modulate the migration of ASM cells (1, 5,6). Taken together, these studies suggest that the migration ofASM cells contributes to ASM cell hyperplasia under asthma-related conditions (1). Although the role of HASM hyperplasiain asthma is well established, the molecular mechanisms regu-lating the migration of HASM cells under asthma-related con-ditions are poorly understood (1).
Evidence demonstrates that cyclic adenosine monophosphate(cAMP)–inducing agents, including the long-acting b2-adrenergicreceptor (b2AR) agonist formoterol, inhibit the agonist-dependentmigration of ASM cells (5, 7). We previously reported that thecAMP-inducing agents salmeterol, cilomolast, and prostaglandinE2 (PGE2) attenuate the migration of HASM cells (6) and induceactin depolimerization in HASM cells (8). However, the mecha-nisms by which b2AR agonists modulate the actin cytoskeletonand inhibit cell migration remain unknown.
Cell migration requires actin cytoskeleton remodeling, in-cluding the disassembly of actin stress fibers and the formationof actin branching at the lamellipodia protrusion at the leadingedge of migrating ASM cells. Vasodilator-stimulated phospho-protein (VASP) belongs to a conserved family of actin-regulatory proteins that regulate actin stress fiber assemblyduring cell migration (9). VASP facilitates the assembly of theactin filaments by serving as an anticapping protein (9). Agentsthat elevate cellular concentrations of cAMP and cyclic guano-sine monophosphate promote the inhibitory phosphorylation ofVASP and attenuate the anticapping activity of VASP (9). Theactivity of VASP is negatively regulated by the inhibitory phos-phorylation of serine 157 (Ser157) by protein kinase A (PKA)
(Received in original form June 24, 2011 and in final form July 21, 2011)
*These authors contributed equally to this work.
This work was supported by GSK (R.A.P.), Sepracor (R.A.P.), and National Insti-
tutes of Health grant HL58506 (R.B.P.).
Correspondence and requests for reprints should be addressed to Reynold A.
Panettieri, Jr., M.D., Airways Biology Initiative, Division of Pulmonary, Allergy,
and Critical Care, Department of Medicine, University of Pennsylvania, 125 South
31st St., TRL Suite 1200, Philadelphia, PA 19104. E-mail: [email protected]
This article has an online supplement, which is accessible from this issue’s table of
contents at www.atsjournals.org
Am J Respir Cell Mol Biol Vol 46, Iss. 1, pp 48–54, Jan 2012
Originally Published in Press as DOI: 10.1165/rcmb.2011-0217OC on July 28, 2011
Internet address: www.atsjournals.org
CLINICAL RELEVANCE
Understanding the cellular and molecular mechanisms ofhuman airway smoothmuscle (HASM) remodeling is criticallyimportant, not only for advancing our knowledge about diseasepathobiology but also in terms of indentifying novel moleculartargets for therapeutic intervention. This study identifiesa mechanism for the inhibitory effects of b2-adrenergic re-ceptor (b2AR) agonists on the migration of HASM cells. Wefound that the b2AR agonist–induced activation of proteinkinase A modulates the phosphorylation of vasodilator-stimulated phosphoprotein, which inhibits the migration ofHASM cells. Furthermore, we found that the prolongedtreatment of HASM cells with b2AR agonists desensitizes theairways to further treatment with the b2AR agonist. Our datasuggest that the prolonged use of b2AR agonists in asthma asa single therapy may promote the development of toleranceto b2AR agonists and the migration of airway smoothmuscle cells.

(9). Although the importance of protein kinase A in cell migra-tion is supported by several reports, published data indicate thatPKA may serve to either promote or inhibit cell motility (10–12), suggesting that the role of PKA in cell migration ishighly cell type–dependent and context-dependent. b2AR ago-nists modulate the activity of PKA via the production of cAMP,but the role of PKA/VASP signaling in the b2AR agonist-dependent migration of HASM cells remains unclear.
In this study, we demonstrate that b2AR agonists modulatethe migration of HASM cells through a b2AR/PKA/VASP sig-naling pathway. We also show that the prolonged exposure ofHASM cells to albuterol abrogates the PKA-specific phosphor-ylation of VASP, and prevents the b2AR agonist-dependentinhibition of HASM cell migration, suggesting the potentialclinical relevance of our findings.
MATERIALS AND METHODS
Cell Culture
HASM cells were dissociated from human tracheas obtained from hu-man lung transplant donors, in accordance with procedures approvedby the Committee on Studies Involving Human Beings at the Universityof Pennsylvania, as described previously (13). Cells were culturedin Ham’s F12 media (Life Technologies, Grand Island, NY), supple-mented with 10% FBS (HyClone Laboratories, Logan, UT), 100 U/mlpenicillin, and 0.1 mg/ml streptomycin. Before the experiments, HASMcells were maintained for 48 hours in serum-free Ham’s F12 media,supplemented with 0.1% BSA.
Immunoblot Analysis
Growth-arrested cells were transfected with siRNA VASP or controlsmall interfering (si) RNA, or treated with different concentrationsof racemic, (R)-albuterol (where “R” stands for rectus) or (S)-albuterol(where “S” stands for sinistre), formoterol, PGE2, or propranolol inthe presence or absence of preincubation with albuterol, followed byimmunoblot analysis with anti-phospho VASP Ser157 or anti-VASPantibodies (Cell Signaling Technology, Inc., Danvers, MA).
Immunocytochemical Analysis
Cells growth-arrested for 48 hours were incubated with 0.01, 0.1, 1, or10 mM formoterol or diluent for 5, 15, or 30 minutes, or with 10 mM(R)-albuterol, (S)-albuterol, or diluent for 5, 15, and 30 minutes, fol-lowed by immunocytochemical analysis with anti-phospho VASPSer157 antibody, as previously described (14). A detailed explanationof the technique is provided in Figure E1 in the online supplement.
Transient Transfection
Cells were transfected with 100 or 250 nM siRNA VASP or controlsiGLO RISC-Free (GLO) siRNA for 48 hours with RNAiFect transfec-tion reagent (Qiagen, Valencia, CA), according to the manufacturer’sprotocol, as described elsewhere (15). The siRNA-induced depletion ofVASP was confirmed by immunoblot analysis with anti-VASP antibody.
Cell Migration Assay
Cell migration was examined using a Boyden chamber apparatus, as wedescribed previously (6, 16). Briefly, HASM cells were growth-arrestedfor 48 hours, transfected with siRNA VASP or control siRNA GLO, orincubated with albuterol. HASM cells were then briefly trypsinized by0.05% trypsin/0.53 mM EDTA, centrifuged at 900 rpm for 10 minutes,and resuspended in serum-free media supplemented with BSA. Cells(5 3 104) were then placed into the upper wells of the Boyden chamber,fitted with an 8-mm pore membrane coated with Vitrogen (AdvancedBioMatrix, San Diego, CA) (100 mg/ml). Agonists or vehicle in serum-free media supplemented with BSA were added to the lower chambers.Cells in the Boyden chamber were incubated for 4 hours at 378C in a 5%
CO2 incubator. Nonmigrated cells were scraped off. The membrane wasfixed with methanol, stained with a Hemacolor stain set (EM Industries,Inc., Gibbstown, NJ), and scanned. Cell migration was analyzed using theGel-Pro analyzer program (Media Cybernetics, Silver Spring, MD).
Statistical Analysis
Data points from individual assays represent mean values 6 SE. Sta-tistically significant differences among groups were assessed accordingto ANOVA (Bonferroni-Dunn test), with values of P , 0.05 con-sidered sufficient to reject the null hypothesis for all analyses. Allexperiments were designed with matched control conditions withineach experiment, to enable statistical comparisons as paired samples.All experiments were performed with a minimum of three differentHASM cell cultures.
RESULTS
Short-Term Treatment with b2AR Agonists and PGE2
Promotes the Phosphorylation of VASP in HASM Cells
Because cAMP-mobilizing agents induce the activation of PKAand the inhibitory phosphorylation of VASP in different typesof cells (9), we examined whether the b2AR agonists albuteroland formoterol modulate the PKA-dependent activity of VASPin HASM cells by using two different techniques, that is, immu-noblot and immunocytochemical analyses with antibodies spe-cifically recognizing VASP phosphorylated at Ser157, a majorsite of PKA phosphorylation.
As shown in Figure 1, a 30-minute incubation with eitheralbuterol or formoterol induced the dose-dependent phosphory-lation of VASP, compared with diluent-treated cells at concen-trations comparable to those for PGE2 (Figures 1A and 1B,respectively). Notably, formoterol induced the phosphorylationof VASP at 10–100 times lower doses compared with albuterol,which is consistent with the systemic dose potency of formoterol,which is 10–100 times higher than that of albuterol (17). Immu-nocytochemical analysis demonstrated that formoterol increasedthe phosphorylation of VASP at Ser157 in a concentration-dependent and time-dependent manner (Figure E1). Similarly,lower doses of formoterol were required to inhibit the migra-tion of HASM cells, compared with albuterol (the half maximalinhibitory concentration was approximately 0.1 mM and1 mM for formoterol and albuterol, respectively) (Figure E2),
Figure 1. Short-term incubation with b2-adrenergic receptor (b2AR)
agonists and prostaglandin E2 (PGE2) promotes the phosphorylation
of vasodilator-stimulated phosphoprotein (VASP) in human airwaysmooth muscle (HASM) cells. HASM cells were growth-arrested for
48 hours and incubated with different concentrations of albuterol
(A), formoterol (B), PGE2, or diluent (0) for 30 minutes, followed byimmunoblot analysis with anti-phospho VASP serine 157 (Ser157) or
anti-total VASP antibodies. Images are representative of three indepen-
dent experiments. P, phosphorylated.
Goncharova, Goncharov, Zhao, et al.: VASP, b2-Receptor Agonists, and HASM Migration 49
suggesting that a negative correlation exists between the phos-phorylation levels of VASP and the migration of HASM cells.
Collectively, these data show that the b2AR agonists albu-terol and formoterol induce the PKA-dependent phosphor-ylation of VASP at Ser157 in HASM cells, and inhibit themigration of HASM cells.
Binding with b2AR Is Required for the Albuterol-Dependent
Phosphorylation of VASP and Inhibition of HASM
Cell Migration
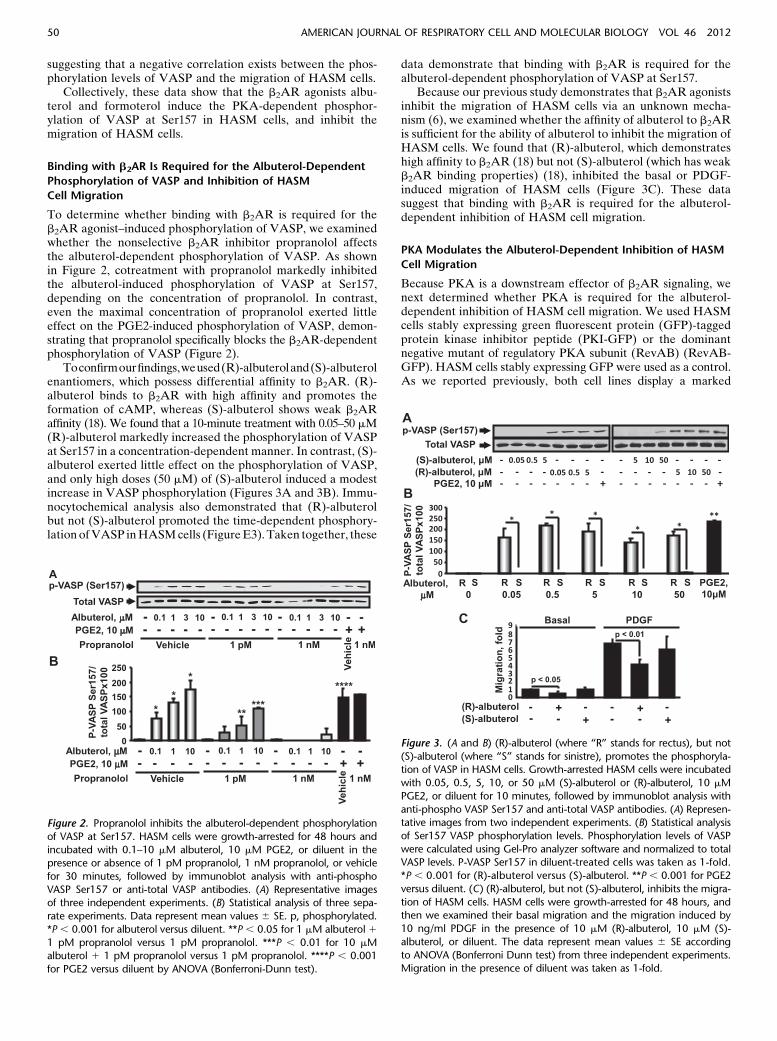
To determine whether binding with b2AR is required for theb2AR agonist–induced phosphorylation of VASP, we examinedwhether the nonselective b2AR inhibitor propranolol affectsthe albuterol-dependent phosphorylation of VASP. As shownin Figure 2, cotreatment with propranolol markedly inhibitedthe albuterol-induced phosphorylation of VASP at Ser157,depending on the concentration of propranolol. In contrast,even the maximal concentration of propranolol exerted littleeffect on the PGE2-induced phosphorylation of VASP, demon-strating that propranolol specifically blocks the b2AR-dependentphosphorylation of VASP (Figure 2).
Toconfirmourfindings,weused(R)-albuteroland(S)-albuterolenantiomers, which possess differential affinity to b2AR. (R)-albuterol binds to b2AR with high affinity and promotes theformation of cAMP, whereas (S)-albuterol shows weak b2ARaffinity (18). We found that a 10-minute treatment with 0.05–50 mM(R)-albuterol markedly increased the phosphorylation of VASPat Ser157 in a concentration-dependent manner. In contrast, (S)-albuterol exerted little effect on the phosphorylation of VASP,and only high doses (50 mM) of (S)-albuterol induced a modestincrease in VASP phosphorylation (Figures 3A and 3B). Immu-nocytochemical analysis also demonstrated that (R)-albuterolbut not (S)-albuterol promoted the time-dependent phosphory-lationofVASP inHASMcells (FigureE3). Taken together, these
data demonstrate that binding with b2AR is required for thealbuterol-dependent phosphorylation of VASP at Ser157.
Because our previous study demonstrates that b2AR agonistsinhibit the migration of HASM cells via an unknown mecha-nism (6), we examined whether the affinity of albuterol to b2ARis sufficient for the ability of albuterol to inhibit the migration ofHASM cells. We found that (R)-albuterol, which demonstrateshigh affinity to b2AR (18) but not (S)-albuterol (which has weakb2AR binding properties) (18), inhibited the basal or PDGF-induced migration of HASM cells (Figure 3C). These datasuggest that binding with b2AR is required for the albuterol-dependent inhibition of HASM cell migration.
PKA Modulates the Albuterol-Dependent Inhibition of HASM
Cell Migration
Because PKA is a downstream effector of b2AR signaling, wenext determined whether PKA is required for the albuterol-dependent inhibition of HASM cell migration. We used HASMcells stably expressing green fluorescent protein (GFP)-taggedprotein kinase inhibitor peptide (PKI-GFP) or the dominantnegative mutant of regulatory PKA subunit (RevAB) (RevAB-GFP). HASM cells stably expressing GFP were used as a control.As we reported previously, both cell lines display a marked
Figure 2. Propranolol inhibits the albuterol-dependent phosphorylation
of VASP at Ser157. HASM cells were growth-arrested for 48 hours and
incubated with 0.1–10 mM albuterol, 10 mM PGE2, or diluent in thepresence or absence of 1 pM propranolol, 1 nM propranolol, or vehicle
for 30 minutes, followed by immunoblot analysis with anti-phospho
VASP Ser157 or anti-total VASP antibodies. (A) Representative imagesof three independent experiments. (B) Statistical analysis of three sepa-
rate experiments. Data represent mean values 6 SE. p, phosphorylated.
*P , 0.001 for albuterol versus diluent. **P , 0.05 for 1 mM albuterol 11 pM propranolol versus 1 pM propranolol. ***P , 0.01 for 10 mMalbuterol 1 1 pM propranolol versus 1 pM propranolol. ****P , 0.001
for PGE2 versus diluent by ANOVA (Bonferroni-Dunn test).
Figure 3. (A and B) (R)-albuterol (where “R” stands for rectus), but not
(S)-albuterol (where “S” stands for sinistre), promotes the phosphoryla-tion of VASP in HASM cells. Growth-arrested HASM cells were incubated
with 0.05, 0.5, 5, 10, or 50 mM (S)-albuterol or (R)-albuterol, 10 mM
PGE2, or diluent for 10 minutes, followed by immunoblot analysis with
anti-phospho VASP Ser157 and anti-total VASP antibodies. (A) Represen-tative images from two independent experiments. (B) Statistical analysis
of Ser157 VASP phosphorylation levels. Phosphorylation levels of VASP
were calculated using Gel-Pro analyzer software and normalized to totalVASP levels. P-VASP Ser157 in diluent-treated cells was taken as 1-fold.
*P , 0.001 for (R)-albuterol versus (S)-albuterol. **P , 0.001 for PGE2
versus diluent. (C) (R)-albuterol, but not (S)-albuterol, inhibits the migra-
tion of HASM cells. HASM cells were growth-arrested for 48 hours, andthen we examined their basal migration and the migration induced by
10 ng/ml PDGF in the presence of 10 mM (R)-albuterol, 10 mM (S)-
albuterol, or diluent. The data represent mean values 6 SE according
to ANOVA (Bonferroni Dunn test) from three independent experiments.Migration in the presence of diluent was taken as 1-fold.
50 AMERICAN JOURNAL OF RESPIRATORY CELL AND MOLECULAR BIOLOGY VOL 46 2012
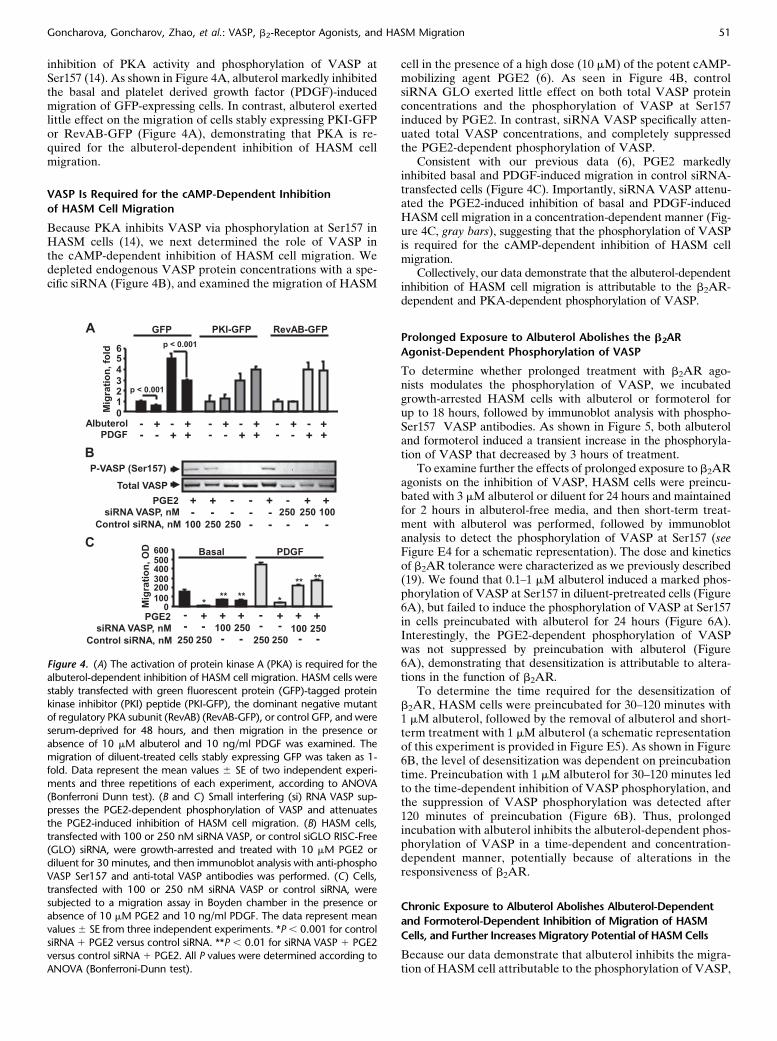
inhibition of PKA activity and phosphorylation of VASP atSer157 (14). As shown in Figure 4A, albuterol markedly inhibitedthe basal and platelet derived growth factor (PDGF)-inducedmigration of GFP-expressing cells. In contrast, albuterol exertedlittle effect on the migration of cells stably expressing PKI-GFPor RevAB-GFP (Figure 4A), demonstrating that PKA is re-quired for the albuterol-dependent inhibition of HASM cellmigration.
VASP Is Required for the cAMP-Dependent Inhibition
of HASM Cell Migration
Because PKA inhibits VASP via phosphorylation at Ser157 inHASM cells (14), we next determined the role of VASP inthe cAMP-dependent inhibition of HASM cell migration. Wedepleted endogenous VASP protein concentrations with a spe-cific siRNA (Figure 4B), and examined the migration of HASM
cell in the presence of a high dose (10 mM) of the potent cAMP-mobilizing agent PGE2 (6). As seen in Figure 4B, controlsiRNA GLO exerted little effect on both total VASP proteinconcentrations and the phosphorylation of VASP at Ser157induced by PGE2. In contrast, siRNA VASP specifically atten-uated total VASP concentrations, and completely suppressedthe PGE2-dependent phosphorylation of VASP.
Consistent with our previous data (6), PGE2 markedlyinhibited basal and PDGF-induced migration in control siRNA-transfected cells (Figure 4C). Importantly, siRNA VASP attenu-ated the PGE2-induced inhibition of basal and PDGF-inducedHASM cell migration in a concentration-dependent manner (Fig-ure 4C, gray bars), suggesting that the phosphorylation of VASPis required for the cAMP-dependent inhibition of HASM cellmigration.
Collectively, our data demonstrate that the albuterol-dependentinhibition of HASM cell migration is attributable to the b2AR-dependent and PKA-dependent phosphorylation of VASP.
Prolonged Exposure to Albuterol Abolishes the b2AR
Agonist-Dependent Phosphorylation of VASP
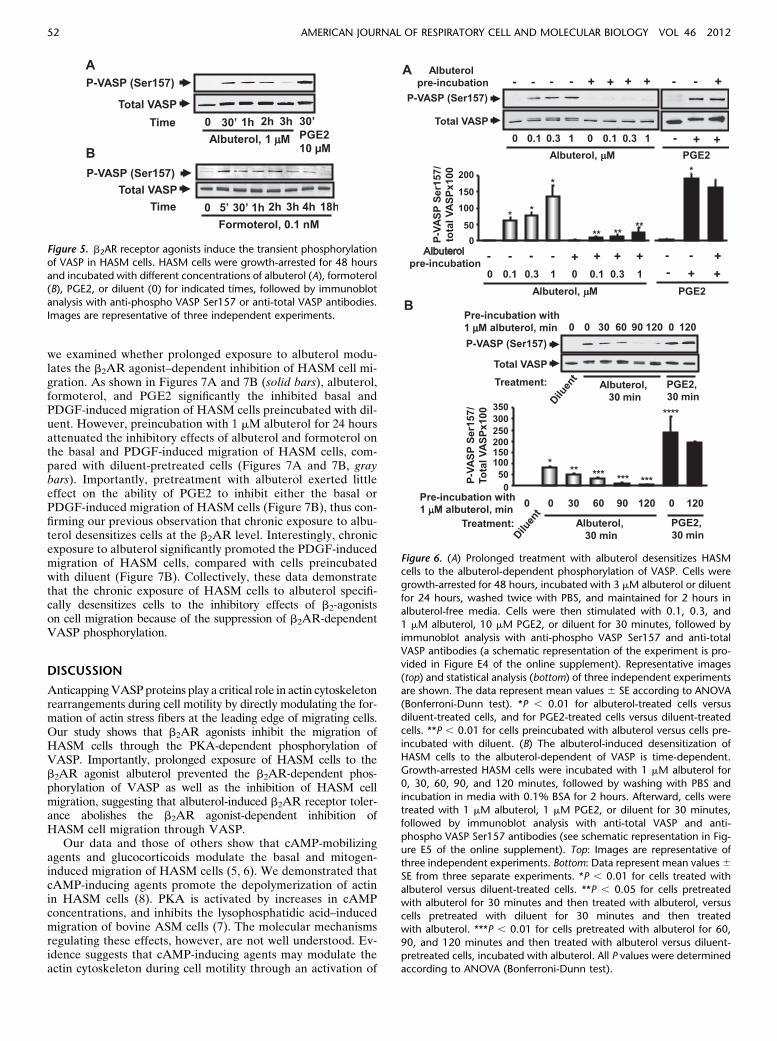
To determine whether prolonged treatment with b2AR ago-nists modulates the phosphorylation of VASP, we incubatedgrowth-arrested HASM cells with albuterol or formoterol forup to 18 hours, followed by immunoblot analysis with phospho-Ser157 VASP antibodies. As shown in Figure 5, both albuteroland formoterol induced a transient increase in the phosphoryla-tion of VASP that decreased by 3 hours of treatment.
To examine further the effects of prolonged exposure to b2ARagonists on the inhibition of VASP, HASM cells were preincu-bated with 3 mM albuterol or diluent for 24 hours and maintainedfor 2 hours in albuterol-free media, and then short-term treat-ment with albuterol was performed, followed by immunoblotanalysis to detect the phosphorylation of VASP at Ser157 (seeFigure E4 for a schematic representation). The dose and kineticsof b2AR tolerance were characterized as we previously described(19). We found that 0.1–1 mM albuterol induced a marked phos-phorylation of VASP at Ser157 in diluent-pretreated cells (Figure6A), but failed to induce the phosphorylation of VASP at Ser157in cells preincubated with albuterol for 24 hours (Figure 6A).Interestingly, the PGE2-dependent phosphorylation of VASPwas not suppressed by preincubation with albuterol (Figure6A), demonstrating that desensitization is attributable to altera-tions in the function of b2AR.
To determine the time required for the desensitization ofb2AR, HASM cells were preincubated for 30–120 minutes with1 mM albuterol, followed by the removal of albuterol and short-term treatment with 1 mM albuterol (a schematic representationof this experiment is provided in Figure E5). As shown in Figure6B, the level of desensitization was dependent on preincubationtime. Preincubation with 1 mM albuterol for 30–120 minutes ledto the time-dependent inhibition of VASP phosphorylation, andthe suppression of VASP phosphorylation was detected after120 minutes of preincubation (Figure 6B). Thus, prolongedincubation with albuterol inhibits the albuterol-dependent phos-phorylation of VASP in a time-dependent and concentration-dependent manner, potentially because of alterations in theresponsiveness of b2AR.
Chronic Exposure to Albuterol Abolishes Albuterol-Dependent
and Formoterol-Dependent Inhibition of Migration of HASM
Cells, and Further IncreasesMigratory Potential of HASMCells
Because our data demonstrate that albuterol inhibits the migra-tion of HASM cell attributable to the phosphorylation of VASP,
Figure 4. (A) The activation of protein kinase A (PKA) is required for the
albuterol-dependent inhibition of HASM cell migration. HASM cells were
stably transfected with green fluorescent protein (GFP)-tagged proteinkinase inhibitor (PKI) peptide (PKI-GFP), the dominant negative mutant
of regulatory PKA subunit (RevAB) (RevAB-GFP), or control GFP, and were
serum-deprived for 48 hours, and then migration in the presence or
absence of 10 mM albuterol and 10 ng/ml PDGF was examined. Themigration of diluent-treated cells stably expressing GFP was taken as 1-
fold. Data represent the mean values 6 SE of two independent experi-
ments and three repetitions of each experiment, according to ANOVA
(Bonferroni Dunn test). (B and C) Small interfering (si) RNA VASP sup-presses the PGE2-dependent phosphorylation of VASP and attenuates
the PGE2-induced inhibition of HASM cell migration. (B) HASM cells,
transfected with 100 or 250 nM siRNA VASP, or control siGLO RISC-Free(GLO) siRNA, were growth-arrested and treated with 10 mM PGE2 or
diluent for 30 minutes, and then immunoblot analysis with anti-phospho
VASP Ser157 and anti-total VASP antibodies was performed. (C) Cells,
transfected with 100 or 250 nM siRNA VASP or control siRNA, weresubjected to a migration assay in Boyden chamber in the presence or
absence of 10 mM PGE2 and 10 ng/ml PDGF. The data represent mean
values6 SE from three independent experiments. *P, 0.001 for control
siRNA 1 PGE2 versus control siRNA. **P , 0.01 for siRNA VASP 1 PGE2versus control siRNA 1 PGE2. All P values were determined according to
ANOVA (Bonferroni-Dunn test).
Goncharova, Goncharov, Zhao, et al.: VASP, b2-Receptor Agonists, and HASM Migration 51
we examined whether prolonged exposure to albuterol modu-lates the b2AR agonist–dependent inhibition of HASM cell mi-gration. As shown in Figures 7A and 7B (solid bars), albuterol,formoterol, and PGE2 significantly the inhibited basal andPDGF-induced migration of HASM cells preincubated with dil-uent. However, preincubation with 1 mM albuterol for 24 hoursattenuated the inhibitory effects of albuterol and formoterol onthe basal and PDGF-induced migration of HASM cells, com-pared with diluent-pretreated cells (Figures 7A and 7B, graybars). Importantly, pretreatment with albuterol exerted littleeffect on the ability of PGE2 to inhibit either the basal orPDGF-induced migration of HASM cells (Figure 7B), thus con-firming our previous observation that chronic exposure to albu-terol desensitizes cells at the b2AR level. Interestingly, chronicexposure to albuterol significantly promoted the PDGF-inducedmigration of HASM cells, compared with cells preincubatedwith diluent (Figure 7B). Collectively, these data demonstratethat the chronic exposure of HASM cells to albuterol specifi-cally desensitizes cells to the inhibitory effects of b2-agonistson cell migration because of the suppression of b2AR-dependentVASP phosphorylation.
DISCUSSION
AnticappingVASPproteins play a critical role in actin cytoskeletonrearrangements during cell motility by directly modulating the for-mation of actin stress fibers at the leading edge of migrating cells.Our study shows that b2AR agonists inhibit the migration ofHASM cells through the PKA-dependent phosphorylation ofVASP. Importantly, prolonged exposure of HASM cells to theb2AR agonist albuterol prevented the b2AR-dependent phos-phorylation of VASP as well as the inhibition of HASM cellmigration, suggesting that albuterol-induced b2AR receptor toler-ance abolishes the b2AR agonist-dependent inhibition ofHASM cell migration through VASP.
Our data and those of others show that cAMP-mobilizingagents and glucocorticoids modulate the basal and mitogen-induced migration of HASM cells (5, 6). We demonstrated thatcAMP-inducing agents promote the depolymerization of actinin HASM cells (8). PKA is activated by increases in cAMPconcentrations, and inhibits the lysophosphatidic acid–inducedmigration of bovine ASM cells (7). The molecular mechanismsregulating these effects, however, are not well understood. Ev-idence suggests that cAMP-inducing agents may modulate theactin cytoskeleton during cell motility through an activation of
Figure 5. b2AR receptor agonists induce the transient phosphorylation
of VASP in HASM cells. HASM cells were growth-arrested for 48 hours
and incubated with different concentrations of albuterol (A), formoterol(B), PGE2, or diluent (0) for indicated times, followed by immunoblot
analysis with anti-phospho VASP Ser157 or anti-total VASP antibodies.
Images are representative of three independent experiments.
Figure 6. (A) Prolonged treatment with albuterol desensitizes HASMcells to the albuterol-dependent phosphorylation of VASP. Cells were
growth-arrested for 48 hours, incubated with 3 mM albuterol or diluent
for 24 hours, washed twice with PBS, and maintained for 2 hours in
albuterol-free media. Cells were then stimulated with 0.1, 0.3, and1 mM albuterol, 10 mM PGE2, or diluent for 30 minutes, followed by
immunoblot analysis with anti-phospho VASP Ser157 and anti-total
VASP antibodies (a schematic representation of the experiment is pro-vided in Figure E4 of the online supplement). Representative images
(top) and statistical analysis (bottom) of three independent experiments
are shown. The data represent mean values 6 SE according to ANOVA
(Bonferroni-Dunn test). *P , 0.01 for albuterol-treated cells versusdiluent-treated cells, and for PGE2-treated cells versus diluent-treated
cells. **P , 0.01 for cells preincubated with albuterol versus cells pre-
incubated with diluent. (B) The albuterol-induced desensitization of
HASM cells to the albuterol-dependent of VASP is time-dependent.Growth-arrested HASM cells were incubated with 1 mM albuterol for
0, 30, 60, 90, and 120 minutes, followed by washing with PBS and
incubation in media with 0.1% BSA for 2 hours. Afterward, cells were
treated with 1 mM albuterol, 1 mM PGE2, or diluent for 30 minutes,followed by immunoblot analysis with anti-total VASP and anti-
phospho VASP Ser157 antibodies (see schematic representation in Fig-
ure E5 of the online supplement). Top: Images are representative ofthree independent experiments. Bottom: Data represent mean values6SE from three separate experiments. *P , 0.01 for cells treated with
albuterol versus diluent-treated cells. **P , 0.05 for cells pretreated
with albuterol for 30 minutes and then treated with albuterol, versuscells pretreated with diluent for 30 minutes and then treated
with albuterol. ***P , 0.01 for cells pretreated with albuterol for 60,
90, and 120 minutes and then treated with albuterol versus diluent-
pretreated cells, incubated with albuterol. All P values were determinedaccording to ANOVA (Bonferroni-Dunn test).
52 AMERICAN JOURNAL OF RESPIRATORY CELL AND MOLECULAR BIOLOGY VOL 46 2012

PKA, which regulates the activity of VASP by specific phos-phorylation at Ser157. The PKA-dependent of VASP was dem-onstrated in rat aortic vascular smooth muscle cells (20). Thestimulation of human neutrophils by a chemoattractant inducesthe PKA-dependent phosphorylation of VASP at Ser157 (21).VASP also plays a critical role in neuronal function by modu-lating the initiation of neurites and the outgrowth and guidanceof axons (22). The negative regulatory function of VASP phos-phorylation in cell migration was also suggested (23).
Here, we provide evidence linking theb2ARagonist–dependentphosphorylation of VASP to the inhibition of HASM cell migra-tion induced by cAMP-mobilizing agents. Our data show thatshort-term treatment with b2AR agonists induces the b2AR-dependent and PKA-dependent phosphorylation of VASP re-quired for the b2AR agonist–induced inhibition of HASM cellmigration.
Importantly, the stimulation of HASM cells with albuterol orformoterol induced a transient phosphorylation of VASP that wascompletely abolished after 3 hours of treatment with albuterol and18 hours of treatment with formoterol. Because b2AR agonistsare currently used as asthma therapeutics, we investigatedwhether HASM cells chronically treated with b2AR agonists willmaintain their responsiveness to the albuterol-dependent andformoterol-dependent phosphorylation of VASP and their inhi-bition of cell migration. Interestingly, a previous report by Hastieand colleagues demonstrated a decreased phosphorylation ofVASP in epithelial cells derived from patients with asthma whohad been chronically treated with b2AR agonists (24). We reportthat prolonged treatment with albuterol decreases the respon-siveness of HASM cells to b2AR agonists. Thus, albuterol failedto induce the phosphorylation of VASP in HASM cells treatedwith albuterol for 24 hours. Further, preincubation with albuterol
for 24 hours reversed the ability of albuterol and formoterol toinhibit the migration of HASM cells. Our data demonstrate thatprolonged treatment with b2AR agonists desensitizes HASM tob2AR agonists, and suggest that the phosphorylation level ofVASP may serve as a potential biomarker of airway responsive-ness to b2AR agonists and b2AR HASM desensitization.
Interestingly, pretreatment with albuterol did not affect theinhibitory effects of PGE2 on the phosphorylation of VASPand of HASM cells, suggesting a differential mechanism ofthe level of receptor or downstream signaling. Our previouslypublished studies demonstrated that PGE2 promotes thecAMP-dependent activation of PKA and phosphorylation ofVASP in HASM cells via the prostaglandin E2 (EP2) receptor(25, 26), which, in contrast to b2AR, is relatively resistant toagonist-specific desensitization. Indeed, the accumulation ofcAMP and of PKA during both acute and chronic with agonistswere markedly greater with PGE2 compared with b2AR ago-nists (14, 25–28) with regard to greater efficiency in the inhibi-tion of HASM cell growth (26, 29) and migration (6). Indeed,our published data show that, in contrast to b2AR agonists thattransiently increase the phosphorylation of VASP but decreasein effectiveness by 3 hours, PGE2 promotes a PKA-dependentphosphorylation of VASP that is sustained for up to 18 hours,suggesting that prolonged exposure to PGE2 does not desensi-tize EP2 receptors (25). Such differences between b-agonistsand PGE2 may be explained by differential mechanisms ofb2AR and EP2 receptor desensitization in HASM cells. Ourprevious data demonstrate that, in HASM cells, the desensitiza-tion of the b2AR receptor is selectively mediated by G protein-coupled receptor kinases 2/3 (GRK2/3) that exert little effect onEP2 receptor functions (26).
Our data demonstrate that the desensitization of VASP tob2AR agonists impairs the inhibitory effects of b2-agonists andpromotes the migration of HASM cells. In terms of clinical sig-nificance, our findings suggest that the chronic use of b2-agonistscould enhance the migration of ASM cells or prime them for anenhanced mitogenic response to growth factors. Ultimately, theenhanced migration of HASM cells and mitogenic responses maypromote an asthma phenotype manifested by irreversible airflowobstruction. Although our data suggest that such mechanismsmay be operative in chronic severe asthma, further clinical stud-ies are needed to characterize the role of VASP definitively in themigration and proliferation of HASM cells in mediating the ir-reversible airflow obstruction of chronic asthma.
Understanding the cellular and molecular mechanisms ofHASM remodeling is critically important, not only for advancingour knowledge of disease pathobiology, but also because suchan understanding may lead to indentifying novel molecular tar-gets for therapeutic intervention. This study identifies a mecha-nism of the inhibitory effects of b2AR agonists on the migrationof HASM cells. We found that the b2AR agonist–inducedactivation of PKA modulates the phosphorylation of VASP thatinhibits the migration of HASM cells. Furthermore, we foundthat the prolonged treatment of HASM with b2AR agonistsdesensitizes airways to further treatment with b2AR agonists.Our data suggest that a prolonged use of b2AR agonists inasthma as single therapy may promote the development of tol-erance to b2AR agonists and the migration of ASM cells.
Author Disclosure: The institution of R.B.P. has received grants and grants pend-ing from GSK for independent industry. R.A.P. has received consultancy fees andlecture fees from Merck and AstraZeneca, and he has received grants and grantspending from Merck and AstraZeneca. E.A.G., D.A.G., V.P.K., and H.Z. do nothave a financial relationship with a commercial entity that has an interest in thesubject of this manuscript.
Acknowledgments: Human tissue was obtained from the National Disease Re-search Interchange (Philadelphia, PA).
Figure 7. Prolonged exposure to albuterol prevents the albuterol-dependent and formoterol-dependent, but not PGE2-dependent, inhi-
bition of HASM cell migration. Cells were serum-deprived for 24 hours
and preincubated with 1 mM albuterol or vehicle for 24 hours, and
then basal (A) or PDGF-induced (B) migration in the Boyden chamberwas performed for 4 hours in the presence of 1 mM albuterol, 1 mM
formoterol, 1 mM PGE2, or diluent. The data represent mean values 6SE from three separate experiments, with three repetitions of eachexperiment. *P , 0.05 for albuterol and formoterol vs. diluent; **P ,0.001 for PGE2 vs. diluent by ANOVA (Bonferroni-Dunn test). The
migration of vehicle-pretreated cells in the presence of diluent was
taken as 1-fold.
Goncharova, Goncharov, Zhao, et al.: VASP, b2-Receptor Agonists, and HASM Migration 53

References
1. Gerthoffer WT. Migration of airway smooth muscle cells. Proc Am
Thorac Soc 2008;5:97–105.
2. Bentley JK, Hershenson MB. Airway smooth muscle growth in asthma:
proliferation, hypertrophy, and migration. Proc Am Thorac Soc 2008;
5:89–96.
3. Joubert P, Hamid Q. Role of airway smooth muscle in airway remod-
eling. J Allergy Clin Immunol 2005;116:713–716.
4. Gizycki MJ, Adelroth E, Rogers AV, O’Byrne PM, Jeffery PK. Myofi-
broblast involvement in the allergen-induced late response in mild
atopic asthma. Am J Respir Cell Mol Biol 1997;16:664–673.
5. Carlin SM, Roth M, Black JL. Urokinase potentiates PDGF-induced
chemotaxis of human airway smooth muscle cells. Am J Physiol
Lung Cell Mol Physiol 2003;284:L1020–L1026.
6. Goncharova EA, Billington CK, Irani C, Vorotnikov AV, Tkachuk VA,
Penn RB, Krymskaya VP, Panettieri RA Jr. Cyclic AMP–mobilizing
agents and glucocorticoids modulate human smooth muscle cell mi-
gration. Am J Respir Cell Mol Biol 2003;29:19–27.
7. HirakawaM, Karashima Y,Watanabe M, Kimura C, Ito Y, Oike M. Protein
kinase A inhibits lysophosphatidic acid–induced migration of airway
smooth muscle cells. J Pharmacol Exp Ther 2007;321:1102–1108.
8. Hirshman CA, Zhu D, Pertel T, Panettieri RA Jr, Emala CW. Isopro-
terenol induces actin depolymerization in human airway smooth
muscle cells via activation of an SRC kinase and GS. Am J Physiol
Lung Cell Mol Physiol 2005;288:L924–L931.
9. Krause M, Dent EW, Bear JE, Loureiro JJ, Gertler FB. ENA/VASP
proteins: regulators of the actin cytoskeleton and cell migration. Annu
Rev Cell Dev Biol 2003;19:541–564.
10. Iwasaki T, Chen JD, Kim JP, Wynn KC, Woodley DT. Dibutyryl cyclic
AMP modulates keratinocyte migration without alteration of integrin
expression. J Invest Dermatol 1994;102:891–897.
11. Edin ML, Howe AK, Juliano RL. Inhibition of PKA blocks fibro-
blast migration in response to growth factors. Exp Cell Res 2001;270:
214–222.
12. Ydrenius L, Molony L, Ng-Sikorski J, Andersson T. Dual action of
cAMP-dependent protein kinase on granulocyte movement. Biochem
Biophys Res Commun 1997;235:445–450.
13. Panettieri RA Jr, Murray RK, DePalo LR, Yadvish PA, Kotlikoff MI. A
human airway smooth muscle cell line that retains physiological re-
sponsiveness. Am J Physiol Cell Physiol 1989;256:C329–C335.
14. Guo M, Pascual RM, Wang S, Fontana MF, Valancius CA, Panettieri
RA Jr, Tilley SL, Penn RB. Cytokines regulate beta-2–adrenergic
receptor responsiveness in airway smooth muscle via multiple PKA-
and EP2 receptor–dependent mechanisms. Biochemistry 2005;44:
13771–13782.
15. Goncharova EA, Goncharov DA, Damera G, Tliba O, Amrani Y,
Panettieri RA Jr, Krymskaya VP. Signal transducer and activator of
transcription 3 is required for abnormal proliferation and survival of
TSC2-deficient cells: relevance to pulmonary lymphangioleiomyo-
matosis. Mol Pharmacol 2009;76:766–777.
16. Goncharova EA, Goncharov DA, Krymskaya VP. Assays for in vitro
monitoring of human airway smooth muscle (ASM) and human pul-
monary arterial vascular smooth muscle (VSM) cell migration. Nat
Protoc 2007;1:2933–2939.
17. Rosenborg J, Bengtsson T, Larsson P, Blomgren A, Persson G, Lotvall J.
Relative systemic dose potency and tolerability of inhaled formoterol
and salbutamol in healthy subjects and asthmatics. Eur J Clin Phar-
macol 2000;56:363–370.
18. Page CP, Morley J. Contrasting properties of albuterol stereoisomers.
J Allergy Clin Immunol 1999;104:S31–S41.
19. Cooper PR, Panettieri RA Jr . Steroids completely reverse albuterol-
induced [beta]2-adrenergic receptor tolerance in human small air-
ways. J Allergy Clin Immunol 2008;122:734–740.
20. Gros R, Ding Q, Chorazyczewski J, Pickering JG, Limbird LE, Feldman
RD. Adenylyl cyclase isoform–selective regulation of vascular smooth
muscle proliferation and cytoskeletal reorganization. Circ Res 2006;
99:845–852.
21. Eckert RE, Jones SL. Regulation of VASP serine 157 phosphorylation in
human neutrophils after stimulation by a chemoattractant. J Leukoc
Biol 2007;82:1311–1321.
22. Drees F, Gertler FB. ENA/VASP: proteins at the tip of the nervous
system. Curr Opin Neurobiol 2008;18:53–59.
23. Machesky LM. Putting on the brakes: a negative regulatory function for
ENA/VASP proteins in cell migration. Cell 2000;101:685–688.
24. Hastie AT, Wu M, Foster GC, Hawkins GA, Batra V, Rybinski KA,
Cirelli R, Zangrilli JG, Peters SP. Alterations in vasodilator-
stimulated phosphoprotein (VASP) phosphorylation: associations
with asthmatic phenotype, airway inflammation and beta2-agonist
use. Respir Res 2006;7:25.
25. Yan H, Deshpande DA, Misior AM, Miles MC, Saxena H, Riemer EC,
Pascual RM, Panettieri RA, Penn RB. Anti-mitogenic effects of
b-agonists and PGE2 on airway smooth muscle are PKA dependent.
FASEB J 2011;25:389–397.
26. Kong KC, Gandhi U, Martin TJ, Anz CB, Yan H, Misior AM, Pascual
RM, Deshpande DA, Penn RB. Endogenous Gs-coupled receptors in
smooth muscle exhibit differential susceptibility to GRK2/3-mediated
desensitization. Biochemistry 2008;47:9279–9288.
27. Penn RB, Panettieri RA, Benovic JL. Mechanisms of acute desensiti-
zation of the beta 2AR–adenylyl cyclase pathway in human airway
smooth muscle. Am J Respir Cell Mol Biol 1998;19:338–348.
28. Penn RB, Pascual RM, Kim YM, Mundell SJ, Krymskaya VP, Pan-
ettieri RA, Benovic JL. Arrestin specificity for G protein–coupled
receptors in human airway smooth muscle. J Biol Chem 2001;276:
32648–32656.
29. Misior AM, Yan H, Pascual RM, Deshpande DA, Panettieri RA, Penn
RB. Mitogenic effects of cytokines on smooth muscle are critically
dependent on protein kinase A and are unmasked by steroids and
cyclooxygenase inhibitors. Mol Pharmacol 2008;73:566–574.
54 AMERICAN JOURNAL OF RESPIRATORY CELL AND MOLECULAR BIOLOGY VOL 46 2012


















