
Astudy ofthe intracellular signalling events involved ...
221
A study of the intracellular signalling events involved in the zona pellucida- induced acrosome reaction in human spermatozoa by SIMON STEPHANUS DU PLESSIS Dissertation presented for the degree of Doctor of Philosophy at the Faculty of Health Sciences, University of Stellenbosch Promotors: Prof DR Franken and Dr C Page December 2002
Transcript of Astudy ofthe intracellular signalling events involved ...

A study of the intracellular signallingevents involved in the zona pellucida-induced acrosome reaction in human
spermatozoa
by
SIMON STEPHANUS DU PLESSIS
Dissertation presented for the degree of
Doctor of Philosophy
at the Faculty of Health Sciences, University of Stellenbosch
Promotors:
Prof DR Franken and Dr C Page
December 2002

ii
DECLARA TION
I, the undersigned, hereby declare that the work in this dissertation is my own original
work and that I have not previously in its entirety or in part submitted it at any
university for a degree.
Dal
Stellenbosch University http://scholar.sun.ac.za

iii
ABSTRACT
In this study the author presents new data that will shed light on the fairly nebulous
knowledge of intracellular pathways involved in the physiologically induced acrosome
reaction and the subsequent events leading to fertilization. The zona pellucida-
induced acrosome reaction, sperm-zona interaction as well as various sperm motion
characteristics were investigated.
The first part of the study focussed on the role of extracellular signal regulated kinase
(ERK), a member of the family of mitogen activated protein kinases, during the zona
pellucida-induced acrosome reaction and sperm-oocyte interaction. It was shown that
the inhibition of ERK significantly reduced the zona pellucida-induced acrosome
reaction as measured by fluorescent microscopy. This suggests that ERKs are
directly or indirectly involved in the signal transduction pathway through which the
human sperm acrosome reaction is induced by the zona pellucida. In an attempt to
provide further proof that ERK was involved in human acrosome signalling hemizona
assays were employed to test sperm-oocyte binding. From these sperm-oocyte-
binding experiments it was clear that the inhibition of ERK leads to increased binding.
These results support the idea that the zona pellucida-induced acrosome reaction,
and therefore the physiologically relevant exocytotic event, is regulated by an ERK-
mediated signal transduction process.
In the second part of the study the significance of phosphatidylinositol 3-kinase
(PI3K) in the process of human sperm motility, acrosome reaction and sperm-oocyte
binding, was investigated by employing the specific PI3K, LY294002. PI3K inhibition
increased the percentage motility and percentage progressive motility in
Stellenbosch University http://scholar.sun.ac.za

iv
asthenozoospermia patients. Under the present laboratory conditions PI3K inhibition
furthermore did not influence the acrosome reaction, whilst it enhanced sperm-oocyte
binding. These results therefore imply that PI3K negatively affect sperm motility and
zona-binding.
In the last part of the study the possible effects of intracellular cGMP accumulation
via acute in vivo sildenafil citrate (ViagraTM) administration was investigated on
seminal parameters, induction of the acrosome reaction, sperm-oocyte binding and
sperm motility. As was expected no changes in the macro- and microscopically
seminal parameters were caused by sildenafil citrate when compared to placebo.
Furthermore the acrosome reaction was also not initiated or potentiated by sildenafil
citrate at concentrations of 50mg orally. Sperm-oocyte binding, smooth path velocity,
straight line velocity and the percentage rapid cells all increased after sildenafil citrate
treatment.
From these results it appear that there are various role players in the zona pellucida-
induced acrosome reaction intracellular signalling system. A thorough understanding
of such signal transduction systems and the crosstalk in-between will ultimately yield
information regarding the nature of receptors to which these signal transduction
pathways are coupled in human spermatozoa as well as the intracellular effectors
that ultimately regulate sperm function. Moreover, an understanding of these
regulatory pathways will be essential for the future development of clinical
approaches designed to enhance or preclude fertilization.
Stellenbosch University http://scholar.sun.ac.za

v
OPSOMMING
Die outeur lê in hierdie studie nuwe data voor ten einde meer lig te werp op die
relatief vae veld van intrasellulêre seintransduksie paaie betrokke by die fisiologies-
geïnduseerde akrosoomreaksie en die daaropvolgende gebeure wat aanleiding gee
tot bevrugting. Die zona pellucida-geïnduseerde akrosoomreaksie, sperm-zona
interaksie sowel as spermmotiliteitseienskappe is ondersoek.
Die eerste gedeelte van die studie fokus op die rol van ekstrasellulêre-
seingereguleerde-kinase (ERK), 'n lid van die familie van mitogeen-geaktiveerde
proteïenkinases, tydens die zona pellucida-geïnduseerde akrosoomreaksie en
sperm-oosiet interaksie. Daar word aangetoon dat die inhibisie van ERK die zona
pellucida geïnduseerde akrosoomreaksie, soos gemeet met behulp van fluorosensie
mikroskopie, betekenisvol verminder. Dit suggereer dat ERK direk of indirek betrokke
is by die seintransduksie paaie waardeur die akrosoomreaksie van die menslike
sperm deur die zona pellucida geïnduseer word. In 'n poging om onomwonde te
bewys dat ERK betrokke is by menslike akrosoom-seintransduksie, is hemizona
essais gebruik om sperm-oesiet binding te bepaal. Van hierdie sperm-oosiet binding-
eksperimente is dit duidelik dat die inhibisie van ERK aanleiding gee tot verhoogde
binding. Hierdie resultate ondersteun dus die idee dat die zona pellucida-
geïnduseerde akrosoomreaksie en dus die fisiologies relevante eksositotiese
gebeurtenis gereguleer word deur 'n ERK-gemediëerde seintransduksie proses.
In die tweede gedeelte van die studie is die belang van fosfatidielinositol 3-kinase
(PI3K) in die proses van menslike spermmotiliteit, akrosoomreaksie en sperm-oesiet
binding ondersoek deur van die spesifieke PI3K inhibitor LY294002, gebruik te maak.
Stellenbosch University http://scholar.sun.ac.za

vi
Pl3K-inhibisie het die persentasie motiliteit en progressiewe motiliteit by
astenozoospermiese pasiënte verhoog. Onder hierdie laboratoriumtoestande het
Pl3K-inhibisie nie die akrosoomreaksie beïnvloed nie, terwyl sperm-oosiet binding
verhoog is. Hierdie resultate beteken dus dat PI3K spermmotiliteit en zona-binding
negatief beïnvloed.
In die laaste gedeelte van die studie is die effekte van intrasllulêre cGMP
akkumulasie deur akute in vivo sildenafil sitraat (ViagraTM) toediening op seminale
parameters, induksie van die akrosoomreaksie, sperm-oesiet binding en
spermmotiliteit ondersoek. Soos verwag is geen veranderinge in die makro- en
mikroskopiese seminale parameters deur sildenafil sitraat in vergelyking met plasebo
veroorsaak nie. Verder is die akrosoomreaksies ook nie deur 50mg orale sildenafil
sitraat geïnisieer of potensieer nie. Sperm-oosiet binding, geplaneerde snelheid,
reguitlyn snelheid en persentasie vinnigbewegende selle was almal vehoog na
sildenafil sitraat behandeling.
Uit hierdie resultate blyk dit dat daar verskeie rolspelers in die zona pellucida-
geïnduseerde akrosoomreaksie is. 'n Deeglike insig van al hierdie seintransduksie-
paaie en die onderlinge kruiskontak tussen mekaar sal uiteindelik die nodige inligting
rakende die reseptore waaraan hierdie seintransduksie paaie gekoppel is, verskaf
sowel as die intrasellulêre effektore wat uiteindelik spermfunksie beheer. Nog te
meer sal die begrip van hierdie regulatoriese paaie verder noodsaaklik wees vir die
toekomstige ontwikkeling van kliniese benaderings om bevrugting te bevorder of te
voorkom.
Stellenbosch University http://scholar.sun.ac.za

vii
This dissertation is dedicated to
Wendy and Christopher
Without your tremendous patience, support and love
I would not have been able to
successfully complete
this study.
"The art of love ... is largely the art of persistence."
Stellenbosch University http://scholar.sun.ac.za

viii
ACKNOWLEDGEMENTS
I wish to extend my most sincere gratitude and appreciation to the following people
for their contributions to the successful completion of this study:
Prof Daniel Franken for his guidance, encouragement, support, friendship and many
happy hours spent together in the laboratory;
Dr Carine Page for her guidance and help in the preparation of this manuscript;
My colleagues in the Department of Medical Physiology for encouraging me to
pursue the opportunity of furthering my studies;
The University of Stellenbosch for providing the research facilities;
Schering Aktiengesellschaft and the Harry Crossley Foundation for financial
support in covering research expenses;
Wendy, Christopher, my parents, family and friends for showing a keen interest in
my research and creating a support structure that enabled me to complete this study;
All those instrumental in my growth as a scientist.
Stellenbosch University http://scholar.sun.ac.za

ix
TABLE OF CONTENTS
Declaration
Abstract
Opsomming
Acknowledgements
List of tables
List of figures
Alphabetical list of abbreviations
Page
ii
iii
v
viii
xiv
xv
xvii
CHAPTER 1: INTRODUCTION AND STATEMENT OF PROBLEM
1.1 Introduction
1.2 Objectives and statement of the problem
1.3 Plan of study
1.4 Conclusion
1
2
2
4
CHAPTER 2: LITERATURE REVIEW
2.1 Introduction
2.2 Capacitation
2.2.1 Cholesterol efflux and changes in membrane lipids and
phospholipids during capacitation
2.2.2 Ion fluxes and the regulation of sperm plasma membrane
potential
2.2.2.1 Modification in concentration of intracellular calcium and 17
5
6
12
15
other ions during capacitation
Stellenbosch University http://scholar.sun.ac.za

x
2.2.3 Changes in protein phosphorylation and protein kinase activity 23
during capacitation
2.2.3.1 Involvement of AC/cAMP/PKA pathway in capacitation 24
2.2.3.2 Involvement of PKC in capacitation 25
2.2.3.3 Involvement of tyrosine phosphorylation in capacitation 25
2.2.3.4 Crosstalk between different signalling events during 31
sperm capacitation
2.2.4 Consequences of capacitation on sperm function 33
2.3 The sperm acrosome 35
2.3.1 The acrosome reaction 36
2.3.1.1 Increase in intracellular calcium during the acrosome 43
reaction
2.3.1.2 Phospholipases activation during acrosome reaction 45
2.3.1.3 Involvement of protein kinases in acrosome reaction 46
process
2.4 Motility 49
2.4.1 Factors influencing sperm motility 52
2.4.1.1 Cyclic adenosine mono-phosphate 52
2.4.1.2 Adenylate cyclase 54
2.5 Summary 54
2.6 References 57
CHAPTER 3: MATERIALS AND METHODS
3.1 Introduction
3.2.1 Preparation of human tubal fluid culture medium
87
87
Stellenbosch University http://scholar.sun.ac.za

xi
3.2.2 Semen collection 88
3.2.3 Oocyte collection and storage 88
3.2.4 Solubilized zona pellucida preparation 893.2.5 Assessment of the acrosome reaction 893.2.6 Hemizona binding assay 91
3.2.6.1 Bisecting of oocytes 913.2.6.2 Competitive sperm-binding to the hemizona 92
3.2.7 Computer assisted sperm analyses 943.2.8 Statistical analyses 943.2.9 References 95
CHAPTER 4: THE ZONA PELLUCIDA-INDUCED ACROSOME REACTION
OF HUMAN SPERMATOZOA INVOLVES EXTRACELLULAR SIGNAL-
REGULATED KINASE ACTIVATION
Summary
Introduction
Materials and methods
Results
Discussion
References
CHAPTER 5: EXTRACELLULAR SIGNAL-REGULATED KINASE
ACTIVATION INVOLVED IN HUMAN SPERM-ZONA PELLUCIDA BINDING
Summary
Introduction
Materials and methods
979799102104
109
114114116
Stellenbosch University http://scholar.sun.ac.za

xii
Results
Discussion
References
119
121
124
CHAPTER 6: PHOSPHATIDYLINOSITOL 3-KINASE INHIBITION ENHANCES
HUMAN SPERM MOTILITY AND SPERM-ZONA PELLUCIDA BINDING
Summary 129
Introduction 129
Materials and methods 132
Results 135
Discussion 141
References 148
CHAPTER 7: THE EFFECT OF ACUTE IN VIVO SILDENAFIL CITRATE
(VIAGRA™) TREATMENT ON SEMEN PARAMETERS AND SPERM
FUNCTION
Summary 154
Introduction 154
Materials and methods 159
Results 164
Discussion 172
References 177
CHAPTER 8: SUMMARY, RECOMMENDATIONS AND CONCLUSIONS
8.1 Conclusion 182
Stellenbosch University http://scholar.sun.ac.za

xiii
8.1.1 Motility 1828.1.2 Acrosome reaction 184
8.2 Recommendations 1888.3 Future research 1898.4 References 191
Stellenbosch University http://scholar.sun.ac.za

xiv
LIST OF TABLES
Page
CHAPTER2
Table I Molecules that can induce the acrosome reaction in vivo. 40
CHAPTER 5
Table I Sperm-zona binding results after P0098059 treatment followed by 120
exposure to calcium ionophore and solubilized zona pellucida.
Table II Sperm-zona binding results expressed as a hemizona index (HZI) 121
after P0098059 (PO) treatment followed by exposure to calcium
ionophore (A23187) and solubilized zona pellucida (ZP).
CHAPTER 6
Table I Sperm kinematics results of all the samples as well as when 137
CHAPTER 7
Table I
Table II
Table III
Table IV
Table V
divided into normozoospermic and asthenozoospermic donors in
the presence and absence of LY294002.
Incidence of adverse events following treatment with placebo or 165
sildenafil citrate (n=20).
Initial macroscopic appearance and evaluation of semen after 165
either placebo or sildenafil citrate administration (n=20).
Effects of acute in vivo administration of sildenafil citrate on 166
microscopical secondary semen analysis parameters (n=20).
The effect of in vitro intracellular cGMP elevation by the addition of 167
8-Br-cGMP (20j.JM)on eliciting of the acrosome reaction (n=6).
Effects of 8-Br-cGMP (in vitro) and sildenafil (in vivo) on different
sperm motility parameters as measured by CASA.
171
Stellenbosch University http://scholar.sun.ac.za

xv
LIST OF FIGURES
CHAPTER2
Figure 1
Figure 2
Figure 3
Figure 4
Figure 5
Page
Schematic representation of the main events occurring under 8
conditions leading to capacitation and development of
hyperactivated motility of human spermatozoa in vitro.
Working model displaying the transmembrane and intracellular 11
signalling pathways to playa role in regulating sperm capacitation.
Regulation of protein tyrosine phosphorylation by a cAMP/PKA- 30
dependent pathway.
Crosstalk between signalling pathways involved in capacitation. 32
Diagram illustrating the main signal transduction pathways 42
activated during the process of acrosome reaction in response to
zona protein 3 (ZP3).
51Figure 6 Motion parameters of a single sperm track.
CHAPTER 3
Figure 1 Patterns recorded during FITC-PSA acrosome staining. 90
Figure 2 The competitive hemizona assay. 93
CHAPTER4
Figure 1 Influence of the MEK-inhibitor PD098059 (PO) on the acrosome 103
reaction (Mean±SE) mediated by A23187.
Figure 2
Figure 3
Influence of the MEK-inhibitor PD098059 (PO) on the acrosome 104
reaction (Mean±SE) mediated by ZP.
Possible interactions between the different signal transduction
pathways invoked during the acrosome reaction.
106
Stellenbosch University http://scholar.sun.ac.za

xvi
CHAPTER6
Figure 1 Correlation between beat cross frequency (BCF) and progressive 138
motility (PM) of pooled experiments (n=36).
Figure 2 Correlation between beat cross frequency (BCF) and amplitude of 138
lateral head displacement (ALH) of pooled experiments (n=36).
Figure 3 Histogram showing the percentage acrosome reaction (mean±SE) 140
of control (C) spermatozoa and spermatozoa after exposure to
zona pellucida (ZP), PI3K antagonist LY294002 (LY) or both ZP
and LY294002 (ZP+LY).
Figure 4 Histogram showing the number of control and LY294002 pre- 141
treated (Test) spermatozoa (mean±SE) tightly bound to each
hemizona respectively (n=18).
Figure 5 Possible interactions between the different signal transduction 147
pathways invoked during the acrosome reaction.
CHAPTER 7
Figure 1 Enhancement of penile erection by sildenafil citrate. 158
Figure 2 The effects of double-blind placebo or 50mg-sildenafil citrate 168
administration on the sperm acrosome reaction (n=20).
Figure 3 The effect of double-blind placebo or 50mg-sildenafil citrate 169
administration on the average number of spermatozoa tightly/firmly
bound to each hemizona (n=10; P=0.281).
CHAPTER 8
Figure 1 Hypothesized signal transduction pathways and possible 187
interactions between them.
Stellenbosch University http://scholar.sun.ac.za

xvii
ALPHABETICAL LIST OF ABBREVIATIONS
AA
AC
AKAPs
AlHAR8-Br-cGMP
BSA
BCF
[Ca2+]1
cAMP
CASA
cGMP
DAG
OFDMSO
ED
ERKFITC-PSA
FPP
GABAA
gluNAc
G-protein
GTP
HCl
= Arachidonic acid
= Adenylate cyclase
= A kinase-anchoring proteins
= Amplitude of lateral amplitude
= Acrosome reaction
= 8-bromo-cGMP
= Bovine serum albumin
= Beat-cross frequency
= Intracellular free ionized calcium concentration
= cyclic 3' ,5' adenosine monophosphate
= Computer assisted semen analysis
= cyclic 3',5' guanosine monophosphate
= Diacylglycerol
= Decapacitation factors
= Dimethylsulfoxide/sucrose
= Erectile dysfunction
= Extracellular signal-regulated kinases
= Fluorescein-labeled Pisum Sativum agglutinin
= Fertilization-promoting peptide
= y-aminobutyric acid A
= N-acetyl-a-D-glucosamine
= Guanine nucleotide binding protein
= Guanosine triphosphate
= Hydrochloric acid
Stellenbosch University http://scholar.sun.ac.za

xviii
HC03- = Bicarbonate
HTF = Human tubal fluid medium
HZA = Hemizona assay
HZI = Hemizona Index
ICSI = Intracytoplasmic sperm injection
IP3 = Inositol-1, 4, 5-triphosphate
IVF = In vitro fertilization
LC = Lyso-phosphatidylcholine
LIN = Linearity
LVA = low voltage activated
MAP = Microtubule-associated proteins
MAPK = Mitogen-activated protein kinases
MEK = ERK kinase
NANC = Non-adrenergic non-cholinergic
NaOH = Sodium hydroxide
NO = Nitric oxide
P = Progesterone
PA = Phosphatic acid
PBS = Phosphate buffered saline
POEs = Phosphodiesterases
POE5 = Phosphodiesterase type 5
PI3-K = Phosphatidylinositol 3-kinase
PIP2 = Phosphatidyl-inositol biphosphate
PL = Phospholipids
PLA2 = Phospholipase A2
Stellenbosch University http://scholar.sun.ac.za

PlCPLO
PKA
PKC
PKG
Ptdlns
PTK
ras-MEK-MAPK
ROS
SNARE
STR
TK
TKR
VAP
VClVOCC
VOCCT
VSlWHO
ZP
ZP3
ZRK
xix
= Phospholipase C
= Phospholipase 0
= Protein kinase A
= Protein kinase C
= Protein kinase G
= Phosphatidylinositol
= Protein tyrosine kinases
= Mitogen-activated protein kinase
= Reactive oxygen species
= Soluble N-ethylmaleimide-sensitive attachment protein
receptors
= Straightness
= Tyrosine kinase(s)
= Tyrosine kinase receptor
= Average path velocity
= Curvilinear velocity (Track speed)
= Voltage-operated calcium channels
= T-type voltage-operated calcium channels
= Straight-line (progressive) velocity
= World Health Organisation
= Zona pellucida
= Zona pellucida glycoprotein 3
= Zona receptor kinase
Stellenbosch University http://scholar.sun.ac.za

"The great tragedy of science ...
the slaying of a beautiful hypothesis by an
ugly fact."
- Thomas Huxley-
Stellenbosch University http://scholar.sun.ac.za

1
CHAPTER 1
INTRODUCTION AND STATEMENT OF PROBLEM
1.1 Introduction
The development of the fertilization competent state of the spermatozoon occurs
through a series of poorly understood processes. Successful fertilization involves
several steps including (1) movement/transit from the site of ejaculation to the site of
fertilization; (2) sperm capacitation in the female genital tract; (3) binding of
capacitated spermatozoa to the oocyte's extracellular coat, the zona pellucida (ZP);
(4) induction of the acrosome reaction; (5) penetration of the ZP; and (6) fusion of the
spermatozoon with the egg vitelline membrane.
Sperm-egg interaction is a carbohydrate-mediated species-specific event that
initiates a signal transduction cascade resulting in the exocytosis of sperm acrosomal
contents (i.e. the acrosome reaction). This step is believed to be a prerequisite that
enables the acrosome reacted spermatozoa to penetrate the ZP and fertilize the egg.
Researchers only recently started to investigate the intracellular mechanisms
resulting in acrosomal exocytosis (i.e. fusion and vesiculation of the sperm plasma
membrane and outer acrosomal membrane, allowing the exposure and release of the
acrosomal contents). Although various studies have been published, very little work
has been done on the intracellular signalling pathways elicited by the physiologically-
induced, i.e. the zona pellucida-induced, acrosome reaction as most researchers
make use of ligands mimicking zona pellucida action, thus ultimately leading to
conflicting results.
Stellenbosch University http://scholar.sun.ac.za

2
1.2 Objectives and statement of the problem
Against this introductory perspective the overall objective of this research study is to
present additional data that will assist reproductive biologists and clinicians in the
ongoing quest to unravel the intracellular pathways involved in the physiologically
induced acrosome reaction. The scarcity of human zonae pellucidae has placed the
investigation into the physiological acrosome reaction out of reach of most
reproductive biologists. Due to our fortunate accessibility to human zonae pellucidae
we could subsequently persue investigations into this reaction.
The aim of this study was to investigate different members and mechanisms by which
various intracellular signalling systems are activated during the zona pellucida-
induced acrosome reaction and their specific roles in the subsequent events leading
to fertilization. Sperm-zona interaction as well as various sperm motion
characteristics was also investigated. All these studies were performed on human
spermatozoa by making use of different signal transduction inhibitors.
1.3 Plan of study
This research project includes aspects that relate to intracellular signal transduction
processes involved in human sperm function such as inducibility of the acrosome
reaction, sperm-oocyte binding and motility.
As a background to the study, a broad overview of the current literature on
capacitation, acrosome reaction and kinematics of human spermatozoa is provided in
chapter two. Following this the basic materials used and methods followed during the
research project are outlined in Chapter 3.
Stellenbosch University http://scholar.sun.ac.za

3
The experiments have been divided into four research papers that are presented as
separate chapters. Each article has a specific summary, introduction, material and
methods, results, discussion and reference section pertaining to the study concerned.
Although each of these chapters is a complete and separate experiment, it is
important to remember that the data remain closely related, since all the results have
a mutual goal i.e. to further our understanding of the human sperm function in the
presence of zona pellucida.
The central theme of Chapters 4 and 5 will focus on the role of extracellular signal-
regulated kinase activation in human spermatozoa during the zona pellucida-induced
acrosome reaction and its involvement in sperm-zona pellucida binding.
Chapter 6 examines the ways by which phosphatidylinositol 3-kinase inhibition
enhance sperm motility parameters, while the effects of this inhibition on the zona
pellucida-induced acrosome reaction and sperm-oocyte binding was also addressed.
The effect of sildenafil citrate on the initiation of the zona pellucida-induced acrosome
reaction, sperm-oocyte binding and motility of human spermatozoa is the subject of
Chapter 7.
The final chapter (Chapter 8) gives a retrospective look at the study. Certain aspects
of the project will be highlighted and relevant suggestions will be made.
Stellenbosch University http://scholar.sun.ac.za

4
1.4 Conclusion
A thorough understanding of signal transduction in human spermatozoa will
ultimately yield information regarding the nature of receptors to which these signal
transduction pathways are coupled as well as the intracellular effectors that ultimately
regulate sperm function. Moreover, an understanding of these regulatory pathways
will be essential for the future development of clinical approaches designed to
enhance or preclude fertilization.
Stellenbosch University http://scholar.sun.ac.za

"Live as if your were to die tomorrow.
Learn as if you were to live forever."
- Mahatma Gandhi -
Stellenbosch University http://scholar.sun.ac.za

5
CHAPTER 2
LITERATURE REVIEW
2.1 Introduction
In mammals testicular sperm are morphologically differentiated but are neither
progressively motile nor able to fertilize an egg. Although the ability to move forward
is acquired during epididymal maturation, sperm are fertilization incompetent until
after a finite period of residence in the female reproductive tract (Visconti et al.,
2002). Two processes, namely capacitation and acrosome reaction (AR) are of
fundamental importance in the fertilization of the oocyte by the spermatozoon.
Physiologically occurring in the female genital tract, capacitation is a complex
process, which renders the sperm cell capable for specific interaction with the oocyte.
During capacitation, modification of membrane characteristics, enzyme activity and
motility properties of spermatozoa render these cells able to penetrate oocyte
investments and responsive to stimuli that induces the AR prior to fertilization. The
physiological AR occurs upon interaction of the spermatozoon with the zona pellucida
(ZP) and specifically zona pellucida protein 3 (ZP3). This is followed by liberation of
several acrosomal enzymes and other constituents that facilitate penetration of the
zona and expose molecules on the sperm equatorial segment that allows fusion of
the sperm membrane with the oolemma (Baldi et al., 2000). The molecular
mechanisms and the signal transduction pathways mediating the processes of
capacitation and AR have been partially defined, and appear to involve modifications
of intracellular calcium and other ions, lipid transfer and phospholipid remodelling in
the sperm's plasma membrane as well as changes in protein phosphorylation. Some
of the kinases and phosphorylated proteins that are involved in the processes of
Stellenbosch University http://scholar.sun.ac.za

6
capacitation and AR have been characterised, while characterisation of sperm
receptors to physiological inducers of the AR is in progress. The main signal
transduction pathways involved in capacitation and AR will subsequently be
summarised, as well as the various motility parameters and factors that might
influence it.
2.2 Capacitation
The process of capacitation consists of a series of functional biochemical and
biophysical modifications that render the ejaculated spermatozoa competent for
fertilization of the oocyte. This fundamental process normally takes place in the
female genital tract during the migration of spermatozoa to the site of fertilization as a
consequence of specific interactions between sperm and epithelial tubal cells
(Yanagimachi, 1994). However, under appropriate conditions, capacitation can also
be induced in vitro (Yanagimachi, 1994). Most of our knowledge regarding this
process has in fact been derived from in vitro studies. Following ejaculation, the
sperm surface is surrounded by molecules, known as decapacitation factors (OF),
that, until released, keeps the sperm in a non-capacitated state. These factors
associate with spermatozoa following contact with seminal fluids and are
progressively released from the sperm surface during capacitation. OFs thus
modulates capacitation thereby leading spermatozoa to the maximal fertilizing ability
at the site of fertilization (Fraser, 1999). It has been supposed that OFs, once
attached to the sperm surface, activate an intracellular Ca2+ -ATPase maintaining low
intracellular calcium concentration ([Ca2+]i) (Adeoya-Osiguwa & Fraser, 1996) and
when they are released from the surface [Ca2+]i increases. Several molecules have
been indicated as possible OFs (Yanagimachi, 1994).
Stellenbosch University http://scholar.sun.ac.za

7
It was recently demonstrated that two proteins present in seminal plasma, uteroglobin
and transglutaminase inhibit sperm capacitation and motility, thus representing two
possible OF candidates (Luconi et aI., 2000). Another sperm inhibitory factor present
in seminal fluid is cholesterol, which is known to inhibit several sperm functions,
including capacitation (Cross, 1996; Khorasani et al., 2000). In seminal plasma there
are also molecules that can stimulate the fertilizing ability of spermatozoa such as
fertilization-promoting peptide (FPP). FPP is a small peptide that promotes
capacitation (Funahashi et aI., 2000; Fraser, 1998) and inhibits spontaneous
acrosome loss, thereby retaining fertilization potential of sperm until the site of
fertilization is reached.
Capacitation is associated with the development of a distinct motility pattern called
hyperactivation (Yanagimachi, 1994), which is characterised by pronounced flagellar
movements, marked lateral excursion of the sperm head and a non linear trajectory.
Whether the development of hyperactivated motility is related to the biochemical and
biophysical changes occurring during the process of capacitation is still a matter of
debate. It is worth noting that signal transduction mechanisms involved in the
development of this special sperm motility pattern are similar to those described to
occur during capacitation (see later). It has recently been shown that, during in vitro
capacitation, modifications of mitochondrial morphology, which may be relevant for
the development of the hyperactivated motility pattern, also occur (Vorup-Jensen et
al., 1999). An additional manifestation of sperm capacitation is the acquisition of the
ability to undergo the AR in response to physiological stimuli such as ZP3 and
progesterone (P). The responsiveness of spermatozoa to ZP3 (Florman, 1994), P
Stellenbosch University http://scholar.sun.ac.za

8
(Baldi et al., 1998) and other stimuli of the AR increases during capacitation, assuring
maximal responsiveness at the site of fertilization. Capacitation involves
modifications in sperm surface protein distribution, alterations in plasma membrane
characteristics, changes in enzymatic activities and modulation of expression of
intracellular constituents (Yanagimachi, 1994). These events are made possible by
activation of a cascade of signalling pathways (schematised in Figure 1) effected by
unknown mediators during transit of sperm in the female reproductive tract or during
incubation in vitro in defined media.
ROS NO
PKA ~ cAMP•PTK Tyrosine phosphorylation+)/ il??.
HYPERACTIVA TED < Capacitation
PKC
outside
o Cholesterolacceptor
Cholesterol-+t Membranefluidity membrane
inside
PLAy
Figure 1. Schematic representation of the main events occurring under conditions
leading to capacitation and development of hyperactivated motility of human
Stellenbosch University http://scholar.sun.ac.za

9
spermatozoa in vitro. Changes in membrane permeability to several ions have been
described, among these Ca2+ and bicarbonate (HC03-), whose influx increase during
capacitation, have been reported to exert a primary role in the process. Membrane
fluidity increases due to the loss of cholesterol from the membrane, which may be
accelerated by the presence of a cholesterol acceptor in the external medium.
Remodelling of sperm membrane phospholipids (PL) and activation of
phospholipases (PLA-2 and PLCy-1) have also been shown: in particular, increased
synthesis of phosphatidylcholine from phosphatidyl-ethanolamine,
phosphatidylinositol and Iyso-phosphatidylcholine (LC) have been documented. A
time-dependent increase of tyrosine phosphorylation of proteins is associated with
development of capacitation. The increase of tyrosine phosphorylation is primarily
dependent on the increase in bicarbonate, which, in turn, activates adenyl cyclase
(AC) with increased generation of cAMP and subsequent activation of protein kinase
A (PKA). PKA activation leads to the activation of sperm tyrosine kinase(s) (TK). On
the contrary, Ca2+ inhibits tyrosine phosphorylation during capacitation. Other
possible physiological modulators of tyrosine phosphorylation during capacitation are
reactive oxygen species (ROS) that may be generated from spermatozoa or
leucocytes present in the ejaculate and nitric oxide (NO). An involvement of protein
kinase C (PKC) and ras-MEK-MAPK (mitogen-activated protein kinase) pathways
has also been reported. (From: Baldi et ai., 2000)
UNIVERSITEIT STELLENBOSCHBIBLIOTEEK
Stellenbosch University http://scholar.sun.ac.za

10
All these pathways may then crosstalk among each other determining the
development of capacitation. However, the exact relationship among these
modifications is not yet completely understood. As of yet there is no well-defined
method that allows distinction of capacitated from noncapacitated spermatozoa.
Capacitation does not occur synchronously in spermatozoa (Cohen-Dayag et a/.,
1995). In addition, capacitation is transient and already capacitated spermatozoa
cannot be capacitated again (Cohen-Dayag et a/., 1995). These complexities in in
vitro capacitation make it difficult to appropriately interpret in vitro studies. To
facilitate consideration of the complex cascade of molecular events that occur during
capacitation, a discussion of this process may be divided into events that initiate
capacitation and events that are a consequence of this process. Molecular events
implicated in the initiation of capacitation include: removal of cholesterol from the
sperm plasma membrane and changes in lipid distribution and composition; ion
fluxes resulting in alteration of sperm membrane potential as well as concentration of
intracellular calcium and other ions; and changes in protein phosphorylation and
kinase activities with emphasis on an increased tyrosine phosphorylation of proteins
involved in induction of hyperactivation and the AR. A working model for the initiation
of capacitation based on recent work is presented in Figure 2.
Stellenbosch University http://scholar.sun.ac.za

11
Cholesterol Efflux
Hyperpolarization ofSperm Plasma
Membrane Potential
PKA Activation
Protein TyrosinePhosphorylation
Adenylate CyclaseActivation
Figure 2. Working model displaying the transmembrane and intracellular signalling
pathways hypothesised to playa role in regulating sperm capacitation. This model is
based on work from a number of laboratories (Visconti et aI., 2002).
Stellenbosch University http://scholar.sun.ac.za

12
2.2.1 Cholesterol efflux and changes in membrane lipids and phospholipids
during capacitation
Changes in the distribution and composition of plasma membrane lipids and PL are
an important feature of sperm capacitation. These changes lead to an increase in the
membrane fluidity and to changes in the architecture and composition of the plasma
membrane (Yanagimachi, 1994). Serum albumin, an essential component of in vitro
capacitation media, is believed to function as a sink for cholesterol by removing it
from the sperm plasma membrane (Davis et al., 1979; Davis et al., 1980; Davis,
1981; Go & Wolf 1985; Langlais & Roberts, 1985; Suzuki & Yanagimachi, 1989;
Cross, 1996; Cross, 1998). Although serum albumin may have other roles during
capacitation (Espinosa et ai., 2000), its ability to facilitate cholesterol efflux is required
for capacitation. For example, capacitation is inhibited by the addition of cholesterol
and/or cholesterol analogues to the capacitation medium (Visconti et ai., 1999b).
Furthermore, serum albumin can be substituted in in vitro capacitation media with
cholesterol-binding compounds such as high density lipoproteins (HDL) (Therien et
ai., 1997; Visconti et ai., 1999b) and cyclodextrins (Choi & Toyoda, 1998; Cross,
1999; Osheroff et al., 1999; Visconti et al., 1999a) to induce capacitation. The
removal of cholesterol and likely other sterols (e.g. desmosterol) (Visconti et ai.,
1999b) from the plasma membrane is upstream of multiple signalling events intrinsic
to the capacitation process. Visconti and co-workers (1999) have demonstrated that
heptasaccharides (cholesterol-binding molecules) promote the release of cholesterol
from the plasma membrane of mouse sperm in the absence of bovine serum albumin
(BSA), increase tyrosine phosphorylation and promote capacitation of mouse sperm
as measured by the ability of the ZP to induce the AR and by successful fertilization
in vitro. These data suggest that cholesterol release is the signal that activates
Stellenbosch University http://scholar.sun.ac.za

13
membrane signal transduction pathways related to capacitation (Visconti et al.,
1999).
The removal of sterols decreases the cholesterol:phospholipid molar ratio in the
sperm plasma membrane as assessed by different criteria (Davis, 1981; Tesarik &
Flechon, 1986; Hoshi et al., 1990; Cross 1998). This could account for the membrane
fluidity changes (Wolf & Cardullo, 1991; Wolf et al., 1986) and redistribution of
membrane proteins, observed with lectins (Cross & Overstreet, 1987) and antibodies
(Shalgi et al., 1990; Rochwerger & Cuasnicu, 1992) that occur during capacitation.
From the standpoint of cell signalling, capacitation-associated alterations, in the
transbilayer phospholipid behaviour resulting in membrane lipid disorders, were
recently reported to occur through a cAMP-dependent pathway after exposure of
boar sperm to HC03- (GadelIa & Harrison, 2000). Therefore, multiple plasma
membrane modifications appear to contribute to the process of capacitation. It is also
important to consider what component of the female tract fluid might serve as a
cholesterol acceptor in vivo. Since fluids of the female tract are partially derived from
serum, serum-associated sterol acceptors could function in vivo. The identity of such
acceptors remains to be clarified.
The total amount of PL does not appear to change considerably during capacitation
(Yanagimachi, 1994). However, capacitation is associated with an increase of
phospholipid methylation and increased synthesis of phosphatidylcholine from
phosphatidylethanolamine (Llanos & Meizei, 1983). Incubation of spermatozoa under
capacitating conditions, in the presence of bicarbonate, does not alter phospholipid
distribution (Harrison & GadelIa, 1995). Such conditions, however, strongly inhibits
Stellenbosch University http://scholar.sun.ac.za

14
phospholipid transfer and leads to a slow increase of phosphatidylcholine
concentration in the inner leaflet of the membrane (Harrison & GadelIa, 1995).
Recently, GadelIa and Harrison (2000) have shown that the inclusion of bicarbonate
in the capacitating medium increases the translocation of phosphatidylcholine and
sphingomyelin from the outer to the inner leaflet of the membrane, possibly due to
activation of a bi-directional translocase (scramblase). Levels of phosphatidylinositol
and LC increase during capacitation in vivo in porcine sperm (Snyder & Clegg, 1975).
In view of the fusogenic properties of Iysophospholipids, an increase in their relative
amount may be relevant to prepare the sperm for the AR.
The molecular basis of signalling events induced by cholesterol efflux from sperm is
not well understood. In somatic cells, cholesterol removal is thought to disrupt lipid
rafts activating signalling events involving tyrosine kinases (TK), guanine nucleotide
binding proteins (G-proteins), and/or other molecules (Kabouridis et aI., 1997; Brown
& London, 1998; Roy et aI., 1999). The activation of similar signalling events during
capacitation in sperm correlates to the removal of cholesterol from the plasma
membrane. In sperm, cholesterol may likewise be concentrated in specialised plasma
membrane microdomains such as lipid rafts and caveolae (Brown & London, 1998).
This idea is supported by the recent finding that one important component of
caveolae, caveolin 1, is present in the plasma membrane overlying the acrosomal
region and the flagellum of mouse and guinea pig sperm (Travis et aI., 2000). The
hypothesis that lipid rafts and caveolae concentrate signalling complexes in the
sperm plasma membrane warrants continued investigation.
Stellenbosch University http://scholar.sun.ac.za

15
2.2.2 Ion fluxes and the regulation of sperm plasma membrane potential
During transit through the male and female reproductive tracts, sperm are exposed to
significant changes in the extracellular milieu, including variations in extracellular ion
concentrations and osmolarity. For example, caudal epididymal sperm are stored in
an environment that contains high K+, low Na+ and very low HC03- concentrations
(Brooks, 1983; Setchell et a/., 1994). These ion concentrations radically change when
the sperm are ejaculated, first in the seminal fluid and then in the female tract, where
the K+ is significantly reduced and the Na+ and HC03- concentrations are significantly
increased (Brooks, 1983; Setchell et a/., 1994; Yanagimachi, 1994). These dramatic
shifts in extracellular ion concentrations trigger modulations in intracellular ion
concentrations and consequently lead to an altered membrane potential of the sperm
plasma membrane.
Changes in intracellular ion concentrations have been associated with different
aspects of sperm function such as sperm motility in trout sperm (Morisawa & Suzuki,
1980; Gatti et a/., 1990), capacitation in mammalian sperm (Visconti et a/., 1995a;
Zeng et a/., 1995; Arnouit et a/., 1999) and the acrosome reaction in sperm from
multiple species (Arnouit et a/., 1996; Darszon et a/., 1999). The dramatic influence of
the external ion composition and the effect of channel blockers on sperm motility,
capacitation, and the AR strongly suggest that ion channels actively participate in the
regulation of sperm function. Ion channels can catalyse the flow of millions of ions
through the non-conducting lipid bilayer; therefore, a few ion channels can cause
changes in a small cell like the sperm within milliseconds (Darszon et aI., 1999). Ion
concentrations determine the plasma membrane potential through ion-selective
channels and control the extent of channel activity and ion flow. The plasma
Stellenbosch University http://scholar.sun.ac.za

16
membrane potential can also regulate ion channel activity as well as second
messenger levels. For example, in trout sperm, changes in the plasma membrane
potential by changes in extracellular K+ concentration modulate sperm motility
through a cAMP pathway (Morisawa & Ishida, 1987). Moreover, Beltran et ai., (1996),
demonstrated in sea urchin sperm that cAMP synthesis could be regulated by
changes in membrane potential.
In vitro, the resting membrane potential is determined by the relative permeabilities of
the sperm plasma membrane for ions that constitute the capacitation media. Under
normal conditions, sperm maintain an internal ion concentration markedly different
from that in the extracellular medium and these differences establish the resting
plasma membrane potential. The ion composition of capacitation media mimics that
of oviductal fluid (Yanagimachi, 1994). These media are high in Na+ (about 130 mM)
and cr (about 100 mM), but low in K+ (about 5.9 mM). Capacitation media also
contain Ca2+ (about 2.7 mM) and HC03- (10-25 mM). In contrast, intracellular fluids of
sperm have a low concentration of Na+ (about 14 mM) and high concentration of K+
(about 90-120 mM) (Babcock, 1983; Zeng et ai., 1995). The free intracellular Ca2+
concentration of non-capacitated sperm is approximately 0.1 M or less, but during the
AR it may increase to approximately 10M (Bailey & Storey, 1994; Arnouit et al.,
1999). To date, the intracellular concentrations of cr and HC03- in sperm have not
been determined. These differences between extracellular and intracellular ion
concentrations are established by the respective ion permeabilities and determine the
resting membrane potential in mammalian sperm.
Stellenbosch University http://scholar.sun.ac.za

17
Recently, Zeng et a/., (1995) demonstrated that capacitation is accompanied by
hyperpolarization of the sperm plasma membrane. Membrane hyperpolarization may
be partially due to an enhanced K+ permeability related to the decrease in inhibitory
modulation of K+channels during capacitation (Zeng et a/., 1995). Since capacitation
prepares the sperm for the AR, capacitation-associated hyperpolarization may
regulate the ability of sperm to generate transient Ca2+ elevations during AR by
physiological agonists (e.g. ZP of the egg or P). This hypothesis is consistent with the
presence of low voltage activated (LVA) Ca2+ T-channels in spermatogenic cells
(Arnouit et a/., 1996; Lievano et a/., 1996) that may also be present in mature sperm.
A signature property of LVA Ca2+ channels is a low threshold for voltage-dependent
inactivation. These Ca2+ channels are inactivated at holding potentials between -80
and -60 mV and cannot be activated readily from more positive holding potentials
(Arnouit et a/., 1996; Lievano et al., 1996). Thus, if LVA Ca2+ T-channels are involved
in the regulation of the AR, sperm must maintain a hyperpolarized membrane
potential during the early stages of interaction with the egg (Arnouit et a/., 1999;
Florman et al., 1998). Presently, little is known about the regulation of capacitation-
associated hyperpolarization. In order to understand the ionic basis of these changes
in sperm plasma membrane potential, it will be necessary to analyse how the
aforementioned ion permeabilities change during capacitation.
2.2.2.1 Modification in concentration of intracellular calcium and other ions
during capacitation (HC03-, Ca2+, and the cAMP pathway)
Numerous studies have demonstrated that capacitation is Ca2+ -dependent
(DasGupta et al., 1993; Visconti et a/., 1995a). The initiation and/or regulation of
capacitation by Ca2+ occur via different targets, some of which are involved with
Stellenbosch University http://scholar.sun.ac.za

18
cAMP metabolism. Since in sperm Ca2+/calmodulin can activate both the synthesis of
cAMP by adenylate cyclase (Gross et aI., 1987), as well as degradation by cAMP
cyclic nucleotide phosphodiesterase (Wasco & Orr, 1984), it is not surprising that
Ca2+ has both positive and negative actions on capacitation and related signalling
events. In this respect, Ca2+ has a positive effect on mouse sperm by inducing
capacitation-associated changes in protein tyrosine phosphorylation (Visconti et al.,
1995a). In contrast, Ca2+ has been demonstrated to inhibit protein tyrosine
phosphorylation in human sperm during the first 2 h of in vitro capacitation (Carrera
et aI., 1996; Luconi et aI., 1996). An increase in intracellular sperm Ca2+ during
capacitation has been described by some investigators, whereas others have shown
that no changes in Ca2+ levels occur during this maturational event (Yanagimachi,
1994). This ambiguity could be due, in part, to the well-demonstrated action of Ca2+
on the AR and to the inherent difficulties in differentiating between these events.
However, the action of Ca2+ at the level of effector enzymes involved in sperm signal
transduction suggests that this divalent cation is likely to play an important role in
capacitation.
Modification of intracellular concentration of calcium ions ([Ca2+]i) is the most fully
characterised biochemical event during capacitation. An increase in the concentration
of Ca2+ during capacitation has been demonstrated in several mammalian species
(Yanagimachi, 1994) including human (Baldi et aI., 1991; Garcia & Meizei, 1999).
Extracellular Ca2+ is indeed one of the necessary constituents for the completion of
capacitation of spermatozoa in vitro (Yanagimachi, 1994). In spermatozoa [Ca2+]i is
regulated by a Ca2+-ATPase (acting as a Ca2+ extrusion pump) (Fraser & McDermott,
1992), Ca2+/H+ exchanger system and Na+/Ca2+ antiporter (acting as Ca2+ entrance
Stellenbosch University http://scholar.sun.ac.za

19
systems) in the plasma membrane (Fraser, 1995), and by putative intracellular Ca2+
stores, whose presence in human sperm has been suggested by several evidences
(Blackmore, 1992; Walensky & Sneider, 1995; Dragileva et ai., 1999; O'Toole et ai.,
2000). However, the role of intracellular calcium stores in the physiology of
spermatozoa is still questioned (O'Toole et ai., 2000; Kirkman-Brown et ai., 2000;
Kobori et al., 2000). Recent data (Dragileva et al., 1999) indicate that cytosolic Ca2+
is actively transported into the acrosome by an ATP-dependent, thapsigargin-
sensitive pump and that it may be released from the acrosome through an inositol-
1,4,5-triphosphate (IP3)-gated calcium channel. More recently, the existence of store-
operated calcium channels that mediate sustained calcium increase in response to
ZP3 in mouse sperm has been demonstrated (O'Toole et ai., 2000). The location of
such stores remains to be demonstrated since endoplasmic reticulum is not present
in mature spermatozoa and the acrosome does not appear (according to recent
studies performed in individual spermatozoa) to retain significant amounts of calcium
(Kirkman-Brown et ai., 2000; Kobori et ai., 2000). It has been hypothesised that
modulation of the activity of the Ca2+-extrusion system, in particular Ca2+-ATPase,
occurs during capacitation leading to an increase in intracellular Ca2+ (Fraser &
McDermott, 1992; Fraser, 1995). Drugs such as quercetin, that inhibit Ca2+-ATPase,
accelerate capacitation (Fraser & McDermott, 1992; Fraser, 1995). A Na+/Ca2+
exchanger is present in mammalian sperm, however, its role in controlling
intracellular Ca2+ during capacitation is not clear. Similarly, although voltage-operated
calcium channels (VOCG) have been demonstrated in mammalian spermatozoa
(Benoff, 1998), their role in regulating intracellular calcium concentration during
capacitation is not clear. Moreover, although the presence of T-type voltage-operated
calcium channels (VOCCT) in germ cells has been unequivocally demonstrated
Stellenbosch University http://scholar.sun.ac.za

20
(Arnouit et al., 1996; Darszon et al., 1999) and their involvement in the effect of ZP3
on mature sperm indicated (Arnouit et ai., 1996, O'Toole et ai., 2000), the role of
these channels in the process of capacitation is far from being defined. However, it
seems quite clear that sperm VOCCT may be modulated during capacitation. Indeed,
during this process the sperm membrane becomes hyperpolarized due to enhanced
K+ permeability (Brewis et al., 2000). This hyperpolarization may act to prime VOCCT
from an inactivated state to a closed one, which can be activated by an agonist
inducing depolarisation such as ZP3 (Arnouit et ai., 1998). Downstream targets of
calcium include the calcium-binding protein calmodulin, whose involvement in sperm
capacitation has been recently demonstrated (Si & Olds-Clarke, 2000). Besides Ca2+,
intracellular K+ (Zeng et ai., 1995), Na+ (Hyne et ai., 1985) and cr (Fraser, 1995)
concentrations have been shown to be modulated during capacitation. The increase
of intracellular Na+ appears to be important for capacitation, since the Na+ ionophore,
monensin, promotes this process in mouse sperm (Fraser, 1995). The intracellular
concentration in zinc ion decreases in the acrosome of hamster spermatozoa during
capacitation (Andrews et ai., 1994). In addition, incubation of spermatozoa in a zinc-
containing medium inhibits the process (Andrews et ai., 1994). These findings
suggest that zinc may play a role in destabilisation of plasma membrane during
capacitation (Andrews et ai., 1994).
A rise in intracellular pH has been reported during capacitation of bovine sperm
(Vredenburgh-Wilberg and Parrish, 1995). However, the role of pH in sperm
capacitation is not yet clear, since an artificial increase in intracellular pH in
spermatozoa does not accelerate the process (Fraser, 1995). Cross et al. (1997)
have shown that a cholesterol efflux from the plasma membrane, during the
Stellenbosch University http://scholar.sun.ac.za

21
capacitation of human spermatozoa, determines a rise in intracellular pH and
responsiveness to P. Recent data have demonstrated the presence of a Na+
dependent Cr/HC03- exchanger that regulates bicarbonate transport and motility in
mature sperm (Zeng et al., 1996), however the role of this pump in capacitation is not
clear. It has been shown that bicarbonate regulates adenylate cyclase (AC) activity
and cAMP metabolism (Visconti et al., 1990) and is necessary for tyrosine
phosphorylation of proteins during capacitation (see later). It is of interest that
bicarbonate concentration is low in the epididymis and high in the seminal plasma
and in the oviduct indicating that modifications of bicarbonate concentration in
reproductive tracts play an important role in the suppression of capacitation in the
epididymis and in the promotion of this process in the female reproductive tract
(Wassarman, 1999).
Numerous studies have demonstrated that capacitation is HC03--dependent (Lee &
Storey, 1986; Neill & Olds-Clarke, 1987; Boatman & Robbins, 1991; Shi & Roldan,
1995; Visconti et ai., 1995a). Little is known about the mechanisms of HC03-
transport in sperm. However, the ability of 4,4'-diidothiocyanatostilbene-2,2'-disulfonic
acid (OIOS), a well-known inhibitor of anion transporters, to inhibit the actions of
HC03- on various sperm functions suggests that sperm contain functional anion
transporters (Okamura et al., 1988; Visconti et al., 1990; Spira & Breitbart,
1992;Visconti et ai., 1999c).
The transmembrane movement of HC03- anions into sperm could be responsible for
the known increase in intracellular pH that is observed during capacitation (Uguz et
al., 1994; Zeng et al., 1996). An additional target for the action of this anion could be
Stellenbosch University http://scholar.sun.ac.za

22
the regulation of sperm cAMP metabolism, since the synthesis of cAMP by
mammalian sperm AC is markedly stimulated by HC03- (Okamura et aI., 1985; Garty
& Salomon, 1987; Visconti et aI., 1990). The increase in cAMP during capacitation
and the stimulation of AC activity in sperm by increased levels of intracellular HC03-
and Ca2+ implicate a role for this enzyme and the cAMP-signalling pathway in
capacitation. AC in sperm has been the subject of multiple studies, but whether one
or more proteins represent it remains controversial.
Sperm AC has unique properties when compared with somatic cell cyclases (Garbers
& Kopf, 1980; Leclerc & Kopf, 1995). For example, unlike somatic cell cyclases,
responses of sperm AC to agents that modulate stimulatory GTP binding proteins
(Gs), such as cholera toxin, AIF4- or GTP analogues, are weak or completely absent.
Since no cholera toxin-ADP ribosylated substrate has been detected in mammalian
sperm, the low sensitivity to G-protein effectors could be due to the lack of Gs protein
in these cells (Hildebrandt et aI., 1985). Another possibility is that the sperm AC is
unable to interact with Gs proteins due to differences in cyclase tertiary structure. As
mentioned above, an important property of the sperm AC is its regulation by HC03-
anion (Okamura et al., 1985).
Recent studies suggest that the sperm AC is a post-translationally modified form of
the testicular soluble AC (Buck et al., 1999). Similar to the sperm AC activity, the
enzymatic activity of recombinant testicular soluble AC is stimulated by HC03- anions
(Chen et al., 2000). In addition, antibodies against the catalytic domain of the
testicular soluble AC recognised two sperm proteins in corresponding to the deduced
molecular masses of the processed and unprocessed forms of the testicular enzyme
Stellenbosch University http://scholar.sun.ac.za

23
(Chen et al., 2000) suggesting that this cyclase remains associated with sperm after
spermatogenesis. Interestingly, the sequence from the catalytic domain of this
cyclase has sequence homology to cyanobacterial AC and the cyanobacterial
cyclase is also HC03--dependent (Chen et aI., 2000). Although the testis cyclase has
been found in the soluble fraction, it is significant that cyclase activity identified in
mammalian sperm remains associated with the particulate membrane fraction.
Therefore, the testicular soluble AC found would be predicted to have a mechanism
allowing for translocation from the cytosol to the membrane at some point during
spermatogenesis.
2.2.3 Changes in protein phosphorylation and protein kinase activity during
capacitation
Protein phosphorylation during sperm capacitation has been widely studied in the last
five years, leading to the generation of more than 100 papers in the international
literature. The best studied kinases involved in capacitation are Ca2+-calmodulin
activated kinases, cAMP-dependent kinases (pKA), calcium and phospholipid
activated protein kinase (PKC), which induce phosphorylation of proteins in serine
and threonine residues, and TKs, which phosphorylate proteins in tyrosine residues.
The involvement of kinase pathways in the process of capacitation has been mainly
studied by using inhibitors of such pathways. However, specificity of kinase inhibitors
is doubtful and many of them may affect other intracellular signal transduction
pathways. In this light, caution should be applied in interpreting results of these
studies (Baldi et al., 2000).
Stellenbosch University http://scholar.sun.ac.za

24
2.2.3.1 Involvement of AC/cAMP/PKA pathway in capacitation
A spontaneous increase of cAMP during capacitation has been demonstrated (White
& Aitken, 1989) and inhibitors of PKA, the serine-threonine kinase activated by this
pathway, inhibit capacitation (Aitken et ai., 1998). Pentoxifylline, which promotes an
increase in cAMP by inhibiting sperm phosphodiesterases, induces capacitation (Ain
et ai., 1999). It has been well established that Ca2+ and HC03- stimulate AC but the
exact mechanism by which these ions activate AC is not clear (Visconti et al., 1990;
Garty & Salomon, 1987). cAMP generated during capacitation and subsequent
activation of PKA appears to play a key role in the increase of tyrosine
phosphorylation during capacitation (Visconti et ai., 1999, also see later). Since PKA
may phosphorylate several cellular substrates, its sequestration in specific cellular
compartments is necessary to spatially restrict its action, thus ensuring specificity of
functions. Compartmentalisation of PKA is accomplished by A-kinase-anchoring
proteins (AKAPs) which have been recently characterised in sperm of different
species (Carerra et al., 1996; Moss et ai., 1999; Vijayaraghavan et ai., 1999). Their
specific localisation to the tail of the sperm indicates a role for these proteins in the
modulation of sperm motility (Vijayaraghavan et ai., 1999). Recently, a role for the
AC/cAMP/PKA pathway in plasma membrane lipid remodelling, occurring during
capacitation, has been demonstrated (Harrison & Miller, 2000). Yet, many questions
still remain unanswered concerning the AC/cAMP/PKA pathway in human sperm.
Although the specific expression of a soluble AC has been shown in male germ cells
(Sinclair et ai., 2000), the types of phosphodiesterases (PDEs) present in these cells
remain to be defined. Recently, Richter et al. (1999) detected, the presence of mRNA
transcripts for several PDEs in ejaculated human spermatozoa, but no conclusions
Stellenbosch University http://scholar.sun.ac.za

25
can be drawn concerning the type of POE specifically expressed in mature
spermatozoa.
2.2.3.2 Involvement of PKC in capacitation
The presence of PKC in mammalian spermatozoa and its role in sperm motility and
the process of AR are documented (Breitbart & Noar, 1999), but PKC activity is very
low compared to somatic cells (Breitbart & Noar, 1999, Bonaccorsi et a/., 1998). The
role of this enzyme during capacitation is poorly understood. Early studies
demonstrated that stimulation of PKC with phorbol esters accelerates the process of
capacitation (Rotem et a/., 1992). This effect was inhibited by PKC inhibitors,
suggesting that PKC may be involved in capacitation (Rotem et a/., 1992). In
addition, PKC may also be involved in epidermal growth factor-induced capacitation
(Furuya et a/., 1993). All these studies were performed using high levels of phorbol
esters as PKC inducers. Although it was shown that PKC activity of human sperm
can be stimulated by a phorbol ester (Bonaccorsi et a/., 1998), additional effects of
these tumour promoters on other sperm kinases cannot be excluded. Recently, the
role of PKC in capacitation has been questioned (Ain et a/., 1999).
2.2.3.3 Involvement of tyrosine phosphorylation in capacitation
The first evidence for the presence of tyrosine phosphorylated proteins in mammalian
spermatozoa dates back to 1989 (Leyton & Saling, 1989). Using anti-
phosphotyrosine antibodies, Leyton and Saling (1989) identified three different
phosphoproteins at 52, 75, and 95 kOa in the mouse spermatozoa. The 95 kOa
protein was tyrosine phosphorylated under all experimental conditions and including
interaction of spermatozoa with solubilized ZP proteins (Leyton & Sailing, 1989).
Stellenbosch University http://scholar.sun.ac.za

26
Capacitation is characterised by a spontaneous, time-dependent increase of tyrosine
phosphorylation of different proteins (Luconi et al., 1995; Visconti et al., 1995). The
main tyrosine phosphorylated proteins are in the range of 95-100 kDa (Leyton &
Saling, 1989; Luconi et al., 1995; Visconti et al., 1995; Luconi et al., 1996; Osheroff
et al., 1999). Capacitation-associated changes in protein tyrosine phosphorylation
have been demonstrated in multiple species including the mouse (Visconti et al.,
1995a), bovine (Galantino-Homer et al., 1997), human (Leclerc & Kopf, 1995;
Osheroff et ai., 1999), pig (Kalab et ai., 1998) and hamster (Devi et ai., 1999; Visconti
et al. 1999c; Jha & Sjivaji, 2001). In the mouse, in vitro capacitation of caudal
epididymal sperm promotes tyrosine phosphorylation of a subset of proteins between
Mr 40000 and 120000 (Visconti et al., 1995a). At least in the mouse, the increase of
protein tyrosine phosphorylation is dependent on the presence of Ca2+, NaHC03- and
BSA (Visconti et ai., 1995). Specifically, the absence of anyone of these media
constituents prevents both protein tyrosine phosphorylation and capacitation. It is
necessary to mention that the effect of media constituents on protein tyrosine
phosphorylation and capacitation varies slightly from species to species (Visconti et
ai., 1999c; Jha & Sjivaji, 2001). It is worth to note that some of these results have not
been confirmed in human sperm. Indeed, in humans, Ca2+ and tyrosine
phosphorylation seem to be inversely related and protein tyrosine phosphorylation is
enhanced in calcium free medium (Luconi et al., 1996; Carrera et al., 1996) indicating
that calcium might induce the activation of tyrosine phosphatases (Carrera et al.,
1996). The dependence of in vitro protein tyrosine phosphorylation on serum
albumin, or other cholesterol acceptors, indicates a correlation between cholesterol
efflux and cAMP-induced tyrosine phosphorylation. It has been demonstrated that in
BSA-deprived media protein tyrosine phosphorylation and sperm capacitation are
Stellenbosch University http://scholar.sun.ac.za

27
inhibited (Visconti et al., 1995; Osheroff, 1999). Whether cholesterol removal is
upstream from or co-incidental with the action of Ca2+ and/or NaHC03" is not
presently known. It is hypothesised that cholesterol removal, with a resultant change
in sperm plasma membrane fluidity, modulates Ca2+ and/or HC03" ion fluxes leading
to AC activation; this hypothesis remains to be tested.
The increase in protein tyrosine phosphorylation is regulated by an AC/cAMP-
dependent pathway that involves PKA in sperm from the mouse (Visconti et ai.,
1995b), bull (Galanti no-Homer et ai., 1997), human (Leclerc et ai., 1996; Osheroff et
ai., 1999), boar (Kalab et ai., 1998) and hamster (Visconti et ai., 1999c), as well as by
reactive oxygen species generated at the beginning of capacitation (LeClerc et al.,
1998; see later). In addition, recent data suggest a role for nitric oxide generated by
human spermatozoa in the promotion of capacitation (Herrero et al., 1998;
Francavilla et al., 2000) and the increase of tyrosine phosphorylation (Herrero et al.,
1998).
The involvement of PKA is indicated since inhibitors of PKA activity are able to inhibit
tyrosine phosphorylation as well as capacitation. Since PKA is not able to
phosphorylate proteins on tyrosine residues, an intermediate TK may be involved.
Figure 3 summarises three possible mechanisms: (1) direct or indirect stimulation of
a TK by PKA; (2) direct or indirect inhibition of a phosphotyrosine phosphatase; and
(3) direct or indirect phosphorylation of proteins by PKA on serine or threonine
residues that prime these proteins for subsequent phosphorylation of tyrosine
residues. These distinct possibilities are currently being explored in different
Stellenbosch University http://scholar.sun.ac.za

28
laboratories. The identification of the enzymes responsible for the tyrosine
phosphorylation pathway(s) will improve our knowledge of the capacitation process.
As stated above, in human sperm the highest degree of tyrosine phosphorylation was
found in a protein of 95-97 kDa (Leyton & Saling, 1989; Luconi ef al., 1995; Visconti
ef al., 1995; Luconi ef al., 1996; Osheroff, 1999). A tyrosine-phosphorylated protein in
this molecular weight range was previously indicated as the possible sperm receptor
for ZP3, zona receptor kinase (ZRK) (Burks ef al., 1995). This protein has been
characterised, partially cloned and sequenced. Its sequence shows a 55% homology
with the receptor-like PTK c-eyk (Burks ef aI., 1995) and 97-100% homology with the
proto-oncogene c-mer (Bork ef al., 1996). Among the other tyrosine phosphorylated
proteins in the same molecular weight range, an AKAP specifically expressed at the
tail level has been recently identified (Carrera ef al., 1996; Moss ef al., 1999;
Vijayaraghavan ef al., 1999). This protein may be involved in the development of the
hyperactivated motility pattern. An extracellular signal regulated kinase pair (ERK-1
and ERK-2) of sperm proteins that are phosphorylated on tyrosine and activated
during sperm capacitation was also identified (Luconi ef al., 1998; Luconi ef al.,
1998). Their inhibition with a pharmacological compound suppresses capacitation
(Luconi ef al., 1998), indicating a role for these proteins in the process.
Immunofluorescence labelling of phosphotyrosine residues, indicated that
capacitation as well as exposure to zona proteins increased the degree of tyrosine
phosphorylation in each spermatozoon and the number of sperm cells
phosphorylated in the acrosomal region of the sperm head (Naz ef aI., 1991).
Incubation of spermatozoa with antiphosphotyrosine antibodies or inhibition of TK
activity inhibited zona-free hamster egg penetration (Naz ef al., 1991), prevented AR
Stellenbosch University http://scholar.sun.ac.za

29
and blocked fertilization (Leyton et ai., 1992). However, whether tyrosine
phosphorylation is fundamental for the development of the capacitated state or is
simply associated to the phenomenon stay a matter of debate. It was also shown that
erbstatin, a potent inhibitor of TK, did not inhibit capacitation measured as the ability
of spermatozoa to respond to P (Luconi et ai., 1996). Similarly, Ain et al. (1999), have
reported that thyrphostin A-47, a PTK inhibitor, does not inhibit pentoxifylline-
stimulated capacitation, although it suppress the AR stimulated by this agent. To
understand the role of tyrosine phosphorylation in the process of capacitation, we will
probably need to wait for the characterisation of many, if not all, of the tyrosine
phosphorylated proteins as well as the TKs that are activated during the process
(Baldi et al., 2000).
Stellenbosch University http://scholar.sun.ac.za

30
Protein Kinase A
t Tyrosine Kinase SerlThrPhosphorylation
I Phosphotyrosine'" Phosphatase
TyrosinePhosphorylation
Other kinases
Figure. 3. Regulation of protein tyrosine phosphorylation by a cA,MPIPKA-dependentltp
pathway. (1) A protein tyrosine kinase is stimulated through direct phosphorylation by
PKA or by an enzymatic cascade that involves phosphorylation by PKA. (2) A
phosphotyrosine phosphatase is inhibited through direct phosphorylation by PKA or
by an enzymatic cascade that involves phosphorylation by PKA. (3) Protein targets
become substrates for protein tyrosine activity after phosphorylation in ser or thr
residues by PKA (Visconti et aI., 2002).
Stellenbosch University http://scholar.sun.ac.za

31
2.2.3.4 Crosstalk between different signalling events during sperm capacitation
Cyclic AMP appears to be a central regulator of several sperm processes, such as
motility (Eddy & O'Brien, 1994), capacitation (Visconti et al., 1995b; Visconti et al.,
1997), and the AR (De Jonge et al., 1991a; Leclerc & Kopf, 1995; Garde & Roldan,
2000). Changes in membrane potential are also involved in these sperm functions
(Morisawa & Suzuki, 1980; Florman et al., 1992; Zeng et al., 1995; Arnouit et al.,
1996; Darszon et al., 1999). Presently, it is not known whether these two
(capacitation and AR) signalling events interact. However, it is likely that these two
processes are related, since capacitation is accompanied by both hyperpolarization
of the plasma membrane and an increase in cAMP synthesis. Supporting this idea,
the presence of a membrane potential-regulated AC has been reported in non-
mammalian species (Beltran et aI., 1996). In addition, although PKA is the main
downstream effector for cAMP in sperm, the view that PKA mediates all of the effects
of cAMP has been amended with the discovery of new types of cyclic nucleotide
receptors. These receptors include cyclic-nucleotide-gated channels, exchange
factors (Kawasaki et al., 1998), a cGMP binding cyclic nucleotide POE, and
extracellular cAMP receptors (Shabb & Corbin, 1992). Cyclic nucleotide-gated
channels were identified in sea urchin (Gauss et al., 1998) and mammalian sperm
(Weyand et al., 1994) with specificity for cAMP and cGMP, respectively. Altogether,
these data support the idea that crosstalk might occur between modification in
membrane potential and the cAMP signalling pathway during capacitation. Alternative
possibilities are summarised in Figure 4.
Stellenbosch University http://scholar.sun.ac.za

32:--------------------------------------------: (3), ,
eSA --'1 eSA-chol. ,(a)
- ~ I ~b)..-fr- L__ C_h_O_I.__ Hyperpolarization ~ .-} tAC -tcAMP - tPKA
1 (2) tHCO;
*/+tHC03-
tAC --. tcAMP --. tPKA --. Hyperpolarization
CAMP-gated channel 1
Figure 4. Crosstalk between signalling pathways involved in capacitation. (1)
Hyperpolarization is upstream to the increase in cAMP synthesis. Cholesterol
removal regulates sperm plasma membrane potential through a I<" channel or
through the increase in anionic permeability, (a) hyperpolarization may regulate AC
activity as described in sea urchin and trout sperm (Beltran et aI., 1996; Morisawa &
Ishida, 1987) or (b) hyperpolarization may regulate HC03- permeability and in this
way activate AC. (2) Hyperpolarization is downstream of the increase in cAMP
synthesis. In this model, cholesterol removal regulates an HHC03- permeability and
HC03- stimulates AC, cAMP could then activate a cyclic nucleotide gated channel
directly or indirectly through phosphorylation by PKA leading to plasma membrane
hyperpolarization. (3) Hyperpolarization and cAMP synthesis are independently
associated with capacitation.
Stellenbosch University http://scholar.sun.ac.za

33
2.2.3.5 Consequences of capacitation on sperm function
Although fertilization still represents the endpoint of sperm capacitation, the ability of
the sperm to undergo a regulated AR (e.g. in response to the ZP or P) can be taken
as an earlier, upstream endpoint of capacitation (Florman & Babcock, 1991; Visconti
et al., 1998). Capacitation is also correlated with changes in sperm motility patterns,
designated as hyperactivation, in a number of species (Suarez, 1996; Yanagimachi,
1994). When one attempts to understand the process of capacitation at the molecular
level, events occurring both in the sperm head (i.e. AR) and in the tail (i.e. motility
changes) must be considered. Therefore, one may postulate that components of both
the sperm exocytotic and motility machinery are modified during capacitation. Some
of these alterations may involve changes in the phosphorylation status of certain
proteins, changes in protein localisation, and/or modification of protein-protein
interactions. Experiments leading to the identification and characterisation of these
effector molecules will further increase our understanding of capacitation.
To understand the link between capacitation and the AR, a better knowledge of the
mechanisms that regulate this exocytotic event in sperm is necessary. Exocytosis is
a tightly regulated, complex process that involves fusion of subcellular vesicles with
the overlying plasma membrane and release of vesicular contents. Recent evidence
suggests that membrane fusion is governed by a few conserved protein families
regardless of whether membrane fusion occurs between intracellular organelles or
between trafficking vesicles and the plasma membrane. Proteins involved in fusion
events include a family of proteins commonly referred to as SNARE proteins (soluble
N-ethylmaleimide-sensitive attachment protein receptors) (Jahn & Sudhof, 1999).
Stellenbosch University http://scholar.sun.ac.za

34
Sperm homologues of SNARE proteins as well as SNARE-associated proteins, such
as Rab 3A and NSF, have been detected in sea urchin (Schulz et ai., 1997; Schulz et
al., 1998) and mammalian sperm (Michaut et al., 2000; Ramalho-Santos et al., 2000;
Yunes et al., 2000). These observations support the idea that the sperm AR might be
regulated in similar ways to exocytotic processes in somatic cells. Since capacitation
is necessary for exocytosis in mammalian sperm, elucidation of the mechanisms
regulating the AR will also increase our understanding of capacitation.
Capacitation is also linked to events that occur in the sperm flagellum. For example,
two members of the AKAP family located in the fibrous sheath become
phosphorylated at tyrosine residues during human sperm capacitation (Carrera et al.,
1996; Mandai et ai., 1999; Vijayaraghavan et ai., 1999). AKAPs represent a growing
family of scaffolding proteins that function to tether the regulatory subunits of PKA
and signalling enzymes, such as calcineurin and PKC, to organelles or cytoskeletal
elements. These proteins permit the precise control of signal transduction in discrete
regions of the cell (Pawson & Scott, 1997). Tyrosine phosphorylation of AKAPs might
alter the biochemical and biophysical properties of these proteins and the fibrous
sheath and thus, contribute to the regulation of events associated with flagellar
bending, including changes in tail wave amplitude during hyperactivation.
Stellenbosch University http://scholar.sun.ac.za

35
2.3 The sperm acrosome
The acrosome plays an important role at the site of sperm-zona (egg) binding during
the fertilizing process. Clinical studies have identified a distinct group of men whose
infertility is associated with abnormal AR (Benoff, 1997). Since the acrosome acts in
concert with the plasma membrane overlying the acrosome during the early events of
fertilization, any discussion on its formation and organisation will contribute to our
understanding of its functional significance.
The acrosome is a Golgi-derived secretory granule that is formed during an early
stage of spermiogenesis. It resembles the cellular lysosome, a bag like structure that
normally functions in intracellular digestive and defensive mechanisms, in three
different ways. First, both the acrosome and the lysosome are derived from the Golgi
apparatus. Secondly, both organelles stain a bright orange red colour with acridine
orange, indicating an acidic pH within the organelles. Finally the two organelles
contain several common enzymes such as acid glycohydrolases, proteases,
esterases, acid phosphatases and aryl sulfatases (Tuisiani et al., 1998; Zaneveld &
De Jonge, 1991). Despite these similarities the acrosome has some distinctive
features. The sperm acrosome is a sac-like structure surrounded by inner and outer
acrosomal membranes. Immediately after sperm (receptor)-zona ligand) binding, the
outer acrosomal membrane fuse with the overlying plasma membrane, releasing the
acrosomal contents (glycohydrolases, proteases, etc.) at the site of sperm-egg
binding (acrosomal exocytosis). The acrosome is also different from the cellular
lysosome in that it contains antigens such as acrosin (Saling, 1989), acrogranin
(Anakwe & Gerton, 1990), and sperm protein AM67 (Foster et aI., 1997). Because of
Stellenbosch University http://scholar.sun.ac.za

36
these differences and its exocytotic properties, the sperm acrosome is considered
analogous to a secretory granule (Eddy & O'Brien, 1994).
Several biochemical and ultrastructural studies have provided evidence for the
involvement of cytoskeletal domains such as actin (Talbot & Kleve, 1978), calmodulin
(Camatini & Casale, 1987), and a-spectrin-like antigens (Virtanen et aI., 1984) in the
organisation of the acrosome. In addition, the organelle contains filamentous
structures primarily associated with the outer acrosomal membrane (Olson et al.,
1987). However the functional significance of the filamentous structures, if any, is not
yet known (Abou-Haila & Tulsiani, 2000).
Today, most researchers agree that the powerful hydrolytic enzymes
(glycohydrolases, proteases, etc) released at the site of sperm-egg binding, along
with the enhanced thrust generated by the hyperactivated beat pattern of the bound
spermatozoa (Katz & Drobnis, 1990), are the important factors regulating the
penetration of ZP and fusion of the gametes.
2.3.1 The acrosome reaction (AR)
In all mammals, sperm cells are required to fertilize oocytes, thereby providing a
haploid set of chromosomes with a paternal pattern of genomic imprinting needed for
normal development and triggering oocyte activation (Loeb, 1915; Austin, 1954;
Yanagimachi, 1994). It has long been known that successful fertilization is dependent
on the extracellular ionic environment, in large part because this can modify the
intracellular composition of gametes. The first observation was made in the sea
Stellenbosch University http://scholar.sun.ac.za

37
urchin when it was noted that fertilization did not occur in the absence of extracellular
Ca2+ (Loeb, 1952) due to failure of the AR to occur. After the development of
successful culture systems for mammalian gametes, it was possible to demonstrate
that mammalian sperm fertilizing ability, like that of invertebrate sperm, can be
modulated by alterations in extracellular components (Fraser, 1995).
The AR is an exocytotic process physiologically induced by ligand (ZP3)-receptor
interaction, consisting in multiple fusions between the outer acrosomal membrane
and the overlaying plasma membrane leading to the release of acrosomal enzymes
and exposure of the molecules present on the inner acrosomal membrane surface
that mediate fusion with the oolemma. The sperm acrosome is a Golgi-derived
structure forming a cap over the anterior region of the nucleus that contains many
hydrolytic enzymes and consists of an anterior cap and a posterior region called the
equatorial segment (Yanagimachi, 1994). As mentioned previously, only capacitated
sperm are physiologically able to undergo the AR in response to physiological stimuli.
It is thus conceivable that the two process, capacitation and AR, are sequentially and
functionally linked such that several of the effectors involved in mediating intracellular
signalling activated by AR start to be tuned during capacitation. For instance, the
increase of intracellular calcium levels and tyrosine phosphorylation of proteins
accompanying capacitation is also essential for the subsequent exocytosis in
response to P (Baldi et a/., 1991; Aitken et a/., 1998). The AR consists of the
development of multiple fenestrations between the outer acrosomal membrane and
the plasma membrane of the spermatozoa (Yanagimachi, 1994). This lead to the
release of the enzymatic content of the acrosome and to the exposure of the
enzymes bound to the inner membrane adjacent to the nuclear envelope
Stellenbosch University http://scholar.sun.ac.za

38
(Yanagimachi, 1994). In the absence of any specific stimuli only a low percentage of
human spermatozoa can undergo the AR (Leyton & Saling, 1989). It has been
suggested that self-aggregation of the sperm receptor for ZP may account for this
spontaneous acrosome reaction (Saling, 1989). A wide variety of molecules present
on the surface of the sperm have been proposed as putative candidates for the ZP3
receptor (Wassarman, 1999). The involvement of different receptors in different
species or different binding affinities, or even multiple receptors that may co-operate
in sequence to induce AR may justify this rather long list of possible candidates.
However, none of these molecules has been definitively recognised as "the sperm-
egg receptor" (Wassarman, 1999). Progesterone and a highly conserved ZP
glycoprotein termed ZP3 have been identified as natural oocyte-associated AR-
inducing ligands, and their sequential action has been shown to support the
occurrence of the physiological AR (Melendrez et ai., 1994; Roldan et ai., 1994).
There are also apparent divergences between the two pathways because the one
used by the ZP ligand involves a pertussis toxin sensitive G-protein (Franken et ai.,
1993) whereas that used by P does not (Tesarik et ai., 1993). Moreover, P also
stimulates transmembrane chloride fluxes employing a plasma membrane channel
sharing some, but not all properties with neuronal y-aminobutyric acid A (GABAA)
receptor (Wistrom & Meizei, 1993; Blackmore et ai., 1994; Shi & Roldan, 1995).
It is well known that P opens a sperm plasma membrane calcium channel and
activates phosphotyrosine kinase independently of each other (Mendoza et al.,
1995). Progesterone therefore reacts with a multiple-receptor system on the surface
and this system co-operates with that used by ZP3 to control the physiological AR.
Stellenbosch University http://scholar.sun.ac.za

39
Each of the respective receptors alone can eventually induce some of the AR events
and in some cases complete acrosomal exocytosis.
The AR can also be physiologically induced by P. It is present at high levels in the
cumulus matrix, surrounding the oocyte, that must be crossed by sperm in order to
reach the ZP. This steroid has been described to affect several other sperm
functions, including capacitation, motility and the priming effect on the ZP3-induced
AR through stimulation of a rapid nongenomic signalling pathway mediated by P
receptors present on the sperm surface (for further details on effects of P see Baldi et
a/., 1998; Bray et a/., 1999). P-induced AR is inhibited by 17b-estradiol through
interaction with a specific nongenomic estrogen receptor on the sperm plasma
membrane (Luconi et a/., 1999). This suggest that estradiol, which is present at
micromolar levels in the follicular fluid, may act as a physiological modulator of P
action on sperm assuring the appropriate timing of activation in the fertilization
process. Other agonists proposed to act as in vivo inducers of the AR are listed in
Table 1.
Stellenbosch University http://scholar.sun.ac.za

40
Table 1. Molecules that can induce the acrosome reaction in vitro.
AGONIST REFERENCEZona pellucida protein 3 (ZP3) Yanagimachi, 1994; Saling, 1989
Prostaglandin E1 Schaefer et al., 1998
ATP Foresta et al., 1993
Dibutyryl cAMP (cAMP analogues) De Jonge et al., 1991b
Progesterone & 17bOH-progesterone Baldi et al., 1998; Bray et al., 1999
Platelet-activating factor Sengoku et al., 1992; Krausz et al., 1994;
Atrial natriuretic peptide Breitbart & Naor, 1999
Epidermal growth factor Furuya et al., 1993
Serum albumin Yanagimachi, 1994
Manosylated bovine serum albumin Benoff et al., 1997a
A23187 Fênichel et al., 1989
GABA Shi et al., 1997
Forskolin De Jonge et al., 1991b
Pentoxifylline Gearon et al., 1994
1-oleoyl-2-acetylglycerol (DAG analogue) Rotem et al., 1992
Phorbol diesters (PMA, TPA) De Jonge et al., 1991 b; Lee et al., 1987
Thapsigargin Meizei & Turner, 1993
Stellenbosch University http://scholar.sun.ac.za

41
Many molecular mechanisms have been demonstrated to be active during AR
(Figure 5), but for many of them a precise cause-effect relationship has not yet been
defined, such that we still do not know whether they are essential for the process of
fertilization or whether they are simply associated with the process. The fundamental
difference between P- and ZP3-induced AR stands on the nature of their receptors.
In fact, in the case of ZP3, its action is mediated by receptor recruitment of G-
proteins which are not involved in a P stimulated cascade (Wassarman, 1999). One
of the first events that occur in spermatozoa following stimulation with ZP3 and P is
receptor aggregation (Saling, 1989; Tesarik & Mendoza, 1993). This is followed by a
cascade of downstream membrane and cytosolic signalling factors involved in
induction of AR (schematised Figure 5). Among them, the roles of calcium,
phospholipases and protein kinases are discussed below.
Stellenbosch University http://scholar.sun.ac.za

42
Na'~']ljInside
1]--=, \, pH, ••.........
ACROSOME REACTION
Figure 5. Diagram illustrating the main signal transduction pathways activated during
the process of AR in response to zona protein 3 (ZP3). Following interaction with the
agonist, aggregation of receptors for ZP3 (ZRK) induces TK activation (which
increases protein tyrosine phosphorylation) and autophosphorylation of the receptor.
Activation of phosphatidylinositol-3kinase (PI3K) has been also reported. A guanine
nucleotide binding protein (G-protein) transduces the signal interacting with
membrane-bound enzymes like phospholipase C (PLC) and AC. Activation of these
two enzymes lead to increased generation of the second messengers cyclic
adenosine monophosphate (cAMP), IP3 and diacylglycerol (DAG). A consequence of
the increase of second messengers is the activation of protein kinases such as
cAMP-dependent kinase (PKA) and Ca2+ and phospholipid-dependent kinase (PKC)
with increased protein phosphorylation. cAMP-dependent influx of sodium (Ns') has
~......... 1
Stellenbosch University http://scholar.sun.ac.za

43
been reported. IP3 may increase intracellular Ca2+ by liberation of the ion from
intracellular Ca2+ stores. The increase of intracellular Ca2+ consequent to activation of
ZP3 receptors is completely due to influx from the extracellular medium, is dependent
on activation of G-proteins, involves voltage-dependent Ca2+ channels and is
accompanied by an efflux of H+ which determines a rise of intracellular pH (pHi).
Partial dependence of Ca2+ -intiux from TK activation has been reported. Ca2+
dependent activation of phospholipase A2 (PLA2) and phospholipase 0 (PLO) [with
increased generation of other second messengers as arachidonic acid (AA), Iyso-
phosphatidylcholine (LC) and phosphatidic acid (PA) from membrane phospholipids
(PL)] have also been described to occur during AR. (From: Baldi et aI., 2000)
2.3.1.1 Increase in intracellular calcium during acrosome reaction
Calcium plays a central role in receptor-mediated response and membrane fusion
processes in spermatozoa (Yanagimachi, 1994). Calcium ionophores are the most
widely used non-physiological inducers of AR (Yanagimachi, 1994). Although the AR
can be induced in the absence of extracellular calcium with some agonists (Foresta
et aI., 1993; Krausz et aI., 1994; Bielfield et aI., 1994) stimulation of calcium fluxes is
one of the earlier responses activated by most stimuli that induce the AR. The shape
of the calcium wave is different during P- and ZP- stimulation. P elicits a biphasic
calcium wave, consisting of a rapid initial peak followed by a long lasting plateau
phase (Baldi et al., 1998). While ZP3 induce a slow and sustained increase of
calcium (Florman, 1994), resembling the second phase of P. However, it has been
shown, using a different methodological approach, that ZP3 may induce a rapid
increase of intracellular calcium concentrations occurring within milliseconds after
stimulation (Arnouit et al., 1999). Parallel inhibition of both AR and the second phase
Stellenbosch University http://scholar.sun.ac.za

44
of calcium response to P by both TK blockers and a previous administration of
estradiol, suggests the plateau phase to be responsible for agonist- induced AR
(Bonaccorsi et a/., 1995; Tesarik et a/., 1996; Bray et a/., 1999). Recently, a direct
relationship between the sustained calcium phase and AR in response to ZP3 has
been demonstrated (O'Toole et a/., 2000). Concerning P, such a relationship is less
apparent, since the percentage of sperm undergoing AR in response to the steroid is
smaller than that where a sustained response is observed (Kirkman-Brown et a/.,
2000; Kobori et a/., 2000). Although extracellular calcium depletion totally prevents
both ZP3 and P-induced intracellular calcium increase, recent data suggest the
possible involvement of intracellular calcium stores in sperm the AR (Walensky &
Snyder, 1995; Dragileva et a/., 1999; O'Toole et a/., 2000). In particular, it has been
shown that calcium entry during the sustained phase in response to ZP3 is due to
activation of store-operated channels (O'Toole et a/., 2000). While membrane
voltage-operated calcium channels of T-type have been demonstrated in mediating
ZP-induced calcium influx (Arnouit et a/., 1996; Darszon et a/., 1999), the nature of P-
stimulated calcium channels is still a matter of discussion (Garcia & Meizei, 1999;
Blackmore & Eisoldt, 1999; Patrat et a/., 2000). Calcium plays a key role in the fusion
events in the sperm membrane (Watson et a/., 1995). Using a pyroantimonate-
osmium fixation technique, the temporal and spatial location of intracellular calcium
granules was monitored during AR in ram spermatozoa (Watson et a/., 1995). Ca2+ is
initially associated with the outer acrosomal membrane. As the process progresses,
Ca2+ associates with the fusion sites between the outer acrosomal membrane and
the plasma membrane anteriorly to the equatorial segment. At later stages, Ca2+ is
localised in both post acrosomal dense lamina and on the outer acrosomal
Stellenbosch University http://scholar.sun.ac.za

45
membrane under the equatorial segment. These findings suggest that Ca2+ may be
implicated in the fusion process (Watson et al., 1995).
Increase of intracellular Ca2+ in response to ZP3 and P is associated with an efflux of
H+ and a rise in intracellular pH (Florman et al., 1989; Garcia & Meizei, 1996; Brook
et al., 1996). P has been shown to rapidly stimulate sodium influx (patrat et al., 2000)
and the presence of sodium in the extracellular medium is absolutely required for
induction of the AR by the steroid (Patrat et al., 2000; Garcia & Meizei, 1996). Sperm
intracellular pH may be regulated by a Na+IH+ exchanger (Garcia & Meizei, 1999) as
well as by Cr/HC03- (Holappa et al., 1999), whose presence have been
demonstrated in germ cells.
2.3.1.2 Phospholipase activation during acrosome reaction
The presence of PlA2, PLC and other phospholipases have been demonstrated in
mammalian spermatozoa (Roldan, 1998). The roles and activities of these enzymes
in human sperm capacitation and AR have been recently reviewed (Roldan, 1998). It
was also shown previously that during the AR in boar spermatozoa, the amount of
DAG and free fatty acids increases (Nikolopoulou et al., 1986). This finding was
consistent with activation of phospholipases (Roldan, 1998). In fact, increase of
[Ca2+]i stimulated by ionophores and P leads to activation of PLC in human
spermatozoa (Roldan, 1998). Such activation leads to an intracellular increase in the
amount of IP3 and DAG. Similarly, ZP3 activates the phosphoinositide specific
enzyme phospholipase Cg1 by virtue of tyrosine phosphorylation and leads to its
translocation from cytosol to particulate fractions (Tomes et al., 1996). Presence and
activation of phosphatidylcholine-specific PLC during the AR has also been
Stellenbosch University http://scholar.sun.ac.za

46
demonstrated (Roldan, 1998). DAG and IP3, produced by calcium-dependent
activation of PLC, may be respectively involved in the regulation of PKC activity and
release of calcium from putative sperm intracellular stores (Walensky & Snyder,
1995; Breitbart & Noar, 1999).
AR induced by ionophores and P also leads to the activation of PLA2 (Roldan, 1998).
This activation is associated with generation of lipid metabolites, such as arachidonic
acid and Iysophospholipids. Phosphatidylcholine, Iysophospholipids, and unsaturated
fatty acids, such as arachidonic acid, are potent inducers of AR (Cross, 1994;
Fleming & Yanagimachi, 1981; Kyono et al., 1984) and may be implicated in the
fusion process that occurs during AR. Moreover, LC generated from PLA2 activation
may act as a substrate for generation of platelet-activating factor (PAF) (Baldi et al.,
1993; Kumar et al., 1988). This phospholipid, which is synthesised in response to P
(Baldi et al., 1993), may further enhance AR (Krausz et al., 1994; Fukuda et al.,
1994; Huo & Yang, 2000). In vitro treatment of human sperm with PAF (1-0-alkyl-2-
acetyl-sn-glycero-3-phosphocholine) enhances penetration of zona free hamster
oocytes (Minhas, 1993). In animal studies, in vitro treatment of sperm with PAF
significantly improves fertilization rate using both intra cytoplasmic sperm injection
(ICSI) and in vitro fertilization (IVF) techniques without showing detrimental effects on
subsequent embryo development (Minhas et al., 1996; Lee et al., 1997).
2.3.1.3 Involvement of protein kinases in acrosome reaction process
During AR, activation of protein kinases (serine-threonine kinases as PKC, PKA and
protein kinase G [PKG], and TKs) is downstream to the production and/or activation
of early second messengers. Early evidence for the involvement of PKA in the AR,
Stellenbosch University http://scholar.sun.ac.za

47
demonstrated that AC activity and cAMP generation increase during this process (De
Jonge et ai., 1991a). Adenyl cyclase stimulators, such as forskolin, and the cAMP
analogue dibutyryl cAMP, induce AR in a dose-dependent manner in mammalian
sperm (Anderson et ai., 1992; Garde & Roldan, 2000; Harrison & Meizei, 2000;
Kobori et ai., 2000). In addition, PKA inhibitors have been reported to suppress the
AR induced by P (Harrison & Meizei, 2000), although controversial results are
present in the literature concerning an increase of cAMP synthesis in response to P
(Parinaud & Milhet, 1996; Schaefer et al., 1998). Taken together, these findings
suggest that PKA activation may be involved in the AR process (Baldi et al., 2000).
The involvement of PKC in the process of AR is still under debate. Although
activators such as phorbol esters and synthetic DAG induce the AR, PKC activity in
human sperm is very low and the identity of PKC substrates is still under
investigation. Induction of AR by solubilized ZP was partially reduced by pre-
treatment with inhibitors of PKA, PKC and PKG tested separately, while combinations
of them caused a significantly greater inhibition (Bielfield et ai., 1994). These results
suggest a concomitant role for PKA, PKC and PKG in the human ZP-induced AR
(Bielfield et ai., 1994). Both P (O'Toole et ai., 1996; Bonaccorsi et ai., 1998) and ZP
(Liu & Baker, 1997) increase sperm PKC activity. Such activation is downstream to
calcium influx (P (Bonaccorsi et ai., 1998), since it can be prevented by calcium
channel blockers (O'Toole et ai., 1996).
ZP3 induced an increase in tyrosine phosphorylation of sperm proteins (Ley ton &
Saling, 1989; Burks et al., 1995). A major tyrosine phosphorylated protein in
mammalian spermatozoa was 95-97 kDa (Leyton & Saling, 1989; Burks et ai., 1995).
Stellenbosch University http://scholar.sun.ac.za

48
Proteins in this molecular weight range appear to undergo phosphorylation during
capacitation (see above) and in response to P (Baldi et a/., 1998). Recombinant
human ZP3 produced by genetic engineering has been demonstrated to induce AR
and tyrosine phosphorylation of the 95-110 kDa protein in human sperm (Brewis et
a/., 1998), perfectly mimicking the physiological action of this glycoprotein. Known
inhibitors of TK, such as genistein and tyrphostin-47, block ZP3-induced AR (Leyton
et a/., 1992). Both the TK inhibitors tyrphostin A48 and pertussis toxin suppress ZP3-
induced calcium influx in mouse spermatozoa (Bailey & Storey, 1994). Using a
similar pharmacological approach, Tesarik et al. (1993) and Luconi et al. (1995)
showed involvement of TKs in the P-mediated AR. Activation of TK also seem to be
involved in the plateau phase of increase in the amount of intracellular Ca2+ in
response to P (Bonaccorsi et a/., 1995; Tesarik et a/., 1996). The mechanisms
through which TKs are activated in sperm during the process of AR are still under
investigation. Although during capacitation activation of the AC/cAMP/PKA pathway
appears to be highly involved, activation of this pathway in response to stimuli that
induce AR is less clear (Parinaud & Milhet, 1996; Schaefer et a/., 1998). The
involvement of reactive oxygen species, generated by sperm in response to stimuli
that induce AR, in tyrosine phosphorylation has been suggested by a few
researchers (de Lamirande et a/., 1998; Fisher et a/., 1998).
Recent data (Fisher et a/., 1998) indicate the involvement of PI3K in the AR
stimulated by agents that mimic ZP3. These include mannose-BSA and polyclonal
antibodies raised against the p95 protein (indicated as the possible sperm ZP3
receptor; see ZRK above), but not by P or ionophores (Baldi et a/., 2000).
Stellenbosch University http://scholar.sun.ac.za

49
2.4 Motility
Human spermatozoa emerge highly differentiated from the testes, but as for all other
eutheria, they are motionless or feebly motile. This immotility is apparently due to the
"immaturity" of the plasmalemma, as demembrated spermatozoa can be induced,
under appropriate conditions to move almost as actively as mature spermatozoa from
the cauda (Mohri & Yanagimachi, 1980). The spermatozoa continue to develop
during their passage through the epididymis. Only at ejaculation, when they are
mixed with the secretions of the accessory glands, do spermatozoa undergo motility
activation, acquiring a mature motility pattern and fertilizing ability.
Sperm motility plays an important role in transport of the spermatozoa in the female
reproductive tract before fertilization (Yanagimachi, 1969). Sperm motility is a
complex phenomenon, the understanding of which requires integration of cell biology
with reproductive physiology, biochemistry, biophysics and clinical andrology. The
spermatozoon is an intricate motile cell, whose motility depends on a flagellum that
develops the propulsive force for swimming. The likelihood of achieving a pregnancy
increases with decreasing proportions of immotile spermatozoa and with increasing
quality of sperm progression (Bostofte et al., 1983; Bostofte et al., 1984).
Over the years a plethora of terminology has appeared to describe the various
movement characteristics of motile spermatozoa. Consensus on these parameters
was reached at the "Automated Sperm Motility Analysis" workshop held at the
American Society of Andrology's Annual Meeting in Houston (Tx, USA) during March
1988 (Mortimer 1990). These parameters have now been standardised and are
currently used in motion analysis (Owen & Katz, 1993). These parameters include:
Stellenbosch University http://scholar.sun.ac.za

50
Curvilinear velocity (VCL): The VCl (urn/s) is the time-average velocity of a sperm
head along its actual curvilinear path.
Straight line velocity (VSL): The VSl (um/s) is the time-average linear velocity of a
sperm head along the straight line between the start and the end of the observed
track.
Average path velocity (VAP): The VAP (!-ImIs) is the time-average velocity of a sperm
head along its average path. This path is computed by smoothing the actual path
according to algorithms in the CASA instrument.
Amplitude of lateral head displacement (ALH): The AlH (urn) is the magnitude of
lateral displacement/deviation of a sperm head about its average path. It can be
expressed as a maximum or an average of such displacements.
Beat-cross frequency (BCF): The BCF (Hz) is the average rate at which the sperm's
curvilinear path crosses its average path.
Straightness (STR): The STR (%) is the ratio of VSLlVAP and measures the
straightness of the average path.
Linearity (Lin): The Lin (%) is reported as the ratio of VSLlVCl and measures the
linearity of the curvilinear path. The value range from 0 to 100 with a value of 100
representing cells swimming in a straight line.
All these motility parameters, as depicted in Figure 6, give a clear indication of
movement characteristics of spermatozoa (WHO, 1999).
Stellenbosch University http://scholar.sun.ac.za

51
vel
Figure 6: Motion parameters of a single sperm track, where ALH = amplitude of
lateral head displacement; BGF = beat-cross frequency; VAP = average path
velocity; VGL = curvilinear velocity and VSL = straight line velocity.
Stellenbosch University http://scholar.sun.ac.za

52
2.4.1 Factors influencing sperm motility
Enhanced sperm motility values have been associated with improved conception
rates (Edvinsson et a/., 1983). In this regard, efforts have been directed towards
improvement of poor motility of sperm as well as the inducement of motility of
immotile cells. The practice of mixing sperm with a motility stimulating agent for
improving the successful rate of artificial insemination has been reported (Yovich et
al., 1988).
2.4.1.1 Cyclic adenosine mono-phosphate (cAMP)
Cyclic nucleotides play a major role in sperm capacitation, metabolism, AR and
motility (Tash & Means, 1983). This was first observed when it was noted that
caffeine and other methyl-xanthines, which act as cyclic nucleotide PDE inhibitors
and associated with an increase in intracellular cAMP, can stimulate sperm motility
and metabolic activity (Hoskins, 1973).
Garbers et al. (1971) first showed that the addition of dibutryl cAMP stimulated the
motility of bovine caudal epididymal spermatozoa. cAMP was shown to have effects
on the metabolism and motility of sperm (Tash & Means, 1982). Since then the cAMP
system of spermatozoa has been extensively studied.
Elevation of intracellular cAMP levels appear to be related to:
... the acquisition of the potential for motility during epididymal maturation;
... the actual initiation of motility upon ejaculation; and
...the surge of increased motility associated with capacitation.
Stellenbosch University http://scholar.sun.ac.za

53
The cAMP content of diluted spermatozoa increases two fold as sperm traverse the
epididymis (Hoskins et ai., 1974). This seems reasonable since the intracellular
concentrations of cAMP correlates directly with the degree of motility (Hoskins &
Casillas, 1975). Furthermore, a 25% decrease in cAMP levels is observed as washed
bovine caudal sperm become immotile as compared to the initial motility (Hoskins et
ai., 1974). During ejaculation, as bovine caudal epididymal spermatozoa are mixed
with secretions from the accessory glands, cAMP levels double within 30 seconds
(Cascieri et ai., 1976). A similar rise in cAMP concentration has been reported in
hamster spermatozoa (Morton et ai., 1974).
cAMP has also been implicated with sperm motility during capacitation and AR
(Rosado et ai., 1974). Vigorous motility is commonly observed concomitant with
capacitation and is believed to be essential for penetration of the egg following the
AR (Yanagimachi, 1970). Reyes et al. (1977) showed that the addition of exogenous
cAMP to rabbit or human spermatozoa more than doubled the rate of capacitation in
vitro.
cAMP appears to activate cAMP dependant kinase, which phosphorylates key
proteins that are required for motility (Lindemann & Kanous, 1989). Deactivation
occurs when these proteins are dephosphorylated by phosphoprotein phosphatase.
The phosphorylation of several integral axonemal proteins has been reported to be
cAMP-dependant (Tash et al., 1984; Horowitz et al. 1988).
Stellenbosch University http://scholar.sun.ac.za

54
2.4.1.2 Adenylate cyclase
Mammalian sperm possess both AC and POEs, which are involved in the regulation
of the intracellular concentration of cAMP (Bhatnagar et al., 1982) and also possess
cAMP-dependant protein kinase through which the physiological action of cAMP is
mainly mediated. Pariset et al. (1983) reported that there is a positive correlation
between the motility index and the cAMP-dependant protein kinase activity of human
spermatozoa.
A significant positive correlation was found between sperm motility and cAMP content
and AC activity in human spermatozoa (Ishikawa et al., 1989). These results suggest
that sperm motility is controlled by the AC activity in spermatozoa and that the
disturbance of sperm motility in infertile men is probably caused by reduced activity of
AC.
2.5 Summary
Mammalian sperm are not immediately fertile upon release from the male
reproductive tract, despite their ability to exhibit vigorous motility. They require a
species dependent period of time during which they undergo a series of changes,
collectively referred to as capacitation (Austin, 1952; Yanagimachi, 1994), that are
needed for cells to become fully competent to fertilize an oocyte. When capacitated,
mammalian sperm can (i) express hyperactivated motility, the very vigorous, thrusting
pattern of motility that is needed for penetration of the oocyte investments and (ii)
interact with the oocytes (including cumulus cells, folucular fluid and ZP) to undergo
the AR. Capacitation is a unique feature of mammalian sperm that occurs in the
female tract and is essential for fertilization. During capacitation, sperm acquire
Stellenbosch University http://scholar.sun.ac.za

55
hyperactive motility and the ability to undergo a regulated AR. Capacitation is
accompanied by changes in lipid composition of the plasma membrane as well as by
protein post-translational modifications regulated by several signalling pathways.
Studying in vitro capacitation has allowed characterisation of a number of
biochemical events that occur in human spermatozoa. It is not clear whether the
same or similar events occur during capacitation in vivo. P, PAF and other effectors
present in the follicular fluid and/or cumulus matrix, could facilitate capacitation or
prime the AR in vivo. For instance, the clinical significance of P effects has been
shown by several studies showing a significant correlation of response to P with
sperm fertilizing ability as well as reduced or absent response to P in infertile subjects
(Baldi et a/., 1998; Bray et aI., 1999). Moreover, treatment of human sperm with P
has been shown to enhance hamster oocyte penetration (Aitken et aI., 1996). These
studies indicate that sperm P receptor, as well as the one that mediates ZP3
stimulation, is involved in the process of fertilization representing a possible target for
developing pharmacological strategies to potentiate sperm fertilizing ability in
assisted reproductive techniques or contraceptive molecules. Functional Preceptors
on the human sperm surface have been recently characterised (Luconi et al., 1998;
Falkenstein et aI., 1999). Sequencing of the receptor and cloning of the encoding
gene will represent next steps in such characterisation.
The AR is an exocytotic event that promotes interaction and penetration through the
ZP and confers fusogenic properties on the remaining plasma membrane in the
sperm head (Yanagimachi, 1994).
Stellenbosch University http://scholar.sun.ac.za

56
It has been suggested that the importance of capacitation may actually be to prevent
sperm from becoming fertile too quickly, given that spermatozoa are deposited into
the lower regions of the female reproductive tract and still need to travel some
considerable distance in order to reach the site of fertilization (Bedford, 1983). The
AR must therefore also be accurately timed to ensure fertilization, since a premature
AR leads to the loss of ZP recognition sites from the sperm surface and thus impairs
sperm-zona pellucida binding (Franken et aI., 1993). On the other hand inability of
zona bound spermatozoa to activate the AR also prevents zona penetration.
Although work emanating from multiple laboratories is leading to a better
understanding of capacitation, AR and motility, several of the proteins involved in
these processes remain to be characterised.
Stellenbosch University http://scholar.sun.ac.za

57
References
Abou-Halla A, Tulsiani DR (2000) Mammalian sperm acrosome: formation, contents
and function. Arch Biochem Biophys 379(2): 173-182.
Adeoya-Osiguwa S & Fraser LR (1996) Evidence for Ca2+-dependent ATPase
activity, stimulated by decapacitation factor and calmodulin, in mouse sperm.
Mol Reprod.Dev 44, 111-120.
Ain R, Uma Devi K, Shivaji S, Seshagiri PB (1999) Pentoxifylline-stimulated
capacitation and acrosome reaction in hamster spermatozoa: involvement of
intracellular signalling molecules. Mol Hum Reprod 5, 618-626.
Aitken RJ, Buckingham DW, Irvine DS (1996) The extragenomic action of
progesterone on human spermatozoa: evidence for ubiquitous response that is
rapidly down-regulated. Endocrinology 137, 3999-4009.
Aitken RJ, Harkiss D, Knox W, Paterson M, Irvine DS (1998a) A novel signal
transduction cascade in capacitating human spermatozoa characterised by a
redox-regulated, cAMP-mediated induction of tyrosine phosphorylation. J Cell
Sci 111,645-656.
Aitken RJ, Harkiss D, Knox W, Paterson M, Irvine DS (1998b) On the cellular
mechanisms by which the bicarbonate ion mediates the extragenomic action of
progesterone on human spermatozoa. Bioi Reprod 58, 186-96.
Anakwe 00, Gerton GL (1990) Acrosome biogenesis begins during meiosis:
evidence from the synthesis and distribution of an acrosomal glycoprotein,
acrogranin, during guinea pig spermatogenesis. Bioi Reprod 42:317-328.
Stellenbosch University http://scholar.sun.ac.za

58
Anderson RA, Feathergill KA, De Jonge CJ, Mack SR, Zaneveld LJO (1992)
Facilitative effect of pulsed addition of dibutyryl cAMP on the acrosome reaction
of noncapacitated spermatozoa. JAndrol 13, 398-408.
Andrews JC, Nolan JP, Hammerstedt RH, Bavister BO (1994) Role of zinc during
hamster sperm capacitation. Bioi Reprod 51, 1238-1247.
Arnouit C, Cardullo RA, Lemos JR, Florman HM (1996) Activation of mouse sperm T-
type Ca2+ channels by adhesion to egg zona pellucida. Proc. Natl. Acad. Sci.
USA 93,13004-13009.
Arnouit C, Kazam lG, Visconti PE, Kopf GS, Villaz M, Florman HM (1999) Control of
the low voltage-activated calcium channel of mouse sperm by egg ZP3 and by
membrane hyperpolarization during capacitation. Proc Natl Acad Sci USA 96,
6757-6762.
Arnouit C, Villaz M, Florman HM (1998) Pharmacological properties of the T-type
Ca2+ current of mouse spermatogenetic cells. Mol Pharmacol 53, 1104-1111.
Austin CR (1952) The capacitation of the mammalian sperm. Nature 170:326.
Babcock OF (1983) Examination of the intracellular ionic environment and of
ionophore action by null point measurements employing the fluorescein
chromophore. J. BioI. Chemo258, pp. 6380-6389.
Bailey JL, Storey TB (1994) Calcium influx into mouse spermatozoa activated by
solubilized mouse zona pe/lucida, monitored with calcium fluorescent indicator,
Fluo-3. Inhibition of the influx by three inhibitors of the zona pellucida-induced
acrosome reaction: tyrphostin A48, pertussis toxin and 3-quinuclidinyl enzilate.
Mol Reprod Oev 39, 297-308.
Stellenbosch University http://scholar.sun.ac.za

59
Baldi E, Casano R, Falsetti C, Krausz C, Maggi M & Forti G (1991) Intracellular
calcium accumulation and responsiveness to progesterone in capacitating
human spermatozoa. JAndrol 12, 323-330.
Baldi E, Falsetti C, Krausz C, Gervasi G, Carloni V, CasanoR, Forti G (1993)
Stimulation of platelet-activating factor synthesis by progesterone and A23187
in human spermatozoa. Biochem J 292, 209-216.
Baldi E, Luconi M, Bonaccorsi L & Forti G (1998) Nongenomic effects of
progesterone on spermatozoa: mechanisms of signal transduction and clinical
implications. Front Biosci 3, d1051-1059.
Baldi E, Luconi M, Bonaccorsi L, Muratori M, Forti G (2000) Intracellular events and
signalling pathways involved in sperm acquisition of fertilizing capacity and
acrosome reaction. Front Biosci 5, e110-123.
Bedford JM (1983) Significance of the need for sperm capacitation before fertilization
in eutherian mammals. Bioi Reprod 28:108-1290.
Beltran C, Zapata 0, Darszon A (1996) Membrane potential regulates sea urchin
sperm adenylylcyclase. Biochemistry 35, pp. 7591-7598.
Benoff S (1997) Carbohydrates and fertilization: an overview. Mol. Hum. Reprod 3:
599-637.
Benoff S (1998) Voltage-dependent calcium channels in mammalian spermatozoa.
Front Biosci 3, d1220-d1240.
Benoff S, Hurley IR, Mandel FS, Cooper GW, Hershlag A (1997a) Induction of the
human sperm acrosome reaction with mannose-containing neoglycoprotein
ligands. Mol Hum Reprod 3: 827-837.
Stellenbosch University http://scholar.sun.ac.za

60
Bhatnagar SK, Anand SR (1982) Cyclic nucleotide phosphodiesterase activity in
midpiece and tail of buffalo spermatozoa and its role in sperm motility. Biochem
Biophys Acta 716:133.
Bielfeld PR, Anderson A, Mack SR, De Jonge CJ, Zaneveld LJD (1994) Are
capacitation or calcium ion influx required for the human sperm acrosome
reaction? Fertil Steril62, 1255-1261.
Blackmore PF (1992) Thapsigargin elevates and potentiates the ability of
progesterone to increase intracellular free calcium in human sperm: possible
role of perinuclear calcium. Cell Calcium 14, 53-60.
Blackmore PF, Eisoldt S (1999) The neoglycoprotein mannose-bovine serum
albumin, but not progesterone, activates T-type calcium channels in human
spermatozoa. Mol Hum Reprod 5:498-506.
Blackmore PF, 1mWB, Bleasdale JE (1994) The cell surface progesterone receptor
which stimulates calcium influx in human sperm is unlike the A ring reduced
steroid site in the GABAA receptor/chloride channel. Mol Cell Endocrinol
104:237-243.
Boatman DE, Robbins RS (1991) Bicarbonate: carbon-dioxide regulation of sperm
capacitation, hyperactivated motility, and acrosome reactions. BioI. Reprod. 44,
pp.806-813.
Bonaccorsi L, Krausz C, Pecchioli P, Forti G, Baldi E (1998) Progesterone-stimulated
intracellular calcium increase in human spermatozoa is calcium-indipendent.
Mol Hum Reprod 4,259-268.
Bonaccorsi L, Luconi M, Forti G, Baldi E (1995) Tyrosine kinase inhibition reduces
the plateau phase of the calcium increase in response to progesterone in
human spermatozoa. FEBS Lett 364, 83-86.
Stellenbosch University http://scholar.sun.ac.za

61
Bork P, Tsai J-Y, Silver lM, Saling P, Carballada R, Burks 0, Moore H (1996)
Sperm-egg binding protein or proto-oncogene? Science 271, 1431-1435.
Bostofte E, Serup J, Rebbe H (1983) Relation between spermatozoa motility and
pregnancies obtained during a twenty-year follow-up period spermatozoa
motility and fertility. Andrologia 15(6):682-686.
Bostofte E, Serup J, Rebbe H (1984) Relation between number of immobile
spermatozoa and pregnancies obtained during a twenty-year follow-up period
immobile spermatozoa and fertility. Andrologia 16(2):136-140.
Bray C, Brown JCK, Publicover, S Barratt ClR (1999): Progesterone interaction with
sperm plasma membrane, calcium influx and induction of the acrosome
reaction. Reprod Med Rev 7:81-93.
Breitbart H, Naor Z (1999) Protein kinase in mammalian sperm capacitation and the
acrosome reaction. Rev Reprod 4, 151-159.
Brewis lA, Clayton R, Browes CE, Martin M, Barratt Cl, Hornby OP, Moore HO
(1998) Tyrosine phosphorylation of a 95 kOa protein and induction of the
crosome reaction in human spermatozoa by recombinant human zona pellucida
glycoprotein 3. Mol Hum Reprod 4, 1136-1144.
Brewis lA, Morton IE, Mohammad SN, Browes CE, Moore HO (2000) Measurement
of intracellular calcium concentration and plasma membrane potential in human
spermatozoa using flow cytometry. JAndrol 21, 23849.
Brook PF, lawry, CookelO, Barratt Cl (1996) Measurement of intracellular pH in
human spermatozoa by flow cytometry with the benzo[c]xanthene dye SNAFl-
1: a novel, single excitation, dual emission, molecular probe. Mol Hum Reprod
2, 18-25.
Stellenbosch University http://scholar.sun.ac.za

62
Brooks DE (1983), Epididymal functions and their hormonal regulation. Aust. J. BioI.
Sci. 36, pp. 205-221.
Brown DA, London E (1998) Functions of lipid rafts in biological membranes. Annu.
Rev. Cell Dev. BioI. 14, pp. 111-136.
Buck J, Sinclair ML, Schapal L, Cann MJ, Levin LR (1999) Cytosolic adenylyl cyclase
defines a unique signaling molecule in mammals. Proc. Natl. Acad. Sci. USA
96, pp. 79-84.
Burks OJ, Carballada R, Moore HOM, Saling PM (1995) Interaction of a tyrosine
kinase from human sperm with the zona pellucida at fertilization. Science 269,
83-86.
Camatini M, Casale A (1987) Actin and calmodulin coexist in the equatorial segment
of ejaculated boar sperm Gamete Res 42:311-318.
Carrera A, Moss J, Ning XP, Gerton GL, Tesarik J, Kopf GS, Moss SB (1996)
Regulation of protein tyrosine phosphorylation in human sperm by a
calcium\calmodulin dependent mechanism: identification of a A kinase anchor
proteins as a major substrates for tyrosine phosphorylation. Dev Bioi 180, 284-
296.
Cascieri M, Amann RP, Hammerstedt RH (1976) Adenine nucleotide changes at
initiation of bull sperm motility. J Bioi Chem 251: 787.
Chen Y, Cann MJ, Litvin TN, lourgenko V, Sinclair ML, Levin LR, Buck J (2000)
Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor
[see comments]. Science 289, pp. 625-628.
Choi YH, Toyoda Y (1998) Cyclodextrin removes cholesterol from mouse sperm and
induces capacitation in a protein-free medium. BioI. Reprod. 59, pp. 1328-1333.
Stellenbosch University http://scholar.sun.ac.za

63
Cohen-Dayag A, Eisenbach M (1994) Potential assays for sperm capacitation in
mammals [published errata appear in Am J Physiol 1995 Mar;268(3 Pt
1):section C following table of contents and 1995 Jun;268(6 Pt 3):section C
following table of contents]. Am. J. Physiol. 267, pp. C1167-C1176.
Cohen-Dayag A, Tur-Kaspa I, Dor J, Mashiach S & Eisenbach M (1995) Sperm
capacitation in humans is transient and correlates with chemotactic
responsiveness to follicular factors. Proc Natl Acad Sci USA 92, 11039-11043.
Cross Nl (1994) Phosphatidylcholine enhances the acrosomal responsiveness of
human spermatozoa. JAndrol 15, 484-488.
Cross Nl (1996) Effect of cholesterol and other sterols on human sperm acrosomal
responsiveness. Mol Reprod Dev 45,212-217.
Cross Nl (1998) Role of cholesterol in sperm capacitation. Bioi Reprod 59, 7-11.
Cross Nl (1999) Effect of methyl-beta-cyclodextrin on the acrosomal responsiveness
of human sperm. Mol. Reprod. Dev. 53, pp. 92-98.
Cross Nl, Overstreet JW (1987) Glycoconjugates of the human sperm surface:
distribution and alterations that accompany capacitation in vitro. Gamete Res.
16, pp. 23-35
Cross Nl, Razy-Faulkner P (1997) Control of human sperm intracellular pH bu
cholesterol and its relationship to the response of the acrosome to
progesterone. Bioi Reprod 56, 1169-1174.
Darszon A, Labarea P, Nishigaki T, Espinosa F (1999) Ion channels in sperm
physiology. Physiol Rev 79,481-510.
DasGupta S, Mills Cl, Fraser lR (1993) Ca2+-related changes in the capacitation
state of human spermatozoa assessed by a chlortetracycline fluorescence
assay. J. Reprod. Fertil. 99, pp. 135-143.
Stellenbosch University http://scholar.sun.ac.za

64
Davis BK (1981) Timing of fertilization in mammals: sperm cholesterol/phospholipid
ratio as a determinant of the capacitation interval. Proc. Natl. Acad. Sci. USA
78, pp. 7560-7564.
Davis BK, Byrne R, Bedigian K (1980) Studies on the mechanism of capacitation:
albumin-mediated changes in plasma membrane lipids during in vitro incubation
of rat sperm cells. Proc. Natl. Acad. Sci. USA 77, pp. 1546-1550.
Davis BK, Byrne R, Hungund B (1979) Studies on the mechanism of capacitation. II.
Evidence for lipid transfer between plasma membrane of rat sperm and serum
albumin during capacitation in vitro. Biochim. Biophys. Acta. 558, pp. 257-266.
De Jonge C, Han HL, Lawrie H, Mack SR, Zaneveld LJD (1991a) Modulation of the
human sperm acrosome reaction by effector of adenylate cyclase/cyclic AMP
second messanger pathway. J Exp Zool 258, 113-125.
De Jonge C, Han HL, Mack SR, Zanefeld LJD (1991b) Effect of phorbol diesters,
synthetic diacylglecerols, and a protein kinase C inhibitor on the human sperm
acrosome reaction, J Androl12: 62-70.
De Lamirande E, Harakat A, Gagnon C (1998) Human sperm capacitation induced by
biological fluids and progesterone, but not by NADH or NADPH, is associated
with the production of superoxide anion. J Androl19, 215-225.
De Lamirande E, Leclerc P, Gagnon C (1997) Capacitation as a regulatory event that
primes spermatozoa for the acrosome reaction and fertilization. Mol. Hum.
Reprod. 3, pp. 175-194.
De Lamirande E, Tsai C, Harakat A, Gagnon C (1998) Involvement of reactive
oxygen species in human sperm arcosome reaction induced by A23187,
Iysophosphatidylcholine, and biological fluid ultrafiltrates. JAndrol 19, 585-594.
Stellenbosch University http://scholar.sun.ac.za

65
Devi KU, Jha K, Shivaji S (1999) Plasma membrane-associated protein tyrosine
phosphatase activity in hamster spermatozoa. Mol. Reprod. Dev. 53 pp. 42-50.
Dragileva E, Rubinstein S, Breitbart H (1999) Intracellular Ca2+-Mg2+-ATPase
regulates calcium influx and acrosomal exocytosis in bull and ram spermatozoa.
Bioi Reprod 61,1226-34.
Eddy EM, O'Brien DA (1994) The spermatozoon. In: E. Knobil and J.D. Neill, Editors,
The Physiology of Reproduction 1, Raven Press, New York, pp. 29-77.
Edvinsson A, Bergman B, Steen Y, Nillson S (1983) Characteristics of donor semen
and cervical mucus at the time of conception. Fertil Steril 39:327.
Espinosa F, Lopez-Gonzalez I, Munoz-Garay C, Felix R, De la Vega-Beltran JL, Kopf
GS, Visconti PE, Darszon A (2000) Dual regulation of the T-type Ca2+current
by serum albumin and beta-estradiol in mammalian spermatogenic cells. FEBS
Lett. 475, pp. 251-256.
Falkenstein E, Heck M, Gerdes 0, Grube 0, Christ M, Weigel M, Buddhikot M, Meizei
S, Wehling M (1999) Specific progesterone binding to a membrane protein and
related nongenomie effects on Ca2+-fluxes in sperm. Endocrinology 140, 5999-
6002.
Fênichel P, Hsi BL, Farahifar 0, Donzeau M, Barrier-Delpech N, Yeh CJG (1989)
Evaluation of the sperm acrosome reaction using a monoclonal antibody, GB24,
and the fluorescence activated cell sorter. J Reprod Fert 87:699-706.
Fisher HM, Brewis lA, Barratt CL, Cooke ID, Moore HO (1998) Phosphoinositide 3-
kinase is involved in the induction of the human sperm acrosome reaction
downstream of tyrosine phosphorylation. Mol Hum Reprod 4, 849-855.
Fleming AD, Yanagimachi R (1981) Effects of various lipids on the acrosome reaction
and fertilizing capacity of guinea pig spermatozoa with special reference to the
Stellenbosch University http://scholar.sun.ac.za

66
possible involvement of Iysophospholipids in the acrosome reaction. Gamete
Res 4, 253-273.
Florman HM (1994) Sequential focal and global elevations of sperm intracellular Ca2+
are initiated by the zona pellucida during acrosomal exocytosis. Dev Bioi 165,
152-164.
Florman HM, Arnouit C, Kazam lG, Li C, O'Toole CM (1998) A perspective on the
control of mammalian fertilization by egg-activated ion channels in sperm: a tale
of two channels. BioI. Reprod. 59, pp. 12-16.
Florman HM, Babcock DF (1991) Progress towards understanding the molecular
basis of capacitation. In: Wassarman, P.M. (Ed.), Chemistry of Fertilization.
CRC Press, Boca Raton, FL, pp. 105-132.
Florman HM, Corron ME, Kim TD, Babcock DF (1992) Activation of voltage-
dependent calcium channels of mammalian sperm is required for zona
pellucida-induced acrosomal exocytosis. Dev. BioI. 152, pp. 304-314.
Florman HM, Tombes RM, First NL, Babcock DF (1989) An adhesion-associated
agonist from the zona pellucida activates G protein-promoted elevations of
internal Ca and pH that mediate mammalian sperm acrosomal exocytosis. Dev
Bioi 135, 133-146.
Foresta C, Rossato M, Di Virgilio F (1993) Extracellular ATP is a trigger for the
acrosome reaction in human spermatozoa. J Bioi Chem 267, 19443-19447.
Foster JA, Friday BB, Maulif MT, Blobel C, Winfrey VP, Olson GE, Kim K-S, Gerton
GL (1997) J Bioi Chem 272: 12714-12722.
Francavilla F, Santucci R, Macerola B, Ruvolo G, Romano R (2000) Nitric oxide
synthase inhibition in human sperm affects sperm-oocyte fusion but not zona
pellucida binding. Bioi Reprod 63, 425-429.
Stellenbosch University http://scholar.sun.ac.za

67
Franken DR, Kruger TF, Lombard SC, Oehninger SC, Acosta AA, Hodgen GD (1993)
The ability of the hemizona assay to predict human fertilization in vitro in
different IVF/GIFT cycles. Hum Reprod 8: 1240-1244.
Franken DR, Morales PJ, Habenicht UF (1996) Inhibition of G protein in human
sperm and its influence on acrosome reaction and zona pellucida binding. Fertil
Steril. 66(6):1009-11.
Fraser L (1995a) Cellular biology of capacitation and he acrosome reaction. In:
Modern Andrology. Ombelet W, Vereecken A (eds) Human Reproduction Vol10
Suppl 1: 22-30.
Fraser LR (1995b) Mechanisms regulating capacitation and the acrosome reaction.
In: Human sperm acrosome reaction. Eds: Fenichel P., Parinaud J.,
Colloque/INSERIM John Libbey Eurotext Ltd, Montrouge, France, vol. 236, 17-
33.
Fraser LR (1998) Role of fertilization promoting peptide (FPP) in modulating
mammalian sperm function. Front Biosci 3, D1187-191.
Fraser LR (1999) New insights into possible causes of male infertility. Human.
Reprod 14,38-46.
Fraser LR, McDermott CA (1992) Ca2+-related changes in the mouse sperm
capacitation state: a possible role for Ca2+-ATPase. J Reprod Fertil 96, 363-
377.
Fukuda A, Roudebusk WE, Thatcher SS (1994) Platelet-activating factor enhances
the acrosome reaction, fertilization in vitro by subzonal sperm injection and
resulting embryonic development in the rabbit. Human Reprod 9, 94-99.
Funahashi H, Asano A, Fujiwara T, Nagai T, Niwa K & Fraser LR (2000) Both
fertilization promoting peptide and adenosine stimulate capacitation but inhibit
Stellenbosch University http://scholar.sun.ac.za

68
spontaneous acrosome loss in ejaculated boar spermatozoa in vitro. Mol
Reprod Dev 55, 117-124.
Furuya S, Endo Y, Oba M, Suzuki S (1993) Effects of modulators of protein kinase C
on human sperm capacitation. Fertil Steril 59, 1285-1290.
GadelIa BM, Harrison RAP (2000) The capacitating agent bicarbonate induces
protein kinase A-dependent changes in phospholipid transbilayer behavior in
the sperm plasma membrane. Development 127, 2407-2420.
Galantino-Homer HL, Visconti PE, Kopf GS (1997) Regulation of protein tyrosine
phosphorylation during bovine sperm capacitation by a cyclic adenosine 3'5'-
monophosphate-dependent pathway. BioI. Reprod. 56, pp. 707-719.
Garbers DL, Kopf GS (1980) The regulation of spermatozoa by calcium and cyclic
nucleotides. Adv. Cyclic Nucleotide Res. 13, pp. 251-306.
Garbers DL, Lust WD, First NL, Lardy HA (1971) Effects of phosphodiesterase
inhibitors and cyclic nucleotides on sperm respiration and motility. Biochemistry
10:1825.
Garcia MA, Meizei S (1996) Importance of sodium ion to the progesterone-initiated
acrosome reaction in human sperm. Mol Reprod Dev 45, 513-520.
Garcia MA, Meizei S (1999a) Progesterone-mediated calcium influx and acrosome
reaction of human spermatozoa: pharmacological investigation of T-type
calcium channels. Bioi Reprod 60, 102-9.
Garcia MA, Meizei S (1999b) Regulation of intracellular pH in capacitated human
spermatozoa by a Na+/H+ exchanger. Mol Reprod Dev 52, 189-195.
Garde J, Roldan ER (2000) Stimulation of Ca2+-dependent exocytosis of the sperm
acrosome by cAMP acting downstream of phospholipase A2. J. Reprod. Fertil.
118, pp. 57-68.
Stellenbosch University http://scholar.sun.ac.za

69
Garty NB, Salomon Y (1987) Stimulation of partially purified adenylate cyclase from
bull sperm by bicarbonate. FEBS Lett. 218 pp. 148-152.
Gatti JL, Billard R, Christen R (1990) Ionic regulation of the plasma membrane
potential of rainbow trout (Salmo gairdneri) spermatozoa: role in the initiation of
sperm motility. J. Cell Physiol. 143, pp. 546-554.
Gauss R, Seifert R, Kaupp UB (1998) Molecular identification of a hyperpolarization-
activated channel in sea urchin sperm. Nature 393, pp. 583587.
Gearon CM, Mortimer D, Chapman MG, Forman RG (1994) Artificial induction of the
acrosome reaction in human spermatozoa. Hum Reprod 9: 77-82.
Go KJ, Wolf DP, (1985) Albumin-mediated changes in sperm sterol content during
capacitation. BioI. Reprod. 32, pp. 145-153.
Gross MK, Toscano DG, Toscano WA (1987) Calmodulin-mediated adenylate
cyclase from mammalian sperm. J. BioI. Chemo262, pp. 8672-8676.
Harrison DA, Meizei S (2000) Involvement of protein kinase A and A kinase
anchoring protein in the progesterone-initiated human sperm acrosome
reaction. Bioi Reprod 62,811-20.
Harrison RA (1996) Capacitation mechanisms, and the role of capacitation as seen in
eutherian mammals. Reprod. Fertil. Dev. 8, pp. 581-594.
Harrison RA, Miller NG (2000) cAMP-dependent protein kinase control of plasma
membrane lipid architecture in boar sperm. Mol Reprod Dev 55,220-228.
Harrison RAP, GadelIa BM (1995) Membrane changes during capacitation with
special reference to lipid architecture. In: Human sperm acrosome reaction.
Eds: Fenichel P., Parinaud J., Colloque INSERIM/John Libbey Eurotext Ltd,
Montrouge, France, vol. 236, 45-65.
Stellenbosch University http://scholar.sun.ac.za

70
Herness EA, Naz RK (1999) Presence and tyrosine phosphorylation of c-met
receptor in human sperm. JAndrol 20, 640-647.
Herrero MB, de Lamirande E, Gagnon C (1999) Nitric oxide regulates human sperm
capacitation and protein-tyrosine phosphorylation in vitro. Bioi Reprod 61, 575-
581.
Hildebrandt JD, Codina J, Tash JS, Kirchick HJ, Lipschultz L, Sekura RO,
Birnbaumer L (1985) The membrane-bound spermatozoal adenylyl cyclase
system does not share coupling characteristics with somatic cell adenylyl
cyclases. Endocrinology 116, pp. 1357-1366.
Holappa K, Mustonen M, Parvinen M, Vihko P, Rajaniemi H, Kellokumpu SP (1999)
Primary structure of a sperm cell anion exchanger and its messenger
ribonucleic acid expression during spermatogenesis. Bioi Reprod 61,981-986.
Horowitz JA, Wasco W, Leiser M (1988) Interaction of the regulatory subunit of type II
cAMP dependant protein kinase with mammalian sperm flagellum. J Bioi Chem
263:2098.
Hoshi K, Aita T, Yanagida K, Yoshimatsu N, Sato A (1990) Variation in the
cholesterol/phospholipid ratio in human spermatozoa and its relationship with
capacitation. Hum. Reprod. 5, pp. 71-74.
Hoskins DO (1973) Adenosine nucleotide mediation of fructolysis and motility in
bovine epididymal spermatozoa. J Bioi Chem 248:1135.
Hoskins DO, Casillas ER (1975) Functions of cyclic nucleotides in mammalian
spermatozoa. In: Handbook of physiology, Section 7, Vol 5. Geiger S (eds)
Williams and Wilkins Company, Baltimore, pp453.
Hoskins DO, Stephen OT, Hall ML (1974) cAMP and protein kinase in developing
bovine spermatozoa. J Reprod Fertil 37:131.
Stellenbosch University http://scholar.sun.ac.za

71
Huo LJ, Yang ZM (2000) Effects of platelet activating factor on capacitation and
acrosome reaction in mouse spermatozoa. Mol Reprod Dev 56, 436-440.
Hyne RV, Edwards KP, Smith JD (1985) Changes in guinea pig sperm intracellular
sodium and potassium content during capacitation and treatment with
monovalent ionophore. Gamete Res 12, 65-73.
Ishikawa H, Tomomasa H, Yoshii, Koisa K, Tajima Y, Okamura N, Sugita Y (1989)
Correlation between the sperm motility and adenylate cyclase activity in infertile
men. Andrologia 21 :437.
Jahn R, Sudhof TC (1999) Membrane fusion and exocytosis. Annu. Rev. Biochem.
68, pp. 863-911.
Jha K, Shivaji S (2001) Capacitation-associated changes in protein tyrosine
phosphorylation, hyperactivation and acrosome reaction in hamster
spermatozoa. Andrologia 33 pp. 95-104.
Kabouridis PS, Magee AI, Ley SC (1997) S-acylation of LCK protein tyrosine kinase
is essential for its signalling function in T lymphocytes. Embo. J. 16, pp. 4983-
4998.
Kalab P, Peknicova J, Geussova G, Moos J (1998) Regulation of protein tyrosine
phosphorylation in boar sperm through a cAMP-dependent pathway. Mol.
Reprod. Dev. 51, pp. 304-314.
Katz OF, Drobnis EZ (1990) In: Fertilization in mammals. Bavister BO, Cummins J,
Roldan ERS (eds) Serono Symposium, Norwell, MA 125-137.
Kawasaki H, Springett GM, Mochizuki N, Toki S, Nakaya M, Matsuda M, Housman
DE, Graybiel AM (1998) A family of cAMP-binding proteins that directly activate
Rap1. Science 282, pp. 2275-2279.
Stellenbosch University http://scholar.sun.ac.za

72
Khorasani AM, Cheung AP & Lee CY (2000) Cholesterol inhibitory effects on human
sperm-induced acrosome reaction. JAndrol 21, 586-594.
Kirkman-Brown JC, Bray C, Stewart PM, Barratt CLR, Publicover SJ (2000) Biphasic
elevation of [Ca2+]i in individual human spermatozoa exposed to progesterone.
Dev Bioi 222, 326-335.
Kobori H, Miyazaki S, Kuwabara Y (2000) Characterization of intracellular Ca2+
increase in response to progesterone and cyclic nucleotides in mouse
spermatozoa. Bioi Reprod 63, 113-120.
Krausz C, Gervasi G, Forti G, Baldi E (1994) Effect of platelet-activating factor on
motility and acrosome reaction of human spermatozoa. Human Reprod 9, 471-
476.
Kumar R, Harper MJK, Hanahan DJ (1988) Occurrence of platelet-activating factor in
rabbit spermatozoa. Arch Biochem Biophys 260, 497-502.
Kyono K, Hoshi K, Saito A, Tsuiki A, Hoshiai H, Suzuki M (1984) Effects of
phospholipase A2, Iysophosphatidylcholine, and fatty acid on the acrosome
reaction of human spermatozoa. J Exp Med 144, 257-263.
Langlais J,Roberts JD (1985) A molecular membrane model of sperm capacitation
and the acrosome reaction of mammalian spermatozoa. Gamete Res. 12, pp.
183-224.
Leclerc P, de Lamirande E, Gagnon C (1996) Cyclic adenosine 3',5'monophosphate-
dependent regulation of protein tyrosine phosphorylation in relation to human
sperm capacitation and motility. BioI. Reprod. 55, pp. 684-692.
Leclerc P, de Lamirande E, Gagnon C (1998) Interaction between Ca2+, cyclic 3',5'
adenosine monophosphate, the superoxide anion, and tyrosine phosphorylation
pathways in the regulation of human sperm capacitation. JAndrol 19, 434-443.
Stellenbosch University http://scholar.sun.ac.za

73
Leclerc P, Kopf GS (1995) Mouse sperm adenylyl cyclase: general properties and
regulation by the zona pellucida. BioI. Reprod. 52, pp. 1227-1233.
Lee DR, Lee JE, Yoon HS, Roh SI (1997) Induction of acrosome reaction in human
spermatozoa accelerates the time of pronucleus formation of hamster oocytes
after intracytoplasmatic sperm injection. Fertil Steril 67,315-320.
Lee MA, Kopf GS, Storey BT (1987) Effects of phorbol esters and a diacylglycerol on
the mouse sperm acrosome reaction induced by the zona pellucida. Bioi
Reprod 36: 617-627.
Lee MA, Storey BT (1986) Bicarbonate is essential for fertilization of mouse eggs:
mouse sperm require it to undergo the acrosome reaction. BioI. Reprod. 34, pp.
349-356.
Leyton L, LeGuen P, Bunch 0, Saling PM (1992) Regulation of mouse gamete
interaction by sperm tyrosine kinase. Proe Natl Acad Sci USA 89, 11692-11695
Leyton L, Saling P (1989a): 95 kd sperm proteins bind ZP3 and serve as tyrosine
kinase substrates in response to zona binding. Cell 57, 1123-1130.
Leyton L, Saling PM (1989b) Evidence that aggregation of mouse sperm receptors by
ZP3 triggers the acrosome reaction. J Cell Bio1108, 2163-2168.
Lievano A, Santi CM, Serrano CJ, Trevino CL, Bellve AR, Hernandez-Cruz A,
Darszon A (1996) T-type Ca2+ channels and alpha1E expression in
spermatogenic cells, and their possible relevance to the sperm acrosome
reaction. FEBS Lett. 388, pp. 150-154.
Lindeman CB, Kanous KS (1989) Regulation of sperm motility. Arch Androl 23:1.
Liu DY, Baker HW (1997) Protein kinase C plays an important role in he human zona
pellucida -induced acrosome reaction. Mol Hum Reprod 3, 1037-1043.
Stellenbosch University http://scholar.sun.ac.za

74
Llanos MN, Meizei S (1983) Phospholipid methylation increases during capacitation
of golden hamster sperm in vitro. Bioi Reprod 28, 1043-1051.
Loeb J (1915) On the nature of conditions which determine or prevent entrance of the
spermatozoa into the egg. Amer Nat 49:257-85.
Luconi M, Barni T, Vannelli GB, Krausz C, Marra F, Benedetti PA, Evangelista V,
Francavilla S, Properzi G, Forti G, Baldi E (1998) Extracellular-signal regulated
kinases (ERKs) modulate capacitation of human spermatozoa. Bioi Reprod 58,
1476-1489.
Luconi M, Bonaccorsi L, Krausz C, Gervasi G, Forti G, Baldi E (1995) Stimulation of
protein tyrosine phosphorylation by platelet-activating factor and progesterone
in human spermatozoa. Mol Cell Endocrinol 108, 35-42.
Luconi M, Bonaccorsi L, Maggi M, Pecchioli P, Krausz C, Forti G, Baldi E (1998)
Identification and characterization of functional nongenomic progesterone
receptors on human sperm membrane. J Clin Endocrinol Metab 83, 877-890.
Luconi M, Krausz C, Barni T, Vannelli GB, Forti G, Baldi E (1998) Progesterone
stimulates p42 extracellular signal-regulated kinase (p42erk) in human
spermatozoa. Mol Hum Reprod 4,251-258.
Luconi M, Krausz C, Forti G, Baldi E (1996) Extracellular calcium negatively
modulates tyrosine phosphorylation and tyrosine kinase activity during
capacitation of human spermatozoa. Bioi Reprod 55, 207-216.
Luconi M, Muratori M, Forti G, Baldi E (1999) Identification and characterization of a
novel functional estrogen receptor on human sperm membrane which interferes
with progesterone effects. J Clin Endocrin Metab 84,1670-1678.
Stellenbosch University http://scholar.sun.ac.za

75
Luconi M, Muratori M, Maggi M, Pecchioli P, Peri A, Mancini M, Filimberti E, Forti G &
Baldi E (2000) Uteroglobin and transglutaminase modulate human sperm
functions. JAndrol 21, 676-688.
Mandai A, Naaby-Hansen S, Wolkowicz MJ, Klotz K, Shetty J, Retief JD, Coonrod
SA, Kinter M, Sherman N, Cesar F, Flickinger CJ, Herr JC (1999) FSP95, a
testis-specific 95-kilodalton fibrous sheath antigen that undergoes tyrosine
phosphorylation in capacitated human spermatozoa. BioI. Reprod. 61, pp. 1184-
1197.
Meizei S, Turner KO (1993) Initiation of the sperm acrosome reaction by
thapsigargin. J Exp Zool 267:350-355.
Melendrez CS, Meizei S, Berger T (1994) Comparison of the ability of progesterone
and heat solubilized porcine zona pellucida to initiate the porcine perm
acrosome reaction in vitro. Mol Reprod Dev 39: 433-438.
Mendoza, Solar A, Tesarik J (1995) Nongenomie steroid action: independent
targeting of a plasma membrane calcium channel and a tyrosine kinase.
Biochem Biophysic Res Comm 210: 518-523.
Michaut M, Tomes CN, De Bias G, Yunes R, Mayorga LS (2000) Calcium-triggered
acrosomal exocytosis in human spermatozoa requires the coordinated
activation of Rab3A and N-ethylmaleimide-sensitive factor. Proc.Natl. Acad. Sci.
USA 97, pp. 9996-10001.
Minhas BS (1993) Platelet activating factor treatment of human spermatozoa
enhances fertilization potential. Am JObstet Gynecol 168, 1314-1317.
Minhas BS, Ripps BA, Zhu YP, Kim HN, Burwinkel TH, Gleicher N (1996) Platelet
activating factor and conception. Am J Reprod Immunol 35, 267-271.
Stellenbosch University http://scholar.sun.ac.za

76
Mohri H, Yanagimachi R (1980) Characteristics of motor apparatus in testicular,
epididymal and ejaculated spermatozoa. A study using demembranated sperm
models. Exp Cell Res 127(1):191-196.
Morisawa M, Ishida K (1987) Short-term changes in levels of cyclic AMP, adenylate
cyclase, and phosphodiesterase during the initiation of sperm motility in rainbow
trout. J. Exp. Zool. 242, pp. 199-204.
Morisawa M, Suzuki K (1980) Osmolality and potassium ion: their roles in initiation of
sperm motility in teleosts. Science 210, pp. 1145-1147.
Mortimer D (1990) Objective analysis of sperm motility and kinematics. In: The CRC
Handbook of Laboratory Diagnosis and Treatment of Infertility. pp
Morton BJ, Harrigan-Lum J, Albagii L, Jooss T (1974) The activation of motility in
quiescent hamsterspermatozoa from the epididymis by calcium and cyclic
nucleotides. Biochem Biophys Res Commun 56:372.
Moss SB, Turner SMO, Burkert KL, VanScoy Butt H, Gerton GL (1999) Conservation
and function of a bovine sperm A-kinase anchor protein homologous to mouse
AKAP82. Bioi Reprod 61,335-342.
Naz RK, Ahmad K (1992) Presence of expression products of c-erbB-1 and c-erbB-
2/HER2 genes on mammalian sperm cell, and effects of their regulation on
fertilization. J Reprod Immunol 21, 223-239.
Naz RK, Ahmad K, Kaplan P (1992) Expression and function of ras-proto-oncogene
proteins in human sperm cells. J Cell Sci 102, 487-494.
Naz RK, Ahmad K, Kaplan P (1993) Involvement of cyclins and cdc2 serine/threonine
protein kinase in human sperm cell function. Bioi Reprod 48, 720-728.
Naz RK, Ahmad K, Kumar R (1991) Role of membrane phosphotyrosine proteins in
human spermatozoa function. J Cell Sci 99, 157-165.
Stellenbosch University http://scholar.sun.ac.za

77
Neill JM, Olds-Clarke P (1987) A computer-assisted assay for mouse sperm
hyperactivation demonstrates that bicarbonate but not bovine serum albumin is
required. Gamete Res. 18, pp. 121-140.
Nikolopoulou M, Soucek DA, Vary JC (1986) Modulation of the lipid composition of
boar sperm plasma membranes during an acrosome reaction in vitro. Arch
Biochem Biophys 250,30-37.
Okamura N, Tajima Y, Soejima A, Masuda H, Sugita Y (1985) Sodium bicarbonate in
seminal plasma stimulates the motility of mammalian spermatozoa through
direct activation of adenylate cyclase. J. BioI. Chemo260, pp. 9699-9705.
Okamura N, Tajima Y, Sugita Y (1988) Decrease in bicarbonate transport activities
during epididymal maturation of porcine sperm. Biochem. Biophys. Res.
Commun. 157, pp. 1280-1287.
Olson GE, Winfrey WP, Winter MA, Davenport GR (1987) Outer acrosomal
membrane of guinea pig spermatozoa: isolation and structural characterization.
Gamete Res 17:77-94.
Osheroff JE, Visconti PE, Valenzuela JP, Travis AJ, Alvarez J, Kopf GS (1999)
Regulation of human sperm capacitation by a cholesterol efflux-stimulated
signal transduction pathway leading to protein kinase A-mediated up-regulation
of protein tyrosine phosphorylation. Mol Hum Reprod 5, 1017-1026.
O'Toole CMB, Arnouit C, Darszon A, Steinhardt RA, Florman HM (2000) Ca2+ entry
through store-operated channels in mouse sperm is initiated by egg ZP3 and
drives the acrosome reaction. Mol Bioi Cell 11, 1571-1584.
O'Toole CMB, Roldan ERS, Fraser LR (1996) Protein kinase C activation during
progesterone-stimulated acrosomal exocytosis in human spermatozoa. Mol
Hum Reprod 2,921-927.
Stellenbosch University http://scholar.sun.ac.za

78
Owen DH, Katz DF (1993) Sampling factors influencing accuracy of sperm kinematic
analysis. J Androl14: 210.
Parinaud J, Milhet P (1996) Progesterone induces Ca++-dependent 3',5'-cyclic
adenosine-monophosphate increase in human sperm. J Clin Endocrinol Metab
81: 1357-1360.
Pariset CC, Roussel C Weinman SJ, Demaille JG (1983) Calmodulin intracellular
concentration and cAMP dependant proetin kinase activity in human sperm
samples in relation to sperm morphology and motility. Gamete Res 8:171.
Patrat C, Serres C, Jouannet P (2000) Induction of a sodium ion influx by
progesterone in human spermatozoa. Bioi Reprod 62, 1380-1386.
Pawson T, Scott JD (1997) Signaling through scaffold, anchoring, and adaptor
proteins. Science 278, pp. 2075-2080.
Ramalho-Santos J, Moreno RD, Sutovsky P, Chan AW, Hewitson L, Wessel GM,
Simerly CR, Schatten G (2000) SNAREs in mammalian sperm: possible
implications for fertilization. Dev. BioI. 223, pp. 54-69.
Reyes A, Goicoechea B, Rosado A (1977) In vitro capacitation of mammalian
spermatozoa by the calcium A23187. Fertil Steril 28:356.
Richter W, Dettmer D, Glander H (1999) Detection of mRNA transcripts of cyclic
nucleotide phosphodiesterase subtypes in ejaculated human spermatozoa. Mol
Hum Reprod 5,732-736.
Rochwerger L, Cuasnicu PS (1992) Redistribution of a rat sperm epididymal
glycoprotein after in vitro and in vivo capacitation. Mol. Reprod. Dev. 31, pp. 34-
41.
Roldan ER (1998) Role of phospholipases during sperm acrosomal exocytosis. Front
Biosci 3, D1109-1119.
Stellenbosch University http://scholar.sun.ac.za

79
Roldan ERS, Murase T, Shi Q-X (1994) Exocytosis in spermatozoa in response to
progesterone and zona pellucida. Science 266: 1578-1581.
Rosado AJ, Hicks JJ, Reyes A, Bianco I (1974) Capacitation in vitro of rabbit
spermatozoa with cyclic adenosine monophosphate and human follicular fluid.
Fertil Steril 25: 821.
Rotem G, Paz F, Homonnai ZT, Kalina M, Lax J, Breitbart H, Naor Z (1992) Ca2+-
independent induction of acrosome reaction by protein kinase C in human
sperm. Endocrinology 131, 2235-2243.
Roy S, Luetterforst R, Harding A, Apolloni A, Etheridge M, Stang E, Rolls B, Hancock
JF, Parton RG (1999) Dominant-negative caveolin inhibits H-Ras function by
disrupting cholesterol-rich plasma membrane domains [see comments]. Nat.
Cell BioI. 1, pp. 98-105.
Saling PM (1989) Mammalian sperm interaction with extracellular matrices of the
egg. In: Oxford Review of Reproductive Biology, Ed: Milligan S.R., Oxford:
Oxford University Press; pages:339-388.
Schaefer M, Hofmann T, Schiltz G, Gudermann T (1988) A new prostaglandin E
receptor mediates calcium influx and acrosome reaction in human
spermatozoa. Proc Natl Acad Sci USA 95, 3008-3013
Schulz JR, Sasaki JD, Vacquier VD (1998) Increased association of synaptosome-
associated protein of 25 kDa with syntaxin and vesicle-associated membrane
protein following acrosomal exocytosis of sea urchin sperm. J. BioI. Chemo273,
pp.24355-24359.
Schulz JR, Wessel GM, Vacquier VD (1997) The exocytosis regulatory proteins
syntaxin and VAMP are shed from sea urchin sperm during the acrosome
reaction. Dev. BioI. 191, pp. 80-87.
Stellenbosch University http://scholar.sun.ac.za

80
Sengoku K, Ishikawa M, Tamate K, Shimizu T (1992) Effects of platelet activating
factor on mouse sperm function. JAssist Reprod Genet 9:447-453.
Setchell BP, Maddocks S, Brooks DE (1994) Anatomy, vasculature, innervation, and
fluids of the male reproductive tract. In: E. Knobil and J.D. Neill, Editors, The
Physiology of Reproduction 1, Raven Press, New York, pp. 1063-1175.
Shabb JB, Corbin JD (1992) Cyclic nucleotide-binding domains in proteins having
diverse functions. J Bioi Chem 257(9): 5723-5726.
Shalgi R, Matityahu A, Gaunt SJ, Jones R (1990) Antigens on rat spermatozoa with a
potential role in fertilization. Mol Reprod Dev 25(3): 286-296.
Shi Q-X, Roldan ERS (1995) Evidence that a GBAA-like receptor is involved in
progesterone-induced acrosomal exocytosis in mouse spermatozoa. Bioi
Reprod 52:373-381.
Shi ax, Yuan Y-Y, Roldan RS (1997) y-aminobutyric acid (GABA) induces the
acrosome reaction in human spermatozoa. Mol Hum Reprod 3: 677-683.
Si Y, Olds-Clarke P (2000) Evidence for the involvement of calmodulin in mouse
sperm capacitation. Bioi Reprod 62, 1231-1239.
Sinclair ML, Wang XY, Mattia M, Conti M, Buck J, Wolgemuth DJ, Levin LR (2000)
Specific expression of soluble adenylyl cyclase in male germ cells. Mol Reprod
Dev 56,6-11.
Snyder DR, Clegg ED (1975) Alteration of phospholipids in porcine spermatozoa
during in vivo uterus and oviduct incubation. J Anim Sci 40,269-274.
Spira B, Breitbart H (1992), The role of anion channels in the mechanism of
acrosome reaction in bull spermatozoa. Biochim. Biophys. Acta. 1109, pp. 65-
73.
Suarez SS (1996) Hyperactivated motility in sperm. J. Androl. 17, pp. 331-335.
Stellenbosch University http://scholar.sun.ac.za

81
Suzuki F, Yanagimachi (1989) Changes in the distribution of intramembranous
particles and filipin-reactive membrane sterols during in vitro capacitation of
golden hamster spermatozoa. Gamete Res. 23, pp. 335-347.
Talbot P, Kleve MG (1978) Hamster sperm cross react with antiactin. J Exp Zool 204:
131-136.
Tash JS, Kakar SS, Means AR (1984) Flagellar motility requires the cAMP
dependant phosphorylation of a heat stable NP-40 soluble 56kd protein,
axokinin. Cell 38:551.
Tash JS, Means AR (1982) Regulation of protein phosphorylation by cyclic
adenosine monophosphate and calcium. Bioi Reprod 26:745.
Tash JS, Means AR (1983) Cyclic adenosine 3',5'monophosphate, calcium and
protein phosphorylation in flagellar movement. Bioi Reprod 28:75.
Tesarik J, Carreras A, Mendoza C (1996) Single cell analysis of tyrosine kinase
dependent and independent Ca2+ fluxes in progesterone induced acrosome
reaction. Mol Hum Reprod 2,225-232.
Tesarik J, Flechon JE (1986) Distribution of sterols and anionic lipids in human sperm
plasma membrane: effects of in vitro capacitation. J. Ultrastruct. Mol. Struct.
Res. 97, pp. 227-237.
Tesarik J, Mendoza C (1993) Insights into the function of sperm surface
progesterone receptor: Evidence of ligand induced receptor aggregation and
implication of proteolysis. Exp Cell Res 205, 111-117.
Tesarik J, Moos J, Mendoza C (1993) Stimulation of protein tyrosine phosphorylation
by a progesterone receptor on the cell surface of human sperm. Endocrinol 133,
328-335.
Stellenbosch University http://scholar.sun.ac.za

82
Therien I, Soubeyrand S, Manjunath P (1997) Major proteins of bovine seminal
plasma modulate sperm capacitation by high-density lipoprotein. BioI. Reprod.
57, pp. 1080-1088.
Tomes CN, McMaster CR, Saling PM (1996) Activation of mouse PIP2-PLC by zona
pellucida is modulated by tyrosine phosphorylation. Mol Reprod Dev 43, 196-
204.
Travis AJ, Vargas LA, Merdiushev T, Moss SB, Hunnicutt GR, Kopf GS (2000)
Expression and localization of caveolin-1 in mouse and guinea pig
spermatozoa. Mol. BioI. Cell. 11, P 121a.
Tulsiani DRP, Abou-Halla A, Loeser CR, Pereira BMJ (1998) The biological and
functional significance of the sperm acrosome and acrosomal enzymes in
mammalian fertilization. Exp Cell Res 240: 151-164.
Uguz C, Vredenburgh WL, Parrish JJ (1994) Heparin-induced capacitation but not
intracellular alkalinization of bovine sperm is inhibited by Rp-adenosine-3',5'-
cyclic monophosphorothioate. BioI. Reprod. 51, pp. 1031-1039.
Vijayaraghavan S, Liberty GA, Mohan J, Winfrey VP, Olson GE, Carr OW (1999)
Isolation and moleculatr characterization of AKAP110, a novel, sperm-specific
protein kinase A-anchoring protein. Mol Endocrinol13, 705-717.
Virtanen I, Bradley RA, Paasivuo R, Lehto VP (1984) J Cell Bioi 99:1083-1091.
Visconti PE, Bailey JL, Moore GD, Pan 0, aids-Clarke P, Kopf GS (1995a)
Capacitation of mouse spermatozoa: I. Correlation between the capacitation
state and protein tyrosine phosphorylation. Development. 121, pp. 1129-1137.
Visconti PE, Galantino-Homer H, Moore GD, Bailey JL, Ning X, Fornes M, Kopf GS
(1998) The molecular basis of sperm capacitation. J. Androl. 19, pp. 242-248.
Stellenbosch University http://scholar.sun.ac.za

83
Visconti PE, Galantino-Homer H, Ning X, Moore GD, Valenzuela JP, Jorgez CJ,
Alvarez JG, Kopf GS (1999a) Cholesterol efflux-mediated signal transduction in
mammalian sperm: beta-cyclodextrins initiate transmembrane signaling leading
to an increase in protein tyrosine phosphorylation and capacitation. J. BioI.
Chemo274, pp. 3235-3242.
Visconti PE, Johnson LR, Oyaski M, Fornes M, Moss SB, Gerton GL, Kopf GS (1997)
Regulation, localization, and anchoring of protein kinase A subunits during
mouse sperm capacitation. Dev. BioI. 192, pp. 351-363.
Visconti PE, Kopf GE (1998) Regulation of protein phosphorylation during sperm
capacitation. BioI. Reprod. 59, pp. 1-6.
Visconti PE, Moore GD, Bailey JL, Leclerc P, Connors SA, Pan D, Olds-Clarke P,
Kopf GS (1995b) Capacitation of mouse spermatozoa: II. Protein tyrosine
phosphorylation and capacitation are regulated by a cAMP-dependent pathway.
Development 121, pp. 1139-1150.
Visconti PE, Muschietti JP, Flawia MM, Tezon JG (1990) Bicarbonate dependence of
cAMP accumulation induced by phorbol esters in hamster spermatozoa.
Biochim Biophys Acta 1054, 231-236.
Visconti PE, Ning X, Fornes MW, Alvarez JG, Stein P, Connors SA, Kopf GS (1999b)
Cholesterol efflux-mediated signal transduction in mammalian sperm:
cholesterol release signals an increase in protein tyrosine phosphorylation
during mouse sperm capacitation. Dev. BioI. 214, pp. 429-443.
Visconti PE, Stewart-Savage J, Blasco A, Battaglia L, Miranda P, Kopf GS, Tezon JG
(1999c) Roles of bicarbonate, cAMP, and tyrosine phosphorylation on
capacitation and the spontaneous acrosome reaction of hamster sperm. Bioi
Reprod 61, 76-84.
Stellenbosch University http://scholar.sun.ac.za

84
Visconti PE, Westbrook VA, Chertihin 0 Demarco 0 Sleight S, Diekman AB (2002)
Novel signalling pathways involved in sperm acquisition of fertilizing capacity. J
Reprod Immun 53:133-150.
Vorup-Jensen T, Hjort T, Abraham-Peskir JV, Guttmann P, Jensenius JC, Uggerhoj
E & Medenwaldt: R (1999) X-ray microscopy of human spermatozoa shows
change of mitochondrial morphology after capacitation. Hum Reprod 14, 880-
884.
Vredenburgh-Wilberg WL, Parrish JJ (1995) Intracellular pH of bovine sperm
increases during capacitation. Mol Reprod Dev 40, 490-592.
Walensky LD, Snyder SH (1995) Inositol 1,4,5-trisphosphate receptors selectively
localized to the acrosomes of mammalian sperm. J Cell Bioi 130, 857-869.
Wasco WM, Orr GS (1984) Function of calmodulin in mammalian sperm: presence of
a calmodulin-dependent cyclic nucleotide phosphodiesterase associated with
demembranated rat caudal epididymal sperm. Biochem. Biophys. Res.
Commun. 118, pp. 636-642.
Wassarman PM (1999) Mammalian fertilization: molecular aspects of gamete
adhesion, exocytosis, and fusion. Cell96, 175-183.
Watson PF, Plummer JM, Jones PS, Bredl JCS (1995) Localization of intracellular
calcium during the acrosome reaction in ram spermatozoa. Mol Reprod Dev 41,
513-520.
Weyand I, Godde M, Frings S, Weiner J, Muller F, Altenhofen W, Hatt H, Kaupp UB
(1994) Cloning and functional expression of a cyclic-nucleotide-gated channel
from mammalian sperm. Nature 368, pp. 859-863.
Stellenbosch University http://scholar.sun.ac.za

85
White DR, Aitken RJ (1989) Relationship between calcium, cyclic AMP, ATP and
intracellular pH and the capacity of hamster spermatozoa to express
hyperactivated motility. Gamete Res 22, 163-177.
Wistrom CA, Meizei S (1993) Evidence suggesting involvement of a unique human
steroid receptor/Ct channel complex in the progesterone initiated acrosome
reaction. Oev Bioi 159:679-690.
Wolf DE, Cardullo RA (1991) Physiological properties of the mammalian sperm
plasma membrane. In: B. Baccetti, Editor, Comparative Spermatology 20 Years
After, Raven Press, New York, pp. 599-604.
Wolf DE, Hagopian SS, Ishijima S (1986) Changes in sperm plasma membrane lipid
diffusibility after hyperactivation during in vitro capacitation in the mouse. J. Cell
BioI. 102, pp. 1372-1377.
World Health Organisation (1999) Collection and examination of human semen. In:
WHO Laboratory Manual for the Examination of Human Semen and Semen-
Cervical Mucus Interaction. 4th ed. Cambridge University Press, London, pp91-
92.
Yanagimachi R (1969) In vitro acrosome reaction and capacitation of golden hamster
spermatozoa by bovine follicular fluid and its fractions. J Exp Zool 170:269.
Yanagimachi R (1970) The movement of golden hamster spermatozoa before and
after capacitation. J Reprod Fertil 23:193.
Yanagimachi R (1994) Mammalian fertilization. In: The physiology of Reproduction.
Vol1 2nd ed. Knobil E, Neil JD (eds). Raven Press, New York, pp 189-317.
Yovich JM, Edirisinghe WR, Cummins JM, Yovich JL (1988) Preliminary results using
pentoxifyline in a pronuclear stage tubal transfer (PROST) program for severe
male factor infertility. Fertil Steril 50:179.
Stellenbosch University http://scholar.sun.ac.za

86
Yunes R, Michaut M, Tomes C, Mayorga LS (2000) Rab3A triggers the acrosome
reaction in permeabilized human spermatozoa. BioI. Reprod. 62, pp. 1084-
1089.
Zanefeld LJD, De Jonge (1991) In: A comparative overview of mammalian
fertilization. Dunbar B, O'Rand M (eds). Plenum Press, New Yor, pp 1265-1274.
Zeng Y, Clark EN, Florman HM (1995) Sperm membrane potential: hyperpolarization
during capacitation regulates zona pellucida-dependent acrosomal secretion.
Dev Bioi 171, 554-563.
Zeng Y, Oberdorf JA, Florman HM (1996) pH regulation in mouse sperm:
Identification of Na+, cr and HC03- dependent and arylaminobenzoate-
dependent regulatory mechanisms and characterization of their role in sperm
capacitation. Dev Bioi 171, 510-520.
Stellenbosch University http://scholar.sun.ac.za

"All life is an experiment."
- Ralph Waldo Emerson -
Stellenbosch University http://scholar.sun.ac.za

87
CHAPTER 3
MATERIALS AND METHODS
3.1 Introduction
The material and methods used for each study is outlined in the individual articles as
presented in the following chapters. The methods differed slightly between the four
studies, however the basic, common and shared materials and procedures used will
subsequently be discussed.
3.2.1 Preparation of Human tubal fluid (HTF) culture medium
This simple culture medium was developed from the composition of human oviduct
fluid (Quin et al., 1985) and has proved useful for in vitro studies and clinical
applications including IVF, GIFT and lUI. Chemicals used should be at least of
analytical grade and the water to be used of tissue culture grade. The recipe for the
preparation of HTF are as follows:
1. Dissolve the following chemicals in about 600ml tissue culture grade water in a
1000-ml volumetric flask: 5.938g NaCI; 0.350g KCI; 0.049g MgS04.7H20; 0.050g
KH2P04; 2.100g NaHC03; 0.036g Na pyruvate; 0.501g Glucose; 0.003g Phenol
red; 3.136 ml Na lactate (60% syrup)
2. Separately dissolve 0.300g CaC12.2H20 in 100mi tissue culture grade water and
add slowly to the rest.
3. Add Penicillin/Streptopen (75mg)
4. Make up to 1OOOmlwith additional culture grade water and mix thoroughly.
5. Adjust pH to 7.5-7.6
6. Check that osmolarity is between 280-290 mOsm.
Stellenbosch University http://scholar.sun.ac.za

88
7. Filter-sterilise into plastic containers under positive pressure.
8. Store at 4°C
9. Warm to 37°C before use
Add 3% BSA if the medium is to be used as a capacitation medium.
3.2.2 Semen collection
Semen samples were obtained from patients attending the in vitro fertilization
programme at Tygerberg hospital and from healthy donors. All semen samples were
collected by masturbation after 2-3 days of sexual abstinence (WHO, 1999).
Collections were made in sterile plastic containers after which the semen was
allowed to liquify for 30 minutes at 37°C.
3.2.3 Oocyte collection and storage
All oocytes used were nonliving, with no developmental potential. Oocytes were
obtained from ovarian tissue that was collected post mortem. This procedure fully
complies with the legal and ethical guidelines of the South African Medical Research
Council. Ovarian tissue was excised within 24 hours of death and subsequently
macerated/manually dissected (Overstreet et al., 1980) and flushed with phosphate
buffered saline (PBS, GIBCO, Grand Island, NY). The PBS flushing was stored at
4°C for no longer than 48 hours. Zona intact oocytes denuded of granulosa cells
were recovered under microscope and placed directly into a 1.5M solution of MgCI2
and stored at 4°C under mineral oil (E.R. Squib & Sons Inc, Princeton, NJ) for
immediate use. Oocytes were also stored in a dimethylsulfoxide/sucrose (DMSO)
solution at -196°C in liquid nitrogen (Hammit et ai., 1991). Twenty four hours prior to
Stellenbosch University http://scholar.sun.ac.za

89
each test, oocytes were removed from storage and thawed at 37°C. Retrieved
oocytes were placed in 0.25M sucrose and 3% BSA in HTF.
3.2.4 Solubilized zona pellucida preparation
On the day of the experiment, 20 oocytes were placed in a drop on a petri dish with a
glass drawn Pasteur micro pipette, after which the HTF was removed under
microscopic vision (Olympus SZ40; Wirsam Scientific, Cape Town, South Africa),
leaving only the 20 oocytes on the petri dish. A total volume of 5JlIof 10mM HCl was
then added to the oocytes on the petri dish in order to dissolve the zonae at room
temperature. Solubilization of the ZP was microscopically observed and controlled
(45-60min). Following solubilization, the reaction was neutralised by adding 5JlI of
10mM NaOH rendering a final volume of 10Jll. Solubilized ZP solutions with
concentrations of 2 ZP/JlIwere stored up to one week at 4°C.
3.2.5 Assessment of the acrosome reaction
Spermatozoa from the different experiments were fixed on separate spots of spotted
slides (MAGV, Germany, XER 201B) and air-dried. Each spot was flooded with
fluorescein-labeled Pisum Sativum agglutinin solution (FITC-PSA; 125Jlg/ml; l-0770;
Sigma, Cape Town, South Africa) and left in the dark for 30 minutes. After 30 minutes
of incubation the excess FITC-PSA was rinsed of in a beaker containing distilled
water. Mounting medium and a cover slip was placed on the slides and then
evaluated within 24 hours under a fluorescence microscope. Two individuals using
the blind scoring method microscopically scored a minimum of 200 spermatozoa for
each different point. The following staining patterns were evaluated as acrosome
reacted spermatozoa; (i) patchy staining on acrosomal region, (ii) distinct staining in
Stellenbosch University http://scholar.sun.ac.za

90
the equatorial region occurring as an equatorial bar and (iii) no staining over the
entire sperm surface. Spermatozoa with patchy FITC-PSA staining were classified as
a population of sperm where the acrosome reaction was initiated and all were
classified as acrosome reacted (Figure 1).
Unreacted Patchy Equatorial Reacted
All classified as "Reacted"
Figure 1:Patterns recorded during FITC-PSA acrosome staining procedures.
Stellenbosch University http://scholar.sun.ac.za

91
3.2.6 Hemizona binding assay
This functional assay assesses tight binding of sperm to the outer surface of the zona
pellucida hemisphere. It allows a controlled comparison of sperm-zona binding
between control and test spermatozoa, as the zonae surfaces are functionally equal.
3.2.6.1 Bisecting of oocytes
A complete micromanipulation system (Narishige, Tokyo, Japan) was used for
bisecting the oocytes. An inverted phase-contrast microscope (Nikon Diaphot,
Garden City, NY) was equipped with a pair of Narishige micromanipulators (model
MO 102), the connecting tubing filled with mineral oil, and a #11 microscalpel blade
attached to the manipulator. A 100mm petri dish (Falcon #25382, Falcon plastics,
Oxnard, CA) served as the cutting chamber. Culture medium was poured into the
dish to a depth of 3mm. Horizontal grooves were cut into the plastic in order to
provide support in holding the egg while cutting. The egg was transferred to the
working area of the dish using a finely drawn Pasteur glass pipette. Using a total
magnification of 200x, the blade was centred and then slowly lowered, first partially
flattening the egg and then finally initiating a midline cut into the zona. A further
lowering of the blade, along with 1 to 2 side to side excursions, produce two cleanly
cut hemizonae. Vigorous pipetting was used to then dislodge the dense ooplasma
inside each hemizona. Only one egg was cut at a time to ensure that matched
hemizona remained paired for subsequent sperm binding tests. Each hemizona pair
was placed in a 501J1droplet of medium in a petri dish, covered with mineral oil and
stored at 4°C.
Stellenbosch University http://scholar.sun.ac.za

92
3.2.6.2 Competitive sperm-binding to the hemizona
A 501J1sperm droplet containing O.5x106 spermimi from both the test and control
samples were placed in a marked tissue culture dish. One hemizona was placed in
the one droplet and the matching hemizona in the other (Figure 2). The droplets were
covered with mineral oil in order to prevent evaporation and dehydration of the sperm
droplets and co-incubated at 37°C and 5% C02 for 4 hours. After co-incubation, the
hemizona from each droplet was transferred into culture medium (HTF) droplets
respectively. The hemizonae were then rinsed by vigorously pipetting five times with
a finely drawn glass pipette in order to dislodge loosely associated spermatozoa. The
number of spermatozoa firmly bound to the outer surface of each hemizona was
counted under phase contrast microscopy (Nikon TMS-F, Research Inst.
Johannesburg, South Africa) at a 200x magnification. The number of sperm bound in
the test sample was divided by the number of sperm bound in the control sample and
expressed as a percentage to give a hemizona index (HZI).
Stellenbosch University http://scholar.sun.ac.za

93
HEMIZONA SPERM BINDING ASSAY
Human oocyte.. ..~ rri\ Oocyte is bisected into~ ~ 2 identical Hemizonae
Jl' "
Test Control
Hemizonae are incubated in test and~ spermpopulationsfor4&
The number of spermfirmly bound to Hemizona are counted and sperm-zona binding
is expressed as Hemizona index
Zona bound sperm (test) x 100Binding index:Zona bound sperm (control)
Figure 2: The competitive hemizona sperm binding assay.
Stellenbosch University http://scholar.sun.ac.za

94
3.2.7 Computer assisted semen analysis (CASA)
Sperm motility/kinematics were determined with the Hamilton-Thorne IVOS analyser
(Hamilton-Thorne Research, Beverly, MA) with standard set-up parameters. The
analyser settings were as follows: 30 frames/60 Hz; minimum contrast, 80; minimum
cell size, 2; minimum static contrast, 30; low average path velocity (VAP) cut-off, 5
mts; low VSL cut-off, 11 mts; head size, nonmotile, 3; head intensity, nonmotile, 160;
static head size, 1.01-2.91; static head intensity, 0.60-1.40; slow cells not motile;
magnification, 2.01, and temperature, 3rC.
Low sperm counts «10 x 106/mL) were confirmed by manual analysis, while high
counts «50 x 106/mL) were diluted 1:1 with HTF medium for accurate determination
of count by the CASA (IVOS) system.
3.2.8 Statistical Analyses
All statistical evaluations and tests were carried out using GraphPad Prism 2.01.
Data are expressed as mean±SE. Student's t-test for paired data was used to
compare the results of all the acrosome and motility studies, while Pearson's test was
used to perform correlation tests. The Mann-Whitney U test for nonparametrie data
was used to compare binding results. A Hemizonae Index was also calculated for
matched hemizonae assay results and expressed as a mean percentage where HZI
- Test sperm bound to hemizona/ 100) P I I I th- Control sperm bound to hemizona X . -va ues equa or ess an
0.05 were considered statistically significant.
Stellenbosch University http://scholar.sun.ac.za

95
References
Hammit DG, Syrop CH, Walker DL, Bennet MR (1991). Conditions of oocyte storage
and use of non inseminated, non-fertilized oocytes for the hemizona assay.
Fertil SteriI60:131-136.
Overstreet JW, Yanagimachi R, Katz OF, Hayashi K, Hanson FW (1980) Penetration
of human spermatozoa into the human zona peJlucida and the zona-free
hamster egg: a study of fertile donors and infertile patients.Fertil Steril.
33(5):534-42.
Quin P, Kerin JF, Warnes GM (1985). Improved pregnancy rate in human in vitro
fertilization with the use of a medium based on the composition of human tubal
fluid. Fertil Steril 44:493-498.
World Health Organisation (1999) Collection and examination of human semen. In:
WHO Laboratory Manual for the Examination of Human Semen and Semen-
Cervical Mucus Interaction. 4th ed. Cambridge University Press, London, pp91-
92.
Stellenbosch University http://scholar.sun.ac.za

"There is one thing even more vital to
science than intelligent methods; and that
is, the sincere desire to find out the truth,
whatever it may be."
- Charles Sanders Pierce -
Stellenbosch University http://scholar.sun.ac.za

96
CHAPTER4
THE ZONA PELLUCIDA-INDUCED ACROSOME REACTION OF
HUMAN SPERMATOZOA INVOLVES EXTRACELLULAR SIGNAL-
REGULATED KINASE ACTIVATION
ANDROLOGIA 2001 Vol 33 no. 6 pp337-342
SS du Plessis1,*, C page1 and DR Franken2
1Department of Medical Physiology and Biochemistry, University of Stellenbosch,
P.O.Box 19063, Tygerberg, 7505, South Africa.
2Department of Obstetries and Gynaecology, University of Stellenbosch, Tygerberg
Hospital, Tygerberg, 7505, South Africa.
Running head: Role for ERK during acrosome reaction
Keywords: spermatozoa, acrosome, ERK, zona pellucida
'Corresponding author. SS du Plessis, Department of Medical Physiology and
Biochemistry, University of Stellenbosch, P.O.Box 19063, Tygerberg, 7505, South
Africa; Tel.: +27-(0) 21-938 9388; fax: +27-(0) 21-938 9476; e-mail:
Stellenbosch University http://scholar.sun.ac.za

97
Summary
Extracellular signal-regulated kinases (ERK), a member of the family of mitogen-
activated protein kinases (MAPK) are cytoplasmic and nuclear serine/threonine
kinases involved in signal transduction of several extracellular effectors. Recent
evidence indicates the presence of p21Ras and the phosphorylation of ERK-1 and
ERK-2, suggesting the occurrence of the Ras/ERK cascade in mammalian
spermatozoa. We report here on the biological role of ERKs during the acrosome
reaction, on stimulation with zona pellucida (ZP), in human spermatozoa. The
mitogen-activated protein kinase inhibitor PD098059 was used as a pharmacological
tool to study the involvement of extracellular signal-regulated kinases in the induction
of the acrosome reaction in human spermatozoa. This compound significantly
inhibited both the ZP- and A23187-induced acrosome reactions. These results
suggest that ERKs are involved in the signal transduction pathway through which ZP
stimulation works during the process of fertilization.
Introd uction
The development of the fertilization-competent state of the spermatozoon occurs
through a series of poorly understood processes including capacitation and
acrosomal reaction (AR). During the process of capacitation spermatozoa acquire the
ability to fertilize the oocyte. Capacitation is characterised by a series of profound
membrane and metabolic transformations in spermatozoa that increase their ability to
respond to physiological stimuli of the acrosome reaction (Yanagimachi, 1994) [1].
The AR is a specialised exocytotic event consisting of multiple fusion's and
fenestration's of the outer acrosomal membrane and the overlying plasma membrane
that lead to subsequent release of the acrosomal enzymes that help the spermatozoa
Stellenbosch University http://scholar.sun.ac.za

98
to penetrate the zona pellucida (Yanagimachi, 1994). These two activational
processes appear to be regulated by intracellular signalling systems similar to those
utilised by somatic cells (Kopf et al., 1995). Although the precise signal transduction
pathways have not been fully elucidated (Baldi et al., 1996) it is known that both
processes are characterised by increases in intracellular calcium concentrations and
phosphorylation of proteins including that in tyrosine residues (Leyton & Saling, 1989;
Carr & Acott, 1990; Naz et aI., 1991; Duncan & Fraser, 1993; Burks et aI., 1995;
Luconi et aI., 1995; Visconti et aI., 1995; Luconi et al. 1996).
Several reports from studies in somatic cells focused attention on the presence and
activation of the mitogen-activated protein kinase (MAPK) family of kinases in
response to ligands binding both G-protein coupled receptors as well as protein
tyrosine kinases (PTK) receptors. The MAPK's are a family of serine/threonine
kinases with multiple membrane, cytosolic and nuclear substrates, the activation of
which lead to an array of responses. This include the activation of (gene transcription
via translocation of MAPK into the nucleus) genes encoding for protective proteins in
response to stress, cell proliferation, cell differentiation, apoptosis and exocytosis
(Offermans et aI., 1994; Page & Doubell, 1996; Canman & Kastan, 1996). It is also
known that the activation of the MAPK family of kinases can be up- or down
regulated through crosstalk with other signalling pathways i.e. protein kinase A
(PKA), protein kinase C (PKC) and PTK in many cell types (Bogoyevitch et aI., 1994;
Burgering & Bos, 1995). Extracellular signal-regulated kinases, ERK-1 (p44 MAPK)
and ERK-2 (p42 MAPK), are the members of this family studied the most in somatic
cells. The presence and biological activity of p21 Ras has recently been
demonstrated in human spermatozoa (Naz et al., 1992). Recent characterisation of a
Stellenbosch University http://scholar.sun.ac.za

99
boar sperm protein kinase with characteristics similar to those of ERK-2 (Berruti,
1994), as well as demonstrating the presence of ERK's in human spermatozoa
(Luconi et ai., 1998) and the activation thereof during in vitro capacitation lead to the
hypothesis that the Ras/Raf/ERK cascade may be involved in the capacitation and/or
AR in human spermatozoa during zona pellucida (ZP) stimulation.
As of yet very little is known about ZP-mediated sperm signal transduction in the
human, due, for the most part, to an inability to obtain sufficient quantities of human
ZP for experimental purposes. The human ZP has been shown to bind human
spermatozoa and to induce the AR of spermatozoa (Cross et ai., 1988; Morales et
ai., 1989).
The present study aimed to determine the regulatory role of MAPK and in particular
ERK during the acrosome reaction in human spermatozoa.
Materials and Methods
Preparation of sperm sam pies
Semen samples were collected from normozoospermic donors by masturbation after
2-3 days of sexual abstinence. Semen samples were analysed according to the
World Health Organisation criteria (WHO, 1992) together with strict sperm
morphology assessment (Kruger et ai., 1986). Motile sperm fractions were collected
from samples using a slightly modified double-wash swim-up technique. Retrieved
sperm samples were resuspended in synthetic human tubal fluid medium (HTF)
(Quin et ai., 1985) supplemented with 3% bovine serum albumin (BSA; Seravac,
Stellenbosch University http://scholar.sun.ac.za

100
Cape Town, South Africa) to a sperm concentration of 10x106 cells/ml. Before the
onset of AR studies, sperm samples were allowed to capacitate at 37°C in 5% CO2
for 3 hours in HTF-BSA. Prepared sperm samples were incubated in the presence or
absence of the MEK-inhibitor, PD098059 (P-215, Sigma, Cape Town, South Africa)
at a concentration of 50/J.Mfor 90 minutes at 37°C.
Preparation of solubilized ZP
Oocytes were retrieved from post mortem derived ovarian material. (The project fully
complies with the legal and ethical guidelines of the South African Medical Research
Council.) Oocytes were stored in a dimethylsulfoxide/sucrose solution at -196°C in
liquid nitrogen (Hammit et al., 1991). Twenty four hours prior to each test, oocytes
were removed from storage and thawed at 37°C. Retrieved oocytes were placed in
0.25M sucrose and 3% BSA in HTF. On the day of the experiment, 20 oocytes were
placed in a drop on a petri dish, after which the HTF was removed under microscopic
vision (Olympus SZ40; Wirsam Scientific, Cape Town, South Africa), leaving only the
20 oocytes on the petri dish. A total volume of 5/J.1of 10mM HCl was then added to
the oocytes on the petri dish; solubilization of the ZP was microscopically observed
and controlled. Following solubilization, 5/J.1of 10mM NaOH was added to the
solubilized ZP, to render a final zona volume of 10/J.1containing 2 ZP//J.1.The final ZP
concentration, after the addition of spermatozoa, was 0.67 ZP//J.1.
Acrosome reaction studies
Acrosomal statuses of spermatozoa stimulated with (i) 10/J.MA23187 (C-7522,
Sigma, Cape Town, South Africa) for 20 minutes and (ii) 0.67ZP//J.1for 60 minutes in
either the presence or absence of PD098059 (pre-treatment) were determined
Stellenbosch University http://scholar.sun.ac.za

101
according to procedures published elsewhere (Cross et ai., 1988; Morales et ai.,
1989, Franken et al., 2000) and compared to that of control samples. Control
samples were allowed to spontaneously acrosome react.
Prior to aspiration into Teflon tips during the micro-assay (Franken et al., 2000), the
sperm/ZP suspensions were gently mixed in a well of a 60 well micro-titre plate
(Microtest plate cat No. P43, Laboratory and Scientific, Cape Town South Africa).
Aspirating HTF droplets into both sides of the Teflon tip sealed off the sperm
suspensions and prevented evaporation from the tip. Each sperm/ZP suspension
was separated from the HTF droplets by air bubbles on both sides.
Progressive motility for both acrosome reaction techniques was monitored before and
after the incubation periods. Sperm droplets were carefully placed on separate spots
of spotted slides (MAGV, Germany, XER 201B) and immediately evaluated for
percentage live sperm under inverted phase contrast microscope (Nikon TMS-F,
Research Inst. Johannesburg, South Africa). Sperm samples were obtained after
swim-up and only samples with a progressive motility of more than 80%, according to
the World Health Organisation criteria (WHO, 1999), were subsequently used in the
experiments.
Spermatozoa from the different experiments were fixed and air-dried, after which
acrosomal status was determined using fluorescein-labeled Pisum Sativum agglutinin
(FITC-PSA; 125f.!g/ml; L-0770; Sigma, Cape Town, South Africa). Two individuals
using the blind scoring method scored a minimum of 200 spermatozoa for each
different point. The following staining patterns were evaluated as acrosome reacted
Stellenbosch University http://scholar.sun.ac.za

102
spermatozoa; (i) patchy staining on acrosomal region, (ii) distinct staining in the
equatorial region occurring as an equatorial bar and (iii) no staining over the entire
sperm surface. Spermatozoa with patchy FITC-PSA staining were classified as a
population of sperm where the acrosome reaction was initiated and all were classified
as acrosome reacted.
Statistical Analysis
The percentage acrosome reacted sperm were compared using Student's paired t
test. p-values equal or less than 0.05 were considered statistically significant.
Results
Acrosome reactions induced spontaneously were completely insensitive to the pre-
treatment of spermatozoa with P0098059 and the mean percentage of acrosome-
reacted spermatozoa remained. As expected, A23187 stimulation induced the AR
and the mean percentage of acrosome-reacted spermatozoa increased with 18.8%,
which was significantly higher than that of the control values (p<0.05). However, after
PO pre-treatment, A23187 stimulation did not induce the AR reaction significantly
more than control values (Figure 1).
ZP stimulation also induced the AR and increased the mean percentage of acrosome
reacted spermatozoa significantly by 9.2% (p<0.05). P0098059 pre-treatment of
spermatozoa resulted in a complete inhibition of ZP stimulated AR and the mean
percentage of acrosome reacted spermatozoa stayed at basal values (13.5%)
(Figure 2).
Stellenbosch University http://scholar.sun.ac.za

103
25.0
20.0 -
15.0 -r::t::« 10.0 -::::e0
5.0*
0.0 I-5.0 PD
*
Figure 1. Influence of the MEK-inhibitor P0098059 (PO) on the acrosome reaction
(Mean±SE) mediated by A23187 (difference between acrosome reaction and the
spontaneous percentage acrosome reaction of 23.9±2.2%). (n=10)
* p<O.05 compared with A23187
Capacitation seems to have been completed before the incubation of the
spermatozoa with the MEK-inhibtor (PD098059) since these cells were able to
acrosome react in response to both ZP and A23187 stimulation (results not shown).
Stellenbosch University http://scholar.sun.ac.za

104
25.0
20.0 -
15.0a::<C 10.0
5.0 -
0.0 --~-
-5.0 -
* *
___PO _ ZP- :{-PD +_ZP
I
Figure 2. Influence of the MEK-inhibitor P0098059 (PO) on the acrosome reaction
(Mean±SE) mediated by ZP (difference between acrosome reaction and the
spontaneous percentage acrosome reaction of 13.5±2.0%). (n=3)
* p<O.001 compared with ZP
Discussion
The present paper shows that ERK activation plays a biological role in the ability of
human spermatozoa to undergo the acrosome reaction, since the ability of these
cells to acrosome react in response to ZP and the calcium ionophore, A23187, is
strongly inhibited in the presence of the MAPK (ERK) cascade inhibitor, PD098059.
To understand the signalling pathways through which both receptor induced (ZP3
binding to the ZP membrane receptor) and non-receptor induced (opening of calcium
channels by an ionophore) mechanisms could elicit the acrosome reaction, a
modified version of the recently hypothesized scheme of Breitbart and Spungin
Stellenbosch University http://scholar.sun.ac.za

105
(1997) will be referred to (Figure 3). The ZP3 glycoprotein binds to at least two
receptors in the plasma membrane. One receptor is a Gj-coupled receptor that
activates phospholipase C (PLC) ~1. The other receptor is a tyrosine kinase receptor
(TKR) coupled to PLCy Binding to the Gj-coupled receptor would regulate adenylate
cyclase (AC) leading to the elevation of cAMP and PKA activation. PKA activates a
voltage-dependent Ca2+ channel in the outer acrosomal membrane, which releases
Ca2+ from the interior of the acrosome to the cytosol. This relative small rise in [Ca2+]j
could result in the activation of PLCy The products of phosphatidyl-inositol
biphosphate (PIP2) hydrolysis by PLCf11 and PLCy, diacylglycerol (DAG) and
inositoltrisphosphate (IP3) lead to PKC translocation to the plasma membrane and its
subsequent activation. This increase in [Ca2+]j can be mimicked by the addition of a
calcium ionophore (e.g. A23187), which will also result in the activation of PLCy, and
PKC activity. PKC opens a voltage-dependent Ca2+ channel in the plasma
membrane, increasing the [Ca2+]1 even more. PKC activation also results in the
activation of ERK, through its ability to phosphorylate and activate an upstream
mediator of the ERK cascade, Raf. Raf will activate and phosphorylate the ERK
kinase, MEK, which in turn activates ERK. The Gj and TKR can also activate a
Na+/H+ exchanger, leading to alkalization (pH increase) of the cytosol. The increase
in [Ca2+]j and pH will lead to membrane fusion and acrosomal exocytosis.
Stellenbosch University http://scholar.sun.ac.za

106
ZP3
Plasma membrane
Ras
Outer acrosomal membrane
Figure 3. Possible interactions between the different signal transduction pathways
invoked during the acrosome reaction. (ZP3 = zona pellucida glycoprotein; R = G;-
coupled receptor; TKR = tyrosine kinase receptor; dashed lines = hypothesised
activation of ERK) (Modified from Breitbart & Spungin, 1997)
In support of this hypothesis, it was previously shown in our laboratories that the ZP-
induced AR appears to be mediated through a G-protein-mediated signal
transduction process after functional inactivation of the Gi-protein receptor by
Stellenbosch University http://scholar.sun.ac.za

107
pertussis toxin (Bastiaan et ai., 1999). Aitken et al. (1996) have also shown that the
addition of the tyrosine kinase inhibitor, genistein, inhibited capacitation, probably by
inhibiting the protein tyrosine kinase ZP receptor. In agreement with the hypothesis,
Luconi et al. (1998) also showed that in vitro capacitation stimulates a sustained and
concomitant increase in tyrosine phosphorylation and kinase activity of ERKs,
indicating the activation of these enzymes during capacitation. It was also shown that
the ability of human spermatozoa to undergo the AR in response to progesterone
were strongly inhibited when capacitation was performed in the presence of the
MAPK cascade inhibitor PD098059 (Luconi et ai., 1998).
The cellular targets of activated ERKs include, nuclear-, cytosolic-, cytoskeletal- and
membrane proteins (Bokemeyer et ai., 1996, Gonzales et ai., 1993). In mitotic cells,
the main target of activated ERKs, is the nucleus, where it phosphorylates
transcriptional factors, ultimately resulting in de novo protein synthesis and cell
proliferation. In non-mitotic mature spermatozoa the nucleus plays hardly, if any, a
role in the modulation of their biological function and therefore a nucleur translocation
of ERKs seem unlikely. Thus the presence of ERKs at the level of the equatorial
segment after the AR (Luconi et ai., 1998) might be related to the regulation and/or
activation of proteins that mediate binding and fusion between the sperm and egg
plasma membranes. It has also been demonstrated that a conformational
rearrangement of proteins, which appear to play an important role in the sperm-
oocyte fusion at the level of the equatorial membrane, take place after stimulation of
the AR (Allen & Green, 1995).
Stellenbosch University http://scholar.sun.ac.za

108
The downstream targets of ERKs during capacitation and the AR of spermatozoa
remain to be defined. Seeing that both capacitation and AR are earmarked by the
rearrangement of cytoskeletal elements, a possible target might be cytoskeletal
elements such as microtubule-associated proteins (MAP), whose phosphorylation
may be important in the cytoskeletal rearrangements that occur during capacitation
(Duncan & Fraser, 1993).
In conclusion, our data demonstrates that ERKs are directly or indirectly involved in
the acrosome reaction induced by human zona pellucida.
Acknowledgements
This study was supported in part by the Technology and Human Resources for
Industry Programme (THRIP), managed by the National Research Foundation (NRF)
and funded by the Department of Trade and Industry (DTI).
Stellenbosch University http://scholar.sun.ac.za

109
References
Allen CA, Green OPL (1995). Monoclonal antibodies which recognise equatorial
segment epitopes presented de novo following the A23187-induced acrosome
reaction of guinea pig sperm. J Cell Sci 108:767-777.
Aitken RJ, Buckingham OW, Harkiss 0 (1996). The extragenomic action of human
spermatozoa is influenced by redox regulated changes in tyrosine
phosphorylation during capacitation. Mol Cell Endocrinol 17:83-93.
Baldi E, Luconi M, Bonaccorsi L, Krausz C, Forti G (1996). Human sperm activation
during capacitation and acrosome reaction: role of calcium, protein
phosphorylation and lipid remodelling pathways. Front Biosci 1:189-205.
Bastiaan H, Franken DR, Wranz P (1999). G-Protein regulation of the solubilised
Human Zona Pellucida-Mediated Acrosome Reaction and Zona Pellucida
Binding. JAssist Reprod Gen 16(6): 332-336.
Berruti G (1994). Biochemical characterisation of the boar sperm 42 kilodalton protein
tyrosine kinase: its potential for tyrosine as well as serine phosphorylation
towards microtubule-associated protein 2 and histone H 2B. Mol Reprod Oev
38:386-392.
Bogoyevitch MA, Glennon PE, Andersson MB, Clerk A, Lazon A, Marshall CJ, Parker
PJ, Sugden PH (1994). ET1 and fibroblast growth factors stimulate the MAP-
kinase signalling cascade in cardiac myocytes. The potential role of the
cascade in the integration of the two signalling pathways leading to myocyte
hypertrophy. J Bioi Chem 269(2):1110-1119.
Bokemeyer 0, Sorokin A, Dunn MJ (1996). Multiple intracellular MAP kinase
signalling cascade. Kidney Int 49:1187-1198.
Stellenbosch University http://scholar.sun.ac.za

110
Breitbart H, Spungin B (1997). The biochemistry of the acrosome reaction. Mol Hum
Reprod 3(3):195-202.
Burgering BM, Bos JL (1995). Regulation of RAS mediated signalling: More than one
way to skin a cat. Trends Biochem Sci 20:18-22.
Burks OJ, Carballada R, Moore HOM, Saling PM (1995). Interaction of a tyrosine
kinase from human sperm with the zona pellucida at fertilization. Science
269:83-86.
Canman CE, Kastan MB (1996). Three paths to stress relief. Nature 384:213-214.
Carr OW, Acott TS (1990). The phosphorylation of a putative sperm micro tubule-
associated protein 2 (MAP2) is uniquely sensitive to regulation. Bioi Reprod
43:795-805.
Cross NL, Morales P, Overstreet JW, Hanson FW (1988). Induction of acrosome
reactions by the human zona pellucida. Bioi Reprod 38:235-244.
Duncan AE, Fraser LR (1993). Cyclic AMP-dependent phosphorylation of epididymal
mouse sperm proteins during capacitation in vitro: identification of an Mr 95 000
phosphotyrosine-containing protein. J Reprod Fertil 97:287-299.
Franken DR, Bastiaan HS, Oehninger SC (2000) Physiological induction of the
acrosome reaction in human sperm: Validation of a micro-assay using minimal
volumes of solubilised, homologous zona pellucida. JAssist Reprod Gen
17(3):156-161.
Gonzales FA, Seth A, Raden DL, Bowman OS, Fay FS, Davis RJ (1993). Serum-
induced translocation of mitogen-activated protein kinase to cell surface ruffling
membrane and the nucleus. J Cell Bioi 122:1089-1101.
Stellenbosch University http://scholar.sun.ac.za

111
Hammit DG, Syrop CH, Walker DL, Bennet MR (1991). Conditions of oocyte storage
and use of non inseminated, non-fertilized oocytes for the hemizona assay.
Fertil SteriI60:131-136.
Kopf GS, Visconti PE, Moos J, Galentino-Homer HL, Ning XP (1995). Integration of
tyrosine kinase and G protein-mediated signal transduction in the regulation of
mammalian sperm function. In: Human sperm acrosome reaction. Eds Fenichel
& Parinaud 236:191-216.
Kruger TF, Menkveld R, Stander FSH, Lombard CJ, Van der Merwe JP, Van Zyl JA,
Smith K (1986). Sperm morphologic features as a prognostic factor in in vitro
fertilization. Fertil Steril 46:1118-1123.
Leyton L, Saling PM (1989). 95 kd sperm proteins bind ZP3 and serve as tyrosine
kinase substrates I response to zona binding. Cell 57:1123-1130.
Luconi M, Barni, T, Vanelli GB, Krausz C, Marra F, Benedetti PA, Evangelista V,
Francavilla S, Properzi G, Forti G, Baldi E (1998). Extracellular regulated
Signal-Kinases Modulate Capacitation of Human Spermatozoa. Bioi Reprod
58:1476-1489.
Luconi M, Bonaccorsi L, Krausz C, Gervasi G, Forti G, Baldi E (1995). Stimulation of
protein tyrosine phosphorylation by platelet-activating factor and progesterone
in human spermatozoa. Mol Cell Endocrinol 108:35-42.
Luconi M, Krausz C, Forti G, Baldi E (1996). Extracellular calcium negatively
modulates tyrosine phosphorylation and tyrosine kinase activity during
capacitation of human spermatozoa. Bioi Reprod 55:207-216.
Morales P, Cross NL, Overstreet JW, Hanson FW (1989). Acrosome-intact and
acrosome-reacted human sperm can initiate binding to the zona pellucida. Dev
Bioi 133: 385-392.
Stellenbosch University http://scholar.sun.ac.za

112
Naz RK, Ahmad K, Kaplan P (1992). Expression and function of ras-proto-oncogene
proteins in human sperm cells. J Cell Sci 102:487-494.
Naz RK, Ahmad K, Kumar R (1991). Role of membrane phosphotyrosine proteins in
human spermatozoa function. J Cell Sci 99:157-165.
Offermans S, Jones SV, Bombien E, Schultz G (1994). Stimulation of MAPK activity
of different secretory stimuli in rat basophilic leukemia cells. J Immunol 152:250-
261.
Page C, Doubell AF (1996). Mitogen-activated protein kinase (MAPK) in cardiac
tissue. Mol Cell Bio 157:49-57.
Quin P, Kerin JF, Warnes GM (1985). Improved pregnancy rate in human in vitro
fertilization with the use of a medium based on the composition of human tubal
fluid. Fertil Steril 44:493-498.
Visconti PE, Bailey JL, Moore GD, Pan 0, Olds-Clarke P, Kopf GS (1995).
Capacitation of mouse spermatozoa. I: Correlation between the capacitation
state and protein tyrosine phosphorylation. Development 121:1129-1137.
World Health Organisation (1992). WHO Laboratory Manual for the Examination of
Human Semen and Semen-Cervical Mucus Interaction, 3rd ed. Cambridge
University Press, Cambridge pp3-26.
World Health Organisation (1999). WHO Laboratory Manual for the Examination of
Human Semen and Semen-Cervical Mucus Interaction, 4th ed. Cambridge
University Press, Cambridge p9.
Yanagimachi R (1994). Mammalian fertilization. In: The physiology of Reproduction.
Knobil E, Neil JD (eds), Vol1 2nd ed. New York: Raven Press, 189-317.
Stellenbosch University http://scholar.sun.ac.za

"A scientific truth does not triumph by
convincing its opponents and making them
see the light, but rather because its
opponents eventually die and a new
generation grows up that is familiar with it."
- Maxwell Planck -
Stellenbosch University http://scholar.sun.ac.za

113
CHAPTER 5
EXTRACELLULAR SIGNAL-REGULATED KINASE ACTIVATION
INVOLVED IN HUMAN SPERM-ZONA PELLUCIDA BINDING
ANDROLOG/A 2002 Vo/34 no. 1pp55-59
SS du Plessis":', C page1 and DR Franken2
1Department of Medical Physiology and Biochemistry, University of Stellenbosch,
P.O.Box 19063, Tygerberg, 7505, South Africa.
2Department of Obstetries and Gynaecology, University of Stellenbosch, Tygerberg
Hospital, Tygerberg, 7505, South Africa.
Running head: ERK involved in sperm-zona binding
Keywords: spermatozoa, acrosome, ERK, zona pellucida, hemizona
·Corresponding author. SS du Plessis, Department of Medical Physiology and
Biochemistry, University of Stellenbosch, P.O.Box 19063, Tygerberg, 7505, South
Africa; Tel.: +27-(0) 21-938 9388; fax: +27-(0) 21-938 9476; e-mail:
Stellenbosch University http://scholar.sun.ac.za

114
Summary
In a previous study involving the inhibition of the mitogen-activated protein kinase
(MAPK), extracellular signal regulated kinase (ERK), we found that the very specific
MAPK Kinase (MEK) inhibitor, PD098059, inhibited the zona pellucida (ZP) induced
acrosome reaction. Since an intact acrosome on the spermatozoa is a prerequisite to
ensure tight binding to the ZP, we investigated the zona binding potential of
spermatozoa after PD098059 treatment of sperm followed by exposure to solubilized
human ZP and calcium ionophore (A23187). PD098059 treated spermatozoa
exposed to solubilized ZP bound significantly more to the ZP as compared to control
spermatozoa also exposed to solubilized ZP (26.5±3.7 vs. 13.8±2.8, p<O.05). No
significant differences in binding to the ZP were observed between PD098059
treated and untreated sperm populations after A23187 exposure. These results can
be interpreted to support the idea that the ZP-induced AR is the physiologically
relevant exocytotic event since it is the ZP-induced AR, and not the spontaneous
(culture medium) or A23187 induced AR, which appears to be mediated through an
ERK-mediated signal transduction process.
Introd uction
Before a spermatozoon can penetrate and fertilize the oocyte, it has to undergo
various physiological processes i.e. capacitation (series of membrane and metabolic
transformations), acrosome reaction (AR - fusion of the outer acrosomal membrane
with the plasma membrane leading to exocytosis of enzymes and exposure of new
membrane domains), as well as tight binding to the oocyte (species specific binding
of spermatozoa to the zona pellucida - ZP). The human AR, a Ca2+-dependent
exocytotic event that is regulated by voltage-operated calcium ion channels located
Stellenbosch University http://scholar.sun.ac.za

115
in the plasma membrane of the sperm head, is a crucial early step in the fertilization
process, and must be completed prior to fusion with oocytes (Babcock & Pfeiffer,
1987; Florman et aI., 1992; Florman, 1994; Arnouit et aI., 1996; Son et aI., 2000).
Under physiological conditions, sperm voltage operated calcium channels are usually
activated by contact with the ZP (Florman et aI., 1992; Arnouit et aI., 1996), while it
can also be experimentally mediated by progesterone (Foresta et aI., 1993).
The mammalian ZP, and specifically glycoprotein 3 (ZP3), is the main mediator of
sperm-egg recognition, sperm binding and the AR in mammals (Yanagimachi, 1994).
ZP3 induces Ca2+ influx into cytoplasm, leading to increases of intracellular Ca2+ and
pH, and resulting in acrosomal exocytosis (Babcock & Pfeiffer, 1987). The human ZP
has been shown to bind human spermatozoa and to induce the AR of spermatozoa
in both the intact and the solubilized state (Bielfield et aI., 1994; Cross et al., 1988;
Morales et aI., 1989). However, these methods cannot be used routinely in human
studies, due to the scarcity of human material. Instead, several bovine serum albumin
(BSA) neoglyco-proteins were shown to be able to stimulate the AR by interacting
with the putative receptor for ZP3 in human spermatozoa (BrandeIIi et al., 1996;
Blackmore & Eisoldt, 1999).
Extracellular regulated kinase (ERK), a member of the serine/threonine family of
mitogen activated kinases, can be phosphorylated and activated through both G-
protein- and protein tyrosine kinase (PTK) receptor ligand binding in many cell types
(Canman & Kastan, 1996; Page & Doubell, 1996). Both of these receptor types also
play a role in the intracellular signalling systems regulating the processes of
capacitation and AR. In support of this hypothesis, it was previously shown in our
Stellenbosch University http://scholar.sun.ac.za

116
laboratories that the ZP-induced AR appears to be mediated through a G-protein-
mediated signal transduction process after functional inactivation of the o-protein
receptor by pertussis toxin (Bastiaan et a/., 1999; Franken et a/., 1996). Aitken et al.
(1996) have also shown that the addition of the tyrosine kinase inhibitor, genistein,
inhibited capacitation, probably by inhibiting the protein tyrosine kinase ZP receptor.
It has recently been shown that ERK's are present in human spermatozoa (Luconi et
a/., 1998). We have also showed recently in our laboratories that the specific
inhibition thereof significantly reduced the ZP stimulated AR in human spermatozoa
(Du Plessis et a/., 2001).
The notion that ERK might play an important regulatory role in sperm ZP binding and
thus sperm-oocyte binding was thus further pursued during this study.
Materials and Methods
Preparation of sperm sam pies
Semen samples were collected from normozoospermic donors by masturbation after
2-3 days of sexual abstinence. Semen samples were analysed according to the
World Health Organisation criteria (WHO, 1999) together with strict sperm
morphology assessment (Kruger et a/., 1986). Motile sperm fractions were collected
from samples using a slightly modified double-wash swim-up technique. Retrieved
sperm samples were resuspended in synthetic human tubule fluid culture medium
(HTF) (Quin et a/., 1985) supplemented with 3% bovine serum albumin (BSA;
Seravac, Cape Town, South Africa) to a sperm concentration of 10x106 cells/ml.
Before the onset of AR studies, sperm samples were allowed to capacitate at 37°C in
Stellenbosch University http://scholar.sun.ac.za

117
5% CO2 for 3 hours in HTF-BSA. Prepared sperm samples were incubated in the
presence or absence of the MEK-inhibitor, PD098059 (P-215, Sigma, Cape Town,
South Africa) at a concentration of 50JlMfor 90 minutes at 37°C.
Preparation of solubilized ZP
Oocytes were retrieved from post mortem derived ovarian material. (The project fully
complies with the legal and ethical guidelines of the South African Medical Research
Council.) Oocytes were stored in a dimethylsulfoxide/sucrose solution at -196°C in
liquid nitrogen (Hammit et a/., 1991). Twenty four hours prior to each test, oocytes
were removed from storage and thawed at 37°C. Retrieved oocytes were placed in
0.25M sucrose and 3% BSA in HTF. On the day of the experiment, 20 oocytes were
placed in a drop on a petri dish, after which the HTF was removed under microscopic
vision (Olympus SZ40; Wirsam Scientific, Cape Town, South Africa), leaving only the
20 oocytes on the petri dish. A total volume of 5JlI of 10mM HCl was then added to
the oocytes on the petri dish; solubilization of the ZP was microscopically observed
and controlled. Following solubilization, 5JlI of 10mM NaOH was added to the
solubilized ZP, to render a final zona volume of 10JlI containing 2 ZP/JlI (Liu and
Baker, 1996). The final ZP concentration, after the addition of spermatozoa, was 0.67
ZP/Jll.
Hemizona Assays/Zona Pellucida Binding
Spermatozoa were stimulated with (i) 10JlM A23187 (C-7522, Sigma, Cape Town,
South Africa) for 20 minutes and (ii) 0.67ZP/JlI for 60 minutes in either the presence
or absence of PD098059 (pre-treatment) to elicit the AR (Cross et a/., 1988; Morales
Stellenbosch University http://scholar.sun.ac.za

118
et a/., 1989, Franken et a/., 2000) and compared to that of control samples. Control
samples were allowed to spontaneously acrosome react.
Prior to aspiration into Teflon tips during the micro-assay (Franken et a/., 2000), the
sperm/ZP and sperm/A23187 suspensions were gently mixed in a well of a 60 well
micro-titre plate (Microtest plate cat No. P43, Laboratory and Scientific, Cape Town
South Africa). Aspirating HTF droplets into both sides of the Teflon tip sealed off the
sperm suspensions and prevented evaporation from the tip. Each sperm/ZP and
sperm/A23187 suspension was separated from the HTF droplets by air bubbles on
both sides.
Progressive motility of all samples was monitored before and after the incubation
periods. Sperm droplets were carefully placed on separate spots of spotted slides
(MAGV, Germany, XER 201B) and immediately evaluated for percentage live sperm
under inverted phase contrast microscope (Nikon TMS-F, Research Inst.
Johannesburg, South Africa). Only samples with a progressive motility of more than
80% were subsequently used in the experiments.
Both test and control sperm droplets (15!J.1;10x106sperm/ml) were placed on a petri
dish to which hemizonae were added in a match-controlled fashion. Hemizona
assays (HZA) were performed multi-fold and co-incubation was for four hours.
Following the co-incubation period, hemizonae were removed and washed (5x) to
strip the loosely attached spermatozoa from the hemizonae. Hemizonae were then
evaluated, while the number of spermatozoa tightly bound to the ZP was recorded for
each test and matching control. Two individuals using the blind scoring method
Stellenbosch University http://scholar.sun.ac.za

119
scored the amount of spermatozoa bound for each hemizona assay. Sperm-zona
binding was also calculated for matched hemizonae assay results and expressed as
a mean percentage or Hemizona Index (HZI) where HZI = Test sperm bound to hemizona/control
sperm bound to hemizona X 100.
Statistical Analysis
Sperm-zona binding results were expressed as the mean number of sperm bound
per hemizona. The Mann-Whitney U test for nonparametric data was used to
compare these binding results. p-values equal or less than 0.05 were considered
statistically significant. A Hemizonae Index was also calculated for matched
hemizonae assay results and expressed as a mean percentage. A HZI of 100%
implies that the compound tested did not interfere with the amount of spermatozoa
that bind to the ZP as compared to that of the control spermatozoa.
Results
Since the acrosome plays an important role during the binding and penetration of the
ZP, the zona-binding capacity of PD098059 treated sperm populations was recorded.
Table 1 illustrates the sperm-zona binding data. Results were obtained after
PD098059 treatment of sperm followed by exposure to solubilized human ZP or
calcium ionophore (A23187). Significantly more of the PD098059 treated
spermatozoa bound to the ZP (mean±SE) as compared to control spermatozoa, after
exposure to solubilized ZP (26.5±3.7 Vs. 13.8±2.8; p<0.05; Table 1). No significant
differences in binding to the ZP were observed between PD098059 treated and
untreated sperm populations after A23187 exposure.
Stellenbosch University http://scholar.sun.ac.za

120
Table 1. Sperm-zona binding results after PD098059 treatment followed by exposure
to Calcium lonophore (A23187) and solubilized Zona pellucida (ZP).
Control hemizona assay Test hemizona assay
Spermatozoa treated with different
components alone
Spermatozoa exposed to components
after PD098059 treatment
Component Mean (±SE) no. of
zona-bound
sperm
Component Mean (±SE) no. of
zona-bound
sperm
Culture medium 53.4±10.58 Culture medium 43.3±8.9
(n=15)
A23187
(n=12)
0.67ZP/IlI
(n=12)
13.8±2.8b
(n=12)
A23187
(n=12)
0.67ZP/IlI
(n=12)
22.2±2.534.7±10.2
26.5±3.7C
a Vs. b, p<0.005; b Vs. c, p<0.05
The results of the sperm zona binding of matched hemizonae are expressed as a HZI
in Table 2. It is evident that the MEK-inhibitor, PD098059, did not influence sperm-
zona binding per se as a HZI of 107% was recorded when compared to the binding
potential of control sperm (Table 2. A). A 14% increase in HZI was observed between
PD098059-treated and untreated spermatozoa, after ZP stimulation prior to sperm-
zona binding, when compared to that of their respective control hemizona assays
(Table 2. 0 = 28% Vs Table 2. E = 42%). PD098059 treated sperm populations
exposed to solubilized ZP bound more to each hemizonae as compared to
spermatozoa incubated in a solution containing only ZP (HZI = 240%; Table 2. G).
No real differences could be detected in sperm-zona binding between control and
Stellenbosch University http://scholar.sun.ac.za

121
P0098059-treated spermatozoa after A23187 stimulation when compared to that of
their respective control hemizona assays (Table 2. B = 57% Vs. Table 2. C = 54%).
Table 2. Sperm-zona binding results expressed as a Hemizona Index (HZI) after
P0098059 (PO) treatment followed by exposure to Calcium lonophore (A23187) and
solubilized Zona pellucida (ZP).
# Control hemizona assay Test hemizona assay HZI n
A Culture medium PO + Culture medium 107% 5
B Culture medium A23187 57% 5
C Culture medium PO + A23187 54% 5
0 Culture medium ZP 28% 7
E Culture medium PO+ZP 42% 5
F A23187 PO + A23187 107% 4
G ZP PO+ZP 240% 5
H PO + Culture medium PO+ZP 41% 6
I PO + Culture medium PO + A23187 50% 5
(HZI = es IControl X 100)
Discussion
The present results illustrate the involvement of extracellular regulated kinase in the
ZP stimulated AR (or physiological relevant exocytotic event) as opposed to the non-
physiological A23187 induced event. The increase in binding of P0098059 pre-
treated spermatozoa to the oocyte (13.8±2.8 Vs. 26.5±3.7 and HZI 28% Vs. 42%) is
proof that the ZP induced AR was indeed inhibited and underline the possible
regulatory effect of P0098059 and its subcellular targets on the ZP-induced AR.
Stellenbosch University http://scholar.sun.ac.za

122
PD098059 is a specific inhibitor of the activation of the ERK kinase (MEK), the kinase
upstream of ERK, responsible for the dual phosphorylation and activation of ERK
following G-protein coupled- and protein tyrosine kinase receptor ligation and
activation, both in vitro and in vivo (Alessi et ai., 1995). Aitken et al. (1996) showed
that the addition of genistein, a tyrosine kinase inhibitor, inhibited capacitation,
probably by inhibiting the protein tyrosine kinase ZP receptor, while the functional
inactivation of the Gi-protein receptor by pertussis toxin inhibited the ZP-induced AR
which appear to be mediated through a G-protein mediated signal transduction
process (Bastiaan et ai., 1999). Luconi et al. (1998) also showed that in vitro
capacitation stimulates a sustained and concomitant increase in tyrosine
phosphorylation and kinase activity of ERK; this activation of ERK was strongly
inhibited when capacitation was performed in the presence of the MAPK cascade
inhibitor PD098059 (Luconi et al., 1998).
PD098059 also has effects seemingly unrelated to the inhibition of ERK, as shown by
its functional inactivation of voltage-dependent calcium (Ca2+) channels that
participate in fertilization of the marine worm, Urechis caupo (Gould & Stephano,
2000). Voltage operated calcium channels can be classified into low voltage-
activated and high voltage-activated calcium channels on the basis of thresholds for
activation. High voltage-activated Ca2+ channels having high thresholds can be
further classified by their pharmacological properties into L-, N-, P/Q-, and R-types
(Birnbaurner et ai., 1994; Dunlap et ai., 1995). Widely used high voltage operated
calcium channel antagonists can selectively block these channels. Stimulation of
sperm with ZP depolarises the sperm membrane potential. ZP (specifically ZP3)
stimulation activates a depolarisation mechanism with the characteristics of a poorly
Stellenbosch University http://scholar.sun.ac.za

123
selective cation channel. Pre-treatment of sperm with PD098059 possibly prevents
activation of the Ca2+ -selective channel by ZP3/zonae pellucidae as is the case with
pertussis toxin treatment (Florman et ai., 1995). The results can be interpreted to
support the idea that the ZP-induced AR is the physiologically relevant exocytotic
event since it is the ZP-induced AR, and not the spontaneous (culture medium) or
A23187 induced AR, which appears to be mediated through a ERK-mediated signal
transduction process.
Although it is generally accepted that the spermatozoa must be acrosome-reacted to
complete penetration of the ZP (Franken et al., 1991), the exact site of the AR has
not been defined and appears to differ between species. PD098059 treatment of
human spermatozoa does not affect the ability of spermatozoa to bind to structural
intact human ZP. The results indicate the importance of intact acrosomes on the
spermatozoa to ensure tight binding to the ZP, i.e., those sperm populations with a
decreased AR; namely, the PD098059 treated spermatozoa bound significantly
higher numbers of sperm during HZA conditions (Du Plessis et al., In press).
Acknowledgements
This study was supported in part by the Technology and Human Resources for
Industry Programme (THRIP), managed by the National Research Foundation (NRF)
and funded by the Department of Trade and Industry (DTI).
Stellenbosch University http://scholar.sun.ac.za

124
References
Aitken RJ, Buckingham OW, Harkiss 0 (1996) The extragenomic action of human
spermatozoa is influenced by redox regulated changes in tyrosine
phosphorylation during capacitation. Mol Cell Endocrinol 17: 83-93.
Alessi DR, Cuenda A, Cohen P, Dudley OT, Saltiel AR (1995) PD098059 is a
specific inhibitor of the activation of mitogen-activated protein kinase kinase in
vitro and in vivo. J Bioi Chem 270:27489-27494.
Arnouit C, Cardullo RA, Lemos JR (1996) Activation of mouse sperm T-type Ca2+
channels by adhesion to the egg zona pellueida. Proe Natl Acad Sci USA
93:13004-13009.
Babcock OF, Pfeiffer DR (1987) Independent elevation of cytosolic [Ca2+] and pH of
mammalian sperm by voltage-dependent and pH-sensitive mechanisms. J Bioi
Chem 262:15041-15047.
Bastiaan H, Franken DR, Wranz P (1999) G-protein regulation of the solubilised
human zona pellueida-mediated acrosome reaction and zona pellueida binding.
JAssist Reprod Gen 16(6):332-336.
Bielfield PP, Faridi A, Zaneveld LJD, de Jonge CJ (1994) The zona pellueida-indueed
acrosome reaction of human spermatozoa is mediated by protein kinases. Fertil
Steril 61: 536-541.
Birnbaumer L, Cainpbell KP, Catterall WA (1994) The naming of voltage-gated
calcium channels. Neuron 13:505-506.
Blackmore RF, Eisoldt S (1999) The neoglycoprotein mannose-bovine semen
albumin, but not progesterone, activates T-ty" calcium channels in human
spermatozoa. Mol Hum Reprod 5:498-506.
Stellenbosch University http://scholar.sun.ac.za

125
BrandeIIi A, Mimda PV, Tezon JG (1996) Voltage-dependent calcium channels and
Gi regulatory protein mediate the human sperm acrosomal exocytosis induced
by N-acetylglucosaminyllmannosyl neoglycoproteins. JAndrol 17:522-529.
Canman CE, Kastan MB (1996) Three paths to stress relief. Nature 384:213-214.
Cross NL, Morales P, Overstreet JW, Hanson FW (1988) Induction of acrosome
reactions by the human zona pellucida. Bioi Reprod 38:235-244.
Du Plessis SS, Page C, Franken DR (2001) The Zona pellucida-induced Acrosome
Reaction of Human Spermatozoa involves Extracellular Signal-Regulated
Kinase activation. Andrologia, 33(6):337-342.
Dunlap K, Luchke Jl, Turner TJ (1995) Exocytotic Ca2+ channels in mammalian
central neurons. Trends Neurosci 18:89-98.
Florman H.M. (1994) Sequential focal and global elevations of sperm intracellular
calcium are initiated by the zona pellucida during acrosomal exocytosis. Dev
Bioi 165:152-164.
Florman HM, Corron ME, Kim TDH (1992) Activation of voltage- dependent calcium
channels of nummalian sperm is required for zona pellucida-induced acrosomal
exocytosis. Dev Bioi 152:304-314.
Foresta C, Rossatao M, Di Virgilio F (1993) Ion fluxes through the progesterone-
activated channel of the sperm plasma membrane. Biochem J 294:279-283.
Franken DR, Bastiaan HS, Oehninger SC (2000) Physiological induction of the
acrosome reaction in human sperm: Validation of a micro-assay using minimal
volumes of solubilised, homologous zona pellucida. JAssist Reprod Gen
17(3):156-161.
Stellenbosch University http://scholar.sun.ac.za

126
Franken DR, Morales P, Habenicht UF (1996) Inhibition of G-protein in human sperm
and its influence on acrosome reaction and zona pellucida binding. Fertil Steril
66:1008-1011.
Franken DR, Oosthuizen WT, Cooper S, Kruger TF, Burkman LJ, Coddington CC,
Hodgen GO (1991) Electronmicroscopic evidence on acrosomal status of bound
sperm and their penetration into human hemizonae pellucida after storage in
buffered salt solution. Andrologia 23:205-208.
Gould MC, Stephano JL (2000) Inactivation of Ca2+ action potential channels by the
MEK inhibitor PD98059. Exp Cell Res 260(1):175-179.
Hammit DG, Syrop CH, Walker DL, Bennet MR (1991) Conditions of oocyte storage
and use of non inseminated, non-fertilized oocytes for the hemizona assay.
Fertil SteriI60:131-136.
Kruger TF, Menkveld R, Stander FSH, Lombard CJ, Van der Merwe JP, Van Zyl JA,
Smith K (1986) Sperm morphologic features as a prognostic factor in in vitro
fertilization. Fertil Steril 46: 1118-1123.
Liu DY, Baker HWG (1996) A simple method for assessment of the acrosome
reaction of spermatozoa bound to the zona pellucida: lack of relationship with
ionophore A23187-induced acrosome reaction. Hum. Reprod. 11:551-557.
Luconi M, Barni T, Vanelli GB, Krausz C, Marra F, Benedetti PA, Evangelista V,
Francavilla S, Properzi G, Forti G, Baldi, E (1998) Extracellular regulated signaI-
kinases modulate capacitation of human spermatozoa. Bioi Reprod 58: 1476-
1489.
Morales P, Cross NL, Overstreet JW, Hanson FW (1989) Acrosome-intact and
acrosome-reacted human sperm can initiate binding to the zona pellucida. Dev
Bioi 133: 385-392.
Stellenbosch University http://scholar.sun.ac.za

127
Page C, Doubell AF (1996) Mitogen-activated protein kinase (MAPK) in cardiac
tissue. Mol Cell Bio 157:49-57.
Quin P, Kerin JF, Warnes GM (1985) Improved pregnancy rate in human in vitro
fertilization with the use of a medium base on the composition of human tubal
fluid. Fertil Steril 44:493-498.
Son W-Y, Lee J-H, Han C-H (2000) Acrosome recation of human spermatozoa is
mainly mediated by alfa1H type calcium channels. Mol Hum Reprad 10, 893-
897.
World Health Organisation (1999) Collection and examination of human semen. In:
WHO Laboratory Manual for the Examination of Human Semen and Semen-
Cervical Mucus Interaction. 4th ed. Cambridge University Press, London, pp4-
33.
Yanagimachi R (1994) Mammalian Fertilization. In: The Physiology of Reproduction.
Vo11, 2nd ed. Knobil E, Neil JD (eds). Raven Press, New York, pp 189-317.
Stellenbosch University http://scholar.sun.ac.za

"Every great advance in science has
issued from a new audacity of
imagination. "
- Joh n Dewey -
Stellenbosch University http://scholar.sun.ac.za

128
CHAPTER 6
Phosphatidylinositol 3-kinase inhibition enhances human sperm
motility and sperm-zona pellucida binding.
Archives of Andrology 2002, submitted
SS du Plessis 1,·, DR Franken2, M Luconf and E Baldi3
1Department of Medical Physiology and Biochemistry, University of Stellenbosch,
P.O.Box 19063, Tygerberg, 7505, South Africa.
2Department of Obstetries and Gynaecology, University of Stellenbosch, Tygerberg
Hospital, Tygerberg, 7505, South Africa.
3Dipartimenti di Fisiopatologia Clinica, Unita di Andrologia, Universitá di Firenzi, viale
Pieraccini 6, 1-50139,Firenzi, Italy
Ruqning head: PI3-Ks effect on sperm motility and binding
Keywords: spermatozoa, acrosome, PI3-kinase, zona pellucida, hemizona
'Corresponotnq author. SS du Plessis, Department of Medical Physiology and
Biochemistry, University of Stellenbosch, P.O.Box 19063, Tygerberg, 7505, South
Africa; Tel.: +27-(0) 21-938 9388; fax: +27-(0) 21-938 9476; e-mail:
Stellenbosch University http://scholar.sun.ac.za

129
Summary
Various signalling pathways are involved in the regulation of sperm motility,
capacitation, acrosome reaction and sperm-zona binding. Recent data pointed out an
important role for phosphatidylinositol 3-kinase (PI3K) in human sperm motility,
however no studies as of yet has been done to determine the effect of this inhibitor
on other sperm parameters. Here we investigated the role of PI3K on human sperm
motility, acrosome reaction and sperm-oocyte binding by using the specific PI3K
inhibitor LY294002. We demonstrate that in vitro incubation of washed unselected
spermatozoa with LY294002, increased the percentage motility and progressive
motility in asthenozoospermia patients as evaluated by computer-aided sperm
analysis. The compound furthermore did not influence the acrosome reaction, whilst
it further did enhance sperm-oocyte binding. Our results therefore imply that PI3K
negatively affect sperm motility and oocyte binding. We subsequently suggest a
possible therapeutic role for PI3K inhibitors in the treatment regime for
asthenozoospermia.
Introd uction
Spermatozoon prerequisites for initiation of fertilization include motility, capacitation
and acrosome reaction. Normal motility patterns are needed to deliver the sperm to
the site of fertilization, while concurrently it also has to undergo a series of functional
biochemical and biophysical modifications named capacitation in order to render the
ejaculated spermatozoa competent for fertilization of the oocyte. Finally a timeous
acrosome reaction, which is an exocytotic process physiologically, induced by ligand
(ZP3)-receptor interaction, ultimately lead to sperm-zona binding.
Stellenbosch University http://scholar.sun.ac.za

130
Many patients who attend infertility clinics are diagnosed with asthenozoospermia
that result in impaired fertilization rates in vivo as well as during in vitro procedures
(Oehninger, 2001). Even with the onset of advanced assisted reproductive
techniques like in vitro fertilization (IVF) and intracytoplasmic sperm injection (ICSI),
the quest to increase the fertilizing ability of semen samples is a very relevant and
ongoing process. It is expected that simplified and more cost-efficient therapeutic
modalities will be developed as additional basic (cellular-molecular) and clinical
knowledge is gained (Edirisinghe et a/., 1995; Oehninger, 2001). It is therefore one of
the goals of reproduction physiologists to try and increase the motility of spermatozoa
among asthenozoospermic patients without interfering with the normal physiological
processes needed for fertilization.
Various studies have demonstrated that several pharmacological agents such as
SpermSolute (based on a proteinase - trypsin) and pentoxifylline can definitely
improve sperm motility (Tesarik et a/., 1992; Krausz et a/., 1994; Lanzafame et a/.,
1994; Minhas & Ripps, 1996; Figenschau & Bertheussen, 1999; Terriou et a/., 2000).
However, the possibility of using such agents is limited due to the presence of non-
responding subjects (Krausz et a/., 1994) and the possible toxic effects of these
molecules (Centola et a/., 1995)
Even though it is known that phosphatidylinositol 3-kinase (PI3K) is present in mouse
spermatozoa (Feng et a/., 1998), it was only recently demonstrated that LY294002,
the specific inhibitor of PI3K, increased the progressive motility of spermatozoa in
humans (Luconi et a/., 2001). Dolci et al. (2001) reported that stem cell factor (SCF)
acts as a mitogenic factor in cultured c-kit-expressing spermatogonia and that both
Stellenbosch University http://scholar.sun.ac.za

131
mitogen-activated protein kinase kinase (MEK)- and Pl3K-dependent pathways are
required for the proliferative response. The mitogenic effect is not accompanied by an
increase in total cellular amount of cyclin D3, but it is associated with a rapid change
in its subcellular localisation. It was also shown that SCF is an anti-apoptotic factor
for spermatogonia, but the MEK- or the Pl3K-dependent pathways are not sufficient
on their own to promote the survival response. It was further suggested that the
binding of c-kit to SCF in mature sperm cells may result in the activation of PLCy1
and PI3K, leading to receptor autophosphorylation and ultimately may playa role in
capacitation and/or the acrosome reaction (Feng et al., 1998).
PI3K's are SH2 domain-containing proteins, which is known to be important in
phosphoinositide-mediated intracellular signalling pathways in numerous cell types.
And is heterodymeric, consisting of a p85 regulatory (adapter) subunit and a p110
catalytic subunit. It phosphorylates the 3'-OH group of the inositol ring in inositol
phospholipids and the most important of these is thought to be the conversion of
phosphatidylinositol (Ptdlns)-4,5-bisphosphate to Ptdlns-3,4,5-trisphosphate (Fisher
et al., 1998). PI3K's have been identified in most cells studied. It has been implicated
in the priming of a series of signalling cascades involved in the regulation of many
processes of somatic cells including mitogenesis, differentiation, motility, cell survival,
oocyte maturation, cell chemotaxis, membrane ruffling, DNA synthesis, receptor
internalisation, vesicular trafficking and metabolic control (KapelIer & Cantley, 1994;
Vanhaesebroeck et ai., 1997; Wymann & Pirola, 1998; Anderson et ai., 1999). It has
also been suggested that PI3K might possibly be involved in the AR elicited by the
mannose-BSA agonist by making use of the unrelated PI3K inhibitor, wortmannin
(Fischer et al., 1998).
Stellenbosch University http://scholar.sun.ac.za

132
As of yet no studies have been performed to test the effect of this inhibitor on the ZP
induced AR as well as sperm-zona binding potential. The study aimed to investigate
the role of PI3Ks in human sperm motility, AR and sperm-zona binding by using the
PI3K inhibitor, LY294002.
Materials and Methods
Preparation of sperm samples
Semen samples were collected from volunteers (average age = 25.81±1.09 years;
n=18) and analysed and classified according to the WHO criteria (WHO, 1999).
Only normozoospermic samples were used to perform AR and sperm-oocyte binding
tests. Motile sperm fractions were collected after a double wash in synthetic human
tubal fluid culture medium (HTF) (Quin et a/., 1985) and 60-minute swim-up
technique in HTF supplemented with 3% bovine serum albumin (BSA; Seravae, Cape
Town, South Africa). Retrieved sperm samples were resuspended in synthetic HTF-
BSA to a sperm concentration of 1Ox106 cells/ml. These fractions were then
capacitated in the presence or absence of the PI3K-inhibitor, LY294002 (LY; 101JM;
120min; 37°C, 5% CO2 and 95% humidity).
The semen samples used to perform computer-aided sperm analysis (CASA)
underwent a double wash in HTF. The pellet containing the spermatozoa was then
resuspended to its original volume in HTF + BSA. After preparation these samples
Stellenbosch University http://scholar.sun.ac.za

133
were incubated (37°C, 5% CO2 and 95% humidity) for 2h in the presence or absence
of the PI3K antagonist, lY294002 (10j.lM; 120min).
Preparation of solubilized ZP
Oocytes were retrieved from post mortem derived ovarian material. Approval was
obtained from the Institutional Review Board and fully complies with the legal and
ethical guidelines of the South African Medical Research Council. Retrieved oocytes
were stored in a dimethylsulfoxide/sucrose solution at -196°C in liquid nitrogen
(Hammit et al., 1991). Twenty four hours prior to each test, oocytes were removed
from storage and thawed at 37°C. Retrieved oocytes were placed in 0.25M sucrose
and 3% BSA in HTF. On the day of the experiment, 20 oocytes were placed in a drop
on a petri dish, after which the HTF was removed under microscopic vision (Olympus
SZ40; Wirsam Scientific, Cape Town, South Africa), leaving only the 20 oocytes on
the petri dish. A total volume of 5f..l1of 10mM HCl was then added to the oocytes on
the petri dish; solubilization of the ZP was microscopically observed and controlled.
Following solubilization, 5f..l1of 10mM NaOH was added to the solubilized ZP, to
render a final zona volume of 10f..l1containing 2 ZP/f..ll(Du Plessis, 2001). The final ZP
concentration, after the addition of spermatozoa, was 0.67 ZP/f..ll.
Sperm kinematics
The samples prepared for motility studies were analysed using a Hamilton-Thorne
(Hamilton-Thorne Research, Beverly, MA) IVOS analyser (integrated visual optical
system) with standard set-up parameters. Washed spermatozoa were resuspended
in a capacitation medium containing HTF with 3% BSA. Computer assisted semen
analysis (CASA) was performed on each sample at 3 hours after collection using a
Stellenbosch University http://scholar.sun.ac.za

134
10.0!J.mchamber (Makler) and a sampling rate of 30 frames at 60Hz. A minimum of
100 cells and four fields was analysed for each aliquot. All analyses were performed
at 37°C.
Acrosome reaction studies
Acrosomal statuses of spermatozoa stimulated with 0.67ZP/!J.1for 30 minutes in
either the presence or absence of LY294002 (pre-treatment) were determined
according to procedures published elsewhere (Cross et al., 1988; Morales et ai.,
1989, Franken et ai., 2000) and compared to that of control samples. These
stimulation procedures were performed as micro-assays as previously described
(Franken et ai., 2000).
Spermatozoa from the different experiments were fixed and air-dried, after which the
acrosomal status was determined using fluorescein-labeled Pisum Sativum agglutinin
(FITC-PSA; 125!J.g/ml;L-0770; Sigma, Cape Town, South Africa). Two individuals
scored a minimum of 200 spermatozoa blindly for each different sample. The
following staining patterns were evaluated as acrosome reacted spermatozoa: (i)
patchy staining on acrosomal region, (ii) distinct staining in the equatorial region
occurring as an equatorial bar and (iii) no staining over the entire sperm surface.
Hemizona Assays/Zona Pellucida Binding
Both LY294002 pre-treated (test) and control sperm samples (15!J.1droplets;
1Ox106sperm/ml) were placed on a petri dish to which hemizonae were added in a
match-controlled fashion. Duplicate hemizona assays (HZA) were performed and
sperm-zona binding was assessed after four hours of co-incubation. Following the
Stellenbosch University http://scholar.sun.ac.za

135
co-incubation period, hemizonae were removed and washed (5x) to strip the loosely
attached spermatozoa. Hemizonae were then evaluated, while the number of
spermatozoa tightly bound to the ZP was recorded for each test and matching
control. Experiments where less than 20 spermatozoa bound to a control hemizona
were discarded. Sperm-zona binding was expressed as a percentage or Hemizona
Index (HZI) where HZI - Test sperm bound to hemizonal X 100 A HZI of- Control sperm bound to hemizona .
100% would therefore imply that the compound tested did not interfere with the
amount of spermatozoa that bind to the ZP as compared to that of the control
spermatozoa.
Statistical Analysis
Sperm kinematics results, percentage acrosome reacted sperm and numbers of
sperm bound to the hemizonae are expressed as mean±SE. Student's t-test for
paired data was used to compare the results of the motility, acrosome and hemizona
studies. Pearson's test was used for correlation tests when comparing different
motility parameters. P-values equal or less than 0.05 were considered statistically
significant.
Results
Sperm kinematics
Overall semen samples (n=18) showed a statistically significant increase in
progressive motility after co-incubation with the PI3K inhibitor, LY294002,
(31.88±3.75% vs. 37.31±3.74%, P=0.020). A complete shift in velocity distribution
also occurred. According to CASA more of the static, slow and medium cells became
Stellenbosch University http://scholar.sun.ac.za

136
activated and the rapid population increased from 43.63±4.80% in the control
samples to 50.38±4.20% in the LY294002 pre-treated samples (P=0.025). The rest of
the sperm kinematics parameters did not change statistically significantly after
LY294002 treatment in the donor population as a whole.
The results were subsequently divided into two groups (Table 1) based on the %
motility of each donor's sperm sample. The groups were classified as >50% motility
(normozoospermic) and <50% motility (asthenozoospermic) according to WHO
(1999) criteria. The effect of PI3K inhibition is much more pronounced in the
asthenozoospermic group compared to the normozoospermic group. The percentage
motile and percentage progressive motile spermatozoa increased statistically
significantly by 22% (from 29.25% to 51.25%; P=0.009) and 10.25% (from 15.00% to
25.25%; P=0.008) respectively after LY294002 treatment (Table 1). The velocity
distribution results of this group also showed a statistically significant change with the
rapid moving population increasing by 15.25% (from 21.50% to 36.75%; P=0.010)
and the static population decreasing by 22% (from 70.75% to 48.75%; P=0.009).
An inverse effect occurred in the amplitude of lateral head displacement (ALH)
between the two groups after LY294002 treatment. (The normozoospermic
populations ALH increased from 4.23±0.17lJm to 4.44±0.14IJm (P=0.046) while the
asthenozoospermic populations decreased from 5.00±0.43IJm to 4.38±0.30IJm
(P=0.017)). When a" of the control and LY pre-treated CASA results were pooled and
analysed together a positive correlation (r=0.6487; P<0.0001; ~=0.4208; n=36) was
found between the progressive motility values and the beat cross frequency (BCF)
values (Figure 1). No correlation existed between progressive motility and ALH, while
Stellenbosch University http://scholar.sun.ac.za

137
upon comparing BeF to ALH (Figure 2) a negative correlation was found (r=-O.3462;
P=O.026; ~=O.1199) in this group.
Table 1. Sperm kinematics results of all the samples as well as when divided into
normozoospermic and asthenozoospermic donors in the presence and absence of
LY294002
All samples >50% Motility <50% Motility P
Control LY Control LY Control LY(n=18) (n=18) (n=12) (n=12) (n=6) (n=6)
Motile 62.87±6.14 69.06±4.69 74.08±3.62 75.00±4.38 29.25±1O.44 51.25±9.38 * 0.009(%) * *Progressive 31.88±3.75 37.31±3.74 38.50±3.38 41.33±4.01 15.00±5.48 25.25±6.26 * 0.020(%) * * # # # 0.008
Path velocity 64.23±2.86 65.31±2.50 65.70±2.87 68.23±2.36 59.78±7.99 56.58±5.49(VAP, um/s)
Progressive velocity 54.85±3.09 55.54±2.59 56.71±2.90 58.39±2.28 49.25±9.14 47.00±6.75(VSL, urn/s)
Track speed 100.89±4.07 105.09±2.88 101.36±4.84 106.84±3 .28 99.48±8.54 99.83±5.91(VCL, um/s)
Lateral amplitude 4.43±0.18 4.43±0.12 4.23±0.17 4.44±0.14 5.00±0.43 4.38±0.30 * 0.046(ALH, urn) * * # # #0.017
Beat frequency 26.20±0.97 27.01±1.08 27.17±1.09 27.08±1.25 23.48±1.64 26.80±2.43(BCF, Hz)
Straightness 82.56±146 82.81±1.31 83.50±0.99 83.50±1.01 79.75±5.31 80.75±4.60(STR, %)
Linearity 54. 13±1.96 53.31±1.97 55.83±1.54 55.00±1.50 49.00±6.23 48.25±6.52(LIN, %)
Elongation 69.44±0.94 68.00±0.63 68.33±0.51 67.67±0.75 72.75±3.12 69.00±1.08(%)
Area 4.35±0.15 4.16±0.12 4.42±0.16 4.16±0.14 4.15±0.41 4.18±0.25(urn sq)
Rapid 43.63±4.80 50.38±4.20 51.00±4.11 54.92±4.28 21.50±7.51 36.75±8.26 * 0.025(%) * * # # # 0.010
Static 37.13±6.14 30.88±4.68 25.92±3.61 24.92±4.35 70.75±10.44 48.75±9.38 * 0.009(%) * *
Stellenbosch University http://scholar.sun.ac.za

138
35-N 30:r;-u,0 2510
200
• • •• •- -----+ - .• •
• •10 20 30 40 50 60 70
PM (%)
Figure 1. Correlation between beat cross frequency (BCF) and progressive motility
(PM) of pooled experiments (n=36). (r=0.6487; P<0.0001; 1=0.4208)
35 ~--------------------------------~-N 30:r;-
•• • •• •_.-+- .~- _.....~-• •••
••• •
u,o 2510
203 4 5
ALH (J.lm)
6 7
Figure 2. Correlation between beat cross frequency (BCF) and amplitude of lateral
head displacement (ALH) of pooled experiments (n=36). (-r=0.3462; P=0.026;
1=0.199)
Stellenbosch University http://scholar.sun.ac.za

139
Acrosome reaction
Due to the scarcity of human ZP, the effect of LY294002 on the spontaneous AR or
the ZP-induced physiologically relevant exocytotic event was only determined in a
limited number of normozoospermic sperm samples. ZP stimulation increased the
%AR by a statistically significant 106.1% (P=0.017) in the control sample (from
10.18±1.924% to 20.98±1.805%), while a statistically significant increase of 112.3%
was recorded in the LY pre-treated group (from 8.525±2.234 to 18.1±1.882). Though
decreases in AR were detected in spontaneous (less 16.3%) and ZP induced AR
(less 13.7%) after LY pre-treatment, it was not of significant value. It can thus be
deducted that exposure of spermatozoa to the PI3K inhibitor, LY294002, did not
influence the spontaneous or ZP induced acrosome reactions during these
experiments (n=5; Figure 3).
Stellenbosch University http://scholar.sun.ac.za

140
25
20
0::: 15«:::e 100
5
0C ZP LY ZP+LY
Figure 3. Histogram showing the percentage acrosome reaction (mean±SE) of
control (C) spermatozoa and spermatozoa after exposure to zona pellucida (ZP),
PI3K antagonist LY294002 (LY) or both ZP and LY294002 JZP+L Y). LY did not inhibit
or increase the spontaneous or ZP induced acrosome reactions (n=5). IC vs. ZP
(P=O.017); C vs. LY (P=O.60B); C vs. ZP+L Y (P=O.007), ZP vs. LY (P=O.009); ZP vs.
ZP+LY (P=O.323); LVvs. ZP+L Y (P=O.019)]
Stellenbosch University http://scholar.sun.ac.za

141
Sperm-zona pellucida binding
A statistically significant difference (P=0.024) could be detected between the number
of LY294002 pre-treated spermatozoa (49.72±2.7; range 29.0-80.0 and median =49.5) and control spermatozoa (45.11±2.6; range 28.0-70.0 and median = 44.0) that
bind to the hemizonae's respectively (Figure 4). The ratio between LY pre-treated
(test) and untreated (control) samples produced a HZI of 112% ± 5%. This suggests
a trend that more LY pre-treated spermatozoa did bind to the hemizona.
ca 600 *N 500... 40caE... 30CJ)c.f/) 20....0 10.0Z 0
Control Test
Figure 4. Histogram showing the number of control and LY294002 pre-treated (Test)
spermatozoa (mean±SE) tightly bound to each hemizona respectively (n=18). (*
P=0.024 vs. control)
Discussion
Controversies do exist regarding sperm motility enhancers and their effects on sperm
viability and fertilizing ability. In the present study it was demonstrated that the
addition of LY294002, a very specific inhibitor of phosphatidylinositol 3-kinase, is able
Stellenbosch University http://scholar.sun.ac.za

142to increase the motility and progressive motility of human spermatozoa. These results
are strongly supported by similar studies performed by luconi et al. (2001). We
specifically show that the addition of l Y294002, to double washed sperm samples,
increase progressive motility and the percentage rapid cells in our general donor
population. In addition we demonstrate that our results are even more exaggerated in
the asthenozoospermic population. Not only did the progressive motility and rapid cell
population increase dramatically, but also the percentage motile cells increased while
percentage static cells decreased markedly. luconi et al. (2001) also showed
increases in Vel, VAP and VSl (and other motion parameters) as measured by
eASA. We however did not find any significant increases in these parameters but did
see increasing trends in these parameters among all our sperm populations. Not
finding statistic significance in this study regarding the above mentioned parameters
can be ascribe to making use of smaller sample populations as well as differences in
preparational techniques (e.g. centrifuged, washed, unselected sperm vs. swim-up
selected sperm). The molecular mechanisms underlying the stimulatory effects of
l Y294002 on sperm motility are still poorly understood. The process of development
and maintaining of motility in mammalian spermatozoa is rather complex and
involves the integration of and crosstalk of several signalling pathways, including
adenylate cyclase/cAMP/PKA, calcium and phosphorylation/dephosphorylation of
proteins (Tash and Bracho, 1994). luconi et a/., (2001) speculate that in
spermatozoa PI3K might be involve in phosphorylating or dephosphorylating proteins
involved in motility or in the generation of reactive oxygen species (ROS) which has a
negative impact on motility.
It can also be deducted from our results that BeF is closely related to progressive
motility and thereby an increase in BeF might lead to an increase in progressive
Stellenbosch University http://scholar.sun.ac.za

143motility. It was also shown that BCF and ALH correlates negatively. It can thus be
further deducted that in order for BCF to increase, there are less time for the sperm
head to be displaced sideways and thereby decreasing ALH.
Knowing that the PI3K signalling pathway is involved in eliciting of the AR (Fisher et
a/., 1998), it was therefore important to investigate the effects of LYon this reaction.
In our hands LY294002 did not affect the spontaneous or the ZP induced AR even
though a slight decline in AR was observed in all 5 experiments between the ZP and
the LY+ZP groups. Furthermore no significant (P=0.163) difference in AR was
observed between progesterone stimulated spermatozoa with or without LY
treatment (results not shown). Luconi (personal communication) also found that LY
exerts no effect on both the spontaneous and progesterone stimulated AR, thereby
supporting the current data. Contrary to our findings, Fischer et al. (1998) showed a
decrease in the AR elicited by the mannose-BSA agonist by making use of the
unrelated PI3K inhibitor, wortmannin. Yet wortmannin was found not to inhibit the
acrosome reaction induced by either A23187 or progesterone (Fischer et ai., 1998).
This may imply that the cellular pathways involved in bringing about the acrosome
reaction induced by the different agonists (ZP3, Progesterone, Mannose-BSA,
A23187) do not involve PI3K to the same extend, or perhaps that the need for PI3K
in the pathway is somehow bypassed. It may also imply that LY294002 and
wortmannin have different mechanisms of action or differ in their specificity to inhibit
PI3K. In this regard wortmannin has indeed been shown to inhibit other enzymatic
activities (Cross et a/., 1995) including MAPK and extracellular signal regulated
kinases (ERK) at doses from 250nM (Ferby et ai., 1996). The presence of ERK has
been demonstrated in human spermatozoa (Luconi et al., 1998) and implicated to be
involved in the ZP-induced AR (Du Plessis et ai., 2001). It can therefore be
Stellenbosch University http://scholar.sun.ac.za

144speculated that the decrease in AR in wortmannin treated samples after exposure to
the mannose-BSA agonist may not only be due to PI3K inhibition, but also due to the
possible simultaneous inhibition of ERK. However as half-maximal inhibition of MAPK
activation occurs at 300nM this must be ruled out as Fisher et al. (1998) used the
inhibitor at concentrations ranging between 10nM and 100nM.
When considering that the different agonists (i.e. ZP3, Progesterone, Mannose-BSA)
all stimulate different signalling pathways through binding to different membrane
receptors, it is obvious that it will result in different cellular responses. From the
literature it is evident that various crosstalk mechanisms are involved in PI3K and
RAS/RAF/ERK signalling. It is known that RAS can both activate RAF/ERK as well as
PI3K (Sears & Nevins, 2002; Krasilnikov, 2000). PI3K on the other hand can also
activate the RAS/RAF/ERK pathway through mitogen signalling (Krasilnikov, 2000)
(see Figure 5). Taking into consideration that mannose-BSA stimulation elicit the AR
through both Gi-protein receptor binding (BrandeIIi et al., 1996) and receptor tyrosine
kinase binding (Fisher et al., 1998), and that PI3K is regulated by receptor tyrosine
kinase activity, it can explain why PI3K inhibition can inhibit the mannose-BSA
induced AR (Fisher et ai., 1998). Wortmannin will inhibit PI3K, thereby also inhibiting
the PI3K activated RAS/RAF/ERK pathway needed for eliciting of the AR (Du Plessis,
2001). There is no evidence in the literature that the mannose-BSA tyrosine kinase
receptor is directly linked to the RAS/RAF/ERK pathway, but only through PI3K. In
our experiments we stimulated the AR with solubilized ZP (ZP3), thereby eliciting the
AR through both a Gi-protein receptor and ZRK binding which directly activates
RAS/RAF/ERK and PI3K through RAS (see Figure 5). Inhibition of PI3K by
LY294002 thus only inhibited the PI3K pathway and not the direct RAS/RAF/ERK
pathway activated independent of PI3K signalling or activation through protein kinase
Stellenbosch University http://scholar.sun.ac.za

145C (PKC). The activation of RAS by ZP binding to ZRK can also stimulate PI3K,
thereby enhancing a possible positive feedback on ERK activation (e.g. via PKC) as
well as other mediators in the process of AR signalling (see Figure 5).
A statistically significant increase in sperm-oocyte binding after LY treatment was
also detected. It was also previously shown in our laboratory that the frequency of
sperm/zona pellucida collision rates has an influence on zona binding (OzgOr &
Franken, 1996). This increase in binding can therefore be ascribed to the fact that
more spermatozoa are motile and progressively motile, thereby increasing the
chances of them colliding with the oocyte. Myles and Primakoff (personal
communication) showed similar results in the mouse model by finding that incubation
of mouse sperm and oocyte in the presence of LY significantly increases sperm-zona
binding.
Compared with signalling pathways in many types of somatic cells, the signal
transduction pathways of the mammalian acrosome reaction (pellucida glycoprotein)
are still very poorly understood (Fisher et al., 1998). One of the chief reasons for this
is that the sperm receptor(s) that bind to zona protein 3 (ZP3) and physiologically
induce the acrosome reaction remain to be conclusively identified (Brewis & Moore,
1997). Indeed, in the human, about ten sperm determinants in spermatozoa have
been reported to be involved (listed and referenced in Benoff, 1997). The most likely
scenario is that several sperm determinants are involved in zona binding and function
in concert as part of a complex (Kopf et al., 1995; Brewis & Moore, 1997).
Our results imply that PI3K negatively regulates sperm motility and increase sperm-
zona binding without influencing the acrosome reaction. This suggests that
Stellenbosch University http://scholar.sun.ac.za

146LY294002 can be used as a tool to enhance the motility of sperm samples during
preparation for IVF from patients with low sperm motility, thereby ultimately opening a
new prospective for severe oligoasthenozoospermic males to enter IVF rather than
ICSI programs.
Stellenbosch University http://scholar.sun.ac.za

147
Mannose-BSA
TKR Plasma membrane
\\\\\\\\\\\\\\\\\\
LY294002Wortmann in
Figure 5. Possible interactions between the different signal transduction pathways
invoked during the acrosome reaction. (ZP3 = zona pellucida glycoprotein 3; ZRK =Zona Receptor Kinase; TKR = mannose tyrosine kinase receptor; dashed lines =hypothesised activation of ERK; G;-coupled receptor pathway not shown)
Stellenbosch University http://scholar.sun.ac.za

148
References
Anderson RA, Boronenkov IV, Doughman SO (1999) Phosphatidylinositol phosphate
kinases, a multifaceted family of signalling enzymes. J Bioi Chem 274:9907-
9910.
Benoff S (1997) Carbohydrates and fertilization: an overview. Mol. Hum. Reprod 3:
599-637
Bielfield PP, Faridi A, Zaneveld LJD, de Jonge CJ (1994) The zona pellucida-induced
acrosome reaction of human spermatozoa is mediated by protein kinases. Fertil
Steril 61: 536-541.
BrandeIIi A, Miranda PV, Tezon JG (1996) Voltage dependent calcium channels and
Gi regulatory protein mediate the human sperm acrosomal exocytosis induced
N-acetylglucosaminyl/mannosyl neogglycoproteins. JAndrol 17: 522-529.
Brewis lA, Moore HOM (1997) Molecular mechanisms of gamete recognition and
fusion at fertilization. Hum Reprod 12 (Natl Suppl, JBFS) 2:156-165.
Centola GM, Cartie RJ, Cox C (1995) Differential responses of human sperm to
varying concentrations of pentoxyfylline with demonstration of toxicity. JAndrol
16:136-142.
Cross MJ, Stewart A, Hodgkin MN, et al. (1995) Wortmannin and its structural
analogue demethoxyviridin inhibit stimulated phospholipase A2 activity in Swiss
3T3 cells. Wortmannin is not a specific inhibitor of phosphatidylinositol 3-kinase.
J Bioi Chem 270: 25352-25355.
Cross NL, Morales P, Overstreet JW, Hanson FW (1988) Induction of acrosome
reactions by the human zona pellucida. Bioi Reprod 38:235-244.
Dolci S, Pellegrini M, Di Agostino S, Geremia R, Rossi P (2001) Signaling through
Extracellular Signal-regulated Kinase Is Required for Spermatogonial
Stellenbosch University http://scholar.sun.ac.za

149Proliferative Response to Stem Cell Factor. J Bioi Chem, 276(43): 40225-
40233.
Du Plessis SS, Page C, Franken DR (2001) The Zona pellucida-induced Acrosome
Reaction of Human Spermatozoa involves Extracellular Signal-Regulated
Kinase activation. Andrologia 33(6): 337-342.
Edirisinghe WR, Junk S, Yovich JM, Bootsma B, Yovich JL (1995) Sperm stimulants
can improve fertilization rates in male-factor cases undergoing IVF to the same
extent as micromanipulation by partial sona dissection (PZO) or subzonal
sperm insemination (SUZI): a randomized controlled study. JAssist Reprod
Genet 12(5): 312-318.
Feng H, Sandlow Jl, Sandra A (1998) The c-kit receptor and its possible signalling
transduction pathway in mouse spermatozoa. Mol Reprod Oev49:317-326.
Ferby 1M,Waga I, Hoshino M, et al. (1996) Wortmannin inhibits mitogen-activated
protein kinase activation by platelet-activating factor through a mechanism
independent of p85/p110-type phosphatidylinositol 3-kinase. J Bioi Chem 271:
11684-11688.
Figenschau Y, Bertheussen K (1999) Enzymatic treatment of spermatozoa with a
trypsin solution, SpermSolute: improved motility and enhanced ATP
concentration. Int JAndrol. Oct;22(5):342-344.
Fischer HM, Brewis lA, Barrat CL et aI., (1998) Phosphoinositide 3-kinase is
involved in the induction of the human sperm scrosome reaction downstream
of tyrosine phosphorylation. Mol Hum Reprod 4:849-855.
Franken DR, Bastiaan HS, Oehninger SC (2000) Physiological induction of the
acrosome reaction in human sperm: Validation of a micro-assay using minimal
volumes of solubilised, homologous zona pellucida. JAssist Reprod Gen
17(3):156-161.
Stellenbosch University http://scholar.sun.ac.za

150Franken DR, Oosthuizen WT, Cooper S, Kruger TF, Burkman LJ, Coddington CC,
Hodgen GO (1991) Electronmicroscopie evidence on acrosomal status of bound
sperm and their penetration into human hemizonae pellucida after storage in
buffered salt solution. Andrologia 23:205-208.
Hammit DG, Syrop CH, Walker DL, Bennet MR (1991) Conditions of oocyte storage
and use of non inseminated, non-fertilized oocytes for the hemizona assay.
Fertil SteriI60:131-136.
KapelIer R, Cantley LC (1994) Phosphatidylinositol 3-kinases. Bioessays, 16:565-
576.
Kopf GS, Visconti PE, Moos J et aI., (1995) Integration of tyrosine kinase and G
protein mediated signal transduction pathways in the regulation of mammalian
sperm function. In Fenichel Pand Parinaud J (eds), Human Sperm Acrosome
Reaction. John Libbey, Montrouge, pp191-216.
Krasilnikov MA (2000) Phosphatidylinositol-3 kinase dependent pathways: the role in
control of cell growth, survival, and malignant transformation. Biochemistry
(Mosc) 65(1): 59-67.
Krausz C, Gervasi G, Forti G, Baldi E (1994) Effect of platelet-activating factor on
motility and acrosome reaction of human spermatozoa. Hum Reprod 9:471-476.
Lanzafame F, Chapman MG, Guglielmino A et al. (1994) Pharmacological stimulation
of sperm motility. Hum Reprod 9:192-199.
Luconi M, Barni, T, Vanelli GB, Krausz C, Marra F, Benedetti PA, Evangelista V,
Francavilla S, Properzi G, Forti G, Baldi E (1998). Extracellular regulated
Signal-Kinases Modulate Capacitation of Human Spermatozoa. Bioi Reprod
58:1476-1489.
Stellenbosch University http://scholar.sun.ac.za

151Luconi M, Marra F, Gandini L, Filimberti E, Lenzi A, Forti G, Baldi E (2001)
Phosphatidylinositol 3-kinase inhibition enhances human sperm motility. Hum
Reprod 16(9):1931-1937.
Minhas BS, Ripps BA (1996) Methods for enhancement of sperm function. Front
Biosci 1:E65-E71.
Morales P, Cross NL, Overstreet JW, Hanson FW (1989) Acrosome-intact and
acrosome-reacted human sperm can initiate binding to the zona pellucida. Dev
Bioi 133: 385-392.
Oehninger S (2001) Strategies for the infertile man. Semin Reprod Med 19(3): 231-
237.
Ola B, Afnan M, Sharif K, Papaioannou S, Hammadieh N, Barrat CL (2001) Should
ICSI be the treatment of choice for all cases of in vitro conception?
Considerations of fertilization and embryo development, cost effectiveness and
safety. Hum Reprod 16(12): 2485-2490.
OzgOr K, Franken DR (1996) The influence of sperm/zona pellucida collision rates on
zona binding. Andrologia 28(5): 261-265.
Quin P, Kerin JF, Warnes GM (1985) Improved pregnancy rate in human in vitro
fertilization with the use of a medium base on the composition of human tubal
fluid. Fertil Steril 44:493-498.
Sears RC, Nevins JR (2002) Signaling networks that link cell proliferation and cell
fate. J Bioi Chem 277(14): 11617-11620.
Simpson JL, Lamb DJ (2001) Genetic effects of intracytoplasmic sperm injection.
Semin Reprod Med 19(3): 239-249.
Tash JS, Bracho GE (1994) Regulation of sperm motility: emerging evidence for a
major role for protein phosphatases. JAndrol 15:505-509.
Stellenbosch University http://scholar.sun.ac.za

152Terriou P, Hans E, Giorgetti C, Spach JL, Salzmann J, Urrutia V, Roulier R (2000)
Pentoxifylline initiates motility in spontaneously immotile epididymal and
testicular spermatozoa and allows normal fertilization, pregnancy, and birth after
intracytoplasmic sperm injection. JAssist Reprod Genet. Apr;17(4):194-199.
Tesarik J, Thebault A, Testart J (1992) Effect of pentoxyfylline on sperm movement
characteristics in normozoospermic and asthenozoospermic specimens. Hum
Reprod 7:1257-1263.
Vanhaesebroeck B, Leevers SJ, Panayyotou G, Waterfield MD (1997)
Phosphoinositide 3-kinase: a conserved family of signal transducers. Trends
Biochem Sci 22:267-272.
World Health Organisation (1999) Collection and examination of human semen. In:
WHO Laboratory Manual for the Examination of Human Semen and Semen-
Cervical Mucus Interaction. 4th ed. Cambridge University Press, London, pp4-
33.
Wymann MP, Pirola L (1998) Structure and function of phosphoinositide 3-kinases.
Biochim Biophys 1436:127-150.
Stellenbosch University http://scholar.sun.ac.za

The erection is the last gasp of modern
manhood. If men can't continue to
produce erections, they're going to evolve
themselves right out of the human
.species.
- Time Magazine, 5 May 1998-
Stellenbosch University http://scholar.sun.ac.za

153CHAPTER 7
The effect of acute in vivo sildenafil citrate (VIAGRA™) treatment on
semen parameters and sperm function
Monduzzi Editore 2002, In Press
SS du Plessis 1,*, PS de Jongh 1 and DR Franken2
1Department of Medical Physiology and Biochemistry, University of Stellenbosch,
P.O.Box 19063, Tygerberg, 7505, South Africa.
2Department of Obstetries and Gynaecology, University of Stellenbosch, Tygerberg
Hospital, Tygerberg, 7505, South Africa.
Running head: Effect of sildenafil citrate on sperm parameters and function
Keywords: sildenafil citrate, spermatozoa, acrosome, zona pellucida, motility
*Corresponding author. SS du Plessis, Department of Medical Physiology and
Biochemistry, University of Stellenbosch, P.O.Box 19063, Tygerberg, 7505, South
Africa; Tel.: +27-(0) 21-938 9388; fax: +27-(0) 21-938 9476; e-mail:
Stellenbosch University http://scholar.sun.ac.za

154
Summary
Viagra TM is a powerful treatment for male erectile dysfunction of various aetiologies.
Sildenafil citrate, the active ingredient of Viagra TM, is a specific and potent inhibitor of
phosphodiesterase type 5 and thus enhances the activity of the nitric oxide-cGMP
pathway that promotes penile erection due to cGMP accumulation. In this study we
investigated whether acute in vivo sildenafil citrate (50mg orally) administration
modifies seminal parameters, induction of the acrosome reaction, sperm-oocyte
binding and sperm motility. No changes in the macro- and microscopical seminal
parameters were caused by sildenafil citrate when compared to placebo. Sildenafil
citrate also did not initiate or potentiate the acrosome reaction. Sperm-oocyte
binding, smooth path velocity, straight line velocity and the percentage rapid cells all
increased after sildenafil citrate treatment. These results suggest a clinical usefulness
for sildenafil citrate in the enhancing of fertilizing ability of inherently poor quality
sperm or for treatment during assisted reproductive techniques.
Introduction
Erectile dysfunction (ED) is a widespread condition that has a negative impact on
quality of life, affecting both older and younger men in an estimated 10% of the adult
population (Feldman et ai., 1994). Until recently no effective oral therapy existed and
available treatments were highly cumbersome or invasive (Purvis et aI., 2002).
Sildenafil citrate (ViagraTM, Pfizer), a cyclic nucleotide phosphodiesterase (PDE)-
inhibitor, was the first oral agent to be successfully introduced for the management of
ED in which there is no established organic cause (Boolell et ai., 1996b). When
administered before sexual activity, it produces reliable efficacy, good tolerability and
rapid absorption that yields prompt onset of action (Boolell et al., 1996a).
Stellenbosch University http://scholar.sun.ac.za

155Cyclic nucleotide phosphodiesterases regulate intracellular levels of cyclic 3',5'
adenosine monophosphate (cAMP) and cyclic 3',5' guanosine monophosphate
(cGMP) by hydrolysing them to the corresponding 5' monophosphates (Conti et aI.,
1995). Cyclic nucleotides serve as second messengers for a wide variety of
extracellular signals such as nitric oxide, neurotransmitters, hormones, light and
odourants. As of yet nine different POE iso-enzymes (POE1 to POE9) have been
described and found to be present at various concentrations in human tissues
(Soderling et aI., 1998a; Fabbri et aI., 1999).
Nitric oxide synthase, a cGMP inducer (Lewis et aI., 1996), and two distinct POE
isoforms (POE1 and POE4) are present in human sperm cells (Fisch et aI., 1998). It
was also shown in previous studies that mRNA coding for cAMP-specific POE
(POE4A) isoforms are present in mature rat and mouse germ cells (Naro et al., 1996)
and that the expression of these isoforms is maximal in round spermatids and is
maintained in mature spermatozoa (Soderling et aI., 1998b).
Phosphodiesterase type-5 (POE5) is the predominant POE iso-enzyme for the
degradation of cGMP in the corpus cavernosum. Sildenafil citrate (Viagra™) is a
specific and potent inhibitor of this cGMP-specific POE5 enzyme and thus enhances
the activity of the nitric oxide-cGMP pathway (Figure 1) that promotes penile erection
due to cGMP accumulation (Ballard et al., 1998; Glossman et al., 1999). It also has
minor inhibitory effects on POE6 and POE1 activities (ICsD= 0.034 and 0.281Jmol/l
respectively) (Morales et al., 1998). Sildenafil citrate mimics cGMP and interacts with
the catalytic site of POE. Ouring normal erections sexual stimulation lead to an
increase in nitric oxide production which in turn stimulate cGMP production. These
increased cGMP levels causes trabecular smooth muscle relaxation, cavernosal
Stellenbosch University http://scholar.sun.ac.za

156artery dilatation, increased cavernosal pressure and penile erection. Sildenafil citrate
potently enhances the relaxant effect of nitric oxide on the human corpus
cavernosum by increasing the value of cGMP in this tissue. However sexual
stimulation still remain mandatory in order to produce its pharmacological effect,
since it has no direct relaxant effect on the corpus cavernosum (Fabbri et ai., 1997;
Ballard et ai., 1998; Goldstein et al., 1998). Viagra TM is rapidly absorbed and acts
within 30 minutes and reach maximum plasma concentrations within approximately 1
hour. The absolute bioavailability is 41% due to first-pass metabolism and is rapidly
cleared from the body with a plasma elimination half-life of 3-4 hours (Nichols et ai.,
2002).
It is likely that men with ED will use sildenafil citrate as an aid to procreation,
especially by younger patients who have ED e.g. secondary to spinal cord injury. It is
also known that a degree of ED may be present in the male infertile partner
especially when assisted reproductive techniques are necessary. These men have
difficulties in producing spermatozoa on demand at the time of egg fertilization. Tur-
Kaspa et al. (1999) showed that these forms of temporary erectile dysfunction can be
successfully treated with sildenafil citrate. In this context however, the local affects of
sildenafil citrate on sperm function have not been researched extensively.
A large number of drugs can be transported into the seminal fluid, where they have
direct effects on function, physiology, metabolism or genetic composition of
spermatozoa (Pichini et ai., 1994). Several drugs have been shown to affect sperm
motility in particular; these include compounds with PDE-inhibitory activity that have
the potential to increase motility (Schoenfeld et a/., 1975; Turner et ai., 1978). One
such drug, pentoxifylline, which is a cAMP PDE inhibitor, has been shown to
Stellenbosch University http://scholar.sun.ac.za

157stimulate human sperm motility both in vitro and in vivo (Shen et aI., 1991). It has
also been demonstrated that human sperm cells contain as yet uncharacterised POE
isoforms which are different from POE1 and POE4, and that the in vitro inhibition of
sperm POE1 and POE4 iso-enzymes by specific inhibitors stimulates acrosome
reaction (AR) and sperm motility (Fisch et al., 1998). It is also known that after acute
oral administration of 100mg-sildenafil citrate, the drug reaches a concentration of
0.1-0.3 ,",mol/I in the ejaculate (Pfizer, Viagra™ data sheet). This concentration is
consistent with possible inhibitory actions of sildenafil citrate on sperm POE isoforms
(Fabbri et aI., 1999).
Sildenafil citrate is rapidly becoming the treatment of choice for EO of various
aetiologies even though many of its potential effects are still being investigated
(Edwards and Muirhead, 2002). The drug is relatively lipophilic (log07.4=2.7) and
would therefore be expected to distribute into the seminal fluid (Purvis et aI., 2002)
where it subsequently might have important local effects/actions which still remain
unknown. We therefore conducted a study to investigate and evaluate the effects of
an acute single oral dose of sildenafil citrate (50mg) on a number of seminal, sperm
functional and morphology parameters in young healthy male volunteers.
Stellenbosch University http://scholar.sun.ac.za
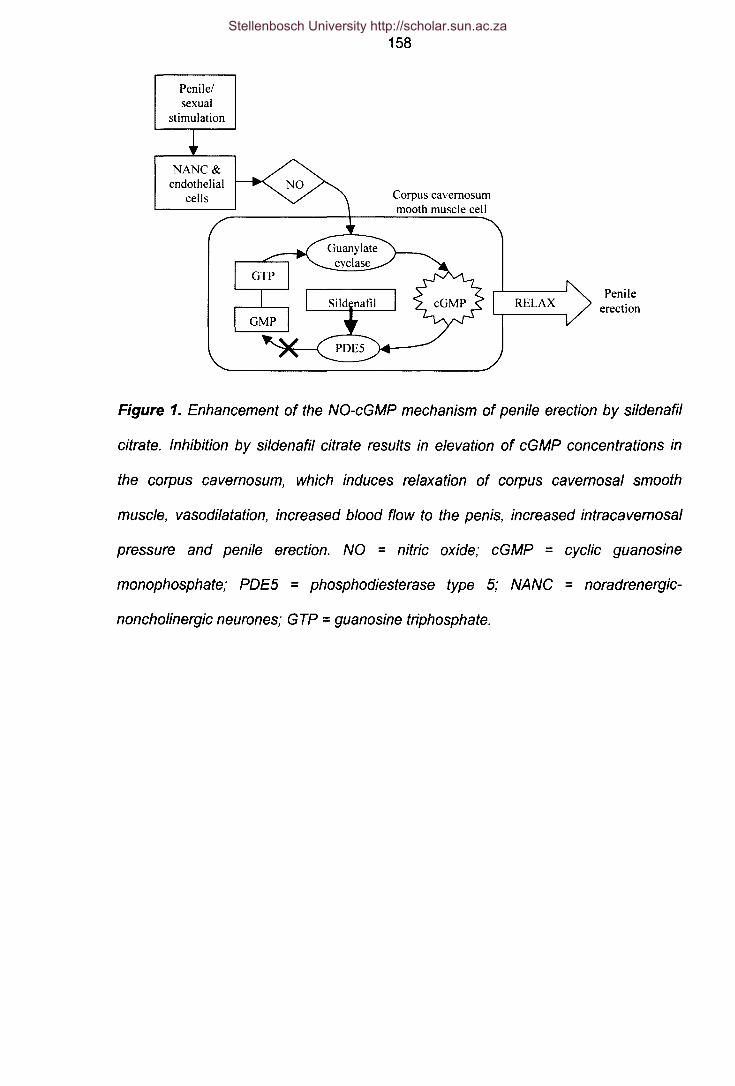
Penile/sexual
stimulation
NANC&endothelial
cells
158
Corpus cavernosummooth muscle cell
PenileerectionRELAX
Figure 1. Enhancement of the NO-cGMP mechanism of penile erection by si/denafil
citrate. Inhibition by sildenafil citrate results in elevation of cGMP concentrations in
the corpus cavernosum, which induces relaxation of corpus cavernosal smooth
muscle, vasodilatation, increased blood flow to the penis, increased intracavernosal
pressure and penile erection. NO = nitric oxide; cGMP = cyclic guanosine
monophosphate; POE5 = phosphodiesterase type 5; NANG = noradrenergic-
noncholinergic neurones; GTP = guanosine triphosphate.
Stellenbosch University http://scholar.sun.ac.za

159
Materials and Methods
Study design and subjects
The study design consisted of a prospective double blind, placebo-controlled,
crossover, two period clinical investigation. The study blind was broken after
completion of all investigations. All subjects enrolled spontaneously in the study after
giving an informed consent to the study protocol, which was approved by the Ethical
Committee of our institution. The inclusion criteria included normal erectile function,
normal electrocardiogram, no prior or concomitant serious illness or consumption of
medications during the 3-month period prior to the study. In a double-blind fashion,
subjects randomly received one of the two treatments, that is SOmg sildenafil citrate
(Viagra™, Pfizer, Sandton, RSA) or placebo to ingest orally. All subjects were asked
not to ejaculate for a minimum of 3 days before each dose of study drug. They were
not permitted to consume products containing alcohol, caffeine or methylxanthines or
cigarette smoking for a period of three days before each session of the study. The
subjects were also required not to use prescription or over-the-counter drugs during
the course of the study. All study medications were taken under supervision with
SOmlof water. The washout period consisted of a 7-day period in which no
medication was given. After this time, all subjects were crossed over to receive the
alternative treatment.
Written informed consent was obtained from each subject before study entry.
Subjects who met the inclusion criteria underwent a pre-study screening in the 3
weeks before the start of the study that consisted of a physical examination,
Stellenbosch University http://scholar.sun.ac.za

160measurement of supine blood pressure and pulse rate, a 12-lead electrocardiogram
and a urine test.
An in vitro study was also conducted in parallel in order to investigate the effects of
increased levels of exogenous cGMP on sperm motility, AR and sperm-oocyte
binding. 8-bromo-cGMP (8-Br-cGMP), a non-hydrolyzable cGMP analogue, was used
to increase intracellular cGMP levels in vitro, as it is lipid soluble and therefore able to
enter the cell quite easily (in contrast to cGMP that cannot penetrate the cell
membrane). The drug dissolved easily in water and was used at a final concentration
of 20IJM(60minutes).
Semen processing I Preparation of sperm samples
Semen samples were collected 1 hour after ingestion of the test drug by means of
masturbation according to the World Health Organisation guidelines (WHO, 1999).
Motile sperm fractions were collected from samples using a slightly modified double-
wash swim-up technique. Retrieved sperm samples were resuspended in synthetic
human tubal fluid culture medium (HTF) (Quin et ai., 1985) supplemented with 3%
bovine serum albumin (BSA; Seravac, Cape Town, South Africa) to a sperm
concentration of 1Ox106 cells/ml. Before the onset of AR and sperm-oocyte binding
studies, sperm samples were allowed to capacitate at 37°C in 5% CO2 for 3 hours in
HTF-BSA. Excess spermatozoa that were not used for motility or AR studies were
cryopreserved (using SpermFreeze; FertiPro N.v, Beernem, Belgium) in 0.5 ml
straws using a slow-freeze method (Mahedevan and Trounson, 1993) in order to
perform matched hemizona assays at a later stage.
Stellenbosch University http://scholar.sun.ac.za

161Macro and microscopical seminal parameters
Ejaculate volume was measured volumetrically and pH was determined with Multistix
Reagent Strips (Bayer Corporation, Tarrytown, NY, USA). Liquefication, viscosity and
appearance of each sample were judged visually. Sperm concentration, sperm count
and sperm head size (area) was acquired by means of computer assisted sperm
analysis (CASA, see later). An assessment of morphology was made after staining
with eosin Y and Harris haemotoxylin and spermatozoa was evaluated according to
World Health Organisation guidelines (WHO, 1999) together with strict sperm
morphology criteria (Kruger et a/., 1986).
Motility studies
Sperm kinematics was assessed after a 3D-minute liquefication period at room
temperature by means of Computer assisted semen analysis (CASA). The analysis
was performed on an Integrated Visual Optical System for sperm analysis (IVOS,
Hamilton Thorne Research, Beverly, MA) with standard set-up parameters. The raw
semen was diluted 1:1 in HTF-BSA and placed in a disposable fixed-depth slide.
(20.0Ilm, Makler) at an image capture rate of a 30 frames at 60Hz. A minimum of 200
cells and three fields was analysed for each aliquot. All analyses were performed at
3rc. The following measurements were performed:
Counts:
Total, Motile, Progressive
% Motile, % Progressively Motile
Rapid, Medium, Slow and Static Cells
Concentrations
Total, Motile, Progressive (millions/ml)
Rapid, Medium, Slow and static Cells (millions/ml)
Stellenbosch University http://scholar.sun.ac.za

162Mean Values
VAP: Smoothed Path Velocity (microns/sec)
VCL: Track Velocity (microns/sec)
VSL: Straight Line Velocity (microns/sec)
AlH: Amplitude of lateral Head Displacement (microns)
BCF: Beat Cross Frequency (hertz)
LIN: Linearity (ratio of VSLlVCl)
STR: Straightness (ratio of VSLlVAP)
Area: Head size (square microns)
Preparation of solubilized ZP
Oocytes were retrieved from post mortem derived ovarian material. (The project fully
complies with the legal and ethical guidelines of the South African Medical Research
Council.) Oocytes were stored in a dimethylsulfoxide/sucrose solution at -196°C in
liquid nitrogen (Hammit et al., 1991). Twenty four hours prior to each test, oocytes
were removed from storage and thawed at 37°C. Retrieved oocytes were placed in
0.25M sucrose and 3% BSA in HTF. On the day of the experiment, 20 oocytes were
placed in a drop on a petri dish, after which the HTF was removed under microscopic
vision (Olympus SZ40; Wirsam Scientific, Cape Town, South Africa), leaving only the
20 oocytes on the petri dish. A total volume of 5JlI of 10mM HCl was then added to
the oocytes on the petri dish; solubilization of the ZP was microscopically observed
and controlled. Following solubilization, 5JlI of 10mM NaOH was added to the
solubilized ZP, to render a final zona volume of 10JlIcontaining 2 ZP/Jll. The final ZP
concentration, after the addition of spermatozoa, was 0.67 ZP/Jll.
Stellenbosch University http://scholar.sun.ac.za

163Acrosome reaction studies
Spermatozoa from both the Viagra or placebo groups were challenged with (i) 10llM
calcium ionophore A23187 (C-7522, Sigma, Cape Town, South Africa) for 30
minutes, (ii) 1lJg/ml progesterone for 30 minutes, (iii) 0.67ZP/1l1for 30 minutes or (iv)
left to spontaneously elicit the AR. The acrosomal status of these spermatozoa were
subsequently determined according to procedures published elsewhere (Cross et al.,
1988; Morales et al., 1989, Franken et al., 2000).
Spermatozoa from the different experiments were fixed and air-dried, after which the
acrosomal status was determined using fluorescein-labeled Pisum Sativum agglutinin
(FITC-PSA; 125Ilg/ml; L-0770; Sigma, Cape Town, South Africa). Two individuals
scored a minimum of 200 spermatozoa blindly for each different sample. The
following staining patterns were evaluated as acrosome reacted spermatozoa: (i)
patchy staining on acrosomal region, (ii) distinct staining in the equatorial region
occurring as an equatorial bar and (iii) no staining over the entire sperm surface.
Hemizona Assays/Zona Pellucida Binding
Straws containing the cryopreserved spermatozoa were thawed rapidly at 3rC for
10 minutes. Cryoprotectant was removed with the dropwise addition of P1 medium
followed by centrifugation at 600 9 for 10 minutes and resuspension in 0.5ml fresh
medium (Mahedevan & Trounson, 1993). Both test (sildenafil citrate) and control
(placebo) sperm droplets (15111;10x106sperm/ml) were placed on a petri dish to
which hemizonae were added in a match-controlled fashion. Hemizona assays (HZA)
were performed in duplicate and co-incubation was for four hours. Following the co-
incubation period, hemizonae were removed and washed (5x) to strip the loosely
attached spermatozoa from the hemizonae. Hemizonae were then evaluated by two
Stellenbosch University http://scholar.sun.ac.za

164individuals using the blind scoring method, while the number of spermatozoa tightly
bound to the ZP was recorded for each test and matching control. Sperm-zona
binding was also expressed as a percentage or a Hemizona Index (HZI) where the
HZI - Test sperm bound to hemizona/ 100 A HZI f 100°/ ould- Control sperm bound to hemizona X. 0 10 W
therefore imply that the compound tested did not interfere with the amount of
spermatozoa that bind to the ZP as compared to that of the control spermatozoa.
Statistical Analysis
All results are expressed as the mean ± the standard error. Data were compared
using Student's t-test for paired data. P-values equal or less than 0.05 were
considered statistically significant. All statistical analysis were performed using
GraphPad PRISM version 2.01.
Results
Demographic characteristics, safety and tolerability
Twenty healthy male volunteers were enrolled in the study. The subjects were all
Caucasian and between 20 and 33 years of age (23.182 ± 0.689). No significant
medical conditions relevant to the study were noted before entry or during the study.
Adverse events considered by the investigator to be related to treatment occurred in
6 of 20 subjects (30%) following administration of sildenafil citrate and in 1 of 20
subjects (5%) following administration of placebo (Table I). The most frequently
reported adverse events were flushing, headache and abnormal vision (blue vision)
and were generally mild to moderate in severity. No subject discontinued treatment
due to adverse effects related to sildenafil citrate.
Stellenbosch University http://scholar.sun.ac.za

165Table I: Incidence of adverse events following treatment with placebo or sildenafil
citrate (n=20).
Placebo Sildenafil 50mg
Treatment related adverse events 1/20 6/20
Flushing 0 3
Headache 0 2
Abnormal vision 0 2
Spontaneous erections 1 1
Macro- and microscopical seminal parameters
Table II shows the macroscopical seminal parameters after acute in vivo sildenafil
citrate or placebo administration. Liquification, appearance and viscosity of the
semen samples from both groups were normal as according to the WHO criteria.
Mean values of semen volume (2.533±0.320ml vs. 2.373±0.313ml) and pH
(8.182±0.076 vs. 8.318±0.076) did not show significant variations between the two
treatment groups.
Table II: Initial macroscopic appearance and evaluation of semen after either
placebo or sildenafil citrate administration (n=20).
Semen parameters Placebo Sildenafil 50mg
Liquefication <60 minutes <60 minutes
Appearance Normal Normal
Viscosity Normal Normal
Ejaculate Volume (ml) 2.533 ± 0.320 2.373 ± 0.313
pH 8.182 ± 0.076 8.318 ± 0.076
p
0.432
0.096
Stellenbosch University http://scholar.sun.ac.za

166
Similarly sildenafil citrate was equivalent to placebo in its effects on all microscopical
secondary semen analysis parameters, including sperm count, sperm concentration
and morphology (Table III). All these variables were also within the normal ranges as
predefined by the study centre. The area of the sperm head size (square microns), as
determined by CASA, was not affected by sildenafil citrate.
Table III: Effects of acute in vivo administration of sildenafil citrate on microscopical
secondary semen analysis parameters (n=20).
Semen parameters Normal Placebo Sildenafil P
values 50mg
Sperm count (x106) 100-500 220.8 ± 45.892 232.2 ± 54.453 0.449
Sperm concentration (x106/ml) 50-150 73.72 ± 13.32 77.52 ± 13.29 0.627
Morphology (% normal forms) >15 15.059 ± 0.961 14.765 ± 0.893 0.281
Area (urn sq) 3.400 ± 0.228 3.350 ± 0.394 0.872
Acrosome reaction
From Table IV it is evident that the addition of exogenous cGMP did not affect the AR
between treatment groups (control vs. 8-Br-cGMP) when left to spontaneously
undergo the AR or when stimulated with A23187 or progesterone. The AR is thus
independent of treatment with 8-Br-cGMP at a concentration of 20!JM.
The results of the various AR studies from the clinical trial are displayed in Figure 2.
When left to spontaneously undergo the AR, the placebo group showed a 13.94 ±
Stellenbosch University http://scholar.sun.ac.za

1672.542% AR while the sildenafil citrate group showed no statistical significant
difference (14.06 ± 2.128%; P = 0.753) in AR. It can be seen that after calcium
ionophore (A23187) stimulation the percentage AR in both groups was statistically
significantly elevated (Placebo: 23.37 ± 2.831%, P=0.0002; Sildenafil: 26.37 ±
3.598%, P=0.0001). Progesterone stimulation had a similar effect and also increased
acrosome reactions in both groups with significant margins to above that of the
spontaneous levels (Placebo: 21.21 ± 3.306%, P=0.004; Sildenafil: 24.90 ± 3.788%,
P=0.007). Exposure to solubilized zona pellucida also increased the acrosome
reactions significantly to above that of spontaneous levels in both groups (Placebo:
25.64 ± 5.797%, P=0.026; Sildenafil: 26.17 ± 3.898%, P = 0.014). The acrosome
reactions between treatment groups (placebo vs. sildenafil) did not differ significantly
after A23187 (P=0.893), progesterone (P=0.728) and zona pellucida (P=0.957)
stimulation and were thus independent of treatment.
Table IV: The effect of in vitro intracellular cGMP elevation by the addition of 8-8r-
cGMP (20fJM) on eliciting of the acrosome reaction (n=6).
Stimuli Control 8-Br-cGMP P
Spontaneous 16.500±3.085 17.167±3.911 0.444
A23187 23.667±3.273 23.000±6.611 0.287
Progesterone 21.333±3.896 20.857±2.852 0.225
Stellenbosch University http://scholar.sun.ac.za

168
35c.2 3013i 25~ 20
i 15ob 10ct:-;;e. 5
1"miii;,l" Placebo_ Sildenafil
n
s A p ZP
Figure 2: The effects of double-blind placebo or 50mg-sildenafil citrate administration
on the sperm acrosome reaction (n=20).
S = spontaneous, A = A23187, P = progesterone, ZP = zona pellucida.
Placebo: S vs. A = 0.0002; S vs. P = 0.004; S vs. ZP = 0.026
Sildenafi/: S vs. A = 0.0001; S vs. P = 0.0073; S vs. ZP = 0.0147
Sperm-zona pellucida binding
Incubation of spermatozoa in the presence of 8-Br-cGMP lead to increased sperm-
zona binding and a HZI of 134% was determined (n=6, in duplicate). This percentage
indicates that more spermatozoa bound to their respective matched hemizonaes after
exposure to 8-Br-cGMP (20IJM).
The number of spermatozoa that bound to each matched hemizona (Figure 3) was
not statistically significantly influenced by acute in vivo sildenafil citrate administration
even though a hemizona index of 148.75% was recorded. These results suggest that
a nearly 49% higher binding rate occurred after acute sildenafil citrate ingestion.
Stellenbosch University http://scholar.sun.ac.za

169
" 40c::::so.c 30Elo.Cl)C.fI) 20....olo.
.! 10E::::sZ
Sildenafil
Figure 3: The effect of double-blind placebo or 50mg-sildenafil citrate administration
on the average number of spermatozoa tightlylfirmly bound to each hemizona (n=10;
P=O.281).
Sperm kinematics results
On average more than 800 cells (817.8±71.92), but not less than 200 cells were
evaluated in order to determine the various sperm motility parameters. The mean
values for the sperm motility parameters were all within the normal ranges as
predefined by the study centre (Table V). No significant difference was observed
between placebo and sildenafil citrate for percentage motile, percentage progressive
motile and percentage static cells. Vel, AlH, BeF, STR and LIN also did not differ
between treatments. It was however found that both VAP (58.61±2.258IJm.s-1 vs.
64.11±2.195IJm.s-\ P=0.047) and VSl (47.28±2.250IJm.s-1vs. 51.92±2.249IJm.s-1,
P=0.046) increased statistically significantly after sildenafil citrate treatment.
Furthermore it was found that after sildenafil citrate treatment a shift occurred from
the medium, slow and static cell populations towards the rapid cell population thereby
Stellenbosch University http://scholar.sun.ac.za

170increasing the percentage rapid cells significantly (44.47±4.782% vs. 50.88±5.180,
P=0.009).
The percentage motile (61.0 ±3.488 vs. 82.00±2.415; P=0.036; n=6) percentage
progressive motile (27.33±2.926 vs. 35.50±4.180; P=0.026; n=6) and percentage
rapid (35.00±3.479 vs. 45.67±4.344; P=0.014; n=6) spermatozoa also increased
significantly after incubation of spermatozoa in the presence of 8-Br-cGMP (201JM,
60minutes).
Stellenbosch University http://scholar.sun.ac.za

171
Table V: Effects of 8-Br-cGMP (in vitro) and sildenafil (in vivo) on different sperm motility parameters as measured by CASA.
Parameter Normal Control 8-Br-cGMP P Placebo Sildenafil Pvalues {n=6} {n=6} {n=20} {n=20}
Motile >50 61.00 ± 3.488 82.00 ± 2.451 0.037 67.39 ± 5.892 68.22 ± 5.673 0.850(%)Progressive motility >15 27.33 ± 2.927 35.50 ± 4.180 0.027 30.89 ± 3.809 33.00 ± 3.801 0.530(%)Smoothed Path Velocity 54.96 ± 3.256 58.58 ± 4.898 0.190 58.61 ± 2.528 64.11 ± 2.195 0.047(VAP - um/s)Straight Line Velocity >25 49.28 ± 3.261 51.93 ± 4.869 0.265 47.28 ± 2.250 51.92 ± 2.249 0.046(VSl - um/s)Track Velocity 72.38 ± 4.325 75.98 ± 4.606 0.288 90.18 ± 3.407 94.64 ± 4.009 0.324(VCl - urn/s)Amplitude of lateral Head 3.10 ± 0.202 3.00 ± 0.089 0.542 3.929 ± 0.1350 4.194 ± 0.2190 0.202Displacement (AlH - urn)Beat Cross Frequency 24.12 ± 1.492 24.03 ± 0.674 0.949 24.11 ± 0.8050 24.59 ± 1.136 0.656(BCF - Hz)Straightness 86.50 ± 1.440 85.67 ± 1.291 0.497 77.20 ± 1.377 78.88 ± 1.252 0.447(STR - %)linearity 64.83 ± 2.518 65.67 ± 2.620 0.662 53.61 ± 1.684 54.61 ± 1.589 0.621(LIN - %)Rapid >25 35.00 ± 3.479 45.67 ± 4.344 0.014 44.47 ± 4.782 50.88 ± 5.180 0.009(%)Static <50 32.33 ± 7.298 22.667 ± 6.547 0.272 37.24 ± 6.822 32.35 ± 7.014 0.239(%)
Stellenbosch University http://scholar.sun.ac.za

172
Discussion
A total of 20 male volunteers were enrolled in this prospective double blind, placebo-
controlled, crossover, two period clinical investigation to determine the
pharmacodynamic effects of sildenafil citrate on sperm function and ejaculate quality.
Sildenafil citrate was well tolerated by the volunteers, producing no serious adverse
events or clinically important changes in vital signs. The most frequently reported
adverse events were generally consistent with those reported in other studies (Purvis
et a/., 2002; Boolell et a/., 1996; Goldstein et a/., 1998; Morales et a/., 1998).
In this study a comparison of acute sildenafil citrate treatment vs. placebo showed no
effect on a wide variety of macroscopical seminal measures including liquefication,
appearance, viscosity, ejaculate volume or pH. Sperm count, sperm concentration,
morphology and area of sperm head size was also not influenced. These results are
in line with those reported in similar studies (Aversa et a/., 2000).
It was shown that exogenous cGMP did not initiate or potentiate the AR of
capacitated spermatozoa, yet it is well known from the literature that cGMP serve as
a signal transducer mediating the AR. When this signalling pathway is activated by
using 8-Br-cGMP, effects are only documented at concentrations of higher than
0.5mM with peak activity at 1mM (Rotem et a/., 1998; Revelli et a/., 2001). This is
most likely due to PKA stimulation because cGMP can interact with the regulatory
subunit of PKA when used at concentrations higher than 100IJM(Lincoln & Cornwell,
1993). In our experiments we used a concentration of 201JM,which did not result in
the initiation of the AR. Capacitated spermatozoa also underwent an AR when
Stellenbosch University http://scholar.sun.ac.za

173
challenged with calcium ionophore, progesterone and zona pellucida plus in
combination with the POE inhibitor sildenafil citrate, but not with sildenafil citrate
alone. These data suggest that sildenafil citrate (50mg orally) by itself cannot initiate
the AR nor can it potentiate the AR of capacitated spermatozoa, as it most probably
cannot elevate intracellular cGMP levels high enough through its inhibition of POE5.
Furthermore, Lefievre et al. (2000) similarly showed that POE inhibitors by
themselves had no effect on the AR.
Fisch et al. (1998) showed by means of immunocytochemical staining that different
types of POE are located at different sites in the spermatozoon e.g. POE4 localises
mainly to the mid-piece, while POE1 is found more prominently in the sperm head.
These observations support the idea that differential POE localisation within the
spermatozoon can allow for selective modulation of sperm function through the
regulation of distinct pools of cAMP (Fisch et al., 1998).
Sildenafil citrate is a controversial member of a drug class (POE inhibitors) previously
shown to either have some effects on sperm motility (Schoenfeld et al., 1975; Turner
et al., 1978; Lefiévre et al., 2000) or no effects on motility at all (Purvis et al., 2000;
Aversa et al., 2000).
In this study a comparison of sildenafil citrate vs. placebo on a wide variety of
kinematics parameters showed no effect on percentage sperm motility, percentage
progressive motility, track velocity, amplitude of lateral head displacement, beat cross
frequency, straightness, linearity or percentage static cells. Borderline statistical
significant changes were however observed in smoothed path velocity and straight-
Stellenbosch University http://scholar.sun.ac.za

174
line velocity after sildenafil citrate administration. The most significant change of note
was observed in the increase in percentage rapid cells after sildenafil citrate
administration. Elevation of intracellular cGMP levels by 8-Br-cGMP also increased
percentage motile, percentage progressively motile and percentage rapid cells
significantly. Both Cuadra et al. (2000) and lefievre et al. (2000) showed that the
inhibition of POES through the addition of sildenafil citrate also enhanced certain
motility parameters. Cuadra et al. (2000) found an increase in sperm progressive
motility and hyperactivation, while lefievre et al. (2000) showed increases in VCl,
AlH and hyperactivation.
The hemizona indexes of 148 % after sildenafil citrate and 134% after 8-Br-cGMP
administration refer to increases in sperm-oocyte binding. These increases in binding
can possibly be explained by the fact that, combined with various increases in
different motion characteristics, more spermatozoa are rapidly motile thereby
increasing the chances of them colliding with the oocyte. This theory fits in with
results previously obtained in our laboratory, that the frequency of sperm/zona
pellucida collision rates has an influence on zona binding (OzgOr& Franken, 1996).
This increase in hemizona index is further proof that sildenafil citrate, and therefore
POE inhibition, does not elicit the AR as only AR intact cells can bind to the oocyte.
lefiévre et al. (2000) discovered that the inhibition of sperm POE by sildenafil citrate
was associated with a significant increase in cAMP and that sperm POE activity
measured with cGMP as substrate was threefold lower than when measured with
cAMP. There was also a dose-dependent increase in cAMP levels in spermatozoa
incubated with sildenafil citrate (lefiévre et ai., 2000). It was demonstrated that
Stellenbosch University http://scholar.sun.ac.za

175
spermatozoa contain POE that can hydrolyse both cAMP and cGMP (Lefiévre et aI.,
2000). However, whereas cAMP has often been linked to sperm function, the role of
cGMP in sperm functions remains still unclear. The intracellular mechanism
responsible for the sildenafil-induced cAMP increase in human spermatozoa is
unknown, but it can be hypothesised that sildenafil citrate could act on POEs other
than type 5 (Lefiévre et al., 2000). Sildenafil citrate induced similar changes in sperm
motility parameters as nonselective POE inhibitors such as pentoxifylline (Calogero et
aI., 1998; Nassar et aI., 1999) caffeine (Cheng & Boettcher, 1981; Leclerc et aI.,
1996) and IBMX (Leclerc et aI., 1996). These observations also suggest that
sildenafil citrate probably acts on types 1 and 4 POE because cAMP has been
implicated in the regulation of sperm motility (Calogero et al., 1998) through
activation of PKA (Yanagimachi, 1994). It was also shown by Lefiévre et al. (2000)
that sildenafil citrate increased capacitation and the associated tyrosine
phosphorylation of p105/81, 2 fibrous sheath proteins. These 2 latter processes are
recognised to be cAMP-dependent (Leclerc et aI., 1996).
In conclusion, inhibition of POE by sildenafil citrate produces an increase in sperm
kinematics parameters as well as sperm-oocyte binding, but no macroscopical or
microscopical changes occurred. It also suggests that POE is not strongly involved in
the events leading to the human sperm AR.
Although the incidence of ED increases with age, younger men are also affected
(Feldman et aI., 1994). As such, it is very likely that a significant number of patients
will use sildenafil citrate as an aid to procreation. When studying erectile dysfunction
drugs the focus was always on obtaining sexual satisfaction and never about sperm
Stellenbosch University http://scholar.sun.ac.za

176
quality. The results of this study clearly indicate that single oral doses of sildenafil
citrate (50mg) can be safely administered without concern on the part of the
physician or the patient regarding adverse effects on sperm or ejaculate quality.
Furthermore, the ability of sildenafil citrate to enhance sperm kinematics results and
sperm-oocyte binding without affecting the AR may be clinically useful. These
findings could have important implications in the use of sildenafil citrate in assisted
reproductive technology to enhance fertilizing potential of inherently poor quality
sperm with less invasive treatments such as intrauterine inseminations, thereby
avoiding more involved procedures such as IVF or ICSI.
Stellenbosch University http://scholar.sun.ac.za

177
References
Aversa A, Mazzilli F, Rossi T, Delfino M, Isidori AM, Fabbri A (2000) Effects of
sildenafil administration on seminal parameters and post-ejaculatory refractory
time in normal males. Hum Reprod 15(1): 131-134.
Ballard SA, Gingell CJ, Tang K, Turner LA, Price ME, Naylor AM (1998). Effects of
sildenafil on the relaxation of human corpus cavernosum tissue in vitro and on
the activities of cyclic nucleotide phosphodiesterase isozymes. J. Urol. 159:
2164-2171.
Boolell M, Allen MJ, Ballard SA et al. (1996a) Sildenafil: an orally active type 5 cyclic
GMP-specific phosphodiesterase inhibitor for the treatment of penile erectile
dysfunction. Int J Impot Res 8: 47-52.
Boolell M, Gepi-Attee S, Gingell JC and Allen MJ (1996b). Sildenafil, a novel effective
oral therapy for male erectile dysfunction. B. J. Urol. 78:257-261.
Calogero AE, Fishel S, Hall J, Ferrara E, Vicari E, Green S, Hunter A, Burrello N,
Thornton S, D'Agata R. (1998) Correlation between intracellular cAMP content,
kinematic parameters and hyperactivation of human spermatozoa after
incubation with pentoxifylline. Hum Reprod 13:911-915.
Cheng CV, Boettcher B (1981) Effect of cAMP, Mn2+ and phosphodiesterase
inhibitors on human sperm motility. Arch Androl 7:313-317.
Conti M, Nemoz G, Sette C and Vicini E (1995) Recent progress in understanding the
hormonal regulation of phosphodiesterases. Endocrine Rev. 16: 370-389.
Cross NL, Morales P, Overstreet JW, Hanson FW (1988). Induction of acrosome
reactions by the human zona pellucida. Bioi Reprod 38:235-244.
Stellenbosch University http://scholar.sun.ac.za

178
Cuadra DL, Chan, PJ, Patton WC, Stewart SC, King A (2000) Type 5
phosphodiesterase regulation of human sperm motility. Am JObstet Gynecol
182(5):1013-1015.
Edwards G, Muirhead GJ (2002) Introduction to supplement. J Clin Pharmacol 53:1S-
3S.
Fabbri A, Aversa A and Isidori A (1997). Erectile dysfunction: an overview. Hum.
Reprod. Update. 3: 455-466.
Fabbri A, Aversa A and Isidori A (1999). Sildenafil and erectile dysfunction. J.
Endocrinol. Invest. 22: 486-491.
Feldman HA, Goldstein I, Hartzichristou DG, Krane RJ, McKinlay JB (1994)
Impotence and its medical and psychosocial correlates: Results of the
Massachusetts Male Aging Study. J Uro1151: 54-61.
Fisch JD, Behr B and Conti M (1998). Enhancement of motility and acrosome
reaction in human spermatozoa: differential activation by type-specific
phosphodiesterase inhibitors. Hum. Reprod. 13: 1248-1254.
Franken DR, Bastiaan HS, Oehninger SC (2000) Physiological induction of the
acrosome reaction in human sperm: Validation of a micro-assay using minimal
volumes of solubilised, homologous zona pellucida. JAssist Reprod Gen
17(3):156-161.
Glossman H, Petrischor G and Bartsch G (1999). Molecular mechanisms of the
effects of sildenafil (VIAGRA) Exp. Gerontol34: 305-318.
Goldstein I, Lue TF, Padma-Nathan H, Rosen RC, Steers WD, Wicker PA (1998)
Oral sildenafil in the treatment of erectile dysfunction. N. Engl. J. Med. 338:
1397-1404.
Stellenbosch University http://scholar.sun.ac.za

179
Hammit DG, Syrop CH, Walker DL, Bennet MR (1991). Conditions of oocyte storage
and use of non inseminated, non-fertilized oocytes for the hemizona assay.
Fertil SteriI60:131-136.
Kruger TF, Menkveld R, Stander FSH, Lombard CJ, Van der Merwe JP, Van Zyl JA,
Smith K (1986). Sperm morphologic features as a prognostic factor in in vitro
fertilization. Fertil Steril 46:1118-1123.
Leclerc P, de Lamirande E, Gagnon C (1996) Cyclic adenosine 3',5' monophosphate-
dependent regulation of protein tyrosine phosphorylation in relation to human
sperm capacitation and motility. Bioi Reprod 55:684-692.
Lefiévre L, De Lamirande E, Gagnon C (2000) The cyclic GMP-specific
phosphodiesterase inhibitor sildenafil, stimulates human sperm motility and
capacitation but not acrosome reaction. JAndrol 21(6): 929-937.
Lewis SE, Donnely ET, Sterling ES, Kennedy MS, Thompson W, Chakravarthy U
(1996). Nitric oxide synthase and nitrite production in human spermatozoa:
evidence that endogenous nitric oxide is beneficial to sperm motility. Mol. Hum.
Reprod. 2:873-878.
Lincoln TH, Cornwell TL (1993) Intracellular cyclic GMP receptor proteins. FASEB J
7:328-338
Mahedevan M, Trounson A (1993). Effect of cryoprotective media and dilution
methods on the preservation of human spermatozoa. Andrologia 15: 355-366.
Morales A, Gingell C, Collins M, Wicker PA, Osterloh IH (1998). Clinical safety of oral
sildenafil citrate (ViagraTM) in the treatment of erectile dysfunction. Int. J. Impot.
Res. 10: 69-74.
Stellenbosch University http://scholar.sun.ac.za

180
Morales P, Cross NL, Overstreet JW, Hanson FW (1989). Acrosome-intact and
acrosome-reacted human sperm can initiate binding to the zona pellucida. Dev
Bioi 133: 385-392.
Naro F, Zang R and Conti M (1996). Developmental regulation of unique adenosine
3'-5'-monophosphate-specific phosphodiesterase variants during rat
spermatogenesis. Endocrinology. 137: 2464-2472.
Nassar A, Mahoney M, Morshedi M, Lin MH, Srisombut C, Oehninger S (1999)
Modulation of sperm tail protein tyrosine phosphorylation by pentoxifylline and
its correlation with hyperactivation and motility. Fertil Steril 71:919-923.
Nichols OJ, Muirhead GJ, Harness JA (2002) Pharmacokinetics of sildenafil citrate
after single oral doses in healthy male subjects: absolute bioavailability, food
effects and dose proportionality. Br J Clin Pharmacol 53: 5S-12S
OzgOr K, Franken DR (1996) The influence of sperm/zona pellucida collision rates on
zona binding. Andrologia 28(5): 261-265.
Pichini S, Zuccaro P, Pacifi R. (1994) Drugs in semen. Clin Pharmacokinet 26:356-
373
Purvis K, Muirhead GJ, Harness JA (2002) The effects of sildenafil citrate on human
sperm function in healthy volunteers. Br J Clin Pharmacol 53:53S-60S.
Quin P, Kerin JF, Warnes GM (1985). Improved pregnancy rate in human in vitro
fertilization with the use of a medium base on the composition of human tubal
fluid. Fertil Steril 44:493-498.
Revelli A, Costamagna C, Moffa F, Aldieri E, Ochetti S, Bosia A, Massobrio M,
Lindblom B, Ghigo 0 (2001) Signalling pathways of nitric oxide-induced
acrosome reaction in human spermatozoa. Bioi Reprod 64(6): 1708-1712.
Stellenbosch University http://scholar.sun.ac.za

181
Rotem R, Zamir N, Keynan N, Barkan D, Breitbart H, Naor Z (1998) Atrial natriuretic
peptide induces acrosomal exocytosis of human spermatozoa. AJP -
Endocrinology and Metabolism: 274(2): E218-E223.
Schoenfeld C, Amelar RD, Dublin L (1975) Stimulation of ejaculated human
spermatozoa by caffeine. Fertil SteriI26:158-161.
Shen MR, Chiang PH, Yang RC, Hong CY Chen SS (1991) Pentoxifylline stimulates
human sperm motility both in vitro and after oral therapy. Br J Clin
PharmacoI31:711-714.
Soderling SH, Bayuga SJ, Beavo JA (1998a). Cloning and characterising of cAMP-
specific cyclic nucleotide phosphodiesterase. Proc. Natl Acad. Sci USA. 95:
8991-8996.
Soderling SH, Bayuga SJ, Beavo JA (1998b).ldentification and characterization of a
novel family of cyclic nucleotide phosphodiesterases. J BioI. Chemo273: 15553-
15558.
Tut-Kaspa I, Segal S and Moffa M, Massobrio M, Meltzer S (1999). Viagra for
temporary erectile dysfunction during treatments with assisted reproductive
technologies: case report. Hum. Reprod. 14: 1783-1784.
Turner EA, De Aparicio NJ, Turner D, Schwartstein L (1978) Effect of two
phosphodiesterase inhibitors, cyclic adenosine 3',5'-monophosphate, and a
beta-blocking agent on human sperm motility. Fertil Steril 29:328-331.
World Health Organisation (1999). WHO Laboratory Manual for the Examination of
Human Semen and Semen-Cervical Mucus Interaction, 4th ed. Cambridge
University Press, Cambridge p.
Yanagimachi R (1994). Mammalian fertilization. In: The physiology of Reproduction.
Knobil E, Neil JD (eds), Vol1 2nd ed. New York: Raven Press, 189-317.
Stellenbosch University http://scholar.sun.ac.za

"Today, if you are not confused, you are
just not thinking clearly."
- U. Peter-
Stellenbosch University http://scholar.sun.ac.za

182
CHAPTER 8
CONCLUSIONS AND RECOMMENDATIONS
8.1 Conclusion
The physiological acrosome reaction occurs upon interaction of the spermatozoon
with the zona pellucida protein ZP3 and is of fundamental importance in the
fertilization of the oocyte by the spermatozoon. This is followed by the liberation of
several acrosomal enzymes and other constituents that facilitate penetration of the
zona and expose molecules on the sperm equatorial segment that allow fusion of the
sperm membrane with the oolemma. The molecular mechanisms and the signal
transduction pathways mediating the processes of motility, capacitation and the
acrosome reaction have been partially defined and appear to involve modifications of
intracellular calcium and other ions, lipid transfer and phospholipid remodelling in the
sperm plasma membrane as well as changes in protein phoshorylation. However
there are still many unanswered questions regarding human sperm interaction with
the oocyte.
8.1.1 Motility
From the results obtained in this study it was evident that:
i) Phosphatidylinositol 3-kinase negatively regulates sperm motility.
ii) Inhibition of phosphodiesterase type 5 by sildenafil citrate, and therefore a possible
accumulation of cGMP in the spermatozoon, lead to an increase in certain sperm
kinematics parameters.
Stellenbosch University http://scholar.sun.ac.za

183
The process of development and maintaining of motility in mammalian spermatozoa
is rather complex and involves the integration and crosstalk of several signalling
pathways, including adenylate cyclase/cAMP/PKA, calcium and
phosphorylation/dephosphorylation of various substrate proteins (Tash and Bracho,
1994).
The molecular mechanisms underlying the stimulatory effects of the PI3K inhibitor,
LY294002, on sperm motility are still poorly understood. At this stage we can only
speculate that in spermatozoa, PI3K might be involved in the phosphorylating or
dephosphorylating of proteins involved in motility. Another possibility is that PI3K is
involved in the generation of reactive oxygen species (ROS), which has a negative
impact on motility (Luconi et ai., 2001).
The increase in motility due to in vivo Viagra TM treatment is a novel finding and only
speculations can be made regarding the intracellular mechanisms underlying the
finding at this stage. Sildenafil citrate induced similar changes in sperm motility
parameters as non-selective POE inhibitors such as pentoxifylline (Calogero et al.,
1998; Nassar et al., 1999), caffeine (Cheng & Boettcher, 1981; Leclerc et al., 1996)
and IBMX (Leclerc et ai., 1996). The intracellular mechanism responsible for the in
vitro sildenafil citrate-induced cAMP increase in human spermatozoa is also
unknown, but it can be hypothesised that sildenafil citrate could act on POEs other
than type 5 (Lefiévre et al., 2000). These observations also suggest that sildenafil
citrate probably acts on types 1 and 4 POEs because cAMP has been implicated in
the regulation of sperm motility (Calogero et al., 1998) through activation of PKA
(Yanagimachi, 1994).
Stellenbosch University http://scholar.sun.ac.za

184
Both of these motility-enhancing effects need to be investigated further in order to
clarify their exact mechanisms of action.
8.1.2 Acrosome reaction
From the results obtained in this study it was evident that:
i) Extracellular signal regulated kinases are directly or indirectly involved in the signal
transduction pathway through which the zona pellucida-induced acrosome reaction
(physiologically relevant exocytotic event) is mediated and not the spontaneous or
calcium ionophore induced acrosome reaction.
ii) Phosphatidylinositol 3-kinase increase sperm-zona binding without influencing the
acrosome reaction.
iii) Inhibition of phosphodiesterase type 5 by sildenafil citrate and therefore possible
accumulation of cGMP produce an increase in sperm-oocyte binding. It also suggests
that phosphodiesterase type 5 is not strongly involved in the events leading to the
human sperm acrosome reaction.
By combining these results with current facts from the literature, a hypothesised
schematised representation of the intracellular signal transduction pathways involved
in the zona pe/lucida-induced acrosome reaction will be shown in Figure 1 and
discussed subsequently.
The ZP3 glycoprotein binds to at least two receptors in the plasma membrane. One
receptor is a Gj-coupled receptor that activates phospholipase C (PLC) r11. The other
receptor is a tyrosine kinase receptor (TKR) coupled to PLCy• Binding to the Gj-
Stellenbosch University http://scholar.sun.ac.za

185
coupled receptor would activate adenylate cyclase (AC) leading to the elevation of
cAMP and PKA activity. PKA activates a voltage-dependent Ca2+channel in the outer
acrosomal membrane, which releases Ca2+from the interior of the acrosome to the
cytosol. This relative small rise in [Ca2+]jcould result in the activation of PLCy.The
products of phosphatidyl-inositol biphosphate (PIP2) hydrolysis by PLCB1and PLCy,
diacylglycerol (DAG) and inositoltrisphosphate (IP3) lead to PKC translocation to the
plasma membrane and its subsequent activation. This increase in [Ca2+]j can be
mimicked by the addition of a calcium ionophore (e.g. A23187), which will also result
in the activation of PLCy, and PKC activity. PKC opens a voltage-dependent Ca2+
channel in the plasma membrane, increasing the [Ca2+]jeven more. PKC activation
also results in the activation of ERK, through its ability to phosphorylate and activate
an upstream mediator of the ERK cascade, Raf. Raf will activate and phosphorylate
the ERK kinase, MEK, which in turn activates ERK. The Gj-coupled receptor and
TKR can also activate a Na+/H+exchanger, leading to alkalisation (pH increase) of
the cytosol. The increase in [Ca2+]j and pH will lead to membrane fusion and
acrosomal exocytosis.
From the literature it is evident that various crosstalk mechanisms are involved in
phosphatidylinositol 3-kinase (PI3K) and RAS/RAF/ERK signalling. It is known that
RAS can both activate RAF/ERK as well as PI3K (Sears & Nevins, 2002; Krasilnikov,
2000). PI3K on the other hand can also activate the RAS/RAF/ERK pathway through
mitogen signalling (Krasilnikov, 2000). In our experiments we stimulated the AR with
solubilized ZP (ZP3), thereby eliciting the AR through both a Gj-protein receptor and
ZRK binding which directly activates RAS/RAF/ERK and PI3K through RAS.
Inhibition of PI3K by LY294002 thus only inhibited the PI3K pathway and not the
Stellenbosch University http://scholar.sun.ac.za

186
direct RAS/RAF/ERK pathway activated independent of PI3K signalling or activation
through PKC. The activation of RAS by ZP binding to ZRK can also stimulate PI3K,
thereby enhancing a possible positive feedback on ERK activation (e.g. via PKC) as
well as other mediators in the process of AR signalling.
It is well known from the literature that cGMP serve as a signal transducer mediating
the AR. When this signalling pathway is activated by using 8-Br-cGMP, effects are
only documented at concentrations of higher than 0.5mM with peak activity at 1mM
(Rotem et al., 1998; Revelli et al., 2001). This is most likely due to PKA stimulation
because cGMP can interact with the regulatory subunit of PKA when used at
concentrations higher than 100IJM(Lincoln and Cornwell, 1993).
Compared with signalling pathways in many types of somatic cells, the signal
transduction pathways of the human acrosome reaction (pellucida glycoprotein) are
still very poorly understood (Fisher et aL, 1998). One of the chief reasons for this is
that the sperm receptor(s) that bind to zona protein 3 (ZP3) and physiologically
induce the acrosome reaction remain to be conclusively identified (Brewis and
Moore, 1997). Indeed, in the human, about ten sperm determinants in spermatozoa
have been reported to be involved. The most likely scenario is that several sperm
determinants are involved in sperm-zona binding and acrosome reaction, functioning
in concert as part of a complex, while crosstalk can also occur between different
intracellular signalling systems (Kopf et aI., 1995; Brewis and Moore, 1997). [In
Fisher et al., 1998]
Stellenbosch University http://scholar.sun.ac.za

187
A thorough understanding of signal transduction in human spermatozoa will
ultimately yield information regarding the nature of receptors to which these signal
transduction pathways are coupled as well as the intracellular effectors that ultimately
regulate sperm function. Moreover, an understanding of these regulatory pathways
will be essential for the future development of clinical approaches designed to
enhance or preclude fertilization.
ZP3
PDES
1
Plasma membrane
Ca'+
[cGMPj > O.SmM -------
Outer acrosomal membrane
Figure 1: Hypothesised signal transduction pathways and possible interactions
between them. (Dashed lines = hypothesised activation/stimulation)
Stellenbosch University http://scholar.sun.ac.za

188
8.2 Recommendations
In the field of reproductive biology the emphasis are for various reasons currently
shifting away from the female and directed more towards finding solutions for male
contraception and male infertility.
Due to the fact that hormonal manipulation (as is the basis of most female
contraceptives) as a male contraceptive has so far not been very successful as it can
lead to various side effects and even render the male permanently infertile, the need
has arisen to look at other fronts. On the other hand, the cure for male infertility factor
due to asthenozoospermia often leads to expensive intracytoplasmic sperm injection
(IeSI) procedures which is still surrounded by controversy regarding the outcome of
the health of the conceptus (Oehninger, 2001; Ola et al., 2001; Simpson & Lamb,
2001). If the possibility exists to increase the motility of spermatozoa without
interfering with the rest of the sperms normal physiological processes needed for
fertilization, these patients can automatically become candidates for relatively less
expensive IVF treatments (Edirisinghe et aI., 1995).
With this in mind the following recommendations can be made regarding solutions at
the level of intracellular signalling in the spermatozoon:
i) It is recommended that in the quest to preclude fertilization or finding a male
contraceptive the possible role of ERK inhibitors and MAPK agonists and antagonists
need be further investigated. Our data demonstrate that ERKs are directly or
indirectly involved in the acrosome reaction induced by human zona pellucida. The
results also indicate the importance of intact acrosomes to ensure tight binding to the
Stellenbosch University http://scholar.sun.ac.za

189
ZP, while it is generally accepted that the spermatozoa must be acrosome-reacted to
complete penetration of the zona pellucida.
ii) Our results imply that PI3K negatively regulates sperm motility and increase
sperm-zona binding without influencing the acrosome reaction. We subsequently
suggest a possible therapeutic role for PI3K inhibitors in the treatment regime for
asthenozoospermia. This suggests that LY294002 can be used as a tool to enhance
the motility of sperm samples during preparation for IVF from patients with low sperm
motility, thereby ultimately opening a new prospective for severe
oligoasthenozoospermic males to enter IVF rather than ICSI programs.
iii) The ability of sildenafil citrate to enhance sperm kinematics results and sperm-
oocyte binding without affecting the acrosome reaction may be clinically useful.
These findings could have important implications in the use of sildenafil citrate in
assisted reproductive techniques in order to enhance the fertilizing potential of
inherently poor quality sperm with less invasive treatments such as intra-uterine
inseminations, thereby avoiding more involved procedures such as IVF or ICSI.
8.3 Future research
The requirement for sperm capacitation as well as the acrosomal status of the
spermatozoa travelling through the cumulus oophorus needs to be studied further.
Although efforts have been made to establish the characteristics of the proteins
present in the sperm surface that acts as the receptor for ZP3 glycoprotein, the exact
function for these proteins in the mechanism associated to the ZP3-induced AR has
not yet been elucidated. Thus future studies should focus on determining the exact
Stellenbosch University http://scholar.sun.ac.za

190
nature of these receptors, their structure and function. The presence of neuronal
receptors (such as GABA-like and glycine receptor) in mammalian spermatozoa with
a role in AR, either induced by progesterone or solubilized ZP needs also be
investigated (Morales and Llanos, 1996). In addition, the AR, a major regulatory
event for mammalian sperm-oocyte interaction leading to fertilization requires further
studies to integrate the information regarding the role of the different signalling
pathways. These studies may provide the precise sequential role of the intracellular
messengers produced after the spermatozoa-ZP interaction has taken place. The
nature of the zona receptor (ZP2) in the acrosomal-reacted spermatozoa and the
acrosomal proteases in the development of the AR and penetration through the ZP
need to be clarified. Investigations of these and other related questions should
unravel the molecular processes involved in the acrosome reaction.
Stellenbosch University http://scholar.sun.ac.za

191
References
Brewis lA, Moore HOM (1997) Molecular mechanisms of gamete recognition and
fusion at fertilization. Hum Reprod 12 (Natl Suppl, JBFS) 2:156-165.
Calogero AE, Fishel S, Hall J, Ferrara E, Vicari E, Green S, Hunter A, Burrello N,
Thornton S, O'Agata R. (1998) Correlation between intracellular cAMP content,
kinematic parameters and hyperactivation of human spermatozoa after
incubation with pentoxifylline. Hum Reprod 13:911-915.
Cheng CV, Boettcher B (1981) Effect of cAMP, Mn2+ and phosphodiesterase
inhibitors on human sperm motility. Arch Androl 7:313-317.
Edirisinghe WR, Junk S, Yovich JM, Bootsma B, Yovich JL (1995) Sperm stimulants
can improve fertilization rates in male-factor cases undergoing IVF to the same
extent as micromanipulation by partial sona dissection (PZO) or subzonal
sperm insemination (SUZI): a randomized controlled study. JAssist Reprod
Genet 12(5): 312-318.
Fisher HM, Brewis lA, Barrat CL et al., (1998) Phosphoinositide 3-kinase is involved
in the induction of the human sperm scrosome reaction downstream of tyrosine
phosphorylation. Mol Hum Reprod 4:849-855.
Kopf GS, Visconti PE, Moos J et al., (1995) Integration of tyrosine kinase and G
protein mediated signal transduction pathways in the regulation of mammalian
sperm function. In Fenichel Pand Parinaud J (eds), Human Sperm Acrosome
Reaction. John Libbey, Montrouge, pp191-216.
Krasilnikov MA (2000) Phosphatidylinositol-3 kinase dependent pathways: the role in
control of cell growth, survival, and malignant transformation. Biochemistry
(Mosc) 65(1): 59-67.
Stellenbosch University http://scholar.sun.ac.za

192
Leclerc P, de Lamirande E, Gagnon C (1996) Cyclic adenosine 3',5' monophosphate-
dependent regulation of protein tyrosine phosphorylation in relation to human
sperm capacitation and motility. Bioi Reprod 55:684-692.
Lefiévre L, De Lamirande E, Gagnon C (2000) The cyclic GMP-specific
phosphodiesterase inhibitor sildenafil, stimulates human sperm motility and
capacitation but not acrosome reaction. JAndrol 21(6): 929-937.
Lincoln TH, Cornwell TL (1993) Intracellular cyclic GMP receptor proteins. FASEB J
7:328-338
Luconi M, Marra F, Gandini L, Filimberti E, Lenzi A, Forti G, Baldi E (2001)
Phosphatidylinositol 3-kinase inhibition enhances human sperm motility. Hum
Reprod 16(9):1931-1937.
Morales P, Llanos M (1996) Interaction of human spermatozoa with the zona
pellucida of oocyte: development of the acrosome reaction. Front Biosc 1:d146-
160.
Nassar A, Mahoney M, Morshedi M, Lin MH, Srisombut C, Oehninger S (1999)
Modulation of sperm tail protein tyrosine phosphorylation by pentoxifylline and
its correlation with hyperactivation and motility. Fertil Steril 71:919-923.
Oehninger S (2001) Strategies for the infertile man. Semin Reprod Med 19(3): 231-
237.
Ola B, Afnan M, Sharif K, Papaioannou S, Hammadieh N, Barrat CL (2001) Should
ICSI be the treatment of choice for all cases of in vitro conception?
Considerations of fertilization and embryo development, cost effectiveness and
safety. Hum Reprod 16(12): 2485-2490.
Stellenbosch University http://scholar.sun.ac.za

193
Revelli A, Costamagna C, Moffa F, Aldieri E, Ochetti S, Bosia A, Massobrio M,
Lindblom B, Ghigo D (2001) Signalling pathways of nitric oxide-induced
acrosome reaction in human spermatozoa. Bioi Reprod 64(6): 1708-1712.
Rotem R, Zamir N, Keynan N, Barkan D, Breitbart H, Naor Z (1998) Atrial natriuretic
peptide induces acrosomal exocytosis of human spermatozoa. AJP -
Endocrinology and Metabolism: 274(2): E218-E223.
Sears RC, Nevins JR (2002) Signaling networks that link cell proliferation and cell
fate. J Bioi Chem 277(14): 11617-11620.
Simpson JL, Lamb DJ (2001) Genetic effects of intracytoplasmic sperm injection.
Semin Reprod Med 19(3): 239-249.
Tash JS, Bracho GE (1994) Regulation of sperm motility: emerging evidence for a
major role for protein phosphatases. JAndrol 15:505-509.
Yanagimachi R (1994). Mammalian fertilization. In: The physiology of Reproduction.
Knobil E, Neil JD (eds), Vol 1, 2nd ed. New York: Raven Press, 189-317.
Stellenbosch University http://scholar.sun.ac.za

"It's not your blue blood, your pedigree or
your college degree. It's what you do with
your life that counts."
- Millard Fuller -
Stellenbosch University http://scholar.sun.ac.za


















