
Autism New Neurobiology
7
NEUROLOGICAL REVIEW SECTION EDITOR: DAVID E. PLEASURE, MD The New Neurobiology of Autism Cortex, Connectivity, and Neuronal Organization Nancy J. Minshew, MD; Diane L. Williams, PhD T his review covers a fraction of the new research developments in autism but estab- li shes the basic el ements of the new neur obiologi c understanding of autism. Autism is a polygenetic developmental neurobiologic disorder with multiorgan system involve- ment, though it predominantly involves central nervous system dysfunction. The evi- dence supports autism as a disorder of the association cortex, both its neurons and their projec- tio ns. In par tic ula r, it is a dis ord er of connec tiv ity , whi ch app ear s, fro m cur ren t evi den ce, to pri mar il y inv olve int rah emi spheric con nec tiv ity . The foc us of connectivity st udies thus far has bee n on white matter, but alterations in functional magnetic resonance imaging activation suggest that intracor- tical connectivity is also likely to be disturbed. Furthermore, the disorder has a broad impact on cognitive and neurologic functioning. Deficits in high-functioning individuals occur in process- ing that places high demands on integration of information and coordination of multiple neural systems. Intact or enhanced abilities share a dependence on low information-processing demands and loc al neur al connect ions. Thi s mul tid omain model wit h sha red charact eri stics pre dicts an un- der ly ing pat hop hys iol ogi c mec han ism tha t impacts the bra in bro adl y, acc ord ing to a common neu - robiologic principle. The multiorgan system involvement and diversity of central nervous system findi ngs sugg est an epige netic mechan ism. ArchNeurol.2007;64(7):945-950 In the Diagnostic and Statistical Manual of Mental Disorders (Fourth Edition), 1 au- tism is theproto type forthe cat eg orycall ed pervasive develo pmenta l disorders. Of the pervasiv e develop mental disorders , autis- tic disorder, Asperger disorder, and per- vasi ve deve lopmental diso rder not other- wis e spe cifi ed are info rma lly refe rred to as the autism spectrum disorders (ASDs), which may affect up to 1% of children. 2 Pervasi ve developmental disorders are characterized by impairments in social ski lls, the communi cati ve use of verb al and non ver bal language, and res tricted and re- petitive behaviors. Behaviors in the re- stric ted and repetitiv e cat eg ory sha re a fo- cus on details and an inability to grasp concepts. 3 Though it is not the focus of this re- vi ew, it is important to appreciate tha t au- ti sm is a po ly ge netic di sorder with a heri- tability index of 0.90. 4 The search for contr ibuti ng ge nes has le d to ma ny pr om- ising but not yet replicated reports of as- sociated genes. A recent study reported a significant genetic asso ciati on betw een au- tism and a C allele in the promoter re- gion of the MET receptor tyrosine kinase gene that codes for a protein that relays signals turning on a cell’s internal signal- ing cascades that can result in increased proliferatio n, motility , differentiatio n, pro- cess outgrowth, or survival. The stron- ges t ass oci ati on occur red in famili es wit h more than 1 affected child. 5 Inheriting 2 cop iesof thi s all ele(the CC geno type ) con- ferred a 2.27 relative risk of autism diag- nosis and a 2-fold decrease in MET pro- mote r acti vity and bind ing of trans cript ion factor complex es. MET signaling is im- Author Affiliations: Departments of Psychiatry and Neurology, University of Pittsburgh School of Medicine, Pittsburgh, Pennsylvania (Dr Minshew); and Department of Speech-Language Pathology, Duquesne University, Pittsburgh (Dr Williams). (REPR INTED) ARCH NEUROL/ VOL 64 (NO. 7), JULY 2007 WWW.ARCHNEUROL.COM 945 ©2007 American Medical Association. All rights reserved. at Capes Consortia, on February 4, 2008 www.archneurol.com Downloaded from
-
Upload
mariana-keres -
Category
Documents
-
view
221 -
download
0
Transcript of Autism New Neurobiology

8/12/2019 Autism New Neurobiology
http://slidepdf.com/reader/full/autism-new-neurobiology 1/7
NEUROLOGICAL REVIEW
SECTION EDITOR: DAVID E. PLEASURE, MD
The New Neurobiology of AutismCortex, Connectivity, and Neuronal Organization
Nancy J. Minshew, MD; Diane L. Williams, PhD
T his review covers a fraction of the new research developments in autism but estab-lishes the basic elements of the new neurobiologic understanding of autism. Autism isa polygenetic developmental neurobiologic disorder with multiorgan system involve-ment, though it predominantly involves central nervous system dysfunction. The evi-
dence supports autism as a disorder of the association cortex, both its neurons and their projec-
tions. In particular, it is a disorder of connectivity, which appears, from current evidence, to primarilyinvolve intrahemispheric connectivity. The focus of connectivity studies thus far has been on whitematter, but alterations in functional magnetic resonance imaging activation suggest that intracor-tical connectivity is also likely to be disturbed. Furthermore, the disorder has a broad impact oncognitive and neurologic functioning. Deficits in high-functioning individuals occur in process-ing that places high demands on integration of information and coordination of multiple neuralsystems. Intact or enhanced abilities share a dependence on low information-processing demandsand local neural connections. This multidomain model with shared characteristics predicts an un-derlying pathophysiologic mechanism that impacts the brain broadly, according to a common neu-robiologic principle. The multiorgan system involvement and diversity of central nervous systemfindingssuggestan epigeneticmechanism. ArchNeurol.2007;64(7):945-950
In the Diagnostic and Statistical Manual of Mental Disorders (Fourth Edition), 1 au-tism is theprototype forthe categorycalledpervasivedevelopmental disorders. Of thepervasive developmental disorders, autis-tic disorder, Asperger disorder, and per-vasive developmental disorder not other-wise specified are informallyreferred to asthe autism spectrum disorders (ASDs),which may affect up to 1% of children. 2
Pervasive developmental disorders arecharacterized by impairments in socialskills, thecommunicativeuseof verbal andnonverbal language,and restrictedand re-petitive behaviors. Behaviors in the re-strictedandrepetitive categoryshare a fo-cus on details and an inability to graspconcepts. 3
Though it is not the focus of this re-view, it is important to appreciate that au-tism is a polygenetic disorder with a heri-tability index of 0.90. 4 The search forcontributing geneshas led to many prom-ising but not yet replicated reports of as-sociated genes. A recent study reported asignificant genetic association betweenau-tism and a C allele in the promoter re-gion of the MET receptor tyrosine kinasegene that codes for a protein that relays
signals turning on a cell’s internal signal-ing cascades that can result in increasedproliferation, motility, differentiation, pro-cess outgrowth, or survival. The stron-gest association occurred in families withmore than 1 affected child. 5 Inheriting 2copiesof this allele(the CC genotype) con-ferred a 2.27 relative risk of autism diag-nosis and a 2-fold decrease in MET pro-moter activityandbindingof transcriptionfactor complexes. MET signaling is im-
Author Affiliations: Departments of Psychiatry and Neurology, University of Pittsburgh School of Medicine, Pittsburgh, Pennsylvania (Dr Minshew); andDepartment of Speech-Language Pathology, Duquesne University, Pittsburgh(Dr Williams).
(REPRINTED) ARCH NEUROL/ VOL 64 (NO. 7), JULY 2007 WWW.ARCHNEUROL.COM945
©2007 American Medical Association. All rights reserved. at Capes Consortia, on February 4, 2008www.archneurol.comDownloaded from

8/12/2019 Autism New Neurobiology
http://slidepdf.com/reader/full/autism-new-neurobiology 2/7
portant in neocortical and cerebellar development, im-
mune systemfunction,andgastrointestinal repair, allsys-tems thathave manifestedsigns of dysfunction in autism.Thus, this gene provides a mechanism for simultaneousmultiorgan dysfunction in autism, compared with sometheories that have hypothesized that immune system orgastrointestinal dysfunction causes braindysfunction andthus autism. This study grew out of prior research im-plicating chromosome 7q31 as a candidate gene regionand a cortical interneuron inhibitory hypothesis 6 basedon minicolumn abnormalities in autism that suggesteda possible deficiency in interneurons. 7 Such work exem-plifies the translational research nowpossibleas a resultof cumulating research findings and the new mecha-nisms identifiedthathave thepotential toexplain thecom-
plex array of findings in autism.Autism is characterized by abnormalities in complexbehavior, language, and cognition with mental retarda-tion in 70% of cases and seizures in 30% of cases, andthe absence of blindness, deafness, and long tract signsin 90% to 95% of cases without any cause for brain dys-function other than autism. From a behavioral neurol-ogy perspective, this constellation suggests generalizeddysfunction of the association cortex, with sparing of pri-mary sensory andmotor cortex andof white matter. Theabsence of clinical signs of focal brain dysfunction com-mon in children with hypoxic ischemic injury, such asvisuospatial deficits and cerebral palsy, further suggestsa distributed neural systemsabnormality. 8 Theabove be-
havioralneurologicalassessment waspossibleby theearly1980s, when secondary cases of autism were distin-guished from idiopathic cases. 9 However, it was not un-til thepast10yearsthat a neurobiology for autismmatch-ing this behavioral neurology emerged.
The contemporary neurobiology of autism began withevidence that group mean head circumference was at the60th to 70th percentile relative to population norms andwas disproportionate to height and weight. Fifteen per-cent to 20% of the autism group had macrocephaly (headcircumference 99th percentile). 10 Although larger head
circumference for height was the most common outcomeand true for the autism group as a whole, it was not uni-versal;headcircumference proportionate toheight andheadcircumference less than height was also observed in au-tism. 10 Thus, many head growth trajectories were consis-tent withautism. Head circumference data from very youngchildren with autismretrospectivelyfound onset of accel-erated head growth by 12 months of age 11 and macro-cephaly in15% to20% ofchildrenby4 to5 years ofage. 10
Structural magnetic resonance imaging studies con-firmed the increase in total brainvolumein autism, whichhad been inferred from the increased head circumfer-ence. The increase in total brain volume was docu-mented beginning at 2 to 4 years of age, 11,12 the earliestage of clinical recognition, and persisted into childhoodbutnot adolescence. 13 The tissuescontributing to thisin-creasewere total cerebral white matter and total corticalgray matter, with the latter contribution varying with theparcellation program used. One influential study of 6-to 11-year-old children parcellated cerebral white mat-ter into an outer zone of radiate white matter composedof intrahemispheric corticocortical connections and aninner zone of bridging and sagittal compartments
(Figure 1 ).14
The inner zone of white matter, espe-cially the corpus callosum and internal capsule, showedno volume increase. Thevolume of theouter radiate whitematter was increased in all cerebrallobes but with a fron-tal predominance. Collectively, these findings were in-terpreted asevidence ofovergrowthofshort-andmedium-range intrahemispheric corticocortical connections withno detectable involvement of interhemispheric connec-tions or connections betweencortexandsubcortical struc-tures. The onset of brain overgrowth coincidedwith theonset of the signs and symptoms of autism, indicatingthat theovergrowth was part of a pathologic process thatdisrupted thedevelopment of normal brain structure andfunction in autism.
Another study examined cortical connectivity in ananalogous way by comparing gyral and sulcal thicknessas indices of short- and longer-distance cortical connec-tions. 15 This study found an overall increase in corticalthickness in a sample of high-functioning 8- to 12-year-old boys with autism compared with typical boys. Cor-tical thickness in sulci (long connections) was greater(analogous to increased volume of outer radiate whitematter) than in gyri (short vertical connections), whichis comparable to thefindings ofHerbert andcolleagues 14
for white matter.A secondconsistent finding of cross-sectional imaging
studies in autism has been a reduction in the size of thecorpus callosum, though the segment affected has var-
ied (D.L.W., unpublished data, 2006). In some studies,the decrease was only present relative to the increase intotal brain volume in autism. In other studies, there wasan absolute decrease in corpus callosum size indepen-dent of brain volume. The contrast between the in-crease in intrahemispheric white matter volume and thelack of change or decrease in corpus callosum size is no-table, indicating that the neurobiological process affect-ing intrahemispheric white matter spares interhemi-spheric white matter. Although there is ample evidenceof intrahemispheric processing deficits in autism, the sta -
Figure 1. Graphical representation of nonuniform growth patterns in autisticbrain. Yellow areas indicate the outer radiate white matter white zone, whichare larger in volume in this study of children with autism than in the controls.The white area represents bridging and sagittal components, which did notdiffer in volume from controls. The volumes of areas in blue were absolutelybut not relatively different. Image courtesy of Martha Herbert, MD, PhD.14
(REPRINTED) ARCH NEUROL/ VOL 64 (NO. 7), JULY 2007 WWW.ARCHNEUROL.COM946
©2007 American Medical Association. All rights reserved. at Capes Consortia, on February 4, 2008www.archneurol.comDownloaded from
8/12/2019 Autism New Neurobiology
http://slidepdf.com/reader/full/autism-new-neurobiology 3/7
tus of interhemispheric processing remains an unan-swered but important research question.
Thenext significant contributionwasthe reportofmini-column abnormalities in autism. 7 Prior to this, there wereno substantive histopathologic abnormalities reported inthe cerebral cortex in autism, though all behavioral neu-rological evidence pointed to it as a primary site of brain
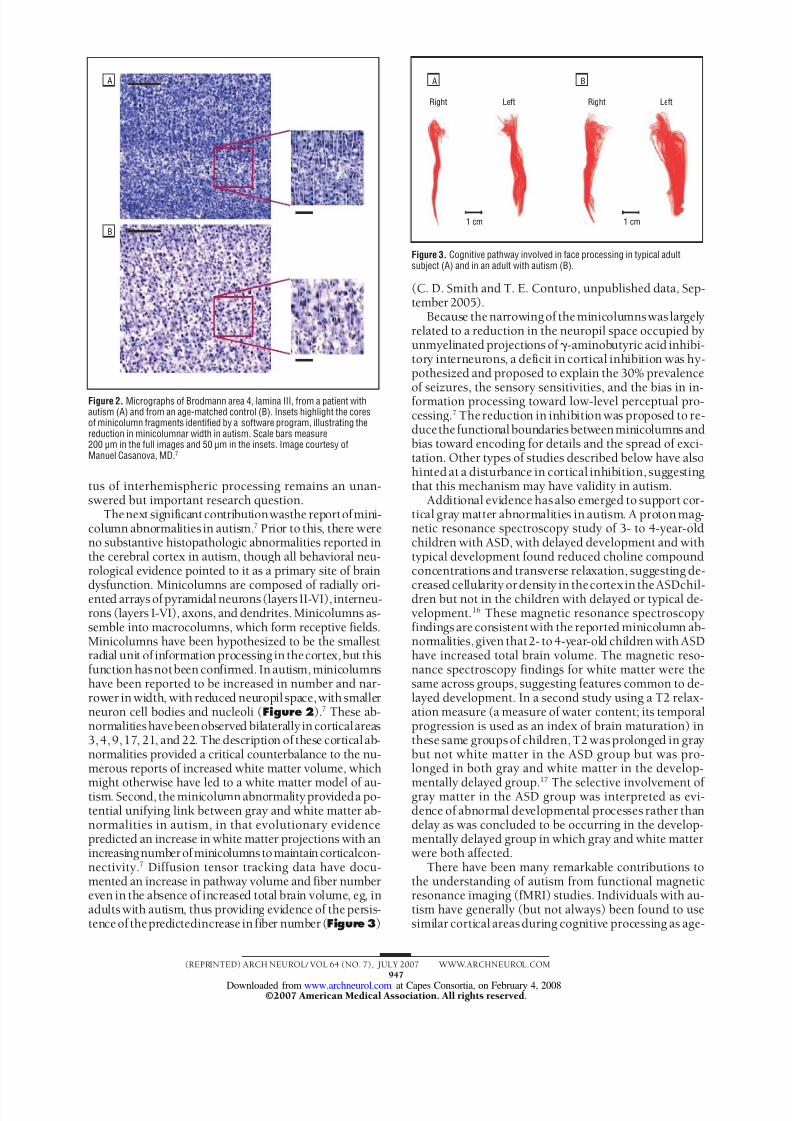
dysfunction. Minicolumns are composed of radially ori-ented arrays ofpyramidal neurons(layers II-VI), interneu-rons (layers I-VI), axons, and dendrites. Minicolumns as-semble into macrocolumns, which form receptive fields.Minicolumns have been hypothesized to be the smallestradial unit of information processing in thecortex,but thisfunction hasnotbeen confirmed. In autism, minicolumnshave been reported to be increased in number and nar-rower in width, with reduced neuropil space,with smallerneuron cell bodies and nucleoli ( Figure 2 ).7 These ab-normalitieshavebeenobserved bilaterally in corticalareas3,4, 9,17, 21, and 22. The description of these corticalab-normalities provided a critical counterbalance to the nu-merous reports of increased white matter volume, which
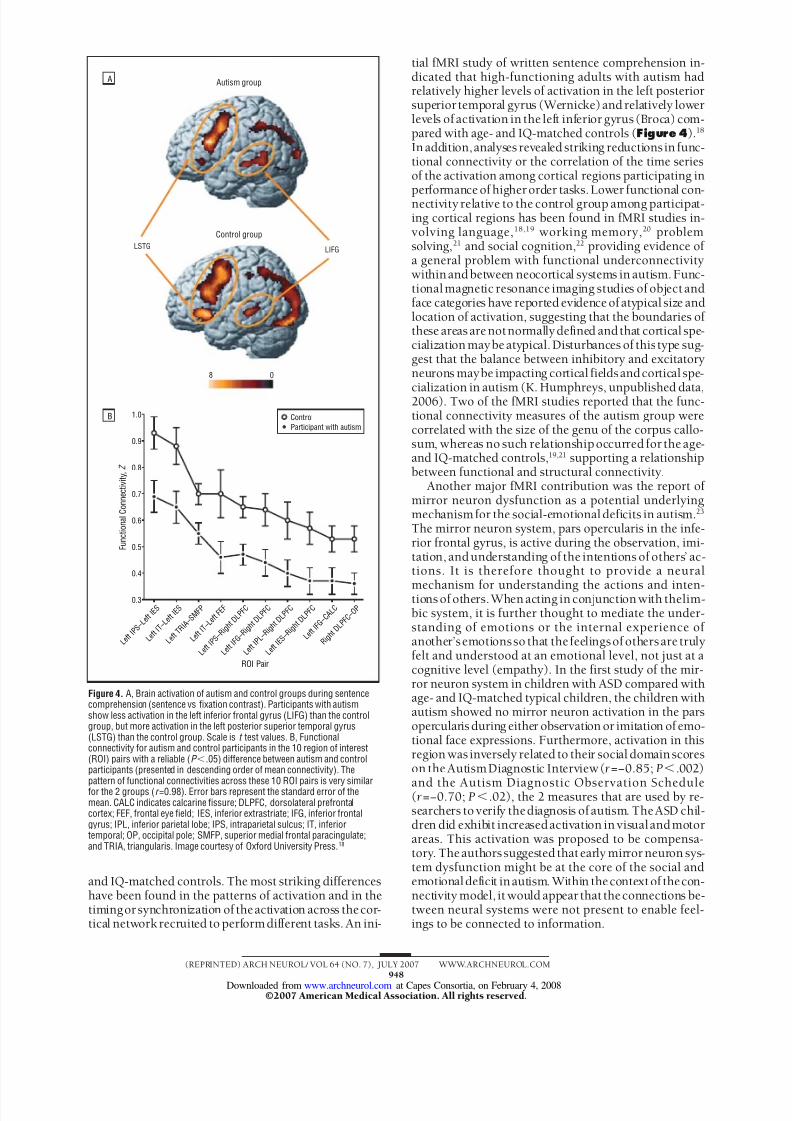
might otherwise have led to a white matter model of au-tism. Second, the minicolumn abnormalityprovideda po-tential unifying link between gray and white matter ab-normalities in autism, in that evolutionary evidencepredicted an increase in white matter projections with anincreasingnumberofminicolumnstomaintaincorticalcon-nectivity. 7 Diffusion tensor tracking data have docu-mented an increase in pathway volume and fiber numbereven in the absence of increased total brain volume, eg, inadults with autism, thus providing evidence of the persis-tenceof thepredictedincrease in fiber number ( Figure 3 )
(C. D. Smith and T. E. Conturo, unpublished data, Sep-tember 2005).
Because thenarrowingof theminicolumnswas largelyrelated to a reduction in the neuropil space occupied byunmyelinated projections of -aminobutyric acid inhibi-tory interneurons, a deficit in cortical inhibition was hy-pothesized and proposed to explain the 30% prevalence
of seizures, the sensory sensitivities, and the bias in in-formation processing toward low-level perceptual pro-cessing. 7 Thereduction in inhibition was proposed to re-ducethe functionalboundariesbetweenminicolumns andbias toward encoding for details and the spread of exci-tation. Other types of studies described below have alsohinted at a disturbance in cortical inhibition, suggestingthat this mechanism may have validity in autism.
Additional evidence hasalso emerged to support cor-tical gray matter abnormalities in autism. A protonmag-netic resonance spectroscopy study of 3- to 4-year-oldchildren with ASD, with delayed development and withtypical development found reduced choline compoundconcentrations and transverse relaxation, suggesting de-
creased cellularity ordensity in thecortexin theASDchil-dren but not in the children with delayed or typical de-velopment. 16 These magnetic resonance spectroscopyfindings are consistentwith the reported minicolumn ab-normalities,given that2- to 4-year-old children with ASDhave increased total brain volume. The magnetic reso-nance spectroscopy findings for white matter were thesame across groups, suggesting features common to de-layed development. In a second study using a T2 relax-ation measure (a measure of water content; its temporalprogression is used as an index of brain maturation) inthese same groupsof children,T2 wasprolonged in graybut not white matter in the ASD group but was pro-longed in both gray and white matter in the develop-
mentally delayed group.17
The selective involvement of gray matter in the ASD group was interpreted as evi-dence of abnormal developmental processes rather thandelay as was concluded to be occurring in the develop-mentally delayed group in which gray and white matterwere both affected.
There have been many remarkable contributions tothe understanding of autism from functional magneticresonance imaging (fMRI) studies. Individuals with au-tism have generally (but not always) been found to usesimilar cortical areasduring cognitive processing as age-
A
B
Figure 2. Micrographs of Brodmann area 4, lamina III, from a patient withautism (A) and from an age-matched control (B). Insets highlight the coresof minicolumn fragments identified by a software program, illustrating thereduction in minicolumnar width in autism. Scale bars measure200 µm in the full images and 50 µm in the insets. Image courtesy ofManuel Casanova, MD.7
Right Left
A
Right Left
B
1 cm 1 cm
Figure 3. Cognitive pathway involved in face processing in typical adultsubject (A) and in an adult with autism (B).
(REPRINTED) ARCH NEUROL/ VOL 64 (NO. 7), JULY 2007 WWW.ARCHNEUROL.COM947
©2007 American Medical Association. All rights reserved. at Capes Consortia, on February 4, 2008www.archneurol.comDownloaded from
8/12/2019 Autism New Neurobiology
http://slidepdf.com/reader/full/autism-new-neurobiology 4/7
and IQ-matched controls. The most striking differenceshave been found in the patterns of activation and in thetimingor synchronization of theactivation across thecor-tical network recruited to performdifferent tasks. An ini-
tial fMRI study of written sentence comprehension in-dicated that high-functioning adults with autism hadrelatively higher levels of activation in the left posteriorsuperior temporal gyrus (Wernicke)and relatively lowerlevels of activation in the left inferior gyrus (Broca) com-pared with age- and IQ-matched controls ( Figure 4 ).18
In addition,analyses revealed striking reductions in func-tional connectivity or the correlation of the time seriesof the activation among cortical regions participating in
performance of higherorder tasks.Lower functional con-nectivity relative to the control group among participat-ing cortical regions has been found in fMRI studies in-volving language, 18,19 working memory, 20 problemsolving, 21 and social cognition, 22 providing evidence of a general problem with functional underconnectivitywithinandbetween neocortical systems in autism.Func-tionalmagnetic resonance imaging studies of object andface categories have reported evidence of atypical size andlocation of activation, suggesting that the boundaries of these areas arenotnormallydefined andthat cortical spe-cialization maybe atypical.Disturbances of this type sug-gest that the balance between inhibitory and excitatoryneurons maybe impacting cortical fieldsandcortical spe-
cialization in autism (K. Humphreys, unpublished data,2006). Two of the fMRI studies reported that the func-tional connectivity measures of the autism group werecorrelated with the size of the genu of the corpus callo-sum, whereas no such relationshipoccurred for the age-and IQ-matched controls, 19,21 supporting a relationshipbetween functional and structural connectivity.
Another major fMRI contribution was the report of mirror neuron dysfunction as a potential underlyingmechanism for the social-emotional deficits in autism. 23
The mirror neuron system, pars opercularis in the infe-rior frontal gyrus, is active during the observation, imi-tation, andunderstanding of the intentions of others’ ac-tions. It is therefore thought to provide a neural
mechanism for understanding the actions and inten-tions of others.When acting in conjunction with thelim-bic system, it is further thought to mediate the under-standing of emotions or the internal experience of another’semotionsso that thefeelingsof others are trulyfelt and understood at an emotional level, not just at acognitive level (empathy). In the first study of the mir-ror neuron system in children with ASD compared withage- and IQ-matched typical children, the children withautism showed no mirror neuron activation in the parsopercularis during either observation or imitation of emo-tional face expressions. Furthermore, activation in thisregion was inversely related to their social domainscoreson the AutismDiagnostic Interview( r =−0.85; P .002)
and the Autism Diagnostic Observation Schedule(r =−0.70; P .02), the 2 measures that are used by re-searchers to verify thediagnosis of autism. TheASD chil-dren did exhibit increasedactivation in visualandmotorareas. This activation was proposed to be compensa-tory. Theauthors suggested that early mirror neuron sys-tem dysfunction might be at the core of the social andemotionaldeficit inautism. Within thecontextof thecon-nectivity model, it would appear that theconnections be-tween neural systems were not present to enable feel-ings to be connected to information.
A
B
Autism group
Control groupLSTG
8 0
LIFG
1.0
0.8
0.9
0.7
0.5
0.4
0.6
0.3
L e f t I P S – L e f t I E S
L e f t I T – L e f t I E S
L e f t T R I A –
S M F P
L e f t I T – L e f t F E F
L e f t I P S – R i g h t D
L P F C
L e f t I F G – R i g h t D
L P F C
L e f t I P L – R i g h t D
L P F C
L e f t I E S – R i g h t D
L P F C
L e f t I F G – C A L
C
R i g h t D L P F C –
O P
ROI Pair
F u n c t i o n a
l C o n n e c t i v
i t y ,
Z
ControlParticipant with autism
Figure 4. A, Brain activation of autism and control groups during sentencecomprehension (sentence vs fixation contrast). Participants with autismshow less activation in the left inferior frontal gyrus (LIFG) than the controlgroup, but more activation in the left posterior superior temporal gyrus(LSTG) than the control group. Scale is t test values. B, Functionalconnectivity for autism and control participants in the 10 region of interest(ROI) pairs with a reliable (P .05) difference between autism and controlparticipants (presented in descending order of mean connectivity). The
pattern of functional connectivities across these 10 ROI pairs is very similarfor the 2 groups (r =0.98). Error bars represent the standard error of themean. CALC indicates calcarine fissure; DLPFC, dorsolateral prefrontalcortex; FEF, frontal eye field; IES, inferior extrastriate; IFG, inferior frontalgyrus; IPL, inferior parietal lobe; IPS, intraparietal sulcus; IT, inferiortemporal; OP, occipital pole; SMFP, superior medial frontal paracingulate;and TRIA, triangularis. Image courtesy of Oxford University Press.18
(REPRINTED) ARCH NEUROL/ VOL 64 (NO. 7), JULY 2007 WWW.ARCHNEUROL.COM948
©2007 American Medical Association. All rights reserved. at Capes Consortia, on February 4, 2008www.archneurol.comDownloaded from

8/12/2019 Autism New Neurobiology
http://slidepdf.com/reader/full/autism-new-neurobiology 5/7
Autismis conventionally defined by 3 symptoms or af-fected neural systems that co-occur for no apparent rea-son. The choice is to view these as separate dimensionscoded forbydifferent genes 24,25 or toseeka commonchar-acteristic sharedbythesignsandsymptoms that mightex-plain their co-occurrence as a syndrome. Two studies ex-amined the profile of neuropsychologic functioning inautismto identifya sharedcharacteristic forthedeficits andfor the intact abilities. 26,27 The first profile study provided
theconceptual foundation forour fMRI studies that led tothe present connectivity model for autism. As the fieldevolved, itwas possible toalsomakethe point with thesec-ondprofilestudythat autismextendedcognitivelyand neu-rologically beyond the diagnostic triad and thus morebroadly involved the brain. Testing of 178 high-functioning autistic and control individuals aged 8 to 40years across both studies revealed deficits in higher corti-cal sensoryperception, skilledmotor abilities, memoryforcomplex stimuli that required detection or application of an organizing strategy, higher order languageabilities (de-tection or creation of story themes, metaphors, infer-ences, or idioms), and concept formation. Spared or en-hanced abilities included attention, elementary sensory
perceptual abilities, elementary motor skills, memory forsimple material dependent on basic associative skills, for-mal language skills, and the rule-learning aspects of ab-straction. The conclusions drawn from this pattern werethat (1) basic information acquisition abilities were in-tact, (2) impaired abilities shared a dependence on highdemandson processing information or integration, (3) in-tact abilities shared a dependence on low information-processing demands, and (4) within and across domains,skills or abilities were impacted in proportion to the de-mands for integration of information. These observationsled to the proposal that autism was a disorder of complexinformationprocessing withintact or enhancedsimple in-formationprocessing. 28 This constructprovided a reason-
able account for theco-occurrence of the signs andsymp-toms as a syndrome. It was also a valuable model forintervention and teaching. From a neurobiological per-spective, this concept suggested a developmental dis-turbance in neuronal organizational events with preser-vation or overdevelopment of local circuitry andunderdevelopment of the connections within and be-tweencorticalsystemsinhigh-functioning individualswithautism. In low-functioning individuals with autism, thereappears to be little to no development of functional con-nections between primary sensory cortex and associationcortex. Thecommon themesacross the spectrum,regard-less of severity, are cortical connectivity and information-processing capacity.
A secondmajor implication oftheneuropsychologicpro-file studies was that domains outside the classic diagnos-tictriad wereinvolved.Previously,sensoryandmotormani-festations were consideredassociatedsignsandsymptoms,and memory dysfunction (other than the disproved am-nesiahypothesis)wasnotthoughtto bepartofautism.How-ever,the involvementof theseareasoffunctionin the samepattern as the traditionalsignsandsymptoms impliedthatthese domains were impacted by the same neurobiologicprocess and that autism had a far broader impact on thebrain thanwasoriginally conceptualized. Thisconclusion
was reinforced by a large study of postural control fromchildhood to middle age in autism, which revealed de-layed onset of maturation of postural control with failureto achieve adult levels of function. 29 The postural instabil-ity was the result of reduced capacity for multimodal sen-soryintegration(visual, vestibular,andpositionsense).Thisstudy provided timely support for the emerging connec-tivity model of autismbut also for the concept that autismbroadlyaffected the brain, withinformation integration as
a common denominator.Anothernotwell-appreciatedaspect of thisclinical syn-drome is the second wave of deficits that emerges in thesecond decade of life. Frontal circuitry normally ma-tures during thistime period, andoculomotor, fMRI, andneuropsychologic studies of working memory and ex-ecutive function in autism have revealed the emergenceof new deficits in adolescence in autism, with the failureof frontal lobe skills to develop. 30 Previously, it wasthought that adolescents and young adults with autismfell further behind peers in adaptive function in the sec-onddecadebecause life becamemore challenging. In fact,the higher order frontal lobe skills needed to cope withthese life demands failed to develop in autism. This se-
vere frontal dysmaturity likely accounts for the unex-pectedly poor adaptive function in adult life in the ma- jority of high-functioning ASD individuals.
The structural imaging findings have led to the neargeneral acceptance of autismas a disorder originating inthe brain rather than in behavior, a subtle but signifi-cant distinction. (Social theories hold that lack of moti-vation to interact is the initiating event in autism and re-sults in failure of brain circuitry to develop. Thus, theunderdevelopment of fusiform face area is secondary tothe lack of eye contact, rather than underdevelopmentof thecircuitry for nonverbal language and fusiform facearea being a primary event that results in the observedbehavior.)Developmental alterations in whitematter and
their role in altered intrahemispheric connectivity re-flect a new understanding of the involvement of whitematter in childhood disorders and a departure from thetraditional injury model (cerebral palsy). These find-ings, together with fMRI studies, have shifted thinkingaway from autism as a disorder of regional brain dys-function to a model of autism as a large-scale neural sys-tems disorder with alterations in cortical systems con-nectivity. Recent findings have also implicated cerebralgray matter, creating a picture of disturbances in cere-bral gray andwhite matter connectivity.Studiesalso sug-gest that abnormalities in cortical synchronization andin the balance of cortical inhibition and excitation maybe mediating mechanisms. Various genetic mechanisms
arebeginningto be identified to account fornotonly mul-tiorgan involvement but also the diversity of the find-ings within the central nervous system.
The limitations of this review are many. Most no-table is the failure to touch on the richness of the litera-ture related to the social, emotional, gaze, face-proces-sing, andmotion-processingor motiondetectionnetworksand their associated cognitive and neurologic implica-tionsfor autism. Genetics, epigenetics, epidemiology, de-velopmental neurobiology, neurochemistry, and mousemodel contributions have also been neglected.
(REPRINTED) ARCH NEUROL/ VOL 64 (NO. 7), JULY 2007 WWW.ARCHNEUROL.COM949
©2007 American Medical Association. All rights reserved. at Capes Consortia, on February 4, 2008www.archneurol.comDownloaded from

8/12/2019 Autism New Neurobiology
http://slidepdf.com/reader/full/autism-new-neurobiology 6/7
Accepted for Publication: December 12, 2006.Correspondence: NancyJ.Minshew,MD,WebsterHall,Suite300, 3811 O’Hara St, Pittsburgh, PA 15213 ([email protected]).Author Contributions: Both authorscontributedequallyto this review. Study concept anddesign:MinshewandWil-liams. Analysis and interpretation of data: Minshew and Williams. Drafting of the manuscript: Minshew and Wil-liams. Critical revision of the manuscript for important in-
tellectual content: Minshew and Williams. Obtained fund-ing: Minshew and Williams. Administrative, technical, andmaterial support: Minshew and Williams.Financial Disclosure: None reported.Funding/Support: Thepreparationof thismanuscript wassupported by grant U19HD35469 from the National In-stitute of Child Health and Human Development(NICHD), a NICHD/National Institute on Deafness andOther Communication Disorders (NIDCD) Collabora-tive Program of Excellence in Autism. Support wasalso provided by grant K23DC006691 from NIDCD(Dr Williams).Additional Contributions: We express our apprecia-tion to all autism researchers who contributed their ad-
vice about what should be highlighted in this brief re-view and especially to those who provided figures.
REFERENCES
1. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 4th ed. Washington, DC: American Psychiatric Association; 1994.
2. Baird G, Simoff E, Pickles A, et al. Prevalence of disorders of the autism spec-trum in a population cohort of children in south Thames: the special needs andautism project (SNAP). Lancet . 2006;368(9531):210-215.
3. Minshew NJ, Meyer J, Goldstein G. Abstract reasoning in autism: a dissociationbetween concept formation and concept identification.Neuropsychology . 2002;16(3):327-334.
4. Folstein S. Twin andadoptionstudiesin childand adolescent psychiatric disorders.Curr Opin Pediatr . 1996;8(4):339-347.
5. Campbell DB, Sutcliffe JS, Ebert PJ, et al. A genetic variant that disrupts MET
transcription is associated with autism. Proc Natl Acad Sci U S A. 2006;103(45):16834-16839.
6. Levitt P. Disruption of interneuron development. Epilepsia . 2005;46(suppl 7):22-28.
7. Casanova MF, van Kooten IAJ, Switala AE, et al. Minicolumnar abnormalities inautism. Acta Neuropathol (Berl) . 2006;112(3):287-303.
8. Minshew NJ, Payton JB. New perspectives in autism, part II: the differential di-agnosis and neurobiology of autism. Curr Probl Pediatr . 1988;18(11):613-694.
9. Prior MR, Tress B, Hoffman WL, Boldt D. Computed tomographic study of chil-dren with classic autism. Arch Neurol . 1984;41(5):482-484.
10. Lainhart JE, Bigler ED, Bocian M, et al. Head circumference and height in au-
tism: a study by the Collaborative Program of Excellence in Autism. Am J Med Genet A. 2006;140(21):2257-2274.
11. Hazlett HC, Poe M, Gerig G, et al. Magnetic resonance imaging and head cir-cumference study of brain size in autism: birth through age 2 years. Arch Gen Psychiatry . 2005;62(12):1366-1376.
12. Courchesne E,KarnsCM, DavisHR, et al.Unusualbrain growthpatternsin earlylife in patients with autistic disorder: an MRI study. Neurology . 2001;57(2):245-254.
13. Lainhart JE. Advances in autism neuroimaging research for the clinician andgeneticist. Am J Med Genet C Semin Med Genet . 2006;142(1):33-39.
14. Herbert MR, Ziegler DA, Makris N, et al. Localization of white matter volume in-crease in autism and developmental language disorder. Ann Neurol . 2004;55(4):530-540.
15. HardanAY, Muddasani S, VemulapalliM, KeshavanM, MinshewNJ. An MRIstudyof increased cortical thickness in autism. Am J Psychiatry . 2006;163(7):1290-1292.
16. Friedman SD, Shaw DWW, Artru AA, et al. Gray and white matter brain chemis-try in young children with autism. Arch Gen Psychiatry . 2006;63(7):786-794.
17. Petropoulos H, Friedman SD, Shaw DWW, et al. Gray matter abnormalities inautism spectrum disorder revealed by T2 relaxation. Neurology . 2006;67(4):632-636.
18. Just MA, Cherkassky VL, Keller TA, Minshew NJ. Cortical activation and syn-chronization during sentence comprehension in high-functioning autism: evi-dence of underconnectivity. Brain . 2004;127(pt 8):1811-1821.
19. Kana RK, Keller TA, Cherkassky VL, Minshew NJ, Just MA. Sentence compre-hension in autism: thinking in pictures with decreased functional connectivity.Brain . 2006;129(pt 9):2484-2493.
20. Koshino H, Carpenter PA, Minshew NJ, Cherkassky VL, Keller TA, Just MA.
Functionalconnectivityin anfMRIworkingmemory taskin high-functioningautism.Neuroimage . 2005;24(3):810-821.21. CherkasskyVL, Keller TA,Kana RK,MinshewNJ. Functionaland anatomicalcor-
tical underconnectivity in autism: evidence from an fMRI study of an executivefunction task and corpus callosum morphometry. Cerb Cortex . 2007;17(4):951-961.
22. Castelli F, FrithC, Happe F, FrithU. Autism, aspergersyndromeand brainmecha-nisms for the attribution of mental states to animated shapes. Brain . 2002;125(pt 8):1839-1849.
23. DaprettoM, DaviesMS,Pfeifer JH,et al.Understandingemotions inothers:mir-rorneurondysfunctionin childrenwith autism spectrumdisorders. Nat Neurosci .2006;9(1):28-30.
24. Happe F, RonaldA, PlominR. Time togive upon a singleexplanation forautism.Nat Neurosci . 2006;9(10):1218-1220.
25. Belmonte MK, Bourgeron T. Fragile X syndrome and autism at the intersectionof genetic and neural networks. Nat Neurosci . 2006;9(10):1221-1225.
26. Minshew NJ, Goldstein G, Siegel DJ. Neuropsychologic functioning in autism:
profile of a complex information processing disorder. J Int Neuropsychol Soc .1997;3(4):303-316.27. Williams DL, Goldstein G, Minshew NJ. Neuropsychologic functioning in chil-
drenwith autism:furtherevidence fordisorderedcomplex information-processing.Child Neuropsychol . 2006;12(4-5):279-298.
28. Minshew NJ, Williams DL. Brain behavior connections in autism. In: Buron KD,WolfbergP, eds. Educating Learnerson theAutismSpectrum: TranslatingTheory Into Meaningful Practice. Shawnee, KS: Autism Asperger Publishing Co; 2006.
29. MinshewNJ, SungK, Jones B, Furman J. Underdevelopmentof the posturalcon-trol system in autism. Neurology . 2004;63(11):2056-2061.
30. Luna B, Doll SK, Hegedus SJ, Minshew NJ, Sweeney JA. Maturation of execu-tive function in autism. Biol Psychiatry . 2007;61(4):474-481.
(REPRINTED) ARCH NEUROL/ VOL 64 (NO. 7), JULY 2007 WWW.ARCHNEUROL.COM950
©2007 American Medical Association. All rights reserved. at Capes Consortia, on February 4, 2008www.archneurol.comDownloaded from

8/12/2019 Autism New Neurobiology
http://slidepdf.com/reader/full/autism-new-neurobiology 7/7
Accepted for Publication: April 3, 2007.Correspondence: Walter A. Rocca, MD, MPH, Divisionof Epidemiology, Department of Health Sciences Re-search, Mayo Clinic College of Medicine, 200First St SW,Rochester, MN 55905 ([email protected]).Author Contributions: Study concept and design: Rocca,McDonnell, Schaid, and Maraganore. Acquisition of data:Rocca, Bower, Ahlskog,andMaraganore. Analysis and in-terpretationof data: Rocca, Elbaz, Grossardt, McDonnell,
and Schaid. Drafting of the manuscript: Rocca andGrossardt. Critical revision of the manuscript for impor-tant intellectual content: Rocca, Bower, Ahlskog, Elbaz,Grossardt, McDonnell, Schaid, and Maraganore. Statis-tical analysis: Rocca, Elbaz, Grossardt, McDonnell, andSchaid. Obtained funding: Rocca. Administrative, techni-cal, and material support: Rocca. Study supervision:Rocca and Maraganore.Financial Disclosure: None reported.Funding/Support: This study wassupportedby grant R01NS33978 from the National Institutes of Health and wasmade possible by grant R01 AR30582 (Rochester Epi-demiology Project), also from the National Institutes of Health.
Role of the Sponsors: The sponsors of this study had norole in the design and conduct of the study; collection,management, analysis, and interpretation of the data; orpreparation, review, or approval of the manuscript.
REFERENCES
1. Marder K, Tang MX, Alfaro B, et al. Risk of Alzheimer’s disease in relatives ofParkinson’s disease patients with and without dementia. Neurology . 1999;52(4):719-724.
2. Kurz MW, Larsen JP, Kvaloy JT, Aarsland D. Associations between family his-tory of Parkinson’s disease and dementia and risk of dementia in Parkinson’sdisease: a community-based, longitudinal study. Mov Disord . 2006;21(12):2170-2174.
3. Mickel SF, Broste SK, Hiner BC. Lack of overlap in genetic risks for Alzheimer’sdisease and Parkinson’s disease. Neurology . 1997;48(4):942-949.
4. Levy G, Louis ED, Mejia-Santana H, et al. Lack of familial aggregation of Parkin-son disease and Alzheimer disease. Arch Neurol . 2004;61(7):1033-1039.
5. Pankratz N, Byder L, Halter C, et al. Presence of an APOE4 allele results in sig-nificantly earlier onset of Parkinson’s disease and a higher risk with dementia.Mov Disord . 2006;21(1):45-49.
6. ApaydinH, AhlskogJE, Parisi JE,Boeve BF,DicksonDW. Parkinsondiseaseneu-ropathology: later-developingdementiaand lossof the levodoparesponse. Arch Neurol . 2002;59(1):102-112.
7. McKeith IG,DicksonDW, Lowe J,et al.Diagnosis andmanagementof dementiawith Lewy bodies: third report of the DLB Consortium. Neurology . 2005;65(12):1863-1872.
8. Elbaz A, McDonnell SK, Maraganore DM, et al. Validity of family history data onPD: evidence for a family information bias. Neurology . 2003;61(1):11-17.
9. Rocca WA, McDonnell SK, Strain KJ, et al. Familial aggregation of Parkinson’sdisease: the Mayo Clinic Family Study. Ann Neurol . 2004;56(4):495-502.
10. Rocca WA, Peterson BJ, McDonnell SK, et al. The Mayo Clinic Family Study ofParkinson’s Disease: study design, instruments, and sample characteristics.Neuroepidemiology . 2005;24(3):151-167.
11. McDonnell SK, Schaid DJ, Elbaz A, et al. Complex segregation analysis of Parkin-son’s disease: the Mayo Clinic Family Study. Ann Neurol . 2006;59(5):788-795.
12. Bower JH, Maraganore DM, McDonnell SK, Rocca WA. Incidence and distribu-tionof parkinsonismin OlmstedCounty, Minnesota, 1976-1990.Neurology . 1999;52(6):1214-1220.
13. Bower JH, Maraganore DM, McDonnell SK, Rocca WA. Influence of strict, inter-mediate, and broad diagnostic criteria on the age- and sex-specific incidence ofParkinson’s disease. Mov Disord . 2000;15(5):819-825.
14. Elbaz A, Peterson BJ, Yang P, et al. Nonfatal cancer preceding Parkinson’s dis-ease: a case-control study. Epidemiology . 2002;13(2):157-164.
15. Melton LJ III. History of the Rochester Epidemiology Project. Mayo Clin Proc .1996;71(3):266-274.
16. Brandt J, Spencer M, Folstein M. The Telephone Interview for Cognitive Status.Neuropsychiatry Neuropsychol Behav Neurol . 1988;1(2):111-117.
17. Welsh KA, Breitner JCS, Magruder-Habib KM. Detection of dementia in the el-derlyusing telephone screeningof cognitivestatus. Neuropsychiatry Neuropsy- chol Behav Neurol . 1993;6(2):103-110.
18. Plassman BL, Newman TT, Welsh KA, Helms M, Breitner JCS. Properties of theTelephoneInterviewfor CognitiveStatus:applicationin epidemiological andlon-gitudinal studies. Neuropsychiatry Neuropsychol Behav Neurol . 1994;7(3):235-241.
19. Silverman JM, Breitner JC, Mohs RC, Davis KL. Reliability of the family historymethod in genetic studies of Alzheimer’s disease and related dementias. Am J Psychiatry . 1986;143(10):1279-1282.
20. Gallo JJ, Breitner JC. Alzheimer’s disease in the NAS-NRC Registry of agingtwinveterans, IV:performancecharacteristics of a two-stage telephone screening pro-cedure for Alzheimer’s dementia. Psychol Med . 1995;25(6):1211-1219.
21. Beeri MS, Werner P, Davidson M, Schmidler J, Silverman J. Validation of themodifiedtelephone interviewfor cognitivestatus (TICS-m) in Hebrew.IntJ Geri- atr Psychiatry . 2003;18(5):381-386.
22. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 4th ed. Washington, DC: American Psychiatric Association; 1994.
23. Therneau TM, Grambsch PM.ModelingSurvivalData: Extending the Cox Model.New York, NY: Springer; 2000.
24. Lin DY, Wei LJ. The robust inference for the Cox proportional hazards model.J Am Stat Assoc . 1989;84(408):1074-1078.
25. Folstein MF, Folstein SE, McHugh PR. “Mini-mental state”: a practical methodfor grading the cognitivestate of patients for the clinician.J PsychiatrRes . 1975;12(3):189-198.
26. Parashos SA, Maraganore DM, O’Brien PC, Rocca WA. Medical services utiliza-tion and prognosis in Parkinson disease: a population-based study. Mayo Clin Proc . 2002;77(9):918-925.
27. KnopmanDS, PetersenRC, ChaRH, Edland SD,Rocca WA.Incidence andcausesof nondegenerativenonvasculardementia:a population-basedstudy. ArchNeurol .2006;63(2):218-221.
28. Sackett DL. Bias in analytic research. J Chronic Dis . 1979;32(1-2):51-63.29. Galvin JE, Roe CM, Xiong C, Morris JC. Validity and reliability of the AD8 infor-
mant interview in dementia. Neurology . 2006;67(11):1942-1948.30. Fiske A, Gatz M, Aadnoy B, Pedersen NL. Assessing age of dementia onset: va-
lidity of informant reports. Alzheimer Dis Assoc Disord . 2005;19(3):128-134.
Correction
In the article titled “The New Neurobiology of Autism:Cortex, Connectivity, and Neuronal Organization” byMinshew and Williams, published in the July 2007 is-sue ofthe Archives(2007;64[7]:945-950), Figure 4A wasmislabeled. The labels LSTG and LIFG should beswitched, with LIFG on the left and LSTG on the right.
(REPRINTED) ARCH NEUROL / VOL 64 (NO. 10), OCT 2007 WWW.ARCHNEUROL.COM14 64
©2007 American Medical Association. All rights reserved. at Capes Consortia, on February 4, 2008www.archneurol.comDownloaded from


















