
ATP-Binding Cassette Efflux Transporters and Passive...
70
ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2007 Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Pharmacy 67 ATP-Binding Cassette Efflux Transporters and Passive Membrane Permeability in Drug Absorption and Disposition PÄR MATSSON ISSN 1651-6192 ISBN 978-91-554-7058-6 urn:nbn:se:uu:diva-8371
Transcript of ATP-Binding Cassette Efflux Transporters and Passive...

ACTA
UNIVERSITATIS
UPSALIENSIS
UPPSALA
2007
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Pharmacy 67
ATP-Binding Cassette EffluxTransporters and PassiveMembrane Permeability in DrugAbsorption and Disposition
PÄR MATSSON
ISSN 1651-6192ISBN 978-91-554-7058-6urn:nbn:se:uu:diva-8371

���������� �������� �� ������ �������� � �� �������� ������� � ���� ����������������� ������ ���� ��������� �� ������� ������ !����� �"� �##" �� #$%�& '��(� ������ ' ���� ' )(����(� * ������ ' )(������+, -(� �������� .��� ��������� � /����(,
��������
������ ), �##0, 1-)2����� �������� /''��� -��������� �� )������ �������)����������� � ���� 1������ �� ��������, 1��� ����������� ���������, ���������� � ���� ����� � � ������� ���� ������� �� �� ������� � ������� 30, 3" ��, ������, 45�6 $0"2$�2&&720#&"23,
-������ �� �� ����� �(� ����� ' �(� (��� ��� �� � �����8������ '� �(��(����������� ���� ' �����, )������ ������� ������������ �� ������ ����������(����� ��� ��9� ���������� ' �(� �������� ������� ' ������ �� .��� �� ' �(����������� � ������ ������� �� �(� �����8��� ��������� �� ������� '�� �(� ���, 4�(�� �(����� �(� ��� ' 1-)2����� �������� *1��+ ���������� �� ������� ������������ ���� ������� �� �������� .�� �����������, )��������� ���(���� .�� ������ ��'����(� �������� ��������� ������� '� �(��� ������� ���(�����,-(� �'����� ' ��''���� ������� ���(.��� ��������� ' �������� ���� �������
.�� ����������� ���� ����������� ����� ' ��''���� ���������, /���������� ������(�� ������ �(� ������������ ���(.�� ���� ������� ��������� ' �������� ���� ������������������ '� ���������� ������� �����, ���(��� �(� �(����� ' �(� 1�� ���������������� ����� ��������� ����� *��:);1��<�+ �� ���������2��������� ��������� ������ *�:)�;1����+ .�� ������������� ����������� ���� ������������ ������� �������� �(��.��� ������������� ' ����� ����������� �����, 1 ����� ����� ' ��������� ��.�(������ .��� �����'��� ��� ���������� ������ ��� �(��� ������� �������� '� ����2������������� �� ����2������ ������� ������ � �� ���������, -(� ��9���� ' �(��(������ �''����� ��� �(��� ��9� 1�� ���������� ��:)� �:)� �� )2��������� *)��;1����+� �� �(��� �����2�����'�� �(������ .��� '�� � �� ����(�� � (��(�� ����(����.��� �����,- �������=�� �(� ������ .�� (�� ��� � � �������� �.����� ' �(� ��������
'������� ' �������� '� 1�� ��������� �(����� �� ������� ������� ������������,)�������� ��. 1�� ��������� �(������ .��� �����'��� �� ���������� ��������������� .��� �������� '� �(� ��''���� ���� ������� ���(�����, -(��� ���� �� ����������� � ������ � �(� �������=��� ' ����������� �''��� � ����� ���� ��������,
� ������ 1-)2����� ��������� 1�� ���������� )2��� )2���������� 1����� ��:)������� ����� ��������� ������ 1��<�� �:)�� ���������2��������� ��������� ����� ��1����� ������� ������������� ���� �������� 1����� �������� )������ ��''���������������� ���� �������� )>5� ?)>5� @51:
��� ������ � ���� �� � ������� !" #$% ������� ���� ����� �&'(#)*+ ������� �� � �
A )B� ����� �##0
4556 �3&�23�$�45�6 $0"2$�2&&720#&"23��%�%��%��%����2"�0� *(���%;;��,��,��;������C��D��%�%��%��%����2"�0�+

���


Papers discussed
This thesis is based on the following papers, which will be referred to by the Roman numerals assigned below: I Matsson, P.; Bergström, C. A. S.; Nagahara, N.; Tavelin, S.;
Norinder, U. and Artursson, P. Exploring the Role of Different Drug Transport Routes in Permeability Screening. Reproduced with permission from the Journal of Medicinal Chemistry, 2005, 48, 604-613. © 2005 American Chemical Society.
II Matsson, P.; Englund, G.; Ahlin, G.; Bergström, C. A. S.; Norinder, U. and Artursson, P. A Global Drug Inhibition Pattern for the Human ATP-Binding Cassette Transporter Breast Cancer Resistance Protein (ABCG2). Reproduced with permission from the Journal of Pharmacology and Experimental Therapeutics, 2007, 323, 19-30. © 2007 The American Society for Pharmacol-ogy and Experimental Therapeutics.
III Matsson, P.; Pedersen, J.; Norinder, U.; Bergström, C. A. S.; Hoogstraate, J. and Artursson, P. Prediction and Identification of New Drug Interactions with the Human ATP-Binding Cassette Transporter Multidrug-Resistance Associated Protein 2 (MRP2; ABCC2). Submitted.
IV Matsson, P.; Pedersen, J.; Norinder, U.; Bergström, C. A. S. and Artursson, P. Comparison and Prediction of Inhibitors of the Three Major Human ATP-Binding Cassette Transporters P-gp, BCRP and MRP2. In manuscript.


Contents
1. Introduction .........................................................................................11 1.1. Transport of drugs across cellular barriers .................................12
1.1.1. Passive transcellular transport ...............................................13 1.1.2. Paracellular transport .............................................................14 1.1.3. Active transport .....................................................................16
1.2. ABC transporters in drug disposition .........................................16 1.2.1. The structure of ABC transporters.........................................18 1.2.2. P-glycoprotein (P-gp; ABCB1) .............................................19 1.2.3. Multidrug-resistance associated proteins (MRPs; ABCCs)...19 1.2.4. Breast cancer resistance protein (BCRP; ABCG2)................20
1.3. Computational modeling of drug transport processes ................21 1.3.1. Model development ...............................................................21 1.3.2. Selecting the dataset...............................................................22 1.3.3. Describing the chemical structure..........................................23 1.3.4. Relating experimental parameters to chemical structure .......26 1.3.5. Validating the model..............................................................27 1.3.6. Models of drug transport processes .......................................28
2. Aims of the thesis ................................................................................31
3. Methods ...............................................................................................32 3.1. Dataset selection.........................................................................32 3.2. Structural diversity .....................................................................32 3.3. Experimental methods................................................................33
3.3.1. Preparation of filter-immobilized hexadecane membranes (HDMs) ...............................................................................................33 3.3.2. Permeability measurements ...................................................33 3.3.3. Measurements of BCRP efflux inhibition..............................34 3.3.4. Measurements of MRP2 efflux inhibition .............................34
3.4. Computational methods..............................................................35 3.4.1. Generation of molecular descriptors......................................35 3.4.2. Pharmacophore modeling ......................................................35 3.4.3. Statistical analysis..................................................................36
4. Results and discussion .........................................................................38 4.1. Diversity of datasets for drug transport modeling ......................38

4.2. Influence of different transport pathways on intestinal permeability..............................................................................................39 4.3. Identification of inhibitors of BCRP and MRP2 ........................40 4.4. Overlapping inhibitor specificity of P-gp, BCRP and MRP2 ....43 4.5. Computational models of ABC transporter inhibition ...............45
5. Conclusions and future outlook ...........................................................48
6. Svensk populärvetenskaplig sammanfattning......................................50
7. Acknowledgements..............................................................................52
8. References and notes ...........................................................................54

Abbreviations
1D One-dimensional 2/4/A1 Immortalized rat intestinal epithelium
cell line 2D Two-dimensional 3D Three-dimensional ABC ATP-binding cassette ADMET Absorption, distribution, metabolism,
elimination/excretion, toxicity AMP Adenosine monophosphate ANN Artificial neural network ANOVA Analysis of variance ATP Adenosine triphosphate BBB Blood-brain barrier BCRP Breast cancer resistance protein
(ABCG2) Caco-2 Human colon adenocarcinoma cell
line ChemGPS Chemical global positioning system CHO Chinese hamster ovary cell line CNS Central nervous system CoMFA Comparative molecular field analysis CYP Cytochrome P450 FA Fraction of an orally administered
dose that is absorbed HBSS Hank’s balanced salt solution HDM Filter-immobilized hexadecane
membrane HEK293 Human embryonic kidney cell line hERG Human ether-a-go-go-related ion-
channel LLC-PK1 Pig kidney epithelial cell line logD Octanol-buffer partition coefficient logP Octanol-water partition coefficient MDCK Madine-Darby canine kidney cell line MLR Multiple linear regression MMFF Merck molecular force field

MNLR Multiple non-linear regression mRNA Messenger ribonucleic acid MRP Multidrug-resistance associated pro-
tein (ABCC) MW Molecular weight n Number of observations NBD Nucleotide binding domain OPLS Orthogonal PLS OPLS-DA Orthogonal PLS discriminant analy-
sis PCA Principal component analysis P-gp P-glycoprotein (ABCB1) PLS Partial least squares projection to
latent structures PLS-DA PLS discriminant analysis Q2 Cross-validated R2 QSAR Quantitative structure-activity rela-
tionship R2 Coefficient of determination RMSE Root mean square error rs Spearman rank order correlation co-
efficient SLC Solute carrier SVM Support vector machine TEER Trans-epithelial electrical resistance TM Transmembrane TMD Transmembrane domain

11
1. Introduction
Transport into and across the cells of the human body is a prerequisite for the pharmacological action of drugs. An orally administered drug needs to cross the intestinal epithelial cell layer if it is to reach the blood circulation (Figure 1). Many target tissues, such as the central nervous system (CNS), are protected by additional cellular barriers, and for intracellular pharmacol-ogical targets an even greater number of cell membranes need to be crossed before the pharmacological action can be elicited. An equally important role of membrane transport is evident in the elimination of drugs from the body. Transport across cellular membranes determines the access to intracellular drug-metabolizing enzymes in the liver and other metabolizing organs, and cellular transport processes have a pronounced effect on the renal and biliary excretion of the drug.
In 1991, inadequate bioavailability and poor pharmacokinetic profiles were the major causes for the termination of candidate drugs.1 This prompted the inclusion of experimental and computational assays for predicting the absorption and metabolism of drugs in early discovery settings, in parallel with the pharmacological screening.2 A decade later, the proportion of clini-cal drug failures that could be related to pharmacokinetic and bioavailability issues had been significantly reduced.3 Experimental and computational models of intestinal solubility and passive membrane permeability, as well as of hepatic metabolism, probably contributed to this development. How-ever, the reduction of the clinical drug failures was in part counterweighed by an increase in the number of failures attributable to toxicological reasons, underlining the need for further characterization of discovery drugs.
During the past decade, the complexity of membrane transport has be-come increasingly apparent. The characterization of the human genome re-vealed a plethora of proteins facilitating the transmembrane transport of vari-ous solutes. Many of these transport proteins have the capacity to transport drug molecules, and significantly contribute to the pharmacological and toxi-cological activity of drugs.
In this thesis, the impact of passive permeability and active transport on the intestinal absorption, tissue distribution and elimination of drugs was examined with the aim of developing predictive computational models for important drug transport mechanisms. A particular emphasis was placed on drug efflux transporters from the ATP-binding cassette (ABC) transporter family.

12
Disintegration
Permeability
Dissolution
Drug in solution
Degradation Complexation
Drug in systemic circulationFraction absorbed
Gut wall metabolism
Liver metabolism
Absorption
Bioavailability
Biliary excretion
Figure 1. The sequence of events following the administration of a solid dosage form to the gastrointestinal tract. The dissolution and the permeability across the epithelium are the most important determinants of intestinal drug absorption. Other factors that can limit the fraction of an orally administered dose reaching the sys-temic blood circulation include: enzymatic and chemical degradation and complexa-tion in the gut lumen; enzymatic degradation in the intestinal epithelium; and enzy-matic degradation and biliary excretion in the liver.
1.1. Transport of drugs across cellular barriers Several distinct pathways affect the transport of drugs across tissue barriers such as the intestinal epithelium and the blood-brain barrier (BBB) (Figure 2). Although the relative importance of the pathways varies between one tissue and another and for different drugs, the general transport mechanisms are applicable to tissues throughout the human body.
The pathways can be divided in two types: the transcellular pathway, the use of which requires that a drug can cross the lipophilic cell membranes, and the paracellular pathway, in which diffusion occurs through the water-filled pores of the tight junctions between the cells. In addition, both passive and active (energy-dependent) transport can contribute to the permeability of drugs via the transcellular route. The four principal pathways of drug trans-port are presented in Figure 2. All of the transport processes may occur in both directions, depending on local drug concentrations (in the case of pas-sive transport) and the directionality of the relevant transporter (in the case of active transport). The various transport mechanisms are discussed below, using the intestinal epithelium as an example. Examples of in vitro methods used in studies of membrane permeability and active drug transport are given in Box 1.

13
1 2 3 4
A B
1 2 3 4 Figure 2. The drug-relevant transport pathways in the intestinal epithelium: 1, pas-sive transcellular transport; 2, paracellular transport; 3, active efflux; 4, active up-take. Passive permeability can occur in absorptive (A) and secretory (B) directions, depending on local drug concentrations. Correspondingly, drug transport proteins are expressed in both the apical and the basolateral membranes, and mediate active transport in absorptive and secretory directions.
1.1.1. Passive transcellular transport The passive transcellular pathway is the most important absorption pathway for the vast majority of orally administered drugs, and also significantly af-fects the distribution of drugs to other tissues. The first requirement for drug absorption by this route is that the solute permeates the apical cell membrane of the intestinal enterocyte. Enterocytes have a polarized cell membrane with distinct differences in the membrane composition between the apical and basolateral membranes.4, 5 It is generally believed that the apical membrane has a lower permeability than the basolateral membrane, and therefore the former is considered to be the rate-limiting barrier to passive transcellular drug absorption.6 In addition, the composition of the phospholipids and pro-teins of the cell membranes varies from cell type to cell type and may, theo-retically, give rise to different permeabilities in different cell types.
Early models of the transcellular pathway regarded the cell membrane as a homogenous barrier and drug permeability was, therefore, considered to be a two-stage process involving partitioning into the lipid bilayer followed by transmembrane diffusion.7 This ‘solubility-diffusion’ model was later ex-tended with the so-called pH-partitioning theory, which states that only the uncharged form of an electrolytic molecule will partition into the cell mem-brane.8 In these early models, the cell membrane was considered to be an isotropic system. This assumption explains why drug partitioning in simple isotropic solvent systems, such as octanol-water, has been extensively used to predict passive membrane permeability.9-11 However, the lipid bilayer that constitutes the cell membrane is a highly complex system.12 Thus multiple-

14
step permeability models have been proposed to account for the anisotropic nature of the cell membrane.13 Molecular dynamics simulations have shown that drug molecules entering the cell membrane experience different diffu-sion rates in different parts of the membrane, since different inter-molecular forces affect the permeating substances depending on the local environment in the lipid bilayer.12, 14-19
Regardless of whether the ‘solubility-diffusion’ model or a more complex description of the cell membrane is used, passive transcellular drug perme-ability can usually be relatively well described using rather simple molecular properties such as size, charge, polarity and hydrogen bonding.14, 20, 21 Many experimental and computational models of intestinal drug permeability have therefore been based on descriptors of these basic molecular properties.22-24 An important general assumption made in these models is that the passive transcellular route dominates the drug permeation. However, when other transport routes, such as the paracellular or active transcellular routes are involved to a significant extent, they will introduce a bias that reduces the predictivity of the model. The influence of the passive transcellular, paracel-lular, and active drug transport routes on intestinal permeability predictions were investigated in Paper I in this thesis.
1.1.2. Paracellular transport Drugs of small to moderate molecular weight can permeate the intestinal epithelium through pores between the cells, a process known as paracellular transport. The narrow, water-filled pores are created by extracellular tight junction proteins that connect adjacent cells. In the intestine, the paracellular pores only correspond to around 0.01–0.1% of the absorptive surface area, and the average pore sizes have been estimated to be 8–13 Å in the human small intestine,25 decreasing to only about 4 Å in the colon.26 This route is, therefore, mainly accessible to smaller molecules (MW<350 g�mol-1)27-30 and is of significance only if these small drugs are too hydrophilic to distrib-ute into the cell membranes at appreciable rates. Similar to the intestinal epithelium, endothelial cells in the brain microvasculature are also connected by tight junctions.31, 32 This results in a much tighter endothelial layer than in capillaries in other parts of the body, and contributes to the protective role of the blood-brain barrier.
Paracellularly permeating drugs are usually incompletely absorbed since absorption only occurs in the small intestine where the pores are sufficiently large to allow the passage of small solutes. The paracellular pathway of the small intestine has been described as a dual-pore system, with small pores at the villus tips and larger ones further down the crypt-villus axis (Figure 3).33,
34 Given this, it can be hypothesized that a hydrophilic drug that is partition-ing slowly into the intestinal cell membrane will have time to diffuse further down the crypt-villus axis and eventually be absorbed via paracellular pores

15
that are significantly larger than those available at the tips of the villi. Thus, such compounds may exhibit higher in vivo absorption than would be ex-pected from their passive transcellular permeability.
Figure 3. Permeability properties of different parts of the crypt-villus axis. A. Sche-matic representation of the crypt-villus axis, divided into upper villus, basal villus and crypt. B. The effective intestinal surface area that the drug encounters will be determined by the epithelial permeability to the drug. Compounds with poor perme-ability (open circles) will have time to diffuse further down the crypt-villus axis, and can therefore be absorbed across a large surface area (dashed line). In contrast, high-permeability compounds (closed circles) are rapidly absorbed at the villus tips and therefore utilize a smaller absorptive surface area (solid line). The average size of the paracellular pores varies along the crypt-villus axis, with the pores getting nar-rower as the cells mature and move towards the villi tips.33

16
1.1.3. Active transport In addition to the passive diffusion processes described above, transport proteins situated in the membranes of the cell significantly affect the move-ment of substrate drugs across cellular barriers. Of the more than 20 000 genes in the human genome, around 500 encode transport proteins and ion channels.35 Although the majority of these have specific physiological roles in, e.g., nutrient uptake or neuronal recirculation of transmitter substances, an increasing number have been found to have more general transport func-tions. These transporters, which have broader substrate specificity, are often referred to as polyspecific transport proteins, and have the capacity to trans-port drugs.
The ATP-binding cassette (ABC) transporters and the solute carriers (SLC) are the two dominating gene families among the plasma membrane transporters. ABC transporters utilize the energy from ATP hydrolysis to translocate their substrates across cellular membranes.36 Some members of the SLC family mediate facilitated diffusion of their substrates along a con-centration gradient, whereas others are secondary active transporters that use ion gradients to mediate concentrative substrate transport.37, 38 ABC and SLC transporters with the capacity to transport drugs are expressed in tissues throughout the body, and have been shown to significantly affect pharma-cokinetic processes such as intestinal absorption,39, 40 distribution to the CNS,41, 42 and uptake into, and subsequent excretion from the hepatocyte.43 In this thesis, particular emphasis was placed on three major drug-transporting members of the ABC transporter family: P-glycoprotein (P-gp; ABCB1), breast cancer resistance protein (BCRP; ABCG2), and multidrug-resistance associated protein 2 (MRP2; ABCC2).
1.2. ABC transporters in drug disposition ABC transporters constitute one of the largest superfamilies of membrane proteins, with members having been found in all species studied so far.44 In humans, 48 members have been identified, and these have been phylogeneti-cally divided into seven subfamilies (labeled ABCA through ABCG).45 Al-though the exact physiological roles are still unknown for many of the hu-man ABC transporters, several are implicated in genetic disorders such as Tangier disease,46 Stargardt disease,47 Dubin-Johnson syndrome48 and cystic fibrosis.49 The human ABC transporters with affinity for drugs are mainly found in the ABCB, ABCC and ABCG subfamilies. The following sections will briefly review the structure, mechanism, and the roles of the major drug efflux transporters P-gp, BCRP, and the MRPs in drug disposition.

17
Box 1. In vitro methods for studying permeability and active transport
Experimental models of intestinal permeability The most widely used model for determining the intestinal permeability to drugs is the Caco-2 cell line, which is derived from a colon carcinoma and has been shown to differentiate to a phenotype similar to the mature small intestinal entero-cyte.50, 51 Alternative cell lines with slightly different properties include the Caco-2 cell clone TC7 which, in contrast to the parental Caco-2 cells, expresses some drug-metabolizing enzymes,52, 53 and the mucus-producing HT29-MTX clone.54 The 2/4/A1 cell line is an immortalized cell line derived from rat intesti-nal epithelium.55, 56 It has a more leaky, small intestine-like paracellular pathway than that of Caco-2 monolayers, and does not express important drug transporters from the SLC and ABC transporter families.
As an alternative to cultured cells, artificial membranes can be used to study passive drug permeability. These systems are either based on simple organic sol-vents such as hexadecane,57 that mimic the hydrophobic interior of the plasma membrane, or on different mixtures of membrane phospholipids.58, 59
Details of the methodology of permeability assessment using Caco-2 and 2/4/A1 cells and artificial hexadecane membranes are given in Reference 51 and in Section 3.3.1.
Experimental models for studying active drug transport Caco-2 cells express many drug-relevant transport proteins, and can thus be used to determine the overall influence of active transport on the permeability. For mechanistic studies of specific transporters, cell lines transfected with one or more transporters are often preferred. Madine-Darby Canine Kidney (MDCK) cells60 and the pig kidney cell line LLC-PK161 are often used to this end, and since they form monolayers they can be used to study active transport processes in Caco-2-like permeability assays.
Cell lines such as HEK293, CHO, K562 and Saos-2 that do not form confluent monolayers are generally used to assess the intracellular accumulation of fluores-cent or radiolabeled transporter substrates, thereby giving information about ac-tive transport mechanisms.62-65 Efflux transporters are often studied using inside-out membrane vesicles.66-68 The inverted nature of these systems enables direct interaction of investigated drugs with the transporter, without the need for ade-quate membrane permeability to reach intracellular binding sites. As an alterna-tive to following the transport of a fluorescent or radiolabeled probe, ATP con-sumption is often used as a surrogate measure of drug transport in cell-based sys-tems and in membrane vesicles.67, 69

18
1.2.1. The structure of ABC transporters ABC transporters share a common domain organization consisting of two transmembrane domains (TMDs) that form the passageway for the transport of the substrate, and two nucleotide-binding domains (NBD) that provide the energy for the transport by catalyzing ATP hydrolysis.44 In prokaryotes, both ABC importers and ABC exporters have been identified, whereas in eu-karyotic cells the transport is exclusively in the exporting (efflux) direction. Because of the difficulty of crystallizing membrane proteins, still only four high-resolution crystal structures of complete ABC transporter proteins are available, three of which are ABC importers.70 The only available structure of an ABC efflux transporter is that of Sav1866 from Staphylococcus Aureus (Figure 4A).71 Recent biochemical evidence suggest that the domain ar-rangement is similar in Sav1866 and human P-gp,72 and consequently, the structure has received much attention as a template for homology models of human ABC transporters.73-77
ABC efflux transporters contain a conserved core of 12 transmembrane (TM) helices, which are believed to form the transport pathway.44 For P-gp, the secondary structure can be divided into two similar halves, each contain-ing 6 TM helices and one NBD (Figure 4B).78 This basic ABC efflux trans-porter structure is shared by MRP4 and MRP5, whereas MRP1-3 and MRP6 contain an additional aminoterminal domain consisting of 5 TM helices.
H2NNBD NBD
COOH
H2N
NBD NBD
COOH
H2NCOOH
NBD
P-gp, MRP4-5
MRP1-3, MRP6
BCRP
out
in
A B
TMs
NBDs
Figure 4. Structure and membrane topology of ABC efflux transporters. A. Crystal structure of the bacterial ABC exporter Sav1866 (pdb id 2HYD). The transmem-brane (TM) helices and the intracellularly located nucleotide binding domains (NBD) are indicated in the picture; the gray box shows the position of the plasma membrane. B. Predicted membrane topology for the human ABC transporters P-gp, MRP1-6 and BCRP. P-gp, MRP4 and MRP5 consist of the conserved 12 TMs and two NBDs, whereas MRP1-3 and MRP6 have an additional aminoterminal domain consisting of 5 TMs.78 BCRP is a half-transporter, and as such only consists of 6 TMs and one NBD. The position of the membrane is indicated by the gray boxes, and all transporters are depicted with the same orientation, down being towards the inside of the cell.

19
In contrast, BCRP is a so-called half transporter that consists of only 6 TM helices and one NBD. However, biochemical and low-resolution crystallo-graphic data have demonstrated that the functional BCRP unit is formed by two protein molecules in a homodimeric structure,79, 80 and thereby BCRP fulfills the conserved domain architecture.
1.2.2. P-glycoprotein (P-gp; ABCB1) P-gp was the first eukaryotic ABC transporter discovered, and was identified as the cause of a multidrug-resistant phenotype observed in drug-selected cells from Chinese hamster ovaries (CHO).81 Since then, a vast body of lit-erature has been published on the structure, mechanism and physiological and pharmacological roles of P-gp. More than 12 000 papers concerning P-gp had been published by November 2007,82 making this the by far most studied drug transporter.
In spite of this, the physiological role of P-gp is still not completely un-derstood. Suggested endogenous substrates include membrane components such as phosphatidylcholine and cholesterol, as well as steroid hormones and cytokines.83 The expression pattern suggests that P-gp plays a role in tissue protection and detoxification, with high expression in the apical membrane of intestinal enterocytes, in the bile canalicular membrane of the liver and in protective tissue barriers such as the BBB, the blood-testis barrier and in the placental syncytiotrophoblasts that connect maternal and fetal blood circula-tion.84-86
P-gp distinguishes itself from most other transporters by the structural di-versity of its substrates. Most P-gp substrates are lipophilic and uncharged or weakly basic, although acidic and hydrophilic substrates such as meth-otrexate have also been reported.78 The common denominator of P-gp sub-strates is their amphipathic character. It is generally accepted that P-gp binds its substrates from the inner leaflet of the plasma membrane, thereby ex-plaining this preference for surface-active compounds.87-89 This model for drug binding to P-gp was recently corroborated by molecular dynamics simulations of surface active P-gp substrates. The compounds first partition into the lipid membrane, and subsequently exchange their interactions with water or the polar lipid head groups in the membrane-cytosol interface for interactions with polar and charged amino acids in the lipid-water interface region of P-gp.90
1.2.3. Multidrug-resistance associated proteins (MRPs; ABCCs) The ABCC subfamily consists of 12 distinct transporters, of which MRP1-6 are generally considered as drug transporters.78 MRP1 was first identified in a drug-resistant cell line,91 and it was subsequently shown to confer resis-tance to a similar, although not identical panel of cytotoxic drugs as that

20
observed for P-gp.92 In contrast to P-gp, MRP1 is localized to the basolateral membranes of intestinal enterocytes and hepatocytes.93 Typical MRP1 sub-strates are organic anions,91, 92 although substrates lacking negative charges have also been reported.94, 95 Such transport is however dependent on intra-cellular glutathione, and it is generally accepted that non-anionic MRP1 substrates are co-transported with reduced glutathione.
The MRP2 gene was identified on the basis of its similarity to MRP1,96, 97 but it differs from its sibling by co-localizing with P-gp to the apical mem-brane of intestinal epithelium and the hepatocyte.92, 98 The physiological role of hepatic MRP2 is to mediate biliary excretion of conjugated bilirubin and bile salts; genetically impaired MRP2 function results in a conjugated hy-perbilirubinemia known as Dubin-Johnson syndrome.99 The substrate spec-trum of MRP2 is distinct but similar to that of MRP1, with a preference for organic anions and reported co-transport of weakly basic substrates with reduced glutathione.100, 101
Several studies have demonstrated that transport mediated by MRP1 and MRP2 can be stimulated by the addition of other compounds.68, 94, 95, 102 For MRP1, this is exemplified by the marked increase in etoposide and vincris-tine transport observed on the addition of glutathione.94, 95 Conversely, eto-poside and vincristine stimulate the transport of glutathione, indicating that these weakly basic drugs are co-transported with reduced glutathione. For MRP2, transport stimulation is readily apparent from the sigmoidal concen-tration-dependency observed for estradiol-17�-glucuronide. This is indica-tive of positive cooperative transport, i.e., estradiol-17�-glucuronide stimu-lates its own transport at higher concentrations.68, 102 However, the fact that not all MRP2 stimulators are transported themselves led to the proposal that there are two distinct binding sites in MRP2: one transporting site and one stimulating site.68, 103 With this explanatory model, transported stimulators have affinity for both sites, whereas other compounds, with an affinity for only one of the sites, act either as substrates or stimulators.
Immunolocalization experiments have demonstrated the expression of MRP3 and MRP6 in the basolateral membrane of hepatocytes,104, 105 and MRP4 and MRP5 have been detected in the luminal membrane of BBB en-dothelial cells, co-localized with MRP1.106 Although these members of the ABCC family have not been extensively studied, reports indicate a similar substrate specificity of MRP3 and MRP6 to that of MRP1,102, 107, 108 and MRP4 and MRP5 have been shown to transport antiviral nucleotide ana-logs.109, 110
1.2.4. Breast cancer resistance protein (BCRP; ABCG2) BCRP was first cloned in the late 1990s from a doxorubicin-resistant breast cancer cell line.111 Since then, the transporter has been detected in a variety of healthy as well as malignant tissues. Physiological expression of BCRP is

21
found e.g. in intestinal epithelium, in bile canalicular membranes of hepato-cytes, in placental syncytiotrophoblasts and in the ducts and lobules of the breast.112-114 Unlike most other half transporters, which are expressed in in-tracellular membranes, BCRP is localized to the plasma membrane.115 Simi-lar to P-gp the physiological role of BCRP is still unclear, but the localiza-tion to apical membranes of tissue barriers suggests a role in tissue protec-tion.
Since its discovery, BCRP has received considerable attention for its role in resistance to various cytotoxic agents.111, 116 More recently, it has been shown to transport a diverse set of drugs from other therapeutic classes.113,
117, 118 The substrate specificity of BCRP overlaps with both the MRPs and with P-gp, and includes intrinsically anionic drugs such as methotrexate119 and pitavastatin,43 anionic conjugates such as estrone-3-sulfate120 and cati-onic compounds such as mitoxantrone.121
It is becoming increasingly clear that BCRP is one of the major ABC transporters affecting drug disposition. This role was recently exemplified by a 111 times higher systemic exposure to the antiinflammatory drug sulfasa-lazine after oral administration to Bcrp1-knockout mice compared to wild-type mice.39 In addition, the human oral bioavailability of the BCRP sub-strate topotecan was more than doubled after coadministration with the po-tent inhibitor GF120918,122 highlighting the risk of significantly altered drug exposure as a result of BCRP inhibition.
1.3. Computational modeling of drug transport processes
The impact of transport processes on the pharmacological and toxicological action of drugs has become increasingly apparent during the past two dec-ades. As a consequence, the application of methods for predicting the trans-port mechanisms of new drugs has become an integral part of the workflow in early drug discovery. Using mathematical and statistical tools it is possi-ble to relate the transport characteristics to the chemical structure in a com-putational model, and to use these models to predict the properties of un-known, possibly not even synthesized compounds. Such models can then aid in selecting the compounds most suitable for further development.
1.3.1. Model development The procedure of developing predictive computational models, e.g. describ-ing membrane permeability or the binding to a transport protein, can be di-vided into five general steps: i) selecting a representative compound dataset; ii) generating or collecting experimental data for the property of interest; iii)

22
describing the chemical structure of the compounds in the dataset; iv) relat-ing the structural description to the experimental data using a mathematical relationship; and v) validating the predictivity of the model using a test set of compounds that were withheld from the model development (Figure 5). Im-portant aspects of these five steps are discussed in the following sections, and examples of computational models of drug transport are discussed.
Experimentaldata
Moleculardescriptors
Dataset Statistics Model
Validation
Use
Figure 5. The steps in the development of a computational model. i) Selecting the dataset, ii) determining experimental data, iii) describing the molecular structures, iv) connecting the experimental data to the chemical structures by statistical or mathematical tools and v) validating the model. Adapted from Reference 123.
1.3.2. Selecting the dataset The dataset used to generate the model will determine its applicability do-main, i.e., for which compounds the model can be expected to give reliable predictions. Models can be divided into global models, that are applicable to widely differing chemical structures, and local models, that are confined to a much more limited chemical space, e.g., specific series of homologous com-pounds. In general, there is a tradeoff between the applicability domain and the quality of the predictions. Consequently, global models are often used to provide initial, qualitative predictions (with yes/no answers or outcomes categorized in groups such as high, intermediate or low), whereas local mod-els are used to provide more detailed, quantitative predictions. Hence, global models can be used to guide experimental efforts to compound series with favorable characteristics, for which local models providing more exact, quantitative predictions can be developed and applied.
Regardless of whether global or local models are developed, the dataset compounds should be structurally diverse within the chemical space of the model. A number of techniques available for determining structural diversity are reviewed in Reference 124. Furthermore, to ensure accurate predictions, it is important that the dataset is not skewed with regard to the modeled ex-perimental parameter.

23
1.3.3. Describing the chemical structure The chemical structures of the modeled compounds need to be translated into numerical representations before they can be related to the experimental parameter. The approaches adopted for obtaining numerical descriptions of the chemical structures can be divided into four main categories, although this distinction is somewhat arbitrary: i) molecular descriptors that encode structural information about the molecule as a whole or parts of the mole-cule; ii) molecular fingerprints that describe the presence or absence of a list of structural features in the molecule; iii) molecular interaction fields that encode the interaction of the molecule with probes in the surrounding space; and iv) pharmacophore models that describe how well the molecule can be superimposed on a three-dimensional arrangement of important functional groups. A brief overview of these four approaches is given below.
1.3.3.1. Molecular descriptors Molecular descriptors encode properties related to the structure of the mole-cule, such as its size or polarity, or the connectivity between atoms in the molecule. Descriptors can be classified as one-dimensional (1D), two-dimensional (2D) or three-dimensional (3D), depending on the complexity of the structural representation used to calculate them (Figure 6). According to this scheme, 1D descriptors refer to bulk properties of the molecule, such as atom counts or the molecular weight. 2D descriptors are typically related to size, flexibility/rigidity or hydrophilicity/lipophilicity and can be calcu-lated from a two-dimensional representation of the molecule. 3D descriptors are dependent on the conformation of the molecule and encode information about the arrangement of atoms in three-dimensional space, e.g., solvent accessible molecular surface areas and molecular volumes. Structural repre-sentations in the form of wave functions constitute a fourth level of com-plexity, and are used in quantum mechanics calculations of properties related to electron densities.
Ideally, a molecular descriptor should be interpretable, so that conclusions about physicochemical interactions with the surroundings can be drawn, and so that models based on the descriptors can be used to guide medicinal chemistry efforts. In addition, the descriptors should be rapidly calculated, so that they can easily be applied to large chemical libraries. Rule-based meth-ods have increased the speed of generating 3D conformations, facilitating the use of 3D descriptors in quantitative structure-activity relationships (QSARs). However, more sophisticated methods for generating molecular conformations, such as molecular mechanics calculations, are preferred for applications where the spatial orientation of functional groups is vital.

24
2D structure
3D structure
Representation Descriptors
Fragment counts, topological indices
Molecular surface properties, molecular volume,drug-probe interactions (GRID/VolSurf), CoMFA, etc
Atom counts, molecular weight1D structure
C22H28FN3O6S
OH
N
N
CH3
CH3
OH
N
S
CH3
O
OH
CH3O
O
F
Figure 6. The different levels of complexity of molecular structure representations. Examples of molecular descriptors that can be calculated from the different repre-sentations are shown to the right.
1.3.3.2. Pharmacophore models A pharmacophore describes how the structural features that determine the biological effect of a set of molecules are arranged in 3D space (Figure 7A). A number of algorithms for developing pharmacophore models are avail-able, with the common goal being to identify the optimal 3D arrangement of structural features that exist in all ligands.125-131 This is generally carried out in two steps: first the ligands are analyzed to find pharmacophore features that are common to all compounds. These structural features usually include hydrogen bond donors and acceptors, hydrophobic, aromatic, and positively and negatively ionizable features. In a second step, the ligand structures are superimposed onto each other, and different 3D arrangements of the features are scored according to how well they fit the ligands.
The algorithms available can be divided into two types: those that solely determine the overlapping features in a set of active ligands, such as the Catalyst HipHop algorithm,129 and those that also take the affinities of the ligands for the target structure into account, including among others the Phase and Catalyst HypoGen algorithms.128, 131 Since no assumptions about the relative potency of the active compounds are made in the former ap-proach, these algorithms only need the structures of the ligands as an input. In addition to the structures, the latter approach requires the experimentally determined affinities for compounds ranging from completely inactive to highly active. This type of modeling has the advantage of not only classify-ing compounds as active or inactive, but also provide predictions of the ligand potency.

25
Molecular interaction fieldB
OH
N
N
CH3
CH3
OH
N
S
CH3
O
OH
CH3O
O
F
0 1 0 0 0 0 1 0 0 0 0 1 0 0 0 0
Molecular fingerprintCPharmacophore modelA
Figure 7. Approaches for describing the chemical structure in computational models of drug transport. A. Rosuvastatin mapped to a 5-point pharmacophore. Red spheres denote hydrogen bond acceptors, the blue sphere denotes a hydrogen bond donor, the green sphere denotes a hydrophobic feature and the yellow circle denotes an aromatic feature in the pharmacophore. B. Molecular interaction fields are deter-mined by calculating the interaction between the drug and probes placed at regularly spaced grid points (denoted by green spheres) in the surrounding space. C. Molecu-lar fingerprints encode the occurrence of structural characteristics in the molecule. E.g., in substructure fingerprints, each position in a string of binary (1/0) values encodes whether or not a certain structure fragment exists in the molecule.
1.3.3.3. Molecular interaction fields Molecular interaction fields are commonly used in 3D-QSAR models to describe the interactions between a molecule and its target structure in three-dimensional space. Various methods are available for determining the inter-action fields, including the widely used CoMFA132 and GRID133 algorithms. With these methods, energies are calculated for the interaction between the molecule and probes placed at regularly spaced intervals in the surrounding space (Figure 7B). By selecting probes with different physicochemical prop-erties, interaction fields related to steric effects, hydrogen bonding and elec-trostatic interactions can be obtained. Models relating interaction fields to activity can thereby define regions in a molecule in which structural modifi-cations would result in an altered biological effect.
1.3.3.4. Molecular fingerprints Molecular fingerprints can be thought of as a collection of substructural mo-lecular descriptors that together provide a description of the structure of the molecule. The fingerprints are typically encoded using a string of binary values, where each bit position marks the presence or absence of a certain structural characteristic in the molecule (Figure 7C). This approach is often used to encode which of a large number of possible substructure fragments are present in a molecule. Similarly, pharmacophore fingerprints are used to describe which, out of all possible pharmacophore patterns, that exist in the molecule, thereby providing a picture of the potential interactions that a molecule can take part in.

26
1.3.4. Relating experimental parameters to chemical structure Once the experimental data has been assembled and the chemical structures of the compounds in the dataset have been described numerically, the next step of the modeling procedure is to relate the experimental property to the structures using statistical methods. In general, single molecular descriptors are not enough to describe complex cellular processes such as drug transport, and multivariate statistics are therefore usually preferred. The available methods include both linear and nonlinear statistics, as well as different clas-sification modeling techniques. The techniques differ in the types of prob-lems they handle, and in the transparency of the resulting models.
1.3.4.1. Modeling continuous response variables The most common methods used for modeling drug transport processes are the linear multivariate methods, including e.g. multiple linear regression (MLR) 134-136 and partial least squares projection to latent structures (PLS).136, 137 Linear methods have the advantage of being relatively transpar-ent and easy to comprehend. The regression coefficients obtained from linear methods provide easily interpreted information about the relative influence of the descriptors on the experimental parameter. A drawback of MLR and PLS is that they lack the ability to handle complex nonlinear problems in their standard forms, although nonlinear extensions are available to this end.138, 139
Although MLR and PLS are both linear methods, they differ considerably in their handling of the multivariate problem. PLS can be viewed as an aug-mented linear regression method that does not display the general limitations of MLR, including the inability to handle inter-correlated descriptors or data matrices containing more descriptors than compounds.137 These advantages of the PLS methodology made it the statistical method of choice throughout this thesis. In PLS, the molecular descriptors are projected to a limited num-ber of latent ‘supervariables’ or principal components. These latent variables contain the majority of the information originally described by a much larger number of molecular descriptors. Orthogonal PLS (OPLS) is an extension of PLS where the part of the information in the molecular descriptors that is related to the response parameter is collected in predictive principal compo-nents.140 The remaining information is described by components orthogonal to the predictive component.
Among the nonlinear methods used for modeling transport processes, ar-tificial neural networks (ANN) are the most widely employed.136, 141-143 ANNs mimic the connections of neurons in the brain, and relate the molecu-lar descriptors to the experimental parameter through a number of layers of neurons or nodes. This complex pattern of connections means that ANNs are able to describe complex nonlinear variations in the relation between the training set compounds and the experimental parameter. However, one

27
drawback is that the transparency is greatly reduced compared to the linear models, and it is generally not possible to deduce the influence of a specific descriptor on the experimental parameter. In addition, a particular problem associated with most ANN models is their tendency to become overtrained, i.e., the compounds used for model development are perfectly described, but the models lack the ability to predict new data.
1.3.4.2. Discriminant models In some cases the aim of modeling is not to predict an exact binding affinity or transport rate, but rather, to classify compounds into groups according to different activities. For this purpose, various classification or discriminant modeling techniques can be used. These include decision trees, support vec-tor machines (SVM) and PLS-discriminant analysis (PLS-DA).
Decision trees, or recursive partitioning techniques, are used to find branching points in the molecular descriptors that result in an optimal divi-sion of the compounds into different activity classes. The end result is a set of rules, such as ‘if the molecular weight is greater than 450 and the octanol-water partition coefficient is above 3.2, then the compound is active’.
SVMs divide the compounds into classes by identifying hyperplanes in the multidimensional descriptor space that result in the largest margin of separation between the activity groups. Since nonlinear functions can be used to define the separating hyperplanes SVMs can deal with complex nonlinear classification problems, although not without a considerable reduc-tion in the transparency of the models.
Throughout this thesis, classification problems were handled using PLS-DA or OPLS-DA. These are variants of the standard PLS methods and, as such, displays the advantages of PLS, e.g., the ability to handle intercorre-lated descriptors and large descriptor matrices. In PLS-DA and OPLS-DA, the continuous response variable in PLS is replaced with a matrix of dummy variables that describe the class memberships of the compounds using binary values (1/0 or 1/-1). Similar to continuous-variable PLS, the molecular de-scriptors are projected to latent variables, and these are used to divide the compounds into activity classes.
1.3.5. Validating the model Before applying a model to the routine prediction of unknown compounds, the validity of the model must be assessed. The validation gives information about the predictivity of the model, and reveals whether the model is being overtrained on the studied compounds. Three approaches are commonly used for this purpose: cross-validation, permutation of the response parame-ter, and external test sets.
Cross-validation is a common technique for internal validation of a pro-posed model. The dataset is divided into groups (typically between four and

28
seven). One group is removed from the dataset, and a model is constructed from the remaining compounds. This model is used to predict the activities of the withheld compounds, and the procedure is repeated so that each cross-validation group remains outside the model once. The cross-validation coef-ficient of determination (Q2) is then calculated as a measure of how much the predictions deviate from the true values of the response parameter.
Another technique for internal model validation is to randomly permutate the order of the response variable. This results in the experimental values no longer being connected to the correct structural representation of the mole-cules. If the model based on the true order of the experimental parameter is valid, a clear difference in the predictivity compared to the random models will be apparent.
Finally, the most stringent test of a model’s predictivity is to apply it to a test set of compounds that were not included in the modeling procedure. This is usually done by keeping a representative subset outside the modeling, until the final model has been developed and internally validated using cross-validation and/or random permutation.
1.3.6. Models of drug transport processes
1.3.6.1. Intestinal drug absorption Many different approaches have been adopted for the prediction of intestinal drug absorption in silico. In the most direct of these, the absorption of orally administered drugs is modeled directly from their molecular properties.144-149 150-154 However, the interpretation is complicated by the fact that several factors influence oral drug absorption, most importantly the solubility in the gastrointestinal tract and the intestinal drug permeability (Figure 1). In addi-tion, oral absorption has only been characterized in detail for a small set of already approved drugs. Attempts to increase the dataset sizes require the inclusion of compounds with more complex pharmacokinetics, resulting in an increased uncertainty in the estimate of the fraction absorbed (FA).153, 154
An alternative approach is to instead model the discrete rate-limiting steps of oral drug absorption. This results in a greater transparency of the models, providing insight into whether solubility or permeability is rate-limiting for the absorption. Attempts to combine these two discrete rate-limiting steps in computational models of intestinal absorption have been proposed.155-158
Many different statistical methods have been applied to the prediction of permeability. These range from simple linear correlations of single parame-ters,159, 160 through multivariate linear and non-linear regression (MLR/MNLR)161-163 to more complex methodology including principal component analysis (PCA),23 partial least squares (PLS),22, 24, 164-166 genetic algorithms and neural networks.167-169

29
The molecular descriptors tend to be selected to contain information about the basic physicochemical properties believed to control the permeability process, i.e., the lipophilicity (related to the partitioning to the cell mem-brane), size, shape and flexibility (related to steric hindrance of membrane permeation) and polarity (describing the limiting effect of desolvation when the drug partitions from the water phase to the membrane).22, 24, 149, 151, 160, 162,
170-173 Computational models of intestinal permeability and absorption are reviewed in greater detail elsewhere.20, 174
A common limitation of most absorption and permeability models is that the datasets are skewed towards completely absorbed drugs. Models based on such data will, therefore, have limited predictive power when applied to discovery compounds with low to intermediate oral absorption. In addition, models generally rely on the assumption that passive transcellular permeabil-ity is the dominant transport pathway. In many cases, however, the perme-ability of compounds that were initially believed to be passively transported has later turned out to be significantly influenced by active drug transporters. Correspondingly, paracellular diffusion can contribute significantly to the permeability of low-molecular, hydrophilic compounds, further complicating the mechanistic analysis of the model results.
To enable unbiased investigations of the different drug transport pathways affecting intestinal drug permeability, the dataset used in Paper I in this the-sis was selected to evenly cover the entire range of FA in humans (0-100%). In addition, actively transported drugs were included to better reflect the situation in drug discovery where transporter affinities are not always known.
1.3.6.2. Models of drug transporters Similar to the models of intestinal drug permeability, a multitude of model-ing approaches have been applied to the transporter field.175-177 Most model-ing studies have focused on P-gp, with computational models for other ABC and SLC transporters having appeared only in recent years. Transporter models have been developed according to two general approaches: protein-based models, describing the structure of the transport proteins, and ligand-based models, describing the structural features of the drug molecules that determine their interaction with the transporter.
Models that describe the three-dimensional structure of the transporter can be used for ligand docking, in which the interactions involved in ligand-transporter binding are scored using different approaches. However, the lack of high-resolution crystal structures for human transporters so far limits the application of ligand docking procedures in transporter studies. As an alter-native, homology modeling allows the prediction of transporter structures based on their sequence similarity with a known protein structure. This is exemplified by the use of the bacterial ABC exporter Sav1866 (Figure 4A) as a template for several recent homology models of human ABC transport-

30
ers.73-77, 90 However, the sequence identity is primarily found in the ATP-binding domains, whereas the transmembrane domains that contain the ligand recognition sites exhibit larger variations, making high-precision modeling of these regions more difficult. Unfortunately, the only other struc-ture that had been published for a bacterial ABC exporter, that of MsbA from Vibrio Cholera, was recently retracted because of erroneous interpreta-tion of the structure and protein topology.178 As a result of this, homology models based on the MsbA structure need to be reevaluated.
In contrast to protein-based models, ligand-based models do not require knowledge of the three-dimensional structure of the transporter. Instead the drug molecules are examined, with the aim of finding structural similarities between ligands and thereby obtaining information about the transporter binding site. The approaches used to derive ligand-based transporter models differ both in the ways the structures are described and in which statistical methods are used to relate the structures to quantitative measures of trans-porter affinity.66, 179-195 In addition, classification models have been devel-oped that discriminate between transporter substrates and non-substrates or between inhibitors and non-inhibitors.196-200
While a wealth of data is available on drug interactions with P-gp, the ex-perimental datasets for other drug transporters are still relatively small. As a consequence, the validity of many transporter models is restricted to a lim-ited part of the chemical space, and can therefore not be used to predict in-teractions with other classes of compounds. Furthermore, small datasets limit the possibility of conducting thorough model validation using external test sets.
To conclude, the membrane permeability and interactions with transport-ers are major factors influencing the absorption, distribution, metabolism, excretion and toxicity (ADMET) profile of new drugs. It is therefore of great importance to determine these properties at an early stage of the develop-ment process. However, datasets used for the prediction of permeability are often highly skewed towards completely absorbed compounds, resulting in poor precision when models are applied to incompletely absorbed discovery compounds. Studies of drug transporters have generally been focused on P-gp, while considerably less attention has been given to other drug-transporting members of the ABC transporter family.
The focus of this thesis was, therefore, placed on studies of passive and active membrane transport, with the purpose of determining the molecular characteristics which are important for these transport mechanisms. Special consideration was given to the dataset selection. Compounds were selected to avoid skewed datasets and to maximize the structural diversity, so that the models developed would be representative of the chemical space of orally administered drugs. Furthermore, a significant effort was made to increase the datasets of BCRP and MRP2 inhibitors, enabling the development of globally applicable models.

31
2. Aims of the thesis
The general objective of this thesis was to determine the influence of passive permeability and active ABC transporter-mediated efflux on the intestinal absorption and tissue distribution of drugs. The specific aims were:
� To investigate the influence of the passive transcellular, paracellu-lar, and active transport routes in experimental models used for prediction of intestinal drug absorption (Paper I).
� To map the affinity of orally administered drugs for the ABC trans-porters BCRP and MRP2 (Papers II and III), using global com-pound datasets representing the chemical space of oral drugs.
� To find new inhibitors with specificity towards individual or multi-ple ABC transporters (Papers II-IV).
� To describe differences and similarities in the drug affinity patterns of BCRP, MRP2 and P-gp (Paper IV).
� To develop predictive computational models of intestinal drug permeability (Paper I) and of drug affinity for the ABC transporters BCRP (Papers II and IV), MRP2 (Papers III and IV) and P-gp (Paper IV).

32
3. Methods
3.1. Dataset selection To enable development of globally predictive models, all datasets in the thesis were selected to be representative of orally administered drugs. De-pending on the specific aims of each study, different additional inclusion criteria were used.
In Paper I, the aim was to predict intestinal absorption from experimental and computationally predicted permeability determinations, with a particular focus on poorly absorbed drugs. The dataset was, therefore, selected to evenly cover the entire range of FA in humans (0-100%), so that the models would not be biased towards completely absorbed drugs. In Paper II, one aim was to compare the affinity pattern of BCRP to other major ABC trans-porters. Thus, compounds with a reported affinity for other ABC transporters and compounds for which no such reports could be found were included. In Paper III a similar approach was used to study the affinity pattern of MRP2. A benchmark dataset for transporter studies was introduced that included a diverse set of compounds with affinity for a panel of important ABC and SLC transporters, as well as markers of Cytochrome P450 (CYP) metabo-lism and hepatic toxicity. The aim of Paper IV was to compare the inhibitor affinity patterns of P-gp, BCRP and MRP2. For this purpose, high-quality data on inhibition of P-gp was compiled from the literature, and was com-pared with in-house data for BCRP and MRP2 that were determined under comparable experimental conditions.
3.2. Structural diversity The structural diversity of the datasets was assessed with the ChemGPS methodology, using 2D descriptors from the SELMA descriptor collection.201 ChemGPS provides a three-dimensional map of the drug-like chemical space onto which the compounds of interest can be projected.202 In ChemGPS, the principal components calculated from a reference set of drug-like molecules are used as the x, y, and z axes of a three-dimensional coordinate system. The three most important principal components are summarized from a large number of molecular descriptors and represent mainly the size, polarity and flexibility of the molecules. The datasets used in this thesis were projected

33
onto the ChemGPS coordinate system, and their structural diversity was determined by comparing their coverage of ChemGPS chemical space with that of a reference set of 450 orally administered drugs from the Physician’s Desk Reference.203
3.3. Experimental methods
3.3.1. Preparation of filter-immobilized hexadecane membranes (HDMs)
HDMs were prepared using a protocol modified from Wohnsland and Faller.57 A mixture of hexadecane and hexane (5% v/v hexadecane) was added to polycarbonate filter inserts (Transwell Costar, Badhoevedorp, the Netherlands; diameter 12 mm; pore size 0.4μm), and the hexane was evapo-rated for at least one hour prior to conducting the permeability measure-ments. [14C]mannitol-flux was used to measure the plate-to-plate variation of the HDM membranes, using three filters on each 12-filter plate. The trans-epithelial electrical resistance (TEER) was measured repeatedly during the assay setup, and was �5000 Ohms for all filters prepared using the protocol presented above.
3.3.2. Permeability measurements Permeability measurements were performed as described previously.51, 204 Briefly, the drug dissolved in Hank’s balanced salt solution (HBSS) buffered to pH 7.4 was added to the donor side of the membrane or the cells, and buffer not containing the drug was added to the receiver side. The mem-branes were incubated in a humidified atmosphere at 37ºC. At regular time points (every 10–120 min.), samples were removed from the receiver cham-ber and the volume was replaced with fresh, preheated buffer of pH 7.4 without any drug. The filter plates were stirred at a high rate (500 rpm) to minimize the influence of the aqueous boundary layer. The integrity of the membranes was determined for each batch of filters by measuring the mem-brane permeability to [14C]mannitol. Radioactively labeled compounds were analyzed using liquid scintillation counting (1900CA TRICARB, Canberra Packard Instruments, Downers Grove, IL), and a reversed-phase gradient HPLC system was used for the unlabeled compounds.

34
Apparent permeability coefficients (Papp, cm/s) were calculated using the non-sink condition analysis,205 which is also applicable to highly permeable compounds that do not exhibit linear drug flux:
1 1
,0( )app
D RP A t
V VR R
D R D R
M MC t C eV V V V
� � � � �� �� �
�� �� �� � �
Equation 1
where VD is the volume in the donor compartment (0.5 ml), VR is the volume in the receiver compartment (1.5 ml), A is the area of the filter (1.13 cm2), M is the total amount of drug in the system, CR,0 is the drug concentration in the receiver compartment at the start of the time interval and CR(t) is the drug concentration in the receiver compartment at time t from the start of the in-terval.
3.3.3. Measurements of BCRP efflux inhibition Inhibition of BCRP-mediated mitoxantrone efflux was studied in Saos-2 cells transfected with wild-type (Arg482) human BCRP (Saos-2/wtABCG2).65 The cells were analyzed for mRNA expression of other transporters, show-ing negligible expression levels for drug transporters from the ABC and SLC families. The cells were cultured under an atmosphere of 5% CO2 at 37ºC, and were incubated with 1 μM mitoxantrone for 60 minutes, with or without the addition of 50 μM of the compound under study. The intracellular mi-toxantrone fluorescence was analyzed using a BeckmanCoulter FC500 flow cytometer (BeckmanCoulter, Fullerton, CA). To determine if the observed increase in mitoxantrone accumulation was caused by specific inhibition of BCRP, the compounds were also studied in mock-vector transfected cells. The increase in intracellular accumulation of mitoxantrone on co-incubation with the compounds under study was used as a measure of the BCRP inhibi-tion and was normalized to the value obtained using 0.5μM of the potent BCRP inhibitor Ko143 (100% inhibition).
3.3.4. Measurements of MRP2 efflux inhibition Inhibition of [3H]-estradiol-17�-D-glucuronide (E17G) transport into MRP2-overexpressing inverted Sf9 membrane vesicles was studied using a rapid filtration technique modified from Ishikawa et al.206 Briefly, membrane vesi-cles (SOLVO Biotechnology, Budapest, Hungary) were incubated with 50 μM E17G for 10 minutes, with or without the addition of 80 μM of the compound under study. The transport was stopped by addition of ice-cold buffer, and the reaction mixture was immediately filtered through a 96-well glass filter plate with a pore size of 0.65 μm (Millipore Corp., Bedford,

35
MA). The radioactivity associated with the membrane vesicles was meas-ured in a 1900CA TRICARB liquid scintillation counter (Canberra Packard Instruments, Downers Grove, IL). ATP-dependent transport rates were de-termined by subtracting the transport in vesicles incubated with AMP from the rate in ATP-incubated vesicles. The inhibitory effects of the test com-pounds were calculated as the ratio between the ATP-dependent E17G trans-port rates in the presence and absence of the test compound.
3.4. Computational methods
3.4.1. Generation of molecular descriptors Molecular structures were either entered manually into MacroModel207 (Pa-per I) or obtained from SciFinder Scholar208 (Papers II-IV). Corina209 was used to generate 3D structures (Papers II-IV). The structures were then en-ergy minimized in vacuum using MacroModel,207 with a 2000-step Polak-Ribiere conjugate gradient procedure using the MMFF94s force field and a dielectric constant of 1. After performing a pre-minimization of the struc-tures, a 500-step Monte Carlo conformational analysis (Paper I) or a 1000-step low-mode conformational analysis (Paper II) was used to identify low energy conformations. Unique conformations with energy levels lower than 50 kJ/mol above the minimum energy conformation were stored and used for further calculations of dynamic surface areas (Paper I) or for pharmacophore generation (Paper II).
A selection of 2D and 3D molecular descriptors related to the molecular size, polarity, flexibility, charge distribution, connectivity, and electroto-pological state indices was calculated using the programs Molconn-Z210 (Pa-per I), SELMA201 (Papers I and II), DragonX211 (Papers III and IV), ADMET-Predictor212 (Papers II-IV), and HYBOT213 (Papers III and IV). The free mo-lecular surface areas for each different atom type was calculated using the in-house software MAREA214 (Papers I-IV), as described previously.149
3.4.2. Pharmacophore modeling Low-energy conformations obtained from MacroModel were imported into Catalyst.215 Ten common feature pharmacophore hypotheses were developed for the BCRP inhibitors in the training set using the common features algo-rithm (HipHop) with the hydrogen bond acceptor, hydrogen bond donor, and positively ionizable, negatively ionizable, hydrophobic and ring aromatic features as possible pharmacophore features. The hypothesis that was most successful in distinguishing between BCRP inhibitors and non-inhibitors

36
was selected for visualization of the preferential orientation of the BCRP inhibitors.
3.4.3. Statistical analysis In Paper I, Spearman’s rank order correlation coefficients were used to de-termine the ability of the experimental and computational permeability mod-els to rank the dataset compounds according to their oral absorption. The rank order correlation coefficients were calculated as the linear correlation coefficient for two separate rankings of n items, according to:
1
2 2
1 1
( )( )
( ) ( )
n
i ii
s n n
i ii i
R R S Sr
R R S S
�
� �
�
�
� � Equation 2
where Ri and Si are the ranks associated with the x and y-values for com-pound i, R and S are the mean ranks for the x and y-values, respectively, and n is the number of compounds.
PLS, PLS-DA or OPLS-DA, all implemented in Simca-P,216 were used to derive multivariate models in Paper I, IV, and II-III, respectively. All mo-lecular descriptors were mean centered and scaled to unit variance, and any descriptors with a variance close to zero were excluded from the analysis. The models were evaluated using the R2 and the leave-many-out cross-validated Q2 with 7 cross-validation groups. For the discriminant models, the influence of differently sized groups was balanced by replication of the compounds belonging to the smaller classes in the datasets. To avoid bias in the models, all replicates of a molecule were removed simultaneously during the cross-validation procedure.
For the continuous PLS models, the root-mean-square error of prediction (RMSE) was used as a measure of the model predictivity:
� ���
�n
imeasuredipredictedi yy
pnRMSE
1
2,,1
1 Equation 3
where n is the number of observations, p is the number of latent variables in the PLS model, and ymeasured and ypredicted are the experimentally determined and predicted values of the response parameter, respectively.
The models were optimized using a variable selection procedure in which groups of molecular descriptors that did not contain information relevant to the problem (i.e., noise) were removed in a stepwise manner. Descriptors

37
were kept outside the model if removing them resulted in a statistically im-proved model based on the Q2 (for continuous models) or the classification accuracy for the training set (for discriminant models).
The statistical validity of the models was tested using a random permuta-tion test, in which the order of the response variable was randomly changed 100 times (Papers II-IV). Furthermore, test sets consisting of compounds not included in the model development were used to validate the models (Papers I-IV).

38
4. Results and discussion
4.1. Diversity of datasets for drug transport modeling A computational model will only be predictive within the chemical space of the compounds on which it is based. Thus, to assess the quality of a predic-tion, it is essential to know the applicability domain of the model. In this thesis, the aim was to investigate the membrane permeability and ABC transporter inhibition of compounds representing the chemical space of drug-like compounds. The emphasis was placed on orally administered drugs since they comprise the majority of the pharmaceutical market. A Intestinal permeability
Paper IB BCRP inhibition
Paper IIC MRP2 inhibition
Paper IIID ABC transporter inhibition
Paper IV
�100
102030
�6�3036�8
�4
0
4
8
t1t3
t2
�100
102030
�6�3036�8
�4
0
4
8
t1t3
t2
�100
102030
�6�3036�8
�4
0
4
8
t1t3
t2
�100
102030
�6�3036�8
�4
0
4
8
t1t3
t2
Figure 8. Structural diversity of the datasets used in the thesis. The ChemGPS axes mainly represent the size (t1), polarity (t2) and flexibility (t3). Large closed symbols denote training set compounds, large open symbols denote test set compounds, and small gray symbols denote a reference set of orally administered drugs.203
The structural diversity of the compounds was assessed using the ChemGPS methodology (Section 3.2). The datasets applied in the investigations of ABC transporter inhibition (Papers II-IV; n = 123, 191 and 126, respec-tively) covered the volume of the oral drug space satisfactorily (Figure 8B-D). In Paper I, the compounds were selected to evenly cover the entire range in human FA (0-100%; n = 30; Figure 9D). The inclusion of a large propor-tion of hydrophilic, incompletely absorbed compounds resulted in a shift towards lower values in the polarity-related second ChemGPS component (Figure 8A).
B PAMPA artificial membranes
Frac
tion
of c
ompo
unds
(%)
FA (%)
0
20
40
60
0 20 40 60 80 100
80n = 25
A Caco-2 cells
Frac
tion
of c
ompo
unds
(%)
FA (%)
0
20
40
60
0 20 40 60 80 100
80n = 38
C Human intestinal perfusion
Frac
tion
of c
ompo
unds
(%)
FA (%)
0
20
40
60
0 20 40 60 80 100
80n = 36
D Dataset in Paper I
Frac
tion
of c
ompo
unds
(%)
FA (%)
0
20
40
60
0 20 40 60 80 100
80n = 30
Figure 9. The dataset in Paper I was selected to evenly cover the entire range of FA (D). In contrast, literature datasets are often skewed towards high FA (A-C).59, 217-219

39
4.2. Influence of different transport pathways on intestinal permeability
In Paper I, the influence of different transport pathways on drug permeability predictions was studied in experimental permeability screening assays of differing complexity: the epithelial cell lines Caco-2 and 2/4/A1, and filter-immobilized artificial hexadecane membranes (HDM) (Box 1). The 2/4/A1 cells lack functional expression of several important drug transporters, and form monolayers with a more leaky, small intestine-like paracellular path-way than Caco-2 monolayers (Figure 10, top row). The HDMs completely lack active transport pathways, and, in addition, the paracellular pathway is absent in these models.
The Papp in the HDM and Caco-2 models were comparable, especially for the highly permeable compounds, whereas Papp in the small-intestine-like 2/4/A1 model was up to 100-fold higher, resulting in values comparable to those observed in the human jejunum.218, 220 This difference is most likely caused by the larger influence of the paracellular pathway in the 2/4/A1 cell line. The experimentally determined Papp was used to classify compounds as being acceptably or poorly absorbed, and to rank them according to their intestinal absorption. The cell-based models gave better classification results and resulted in considerably stronger rank order correlations between FA and Papp (rs = 0.74 and 0.73 for 2/4/A1 and Caco-2) than the HDM model (rs = 0.47; Figure 10A-C). This suggests an important role of the paracellular pathway in intestinal permeability screening. In line with the experimental results, the computational model based on 2/4/A1 permeability resulted in the best in silico prediction of FA, with a rank order correlation similar to the experimental 2/4/A1 data (rs = 0.85; Figure 10D). The computational 2/4/A1 model also successfully predicted the intestinal absorption of an external test set, resulting in a rank order correlation coefficient of 0.74.
2/4/A1
C
Rank order (Papp,2/4/A1)
0
10
20
30
0 10 20 30
Ran
k o r
der (
%FA
) 1
3
2
Caco-2
B
Rank order (Papp,Caco-2)
Ran
k or
der (
%FA
)
0
10
20
30
0 10 20 30
1
3
2
HDM
A
Rank order (Papp,HDM)
0
10
20
30
0 10 20 30
Ran
k or
der (
%FA
) 1
3
2
D
1
3
2
Rank order (Papp,2/4/A1-calculated)
0
10
20
30
0 10 20 30
Ran
k or
der (
%FA
)
Computational 2/4/A1 model
Figure 10. Rank order correlation between FA and Papp determined experimentally in the HDM (A), Caco-2 (B) and 2/4/A1 (C) models, or calculated using a computa-tional model based on 2/4/A1 Papp (D). The squares denote the training set com-pounds and the circles denote compounds in the test set. Closed symbols denote passively transported compounds, and open ones denote actively transported com-pounds. 1: Glycylsarcosine; 2: methotrexate; 3: digoxin. When these outliers were removed, the 2/4/A1 correlation (C) was excellent (rs = 0.95).

40
Permeability-limiting molecular descriptors related to polarity and hydrogen bond interactions dominated the models of Caco-2 and 2/4/A1 permeability, whereas the model based on HDM Papp had a larger influence of permeabil-ity-driving descriptors related to nonpolar interactions. This difference is probably reflecting the relative simplicity of the HDM model, where the hexadecane describes the diffusion step across the hydrophobic membrane interior.17
Notably, only three significant outliers (digoxin, glycylsarcosine, and methotrexate) were observed in the experimental 2/4/A1 correlation. Glycyl-sarcosine and methotrexate are substrates for uptake transporters,221-223 and the absence of the majority of the active transport pathways in the 2/4/A1 cell line probably explains these false predictions. The underestimation of digoxin FA could be related to the fact that compounds with a relatively low permeability coefficient may have time to encounter a larger absorptive sur-face area in the intestine. This would result in a higher absorption in vivo than is expected from the permeability (see Figure 3B). When the three out-liers were removed the experimental 2/4/A1 rank order correlation was ex-cellent (rs = 0.95).
Interestingly, FA was well predicted for all other compounds in the data-set, despite the fact that several of them are substrates for drug transporters. High intestinal drug concentrations are often observed after oral dosing,222 and the limited effect of active transport on the intestinal absorption of most drugs in Paper I might, therefore, be a result of saturation of the intestinal transporters. However, the results further show that none of the experimental models was able to predict FA for all compounds exhibiting significant ac-tive transport mechanisms in vivo. The influence of active transporters is likely to be greater for compounds that are administered in low, sub-saturating doses, or that rely on uptake transporters to gain access to the en-terocytes. For such compounds, inhibition of important drug transporters by co-administered drugs can markedly affect the intestinal absorption.39, 122, 224
4.3. Identification of inhibitors of BCRP and MRP2 Papers II and III aimed to identify inhibitors of the ABC efflux transporters BCRP (ABCG2; Paper II) and MRP2 (ABCC2; Paper III) using datasets that were representative of orally administered drugs. Inhibition of BCRP was studied in Saos-2 osteosarcoma cells over-expressing human wild-type (Arg482) BCRP, whereas inverted membrane vesicles were selected as a model system for MRP2 inhibition.
Saturable transport was demonstrated for standard substrates of each transporter (Figure 11A and B). Saos-2/wtABCG2 cells effluxed mitoxan-trone with an apparent Km of 18�9 μM (Figure 11A), and the transport was completely inhibited by the potent BCRP inhibitor Ko143 (Figure 11C). For

41
MRP2, a Km of 94�7 μM was determined for the transport of E17G (Figure 11B). The transport rate was sigmoidally related to the substrate concentra-tion, which is in line with previous reports demonstrating positive coopera-tive transport for MRP2.68, 102 MRP2-mediated transport of E17G was com-pletely inhibited by the MRP inhibitor MK571 (Figure 11D).
Figure 11. Characterization of the BCRP and MRP2 inhibition assays. A. Mitoxan-trone efflux from BCRP-expressing Saos-2 cells was saturable and followed Micha-elis-Menten kinetics. B. The sigmoidal concentration dependency of the E17G trans-port in MRP2-expressing inside-out Sf9 membrane vesicles indicated positive coop-erative transport. C. BCRP-mediated mitoxantrone transport was inhibited by the model BCRP inhibitor Ko143. D. MRP2-mediated E17G transport was inhibited by the model MRP inhibitor MK571.
Of the 123 compounds studied for BCRP inhibition, a total of 46 resulted in significantly reduced mitoxantrone efflux (Figure 12A). For MRP2, a some-what lower hit rate was observed, with 42 out of 191 compounds resulting in significantly reduced E17G transport (Figure 12B). Several previously un-known inhibitors were discovered for BCRP (n = 29) and MRP2 (n = 27).
In Paper II, we wanted to study whether BCRP inhibition functions by a similar mechanism to that observed for P-gp, where lipophilic drugs bind to the transporter from within the plasma membrane.87-90 For this purpose, com-

42
pounds spanning a wide range of lipophilicity were included in the study. Notably, no compounds with a value for logD7.4 below 0.5 were identified as inhibitors in the cell assay. The BCRP inhibitors had markedly higher logD7.4 than the non-inhibitors (with a median of 4.0 and 0.7, respectively; Figure 13), indicating that partitioning to the plasma membrane significantly influences inhibitor binding to BCRP. This observation was supported by the fact that the hydrophilic BCRP substrates nitrofurantoin and cimetidine were unable to inhibit BCRP under the standard assay conditions, which is in line with their low membrane permeability. However, at increased concentra-tions, both compounds inhibited BCRP. These results show that a high lipo-
0 100 200 300 400ATP dependent transport (% of control)
Inhibitors27/42 (64%)not previously reported
Stimulators7/13 (54%)not previously reported
A BCRP (Paper II; n = 123) B MRP2 (Paper III; n = 191)
Inhibitors29/46 (63%)not previously reported
Figure 12. Inhibition of BCRP (A) and MRP2 (B) by drug-like compounds. A. The increase in cellular mitoxantrone accumulation on co-incubation with potential BCRP inhibitors was measured for 123 compounds, resulting in the identification of 46 inhibitors. B. The relative rates of E17G transport on co-incubation with potential MRP2 inhibitors were measured for 191 compounds. Forty-two compounds resulted in a reduction in the transport rates of at least 50% (i.e., transport inhibitors). In addition, 13 compounds significantly increased the E17G transport (i.e., transport stimulators). All data are presented as means � SE.

43
philicity is not necessary for the binding of drugs to BCRP per se, but is rather determining whether the compound can reach the binding site in suffi-cient amounts to elicit an inhibitory effect.
Likewise, a higher lipophilicity was generally observed for MRP2 inhibi-tors compared to non-inhibitors (median logD7.4 = 3.4 and 0.8, respectively; Figure 13). However, MRP2 inhibitors that have also been reported to be substrates, and that likely bind competitively to the transport site, were found to be more hydrophilic than non-transported inhibitors (median logD7.4 = 0.5 and 3.6, respectively; Figure 13). This significant difference between trans-ported and non-transported inhibitors suggests that several distinct inhibitory binding sites may exist in MRP2. Further, it is possible that the lipophilic non-transported inhibitors might bind to an intramembranous site similar to that proposed for drug binding to P-gp and BCRP.
-8 -6 -4 -2 0 2 4 6logD7.4
MRP2 inhibitors not reported as substrates
MRP2 inhibitors also reported as substrates
Compounds not inhibiting MRP2
All MRP2 inhibitors
Compounds not inhibiting BCRP
BCRP inhibitors
8
Paper II
Paper III
Figure 13. Distribution of lipophilicity in the datasets in Papers II and III. The dis-tribution in inhibitors of BCRP (Paper II) and MRP2 (Paper III) is shown in white, and the distribution in non-inhibitors is shown in black. The inhibitors differ from the non-inhibitors by a shift to higher lipophilicity. For MRP2, the inhibitors that are also reported as substrates are generally less lipophilic than those not reported as substrates. The boxes show the inter-quartile distances and the median values, and the whiskers show the span between the lowest and the highest value.
4.4. Overlapping inhibitor specificity of P-gp, BCRP and MRP2
The results presented in Papers II and III demonstrated a significant overlap in the inhibitor specificities of BCRP and MRP2. These transporters are co-localized with P-gp in protective tissues throughout the human body, with important roles in restricting the intestinal absorption of foreign compounds and mediating the secretion of drugs and metabolites to bile.84, 112, 225-227 Inhi-bition of P-gp, BCRP and MRP2 can, consequently, lead to serious adverse effects, for instance through the accumulation of toxic drugs and metabolites in the liver.228, 229 However, so far, knowledge of the overlap in the affinity of ABC transporters has been based on scattered observations for single compounds or small compound series.230-234

44
In Paper IV, therefore, we examined the specificity of inhibitors towards these three major ABC efflux transporters using a common dataset of 126 structurally diverse drugs. Inhibition of BCRP was determined for 42 new compounds, and was complemented with data from Paper II. MRP2 data for the same compounds were obtained from Paper III, and P-gp data deter-mined under comparable experimental conditions were collected from the literature.
Of the 126 compounds studied, 70 inhibited at least one of the transport-ers (Figure 14). As many as 46 of the identified inhibitors affected more than one of the transporters in the investigated concentration range. Of these, 22 compounds inhibited P-gp, BCRP and MRP2. These multi-specific inhibi-tors contributed the majority (76%) of the MRP2 inhibitors and a significant proportion of the P-gp and BCRP inhibitors (39% and 41%, respectively). Notably, only about half as many inhibitors were found for MRP2 (n = 29) as for P-gp (n = 56) and BCRP (n = 53). The largest overlap was seen be-tween P-gp and BCRP, with 43 common inhibitors. In contrast, only two and one compounds were shared by P-gp and MRP2 or by BCRP and MRP2, respectively, while not affecting the third transporter. The results in Paper IV thus emphasize the significant affinity overlap previously indicated for these major drug efflux transporters.
22
1
21
2
9
4
11
56
P-gp (ABCB1)
MRP2 (ABCC2)
BCRP (ABCG2)
Non-inhibitors
Figure 14. Overlapping inhibition of P-gp, BCRP and MRP2. In total, 70 (56%) of the compounds investigated inhibited one or more of the transporters. The number of specific inhibitors were 11, 9 and 4 for P-gp, BCRP and MRP2, respectively. A large overlap was observed between P-gp and BCRP, with 43 inhibitors in common.

45
4.5. Computational models of ABC transporter inhibition
To define molecular features important for drug-mediated inhibition of BCRP and MRP2, the experimental data in Papers II and III were used to develop computational models to discriminate between inhibitors and non-inhibitors. For BCRP, the inhibitors could be distinguished from the non-inhibitors using just two molecular descriptors: logD7.4 and the molecular polarizability, the latter descriptor reflecting electron delocalization in con-jugated systems (Figure 15A). Both of these descriptors are correlated to the passive membrane permeability, further emphasizing the importance of membrane partitioning for BCRP inhibition. Despite the simplicity of the developed model, it successfully classified the compounds of the training and the test sets (Table 1).
Table 1. Prediction of BCRP and MRP2 inhibition using computational models. Correctly predicted as Transporter model inhibitors non-inhibitors BCRP Training set 93% 92% (Paper II) Test set 83% 76% MRP2 Training set 86% 86% (Paper III) Test set 72% 71%
-3
-2
-1
0
1
2
3
-6 -4 -2 0 2 4t1P
t 2O
-0.4 -0.2 0
Total structure connectivity index
Number of aromatic bonds
Calculated octanol-water
Maximum hydrogen bond
2nd largest dimension
B MRP2 (Paper III)
OPLS-DA regression coefficients0.2 0.4
A BCRP (Paper II) C MRP2 (Paper III)
logD7.4
Pol
ariz
abili
ty
-8 -6 -4 -2 0 2 4 60
20
40
60
80
100
donor energy
partition coefficient
Figure 15. Computational models of BCRP and MRP2 inhibition (from Papers II and III, respectively). In both A and B, the compounds falling in the shaded area were predicted to be non-inhibitors. Closed symbols denote those compounds which were experimentally determined to be inhibitors, and open symbols denote the non-inhibitors. The training set compounds are shown as squares and the test set com-pounds as circles. C. The five molecular descriptors in the MRP2 inhibition model are related to the lipophilicity (calculated octanol-water partition coefficient), size (2nd largest dimension, total structure connectivity index), aromaticity (number of aromatic bonds) and hydrogen bonding potential. Descriptors with positive coeffi-cients have higher values in inhibitors, and descriptors with negative coefficients have higher values in non-inhibitors.

46
Interestingly, the computational model of MRP2 inhibition developed in Paper III showed a similar importance of lipophilicity and aromaticity as the BCRP model. A larger number of molecular descriptors was needed to dis-criminate between inhibitors and non-inhibitors for MRP2 than for BCRP, possibly reflecting a more complex binding pattern with several drug bind-ing sites in MRP2 (Figure 15C; Table 1).
The molecular features important for inhibition of ABC transporters were further investigated in Paper IV. First, individual models were developed for P-gp, BCRP and MRP2 (Figure 16A-C). The size, shape, lipophilicity and aromaticity were the dominating factors distinguishing inhibitors from non-inhibitors for the dataset in Paper IV, which only partly overlapped with the datasets used in Papers II and III. The similarities between the computational models for each transporter suggested that these molecular properties might be related to the large number of multi-specific inhibitors affecting all three transporters. We therefore modeled the multi-specific inhibitors in independ-ent of the specific ones, confirming the importance of lipophilicity and aro-maticity for the overlapping inhibitors (Figure 16D). The median logD7.4 was 4.6 for the multi-specific inhibitors, indicating that they will accumulate in the plasma membrane. This suggests that this group of inhibitors may inter-act with binding sites located in the membrane-protein interface. The fact that a large number of the multi-specific inhibitors have been reported to be
-0.5 -0.25 0 0.25 0.5
logD7.4
n aromatic bonds
Total structure
ABCInhibitors vs. non-inhibitors
E
Length in
Length in 2nd
n aromatic rings
MRP2C
n aromatic rings
logD7.4
BCRPB
Total structure
logD7.4
P-gpA
Higher valuesHigher values
-0.6 -0.4 -0.2 0 0.2 0.4 0.6PLS regression coefficients
-0.6 -0.4 -0.2 0 0.2 0.4 0.6PLS regression coefficients
-0.6 -0.4 -0.2 0 0.2 0.4 0.6PLS regression coefficients
n aromatic bondsn aromatic rings
logD7.4
n multiple bonds
OverlappingInhibitors vs. non-inhibitors
D
-0.5 -0.25 0 0.25 0.5PLS regression coefficientsPLS regression coefficients
Inhibitors vs.non-inhibitors
Inhibitors vs.non-inhibitors
Inhibitors vs.non-inhibitors
connectivity index largest dimension
(double, triple, aromatic) connectivity index
in non-inhibitors in inhibitorsHigher valuesHigher values
in non-inhibitors in inhibitorsHigher valuesHigher values
in non-inhibitors in inhibitors
Higher valuesHigher valuesin non-inhibitors in overlapping
Higher valuesHigher valuesin non-inhibitors in ABC inhibitors
Figure 16. Prediction of ABC transporter inhibitors from: computational models of P-gp inhibitors (A), BCRP inhibitors (B), MRP2 inhibitors (C) and models of over-lapping (D) and general (E) ABC transporter inhibition. Descriptors related to the size, shape, lipophilicity and aromaticity were important for all three models of individual transporters (A-C). This importance was mirrored in the models of over-lapping and general ABC transporter inhibition (D and E), indicating that these properties might be related to the multi-specific inhibitors.

47
P-gp substrates235 indicates that such a site may be part of the intra-membranous substrate binding pocket in P-gp.
Finally, the large inhibitor overlap and the similarities between the indi-vidual transporter models suggested that it would be possible to derive a model describing general ABC inhibition. We therefore used the same ap-proach as for the individual models to develop a model capable of discrimi-nating between compounds inhibiting any of the three ABC transporters from compounds lacking an inhibitory effect. The resultant model mirrored the importance of lipophilicity and aromaticity revealed in the individual models. The classification accuracy was 82% of the inhibitors and 85% of the non-inhibitors in the test set (Figure 16), which is an improvement to most published models of drug-transporter interaction.196, 197, 200

48
5. Conclusions and future outlook
In this thesis, the role of passive membrane permeability and ABC transport proteins on drug absorption and disposition was investigated. Particular em-phasis was placed on defining the molecular properties of importance for these transport mechanisms. Experimental models that include the paracellu-lar pathway were found to result in improved predictions of intestinal drug absorption, especially for incompletely absorbed drugs. Previously unknown inhibitors of BCRP and MRP2 were identified from datasets that were repre-sentative of orally administered drugs. Finally, the overlap in the inhibitor affinity between P-gp, BCRP and MRP2 was described and a majority of the inhibitors was found to affect more than one of these transporters. The work in this thesis demonstrates the usefulness of simplistic approaches, where different mechanisms are studied in isolation. Such data provide the basis for understanding the intricate interplay affecting cellular drug disposition. The specific conclusions were:
� The paracellular drug transport pathway plays a significant role in the
prediction of the intestinal drug permeability of incompletely absorbed drugs.
� Permeability data from the 2/4/A1 cell model, which has a small intes-tine-like paracellular pathway, gave better classification and ranking of drugs according to their intestinal absorption than the artificial hexadec-ane membrane model which exclusively models the hydrophobic interior of the plasma membrane.
� A large number of structurally diverse drugs inhibited the ABC trans-porters BCRP and MRP2. The chemical space of BCRP inhibitors was similar to that of P-gp, whereas fewer drug-like compounds inhibited MRP2.
� The multi-specific ABC transporter inhibitors, i.e., those that affected P-gp, BCRP and MRP2, were, in general, highly lipophilic weak bases.
� The passive membrane permeability to drugs and the inhibitor affinity for ABC transporters could be modeled using easily interpreted molecu-lar descriptors. The models can be used as predictive filters in drug dis-covery to guide further experimental efforts.

49
The research described in this thesis has led to an increased knowledge of the mechanisms determining the intestinal permeability of incompletely ab-sorbed drugs and of the molecular characteristics of importance for inhibi-tion of the major ABC efflux transporters P-gp, BCRP and MRP2.
In human tissues, the transport mechanisms studied here act in concert with one another and with additional transporters from the ABC and SLC families. A drug can be a substrate of several different transporters and can also influence the disposition of coadministered drugs by inhibiting one or more transporters, resulting in clinically important drug-drug interactions. Furthermore, transporter inhibition can lead to accumulation of toxic drugs and metabolites and subsequent cellular toxicity. A major challenge for fu-ture investigations in this field is, therefore, to clarify the interplay between different transport mechanisms.
Much work also remains to be done in terms of characterizing the influ-ence of membrane transport on intracellular processes. The transport into the cell will affect the presentation of the drug for intracellular pharmacological targets and metabolizing enzymes, thereby influencing both the pharmacoki-netics and the pharmacodynamics of the drug. In addition, many drug trans-porters are expressed in the membranes of intracellular organelles. This can result in widely differing drug concentrations in different subcellular com-partments, sometimes leading to subcellular toxicity.
Approaches to the study of complex interplay between different cellular mechanisms include the use of cells transfected with multiple drug trans-porters and knock-out animal models. Further, pharmacokinetic and systems biology modeling can be applied to cellular drug flux, thereby gaining an insight into the relative importance of different transport pathways. The data presented here for specific transport mechanisms will be useful for selecting suitable model drugs and identifying the optimal experimental conditions for such endeavors.

50
6. Svensk populärvetenskaplig sammanfattning
För att ett läkemedel ska kunna ge den önskade effekten krävs att läkeme-delsmolekylerna når fram till målcellen. Kroppens organ skyddas av olika vävnadsbarriärer, och för att passera dessa måste läkemedlet ta sig in i och igenom kroppens celler. Transporten genom cellernas omgivande membran är därför avgörande för läkemedlets effekt.
En molekyl kan passera genom cellmembranet genom två fundamentalt olika mekanismer: passiv diffusion och aktiv transport. Diffusion innebär att molekylerna transporteras i riktning från en hög koncentration till en låg, till exempel från tarmens inre efter att en tablett svalts, och vidare till blodom-loppet. Vid aktiv transport pumpas läkemedelsmolekylerna istället genom cellmembranet med hjälp av transportproteiner som sitter insprängda i mem-branet. I kroppens alla celler hjälper en mängd olika transportproteiner till att ta upp näringsämnen från blodet och att pumpa ut nedbrytningsprodukter och gifter. I denna avhandling har arbetet fokuserat på tre medlemmar av en av de största familjerna av transportproteiner, ABC-transportörerna. Dessa har en viktig roll i att påverka hur läkemedel absorberas från tarmen, hur läkemedlen fördelas i kroppen, samt hur de utsöndras efter att de givit den önskade effekten.
När en patient tar flera läkemedel samtidigt, kan dessa påverka varandras effekt. Om ett läkemedel hämmar transporten av ett annat, kan det leda till att detta inte når sitt mål och att effekten uteblir. Hämning av transporten kan också göra att giftiga substanser anrikas i kroppens vävnader, vilket kan leda till allvarliga biverkningar. I avhandlingen undersöktes därför ett stort antal olika läkemedel, för att utreda hur de påverkar transportproteinerna breast cancer resistance protein (BCRP) och multidrug-resistance associated pro-tein 2 (MRP2). Ett avsevärt antal läkemedel som används i klinisk praxis visades hämma dessa transportproteiner. Ytterligare undersökningar krävs dock för att visa ifall detta kan leda till allvarliga biverkningar.
De experimentella resultaten användes vidare för att utveckla datorbase-rade modeller som beskriver hur läkemedlens kemiska struktur påverkar om de hämmar transportproteinet eller inte. Bland annat visades att läkemedel som påverkar flera ABC-transportörer är mer fettlösliga än andra läkemedel. Datorbaserade modeller av detta slag kan användas för att förutsäga ifall nya läkemedel kommer påverka transporten av andra läkemedel. På så vis kan

51
läkemedel som riskerar att ge allvarliga biverkningar sorteras bort i ett tidigt stadium av utvecklingsprocessen. Detta leder till att nya, säkrare läkemedel kan utvecklas effektivt och till en lägre kostnad, vilket i slutändan även gyn-nar patienterna.

52
7. Acknowledgements
The studies were carried out at the Department of Pharmacy, Faculty of Pharmacy, Uppsala University, Sweden.
The financial support from the Swedish Research Council (Grant 9478), the Knut and Alice Wallenberg Foundation, the Swedish Foundation for Strategic Research, the Swedish Fund for Research without Animal Experiments, and the Swedish Ani-mal Welfare Agency is gratefully acknowledged. My participation in research con-ferences and courses was made possible by travel grants from IF:s stiftelse, Apote-kare C. D. Carlssons stiftelse, the ULLA Consortium, Rektors Wallenbergmedel, and Anna Maria Lundins stipendiefond.
I wish to express my sincere gratitude to all of you who, in some way, contributed to this thesis:
Mina handledare: professor Per Artursson, för att du alltid strävar efter att göra arbe-tena ett steg bättre. Det gör ont att forska – men det är värt det! Tack också för din kreativitet och entusiasm, oavsett om det gäller transportörer, balroger eller att dra paralleller mellan forskning och Star Wars episod III. Professor Ulf Norinder, för dina entusiastiska utläggningar om storheten i olika statistiska metoder och oduglig-heten hos alla andra som försöker använda dem. Tack också för dina bullar, dina deskriptorer, dina ekvilibristiska email, din stora ödmjukhet och dina aldrig sinande små vänligheter.
Professor Martin Malmsten och professor Göran Alderborn för tillhandahållandet av fina mintgröna lokaler och en mycket god arbetsmiljö.
Min låtsashandledare Dr. Kix Bergström för att du alltid förvarnar innan du kommer till rummet och för att du alltid tar dig tid att läsa, kommentera, hjälpa och skratta – med, inte åt.
Mina övriga medförfattare: Dr. Gunilla Englund, Dr. Staffan Tavelin, Dr. Naoki Nagahara, Gustav Ahlin, och ente minst Jenny Pedersen, för ert engagemang för att göra alla arbeten så bra som de är!
Gamla och nyare medlemmar i cell-, transport- och prediktionsgruppen. Särskilt tack till Dr. Lucia Lazorova, för all kunskap om celler, medier, apparater och annat som du frikostigt delar med dig av. Och framförallt, tack för ditt varma hjärta, för all omtanke och för den glädje du sprider omkring dig. Dr Eva Ragnarsson, för all hjälp när du, och sedan jag, var surrogat-Lucior. Dr. Carola Wassvik, för alla diskussioner om molekyldeskriptorer, korsvalidering och variabelselektion. Johan Gråsjö, för

53
hängivet härledande av allehanda matematiska samband och för starkt kaffe i fika-rummet. Dr. Helena Engman (och Dr. Christer Tammerfors), för den fina introduk-tionen till forskarvärlden jag fick under mitt exjobb.
My students Gustav Ahlin, Aki Heikkinen, Nina Ginman, Jenny Krusemo, Pia Brokhøj, Jenny Olofsson/Pedersen och Emma Hillner, for all the good work you contributed, and for good times in the lab and outside.
Mina rumskamrater, Dr. Nadia Merclin, för att vi aldrig behövde köpa en radio till rummet och för den fina adventljusstaken som gör att vårt rum alltid vinner julpynts-tävlingen. Gustav, för att du delar med dig av din högst varierande musiksmak, för att du aldrig säger nej till en öl, för trevligt resesällskap, om det så är till Göteborg, Kansas City eller Kanazawa, men framför allt, för att du pyntar som en gud!
Eva Nises Ahlgren, Ulla Wästberg-Galik, Harriet Pettersson, Christin Magnusson, Göran Frenning, Elisabet Börjesson, Göran Ocklind, Eva Lide Johansson, Birgitta Rylén och Lotta Wahlberg för all hjälp med praktiska saker.
Nuvarande och frånvarande medarbetare på institutionen för farmaci, särskilt alla doktorander som gjort att det alltid har varit kul att komma till jobbet, och att gå därifrån när det vankas fredagsöl, pingisturneringar, innebandy eller finlandsresor. Tack särskilt till Tobias för att jag fick låna din fotkräm i Paris, och till Peter för alla gånger du kommit och berättat nåt roligt (efter att först stått tyst och väntat en kvart i dörröppningen).
Rector Magnificus för det tidiga stödet på min väg längs den kemiskt vetenskapliga banan.
Kulturföreningen Dragskåpet® och alla andra vänner för allt roligt! Onsdagsölgänget för alla intressanta diskussioner och galna helger.
Johan Unga för alla informativa email om vilket väder det är i Göteborg, och Peter och Maria för att det inte stannar vid ett. Vad sägs om en mirgare i januari? Johan bestämmer plats, Maria bestämmer tid.
Vinprovargänget: Emma, Fredrik, Arvid, Anna, Maria och Fredrik. Tack för alla insiktsfulla vinkommentarer och för er värme! Tack också för att ni alltid låter mig vinna.
Maria, min syster! Tack för att du alltid finns där, och för alla samtal om Dr. Carter och om allt annat.
Min extrafamilj Torgny och Anna-Karin, Linda, Henrik, Molly och Sixten och Jonas (min bundsförvant), för att ni tagit emot mig med sådan värme!
Pappa och mamma, för all kärlek ni gett mig under hela mitt liv. För att ni alltid finns där och hjälper när det behövs, och för att ni alltid uppmuntrat mig. Allt bra kommer från er!
Elin, du betyder allt för mig. Ditt stöd och att du tror på mig har hjälpt mig mer än du någonsin kan ana. Jag älskar dig!

54
8. References and notes
1. Kennedy, T., Managing the drug discovery/development interface. Drug Discov Today
1997, 2, (10), 436-444. 2. Kerns, E. H.; Di, L., Pharmaceutical profiling in drug discovery. Drug Discov Today
2003, 8, (7), 316-323. 3. Kola, I.; Landis, J., Can the pharmaceutical industry reduce attrition rates? Nat Rev
Drug Discov 2004, 3, (8), 711-715. 4. van Meer, G., Polarity and Polarized Transport of Membrane Lipids in a Cultured Epi-
thelium. In Functional Epithelial Cells in Culture, Valentich, J. D., Ed. Alan R. Liss: New York, NY, 1989; Vol. 8, pp 43-69.
5. van Meer, G.; Simons, K., The function of tight junctions in maintaining differences in lipid composition between the apical and the basolateral cell surface domains of MDCK cells. EMBO J 1986, 5, (7), 1455-1464.
6. Reuss, L., Tight Junction Permeability to Ions and Water. In Tight Junctions, 2nd ed.; Anderson, J., Ed. CRC Press: Boca Raton, FL., 2001; pp 61-88.
7. Collander, R., The permeability of Nitella cells to non-electrolytes. Physiol Plant 1954, 7, 420-445.
8. Shore, P.; Brodie, B.; Hogben, C., The gastric secretion of drugs: a pH-partition hy-pothesis. J Pharmacol Exp Ther 1957, 119, 361-369.
9. Avdeef, A., Physicochemical profiling (solubility, permeability and charge state). Curr Top Med Chem 2001, 1, (4), 277-351.
10. Avdeef, A.; Testa, B., Physicochemical profiling in drug research: a brief survey of the state-of-the-art of experimental techniques. Cell Mol Life Sci 2002, 59, (10), 1681-1689.
11. Testa, B.; Carrupt, P.-A.; Gaillard, P.; Billois, F.; Weber, P., Lipophilicity in Molecular Modelling. Pharm Res 1996, 13, (3), 335-343.
12. Marrink, S. J.; Berendsen, H. J. C., Simulation of Water Transport through a Lipid-Membrane. J Phys Chem 1994, 98, (15), 4155-4168.
13. Burton, P. S.; Conradi, R. A.; Hilgers, A. R.; Ho, N. F. H.; Maggiora, L. L., The rela-tionship between peptide structure and transport across epithelial cell monolayers. J Control Release 1992, 19, (1-3), 87-97.
14. Goodwin, J. T.; Conradi, R. A.; Ho, N. F.; Burton, P. S., Physicochemical determinants of passive membrane permeability: role of solute hydrogen-bonding potential and vol-ume. J Med Chem 2001, 44, (22), 3721-3729.
15. Bassolino, D.; Alper, H.; Stouch, T. R., Drug-membrane interactions studied by mo-lecular dynamics simulation: size dependence of diffusion. Drug Des Discov 1996, 13, (3-4), 135-141.

55
16. Jacobs, R. E.; White, S. H., The Nature of the Hydrophobic Binding of Small Peptides at the Bilayer Interface - Implications for the Insertion of Transbilayer Helices. Bio-chemistry 1989, 28, (8), 3421-3437.
17. Marrink, S. J.; Berendsen, H. J. C., Permeation process of small molecules across lipid membranes studied by molecular dynamics simulations. J Phys Chem 1996, 100, (41), 16729-16738.
18. Stouch, T. R.; Alper, H. E.; Bassolino, D., Simulations of Drug Diffusion in Biomem-branes. Comput Aided Mol Des 1995, 589, 127-138.
19. Stouch, T. R.; Bassolino, D. A., Size dependence of permeant diffusion through lipid bilayer membranes: A molecular dynamics simulation study. Biophys J 1996, 70, (2), Tuam5-Tuam5.
20. Artursson, P.; Matsson, P., Absorption Prediction. In Pharmaceutical Profiling in Drug Discovery for Lead Selection, Wang, B., Ed. AAPS Press: Arlington, VA., 2004; pp 3-26.
21. Stenberg, P.; Bergström, C. A.; Luthman, K.; Artursson, P., Theoretical predictions of drug absorption in drug discovery and development. Clin Pharmacokinet 2002, 41, (11), 877-899.
22. Cruciani, C.; Crivori, P.; Carrupt, P. A.; Testa, B., Molecular fields in quantitative structure-permeation relationships: the VolSurf approach. J Mol Struct 2000, 503, (1-2), 17-30.
23. van de Waterbeemd, H.; Camenisch, G.; Folkers, G.; Raevsky, O. A., Estimation of Caco-2 cell permeability using calculated molecular descriptors. Quant Struct-Act Rel 1996, 15, (6), 480-490.
24. Österberg, T.; Norinder, U., Prediction of drug transport processes using simple pa-rameters and PLS statistics. The use of ACD/logP and ACD/ChemSketch descriptors. Eur J Pharm Sci 2001, 12, (3), 327-337.
25. Fine, K. D.; Santa Ana, C. A.; Porter, J. L.; Fordtran, J. S., Effect of changing intestinal flow rate on a measurement of intestinal permeability. Gastroenterology 1995, 108, (4), 983-989.
26. Fihn, B. M.; Jodal, M., Permeability of the proximal and distal rat colon crypt and surface epithelium to hydrophilic molecules. Pflugers Arch 2001, 441, (5), 656-662.
27. Adson, A.; Burton, P. S.; Raub, T. J.; Barsuhn, C. L.; Audus, K. L.; Ho, N. F. H., Pas-sive diffusion of weak organic electrolytes across Caco-2 cell monolayers: Uncoupling the contributions of hydrodynamic, transcellular and paracellular barriers. J Pharm Sci 1995, 84(10), 1197-1204.
28. Flanagan, S. D.; Takahashi, L. H.; Liu, X.; Benet, L. Z., Contributions of saturable active secretion, passive transcellular, and paracellular diffusion to the overall transport of furosemide across adenocarcinoma (Caco-2) cells. J Pharm Sci 2002, 91, (4), 1169-1177.
29. Lee, K.; Thakker, D. R., Saturable transport of H2-antagonists ranitidine and famotidine across Caco-2 cell monolayers. J Pharm Sci 1999, 88, (7), 680-687.
30. Zhou, S. Y.; Piyapolrungroj, N.; Pao, L.; Li, C.; Liu, G.; Zimmermann, E.; Fleisher, D., Regulation of paracellular absorption of cimetidine and 5-aminosalicylate in rat intes-tine. Pharm Res 1999, 16, (11), 1781-1785.
31. Ueno, M., Molecular anatomy of the brain endothelial barrier: an overview of the distri-butional features. Curr Med Chem 2007, 14, (11), 1199-1206.

56
32. Lai, C. H.; Kuo, K. H.; Leo, J. M., Critical role of actin in modulating BBB permeabil-ity. Brain Res Brain Res Rev 2005, 50, (1), 7-13.
33. Fihn, B. M.; Sjöqvist, A.; Jodal, M., Permeability of the rat small intestinal epithelium along the villus-crypt axis: effects of glucose transport. Gastroenterology 2000, 119, (4), 1029-1036.
34. Söderholm, J. D.; Olaison, G.; Kald, A.; Tagesson, C.; Sjödahl, R., Absorption profiles for polyethylene glycols after regional jejunal perfusion and oral load in healthy hu-mans. Dig Dis Sci 1997, 42, (4), 853-857.
35. Venter, J. C.; Adams, M. D.; Myers, E. W.; Li, P. W.; Mural, R. J.; Sutton, G. G.; Smith, H. O.; Yandell, M., et al., The sequence of the human genome. Science 2001, 291, (5507), 1304-1351.
36. Hollenstein, K.; Dawson, R. J.; Locher, K. P., Structure and mechanism of ABC trans-porter proteins. Curr Opin Struct Biol 2007, 17, (4), 412-418.
37. Hediger, M. A.; Romero, M. F.; Peng, J. B.; Rolfs, A.; Takanaga, H.; Bruford, E. A., The ABCs of solute carriers: physiological, pathological and therapeutic implications of human membrane transport proteinsIntroduction. Pflugers Arch 2004, 447, (5), 465-468.
38. Tsuji, A.; Tamai, I., Carrier-mediated intestinal transport of drugs. Pharm Res 1996, 13, (7), 963-977.
39. Zaher, H.; Khan, A. A.; Palandra, J.; Brayman, T. G.; Yu, L.; Ware, J. A., Breast cancer resistance protein (Bcrp/abcg2) is a major determinant of sulfasalazine absorption and elimination in the mouse. Mol Pharm 2006, 3, (1), 55-61.
40. Hoffmeyer, S.; Burk, O.; von Richter, O.; Arnold, H. P.; Brockmoller, J.; Johne, A.; Cascorbi, I.; Gerloff, T., et al., Functional polymorphisms of the human multidrug-resistance gene: multiple sequence variations and correlation of one allele with P-glycoprotein expression and activity in vivo. Proc Natl Acad Sci U S A 2000, 97, (7), 3473-3478.
41. Morimoto, K.; Nakakariya, M.; Shirasaka, Y.; Kakinuma, C.; Fujita, T.; Tamai, I.; Ogihara, T., Oseltamivir (TamifluTM) efflux transport at the blood-brain barrier via P-glycoprotein. Drug Metab Dispos 2007, epub.
42. Schinkel, A. H.; Wagenaar, E.; Mol, C. A.; van Deemter, L., P-glycoprotein in the blood-brain barrier of mice influences the brain penetration and pharmacological activ-ity of many drugs. J Clin Invest 1996, 97, (11), 2517-2524.
43. Hirano, M.; Maeda, K.; Matsushima, S.; Nozaki, Y.; Kusuhara, H.; Sugiyama, Y., Involvement of BCRP (ABCG2) in the biliary excretion of pitavastatin. Mol Pharmacol 2005, 68, (3), 800-807.
44. Linton, K. J.; Higgins, C. F., Structure and function of ABC transporters: the ATP switch provides flexible control. Pflugers Arch 2007, 453, (5), 555-567.
45. Dean, M.; Allikmets, R., Complete characterization of the human ABC gene family. J Bioenerg Biomembr 2001, 33, (6), 475-479.
46. Young, S. G.; Fielding, C. J., The ABCs of cholesterol efflux. Nat Genet 1999, 22, (4), 316-318.
47. de Vree, J. M.; Jacquemin, E.; Sturm, E.; Cresteil, D.; Bosma, P. J.; Aten, J.; Deleuze, J. F.; Desrochers, M., et al., Mutations in the MDR3 gene cause progressive familial intra-hepatic cholestasis. Proc Natl Acad Sci U S A 1998, 95, (1), 282-287.
48. Paulusma, C. C.; Kool, M.; Bosma, P. J.; Scheffer, G. L.; ter Borg, F.; Scheper, R. J.; Tytgat, G. N.; Borst, P., et al., A mutation in the human canalicular multispecific or-

57
ganic anion transporter gene causes the Dubin-Johnson syndrome. Hepatology 1997, 25, (6), 1539-1542.
49. Riordan, J. R.; Rommens, J. M.; Kerem, B.; Alon, N.; Rozmahel, R.; Grzelczak, Z.; Zielenski, J.; Lok, S., et al., Identification of the cystic fibrosis gene: cloning and char-acterization of complementary DNA. Science 1989, 245, (4922), 1066-1073.
50. Artursson, P., Epithelial transport of drugs in cell culture. I: A model for studying the passive diffusion of drugs over intestinal absorptive (Caco-2) cells. J Pharm Sci 1990, 79, (6), 476-482.
51. Hubatsch, I.; Ragnarsson, E. G.; Artursson, P., Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nat Protoc 2007, 2, (9), 2111-2119.
52. Carriere, V.; Lesuffleur, T.; Barbat, A.; Rousset, M.; Dussaulx, E.; Costet, P.; de Wa-ziers, I.; Beaune, P., et al., Expression of cytochrome P-450 3A in HT29-MTX cells and Caco-2 clone TC7. FEBS Lett 1994, 355, (3), 247-250.
53. Raeissi, S. D.; Guo, Z.; Dobson, G. L.; Artursson, P.; Hidalgo, I. J., Comparison of CYP3A activities in a subclone of Caco-2 cells (TC7) and human intestine. Pharm Res 1997, 14, (8), 1019-1025.
54. Pontier, C.; Pachot, J.; Botham, R.; Lenfant, B.; Arnaud, P., HT29-MTX and Caco-2/TC7 monolayers as predictive models for human intestinal absorption: role of the mu-cus layer. J Pharm Sci 2001, 90, (10), 1608-1619.
55. Paul, E. C.; Hochman, J.; Quaroni, A., Conditionally immortalized intestinal epithelial cells: novel approach for study of differentiated enterocytes. Am J Physiol 1993, 265, (1 Pt 1), C266-C278.
56. Tavelin, S.; Milovic, V.; Ocklind, G.; Olsson, S.; Artursson, P., A conditionally immor-talized epithelial cell line for studies of intestinal drug transport. J Pharmacol Exp Ther 1999, 290, (3), 1212-1221.
57. Wohnsland, F.; Faller, B., High-throughput permeability pH profile and high-throughput alkane/water log P with artificial membranes. J Med Chem 2001, 44, (6), 923-930.
58. Avdeef, A.; Bendels, S.; Di, L.; Faller, B.; Kansy, M.; Sugano, K.; Yamauchi, Y., PAMPA-critical factors for better predictions of absorption. J Pharm Sci 2007, 96, (11), 2893-2909.
59. Kansy, M.; Senner, F.; Gubernator, K., Physicochemical high throughput screening: parallel artificial membrane permeation assay in the description of passive absorption processes. J Med Chem 1998, 41, (7), 1007-1010.
60. Borst, P.; Evers, R.; Kool, M.; Wijnholds, J., The multidrug resistance protein family. Biochim Biophys Acta 1999, 1461, (2), 347-357.
61. Smit, J. W.; Weert, B.; Schinkel, A. H.; Meijer, D. K., Heterologous expression of various P-glycoproteins in polarized epithelial cells induces directional transport of small (type 1) and bulky (type 2) cationic drugs. J Pharmacol Exp Ther 1998, 286, (1), 321-327.
62. Kimchi-Sarfaty, C.; Alexander, N. S.; Brittain, S.; Ali, S.; Gottesman, M. M., Transduc-tion of multiple cell types using improved conditions for gene delivery and expression of SV40 pseudovirions packaged in vitro. Biotechniques 2004, 37, (2), 270-275.
63. Urban, T. J.; Gallagher, R. C.; Brown, C.; Castro, R. A.; Lagpacan, L. L.; Brett, C. M.; Taylor, T. R.; Carlson, E. J., et al., Functional genetic diversity in the high-affinity car-nitine transporter OCTN2 (SLC22A5). Mol Pharmacol 2006, 70, (5), 1602-1611.

58
64. Brezden, C. B.; Hedley, D. W.; Rauth, A. M., Constitutive expression of P-glycoprotein as a determinant of loading with fluorescent calcium probes. Cytometry 1994, 17, (4), 343-348.
65. Wierdl, M.; Wall, A.; Morton, C. L.; Sampath, J.; Danks, M. K.; Schuetz, J. D.; Potter, P. M., Carboxylesterase-mediated sensitization of human tumor cells to CPT-11 cannot override ABCG2-mediated drug resistance. Mol Pharmacol 2003, 64, (2), 279-288.
66. Saito, H.; Hirano, H.; Nakagawa, H.; Fukami, T.; Oosumi, K.; Murakami, K.; Kimura, H.; Kouchi, T., et al., A new strategy of high-speed screening and quantitative structure-activity relationship analysis to evaluate human ATP-binding cassette transporter ABCG2-drug interactions. J Pharmacol Exp Ther 2006, 317, (3), 1114-1124.
67. Özvegy, C.; Litman, T.; Szakacs, G.; Nagy, Z.; Bates, S.; Varadi, A.; Sarkadi, B., Func-tional characterization of the human multidrug transporter, ABCG2, expressed in insect cells. Biochem Biophys Res Commun 2001, 285, (1), 111-117.
68. Zelcer, N.; Huisman, M. T.; Reid, G.; Wielinga, P.; Breedveld, P.; Kuil, A.; Knipscheer, P.; Schellens, J. H., et al., Evidence for two interacting ligand binding sites in human multidrug resistance protein 2 (ATP binding cassette C2). J Biol Chem 2003, 278, (26), 23538-23544.
69. Loo, T. W.; Clarke, D. M., Drug-stimulated ATPase activity of human P-glycoprotein is blocked by disulfide cross-linking between the nucleotide-binding sites. J Biol Chem 2000, 275, (26), 19435-19438.
70. Higgins, C. F., Multiple molecular mechanisms for multidrug resistance transporters. Nature 2007, 446, (7137), 749-757.
71. Dawson, R. J.; Locher, K. P., Structure of a bacterial multidrug ABC transporter. Na-ture 2006, 443, (7108), 180-185.
72. Zolnerciks, J. K.; Wooding, C.; Linton, K. J., Evidence for a Sav1866-like architecture for the human multidrug transporter P-glycoprotein. Faseb J 2007, 21, (14), 3937-3948.
73. Shilling, R. A.; Venter, H.; Velamakanni, S.; Bapna, A.; Woebking, B.; Shahi, S.; van Veen, H. W., New light on multidrug binding by an ATP-binding-cassette transporter. Trends Pharmacol Sci 2006, 27, (4), 195-203.
74. Ravna, A. W.; Sylte, I.; Sager, G., Molecular model of the outward facing state of the human P-glycoprotein (ABCB1), and comparison to a model of the human MRP5 (ABCC5). Theor Biol Med Model 2007, 4, (1), epub.
75. O'Mara, M. L.; Tieleman, D. P., P-glycoprotein models of the apo and ATP-bound states based on homology with Sav1866 and MalK. FEBS Lett 2007, 581, (22), 4217-4222.
76. Sakurai, A.; Onishi, Y.; Hirano, H.; Seigneuret, M.; Obanayama, K.; Kim, G.; Liew, E. L.; Sakaeda, T., et al., Quantitative structure--activity relationship analysis and molecu-lar dynamics simulation to functionally validate nonsynonymous polymorphisms of human ABC transporter ABCB1 (P-glycoprotein/MDR1). Biochemistry 2007, 46, (26), 7678-7693.
77. Degorter, M. K.; Conseil, G.; Deeley, R. G.; Campbell, R. L.; Cole, S. P., Molecular modeling of the human multidrug resistance protein 1 (MRP1/ABCC1). Biochem Bio-phys Res Commun 2008, 365, (1), 29-34.
78. Schinkel, A. H.; Jonker, J. W., Mammalian drug efflux transporters of the ATP binding cassette (ABC) family: an overview. Adv Drug Deliv Rev 2003, 55, (1), 3-29.

59
79. McDevitt, C. A.; Collins, R. F.; Conway, M.; Modok, S.; Storm, J.; Kerr, I. D.; Ford, R. C.; Callaghan, R., Purification and 3D structural analysis of oligomeric human mul-tidrug transporter ABCG2. Structure 2006, 14, (11), 1623-1632.
80. Henriksen, U.; Fog, J. U.; Litman, T.; Gether, U., Identification of intra- and intermo-lecular disulfide bridges in the multidrug resistance transporter ABCG2. J Biol Chem 2005, 280, (44), 36926-36934.
81. Juliano, R. L.; Ling, V., A surface glycoprotein modulating drug permeability in Chi-nese hamster ovary cell mutants. Biochim Biophys Acta 1976, 455, (1), 152-162.
82. Based on a search of the PubMed literature database (http://www.ncbi.nlm.nih.gov), using the search terms (P-glycoprotein OR P-gp OR mdr1 OR ABCB1).
83. Hoffmann, U.; Kroemer, H. K., The ABC transporters MDR1 and MRP2: multiple functions in disposition of xenobiotics and drug resistance. Drug Metab Rev 2004, 36, (3-4), 669-701.
84. Thiebaut, F.; Tsuruo, T.; Hamada, H.; Gottesman, M. M.; Pastan, I.; Willingham, M. C., Cellular localization of the multidrug-resistance gene product P-glycoprotein in normal human tissues. Proc Natl Acad Sci U S A 1987, 84, (21), 7735-7738.
85. Thiebaut, F.; Tsuruo, T.; Hamada, H.; Gottesman, M. M.; Pastan, I.; Willingham, M. C., Immunohistochemical localization in normal tissues of different epitopes in the mul-tidrug transport protein P170: evidence for localization in brain capillaries and crossre-activity of one antibody with a muscle protein. J Histochem Cytochem 1989, 37, (2), 159-164.
86. Fojo, A. T.; Ueda, K.; Slamon, D. J.; Poplack, D. G.; Gottesman, M. M.; Pastan, I., Expression of a multidrug-resistance gene in human tumors and tissues. Proc Natl Acad Sci U S A 1987, 84, (1), 265-269.
87. Shapiro, A. B.; Corder, A. B.; Ling, V., P-glycoprotein-mediated Hoechst 33342 trans-port out of the lipid bilayer. Eur J Biochem 1997, 250, (1), 115-121.
88. Homolya, L.; Hollo, Z.; Germann, U. A.; Pastan, I.; Gottesman, M. M.; Sarkadi, B., Fluorescent cellular indicators are extruded by the multidrug resistance protein. J Biol Chem 1993, 268, (29), 21493-21496.
89. Gatlik-Landwojtowicz, E.; Äänismaa, P.; Seelig, A., Quantification and characterization of P-glycoprotein-substrate interactions. Biochemistry 2006, 45, (9), 3020-3032.
90. Omote, H.; Al-Shawi, M. K., Interaction of transported drugs with the lipid bilayer and P-glycoprotein through a solvation exchange mechanism. Biophys J 2006, 90, (11), 4046-4059.
91. Cole, S. P. C.; Bhardwaj, G.; Gerlach, J. H.; Mackie, J. E.; Grant, C. E.; Almquist, K. C.; Stewart, A. J.; Kurz, E. U., et al., Overexpression of a transporter gene in a mul-tidrug-resistant human lung cancer cell line. Science 1992, 258, (5088), 1650-1654.
92. Borst, P.; Evers, R.; Kool, M.; Wijnholds, J., A family of drug transporters: the mul-tidrug resistance-associated proteins. J Natl Cancer Institute 2000, 92, (16), 1295-302.
93. Evers, R.; Zaman, G. J. R.; Van Deemter, L.; Jansen, H.; Calafat, J.; Oomen, L. C. J. M.; Oude Elferink, R. P. J.; Borst, P., et al., Basolateral localization and export activity of the human multidrug resistance - associated protein in polarized pig kidney cells. J Clin Invest 1996, 97, (5), 1211-1218.
94. Loe, D. W.; Deeley, R. G.; Cole, S. P. C., Characterization of vincristine transport by the M(r) 190,000 multidrug resistance protein (MRP): Evidence for cotransport with re-duced glutathione. Cancer Res 1998, 58, (22), 5130-5136.

60
95. Rappa, G.; Lorico, A.; Flavell, R. A.; Sartorelli, A. C., Evidence that the multidrug resistance protein (MRP) functions as a co- transporter of glutathione and natural prod-uct toxins. Cancer Res 1997, 57, (23), 5232-5237.
96. Mayer, R.; Kartenbeck, J.; Büchler, M.; Jedlitschky, G.; Leier, I.; Keppler, D., Expres-sion of the MRP gene-encoded conjugate export pump in liver and its selective absence from the canalicular membrane in transport-deficient mutant hepatocytes. J Cell Biol 1995, 131, (1), 137-150.
97. Paulusma, C. C.; Bosma, P. J.; Zaman, G. J. R.; Bakker, C. T. M.; Otter, M.; Scheffer, G. L.; Scheper, R. J.; Borst, P., et al., Congenital jaundice in rats with a mutation in a multidrug resistance-associated protein gene. Science 1996, 271, (5252), 1126-1128.
98. Keppler, D.; König, J., Expression and localization of the conjugate export pump en-coded by the MRP2 (cMRP/cMOAT) gene in liver. FASEB J 1997, 11, (7), 509-516.
99. Keitel, V.; Nies, A. T.; Brom, M.; Hummel-Eisenbeiss, J.; Spring, H.; Keppler, D., A common Dubin-Johnson syndrome mutation impairs protein maturation and transport activity of MRP2 (ABCC2). Am J Physiol Gastrointest Liver Physiol 2003, 284, (1), G165-G174.
100. Evers, R.; de Haas, M.; Sparidans, R.; Beijnen, J.; Wielinga, P. R.; Lankelma, J.; Borst, P., Vinblastine and sulfinpyrazone export by the multidrug resistance protein MRP2 is associated with glutathione export. Br J Cancer 2000, 83, (3), 375-383.
101. Bakos, E.; Evers, R.; Sinko, E.; Varadi, A.; Borst, P.; Sarkadi, B., Interactions of the human multidrug resistance proteins MRP1 and MRP2 with organic anions. Mol Phar-macol 2000, 57, (4), 760-768.
102. Bodo, A.; Bakos, E.; Szeri, F.; Varadi, A.; Sarkadi, B., Differential modulation of the human liver conjugate transporters MRP2 and MRP3 by bile acids and organic anions. J Biol Chem 2003, 278, (26), 23529-23537.
103. Borst, P.; Zelcer, N.; van de Wetering, K.; Poolman, B., On the putative co-transport of drugs by multidrug resistance proteins. FEBS Lett 2006, 580, (4), 1085-1093.
104. Donner, M. G.; Keppler, D., Up-regulation of basolateral multidrug resistance protein 3 (Mrp3) in cholestatic rat liver. Hepatology 2001, 34, (2), 351-359.
105. Beck, K.; Hayashi, K.; Dang, K.; Hayashi, M.; Boyd, C. D., Analysis of ABCC6 (MRP6) in normal human tissues. Histochem Cell Biol 2005, 123, (4-5), 517-528.
106. Nies, A. T.; Jedlitschky, G.; König, J.; Herold-Mende, C.; Steiner, H. H.; Schmitt, H. P.; Keppler, D., Expression and immunolocalization of the multidrug resistance proteins, MRP1-MRP6 (ABCC1-ABCC6), in human brain. Neuroscience 2004, 129, (2), 349-360.
107. Letourneau, I. J.; Slot, A. J.; Deeley, R. G.; Cole, S. P., Mutational Analysis of a Highly Conserved Proline Residue in MRP1, MRP2, and MRP3 Reveals a Partially Conserved Function. Drug Metab Dispos 2007, 35, (8), 1372-1379.
108. Haimeur, A.; Conseil, G.; Deeley, R. G.; Cole, S. P., The MRP-related and BCRP/ABCG2 multidrug resistance proteins: biology, substrate specificity and regula-tion. Curr Drug Metab 2004, 5, (1), 21-53.
109. Schuetz, J. D.; Connelly, M. C.; Sun, D.; Paibir, S. G.; Flynn, P. M.; Srinivas, R. V.; Kumar, A.; Fridland, A., MRP4: A previously unidentified factor in resistance to nu-cleoside-based antiviral drugs. Nat Med 1999, 5, (9), 1048-1051.
110. Wijnholds, J.; Mol, C. A. A. M.; Van Deemter, L.; De Haas, M.; Scheffer, G. L.; Baas, F.; Beijnen, J. H.; Scheper, R. J., et al., Multidrug-resistance protein 5 is a multispecific

61
organic anion transporter able to transport nucleotide analogs. Proc Natl Acad Sci U S A 2000, 97, (13), 7476-7481.
111. Doyle, L. A.; Yang, W.; Abruzzo, L. V.; Krogmann, T.; Gao, Y.; Rishi, A. K.; Ross, D. D., A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc Natl Acad Sci U S A 1998, 95, (26), 15665-15670.
112. Maliepaard, M.; Scheffer, G. L.; Faneyte, I. F.; van Gastelen, M. A.; Pijnenborg, A. C.; Schinkel, A. H.; van De Vijver, M. J.; Scheper, R. J., et al., Subcellular localization and distribution of the breast cancer resistance protein transporter in normal human tissues. Cancer Res 2001, 61, (8), 3458-3464.
113. Jonker, J. W.; Merino, G.; Musters, S.; van Herwaarden, A. E.; Bolscher, E.; Wagenaar, E.; Mesman, E.; Dale, T. C., et al., The breast cancer resistance protein BCRP (ABCG2) concentrates drugs and carcinogenic xenotoxins into milk. Nat Med 2005, 11, (2), 127-129.
114. Cooray, H. C.; Blackmore, C. G.; Maskell, L.; Barrand, M. A., Localisation of breast cancer resistance protein in microvessel endothelium of human brain. Neuroreport 2002, 13, (16), 2059-2063.
115. Rocchi, E.; Khodjakov, A.; Volk, E. L.; Yang, C. H.; Litman, T.; Bates, S. E.; Schnei-der, E., The product of the ABC half-transporter gene ABCG2 (BCRP/MXR/ABCP) is expressed in the plasma membrane. Biochem Biophys Res Commun 2000, 271, (1), 42-46.
116. Krishnamurthy, P.; Schuetz, J. D., Role of ABCG2/BCRP in biology and medicine. Annu Rev Pharmacol Toxicol 2006, 46, 381-410.
117. Gupta, A.; Zhang, Y.; Unadkat, J. D.; Mao, Q., HIV protease inhibitors are inhibitors but not substrates of the human breast cancer resistance protein (BCRP/ABCG2). J Pharmacol Exp Ther 2004, 310, (1), 334-341.
118. Zhang, Y.; Gupta, A.; Wang, H.; Zhou, L.; Vethanayagam, R. R.; Unadkat, J. D.; Mao, Q., BCRP transports dipyridamole and is inhibited by calcium channel blockers. Pharm Res 2005, 22, (12), 2023-2034.
119. Volk, E. L.; Rohde, K.; Rhee, M.; McGuire, J. J.; Doyle, L. A.; Ross, D. D.; Schneider, E., Methotrexate cross-resistance in a mitoxantrone-selected multidrug-resistant MCF7 breast cancer cell line is attributable to enhanced energy-dependent drug efflux. Cancer Res 2000, 60, (13), 3514-3521.
120. Imai, Y.; Asada, S.; Tsukahara, S.; Ishikawa, E.; Tsuruo, T.; Sugimoto, Y., Breast can-cer resistance protein exports sulfated estrogens but not free estrogens. Mol Pharmacol 2003, 64, (3), 610-618.
121. Maliepaard, M.; van Gastelen, M. A.; de Jong, L. A.; Pluim, D.; van Waardenburg, R. C.; Ruevekamp-Helmers, M. C.; Floot, B. G.; Schellens, J. H., Overexpression of the BCRP/MXR/ABCP gene in a topotecan-selected ovarian tumor cell line. Cancer Res 1999, 59, (18), 4559-4563.
122. Kruijtzer, C. M.; Beijnen, J. H.; Rosing, H.; ten Bokkel Huinink, W. W.; Schot, M.; Jewell, R. C.; Paul, E. M.; Schellens, J. H., Increased oral bioavailability of topotecan in combination with the breast cancer resistance protein and P-glycoprotein inhibitor GF120918. J Clin Oncol 2002, 20, (13), 2943-2950.
123. Wassvik, C. M. Computational Analysis of Aqueous Drug Solubility – Influence of the Solid State. http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-7334. Uppsala University, Sweden, 2006.

62
124. Kitchen, D. B.; Stahura, F. L.; Bajorath, J., Computational techniques for diversity analysis and compound classification. Mini Rev Med Chem 2004, 4, (10), 1029-1039.
125. Martin, Y. C.; Bures, M. G.; Danaher, E. A.; DeLazzer, J.; Lico, I.; Pavlik, P. A., A fast new approach to pharmacophore mapping and its application to dopaminergic and ben-zodiazepine agonists. J Comput Aided Mol Des 1993, 7, (1), 83-102.
126. Jones, G.; Willett, P.; Glen, R. C., A genetic algorithm for flexible molecular overlay and pharmacophore elucidation. J Comput Aided Mol Des 1995, 9, (6), 532-549.
127. van Drie, J. H., Pharmacophore discovery--lessons learned. Curr Pharm Des 2003, 9, (20), 1649-1664.
128. Evans, D. A.; Doman, T. N.; Thorner, D. A.; Bodkin, M. J., 3D QSAR methods: Phase and Catalyst compared. J Chem Inf Model 2007, 47, (3), 1248-1257.
129. Clement, O. O.; Mehl, A. T., HipHop: pharmacophore based on multiple common feature alignments. In Pharmacophore Perception, Development and Use in Drug De-sign, Guner, O. F., Ed. IUL: San Diego, 2000; pp 69-84.
130. Li, H.; Sutter, J. M.; Hoffmann, R., HypoGen: an automated system for generating 3D predictive pharmacophore models. In Pharmacophore Perception, Development and Use in Drug Design, Guner, O. F., Ed. IUL: San Diego, 2000; pp 173-189.
131. Dixon, S. L.; Smondyrev, A. M.; Rao, S. N., PHASE: a novel approach to pharma-cophore modeling and 3D database searching. Chem Biol Drug Des 2006, 67, (5), 370-372.
132. Cramer, R. D., Comparative Molecular-Field Analysis (Comfa). 1. Effect of Shape on Binding of Steroids to Carrier Proteins. J Am Chem Soc 1988, 110, (18), 5959-5967.
133. Cruciani, G.; Watson, K. A., Comparative Molecular-Field Analysis Using Grid Force-Field and Golpe Variable Selection Methods in a Study of Inhibitors of Glycogen-Phosphorylase-B. J Med Chem 1994, 37, (16), 2589-2601.
134. Legendre, A. M., Sur la Méthode des moindres quarrés. In Nouvelles méthodes pour la détermination des orbites des comètes., Paris, 1805.
135. Gauss, C. F., Theoria motus corporum coelestium in sectionibus conicis solem ambien-tum. Göttingen, 1809.
136. Hastie, T.; Tibshirani, R.; Friedman, R., The elements of statistical learning. Data min-ing, inference, and prediction. Springer: New York, 2001.
137. Wold, S., PLS-regression: a basic tool of chemometrics. Chemometr Intell Lab 2001, 58, (2), 109-130.
138. Berglund, A.; Kettaneh, N.; Uppgard, L. L.; Wold, S.; Bendwell, N.; Cameron, D. R., The GIFI approach to non-linear PLS modeling. J Chemometr 2001, 15, (4), 321-336.
139. Eriksson, L.; Johansson, E.; Lindgren, F.; Wold, S., GIFI-PLS: Modeling of non-linearities and discontinuities in QSAR. Quant Struct-Act Rel 2000, 19, (4), 345-355.
140. Trygg, J., Orthogonal projections to latent structures (O-PLS). J Chemometr 2002, 16, (3), 119-128.
141. McCulloch, W. S., A Logical Calculus of the Ideas Immanent in Nervous Activity (Reprinted from Bulletin of Mathematical Biophysics, Vol 5, Pg 115-133, 1943). Bull Math Biol 1990, 52, (1-2), 99-115.
142. Cheng, B., Neural Networks - a Review from a Statistical Perspective. Stat Sci 1994, 9, (1), 2-30.
143. Gasteiger, J., Neural Networks in Chemistry. Angew Chem Int Ed Engl 1993, 32, (4), 503-527.

63
144. Bai, J. P.; Utis, A.; Crippen, G.; He, H. D.; Fischer, V.; Tullman, R.; Yin, H. Q.; Hsu, C. P., et al., Use of classification regression tree in predicting oral absorption in humans. J Chem Inf Comput Sci 2004, 44, (6), 2061-2069.
145. Balakin, K. V.; Ivanenkov, Y. A.; Savchuk, N. P.; Ivashchenko, A. A.; Ekins, S., Com-prehensive computational assessment of ADME properties using mapping techniques. Curr Drug Discov Technol 2005, 2, (2), 99-113.
146. Linnankoski, J.; Makela, J. M.; Ranta, V. P.; Urtti, A.; Yliperttula, M., Computational prediction of oral drug absorption based on absorption rate constants in humans. J Med Chem 2006, 49, (12), 3674-3681.
147. Bergström, C. A. S., Computational models to predict aqueous drug solubility, perme-ability and intestinal absorption. Expert Opin Drug Metab Toxicol 2005, 1, (4), 613-627.
148. Jung, E.; Kim, J.; Kim, M.; Jung, D. H.; Rhee, H.; Shin, J. M.; Choi, K.; Kang, S. K., et al., Artificial neural network models for prediction of intestinal permeability of oli-gopeptides. BMC Bioinformatics 2007, 8, 245.
149. Palm, K.; Stenberg, P.; Luthman, K.; Artursson, P., Polar molecular surface properties predict the intestinal absorption of drugs in humans. Pharm Res 1997, 14, (5), 568-571.
150. Raevsky, O. A.; Schaper, K. J., Quantitative estimation of hydrogen bond contribution to permeability and absorption processes of some chemicals and drugs. Eur J Med Chem 1998, 33, (10), 799-807.
151. Wessel, M. D.; Jurs, P. C.; Tolan, J. W.; Muskal, S. M., Prediction of human intestinal absorption of drug compounds from molecular structure. J Chem Inf Comput Sci 1998, 38, (4), 726-735.
152. Egan, W. J.; Merz, K. M., Jr.; Baldwin, J. J., Prediction of drug absorption using multi-variate statistics. J Med Chem 2000, 43, (21), 3867-3877.
153. Zhao, Y. H.; Le, J.; Abraham, M. H.; Hersey, A.; Eddershaw, P. J.; Luscombe, C. N.; Boutina, D.; Beck, G., et al., Evaluation of human intestinal absorption data and subse-quent derivation of a quantitative structure-activity relationship (QSAR) with the Abra-ham descriptors. J Pharm Sci 2001, 90, (6), 749-784.
154. Zhao, Y. H.; Le, J.; Abraham, M. H.; Hersey, A.; Eddershaw, P. J.; Luscombe, C. N.; Butina, D.; Beck, G., et al., Erratum: evaluation of human intestinal absorption data and subsequent derivation of a quantitative structure-activity relationship (QSAR) with the abraham descriptors. J Pharm Sci 2002, 91, (2), 605.
155. Amidon, G. L.; Lennernäs, H.; Shah, V. P.; Crison, J. R., A Theroretical Basis for a Biopharmaceutic Drug Classification: The Correlation of in Vitro Drug Product Disso-lution and in Vivo Bioavailability. Pharm Res 1995, 12, (3), 413-420.
156. Bergström, C. A.; Strafford, M.; Lazorova, L.; Avdeef, A.; Luthman, K.; Artursson, P., Absorption classification of oral drugs based on molecular surface properties. J Med Chem 2003, 46, (4), 558-570.
157. Khandelwal, A.; Bahadduri, P. M.; Chang, C.; Polli, J. E.; Swaan, P. W.; Ekins, S., Computational models to assign biopharmaceutics drug disposition classification from molecular structure. Pharm Res 2007, 24, (12), 2249-2262.
158. Kortejärvi, H.; Urtti, A.; Yliperttula, M., Pharmacokinetic simulation of biowaiver criteria: the effects of gastric emptying, dissolution, absorption and elimination rates. Eur J Pharm Sci 2007, 30, (2), 155-166.
159. Palm, K.; Luthman, K.; Ungell, A. L.; Strandlund, G.; Artursson, P., Correlation of drug absorption with molecular surface properties. J Pharm Sci 1996, 85, (1), 32-39.

64
160. Stewart, B. H.; Chung, F. Y.; Tait, B.; Blankley, C. J.; Chan, O. H.; John, C., Hydro-phobicity of HIV protease inhibitors by immobilized artificial membrane chromatogra-phy: application and significance to drug transport. Pharm Res 1998, 15, (9), 1401-1406.
161. Palm, K.; Luthman, K.; Ungell, A. L.; Strandlund, G.; Beigi, F.; Lundahl, P.; Artursson, P., Evaluation of dynamic polar molecular surface area as predictor of drug absorption: comparison with other computational and experimental predictors. J Med Chem 1998, 41, (27), 5382-5392.
162. Krarup, L. H.; Christensen, I. T.; Hovgaard, L.; Frokjaer, S., Predicting drug absorption from molecular surface properties based on molecular dynamics simulations. Pharm Res 1998, 15, (7), 972-978.
163. Ertl, P.; Rohde, B.; Selzer, P., Fast calculation of molecular polar surface area as a sum of fragment-based contributions and its application to the prediction of drug transport properties. J Med Chem 2000, 43, (20), 3714-3717.
164. Norinder, U.; Österberg, T.; Artursson, P., Theoretical calculation and prediction of Caco-2 cell permeability using MolSurf parametrization and PLS statistics. Pharm Res 1997, 14, (12), 1786-1791.
165. Bravi, G.; Wikel, J. H., Application of MS-WHIM descriptors: 3. Prediction of molecu-lar properties. Quant Struct-Act Rel 2000, 19, (1), 39-49.
166. Stenberg, P.; Norinder, U.; Luthman, K.; Artursson, P., Experimental and computational screening models for the prediction of intestinal drug absorption. J Med Chem 2001, 44, (12), 1927-1937.
167. Ekins, S.; Durst, G. L.; Stratford, R. E.; Thorner, D. A.; Lewis, R.; Loncharich, R. J.; Wikel, J. H., Three-dimensional quantitative structure-permeability relationship analysis for a series of inhibitors of rhinovirus replication. J Chem Inf Comput Sci 2001, 41, (6), 1578-1586.
168. Klon, A. E.; Lowrie, J. F.; Diller, D. J., Improved naive Bayesian modeling of numeri-cal data for absorption, distribution, metabolism and excretion (ADME) property pre-diction. J Chem Inf Model 2006, 46, (5), 1945-1956.
169. Garg, P.; Verma, J., In silico prediction of blood brain barrier permeability: an Artificial Neural Network model. J Chem Inf Model 2006, 46, (1), 289-297.
170. Egan, W. J.; Lauri, G., Prediction of intestinal permeability. Adv Drug Deliv Rev 2002, 54, (3), 273-289.
171. Oprea, T. I.; Gottfries, J., Toward minimalistic modeling of oral drug absorption. J Mol Graph Model 1999, 17, (5-6), 261-274.
172. Andrews, C. W.; Bennett, L.; Yu, L. X., Predicting human oral bioavailability of a compound: development of a novel quantitative structure-bioavailability relationship. Pharm Res 2000, 17, (6), 639-644.
173. Winiwarter, S.; Ax, F.; Lennernäs, H.; Hallberg, A.; Pettersson, C.; Karlén, A., Hydro-gen bonding descriptors in the prediction of human in vivo intestinal permeability. J Mol Graph Model 2003, 21, (4), 273-287.
174. Norinder, U.; Bergström, C. A., Prediction of ADMET Properties. ChemMedChem 2006, 1, (9), 920-937.
175. Chang, C.; Swaan, P. W., Computational approaches to modeling drug transporters. Eur J Pharm Sci 2006, 27, (5), 411-424.
176. Zhang, E. Y.; Phelps, M. A.; Cheng, C.; Ekins, S.; Swaan, P. W., Modeling of active transport systems. Adv Drug Deliv Rev 2002, 54, (3), 329-354.

65
177. Chang, C.; Ekins, S.; Bahadduri, P.; Swaan, P. W., Pharmacophore-based discovery of ligands for drug transporters. Adv Drug Deliv Rev 2006, 58, (12-13), 1431-1450.
178. Chang, G., Retraction of "Structure of MsbA from Vibrio cholera: a multidrug resis-tance ABC transporter homolog in a closed conformation" [J. Mol. Biol. (2003) 330 419-430]. J Mol Biol 2007, 369, (2), 596.
179. Zamora, J. M.; Pearce, H. L.; Beck, W. T., Physical-chemical properties shared by compounds that modulate multidrug resistance in human leukemic cells. Mol Pharma-col 1988, 33, (4), 454-462.
180. Österberg, T.; Norinder, U., Theoretical calculation and prediction of P-glycoprotein-interacting drugs using MolSurf parametrization and PLS statistics. Eur J Pharm Sci 2000, 10, (4), 295-303.
181. Suhre, W. M.; Ekins, S.; Chang, C.; Swaan, P. W.; Wright, S. H., Molecular determi-nants of substrate/inhibitor binding to the human and rabbit renal organic cation trans-porters hOCT2 and rbOCT2. Mol Pharmacol 2005, 67, (4), 1067-1077.
182. Bednarczyk, D.; Ekins, S.; Wikel, J. H.; Wright, S. H., Influence of molecular structure on substrate binding to the human organic cation transporter, hOCT1. Mol Pharmacol 2003, 63, (3), 489-498.
183. Ekins, S.; Kim, R. B.; Leake, B. F.; Dantzig, A. H.; Schuetz, E. G.; Lan, L. B.; Yasuda, K.; Shepard, R. L., et al., Application of three-dimensional quantitative structure-activity relationships of P-glycoprotein inhibitors and substrates. Mol Pharmacol 2002, 61, (5), 974-81.
184. Pajeva, I. K.; Wiese, M., Pharmacophore model of drugs involved in P-glycoprotein multidrug resistance: explanation of structural variety (hypothesis). J Med Chem 2002, 45, (26), 5671-5686.
185. Yates, C. R.; Chang, C.; Kearbey, J. D.; Yasuda, K.; Schuetz, E. G.; Miller, D. D.; Dalton, J. T.; Swaan, P. W., Structural determinants of P-glycoprotein-mediated trans-port of glucocorticoids. Pharm Res 2003, 20, (11), 1794-1803.
186. Langer, T.; Eder, M.; Hoffmann, R. D.; Chiba, P.; Ecker, G. F., Lead identification for modulators of multidrug resistance based on in silico screening with a pharmacophoric feature model. Arch Pharm (Weinheim) 2004, 337, (6), 317-327.
187. Wang, Y. H.; Li, Y.; Yang, S. L.; Yang, L., An in silico approach for screening flavon-oids as P-glycoprotein inhibitors based on a Bayesian-regularized neural network. J Comput Aided Mol Des 2005, 19, (3), 137-147.
188. Baringhaus, K. H.; Matter, H.; Stengelin, S.; Kramer, W., Substrate specificity of the ileal and the hepatic Na(+)/bile acid cotransporters of the rabbit. II. A reliable 3D QSAR pharmacophore model for the ileal Na(+)/bile acid cotransporter. J Lipid Res 1999, 40, (12), 2158-2168.
189. Zhuang, X. M.; Xiao, J. H.; Li, J. T.; Zhang, Z. Q.; Ruan, J. X., A simplified model to predict P-glycoprotein interacting drugs from 3D molecular interaction field. Int J Pharm 2006, 309, (1-2), 109-114.
190. Chang, C.; Bahadduri, P. M.; Polli, J. E.; Swaan, P. W.; Ekins, S., Rapid identification of P-glycoprotein substrates and inhibitors. Drug Metab Dispos 2006, 34, (12), 1976-1984.
191. Chang, C.; Pang, K. S.; Swaan, P. W.; Ekins, S., Comparative pharmacophore modeling of organic anion transporting polypeptides: a meta-analysis of rat Oatp1a1 and human OATP1B1. J Pharmacol Exp Ther 2005, 314, (2), 533-541.

66
192. Cianchetta, G.; Singleton, R. W.; Zhang, M.; Wildgoose, M.; Giesing, D.; Fravolini, A.; Cruciani, G.; Vaz, R. J., A pharmacophore hypothesis for P-glycoprotein substrate rec-ognition using GRIND-based 3D-QSAR. J Med Chem 2005, 48, (8), 2927-2935.
193. Hirono, S.; Nakagome, I.; Imai, R.; Maeda, K.; Kusuhara, H.; Sugiyama, Y., Estimation of the three-dimensional pharmacophore of ligands for rat multidrug-resistance-associated protein 2 using ligand-based drug design techniques. Pharm Res 2005, 22, (2), 260-269.
194. Yarim, M.; Moro, S.; Huber, R.; Meier, P. J.; Kaseda, C.; Kashima, T.; Hagenbuch, B.; Folkers, G., Application of QSAR analysis to organic anion transporting polypeptide 1a5 (Oatp1a5) substrates. Bioorg Med Chem 2005, 13, (2), 463-471.
195. Biegel, A.; Gebauer, S.; Hartrodt, B.; Brandsch, M.; Neubert, K.; Thondorf, I., Three-Dimensional Quantitative Structure-Activity Relationship Analyses of Beta-Lactam An-tibiotics and Tripeptides as Substrates of the Mammalian H+/Peptide Cotransporter PEPT1. J Med Chem 2005, 48, (13), 4410-4419.
196. Gombar, V. K.; Polli, J. W.; Humphreys, J. E.; Wring, S. A.; Serabjit-Singh, C. S., Predicting P-glycoprotein substrates by a quantitative structure-activity relationship model. J Pharm Sci 2004, 93, (4), 957-968.
197. Crivori, P.; Reinach, B.; Pezzetta, D.; Poggesi, I., Computational models for identifying potential P-glycoprotein substrates and inhibitors. Mol Pharm 2006, 3, (1), 33-44.
198. Huang, J.; Ma, G.; Muhammad, I.; Cheng, Y., Identifying P-glycoprotein substrates using a support vector machine optimized by a particle swarm. J Chem Inf Model 2007, 47, (4), 1638-1647.
199. Xue, Y.; Yap, C. W.; Sun, L. Z.; Cao, Z. W.; Wang, J. F.; Chen, Y. Z., Prediction of P-glycoprotein substrates by a support vector machine approach. J Chem Inf Comput Sci 2004, 44, (4), 1497-1505.
200. Penzotti, J. E.; Lamb, M. L.; Evensen, E.; Grootenhuis, P. D., A computational ensem-ble pharmacophore model for identifying substrates of P-glycoprotein. J Med Chem 2002, 45, (9), 1737-1740.
201. Olsson, T.; Sherbuhkin, V. Synthesis and Structure Administration, AstraZeneca R&D, Mölndal, Sweden.
202. Oprea, T. I.; Gottfries, J., Chemography: the art of navigating in chemical space. J Comb Chem 2001, 3, (2), 157-166.
203. Physicians' Desk Reference 61 ed.; Medical Economics Data: Montvale, New Jersey, 2007.
204. Artursson, P.; Karlsson, J., Correlation between oral drug absorption in humans and apparent drug permeability coefficients in human intestinal epithelial (Caco-2) cells. Biochem Biophys Res Commun 1991, 175, (3), 880-885.
205. Tavelin, S.; Gråsjö, J.; Taipalensuu, J.; Ocklind, G.; Artursson, P., Applications of epithelial cell culture in studies of drug transport. Methods Mol Biol 2002, 188, 233-272.
206. Ishikawa, T.; Muller, M.; Klunemann, C.; Schaub, T.; Keppler, D., ATP-dependent primary active transport of cysteinyl leukotrienes across liver canalicular membrane. Role of the ATP-dependent transport system for glutathione S-conjugates. J Biol Chem 1990, 265, (31), 19279-19286.
207. MacroModel, Version 6.5 (Paper I) or 9 (Paper II); Schrödinger: San Diego, CA. 208. SciFinder Scholar, 2006; American Chemical Society: Washington DC. 209. Corina, Version 3.0; Molecular Networks: Erlangen, Germany.

67
210. Molconn-Z, Version 3.15s; Hall Associates Consulting: Quincy, MA. 211. DragonX, Version 3; Talete: Milan, Italy. 212. ADMETPredictor, Version 1.2.4; SimulationsPlus: Lancaster, CA. 213. MOLPRO-2001, TimTec: Newark, DE. 214. MAREA, Version 2.4 (Paper I) or 3.02 (Papers II-IV); The program MAREA is avail-
able from the authors upon request. It is provided free of charge for academic users. Contact Christel Bergström (email: [email protected]).
215. Catalyst, Version 4.9; Accelrys: San Diego, CA. 216. Simca-P, Version 10 (Paper I) or 11.5 (Papers II-IV); Umetrics: Umeå, Sweden. 217. Yazdanian, M.; Glynn, S. L.; Wright, J. L.; Hawi, A., Correlating partitioning and
Caco-2 cell permeability of structurally diverse small molecular weight compounds. Pharm Res 1998, 15, (9), 1490-1494.
218. Sun, D.; Lennernäs, H.; Welage, L. S.; Barnett, J. L.; Landowski, C. P.; Foster, D.; Fleisher, D.; Lee, K. D., et al., Comparison of human duodenum and Caco-2 gene ex-pression profiles for 12,000 gene sequences tags and correlation with permeability of 26 drugs. Pharm Res 2002, 19, (10), 1400-1416.
219. Winiwarter, S.; Bonham, N. M.; Ax, F.; Hallberg, A.; Lennernas, H.; Karlen, A., Corre-lation of human jejunal permeability (in vivo) of drugs with experimentally and theo-retically derived parameters. A multivariate data analysis approach. J Med Chem 1998, 41, (25), 4939-4949.
220. Tavelin, S.; Taipalensuu, J.; Söderberg, L.; Morrison, R.; Chong, S.; Artursson, P., Prediction of the oral absorption of low-permeability drugs using small intestine-like 2/4/A1 cell monolayers. Pharm Res 2003, 20, (3), 397-405.
221. Terada, T.; Sawada, K.; Saito, H.; Hashimoto, Y.; Inui, K., Functional characteristics of basolateral peptide transporter in the human intestinal cell line Caco-2. Am J Physiol 1999, 276, (6), G1435-G1441.
222. Chiou, W. L.; Chung, S. M.; Wu, T. C.; Ma, C., A comprehensive account on the role of efflux transporters in the gastrointestinal absorption of 13 commonly used substrate drugs in humans. Int J Clin Pharmacol Ther 2001, 39, (3), 93-101.
223. Goldman, I. D.; Matherly, L. H., The cellular pharmacology of methotrexate. Pharma-col Ther 1985, 28, (1), 77-102.
224. Westphal, K.; Weinbrenner, A.; Giessmann, T.; Stuhr, M.; Franke, G.; Zschiesche, M.; Oertel, R.; Terhaag, B., et al., Oral bioavailability of digoxin is enhanced by talinolol: evidence for involvement of intestinal P-glycoprotein. Clin Pharmacol Ther 2000, 68, (1), 6-12.
225. Dietrich, C. G.; Geier, A.; Oude Elferink, R. P., ABC of oral bioavailability: transport-ers as gatekeepers in the gut. Gut 2003, 52, (12), 1788-1795.
226. Hilgendorf, C.; Ahlin, G.; Seithel, A.; Artursson, P.; Ungell, A. L.; Karlsson, J., Expres-sion of Thirty-six Drug Transporter Genes in Human Intestine, Liver, Kidney, and Or-ganotypic Cell Lines. Drug Metab Dispos 2007, 35, (8), 1333-1340.
227. Harris, M. J.; Kuwano, M.; Webb, M.; Board, P. G., Identification of the apical mem-brane-targeting signal of the multidrug resistance-associated protein 2 (MRP2/MOAT). J Biol Chem 2001, 276, (24), 20876-20881.
228. Smitherman, P. K.; Townsend, A. J.; Kute, T. E.; Morrow, C. S., Role of multidrug resistance protein 2 (MRP2, ABCC2) in alkylating agent detoxification: MRP2 potenti-ates glutathione S-transferase A1-1-mediated resistance to chlorambucil cytotoxicity. J Pharmacol Exp Ther 2004, 308, (1), 260-267.

68
229. Tang, W., Drug metabolite profiling and elucidation of drug-induced hepatotoxicity. Expert Opin Drug Metab Toxicol 2007, 3, (3), 407-420.
230. Breedveld, P.; Pluim, D.; Cipriani, G.; Wielinga, P.; van Tellingen, O.; Schinkel, A. H.; Schellens, J. H., The effect of Bcrp1 (Abcg2) on the in vivo pharmacokinetics and brain penetration of imatinib mesylate (Gleevec): implications for the use of breast cancer re-sistance protein and P-glycoprotein inhibitors to enable the brain penetration of imatinib in patients. Cancer Res 2005, 65, (7), 2577-2582.
231. Janneh, O.; Owen, A.; Chandler, B.; Hartkoorn, R. C.; Hart, C. A.; Bray, P. G.; Ward, S. A.; Back, D. J., et al., Modulation of the intracellular accumulation of saquinavir in peripheral blood mononuclear cells by inhibitors of MRP1, MRP2, P-gp and BCRP. Aids 2005, 19, (18), 2097-2102.
232. Tian, X.; Li, J.; Zamek-Gliszczynski, M. J.; Bridges, A. S.; Zhang, P.; Patel, N. J.; Raub, T. J.; Pollack, G. M., et al., Roles of P-glycoprotein, Bcrp, and Mrp2 in biliary excretion of spiramycin in mice. Antimicrob Agents Chemother 2007, 51, (9), 3230-3234.
233. Su, Y.; Lee, S. H.; Sinko, P. J., Inhibition of efflux transporter ABCG2/BCRP does not restore mitoxantrone sensitivity in irinotecan-selected human leukemia CPT-K5 cells: evidence for multifactorial multidrug resistance. Eur J Pharm Sci 2006, 29, (2), 102-110.
234. Huang, L.; Wang, Y.; Grimm, S., ATP-dependent transport of rosuvastatin in mem-brane vesicles expressing breast cancer resistance protein. Drug Metab Dispos 2006, 34, (5), 738-742.
235. Ozawa, N.; Shimizu, T.; Morita, R.; Yokono, Y.; Ochiai, T.; Munesada, K.; Ohashi, A.; Aida, Y., et al., Transporter database, TP-Search: a web-accessible comprehensive data-base for research in pharmacokinetics of drugs. Pharm Res 2004, 21, (11), 2133-2134.


Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Pharmacy 67
Editor: The Dean of the Faculty of Pharmacy
A doctoral dissertation from the Faculty of Pharmacy, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofPharmacy. (Prior to January, 2005, the series was publishedunder the title “Comprehensive Summaries of UppsalaDissertations from the Faculty of Pharmacy”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-8371
ACTA
UNIVERSITATIS
UPSALIENSIS
UPPSALA
2007










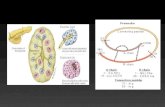
![COMMENTARY Antibiotic Efflux Pumps · the drug efflux pumps in eucaryotic cells ( [7]; drug efflux transporters are classically energized by ATP). The second-ary active transporters,](https://static.fdocuments.in/doc/165x107/6132c0d4dfd10f4dd73aa6ef/commentary-antibiotic-efflux-pumps-the-drug-efflux-pumps-in-eucaryotic-cells-7.jpg)






