
ARQUIPELAGO - UAc · ARQUIPELAGO Life and Marine Sciences ... Netherlands; George R. Sedberry,...
96
Transcript of ARQUIPELAGO - UAc · ARQUIPELAGO Life and Marine Sciences ... Netherlands; George R. Sedberry,...


ARQUIPELAGO Life and Marine Sciences SCOPE ARQUIPELAGO - Life and Marine Sciences, publishes annually original scientific articles, short communications and reviews on the terrestrial and marine environment of Atlantic oceanic islands and seamounts. PUBLISHER University of the Azores Rua da Mãe de Deus, 13A PT – 9501-801 Ponta Delgada, Azores, Portugal. EDITOR Helen Rost Martins Phone: + 351 292 207 400 / 428 - Fax: +351 292 207 811 E-mail: [email protected] INTERNET RESOURCES http://www.arquipelago.info Journal information, instructions to authors and free access to all papers. FINANCIAL SUPPORT Secretaria Regional do Mar, Ciência e Tecnologia, do Governo dos Açores e OMA – Observatório do Mar dos Açores, Horta.
EDITORIAL SECRETARIAT Helen R. Martins, Ruth Higgins, José Gomes-Pereira, Emmanuel Arand. EDITORIAL COMMITTEE Paulo A.V. Borges, Angra do Heroísmo; José Azevedo, Ponta Delgada; João M. Gonçalves, Horta.
ADVISORY BOARD Miguel A. Alcaraz, Barcelona, Spain; Alan B. Bolten, Florida, USA; António B. de Sousa, Lisboa, Portugal; Richard D.M. Nash, Bergen, Norway; Erik Sjögren, Uppsala, Sweden; Charles H.J.M. Fransen, Leiden, Netherlands; George R. Sedberry, Georgia, USA; Hanno Schäfer, Munich, Germany; Tony Pitcher, Vancouver, Canada; João Canning Clode, Funchal, Portugal.
Indexed in: Aquatic Sciences and Fisheries Abstracts (ASFA), BIOSIS Previews,
Zoological Record, Directory of Open Access Journals (DOAJ), Web of Science.
Cover design: Emmanuel Arand. Photo: Myliobatis aquila, Faial Island - Nuno Vasco-Rodrigues.
EXCHANGEThe journal is distributed on exchange basis through the editor.
PURCHASE Separate numbers of the Journal can be purchased from the publisher.
OFFPRINTS Authors receive PDF files of their papers. Orders of paper copies can be sent with the proofs if desired.
INSTRUCTIONS TO AUTHORS General. MSS should be submitted to the editor preferably by e-mail ([email protected]). The text should be ready for printing and carefully checked for all errors, including linguistic ones, before submission. English is the recommended language. Choose Font Times New Roman, size 12, double spaced paragraph, and insert page numbers. MSS that are not in accordance with these and the following instructions will be returned for revision. All MSS will be peer-reviewed by two referees prior to their acceptance by the Editor.
Style. 1) Title and 2) Author's name, followed by 3) Abstract (not exceeding 200 words), up to five key words, and (4) the author's name and professional postal address; 5) The main text; 6) Acknowledgements; 7) References; 8) Appendices; 9) Tables with legends; and the 10) List of figures with legends. For research articles the text (5) must be organized as follows: Introduction; Material and methods; Results; Discussion. The Abstract (3) should convey the scope of the paper, emphasizing the results and conclusions, not merely describing its contents. Limit the authors’ professional address (4) to official filiations only.
References. (7) Should correspond with the examples given below:
a) Book: Etgen, W.M. & P.M. Reaves 1978. Dairy Cattle Feeding and Management (6th edition). John Wiley & Sons Inc. New York. 638 pp.
b) Chapter from a book: O’Dor, R., H. O. Pörtner & R. E. Shadwick 1990. Squid as elite athletes: locomotory, respiratory, and circulatory integration. Pp. 481-503 in: Gilbert, D.L., W.J. Adelman & J.M. Arnold (Eds). Squid As Experimental Animals. Plenum Press, New York-London. 516 pp.
c) Article from a journal: Bentley, M.G., P.J.W. Olive, P.R. Garwood & N.H. Wright 1984. The spawning and spawning mechanism of Nephtys caeca (Fabricius, 1780) and Nephtys homebergi Savigny, 1818 (Annelida: Polychaeta). Sarsia 69: 63-68.
d) Electronic article, from online-only Journal: Woo, K.L. 2006. Testing Visual Sensitivity to the Speed and Direction of Motion in Lizards. Journal of Visualized Experiments [Internet]. Available from: http://www.jove.com/index/details.stp?id=127 (cited 18 February 2007).
Use ampersand (&) for all joint authorships in the reference list and for double authorships in the text. Use “et al.” in the text for references with three or more authors and in the reference list for more than seven authors. Avoid multiple citations in text, preferably including the earliest and most recent work. The citation list should be ordered alphabetically by author’s name and year of publication, followed by increasing number of co-authors. Journal names should be written in full.
Figures and Tables. All figures and large tables should be presented with Font Times New Roman, and separately. Photographs should be clear, have high contrast, and in digital format (tiff or jpg) with 300 dpi, with at least 15x15 cm. No original figures should be submitted before the MS has been accepted.
E-mail Attachment. All text file formats are accepted. Attachments must be identified with the author’s last name, the file name and extension, as follows: Bianchi_text.doc, Bianchi_fig1.tif, Bianchi_table1.xls.
For a complete list of format specifications and instructions to authors consult our website at: http://www.arquipelago.info

11
Gabriel, D., W.E. Schmidt, D.M. Krayesky, D.J. Harris & S. Fredericq 2015. The crustose red algal genus Peyssonnelia (Peyssonneliales, Rhodophyta) in the Azores: from five to one species. Arquipelago. Life and Marine Sciences 32: 1-9.
The family Peyssonneliaceae comprises a worldwide group of non-calcified to calcified, crust-forming red algae of great ecological significance. Of the genera currently recognized in the family, Peyssonnelia has been widely considered to contain the largest number of species, with five members reported for the Azores. Using rbcL as a molecular marker, we here report on the taxonomic identity of recent collections of Peyssonneliaceae from the Azorean islands of São Miguel, Graciosa and Pico, and compare those specimens in a worldwide context. Only a single Peyssonnelia species, P. squamaria, is confirmed for the Azorean archipelago, with three different haplotypes. Although the populations in the Azores are genetically different from those occurring in the Mediterranean, this separation appears to be relatively recent.
Key words: Biodiversity, haplotypes, North Atlantic, phylogeny, rbcL
D. Gabriel (e-mail: [email protected]) & D.J. Harris, Research Center in Biodi-versity and Genetic Resources (CIBIO), University of the Azores, PT-9501-801 Ponta Delgada, Portugal; D.M. Krayesky, Biology Department, Slippery Rock University, Slip-pery Rock, PA 16057-1326, USA; W. E. Schmidt & S. Fredericq, Department of Biology, University of Louisiana at Lafayette, Lafayette, LA 70504-3602, USA.
INTRODUCTION
The family Peyssonneliaceae (Denizot, 1968), recently elevated to ordinal rank (Krayesky et al. 2009), comprises a worldwide group of non-calcified or calcified, crust-forming red algae that are of great ecological significance (Peña & Bar-bara 2013). Of the genera currently recognized in the family, Peyssonnelia (Decaisne, 1841) has been considered the richest in terms of species number (Pueschel & Saunders 2009). A combina-tion of vegetative and reproductive characters are currently used to distinguish species of Peyssonnelia, such as location and degree of cal-cification, variations in crust adherence, mor-phology, anatomy and differences in reproductive development (Maggs & Irvine 1983). The identi-fication of Peyssonnelia species is challenging, resulting in a number of species usually underes-timated or overestimated (Dixon & Saunders
2013). Comparative morphology and DNA se-quence analysis confirm that most species origi-nally reported as belonging in Peyssonnelia in fact belong to other genera (Fredericq et al. 2014) within the Peyssonneliales (Krayesky et al. 2009). Peyssonnelia sensu stricto (following Krayesky et al. 2009) represents species characterized by a hypothallus that cuts off additional cells forming multicellular rhizoids (Krayesky 2007). Recent studies based on worldwide collections indicate that species of Peyssonnelia sensu stricto have a narrow distribution and do not occur in most ocean basins, for example, the Gulf of Mexico (Krayesky et al. 2009; Fredericq et al. 2014). Based on general flora studies, five species of Peyssonnelia have been reported for the Azores (Parente 2010): the generitype P. squamaria ((S.G. Gmelin) Decaisne, 1842) described from Italy; P. rubra ((Greville) J. Agardh, 1851) de-scribed from the Ionian Sea, Greece; P. polymor-

2
Gabriel et al.
3
on with correction (AICc), Akaike information criterion (AIC) and Bayesian information criteri-on (BIC). The alignment was analyzed by Maxi-mum likelihood (ML) as implemented by RAXML v 2.4.4 (Stamatakis 2006) with the above models and partition scheme with 1000 restarts to find the tree with the lowest likelihood score and 1000 Bootstrap (BS) replications. A Bayesian MCMC (Markov Chain Monte Carlo) was also applied to the aligned dataset using MrBayes v. 3.2.5 (Huelsenbeck & Ronquist 2001; Ronquist & Huelsenbeck 2003). The Bayesian analysis consisted of two independent runs of 5 million generations with sampling every 1,000 generations for a total of 10,002 trees. Convergence was visualized using Tracer v1.6 (Rambaut & Drummond 2007) and the first 10 percent of the trees of each run was discarded as the burn-in. The resulting Bayesian Posterior Probabilities derived from the consensus tree were mapped on the ML tree. A distance matrix was also resolved from the branch lengths of the ML tree using the function cophenetic.phylo of the APE Package in R (Paradis et al. 2004). The resulting distance matrix was used to find species boundaries in a stand-alone version of Automatic to Barcode Gap Discovery (ABGD). General Mixed Yule Coalescence (GMYC) mod-el, as implemented by the Splits Package in R (Fujisawa & Barraclough 2013) with a single threshold model, was also used to determine spe-cies boundaries. The requisite ultrametric tree for the GMYC analysis was generated in Beast v 1.8.1 (Drummond et al. 2012) using a relaxed log-normal clock with a constant population coa-lescent as a prior and the best fitting model and partition as described above. MCMC Chains were run for 10 million generations with sampling eve-ry 1000th generation resulting in 10,000 trees. The quality of the run was assessed in Tracer v1.6 (Rambaut & Drummond 2007) to ensure that ef-fective sample size (ESS) values were >200 with the default burn-in (1,000 trees). Tree annotator v 1.8.1 (Drummond et al. 2012) was used to sum-marize the resulting 9001 trees after burning, tar-geting the maximum clade credibility tree with preserved node heights. A statistical parsimony method implemented in the TCS 1.21 software
(Clement et al. 2000) was used to infer genealog-ical relationships among haplotypes. The maxi-mum number of differences resulting from single substitutions among haplotypes was calculated with 95% confidence limits, treating gaps as missing data.
RESULTS
The final dataset was composed of 34 rbcL se-quences of Peyssonneliaceae, 20 of which were newly generated, with 31 sequences representing hitherto confirmed Peyssonnelia species and three sequences of Sonderopelta capensis ((Montagne) Krayesky, 2009) and S. coriacea (Womersley & Sinkora, 1981) (Table 1). Sonderopelta (Womers-ley & Sinkora, 1981) was selected as an outgroup based on previous studies that established the genus as a sister taxon of Peyssonnelia (Kato et al. 2006; Krayesky et al. 2009; Dixon & Saunders 2013). [Note: Sonderopelta has been viewed to be an illegitimate name by Wynne (2011); however, Sonderophycus (Denizot, 1968) is not a valid name (Womersley & Sinkora 1981) in agreement with Article 41.5 of the International Code of Botanical Nomenclature (2012, Melbourne Code)]. The results of the ABGD and GMYC analyses showed the existence of six species of Peyssonnelia within the dataset: P. replicata (Kützing, 1847), P. bornetii (Boudouresque & Denizot, 1973), P. rubra and three closely related species initially identified as P. squamaria (Fig. 2). All Azorean collections belonged to P. squa-maria, along with Mediterranean representatives from Catalonia (Spain), Sicily (Italy), Malta and Greece (not shown). Its two sister clades were only observed in the Mediterranean, and are here referred to as P. coriacea from Malta and P. pol-ymorpha from Sicily. Sequence JX969797, re-ferred to as P. squamaria by Dixon & Saunders (2013), corresponds to material of Sicily that has a unistratose hypothallus layer in contrast to the 2-layered hypothallus of P. squamaria (Boudour-esque & Denizot, 1975). Besides the generitype, four species that are also true Peyssonnelia were only collected in the
2
pha ((Zanardini) F. Schmitz in Falkenberg, 1879) described from the Adriatic Sea; P. coriacea (Feldmann, 1941) described from Tangier, Mo-rocco; and P. rosa-marina (Boudouresque & Denizot, 1973) described from Port-Cros, Medi-terranean France (see Guiry & Guiry 2015). Only one species of Peyssonnelia, P. squamaria (Fig. 1), was recognized for the Azores by Krayesky (2007) and Krayesky et al. (2009) after examina-tion of multiple collections.
Using rbcL as a molecular marker, we report on the taxonomic identity of recent collections of Peyssonneliaceae from the islands of São Miguel, Graciosa and Pico in the Azores, and discuss the connection between the various Azorean haplo-types. The identity of the Azorean specimens is compared with those of Peyssonnelia sensu stric-to in a worldwide context.
Fig. 1. Habit of Peyssonnelia squamaria from the Azores.
MATERIAL AND METHODS
Samples of Peyssonneliaceae were collected in the Azores and the Mediterranean during low tide or by snorkeling and SCUBA diving. Samples were kept in coolers until processed, and then dried in silica gel. Dried samples were ground with mortar and pestle, and total DNA was ex-tracted using DNeasy Plant mini Kits (Qiagen Valencia, CA, USA). All the resulting DNA ex-tracts were deposited in the Seaweed Lab at the University of Louisiana at Lafayette (ULL). Chloroplast-encoded rbcL gene sequences were amplified using PCR primers and protocols de-scribed in Lin et al. (2001) and Gabriel et al. (2010). Resulting PCR products were gel-purified
and sequenced in both directions using Bigdye terminator v 3.1 (Life Technologies Grand Island NY, USA) on the ABI 3130xl genetic analyzer at ULL and assembled with Sequencher v. 5.2 (Gene Codes Corporation). Newly acquired se-quences, in addition to 14 rbcL sequences down-loaded from GenBank, were then manually aligned in Mega v 5.2.2 (Tamura et al. 2011). The subsequent alignment was analyzed in Partition-finder (Lanfear et al. 2012) to determine the best fitting model of evolution and data partition. The analysis resulted in the selection of the General Time Reversible model plus gamma and a pro-portion of invariable sites applied separately to each codon position on the basis of the three in-formation criteria, i.e. Akaike information criteri-
2
pha ((Zanardini) F. Schmitz in Falkenberg, 1879) described from the Adriatic Sea; P. coriacea (Feldmann, 1941) described from Tangier, Mo-rocco; and P. rosa-marina (Boudouresque & Denizot, 1973) described from Port-Cros, Medi-terranean France (see Guiry & Guiry 2015). Only one species of Peyssonnelia, P. squamaria (Fig. 1), was recognized for the Azores by Krayesky (2007) and Krayesky et al. (2009) after examina-tion of multiple collections.
Using rbcL as a molecular marker, we report on the taxonomic identity of recent collections of Peyssonneliaceae from the islands of São Miguel, Graciosa and Pico in the Azores, and discuss the connection between the various Azorean haplo-types. The identity of the Azorean specimens is compared with those of Peyssonnelia sensu stric-to in a worldwide context.
Fig. 1. Habit of Peyssonnelia squamaria from the Azores.
MATERIAL AND METHODS
Samples of Peyssonneliaceae were collected in the Azores and the Mediterranean during low tide or by snorkeling and SCUBA diving. Samples were kept in coolers until processed, and then dried in silica gel. Dried samples were ground with mortar and pestle, and total DNA was ex-tracted using DNeasy Plant mini Kits (Qiagen Valencia, CA, USA). All the resulting DNA ex-tracts were deposited in the Seaweed Lab at the University of Louisiana at Lafayette (ULL). Chloroplast-encoded rbcL gene sequences were amplified using PCR primers and protocols de-scribed in Lin et al. (2001) and Gabriel et al. (2010). Resulting PCR products were gel-purified
and sequenced in both directions using Bigdye terminator v 3.1 (Life Technologies Grand Island NY, USA) on the ABI 3130xl genetic analyzer at ULL and assembled with Sequencher v. 5.2 (Gene Codes Corporation). Newly acquired se-quences, in addition to 14 rbcL sequences down-loaded from GenBank, were then manually aligned in Mega v 5.2.2 (Tamura et al. 2011). The subsequent alignment was analyzed in Partition-finder (Lanfear et al. 2012) to determine the best fitting model of evolution and data partition. The analysis resulted in the selection of the General Time Reversible model plus gamma and a pro-portion of invariable sites applied separately to each codon position on the basis of the three in-formation criteria, i.e. Akaike information criteri-

3
Peyssonnelia in the Azores
3
on with correction (AICc), Akaike information criterion (AIC) and Bayesian information criteri-on (BIC). The alignment was analyzed by Maxi-mum likelihood (ML) as implemented by RAXML v 2.4.4 (Stamatakis 2006) with the above models and partition scheme with 1000 restarts to find the tree with the lowest likelihood score and 1000 Bootstrap (BS) replications. A Bayesian MCMC (Markov Chain Monte Carlo) was also applied to the aligned dataset using MrBayes v. 3.2.5 (Huelsenbeck & Ronquist 2001; Ronquist & Huelsenbeck 2003). The Bayesian analysis consisted of two independent runs of 5 million generations with sampling every 1,000 generations for a total of 10,002 trees. Convergence was visualized using Tracer v1.6 (Rambaut & Drummond 2007) and the first 10 percent of the trees of each run was discarded as the burn-in. The resulting Bayesian Posterior Probabilities derived from the consensus tree were mapped on the ML tree. A distance matrix was also resolved from the branch lengths of the ML tree using the function cophenetic.phylo of the APE Package in R (Paradis et al. 2004). The resulting distance matrix was used to find species boundaries in a stand-alone version of Automatic to Barcode Gap Discovery (ABGD). General Mixed Yule Coalescence (GMYC) mod-el, as implemented by the Splits Package in R (Fujisawa & Barraclough 2013) with a single threshold model, was also used to determine spe-cies boundaries. The requisite ultrametric tree for the GMYC analysis was generated in Beast v 1.8.1 (Drummond et al. 2012) using a relaxed log-normal clock with a constant population coa-lescent as a prior and the best fitting model and partition as described above. MCMC Chains were run for 10 million generations with sampling eve-ry 1000th generation resulting in 10,000 trees. The quality of the run was assessed in Tracer v1.6 (Rambaut & Drummond 2007) to ensure that ef-fective sample size (ESS) values were >200 with the default burn-in (1,000 trees). Tree annotator v 1.8.1 (Drummond et al. 2012) was used to sum-marize the resulting 9001 trees after burning, tar-geting the maximum clade credibility tree with preserved node heights. A statistical parsimony method implemented in the TCS 1.21 software
(Clement et al. 2000) was used to infer genealog-ical relationships among haplotypes. The maxi-mum number of differences resulting from single substitutions among haplotypes was calculated with 95% confidence limits, treating gaps as missing data.
RESULTS
The final dataset was composed of 34 rbcL se-quences of Peyssonneliaceae, 20 of which were newly generated, with 31 sequences representing hitherto confirmed Peyssonnelia species and three sequences of Sonderopelta capensis ((Montagne) Krayesky, 2009) and S. coriacea (Womersley & Sinkora, 1981) (Table 1). Sonderopelta (Womers-ley & Sinkora, 1981) was selected as an outgroup based on previous studies that established the genus as a sister taxon of Peyssonnelia (Kato et al. 2006; Krayesky et al. 2009; Dixon & Saunders 2013). [Note: Sonderopelta has been viewed to be an illegitimate name by Wynne (2011); however, Sonderophycus (Denizot, 1968) is not a valid name (Womersley & Sinkora 1981) in agreement with Article 41.5 of the International Code of Botanical Nomenclature (2012, Melbourne Code)]. The results of the ABGD and GMYC analyses showed the existence of six species of Peyssonnelia within the dataset: P. replicata (Kützing, 1847), P. bornetii (Boudouresque & Denizot, 1973), P. rubra and three closely related species initially identified as P. squamaria (Fig. 2). All Azorean collections belonged to P. squa-maria, along with Mediterranean representatives from Catalonia (Spain), Sicily (Italy), Malta and Greece (not shown). Its two sister clades were only observed in the Mediterranean, and are here referred to as P. coriacea from Malta and P. pol-ymorpha from Sicily. Sequence JX969797, re-ferred to as P. squamaria by Dixon & Saunders (2013), corresponds to material of Sicily that has a unistratose hypothallus layer in contrast to the 2-layered hypothallus of P. squamaria (Boudour-esque & Denizot, 1975). Besides the generitype, four species that are also true Peyssonnelia were only collected in the

4
Gabriel et al.Ta
ble 1
. Sum
mar
y of
spec
imen
s inc
lude
d in
the p
rese
nt st
udy.
Aste
risk
mar
ks n
ewly
gen
erat
ed se
quen
ces.
Lette
rs w
ithin
bra
cket
s ref
er to
the h
aplo
type
s of P
eyss
onne
lia sq
uam
aria
in F
ig. 3
.
Extr
actio
n C
olle
ctio
n nu
mbe
r Ta
xa
Loca
lity
Dep
th
Col
lect
ion
date
C
olle
ctor
A
cces
sion
num
ber
K16
3 SM
G-0
5-24
2 Pe
ysso
nnel
ia sq
uam
aria
(A1)
Sã
o M
igue
l, A
zore
s in
terti
dal
09-ju
l-200
5 EU
3491
77
K16
4 SM
G-0
5-15
2 Pe
ysso
nnel
ia sq
uam
aria
(A1)
Sã
o M
igue
l, A
zore
s 8
m
08-o
ct-2
005
EU34
9178
LAF4
197
MD
0002
162
Peys
sonn
elia
squa
mar
ia (A
1)
Pico
, Azo
res
26 m
10
-aug
-201
1 D
. Gab
riel,
J. M
icae
l K
R732
897*
LAF6
390
MD
0002
066
Peys
sonn
elia
squa
mar
ia (A
1)
São
Mig
uel,
Azo
res
inte
rtida
l 10
-jul-2
011
M.I.
Par
ente
, D. G
abrie
l K
R732
900*
LAF6
393
MD
0001
927
Peys
sonn
elia
squa
mar
ia (A
1)
São
Mig
uel,
Azo
res
15 m
29
-oct
-201
0 A
. Bot
elho
K
R732
899*
LAF6
395
MD
0001
933
Peys
sonn
elia
squa
mar
ia (A
1)
São
Mig
uel,
Azo
res
12 m
05
-jul-2
011
A. B
otel
ho, M
. Dio
nísio
, C.
Lope
s K
R732
898*
K22
9 SM
G-0
6-88
Pe
ysso
nnel
ia sq
uam
aria
(A2)
Sã
o M
igue
l, A
zore
s 16
m
11-a
ug-2
006
EU34
9174
K23
1 G
RW-0
6-10
0 Pe
ysso
nnel
ia sq
uam
aria
(A3)
G
raci
osa,
Azo
res
15 m
22
-jun-
2006
EU
3491
76
K23
0 CA
T-06
-10
Peys
sonn
elia
squa
mar
ia (B
) Ca
talo
nia,
Spai
n in
terti
dal
16-ju
l-200
6 EU
3491
75
LAF5
459
PG-0
8-13
53
Peys
sonn
elia
squa
mar
ia (C
) M
alta
10
m
06-a
ug-2
008
M.I.
Par
ente
, J. M
icae
l K
R732
910*
LAF5
357
PG-0
8-12
10
Peys
sonn
elia
squa
mar
ia (D
) M
alta
15
m
02-a
ug-2
008
M.I.
Par
ente
K
R732
905*
LAF5
360
PG-0
8-11
46
Peys
sonn
elia
squa
mar
ia (D
) M
alta
30
m
02-a
ug-2
008
M.I.
Par
ente
, J. M
icae
l K
R732
909*
LAF5
362
PG-0
8-12
48
Peys
sonn
elia
squa
mar
ia (D
) M
alta
26
m
05-a
ug-2
008
M.I.
Par
ente
, J. M
icae
l K
R732
901*
LAF5
363
PG-0
8-10
88
Peys
sonn
elia
squa
mar
ia (D
) M
alta
30
m
02-a
ug-2
008
M.I.
Par
ente
, J. M
icae
l K
R732
902*
LAF5
455
PG-0
8-11
00
Peys
sonn
elia
squa
mar
ia (D
) M
alta
30
m
02-a
ug-2
008
M.I.
Par
ente
, J. M
icae
l K
R732
907*
LAF5
458
PG-0
8-11
53
Peys
sonn
elia
squa
mar
ia (D
) M
alta
30
m
02-a
ug-2
008
M.I.
Par
ente
, J. M
icae
l K
R732
904*
LAF5
463
PG-0
8-79
5 Pe
ysso
nnel
ia sq
uam
aria
(D)
Sici
ly, I
taly
in
terti
dal
12-m
ar-2
008
M.I.
Par
ente
, R. S
ousa
, J.
Mat
zen
KR7
3290
3*

5
Peyssonnelia in the Azores
Extr
actio
n C
olle
ctio
n nu
mbe
r Ta
xa
Loca
lity
Dep
th
Col
lect
ion
date
C
olle
ctor
A
cces
sion
num
ber
LAF5
464
PG-0
8-75
6 Pe
ysso
nnel
ia sq
uam
aria
(D)
Sici
ly, I
taly
in
terti
dal
12-m
ar-2
008
M.I.
Par
ente
, R. S
ousa
, J.
Mat
zen
KR7
3290
6*
LAF5
465
PG-0
8-74
7 Pe
ysso
nnel
ia sq
uam
aria
(D)
Sici
ly, I
taly
in
terti
dal
12-m
ar-2
008
M.I.
Par
ente
K
R732
908*
LAF5
355
PG-0
8-12
68
Peys
sonn
elia
coria
cea
Mal
ta
26 m
05
-aug
-200
8 M
.I. P
aren
te, J
. Mic
ael
KR7
3291
1*
LAF5
361
PG-0
8-11
73
Peys
sonn
elia
coria
cea
Mal
ta
30 m
02
-aug
-200
8 M
.I. P
aren
te, J
. Mic
ael
KR7
3291
2*
LAF5
457
PG-0
8-12
91
Peys
sonn
elia
coria
cea
Mal
ta
26 m
05
-aug
-200
8 M
.I. P
aren
te, J
. Mic
ael
KR7
3291
3*
LAF5
461
PG-0
8-80
4 Pe
ysso
nnel
ia p
olym
orph
a Si
cily
, Ita
ly
inte
rtida
l 12
-mar
-200
8 M
.I. P
aren
te, R
. Sou
sa, J
. M
atze
n K
R732
915*
LAF5
462
PG-0
8-79
0 Pe
ysso
nnel
ia p
olym
orph
a Si
cily
, Ita
ly
inte
rtida
l 12
-mar
-200
8 M
.I. P
aren
te, R
. Sou
sa, J
. M
atze
n K
R732
916*
- G
WS0
1817
9 Pe
ysso
nnel
ia p
olym
orph
a Si
cily
, Ita
ly
- 10
-apr
-201
0 G
. Fur
nari
JX96
9797
LAF4
199
PG-0
8-55
1 Pe
ysso
nnel
ia p
olym
orph
a Si
cily
, Ita
ly
inte
rtida
l 09
-mar
-200
8 M
.I. P
aren
te
KR7
3291
4*
K16
6 LA
F-8-
2-1-
1-12
Pe
ysso
nnel
ia ru
bra
Ligu
ria, I
taly
3
m
02-a
ug-2
001
B. G
avio
EU
3491
79
K21
7 LA
F-7-
30-1
-1-2
Pe
ysso
nnel
ia b
orne
tii
Ligu
ria, I
taly
3
m
30-ju
l-200
1 B.
Gav
io
EU34
9180
K21
8 LA
F-7-
28-1
-1-2
Pe
ysso
nnel
ia b
orne
tii
Ligu
ria, I
taly
2-
20 m
28
-jul-2
001
B. G
avio
EU
3491
81
K24
1 LA
F-2-
6-01
-2-2
Pe
ysso
nnel
ia re
plic
ata
Kw
aZul
u-N
atal
, Sou
th A
frica
in
terti
dal
06-fe
b-20
01
T. S
chils
EU
3491
82
K24
3 LA
F-7-
23-9
3-1-
1 Pe
ysso
nnel
ia re
plic
ata
Kw
aZul
u-N
atal
, Sou
th A
frica
D
rift
(bea
ch)
23-ju
l-199
3 M
. Hom
mer
sand
EU
3491
83
K21
4 LA
F-2-
6-01
-1-1
5 So
nder
opel
ta ca
pens
is K
waZ
ulu-
Nat
al, S
outh
Afri
ca
30 m
06
-feb-
2001
S.
Fre
deric
q &
O. D
e Cle
rck
EU34
9186
K21
5 LA
F-2-
6-01
-1-1
So
nder
opel
ta ca
pens
is K
waZ
ulu-
Nat
al, S
outh
Afri
ca
30 m
06
-feb-
2001
S.
Fre
deric
q &
O. D
e Cle
rck
EU34
9187
K22
0 LA
F-7-
13-9
5-1-
1 So
nder
opel
ta co
riace
a V
icto
ria, A
ustra
lia
- 13
-jul-1
995
M. H
omm
ersa
nd
EU34
9190
�
6
Gabriel et al.
7
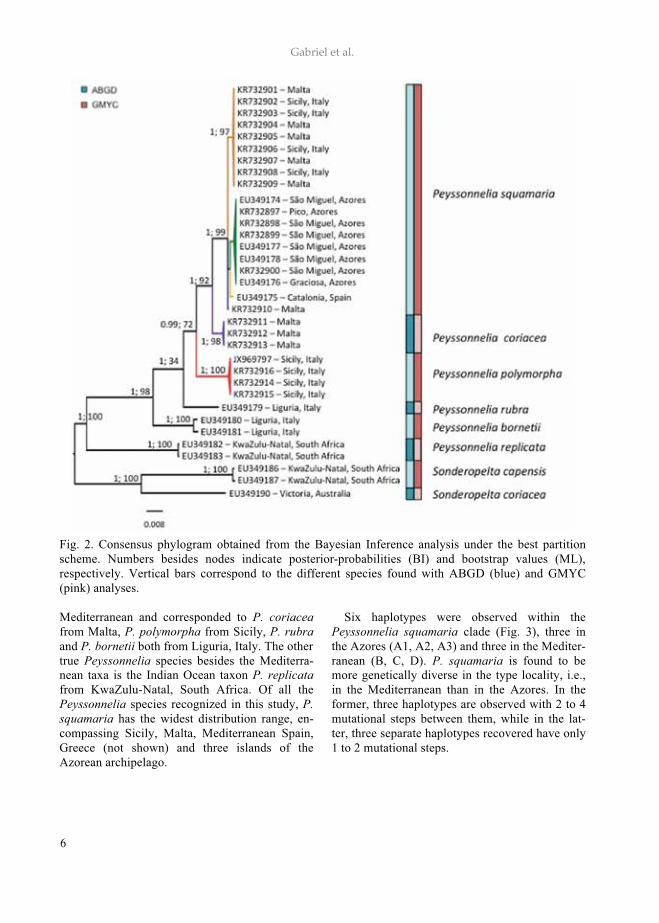
Fig. 3. RbcL haplotype network of Peyssonnelia squamaria and its spatial distribution in the North Atlantic. Haplotype and population sizes are proportional to the number of individual;dach haplotype is represented by a color and a letter (A1 to D). CONCLUSION
Only a single, true Peyssonnelia species is con-firmed for the Azorean Archipelago, in contrast to the five previously reported (Parente 2010). Alt-hough the populations in the Azores are genet-ically different from those occurring in the Medi-terranean, this separation might be relatively re-cent, since the archipelago emerged about 8 My ago (Rumeu et al. 2011). Further studies includ-ing more islands and more samples are necessary to assess the variability of the species within the archipelago and the connection between its popu-lations (Gabriel et al. 2014).
ACKNOWLEDGMENTS
We greatly acknowledge support from a grant from the National Science Foundation Systemat-ics Program (DEB-1027110 to SF). During this research, DG was supported by FCT grant SFRH/BPD/64963/2009. We are grateful to Ma-nuela I. Parente for granting access to her sample collection, to Joana Micael for helping with the haplotype networks, and to António Medeiros for providing the maps.
6
Fig. 2. Consensus phylogram obtained from the Bayesian Inference analysis under the best partition scheme. Numbers besides nodes indicate posterior-probabilities (BI) and bootstrap values (ML), respectively. Vertical bars correspond to the different species found with ABGD (blue) and GMYC (pink) analyses. Mediterranean and corresponded to P. coriacea from Malta, P. polymorpha from Sicily, P. rubra and P. bornetii both from Liguria, Italy. The other true Peyssonnelia species besides the Mediterra-nean taxa is the Indian Ocean taxon P. replicata from KwaZulu-Natal, South Africa. Of all the Peyssonnelia species recognized in this study, P. squamaria has the widest distribution range, en-compassing Sicily, Malta, Mediterranean Spain, Greece (not shown) and three islands of the Azorean archipelago.
Six haplotypes were observed within the Peyssonnelia squamaria clade (Fig. 3), three in the Azores (A1, A2, A3) and three in the Mediter-ranean (B, C, D). P. squamaria is found to be more genetically diverse in the type locality, i.e., in the Mediterranean than in the Azores. In the former, three haplotypes are observed with 2 to 4 mutational steps between them, while in the lat-ter, three separate haplotypes recovered have only 1 to 2 mutational steps.

7
Peyssonnelia in the Azores
7
Fig. 3. RbcL haplotype network of Peyssonnelia squamaria and its spatial distribution in the North Atlantic. Haplotype and population sizes are proportional to the number of individual;dach haplotype is represented by a color and a letter (A1 to D). CONCLUSION
Only a single, true Peyssonnelia species is con-firmed for the Azorean Archipelago, in contrast to the five previously reported (Parente 2010). Alt-hough the populations in the Azores are genet-ically different from those occurring in the Medi-terranean, this separation might be relatively re-cent, since the archipelago emerged about 8 My ago (Rumeu et al. 2011). Further studies includ-ing more islands and more samples are necessary to assess the variability of the species within the archipelago and the connection between its popu-lations (Gabriel et al. 2014).
ACKNOWLEDGMENTS
We greatly acknowledge support from a grant from the National Science Foundation Systemat-ics Program (DEB-1027110 to SF). During this research, DG was supported by FCT grant SFRH/BPD/64963/2009. We are grateful to Ma-nuela I. Parente for granting access to her sample collection, to Joana Micael for helping with the haplotype networks, and to António Medeiros for providing the maps.

8
Gabriel et al.
9
2011. The colonization history of Juniperus brevifolia (Cupressaceae) in the Azores Islands. PLoS ONE 6(11): e27697.
Stamatakis, A. 2006 - RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22: 2688-2690.
Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei & S. Kumar 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum
parsimony methods. Molecular Biology and Evolution 28: 2731-2739.
Womersley, H.B.S. & D. Sinkora 1981. Sonderophycus and the type specimen of Peyssonnelia australis (Cryptonemiales, Rhodophyta). Transactions Royal Society of South Australia 105: 85-87.
Wynne, M.J. 2011. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: third revision. Nova Hedwigia Beihefte 140: 7-166.
Received 2 May 2015. Accepted 12 Aug 2015 Published online 14 Sept 2015.
���������������
8
REFERENCES
Boudouresque, C.-F. & M. Denizot 1975. Révision du genre Peyssonnelia (Rhodophyta) en Médi-terranée. Bulletin Museum Histoire Naturelle Marseille 35: 7-92.
Clement, M., D. Posada & K.A. Crandall 2000. TCS: a computer program to estimate gene genealogies. Molecular Ecology 9: 1657–1659.
Dixon, K.R. & G.W. Saunders 2013. DNA barcoding and phylogenetics of Ramicrusta and Incendia gen. nov., two early diverging lineages of the Peyssonneliaceae (Rhodophyta). Phycologia 52: 82-108.
Drummond A.J., M.A. Suchard, D. Xie & A. Rambaut. 2012. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Molecular Biology and Evolution 29: 1969-1973.
Fredericq, S., N. Arakaki, O. Camacho, D. Gabriel, D. Krayesky, S. Self-Krayesky, G. Rees, J. Richards, T. Sauvage, D. Venera-Ponton & W.E. Schmidt 2014. A dynamic approach to the study of rhodoliths: a case study for the Northwestern Gulf of Mexico. Cryptogamie Algologie 35: 77-98.
Fujisawa, T. & T.C. Barraclough 2013. Delimiting species using single-locus data and the Generalized Mixed Yule Coalescent Approach: A revised method and evaluation on simulated data sets. Systematic Biology 62: 707-724.
Gabriel, D., M.I. Parente, A.I. Neto, M. Raposo, T. Schils & S. Fredericq 2010. Phylogenetic appraisal of the genus Platoma (Nemastomatales, Rhodophyta), including life history and morphological observations on P. cyclocolpum from the Azores. Phycologia 49: 2-21.
Gabriel D., M.I. Parente & S. Fredericq 2014. The algal diaspora. The algal diaspora. Pp. 47-49 in: Carreira, G., R.M. Higgins, F. Cardigos & F.M. Porteiro (Eds). The sea of the Azores: scientific forum for decision support. Arquipelago. Life and Marine Sciences. Supplement 8.
Guiry, M.D. & G.M. Guiry 2015. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; (Cited 07 May 2015).
Huelsenbeck, J.P. & F. Ronquist 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17: 754-5.
International Code of Nomenclature for algae, fungi, and plants (Melbourne Code). 2012. Regnum Vegetabile 154. Koeltz Scientific Books. ISBN 978-3-87429-425-6.
Kato, A., M. Baba, H. Kawai, & M. Masuda 2006. Reassessment of the little-known crustose red algal genus Polystrata (Gigartinales), based on mor-
phology and SSU rDNA sequences. Journal of Phycology 42: 922-933.
Krayesky, D.M. 2007. Phylogenetic and Developmental Studies in Selected Red Algae, with a Focus on Taxa from the Gulf of Mexico. Unpubl. Ph.D. Dissertation, The University of Louisiana at Lafayette, 259 pp.
Krayesky, D., J.N. Norris, P.W. Gabrielson, D. Gabriel & S. Fredericq 2009. A new order of red algae based on the Peyssonneliaceae, with an evaluation of the ordinal classification of the Florideophyceae (Rhodophyta). Proceedings of the Biological Society of Washington 122: 364–391.
Lanfear, R., B. Calcott, S.Y.W Ho & S. Guindon 2012. PartitionFinder: combined selection of partitioning schemes and substitution models for phylogenetic Analyses. Molecular Biology and Evolution 29: 1695-1701.
Lin, S.-M., S. Fredericq & M.H. Hommersand 2001. Systematics of the Delesseriaceae (Ceramiales, Rhodophyta) based on large subunit rDNA and rbcL sequences, including the Phycodryoideae, subfam. nov. Journal of Phycology 37: 881–899.
Maggs, C.A. & L.M. Irvine 1983. Peyssonnelia immersa sp. nov. (Cryptonemiales, Rhodophyta) from the British Isles and France, with a survey of infrageneric classification. British Phycological Journal 18: 219-238.
Paradis, E., J. Claude & K. Strimmer 2004. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 20: 289-290.
Parente, M.I. 2010. List of the marine macroalgae (Rhodophyta, Chlorophyta, Phaeophyceae). In: Borges, P.A.V., A. Costa, R. Cunha, R. Gabriel, V. Gonçalves, A.F. Martins, I. Melo, M. Parente, P. Raposeiro, P. Rodrigues, R.S. Santos, L. Silva, P. Vieira & V. Vieira (Eds) A list of the terrestrial and marine biota from the Azores. pp. 9-33, Princípia, Cascais, 432 pp.
Peña, V. & I. Barbara 2013. Non-coralline crustose algae associated with maerl beds in Portugal: a reappraisal of their diversity in the Atlantic Iberian beds. Botanica Marina 55: 481-493.
Pueschel, C.M. & G.W. Saunders 2009. Ramicrusta textilis sp. nov. (Peyssonneliaceae, Rhodophyta), an anatomically complex Caribbean alga that overgrows corals. Phycologia 48: 480-491.
Rambaut, A. & A. Drummond 2007. Tracer v1.6. Available from: http://tree.bio.ed.ac.uk/software/tracer.
Ronquist, F. & J.P. Huelsenbeck 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics19: 1572-1574.
Rumeu, B., J. Caujapé-Castells, J.L. Blanco-Pastor, R. Jaén-Molina, M. Nogales, R.B. Elias & P. Vargas

9
Peyssonnelia in the Azores
9
2011. The colonization history of Juniperus brevifolia (Cupressaceae) in the Azores Islands. PLoS ONE 6(11): e27697.
Stamatakis, A. 2006 - RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22: 2688-2690.
Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei & S. Kumar 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum
parsimony methods. Molecular Biology and Evolution 28: 2731-2739.
Womersley, H.B.S. & D. Sinkora 1981. Sonderophycus and the type specimen of Peyssonnelia australis (Cryptonemiales, Rhodophyta). Transactions Royal Society of South Australia 105: 85-87.
Wynne, M.J. 2011. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: third revision. Nova Hedwigia Beihefte 140: 7-166.
Received 2 May 2015. Accepted 12 Aug 2015 Published online 14 Sept 2015.

11
Paiva, L.S., E.M.C. Lima, A.I. Neto & J.A.B. Baptista 2015. Screening for angio-tensin I-converting enzyme (ACE) inhibitory activity of enzymatic hydrolysates obtained from Azorean macroalgae. Arquipelago. Life and Marine Sciences 32: 11-17.
As part of ongoing studies into the isolation of antihypertensive peptides, 12 common macroalgae species from Azores were investigated for their potential angiotensin-converting enzyme (ACE) inhibitory properties. After optimization of enzymatic hydrolysis conditions, the anti-ACE activity was determined by high performance liquid chromatog-raphy (HPLC). The species under study include four Rhodophyta (Gelidium microdon, Os-mundea pinnatifida, Porphyra sp. and Pterocladiella capillacea), four Chlorophyta (Chae-tomorpha linum, Codium adhaerens, Ulva compressa and Ulva rigida) and four Heterokon-tophyta (Cystoseira humilis, Fucus spiralis, Padina pavonica and Sargassum cymosum). The bromelain hydrolysates derived from P. capillacea, Porphyra sp., F. spiralis, U. com-pressa, O. pinnatifida and U. rigida were the most active presenting an ACE inhibitory ac-tivity of 48.73 ± 1.58%, 50.96 ± 1.82%, 56.77 ± 1.80%, 59.80 ± 0.95%, 62.59 ± 1.60% and 65.68 ± 0.72%, respectively. These results suggest that some macroalgae from the littoral Azorean sea are potential sources of powerful ACE inhibitor peptides, with potential impact on public health, particularly on the hypertensive patients. These features make these algae valuable to the regional economy and may also provide an incentive for marine biodiversity conservation and valuation of marine algal resources.
Key words: Azores, marine macroalgae, antihypertensive peptides, ACE inhibition, enzy-matic hydrolysis, pepsin, bromelain, HPLC
Lisete S. Paiva (e-mail: [email protected]), Elisabete M.C. Lima & José A.B. Baptista, CITA-A – Research Center for Agricultural Technology & Department of Technological Sciences and Development, University of Azores, 9501-801 Ponta Delgada, S. Miguel, Azores, Portugal; Lisete S. Paiva & Ana I. Neto, Marine Biology Group, Department of Biology, University of Azores, 9501-801 Ponta Delgada, S. Miguel, Azores, Portugal; Ana I. Neto, cE3c – Centre for Ecology, Evolution and Environmental Changes/ Azorean Biodiversity Group and University of Azores, Department of Biology, 9501-801 Ponta Delgada, S. Miguel, Azores, Portugal.
INTRODUCTION
Hypertension, one of the most common serious chronic health problems, affects about 25% of the world’s adult population (Mittal & Singh 2010). This problem of epidemic proportions carries a high risk factor for arteriosclerosis, stroke, myo-
cardial infarction and end-stage renal disease (Zhang et al. 2006). It is well known that the an-giotensin I-converting enzyme (ACE), a zinc-containing metalloenzyme, plays a key role in the control of blood pressure by the rennin-angiotensin system (Fujita et al. 2000). It acts as an exo-peptidase that cleaves a dipeptide of

Anti-ACE activity from macroalgae of the Azores
11
11
Paiva, L.S., E.M.C. Lima, A.I. Neto & J.A.B. Baptista 2015. Screening for angio-tensin I-converting enzyme (ACE) inhibitory activity of enzymatic hydrolysates obtained from Azorean macroalgae. Arquipelago. Life and Marine Sciences 32: 11-17.
As part of ongoing studies into the isolation of antihypertensive peptides, 12 common macroalgae species from Azores were investigated for their potential angiotensin-converting enzyme (ACE) inhibitory properties. After optimization of enzymatic hydrolysis conditions, the anti-ACE activity was determined by high performance liquid chromatog-raphy (HPLC). The species under study include four Rhodophyta (Gelidium microdon, Os-mundea pinnatifida, Porphyra sp. and Pterocladiella capillacea), four Chlorophyta (Chae-tomorpha linum, Codium adhaerens, Ulva compressa and Ulva rigida) and four Heterokon-tophyta (Cystoseira humilis, Fucus spiralis, Padina pavonica and Sargassum cymosum). The bromelain hydrolysates derived from P. capillacea, Porphyra sp., F. spiralis, U. com-pressa, O. pinnatifida and U. rigida were the most active presenting an ACE inhibitory ac-tivity of 48.73 ± 1.58%, 50.96 ± 1.82%, 56.77 ± 1.80%, 59.80 ± 0.95%, 62.59 ± 1.60% and 65.68 ± 0.72%, respectively. These results suggest that some macroalgae from the littoral Azorean sea are potential sources of powerful ACE inhibitor peptides, with potential impact on public health, particularly on the hypertensive patients. These features make these algae valuable to the regional economy and may also provide an incentive for marine biodiversity conservation and valuation of marine algal resources.
Key words: Azores, marine macroalgae, antihypertensive peptides, ACE inhibition, enzy-matic hydrolysis, pepsin, bromelain, HPLC
Lisete S. Paiva (e-mail: [email protected]), Elisabete M.C. Lima & José A.B. Baptista, CITA-A – Research Center for Agricultural Technology & Department of Technological Sciences and Development, University of Azores, 9501-801 Ponta Delgada, S. Miguel, Azores, Portugal; Lisete S. Paiva & Ana I. Neto, Marine Biology Group, Department of Biology, University of Azores, 9501-801 Ponta Delgada, S. Miguel, Azores, Portugal; Ana I. Neto, cE3c – Centre for Ecology, Evolution and Environmental Changes/ Azorean Biodiversity Group and University of Azores, Department of Biology, 9501-801 Ponta Delgada, S. Miguel, Azores, Portugal.
INTRODUCTION
Hypertension, one of the most common serious chronic health problems, affects about 25% of the world’s adult population (Mittal & Singh 2010). This problem of epidemic proportions carries a high risk factor for arteriosclerosis, stroke, myo-
cardial infarction and end-stage renal disease (Zhang et al. 2006). It is well known that the an-giotensin I-converting enzyme (ACE), a zinc-containing metalloenzyme, plays a key role in the control of blood pressure by the rennin-angiotensin system (Fujita et al. 2000). It acts as an exo-peptidase that cleaves a dipeptide of

Paiva et al.
12
Anti-ACE activity from macroalgae of the Azores
13
ENZYME SCREENING FOR THE ESTIMA-TION OF THE HIGHER NUMBER OF PEPTIDES USING FUCUS SPIRALIS Five grams of Fucus spiralis was suspended in 100 mL of distilled water and then homogenized with an Ultra-turrax T50 at 6.000 rpm for 10 min. Five milligrams of each protease were individual-ly added to 10 mL of the homogenate, adjusting substrate/enzyme ratio to 100:1 (w/w) and the mixtures were incubated for 20 h at an appropri-ated pH and temperature for each enzyme reac-tion, using the conditions suggested by the manu-facturer. After hydrolysis, the samples were heat-ed for 15 min in boiling water in order to inacti-vate the proteases. The precipitates were removed by centrifugation (1100 x g) at 4 °C for 10 min. The supernatants were concentrated and lyophi-lized to obtain powder products that were used for the number of peptides determination using HPLC analysis under the following conditions: reverse-phase Zorbax Eclipse XDB-C18 column (4.6 x 150 mm i.d., 5 m) (Agilent Technologies, USA) using a linear gradient of phase A - 0.1% TFA in water and phase B - 0.1% TFA in acetoni-trile (t = 0 min - 0% B and t = 60 min - 40% B) at a flow rate of 1 mL/min and detection by UV at 220 nm.
PREPARATION OF MACROALGAE ENZY-MATIC HYDROLYSATES FOR PRODUC-TION OF ACE INHIBITORY PEPTIDES Twenty grams of the macroalgae samples were suspended in 300 mL of distilled water, homoge-nized with an Ultra-turrax T50 at 6.000 rpm for 10 min and then hydrolysed by proteolytic active enzymes (pepsin and bromelain) using the same reaction conditions described above. The homog-enates were concentrated until an approximate volume of 150 mL and immediately transferred to a 10,000 Da MWCO dialyzing tube. Each perme-ate was concentrated, lyophilized and kept at -80 °C for the ACE inhibitory activity determination.
DETERMINATION OF ACE INHIBITORY ACTIVITY For the assay, 80 L of the enzymatic hydrolysate solution was pre-incubated at 37 °C for 5 min with 20 L ACE (2 mU/mL) enzyme. The mix-ture was subsequently incubated at the same tem-perature with 200 L of the substrate (5 mM
HHL in 100 mM sodium trizma base containing 300 mM NaCl at pH 8.3) during 60 min. The re-action was terminated by adding 250 L of 1 M HCl. The percentage of inhibition was determined by HPLC analysis. An aliquot of 10 L from the reaction mixture was analyzed on a reverse-phase Ultrasphere C18 column (250 x 4.6 mm i.d., 5 m) (Beckman Coulter, Miami, USA) using an iso-cratic elution of MeOH:ACN:0.1% HCl (25:25:50 v/v/v) at a constant flow-rate of 0.5 mL/min and HA and HHL were detected by UV at 228 nm. The percentage of ACE inhibition by macroalgae hydrolysate sample was calculated by the amount of the hippuric acid (HA) released from the substrate hippuryl-L-histidyl-L-leucine (HHL). The extent of inhibition was calculated as follows:
ACE inhibition (%) = [1– (As – Ab)/C] x 100 where As = HA peak area of the sample, Ab = Peak area of blank (no ACE or with ACE inhibi-tor) and C = HA peak area (HHL plus ACE) without sample. The repeatability of the HA HPLC analysis was evaluated in intraday and interday measurements of the retention time by repeated injections (n = 5). The recovery was calculated based on the dif-ference between the total concentration deter-mined in the spiked samples and the concentra-tion observed in the non-spiked samples. STATISTICAL ANALYSIS All determinations were performed in triplicate and the results expressed as means ± standard deviations (SD). Statistical comparison was per-formed by using one-way analysis of variance (ANOVA). A P-values of less than 0.05 (P < 0.05) were considered to be statistically signifi-cant.
RESULTS AND DISCUSSION
The HPLC methodology used to quantify the HA produced during the ACE-catalyzed reaction completely eliminates the interference from HHL during the quantification of the HA as compared with spectrophotometric method of Cushman &
12
C-terminal of angiotensin I to produce the oc-tapeptide angiotensin II, which is a potent vaso-constrictor (Curtiss et al. 1978). Generally, ACE synthetic inhibitors cause a number of undesirable side effects, like cough, hyperkalemia, renal fail-ure, fetal anomalies, angioedema, dysgeusia and skin rashes (Brown & Vaughan 1998). For this reason, there is a trend towards searching for nat-ural ACE inhibitors as alternatives to synthetic drugs. Recently, much attention has been paid to the potential of marine organism constituents as novel therapeutic drug candidates for the treat-ment of hypertension (e.g. Wijesekara & Kim 2010), and some ACE inhibitory peptides have already been isolated from the hydrolyzates of algae species (e.g. Sato et al. 2002). Enzymatic hydrolysis has become a valuable tool for modi-fying the functionality of proteins resulting in a good source of bioactive peptides that may be beneficial for the organism's functions (Korhonen et al. 1998). The Azores Islands, due to their isolated loca-tion in the middle of the Atlantic Ocean associat-ed with a low pollution levels of their seawater (Neto et al. 2009), are a very promising location for marine natural resources that may produce new ingredients with medicine-like effects in treating or preventing certain diseases. Ongoing research investigates the isolation of natural anti-hypertensive agents to search for novel ACE in-hibitory activity peptides from marine algae (Pai-va et al. 2014a,b). The present study analyses, for the first time, the in vitro ACE inhibitory proper-ties of enzymatic hydrolysates obtained from 12 macroalgae species which are abundant in the Azorean intertidal zone (Neto et al. 2006).
MATERIAL AND METHODS
CHEMICALS AND REAGENTS Acetonitrile (ACN) and Methanol (MeOH), HPLC grade, were purchased from Fluka Chemi-ka (Steinheim, Switzerland). Sodium chloride was obtained from E. Merck (Darmstadt, Germa-ny). Deionized water was obtained from a Milli-pore water purification system (Millipore, Bed-ford, MA, USA). Trizma base, zinc chloride, hip-puric acid (HA), hippuryl-L-histidyl-L-leucine
(HHL), hydrochloric acid (HCl) and various commercial enzymes, such as angiotensin I-converting enzyme (ACE) from porcine kidney, bromelain (B4882), chymotrypsin (C-9381), ficin (F6008), pancreatin (P1750), pepsin (77161), peptidase (P7500), protease from Bacillus licheni-formis (P5380), protease from Streptomyces griseus (P6911) and trypsin (T8003) were pur-chased from Sigma-Aldrich (St. Louis, MO, USA). All other reagents used in this study were reagent grade chemicals. MACROALGAL COLLECTION All macroalgae samples used in this study were collected from the littoral zone of São Miguel Island, Azores Archipelago (37°40 N - 25°31 W), Portugal, and voucher specimens were pre-pared and deposited in the Herbarium AZB - Ruy Telles Palhinha of the Department of Biology at the University of Azores, namely: Chaetomorpha linum (O.F. Müller) Kützing (SMG-12-29); Codium adhaerens C. Agardh (SMG-12-28); Cys-toseira humilis Schousboe ex Kützing (SMG-12-26); Fucus spiralis Linnaeus (SMG-13-04); Ge-lidium microdon K tzing (SMG-13-03); Os-mundea pinnatifida (Hudson) Stackhouse (SMG-13-02); Padina pavonica (Linnaeus) Thivy (SMG-13-13); Porphyra sp. C. Agardh (SMG-14-03); Pterocladiella capillacea (S.G. Gmelin) San-telices & Hommersand (SMG-13-05); Ulva com-pressa Linnaeus (SMG-13-15); Ulva rigida C. Agardh (SMG-13-12) and Sargassum cymosum C. Agardh (SMG-12-27). MACROALGAE SAMPLE PREPARATION Within 24 h of collection the macroalgae samples were first washed in seawater to remove encrust-ing material and epiphytes, carefully rinsed with distilled water to remove salts and then air-dried and stored in an air-tight container in a freezer (-80 °C) until further analysis. Prior to the analyti-cal procedures, the samples were defrosted and dried at 45 °C during 48 h (avoiding overheating that could lead to oxidation). Dried triplicate samples were grounded into a fine powder, re-dried at 40 °C and stored in desiccators at a re-frigerated temperature.

Anti-ACE activity from macroalgae of the Azores
13
Anti-ACE activity from macroalgae of the Azores
13
ENZYME SCREENING FOR THE ESTIMA-TION OF THE HIGHER NUMBER OF PEPTIDES USING FUCUS SPIRALIS Five grams of Fucus spiralis was suspended in 100 mL of distilled water and then homogenized with an Ultra-turrax T50 at 6.000 rpm for 10 min. Five milligrams of each protease were individual-ly added to 10 mL of the homogenate, adjusting substrate/enzyme ratio to 100:1 (w/w) and the mixtures were incubated for 20 h at an appropri-ated pH and temperature for each enzyme reac-tion, using the conditions suggested by the manu-facturer. After hydrolysis, the samples were heat-ed for 15 min in boiling water in order to inacti-vate the proteases. The precipitates were removed by centrifugation (1100 x g) at 4 °C for 10 min. The supernatants were concentrated and lyophi-lized to obtain powder products that were used for the number of peptides determination using HPLC analysis under the following conditions: reverse-phase Zorbax Eclipse XDB-C18 column (4.6 x 150 mm i.d., 5 m) (Agilent Technologies, USA) using a linear gradient of phase A - 0.1% TFA in water and phase B - 0.1% TFA in acetoni-trile (t = 0 min - 0% B and t = 60 min - 40% B) at a flow rate of 1 mL/min and detection by UV at 220 nm.
PREPARATION OF MACROALGAE ENZY-MATIC HYDROLYSATES FOR PRODUC-TION OF ACE INHIBITORY PEPTIDES Twenty grams of the macroalgae samples were suspended in 300 mL of distilled water, homoge-nized with an Ultra-turrax T50 at 6.000 rpm for 10 min and then hydrolysed by proteolytic active enzymes (pepsin and bromelain) using the same reaction conditions described above. The homog-enates were concentrated until an approximate volume of 150 mL and immediately transferred to a 10,000 Da MWCO dialyzing tube. Each perme-ate was concentrated, lyophilized and kept at -80 °C for the ACE inhibitory activity determination.
DETERMINATION OF ACE INHIBITORY ACTIVITY For the assay, 80 L of the enzymatic hydrolysate solution was pre-incubated at 37 °C for 5 min with 20 L ACE (2 mU/mL) enzyme. The mix-ture was subsequently incubated at the same tem-perature with 200 L of the substrate (5 mM
HHL in 100 mM sodium trizma base containing 300 mM NaCl at pH 8.3) during 60 min. The re-action was terminated by adding 250 L of 1 M HCl. The percentage of inhibition was determined by HPLC analysis. An aliquot of 10 L from the reaction mixture was analyzed on a reverse-phase Ultrasphere C18 column (250 x 4.6 mm i.d., 5 m) (Beckman Coulter, Miami, USA) using an iso-cratic elution of MeOH:ACN:0.1% HCl (25:25:50 v/v/v) at a constant flow-rate of 0.5 mL/min and HA and HHL were detected by UV at 228 nm. The percentage of ACE inhibition by macroalgae hydrolysate sample was calculated by the amount of the hippuric acid (HA) released from the substrate hippuryl-L-histidyl-L-leucine (HHL). The extent of inhibition was calculated as follows:
ACE inhibition (%) = [1– (As – Ab)/C] x 100 where As = HA peak area of the sample, Ab = Peak area of blank (no ACE or with ACE inhibi-tor) and C = HA peak area (HHL plus ACE) without sample. The repeatability of the HA HPLC analysis was evaluated in intraday and interday measurements of the retention time by repeated injections (n = 5). The recovery was calculated based on the dif-ference between the total concentration deter-mined in the spiked samples and the concentra-tion observed in the non-spiked samples. STATISTICAL ANALYSIS All determinations were performed in triplicate and the results expressed as means ± standard deviations (SD). Statistical comparison was per-formed by using one-way analysis of variance (ANOVA). A P-values of less than 0.05 (P < 0.05) were considered to be statistically signifi-cant.
RESULTS AND DISCUSSION
The HPLC methodology used to quantify the HA produced during the ACE-catalyzed reaction completely eliminates the interference from HHL during the quantification of the HA as compared with spectrophotometric method of Cushman &

Paiva et al.
14
Anti-ACE activity from macroalgae of the Azores
15
Fig. 2. HPLC estimation for the proteolytic activity (number of peptides) derived from different commercial enzymes using the Azorean macroalgae Fucus spiralis. Chromatographic conditions as referred in methods; * Streptomyces griseus, ** Bacillus licheniformis.
Fig. 3. Angiotensin-converting enzyme (ACE) inhibitory activity (%) of six selected Azorean macroal-gae enzymatic hydrolysates. Algal protein not hydrolyzed was used for control. Data are mean ± SD of triplicate experiments.
ACE activity (56.77 ± 1.80%). Remarkable ACE inhibitory activities of enzymatic hydrolysates from some macroalgae have also been reported by other authors, especially from brown and red algal species (e.g. Sato et al. 2002; Athukorala & Jeon 2005; Qu et al. 2010). From these studies,
Qu et al. (2010) reported for Porphyra yezoensis 55% of ACE inhibition that is similar to the value obtained for the studied Porphyra sp. (ca 51%). Research by Cha et al. (2006a,b), but using crude extracts from Korean Porphyra tenera, Ptero-cladiella capillacea, Ulva conglobata and U.
14
Fig. 1. Representative HPLC chromatograms of the angiotensin-converting enzyme inhibition assay. a) More active enzymatic hydrolysate; b) Less active enzymatic hydrolysate; c) Control sample (without inhibitors). Chromatographic conditions as referred in methods. Cheung (1971). Complete baseline separation of HA and HHL was achieved by HPLC in isocratic elution conditions in less than 10 min under the analytical conditions described in methods (see Figure 1) showing a relative standard deviation (RSD) of 1.51% in multiple injections that repre-sents a good repeatability. Another advantage of the HPLC method is the reduction of the time-consuming process of HA extraction into ethyl acetate that is required by the standard Cushman & Cheung (1971) method and also the standard spectrophotometric assay requires at least 250 to 500 L of reaction mixture, compared to the 10
L used in HPLC assay. Therefore, the HPLC provides a simple, rapid and accurate method for the assay of ACE-catalyzed reactions. The estimation of the proteolytic activity (number of peptides) derived from nine commer-cial enzymes using F. spiralis revealed that pep-
sin and bromelain were the enzymes with higher proteolytic activity (Figure 2). From the selected macroalgae species the bro-melain hydrolysates derived from P. capillacea, Porphyra sp., F. spiralis, U. compressa, O. pin-natifida and U. rigida presented an ACE inhibito-ry activity similar and greater than 50% (ranging from 48.73% to 65.68%) (Figure 3). The U. rigida exhibited the strongest anti-ACE activity (65.68 ± 0.72%) and the other Chloro-phyta, U. compressa, presented the third highest anti-ACE activity (59.80 ± 0.95%). The Rhodo-phyta O. pinnatifida presented the second highest anti-ACE activity (62.59 ± 1.60%) and the other red algal species showed lower activity (48.73 ± 1.58% and 50.96 ± 1.82% for P. capillacea and Porphyra sp., respectively). The Heterokonto-phyta F. spiralis, the only active of the selected brown algae, presented the fourth highest anti-
Abs
orba
nce
at 2
28 n
m
Time (min.)

Anti-ACE activity from macroalgae of the Azores
15
Anti-ACE activity from macroalgae of the Azores
15
Fig. 2. HPLC estimation for the proteolytic activity (number of peptides) derived from different commercial enzymes using the Azorean macroalgae Fucus spiralis. Chromatographic conditions as referred in methods; * Streptomyces griseus, ** Bacillus licheniformis.
Fig. 3. Angiotensin-converting enzyme (ACE) inhibitory activity (%) of six selected Azorean macroal-gae enzymatic hydrolysates. Algal protein not hydrolyzed was used for control. Data are mean ± SD of triplicate experiments.
ACE activity (56.77 ± 1.80%). Remarkable ACE inhibitory activities of enzymatic hydrolysates from some macroalgae have also been reported by other authors, especially from brown and red algal species (e.g. Sato et al. 2002; Athukorala & Jeon 2005; Qu et al. 2010). From these studies,
Qu et al. (2010) reported for Porphyra yezoensis 55% of ACE inhibition that is similar to the value obtained for the studied Porphyra sp. (ca 51%). Research by Cha et al. (2006a,b), but using crude extracts from Korean Porphyra tenera, Ptero-cladiella capillacea, Ulva conglobata and U.

Paiva et al.
16
Anti-ACE activity from macroalgae of the Azores
17
Sato, M., T. Hosokawa, T. Yamaguchi, T. Nakano, K. Muramoto, T. Kahara, K. Funayama, A. Kobayashi & T. Nakano 2002. Angiotensin I-converting enzyme inhibitory peptides derived from Wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. Journal of Agricultural and Food Chemistry 50: 6245-6252.
Wijesekara, I. & S.-K. Kim 2010. Angiotensin-I-converting enzyme (ACE) Inhibitors from marine
resources: prospects in the pharmaceutical Industry. Marine Drugs 8: 1080-1093.
Zhang, Y., E.T. Lee, R.B. Devereux, J. Yeh, L.G. Best, R.R. Fabsitz & B.V. Howard 2006. Prehypertension, diabetes, and cardiovascular disease risk in a population-based sample: the Strong Heart Study. Hypertension 47: 410-4.
Received 23 May 2015. Accepted 12 Aug 2015. Published online 22 Sept 2015.
16
pertusa, also reported ACE inhibition but much lower values than the ones obtained for the same genus in the present study. These results empha-size the importance of the enzymatic hydrolysis in the process of ACE inhibitory activity. In conclusion, this study shows a high activity in six Azorean macroalgae suggesting they can be excellent sources of ACE inhibitors, with poten-tial pharmaceutical and nutraceutical applications. Therefore, purification and characterization of their ACE inhibitory peptides from the more potent hydrolysates is already in progress.
ACKNOWLEDGMENTS
This study was financially supported by funds from CIRN (Centro de Investigação de Recursos Naturais, University of the Azores) and by the European Regional Development Fund (ERDF) through the COMPETE - Operational Competi-tiveness Programme and national funds through FCT – Foundation for Science and Technology, under the project “PEst-C/MAR/LA0015/2013, by the Strategic Funding UID/Multi/04423/2013 through national funds provided by FCT – Foun-dation for Science and Technology and European Regional Development Fund (ERDF), in the framework of the programme PT2020 and by cE3c funding (Ref: UID/BIA/00329/2013). Lisete Paiva thanks FRC (Fundo Regional da Ciência) for Doctoral Research Scholarships ref. M3.1.2/F/014/2011, which is co-financed by the European Social Fund through the action-type 4.2.2 from Pro-Emprego.
REFERENCES
Athukorala, Y. & Y.J. Jeon 2005. Screening for angiotensin-1-converting enzyme inhibitory activity of Ecklonia cava. Journal of Food Science and Nutrition 10: 134-139.
Brown, N.J. & D.E. Vaughan 1998. Angiotensin-converting enzyme inhibitors. Circulation 97: 1411-1420.
Cha, S.H., G.N. Ahn, S.J. Heo, K.N. Kim, K.W. Lee, C.B. Song, S.K. Cho & Y.J. Jeon 2006a. Screening of extracts from marine green and brown algae in Jeju for potential marine angiotensin-I converting enzyme (ACE) inhibitory activity. Journal of the
Korean Society of Food Science and Nutrition 35: 307-314.
Cha, S.H., K.W. Lee & Y.J. Jeon 2006b. Screening of extracts from red algae in Jeju for potentials marine angiotensin-I converting enzyme (ACE) inhibitory activity. Algae 21: 343-348.
Curtiss, C., J.N. Chon, T. Vrobel & J.A Francious 1978. Role of the rennin-angiotensin system in the systemic vasoconstriction of chronic congestive heart failure. Circulation 58: 763-70.
Cushman, D.W. & H.S. Cheung 1971. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochemical Pharmacology 20: 1637-1648.
Fujita, H., K. Yokoyama & M. Yoshikawa 2000. Classification and antihypertensive activity of angiotensin I-converting enzyme inhibitory peptides derived from food proteins. Journal of Food Science 65: 564-569.
Korhonen, H., A. Pihlanto-Leppälä, P. Rantamäki & T. Tupasela 1998. Impact of processing on bioactive proteins and peptides. Trends in Food Science &Technology 9: 307-319.
Mittal, B.V. & A.K. Singh 2010. Hypertension in the developing world: challenges and opportunities. American Journal of Kidney Diseases 55: 590-8.
Neto, A.I., I. Tittley & P. M. Raposeiro 2006. Flora Marinha do Litoral dos Açores. Rocky shore marine flora of the Azores. Secretaria Regional do Ambiente e do Mar, Horta, Portugal. 157 pp.
Neto, A.I., V. Brotas, J.M.N. Azevedo, R.F. Patarra, N.M.V. Álvaro, C. Gameiro, A.C.L. Prestes & E.M. Nogueira 2009. Qualidade de águas costeiras do Grupo Oriental do arquipélago dos Açores e proposta de monitorização. [Quality of coastal waters of the Azores Eastern Group and monitoring proposal]. Departamento de Biologia, Universidade dos Açores. iii+70 pp. [In Portuguese]
Paiva, L., E. Lima, A.I. Neto & J. Baptista 2014a. Antihypertensive effect of Azorean macroalgae extracts by the inhibitory activity of the angiotensin I-converting enzyme (ACE) [abstract N016]. EFMC, International Symposium on Medicinal Chemistry, Lisbon, Portugal, 7-11 September 2014. Chemmedchem: 220-221.
Paiva, L., E. Lima, A.I. Neto & J. Baptista 2014b. Investigation of Azorean macroalgae for Angiotensin I-converting enzyme (ACE) inhibitory peptides. Extraction, purification and antihypertensive activity evaluation. Planta Medica 80: 1391-1392.
Qu, W., H. Ma, Z. Pan, L. Luo, Z. Wang & R. He 2010. Preparation and antihypertensive activity of peptides from Porphyra yezoensis. Food Chemistry 123: 14-20.

Anti-ACE activity from macroalgae of the Azores
17
Anti-ACE activity from macroalgae of the Azores
17
Sato, M., T. Hosokawa, T. Yamaguchi, T. Nakano, K. Muramoto, T. Kahara, K. Funayama, A. Kobayashi & T. Nakano 2002. Angiotensin I-converting enzyme inhibitory peptides derived from Wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. Journal of Agricultural and Food Chemistry 50: 6245-6252.
Wijesekara, I. & S.-K. Kim 2010. Angiotensin-I-converting enzyme (ACE) Inhibitors from marine
resources: prospects in the pharmaceutical Industry. Marine Drugs 8: 1080-1093.
Zhang, Y., E.T. Lee, R.B. Devereux, J. Yeh, L.G. Best, R.R. Fabsitz & B.V. Howard 2006. Prehypertension, diabetes, and cardiovascular disease risk in a population-based sample: the Strong Heart Study. Hypertension 47: 410-4.
Received 23 May 2015. Accepted 12 Aug 2015. Published online 22 Sept 2015.

Arquipelago - Life and Marine Sciences ISSN: 0873-4704
19
Arthropods and other Biota associated with the Azorean Trees and Shrubs: Juniperus brevifolia
RUI NUNES, R. GABRIEL, R.B. ELIAS, F. RIGAL, A.O. SOARES, P. CARDOSO & P.A.V. BORGES
Nunes, R., R. Gabriel, R.B. Elias, F. Rigal, A.O. Soares, P. Cardoso & P.A.V. Borges 2015. Arthropods and other Biota associated with the Azorean Trees and Shrubs: Juniperus brevifolia. Arquipelago. Life and Marine Sciences 32: 19-48. Appendix I-IV.
This work aims to characterize the arthropods and other biota (lichens, bryophytes, vascular plants and birds) associated with the Azorean endemic tree, Juniperus brevifolia. This is the first of a series of publications that will (i) provide a comprehensive list of all the biota associated with the main Azorean endemic trees and shrubs, (ii) describe in detail the diver-sity, abundance and spatial patterns of canopy arthropods, and (iii) whenever possible, to extend biodiversity assessments to communities of bryophytes, lichens, vascular plants and vertebrates. We use standardized sampled data from BALA project for canopy arthropods and for the remaining taxa we surveyed literature data and the Herbarium of University of Azores. Juniperus brevifolia occurs in a wide range of elevation belts in Azores and accommodates a remarkable large number of taxa: besides canopy arthropods (161 species) it is also an important substrate to other vascular species (six species), bryophytes (105 spe-cies), lichens (106 species) and also birds (four species). In addition, the species richness and particularly the abundance of endemics are dominant, and the number of conservation concern species for bryophytes is noteworthy (30 out of 70). The complexity of the tree and the high diversity of micro-habitats created the conditions for epiphytic species to easily colonize all parts of the tree, from the bottom, trunk and branches. The communities associ-ated with the Azorean cedar are consequently of a high conservation value and should be further investigated in their ecological dynamics.
Key words: Arthropods, Azorean cedar, bryophytes, host plant, species composition
R. Nunes, R. Gabriel, R.B. Elias, F. Rigal1, P. Cardoso2 & P.A.V Borges (e-mail: [email protected]), CE3C – Centre for Ecology, Evolution and Environmental Changes / Azorean Biodiversity Group and Universidade dos Açores – Department of Agrarian Sci-ences, Rua Capitão João d’Ávila, 9700-042 Angra do Heroísmo, Azores, Portugal; 1 Envi-ronment and Microbiology Team, MELODY group, Université de Pau et des Pays de l’Adour, IPREM UMR CNRS 5254, BP 1155, 64013 Pau Cedex, France; 2 Finnish Museum of Natural History, University of Helsinki, P.O.Box 17 (Pohjoinen Rautatiekatu 13), 00014 Helsinki, Finland; A.O. Soares3 CE3C – Centre for Ecology, Evolution and Environmental Changes / Azorean Biodiversity Group and University of the Azores – Department of Biol-ogy, Rua Madre de Deus, 9501-801 Ponta Delgada, Azores, Portugal.
INTRODUCTION
Azores are known by their low number of indige-nous species (endemic and native non-endemic) of most taxonomic groups, in comparison with the other Macaronesian archipelagos. Moreover, the Azores are probably the archipelago of Maca-
ronesia where the damages caused by historical anthropogenic ecosystem changes are the most apparent (Borges & Hortal 2009; Cardoso et al. 2010; Triantis et al. 2010a,b; Connor et al. 2012, 2013; Rando et al. 2013). Such changes are thought to have caused already the extinction of numerous endemic species, particularly in the

Biota associated with Juniperus brevifolia
19
Arquipelago - Life and Marine Sciences ISSN: 0873-4704
19
Arthropods and other Biota associated with the Azorean Trees and Shrubs: Juniperus brevifolia
RUI NUNES, R. GABRIEL, R.B. ELIAS, F. RIGAL, A.O. SOARES, P. CARDOSO & P.A.V. BORGES
Nunes, R., R. Gabriel, R.B. Elias, F. Rigal, A.O. Soares, P. Cardoso & P.A.V. Borges 2015. Arthropods and other Biota associated with the Azorean Trees and Shrubs: Juniperus brevifolia. Arquipelago. Life and Marine Sciences 32: 19-48. Appendix I-IV.
This work aims to characterize the arthropods and other biota (lichens, bryophytes, vascular plants and birds) associated with the Azorean endemic tree, Juniperus brevifolia. This is the first of a series of publications that will (i) provide a comprehensive list of all the biota associated with the main Azorean endemic trees and shrubs, (ii) describe in detail the diver-sity, abundance and spatial patterns of canopy arthropods, and (iii) whenever possible, to extend biodiversity assessments to communities of bryophytes, lichens, vascular plants and vertebrates. We use standardized sampled data from BALA project for canopy arthropods and for the remaining taxa we surveyed literature data and the Herbarium of University of Azores. Juniperus brevifolia occurs in a wide range of elevation belts in Azores and accommodates a remarkable large number of taxa: besides canopy arthropods (161 species) it is also an important substrate to other vascular species (six species), bryophytes (105 spe-cies), lichens (106 species) and also birds (four species). In addition, the species richness and particularly the abundance of endemics are dominant, and the number of conservation concern species for bryophytes is noteworthy (30 out of 70). The complexity of the tree and the high diversity of micro-habitats created the conditions for epiphytic species to easily colonize all parts of the tree, from the bottom, trunk and branches. The communities associ-ated with the Azorean cedar are consequently of a high conservation value and should be further investigated in their ecological dynamics.
Key words: Arthropods, Azorean cedar, bryophytes, host plant, species composition
R. Nunes, R. Gabriel, R.B. Elias, F. Rigal1, P. Cardoso2 & P.A.V Borges (e-mail: [email protected]), CE3C – Centre for Ecology, Evolution and Environmental Changes / Azorean Biodiversity Group and Universidade dos Açores – Department of Agrarian Sci-ences, Rua Capitão João d’Ávila, 9700-042 Angra do Heroísmo, Azores, Portugal; 1 Envi-ronment and Microbiology Team, MELODY group, Université de Pau et des Pays de l’Adour, IPREM UMR CNRS 5254, BP 1155, 64013 Pau Cedex, France; 2 Finnish Museum of Natural History, University of Helsinki, P.O.Box 17 (Pohjoinen Rautatiekatu 13), 00014 Helsinki, Finland; A.O. Soares3 CE3C – Centre for Ecology, Evolution and Environmental Changes / Azorean Biodiversity Group and University of the Azores – Department of Biol-ogy, Rua Madre de Deus, 9501-801 Ponta Delgada, Azores, Portugal.
INTRODUCTION
Azores are known by their low number of indige-nous species (endemic and native non-endemic) of most taxonomic groups, in comparison with the other Macaronesian archipelagos. Moreover, the Azores are probably the archipelago of Maca-
ronesia where the damages caused by historical anthropogenic ecosystem changes are the most apparent (Borges & Hortal 2009; Cardoso et al. 2010; Triantis et al. 2010a,b; Connor et al. 2012, 2013; Rando et al. 2013). Such changes are thought to have caused already the extinction of numerous endemic species, particularly in the

Nunes et al.
20
Biota associated with Juniperus brevifolia
21
land Portugal and it seems possible that seeds of J. navicularis-like plants or their ancestor werebrought to the Azores by birds from the Iberian Peninsula (Adams 2014). A study by Rumeu et al.(2011) suggested that a single introduction event
likely occurred from Europe and that genetic dif-ferentiation of J. brevifolia post-dated the emer-gence of the oldest island (Santa Maria, 8.12 Ma, [França et al. 2003]).
Figure 1. Juniperus brevifolia (Seub.) Antoine with details of a female branch, with seed cones, and a male branch with male cones. : Fernando Correia (www.efecorreia-artstudio.com). With permission of Azorina – S.A.
Presently, two subspecies are recognized (Elias & Dias 2014): (1) J. brevifolia subsp. maritima is an erect shrub or small tree, with (inverted) pyrami-dal crown; leaves: 5.6–7.6 × 1.4–1.8 mm; seed cones: 7.9–9.5 mm, ripening in the first year; seeds: 1–3 per seed cone, 4.9–6.3 × 3.5–5.1 mm, germinating from February to May; pollination: January–March; distribution: Flores, Terceira, Pico, São Jorge and São Miguel (?), below 100 m
a.s.l.; and (2) J. brevifolia subsp. brevifolia is a tree or shrub; leaves: 4.5–6.7 × 1.1–1.5 mm; seed cones: 6.9–8.7 mm, ripening in the second year; seeds: 2-3 per seed cone, 3.9–5.3 × 2.1–3.7 mm, germinating from March to June; pollination: March–June; distribution: all islands, except Gra-ciosa, usually between 300 m and 1500 m a.s.l. Two varieties have been recognized from the later subspecies (Elias & Dias 2014 ): i) J. brevifolia
Biota associated with Juniperus brevifolia
21
land Portugal and it seems possible that seeds of J. navicularis-like plants or their ancestor werebrought to the Azores by birds from the Iberian Peninsula (Adams 2014). A study by Rumeu et al.(2011) suggested that a single introduction event
likely occurred from Europe and that genetic dif-ferentiation of J. brevifolia post-dated the emer-gence of the oldest island (Santa Maria, 8.12 Ma, [França et al. 2003]).
Figure 1. Juniperus brevifolia (Seub.) Antoine with details of a female branch, with seed cones, and a male branch with male cones. : Fernando Correia (www.efecorreia-artstudio.com). With permission of Azorina – S.A.
Presently, two subspecies are recognized (Elias & Dias 2014): (1) J. brevifolia subsp. maritima is an erect shrub or small tree, with (inverted) pyrami-dal crown; leaves: 5.6–7.6 × 1.4–1.8 mm; seed cones: 7.9–9.5 mm, ripening in the first year; seeds: 1–3 per seed cone, 4.9–6.3 × 3.5–5.1 mm, germinating from February to May; pollination: January–March; distribution: Flores, Terceira, Pico, São Jorge and São Miguel (?), below 100 m
a.s.l.; and (2) J. brevifolia subsp. brevifolia is a tree or shrub; leaves: 4.5–6.7 × 1.1–1.5 mm; seed cones: 6.9–8.7 mm, ripening in the second year; seeds: 2-3 per seed cone, 3.9–5.3 × 2.1–3.7 mm, germinating from March to June; pollination: March–June; distribution: all islands, except Gra-ciosa, usually between 300 m and 1500 m a.s.l. Two varieties have been recognized from the later subspecies (Elias & Dias 2014 ): i) J. brevifolia
Nunes et al.
20
most disturbed islands, where few and minute native forest patches remain (Cardoso et al., 2010; Rando et al. 2013; Terzopoulou et al. 2015). As a consequence, the knowledge of the current distribution and abundance of terrestrial Azorean biotas is a critical step to set conserva-tion programs on the native forest ecosystem (see e.g. Gaspar et al. 2011). Due to the work of sev-eral research groups from the University of the Azores in the last twenty years, many archipelag-ic studies on both indigenous and exotic species have been carried out, such as exhaustive species inventories (Borges et al. 2005b; Borges et al. 2010a), comprehensive biogeographical studies (Whittaker et al. 2008, 2014; Borges & Hortal 2009; Cardoso et al. 2010; Santos et al. 2010; Triantis et al. 2010a,b; Cameron et al. 2013; Ar-anda et al. 2014; Carvalho & Cardoso 2014; Car-valho et al. 2015) and phylogenetic and evolu-tionary studies (Ferreira et al. 2011; Schaefer et al. 2011; Amorim et al. 2012; Rodrigues et al. 2013). In addition, there are several ecological studies at smaller spatial scales focusing on the communities of particular organisms, namely bryophytes (e.g. Gabriel & Bates 2005; Aranda et al. 2015), vascular plants (e.g. Elias & Dias 2009; Rumeu et al. 2011; Marcelino et al. 2013) and arthropods (e.g. Borges et al. 2006; Meijer et al. 2011; Florencio et al. 2013; Raposeiro et al. 2013). However, there are very few studies so far dealing in detail with the biotic communities as-sociated with each of the indigenous host tree species that structure the extant Azorean forests. Among them we emphasise the works on arthro-pod canopy community biodiversity and structure (Markin et al. 1995; Ribeiro et al. 2005; Gaspar et al. 2008; Borges et al. 2008; Ribeiro & Borges 2010) and those dealing with the effects of her-bivory on a specific indigenous host tree species (Vieira et al. 1993; Silva et al. 1995; Ribeiro et al. 2003). Despite these works, there is still insuffi-cient knowledge regarding the biodiversity as-sessment and the spatial structure of the biotas associated with Azorean host trees, which hinders biodiversity conservation and sustainable re-source management (Summervile et al. 2003; Cardoso et al. 2011).
We propose to do a series of publications review-ing the existing knowledge on the biotic commu-nities associated with structurally important na-tive trees and shrubs of the Azores archipelago, namely Juniperus brevifolia (Seub.) Antoine, Ilex perado Aiton subsp. azorica (Loes.) Tutin; Laurus azorica (Seub.) Franco, Erica azorica Hochst. ex Seub., Vaccinium cylindraceum Sm. and Myrsine africana L.. Our main objectives are: i) to provide a comprehensive list of all the biota associated with the main Azorean endemic trees and shrubs, ii) to describe in detail the diversity, abundance and spatial patterns of canopy arthro-pods, since there is already a bulk of standardized data collected with the BALA project – field-work: 1992-2002 and 2003-2004 (e.g. Ribeiro et al. 2005; Gaspar et al. 2008), and iii) whenever possible, to extend our biodiversity assessment to communities of bryophytes, lichens, vascular plants and vertebrates. Given the current rates of biodiversity loss due to global changes (e.g. inva-sive species, habitat erosion and fragmentation, climatic changes), with all their impacts on the ecosystems and associated communities (Butchart et al. 2010, Hooper et al. 2012) we feel that this will be a valuable asset for future researchers and conservation practitioners, offering a baseline that may be used to compare and evaluate future changes in the native Azorean ecosystems and associated species composition. We will start this series of works with the Azorean endemic gymnosperm, Juniperus brevi-folia (Seub.) Antoine, since this is the most com-mon and widespread endemic tree in the extant Azorean native forests (Fig. 1). THE TARGET SPECIES: JUNIPERUS BREVIFOLIA (SEUB.) ANTOINE Juniperus brevifolia, the Azorean cedar or Short leafed juniper, is a dioecious shrub or tree, with a trunk bark brown-purple, exfoliating in strips. Leaves are acicular, closely set, with two broad, white stomatiferous bands on the adaxial surface. Seed cones are subglobose, green and pruinose while young, dark copper-brown when mature. Seeds are free, ovoid and triquetrous (Adams 2014). Sequences from nuclear and chloroplast DNA (trnC-trnD spacer) have placed J. brevifolia in a clade with Juniperus navicularis Gandoger, that is endemic to western coastal areas of main-

Biota associated with Juniperus brevifolia
21
Biota associated with Juniperus brevifolia
21
land Portugal and it seems possible that seeds of J. navicularis-like plants or their ancestor werebrought to the Azores by birds from the Iberian Peninsula (Adams 2014). A study by Rumeu et al.(2011) suggested that a single introduction event
likely occurred from Europe and that genetic dif-ferentiation of J. brevifolia post-dated the emer-gence of the oldest island (Santa Maria, 8.12 Ma, [França et al. 2003]).
Figure 1. Juniperus brevifolia (Seub.) Antoine with details of a female branch, with seed cones, and a male branch with male cones. : Fernando Correia (www.efecorreia-artstudio.com). With permission of Azorina – S.A.
Presently, two subspecies are recognized (Elias & Dias 2014): (1) J. brevifolia subsp. maritima is an erect shrub or small tree, with (inverted) pyrami-dal crown; leaves: 5.6–7.6 × 1.4–1.8 mm; seed cones: 7.9–9.5 mm, ripening in the first year; seeds: 1–3 per seed cone, 4.9–6.3 × 3.5–5.1 mm, germinating from February to May; pollination: January–March; distribution: Flores, Terceira, Pico, São Jorge and São Miguel (?), below 100 m
a.s.l.; and (2) J. brevifolia subsp. brevifolia is a tree or shrub; leaves: 4.5–6.7 × 1.1–1.5 mm; seed cones: 6.9–8.7 mm, ripening in the second year; seeds: 2-3 per seed cone, 3.9–5.3 × 2.1–3.7 mm, germinating from March to June; pollination: March–June; distribution: all islands, except Gra-ciosa, usually between 300 m and 1500 m a.s.l. Two varieties have been recognized from the later subspecies (Elias & Dias 2014 ): i) J. brevifolia
Biota associated with Juniperus brevifolia
21
land Portugal and it seems possible that seeds of J. navicularis-like plants or their ancestor werebrought to the Azores by birds from the Iberian Peninsula (Adams 2014). A study by Rumeu et al.(2011) suggested that a single introduction event
likely occurred from Europe and that genetic dif-ferentiation of J. brevifolia post-dated the emer-gence of the oldest island (Santa Maria, 8.12 Ma, [França et al. 2003]).
Figure 1. Juniperus brevifolia (Seub.) Antoine with details of a female branch, with seed cones, and a male branch with male cones. : Fernando Correia (www.efecorreia-artstudio.com). With permission of Azorina – S.A.
Presently, two subspecies are recognized (Elias & Dias 2014): (1) J. brevifolia subsp. maritima is an erect shrub or small tree, with (inverted) pyrami-dal crown; leaves: 5.6–7.6 × 1.4–1.8 mm; seed cones: 7.9–9.5 mm, ripening in the first year; seeds: 1–3 per seed cone, 4.9–6.3 × 3.5–5.1 mm, germinating from February to May; pollination: January–March; distribution: Flores, Terceira, Pico, São Jorge and São Miguel (?), below 100 m
a.s.l.; and (2) J. brevifolia subsp. brevifolia is a tree or shrub; leaves: 4.5–6.7 × 1.1–1.5 mm; seed cones: 6.9–8.7 mm, ripening in the second year; seeds: 2-3 per seed cone, 3.9–5.3 × 2.1–3.7 mm, germinating from March to June; pollination: March–June; distribution: all islands, except Gra-ciosa, usually between 300 m and 1500 m a.s.l. Two varieties have been recognized from the later subspecies (Elias & Dias 2014 ): i) J. brevifolia

Nunes et al.
22
Biota associated with Juniperus brevifolia
23
Figure 2. Map of the Azores (distances between the island groups and between São Miguel and Santa Maria are not in scale). Sampled reserves: 1 – Morro Alto e Pico da Sé; 2 – Caldeira Funda e Rasa; 3 – Cabeço do Fogo; 4 – Caldeira; 5 – Lagoa do Caiado, 6 – Caveiro, 7 – Mistério da Prainha; 8 – Pico Pinheiro; 9 – Topo; 10 – Serra de Santa Bárbara; 11 – Biscoito da Ferraria; 12 – Terra Brava; 13 – Caldeira de Guilherme Moniz; 14 – Pico Galhar-do; 15 - Graminhais; 16 – Atalhada; 17 – Pico da Vara.
cloud forests are also characterised by a dense cover of mosses and other epiphytes (Gabriel & Bates 2005). In the Azores these forests are shaped by the shallow soil layer, fractured and craggy terrain (Ribeiro et al. 2005) and wind speed. From the work of Frutuoso (1978, 1981, 1987), compiled from documents, oral tradition and from its own travels to the islands, it is clear that for-ests dominated the Azorean landscape at the time of discovery and settlement of the archipelago in the 15th Century. Based on the classical proposal of Dansereau (1970) the vegetation of the Azores could be divided in six altitudinal belts, character-ized by their dominant species: Morella faya (0-300 m); Laurus azorica (300-650 m); Juniperus brevifolia (450-1100 m); Erica azorica (850-1600 m); Calluna vulgaris (1600-2200 m) and (6) li-chens (2200-2350 m). Palynological studies by Connor et al. (2012, 2013) provided us with some very important information about the recent past and historical native vegetation of two Azorean islands, Pico and Flores. These studies added
more evidence that primitive Azorean vegetation was dominated by trees and that human impact had a strong influence on the native vegetation. In fact, as a result of less than 600 years of human settlement more than 95% of the native forest was destroyed by human action (Triantis et al. 2010b; Fernández-Palacios et al. 2011; Silveira, 2013), with most coastal, lowland and submontane for-ests (dominated by Morella faya, Picconia azori-ca, Laurus azorica and Prunus azorica) being replaced, giving way to urban and agricultural areas. The remaining pristine forest continues under threat due to habitat reduction, fragmentation and invasive plants, with clear impacts on the flora and fauna (Borges et al. 2000; Cardoso et al. 2010; Triantis et al. 2010b; Terzopoulou et al. 2015), and is now restricted to a few dispersed high-altitude fragments, with areas between 4 and 1300 ha (Borges et al. 2005a, 2006; Gaspar et al. 2008), totalling some 6000 ha (Gaspar et al. 2008; Gabriel et al. 2011).
Nunes et al.
22
subsp. brevifolia var. brevifolia (the dominant taxa) a small to medium-sized tree, found mostly between 300 and 1000 m a.s.l., in all Islands (ex-cept Graciosa); and ii) J. brevifolia subsp. brevi-folia var. montanum a small prostrate shrub, usu-ally found between 850 and 1500 m a.s.l. in the Islands of Flores, Terceira, Pico, São Jorge, Faial and São Miguel. Overall, this species has wide ecological amplitude and it can be found from the coast to 1500 m a.s.l. (in Pico Mountain). There-fore it has the potential to occupy all islands, at all altitude ranges except Pico (which has a max-imum altitude of 2350 m a.s.l.). Presently, according to Elias (2007), J. brevifo-lia reaches higher values of abundance and fre-quency between 500 and 800 m a.s.l.. Between 800 and 1000 m a.s.l. frequency decreases, and between 1000 and 1500 m a.s.l. abundance also decreases. Low winter temperatures, very high rainfall, strong and frequent winds explain the decreasing abundance. Above 1500 m it is rarely found, possibly due to the frequent snow cover during the winter in Pico Mountain. At lower altitudes, namely between 100 and 500 m (espe-cially up to 300 m) it becomes rarer, not only due to human induced habitat changes but also, in the remaining natural or semi-natural areas due to the competition with tree species with higher stature (e.g. Morella faya, Laurus azorica and Picconia azorica). Furthermore, since J. brevifolia is a pioneer species (Elias & Dias 2009), low fre-quency and intensity of gap creating natural dis-turbances, at these low altitude ranges, might also explain this pattern. Below 100 m a.s.l. there is a small increase in abundance and frequency due to the presence of subsp. maritima in coastal scrubs. Once present in all islands, this species is now extinct in Graciosa and near extinction in Santa Maria. In the islands of São Miguel, São Jorge and Faial, J. brevifolia populations have been greatly reduced or highly fragmented. This de-cline was due to centuries of human exploitation of natural resources. It is well-known that the species was used since human settlement for na-val and house construction, for furniture and as fuel (Frutuoso 1978, 1981, 1987). In addition, extensive dairy farming in the last decades led to widespread clearing of forests for pasture, further decreasing populations of J. brevifolia (Silveira
2013). Nevertheless, it is still possible to find pristine Juniperus forests in Flores, Pico and Ter-ceira (Elias et al. 2011). This species is also found in coastal scrubs, in pioneer and secondary scrubs and in forested peat bogs (Elias & Silva 2008). It is protected both by national and international laws and a few studies have been developed to characterize the genetic variability of the species for conservation and restoration purposes (e.g. Silva 2000; Silva et al. 2011).
METHODS
THE AZOREAN ARCHIPELAGO The Azores are a volcanic archipelago with nine islands (Figure 2), located on the North Atlantic, on a WNW-ESE axis, between 37º - 40ºN and 25º - 31ºW. The archipelago extends for 615 km and is located approximately 1300 km west of main-land Portugal, 1600 km east of North America and 800 km NE from Madeira Island. The islands form three distinct groups: the eastern (Santa Ma-ria and São Miguel), the central (Terceira, Gra-ciosa, São Jorge, Pico and Faial) and the western (Flores and Corvo) groups with different geologi-cal ages, with Santa Maria as the oldest island (8.12 Ma) and Pico as the youngest (0.27 Ma) (França et al. 2003). The archipelago’s climate is affected by the surrounding ocean, namely the effects of the Gulf Stream, as well as by island topography, being mild and very wet, often reaching an average an-nual relative humidity of 95% in the high altitude forests. Annual temperature fluctuations are dampened by the ocean, causing low thermal var-iation throughout the year (Azevedo et al. 2004). AZOREAN VEGETATION Most Azorean extant native forests are montane cloud forests (Elias et al. 2011). They are humid, evergreen forests characterised by short tree stat-ure, dense crown foliage and high tree density. In more exposed and wet environments J. brevifolia is the dominant tree. Otherwise the canopy is usually dominated by J. brevifolia and I. perado subsp. azorica. At lower altitudes, especially in less exposed areas, L. azorica is also a common species (Elias 2007; Elias et al. 2011). Montane

Biota associated with Juniperus brevifolia
23
Biota associated with Juniperus brevifolia
23
Figure 2. Map of the Azores (distances between the island groups and between São Miguel and Santa Maria are not in scale). Sampled reserves: 1 – Morro Alto e Pico da Sé; 2 – Caldeira Funda e Rasa; 3 – Cabeço do Fogo; 4 – Caldeira; 5 – Lagoa do Caiado, 6 – Caveiro, 7 – Mistério da Prainha; 8 – Pico Pinheiro; 9 – Topo; 10 – Serra de Santa Bárbara; 11 – Biscoito da Ferraria; 12 – Terra Brava; 13 – Caldeira de Guilherme Moniz; 14 – Pico Galhar-do; 15 - Graminhais; 16 – Atalhada; 17 – Pico da Vara.
cloud forests are also characterised by a dense cover of mosses and other epiphytes (Gabriel & Bates 2005). In the Azores these forests are shaped by the shallow soil layer, fractured and craggy terrain (Ribeiro et al. 2005) and wind speed. From the work of Frutuoso (1978, 1981, 1987), compiled from documents, oral tradition and from its own travels to the islands, it is clear that for-ests dominated the Azorean landscape at the time of discovery and settlement of the archipelago in the 15th Century. Based on the classical proposal of Dansereau (1970) the vegetation of the Azores could be divided in six altitudinal belts, character-ized by their dominant species: Morella faya (0-300 m); Laurus azorica (300-650 m); Juniperus brevifolia (450-1100 m); Erica azorica (850-1600 m); Calluna vulgaris (1600-2200 m) and (6) li-chens (2200-2350 m). Palynological studies by Connor et al. (2012, 2013) provided us with some very important information about the recent past and historical native vegetation of two Azorean islands, Pico and Flores. These studies added
more evidence that primitive Azorean vegetation was dominated by trees and that human impact had a strong influence on the native vegetation. In fact, as a result of less than 600 years of human settlement more than 95% of the native forest was destroyed by human action (Triantis et al. 2010b; Fernández-Palacios et al. 2011; Silveira, 2013), with most coastal, lowland and submontane for-ests (dominated by Morella faya, Picconia azori-ca, Laurus azorica and Prunus azorica) being replaced, giving way to urban and agricultural areas. The remaining pristine forest continues under threat due to habitat reduction, fragmentation and invasive plants, with clear impacts on the flora and fauna (Borges et al. 2000; Cardoso et al. 2010; Triantis et al. 2010b; Terzopoulou et al. 2015), and is now restricted to a few dispersed high-altitude fragments, with areas between 4 and 1300 ha (Borges et al. 2005a, 2006; Gaspar et al. 2008), totalling some 6000 ha (Gaspar et al. 2008; Gabriel et al. 2011).

Nunes et al.
24
Nunes et al.
24
AZOREAN BIODIVERSITY The Azorean terrestrial biota is currently com-posed by 6199 species and subspecies (about 6147 species), being 455 (414 species) of these endemic to the archipelago (Borges et al. 2010b, with additional updated data for arthropods). A large component of this biota are recent introduc-tions, with 47% of arthropods (excluding taxa whose colonisation status was not possible to determine) and 81% of vascular plants being ex-otic (Borges et al. 2010b, with additional updated data for arthropods). There are currently 2333 arthropod taxa (269 endemics) reported to the Azores, corresponding roughly to 37% of all ter-restrial species. Fungi as a whole are the second most diverse group with 1328 taxa (34 endemics), followed by the vascular plants with 1110 taxa (73 endemics). Bryophytes (sensu lato) account roughly for 7.7% of the biota, with 480 taxa, in-cluding seven Azorean endemics. Land snails and slugs (Mollusca) are also a diverse group, with 114 taxa, including 49 endemics (Borges et al. 2010b, with additional updated data). Excluding fresh-water fishes (13 taxa), all exotic, there are 58 vertebrate species present in the archipelago: 11 of which are mammals (all introduced, except for three species of native bats and the endemic noctule bat, Nyctalus azoreum). Breeding birds are represented by 43 species, including 11 en-demic subspecies and two endemic species (Oceanodroma monteiroi and Pyrrhula murina). Additionally there are also two exotic species of amphibians and two exotic species of reptiles (Borges et al. 2010a). DATA SETS Arthropods The arthropod data set (see Appendix I) consists of the results from the BALA project (Biodiversi-ty of Arthropods of Laurisilva of the Azores), an extensive and standardized sampling protocol applied to native vegetation areas in the years of 1992-2002 and 2003-2004 (Borges et al. 2005a; Ribeiro et al. 2005; Gaspar et al. 2008; see also more details at http://islandlab.uac.pt/proj-ectos/ver.php?id=65). Each forest fragment in-cluded a minimum of four 150 m long transects (the number of transects being dependant on the forest fragment area); besides pitfall traps (not
analysed in this publication, but see Borges et al. 2005a; Gaspar et al. 2008), the two or three most common trees and shrubs per transect were sam-pled every 15 metres (for a total of 10 replicates per transect), using canopy beating. For that pur-pose, a stick and a cloth collector tray, in the shape of an inverted pyramid with a bag at the end, was used. Naturally, Juniper was not availa-ble on all of the transects, and so, for this particu-lar subset of data, each forest fragment included between one and sixteen transects. For a more detailed description of the sampling protocol see Ribeiro et al. (2005) and Gaspar et al. (2008). A total of 16 forest fragments, mostly natural patch-es of Juniperus-Sphagnum woods, Juniperus woods, Juniperus-Ilex forests and Juniperus-Laurus forests were sampled in six islands (Flo-res, Faial, Pico, São Jorge, Terceira and São Mi-guel), for a total of 74 transects and 740 samples made for Juniperus. All samples and identified species are deposited in the EDTP – Entomoteca Dalberto Teixeira Pombo, University of Azores, Angra do Heroísmo, Portugal. Occurrence data was digitized in the ATLANTIS database (http://www.atlantis.angra.uac.pt/) and available also in the Azores Bioportal (http://azores-bioportal. uac.pt/) for the general public. Other taxa Vascular plants The vascular plants list (see Appendix II.1) in-cludes epiphytic species and one hemiparasite of J. brevifolia and was based on the information from Schäfer (2005). Bryophytes The bryophyte data set used in this work includes both herbarium and literature records (Appendix II.2). From all the gathered literature references dealing with Azorean bryophytes (see complete list in Gabriel et al. 2011), 21 specifically men-tioned J. brevifolia as a substrate of either epi-phytic (growing on living bark) or epixylic spe-cies (growing on dead trunks and branches) (see Appendix III). Moreover, data from the collection of bryophytes deposited at the University of the Azores (Angra do Heroísmo) were also included in the study. Some of the herbarium records came from ad hoc inventories, while others, especially
Biota associated with Juniperus brevifolia
25
from Terceira (Gabriel & Bates 2005) and Pico Island (Homem 2005) were obtained from com-munity studies, using standard collection proto-cols on native vegetation areas; bryophyte sam-ples on J. brevifolia were obtained from relevés with 30 cm side, placed at different heights on the trees, allowing the estimation of cover and rich-ness of species (further details on Gabriel 2000). All data are included in the ATLANTIS database (http://www.atlantis.angra.uac.pt/atlantis/common/index.jsf) and also available in the Azores Bi-oportal (http://azoresbioportal.uac.pt/) for the general public. Lichens The lichen data set used in this work includes mainly literature records, although approximately one fifth (22 out of 107 species) of the lichen species associated with J. brevifolia referred in Appendix II.3, may also be found on the Collec-tion of Lichens deposited at the University of the Azores (Angra do Heroísmo). Of the 100 licheno-logical references identified for the Azores, 15 specifically refer Juniperus brevifolia as the sub-strate of the lichens (see Appendix III). Birds The bird data set used in this work (Appendix II.4) follows the literature (see Appendix III). DATA ANALYSES Arthropods For this study, we considered all adult and juve-nile specimens identified to a given morphospe-cies, while excluding all individuals that were not possible to identify or whose classification into morphospecies was dubious. The identification of juvenile/immature specimens is possible for the Azorean fauna due to the relatively low species richness of the archipelago and the long standing experience of one of the authors (PAVB) (see also Ribeiro et al. 2005; Gaspar et al. 2008). For some of the analysis the arthropods were grouped into three colonisation categories: endemic (pre-sent only in the Azores), native non-endemic (“native” for simplification from now on; includ-ing species that, although not endemic, arrived to the islands by natural means), and introduced species, after the criteria of Borges et al. (2010a). We allocated a native status for the taxa without a reliable identification, with the following excep-
tions: i) when the family was known to have only exotic species recorded for the Azores; ii) when the genera was known to be dominated by exotic species in the Azores (based on Borges et al. 2010a). Furthermore, we assigned each species to a functional group (predator, herbivore, sapro-phyte, fungivore) and a feeding mode (external digestion and sucking, chewing and cutting, piercing and sucking, siphoning, not feeding). Data for trophic guild and feeding mode were collated from an extensive literature search of ecological information, including manuscripts with the first descriptions of the species, first spe-cies records for the Azores, brief notes, and eco-logical studies among others. Information was also obtained from experts who have identified the specimens or from experts of a given taxo-nomic group when information for a particular species was not available. For the few taxa identi-fied to morphospecies, we have considered the information of the nearest taxonomic resolution (genus, family). In the case of Terceira island, to avoid pseudo-replication, when transects were sampled in more than one year, replicates were excluded, in a way as to balance the data chronologically. Addition-ally, Terceira was the only island in which some transects had more than 10 samples due to some additional experiments (see Gaspar et al. 2014). As such, and to standardize with the other islands, when transects had more than 10 samples, all samples past the first ten were excluded from the analysis. To ascertain potential differences between the islands regarding species richness, abundance, trophic guild composition and feeding mode, sta-tistical comparison tests (Chi-square) were done with R software (R Core Team, 2015) and the respective post-hoc tests for which pairs of popu-lations differ were computed with the NCStats package (Ogle 2015) by performing all chi-square tests for all pairs of populations (with Bonferroni adjustment for inflation due to multiple compari-sons). In cases where the given classes were pre-sent in only one or two islands, these where ex-cluded from the Chi-square tests. To observe the imbalance caused by the unequal sampling effort in the different islands, sample based accumula-tion curves (not shown), standardized at the min-imum common number of 40 samples were per-

Biota associated with Juniperus brevifolia
25
Biota associated with Juniperus brevifolia
25
from Terceira (Gabriel & Bates 2005) and Pico Island (Homem 2005) were obtained from com-munity studies, using standard collection proto-cols on native vegetation areas; bryophyte sam-ples on J. brevifolia were obtained from relevés with 30 cm side, placed at different heights on the trees, allowing the estimation of cover and rich-ness of species (further details on Gabriel 2000). All data are included in the ATLANTIS database (http://www.atlantis.angra.uac.pt/atlantis/common/index.jsf) and also available in the Azores Bi-oportal (http://azoresbioportal.uac.pt/) for the general public. Lichens The lichen data set used in this work includes mainly literature records, although approximately one fifth (22 out of 107 species) of the lichen species associated with J. brevifolia referred in Appendix II.3, may also be found on the Collec-tion of Lichens deposited at the University of the Azores (Angra do Heroísmo). Of the 100 licheno-logical references identified for the Azores, 15 specifically refer Juniperus brevifolia as the sub-strate of the lichens (see Appendix III). Birds The bird data set used in this work (Appendix II.4) follows the literature (see Appendix III). DATA ANALYSES Arthropods For this study, we considered all adult and juve-nile specimens identified to a given morphospe-cies, while excluding all individuals that were not possible to identify or whose classification into morphospecies was dubious. The identification of juvenile/immature specimens is possible for the Azorean fauna due to the relatively low species richness of the archipelago and the long standing experience of one of the authors (PAVB) (see also Ribeiro et al. 2005; Gaspar et al. 2008). For some of the analysis the arthropods were grouped into three colonisation categories: endemic (pre-sent only in the Azores), native non-endemic (“native” for simplification from now on; includ-ing species that, although not endemic, arrived to the islands by natural means), and introduced species, after the criteria of Borges et al. (2010a). We allocated a native status for the taxa without a reliable identification, with the following excep-
tions: i) when the family was known to have only exotic species recorded for the Azores; ii) when the genera was known to be dominated by exotic species in the Azores (based on Borges et al. 2010a). Furthermore, we assigned each species to a functional group (predator, herbivore, sapro-phyte, fungivore) and a feeding mode (external digestion and sucking, chewing and cutting, piercing and sucking, siphoning, not feeding). Data for trophic guild and feeding mode were collated from an extensive literature search of ecological information, including manuscripts with the first descriptions of the species, first spe-cies records for the Azores, brief notes, and eco-logical studies among others. Information was also obtained from experts who have identified the specimens or from experts of a given taxo-nomic group when information for a particular species was not available. For the few taxa identi-fied to morphospecies, we have considered the information of the nearest taxonomic resolution (genus, family). In the case of Terceira island, to avoid pseudo-replication, when transects were sampled in more than one year, replicates were excluded, in a way as to balance the data chronologically. Addition-ally, Terceira was the only island in which some transects had more than 10 samples due to some additional experiments (see Gaspar et al. 2014). As such, and to standardize with the other islands, when transects had more than 10 samples, all samples past the first ten were excluded from the analysis. To ascertain potential differences between the islands regarding species richness, abundance, trophic guild composition and feeding mode, sta-tistical comparison tests (Chi-square) were done with R software (R Core Team, 2015) and the respective post-hoc tests for which pairs of popu-lations differ were computed with the NCStats package (Ogle 2015) by performing all chi-square tests for all pairs of populations (with Bonferroni adjustment for inflation due to multiple compari-sons). In cases where the given classes were pre-sent in only one or two islands, these where ex-cluded from the Chi-square tests. To observe the imbalance caused by the unequal sampling effort in the different islands, sample based accumula-tion curves (not shown), standardized at the min-imum common number of 40 samples were per-

Nunes et al.
26
Biota associated with Juniperus brevifolia
27
singletons, either at archipelago level (44 species; 27%) or at island level, with particular emphasis for Terceira Island (39 species), where they ac-count for approximately 37% of the species col-lected in juniper canopies (Table 2). The same may be said for the number of uniques, with 49 uniques for the Azores as a whole and 42 uniques in Terceira (40%). Only six species (3.7%) had more than 1000 individuals. The endemic moth Argyresthia atlanticella (Lepidoptera: Yponome-utidae) was the single most abundant species col-lected on the juniper canopies, with 8483 individ-uals (23.1%). The endemic canopy sheet weaver Savigniorrhipis acoreensis (Araneae, Lyniphi-idae) and the native mesh web weaver Lathys dentichelis (Araneae, Dyctinidae) were the most
abundant spiders, with 5758 (15.7%) and 4089 (11.1%) specimens collected, respectively. The other highly diverse and abundant order, the He-miptera, are also represented by the native juniper specialist giant conifer aphid Cinara juniperi (Hemiptera, Lachnidae) with 3085 individuals (8.4%) and by the endemic planthopper from São Miguel Cixius insularis (Hemiptera, Cixiidae) with 1232 specimens (3.4%). The native cock-roach Zetha vestita ranks as the fifth most abun-dant species, with 1509 individuals (4.1%) cap-tured. These six species account for 65.8% of all collected individuals (see also Appendix I for further details), and may be considered as the most common canopy dwellers of the Azorean cedar.
Table 2. Summary table with the Arthropod species richness (S), abundance (N), diversity indexes (Fisher α, Shannon-Wiener), number of singletons and doubletons (species present with one and two individuals), uniques and duplicates (species found only in one and two samples) for the total number of individuals and for each coloni-sation class (Nat, native; End, endemic; Int, introduced), for the entire archipelago and for each individual island. Values in parenthesis indicate the corresponding standardized value, obtained from sample based accumulation curves rarefied at 40 samples. T - Total collected species. FLO – Flores; FAI – Faial; PIC – Pico; SJG – São Jorge; TER – Terceira; SMG – São Miguel.
Azores FLO FAI PIC SJG TER SMG S 161 54 (41.1) 50 (50) 73 (56.3) 62 (55.0) 106 (50.9) 67 (58.8) SNat 62 20 (13.86) 22 (22) 26 (20.2) 22 (19.87) 38 (17.25) 26 (23.72) SEnd 50 27 (21.6) 19 (19) 27 (23.2) 28 (26.3) 35 (20.2) 21 (19.1) SInt 49 7 (5.7) 9 (9) 20 (12.9) 12 (9.9) 33 (13.5) 20 (17.0) N 36729 6297 1681 7401 4119 13657 3574 NNat 11563 655 731 2120 768 6044 1245 NEnd 24159 5577 882 5036 3300 7152 2212 NInt 1007 65 68 245 51 461 117
Fisher α - 8.11 ± 0.42
9.69 ± 0.62
11.25 ± 0.51
10.35 ± 0.54
15.65 ± 0.57
11.70 ± 0.60
Shannon-Wiener - 1,97 2,58 2,71 2,04 2,69 2,25 Species per tree 9.09 ± 3.69 7.64 ± 3.71 9.28 ± 2.92 10.01 ± 4.01 9.47 ± 3.43 9.30 ± 3.29 7.63 ± 4.63 Sampled Trees 739 100 40 140 60 340 59 Singletons T 44 14 (11.1) 14 (14) 13 (11.1) 16 (14.4) 39 (14.3) 16 (16.7) Nat 21 8 7 3 6 14 7 End 8 5 5 4 4 9 2 Int 15 1 2 6 6 16 7 Doubletons T 19 5 (3.8) 4 (4) 3 (5.9) 6 (7.6) 4 (6.0) 14 (16.7) Nat 6 2 1 2 4 1 5 End 3 2 3 0 2 1 6 Int 10 1 0 1 0 2 3 Uniques T 49 15 (12.14) 16 (16) 14 (13.35) 18 (16.93) 42 (17.99) 21 (21.41) Nat 23 8 8 3 8 15 9 End 17 6 6 4 4 10 4 Int 9 1 2 7 6 17 8 Duplicates T 21 7 (4.30) 4 (4) 4 (8.89) 7 (7.99) 6 (7.12) 12 (10.07) Nat 8 3 1 3 4 3 3 End 10 3 2 0 2 1 6 Int 3 1 1 1 1 2 3
Nunes et al.
26
formed in EstimateS (Colwell 2009). This soft-ware was also used to estimate the Fisher α and the Shannon-Wiener index for each island and for the archipelago as a whole. To assess the distribution of species abundanc-es, binning of the species into modified Preston's octaves was performed using the third method described in Gray et al. (2006), using the R pack-age gambin (Matthews et al. 2014). This package was also used to calculate the corresponding Al-pha GamBin values. Nonmetric Multidimensional Scaling (Kruskal 1964) was applied to the data at a transect scale using Bray-Curtis similarities, through R software and packages vegan (Oksanen et al. 2015) and ade4 (Dray & Dufour 2007). Statistical signifi-cance in the dissimilarity between islands was assessed through an analysis of similarities (ANOSIM) (Clarke 1993). To discern the possi-ble bias in the results caused by the extremely unbalanced sampling effort between the islands, the NMDS ordination was also done using the data standardized at four randomly selected tran-sects per island. As a measure of the error associ-ated with this rarefaction of the data for the standardized NMDS analysis, 999 permutations of ANOSIM R2 values, using sets of four ran-domly selected transects per island, were estimat-ed in order to construct a 95% confidence inter-val. The number of shared species between the is-lands and the compositional differences between the islands were assessed using Bray-Curtis simi-larity, through the EstimateS software. The num-ber of shared species and Bray-Curtis similarity values were calculated by using both the full set of data (unequal numbers of transect per island) and 10 standardized sets of data each with 4 tran-sects per island. These standardized sets of data were obtained by numbering each transect in each individual island from 1 to n (where n is the max-imum number of transect in any given island) and then using the R software to generate 10 pseudo-random sets of 4 numbers from 1 to n, for each island. Other taxa Excepting Arthropods, which had enough data to allow the performance of comparative macroeco-
logical analyses, other taxa (Bryophytes [Liver-worts, Mosses], Lichens, Ferns and Birds) were analysed using a descriptive approach, in order to obtain: i) lists of species occurring on Juniperus brevifolia (Appendix II); ii) richness of species per taxonomic group and per island (whenever possible); and iii) richness of endemic, native and exotic species and their proportion regarding the Azorean taxa.
RESULTS
ARTHROPODS Arthropod taxonomical data overview We collected 36729 specimens, corresponding to 161 species, representing four Classes, 15 Orders and 66 Families (Table 1 and Appendix I). Of all the collected species, 50 (31%) are considered endemic, 62 (39%) are native and 49 (30%) are introduced in the archipelago (Table 2).
Table 1. Summary table of the collected arthropod taxa, listing all classes and orders found, with indica-tion of the number of families, species and individuals in each order.
Order / Class Fam. Spp Indiv. Arachnida Pseudoscorpiones 1 1 13 Opiliones 1 1 14 Araneae 12 40 13619 Diplopoda Julida 1 1 177 Chilopoda Lithobiomorpha 1 1 1 Insecta Microcoryphia 2 2 122 Blattodea 1 1 1509 Psocoptera 7 15 2301 Hemiptera 18 40 8840 Thysanoptera 2 9 22 Neuroptera 1 1 149 Coleoptera 11 24 153 Trichoptera 1 1 51 Lepidoptera 7 23 9370 Hymenoptera 1 1 388
Most of the species are rare in the samples, with 90 (56%) having 10 or less individuals. Quite noticeable is the relatively high proportion of

Biota associated with Juniperus brevifolia
27
Biota associated with Juniperus brevifolia
27
singletons, either at archipelago level (44 species; 27%) or at island level, with particular emphasis for Terceira Island (39 species), where they ac-count for approximately 37% of the species col-lected in juniper canopies (Table 2). The same may be said for the number of uniques, with 49 uniques for the Azores as a whole and 42 uniques in Terceira (40%). Only six species (3.7%) had more than 1000 individuals. The endemic moth Argyresthia atlanticella (Lepidoptera: Yponome-utidae) was the single most abundant species col-lected on the juniper canopies, with 8483 individ-uals (23.1%). The endemic canopy sheet weaver Savigniorrhipis acoreensis (Araneae, Lyniphi-idae) and the native mesh web weaver Lathys dentichelis (Araneae, Dyctinidae) were the most
abundant spiders, with 5758 (15.7%) and 4089 (11.1%) specimens collected, respectively. The other highly diverse and abundant order, the He-miptera, are also represented by the native juniper specialist giant conifer aphid Cinara juniperi (Hemiptera, Lachnidae) with 3085 individuals (8.4%) and by the endemic planthopper from São Miguel Cixius insularis (Hemiptera, Cixiidae) with 1232 specimens (3.4%). The native cock-roach Zetha vestita ranks as the fifth most abun-dant species, with 1509 individuals (4.1%) cap-tured. These six species account for 65.8% of all collected individuals (see also Appendix I for further details), and may be considered as the most common canopy dwellers of the Azorean cedar.
Table 2. Summary table with the Arthropod species richness (S), abundance (N), diversity indexes (Fisher α, Shannon-Wiener), number of singletons and doubletons (species present with one and two individuals), uniques and duplicates (species found only in one and two samples) for the total number of individuals and for each coloni-sation class (Nat, native; End, endemic; Int, introduced), for the entire archipelago and for each individual island. Values in parenthesis indicate the corresponding standardized value, obtained from sample based accumulation curves rarefied at 40 samples. T - Total collected species. FLO – Flores; FAI – Faial; PIC – Pico; SJG – São Jorge; TER – Terceira; SMG – São Miguel.
Azores FLO FAI PIC SJG TER SMG S 161 54 (41.1) 50 (50) 73 (56.3) 62 (55.0) 106 (50.9) 67 (58.8) SNat 62 20 (13.86) 22 (22) 26 (20.2) 22 (19.87) 38 (17.25) 26 (23.72) SEnd 50 27 (21.6) 19 (19) 27 (23.2) 28 (26.3) 35 (20.2) 21 (19.1) SInt 49 7 (5.7) 9 (9) 20 (12.9) 12 (9.9) 33 (13.5) 20 (17.0) N 36729 6297 1681 7401 4119 13657 3574 NNat 11563 655 731 2120 768 6044 1245 NEnd 24159 5577 882 5036 3300 7152 2212 NInt 1007 65 68 245 51 461 117
Fisher α - 8.11 ± 0.42
9.69 ± 0.62
11.25 ± 0.51
10.35 ± 0.54
15.65 ± 0.57
11.70 ± 0.60
Shannon-Wiener - 1,97 2,58 2,71 2,04 2,69 2,25 Species per tree 9.09 ± 3.69 7.64 ± 3.71 9.28 ± 2.92 10.01 ± 4.01 9.47 ± 3.43 9.30 ± 3.29 7.63 ± 4.63 Sampled Trees 739 100 40 140 60 340 59 Singletons T 44 14 (11.1) 14 (14) 13 (11.1) 16 (14.4) 39 (14.3) 16 (16.7) Nat 21 8 7 3 6 14 7 End 8 5 5 4 4 9 2 Int 15 1 2 6 6 16 7 Doubletons T 19 5 (3.8) 4 (4) 3 (5.9) 6 (7.6) 4 (6.0) 14 (16.7) Nat 6 2 1 2 4 1 5 End 3 2 3 0 2 1 6 Int 10 1 0 1 0 2 3 Uniques T 49 15 (12.14) 16 (16) 14 (13.35) 18 (16.93) 42 (17.99) 21 (21.41) Nat 23 8 8 3 8 15 9 End 17 6 6 4 4 10 4 Int 9 1 2 7 6 17 8 Duplicates T 21 7 (4.30) 4 (4) 4 (8.89) 7 (7.99) 6 (7.12) 12 (10.07) Nat 8 3 1 3 4 3 3 End 10 3 2 0 2 1 6 Int 3 1 1 1 1 2 3

Nunes et al.
28
Nunes et al.
28
There is a profound imbalance in the sampling effort, with Terceira Island having the highest number of individuals (13657) and comparatively high species richness, but at the same time it has more than double the number of samples of the second most sampled island (Pico Island, Table 2). When standardizing the number of samples, a more homogeneous set of richness and abundance values was obtained (Table 2). Rarity patterns If the species are grouped into modified Preston's octaves, it is possible to observe that for the total number of collected species, for native species and also for the introduced ones it follows a uni-modal distribution, with many rare species and few truly abundant species. On the other hand, the endemics present a more complex multi-modal distribution with some rare and abundant species, and also a large proportion of intermediately abundant species (Fig. 3).
Fig. 3. Modified Preston's octaves histograms for all, endemic, native and introduced species. The three graphs for the three individual colonisation status are scaled equally for the Y axis.
Considering the species in the first quarter of the octaves as truly rare (Gaston 1994), a total of 97 species (or 60% of the total number of species collected on juniper canopies) can be considered as rare in that habitat. If the different colonization classes are considered, 63% of the native species, 36% of the endemic and 53% of the introduced
species are rare in the inventory. If the same bin-ning is done to each island using the total number of species, once again the high number of locally rare species is clear on Faial, São Jorge, Terceira and São Miguel islands. The exceptions are Pico and Flores islands, which show a bimodal distri-bution with many rare and intermediate species (Fig. 4). Repeating the procedure for the islands using the different colonisation classes results in the broad maintenance of the general class pattern (see Fig. 3) in each island.
Fig. 4. Modified Preston's octaves histograms for each of the six sampled islands using all species sorted by island groups. FAI – Faial; PIC – Pico; SJG – São Jor-ge; TER – Terceira; FLO – Flores; SMG – São Miguel. All graphs except the one for Terceira (TER) are scaled equally for the Y axis.
The α GamBin parameter presents a potentially informative ecological diversity metric, because it summarizes the shape of the SAD in a single number. Alpha Gambin values between 0 and 1.5 indicate usually a Log series distribution, between 2 and 2.5 approximates to a Log normal and val-ues higher than 3, more complex multinomial distributions. For deeper explanation of the
Biota associated with Juniperus brevifolia
29
GamBin model see Matthews et al. (2014). So, as a simple and intuitive measure of the shape of the SAD's and to better compare the distinct species distributions in Figures 3 & 4, the values of Alpha GamBin and the approximate shape of the distri-butions as given by the index are presented in Table 3.
Table 3. Alpha GamBin indexes for all, endemic, native and introduced species, as well as for each of the six sampled islands, using all species. FAI – Faial; PIC – Pico; SJG – São Jorge; TER – Terceira; FLO – Flo-res; SMG – São Miguel.
Colonization Class
Alpha Gam-Bin index
Distribution shape
All 0,99 logseries Endemic 2,34 lognormal Native 0,85 logseries Introduced 1,34 logseries
Island Alpha Gam-
Bin index Distribution
shape FAI 1,47 logseries PIC 2,23 lognormal SGJ 1,08 logseries TER 0,88 logseries FLO 1,17 logseries SMG 1,09 logseries
Species richness Analysing the proportion of endemic, native and introduced species, we found that both at the Azores archipelago as a whole and at individual island level, species richness is more or less even-ly distributed amongst the three colonisation cat-egories (endemic, native and introduced) with the exception of Flores, Faial and São Jorge, where less than 20% of the species are introduced (Fig. 5). Nevertheless, there were no statistically signif-icant differences among the islands (Χ2 = 12.571, df = 10, p-value = 0.249). Considering the archipelago as a whole, the orders Araneae, Hemiptera, Coleoptera, Lepidop-tera and Psocoptera were the most diverse, with both Araneae and Hemiptera registering 40 spe-cies each, from the 161 collected. The most diverse families were Linyphiidae –
Araneae with 14 species (six introduced), Staph-ylinidae - Coleoptera with nine species (four in-troduced) and Cixiidae - Hemiptera with a total of seven endemic species. When each island is considered individually, we find that the previous pattern is similar, with Araneae and Hemiptera accounting, in each is-land, for roughly 50% of the collected species (Fig. 5), with a Chi-square test showing no signif-icant differences in the proportions of the 5 most species diverse orders between the islands (Χ2 = 12.38, df = 25, p-value >0.05).
Fig 5. Proportion of species per colonisation status for endemic, native and introduced species at archipelago and island level.
Considering the colonisation status of the species, some deviations from the previous pattern are observed (Fig. 5). For endemics the general pat-tern is maintained along the archipelago, but with higher proportions of spider and true bug species (particularly spiders). The native Hemiptera ac-count for roughly 40% of the collected native species. As with the case of the total species rich-ness, Chi-square tests showed no significant dif-ferences between the islands when considering the share of each Order in the island's species richness (native: Χ2 = 10.30, df = 25, p-value >0.05; endemic: Χ2 = 9.40, df = 25, p-value >0.05; introduced: Χ2 = 22.14, df = 25, p-value >0.05). In the case of the introduced species, Araneae and Coleoptera are the most diverse or-ders at archipelago level, but when looking at each island there is no discernible transversal pattern as was the case for the endemic and native species, notwithstanding the prevalence of spider species on most islands (Fig. 6).

Biota associated with Juniperus brevifolia
29
Biota associated with Juniperus brevifolia
29
GamBin model see Matthews et al. (2014). So, as a simple and intuitive measure of the shape of the SAD's and to better compare the distinct species distributions in Figures 3 & 4, the values of Alpha GamBin and the approximate shape of the distri-butions as given by the index are presented in Table 3.
Table 3. Alpha GamBin indexes for all, endemic, native and introduced species, as well as for each of the six sampled islands, using all species. FAI – Faial; PIC – Pico; SJG – São Jorge; TER – Terceira; FLO – Flo-res; SMG – São Miguel.
Colonization Class
Alpha Gam-Bin index
Distribution shape
All 0,99 logseries Endemic 2,34 lognormal Native 0,85 logseries Introduced 1,34 logseries
Island Alpha Gam-
Bin index Distribution
shape FAI 1,47 logseries PIC 2,23 lognormal SGJ 1,08 logseries TER 0,88 logseries FLO 1,17 logseries SMG 1,09 logseries
Species richness Analysing the proportion of endemic, native and introduced species, we found that both at the Azores archipelago as a whole and at individual island level, species richness is more or less even-ly distributed amongst the three colonisation cat-egories (endemic, native and introduced) with the exception of Flores, Faial and São Jorge, where less than 20% of the species are introduced (Fig. 5). Nevertheless, there were no statistically signif-icant differences among the islands (Χ2 = 12.571, df = 10, p-value = 0.249). Considering the archipelago as a whole, the orders Araneae, Hemiptera, Coleoptera, Lepidop-tera and Psocoptera were the most diverse, with both Araneae and Hemiptera registering 40 spe-cies each, from the 161 collected. The most diverse families were Linyphiidae –
Araneae with 14 species (six introduced), Staph-ylinidae - Coleoptera with nine species (four in-troduced) and Cixiidae - Hemiptera with a total of seven endemic species. When each island is considered individually, we find that the previous pattern is similar, with Araneae and Hemiptera accounting, in each is-land, for roughly 50% of the collected species (Fig. 5), with a Chi-square test showing no signif-icant differences in the proportions of the 5 most species diverse orders between the islands (Χ2 = 12.38, df = 25, p-value >0.05).
Fig 5. Proportion of species per colonisation status for endemic, native and introduced species at archipelago and island level.
Considering the colonisation status of the species, some deviations from the previous pattern are observed (Fig. 5). For endemics the general pat-tern is maintained along the archipelago, but with higher proportions of spider and true bug species (particularly spiders). The native Hemiptera ac-count for roughly 40% of the collected native species. As with the case of the total species rich-ness, Chi-square tests showed no significant dif-ferences between the islands when considering the share of each Order in the island's species richness (native: Χ2 = 10.30, df = 25, p-value >0.05; endemic: Χ2 = 9.40, df = 25, p-value >0.05; introduced: Χ2 = 22.14, df = 25, p-value >0.05). In the case of the introduced species, Araneae and Coleoptera are the most diverse or-ders at archipelago level, but when looking at each island there is no discernible transversal pattern as was the case for the endemic and native species, notwithstanding the prevalence of spider species on most islands (Fig. 6).

Nunes et al.
30
Nunes et al.
30
Fig. 6. Proportion of species per order at archipelago and island level, for all species, endemics, natives and intro-duced species.
Abundance Taking abundance into consideration, the general patterns change. Native species on all islands account for more than 50% of collected individu-als, but nevertheless the proportion of individuals in each colonisation level is significantly different between the islands (Χ2 = 49.84, df = 10, p-value < 0.0001) and samples from Flores are dominated by individuals of endemic species (almost 90%). Also quite strikingly, introduced species account on average for less than 4% of the number of in-dividuals collected on each island (Fig. 7). Post-hoc tests show that Flores was significantly dif-ferent from the remaining islands with the excep-tion of São Jorge. São Jorge was also significant-ly different from Terceira and Faial (see Appen-dix IV, Table 1). Considering the archipelago as a whole, Araneae is the most abundant order, with more than 13500 collected individuals, followed by Lepidoptera and Hemiptera. On the other hand, Coleoptera, despite its relatively high species richness, accounts for much less than 1% of all
collected individuals, with only 153 collected in the entire archipelago. It is also conspicuous the disparity in the proportions of each of the five most abundant orders in each island, with no ap-parent common pattern, which translates into a significant difference between them (Χ2 = 139.79, df = 25, p-value < 0.0001) (see Fig. 8). Post-hoc tests indicate that Terceira and São Miguel were significantly different from most of the other is-lands (see Appendix IV, Table 2). In all the islands spiders and true bugs account for roughly 80% of the collected specimens, but the ratio between them varies in each island, with São Jorge and São Miguel being dominated by spiders whereas in Faial and Flores true bugs ac-count for almost half of the collected individuals. There is also a relatively high abundance of the native cockroach Zetha vestita in all the islands except Faial and Pico. In the case of endemic species, it is quite conspicuous the high propor-tion of Lepidoptera individuals collected in the samples from Flores, Faial, Pico and São Jorge, mainly the ermine moth Argyresthia atlanticella

Biota associated with Juniperus brevifolia
31
Biota associated with Juniperus brevifolia
31
as well as the dominance of Hemiptera in São Miguel (roughly 55% of the collected individuals) particularly the single island endemic Cixius insu-laris. Once more, for the introduced species, there is no discernible pattern of abundance for the islands of the archipelago (see Fig. 8). Chi-square tests confirmed this graphical interpretation by showing significant differences between the is-lands at all colonisation status (native: Χ2 = 112.56, df = 25, p-value < 0.0001; endemic: Χ2 = 203.94, df = 25, p-value < 0.0001; introduced: Χ2 = 182.85, df = 25, p-value < 0.0001). For natives, post-hoc tests show that São Miguel and Faial were significantly different from most of the oth-er islands, while for endemics São Miguel was once more significantly different from the re-maining islands and Terceira was significantly different from Flores, Faial and São Jorge. For
introduced species, only the “Triangle” islands (Faial, Pico and São Jorge) showed no significant differences between them. See Appendix IV, Ta-ble 2 for further details.
Fig. 7. Proportion of abundance per colonisation status for endemic, native and introduced species at archipel-ago and island level.
Fig. 8. Proportion of abundance per order at archipelago and island level, for all species, endemics, natives and introduced species.
Functional groups and feeding modes When considering the functional groups to which the collected individuals belong, 76 species (47%) are herbivores, 60 (37 %) are predators (of which 40 are arachnids) and 24 (15%) are sapro-phytes. There is also one omnivore ant species
and 3 fungivorous species (Fig. 8). Fungivorous arthropods were represented, at archipelago level, by only 10 individuals divided amongst 3 species in Terceira and 2 individuals of one species in São Miguel. We found a great constancy in the proportion of species belonging to each functional
Biota associated with Juniperus brevifolia
31
as well as the dominance of Hemiptera in São Miguel (roughly 55% of the collected individuals) particularly the single island endemic Cixius insu-laris. Once more, for the introduced species, there is no discernible pattern of abundance for the islands of the archipelago (see Fig. 8). Chi-square tests confirmed this graphical interpretation by showing significant differences between the is-lands at all colonisation status (native: Χ2 = 112.56, df = 25, p-value < 0.0001; endemic: Χ2 = 203.94, df = 25, p-value < 0.0001; introduced: Χ2 = 182.85, df = 25, p-value < 0.0001). For natives, post-hoc tests show that São Miguel and Faial were significantly different from most of the oth-er islands, while for endemics São Miguel was once more significantly different from the re-maining islands and Terceira was significantly different from Flores, Faial and São Jorge. For
introduced species, only the “Triangle” islands (Faial, Pico and São Jorge) showed no significant differences between them. See Appendix IV, Ta-ble 2 for further details.
Fig. 7. Proportion of abundance per colonisation status for endemic, native and introduced species at archipel-ago and island level.
Fig. 8. Proportion of abundance per order at archipelago and island level, for all species, endemics, natives and introduced species.
Functional groups and feeding modes When considering the functional groups to which the collected individuals belong, 76 species (47%) are herbivores, 60 (37 %) are predators (of which 40 are arachnids) and 24 (15%) are sapro-phytes. There is also one omnivore ant species
and 3 fungivorous species (Fig. 8). Fungivorous arthropods were represented, at archipelago level, by only 10 individuals divided amongst 3 species in Terceira and 2 individuals of one species in São Miguel. We found a great constancy in the proportion of species belonging to each functional
Biota associated with Juniperus brevifolia
31
as well as the dominance of Hemiptera in São Miguel (roughly 55% of the collected individuals) particularly the single island endemic Cixius insu-laris. Once more, for the introduced species, there is no discernible pattern of abundance for the islands of the archipelago (see Fig. 8). Chi-square tests confirmed this graphical interpretation by showing significant differences between the is-lands at all colonisation status (native: Χ2 = 112.56, df = 25, p-value < 0.0001; endemic: Χ2 = 203.94, df = 25, p-value < 0.0001; introduced: Χ2 = 182.85, df = 25, p-value < 0.0001). For natives, post-hoc tests show that São Miguel and Faial were significantly different from most of the oth-er islands, while for endemics São Miguel was once more significantly different from the re-maining islands and Terceira was significantly different from Flores, Faial and São Jorge. For
introduced species, only the “Triangle” islands (Faial, Pico and São Jorge) showed no significant differences between them. See Appendix IV, Ta-ble 2 for further details.
Fig. 7. Proportion of abundance per colonisation status for endemic, native and introduced species at archipel-ago and island level.
Fig. 8. Proportion of abundance per order at archipelago and island level, for all species, endemics, natives and introduced species.
Functional groups and feeding modes When considering the functional groups to which the collected individuals belong, 76 species (47%) are herbivores, 60 (37 %) are predators (of which 40 are arachnids) and 24 (15%) are sapro-phytes. There is also one omnivore ant species
and 3 fungivorous species (Fig. 8). Fungivorous arthropods were represented, at archipelago level, by only 10 individuals divided amongst 3 species in Terceira and 2 individuals of one species in São Miguel. We found a great constancy in the proportion of species belonging to each functional
Biota associated with Juniperus brevifolia
31
as well as the dominance of Hemiptera in São Miguel (roughly 55% of the collected individuals) particularly the single island endemic Cixius insu-laris. Once more, for the introduced species, there is no discernible pattern of abundance for the islands of the archipelago (see Fig. 8). Chi-square tests confirmed this graphical interpretation by showing significant differences between the is-lands at all colonisation status (native: Χ2 = 112.56, df = 25, p-value < 0.0001; endemic: Χ2 = 203.94, df = 25, p-value < 0.0001; introduced: Χ2 = 182.85, df = 25, p-value < 0.0001). For natives, post-hoc tests show that São Miguel and Faial were significantly different from most of the oth-er islands, while for endemics São Miguel was once more significantly different from the re-maining islands and Terceira was significantly different from Flores, Faial and São Jorge. For
introduced species, only the “Triangle” islands (Faial, Pico and São Jorge) showed no significant differences between them. See Appendix IV, Ta-ble 2 for further details.
Fig. 7. Proportion of abundance per colonisation status for endemic, native and introduced species at archipel-ago and island level.
Fig. 8. Proportion of abundance per order at archipelago and island level, for all species, endemics, natives and introduced species.
Functional groups and feeding modes When considering the functional groups to which the collected individuals belong, 76 species (47%) are herbivores, 60 (37 %) are predators (of which 40 are arachnids) and 24 (15%) are sapro-phytes. There is also one omnivore ant species
and 3 fungivorous species (Fig. 8). Fungivorous arthropods were represented, at archipelago level, by only 10 individuals divided amongst 3 species in Terceira and 2 individuals of one species in São Miguel. We found a great constancy in the proportion of species belonging to each functional

Nunes et al.
32
Biota associated with Juniperus brevifolia
33
Figure 9. Proportion of species and abundance per different functional groups at archipelago and island level. S - Saprophyte, P/H - Omnivorous; P - Predator; H - Herbivore; F - Fungivore.
Fig. 10. Proportion of species and abundance per different feeding modes at archipelago and island level. Ex - External digestion and sucking; Ch - Chewing and cutting; Pi - Piercing and sucking; Si - Siphoning; No - Not feeding.
Biota associated with Juniperus brevifolia
33
Figure 9. Proportion of species and abundance per different functional groups at archipelago and island level. S - Saprophyte, P/H - Omnivorous; P - Predator; H - Herbivore; F - Fungivore.
Fig. 10. Proportion of species and abundance per different feeding modes at archipelago and island level. Ex - External digestion and sucking; Ch - Chewing and cutting; Pi - Piercing and sucking; Si - Siphoning; No - Not feeding.
Nunes et al.
32
groups across all the islands (Chi-square tests showed no significant differences – Χ2 = 11.56, df = 20, p-value > 0.05). In all the islands almost half the species present in juniper trees are herbi-vores, and predators (mostly spiders) represent roughly 35% of the species. Only in São Miguel there are slightly more predator than herbivore species. Saprophytes make up the majority of the remaining species. When looking at the division in functional groups by number of collected individuals (Fig. 9), Chi-square tests showed significant differ-ences between the islands (Χ2 = 64.08, df = 15, p-value < 0.0001). The only common feature is that the herbivores were more abundant than predators and saprophytes combined, with the exception of Terceira, were there were more specimens of predators than those of herbivores in the juniper canopies and in São Miguel, where there is a rela-tively high proportion of saprophytes. Post-hoc tests show that Terceira is significantly different from Faial and Flores, and that São Miguel is significantly different from Faial and São Jorge (see Appendix IV, Table 3). When the feeding mode is considered (Fig. 10), it is possible to observe a repetition of the previ-ous patterns, with the proportions of each catego-ry being more or less conserved across the archi-pelago where species are concerned (and no sig-nificant differences were observed – Χ2 = 13.81, df = 35, p-value >0.05). Considering the propor-tion of collected individuals in each category, once again, no common pattern is observable, with the general pattern for each island following the trends for trophic guild and for the orders when considering the total of individuals. Again, a Chi-square test indicates that the islands are significantly different from each other (Χ2 = 151.48, df = 35, p-value < 0.0001). Post-hoc tests show that São Miguel was significantly different from the other islands. Terceira was also signifi-cantly different from all other islands except Pico (Pico showed no significant differences from the islands of the Central Group nor from Flores). See Appendix IV, Table 4 for further details. Species community similarity The islands from the central group have the high-est proportion of shared species as shown by the higher number of common species and by the
higher values of Bray-Curtis similarity (Table 4). Pico, Terceira and São Jorge islands also have a high number of shared species with Flores, whereas São Miguel has the lowest values of sim-ilarity with the remaining islands. Considering the colonisation status (Table 4), we can say that the previous pattern is probably driven by the endem-ic species, as these maintain and even exacerbate the trends observed for the complete data set, whereas the native species display higher values of Bray-Curtis similarity, and therefore are not spatially structured. Introduced species are in general present in low numbers, display more or less homogeneous values of Bray-Curtis similari-ty and do not seem to conform to a particular spa-tial pattern, appearing spread throughout the is-lands. Using the rarefied set of data (Table 5) we can state that the general similarity trends from the observed data are maintained, with the islands from the Central Group being the most similar and with São Miguel and Flores being the most dissimilar. The rarefaction also smoothed down the effects of the sampling bias for Terceira, as these values are more on a par with those from the remaining islands of the Central Group. To complement our similarity analysis, we ordinated transects according to their similarity values (measured with Bray-Curtis coefficient) using NMDS. The first two axes of the NMDS had a stress value of 0.2 indicating that two di-mensions were enough to give an acceptable rep-resentation of our data. The NMDS using the full set of data (Fig. 11) reveal that: 1) the islands of the Central Group share many species; 2) in the two dimensional space defined by the NMDS, the dispersion of the transects of each island and par-ticularly for Terceira is noticeably low (i.e. tran-sects within an island share many species); 3) São Miguel Island stands apart from the remaining of the archipelago and the scattering of transects is particularly high (i.e. strong heterogeneity be-tween between transects in terms of species com-position); and 4) Flores Island stands slightly apart from the Central Group and diametrically opposite to São Miguel. In general, the configura-tion of islands on the NMDS plot follows their spatial geographical configuration. As expected, the introduced species break this pattern, in ac-cordance with the results from Tables 4 and 5.

Biota associated with Juniperus brevifolia
33
Biota associated with Juniperus brevifolia
33
Figure 9. Proportion of species and abundance per different functional groups at archipelago and island level. S - Saprophyte, P/H - Omnivorous; P - Predator; H - Herbivore; F - Fungivore.
Fig. 10. Proportion of species and abundance per different feeding modes at archipelago and island level. Ex - External digestion and sucking; Ch - Chewing and cutting; Pi - Piercing and sucking; Si - Siphoning; No - Not feeding.
Biota associated with Juniperus brevifolia
33
Figure 9. Proportion of species and abundance per different functional groups at archipelago and island level. S - Saprophyte, P/H - Omnivorous; P - Predator; H - Herbivore; F - Fungivore.
Fig. 10. Proportion of species and abundance per different feeding modes at archipelago and island level. Ex - External digestion and sucking; Ch - Chewing and cutting; Pi - Piercing and sucking; Si - Siphoning; No - Not feeding.

Nunes et al.
34
Biota associated with Juniperus brevifolia
35
Fig. 11. Nonmetric Dimensional Scaling (NMDS) with Bray-Curtis similarities for the six islands using: Total - the full set of data; endemic native and introduced species data.
This separation of São Miguel from the remaining of the archipelago is even more clear when con-sidering only the endemic species and it is equally clear the dissimilarity between transects. This pattern is contrary to the apparent similarity ob-served between the remaining islands. The native communities seem to be fairly similar in each island and also across the archipelago, with the groups showing a great overlap, whereas for the introduced species, all the islands have fairly sim-ilar species compositions, although with some heterogeneity between transects in each island. This visual assessment of the NMDS ordina-tion plots can be statistically tested by using the allied statistical method of Analysis of Similari-ties (ANOSIM). When considering the whole community or only the endemic species, the com-positional dissimilarities seen in the ordination plots are confirmed by significantly high R2 val-ues (Total: R2 = 0.76, p = 0.001; endemic: R2 = 0.83, p = 0.001) obtained. The analysis of similar-
ities for the native species indicates that there are still significant dissimilarities between the species composition of each island, but the relatively lower R2 value hints at a more uniform distribu-tion of the species in the archipelago and/or at a higher intra-island dissimilarity (R2 = 0.39, p = 0.001). The ANOSIM for the introduced species indicates that there are no significant dissimilari-ties between islands/clusters and that the intra-island dissimilarity is high (R2 = 0.02, p = 0.328). Regarding the standardized data, we can observe that the general patterns shown by the full set of data are broadly maintained either with all the individuals or when divided by colonisation sta-tus, as can be seen by the example in Figure 12. In fact, some trends are even more noticeable, such as the differentiation of São Miguel from the remaining of the archipelago or the diametrical opposite position of Flores Island in relation to São Miguel (Fig. 12).
Tabl
e 4.
Su
mm
ary
of th
e re
sults
of
the
shar
ed s
peci
es a
naly
sis
for
the
obse
rved
dat
a us
ing
all s
peci
es, e
ndem
ics
nativ
es a
nd i
ntro
duce
d. M
ain
diag
onal
– n
umbe
r of
spe
cies
pre
sent
in
the
isla
nd, u
pper
hal
f –
num
ber
of s
hare
d sp
ecie
s bet
wee
n th
e is
land
s, lo
wer
hal
f – B
ray-
Cur
tis si
mila
rity
inde
x va
lues
. O
bser
ved
Tot
al
TE
R
FAI
FLO
PI
C
SJG
SM
G
Nat
T
ER
FA
I FL
O
PIC
SJ
G
SMG
T
ER
10
6 41
41
54
50
45
T
ER
38
17
13
19
15
16
FA
I 0,
18
50
33
42
38
34
FAI
0,19
22
12
17
15
16
FL
O
0,36
0,
22
54
38
38
32
FLO
0,
19
0,3
20
14
12
12
PIC
0,
53
0,32
0,
6 73
46
43
PI
C
0,43
0,
49
0,36
26
17
16
SJ
G
0,33
0,
36
0,63
0,
64
62
38
SJG
0,
21
0,43
8 0,
58
0,51
22
14
SM
G
0,19
0,
24
0,14
0,
27
0,24
67
SM
G
0,31
0,
46
0,3
0,61
0,
53
26
End
T
ER
FA
I FL
O
PIC
SJ
G
SMG
In
t T
ER
FA
I FL
O
PIC
SJ
G
SMG
T
ER
35
17
21
22
24
18
T
ER
33
7
7 13
11
11
FA
I 0,
17
19
15
18
16
12
FAI
0,22
9
6 7
7 6
FLO
0,
45
0,19
27
18
20
14
FL
O
0,24
0,
68
7 6
6 6
PIC
0,
59
0,23
0,
67
27
21
17
PIC
0,
47
0,36
0,
32
20
8 10
SJ
G
0,42
0,
33
0,64
0,
7 28
17
SJ
G
0,2
0,71
0,
64
0,26
12
7
SMG
0,
09
0,1
0,1
0,12
0,
14
21
SMG
0,
19
0,2
0,34
0,
3 0,
2 20
Ta
ble
5. S
umm
ary
with
the
geom
etric
mea
ns o
f the
resu
lts fr
om th
e 10
set
s of
rare
fied
shar
ed s
peci
es a
naly
sis
usin
g al
l spe
cies
, en
dem
ics,
nat
ives
and
int
rodu
ced.
Mai
n di
agon
al –
ave
rage
d nu
mbe
r of
spe
cies
pre
sent
in
the
isla
nd, u
pper
hal
f –
aver
aged
nu
mbe
r of s
hare
d sp
ecie
s bet
wee
n th
e is
land
s, lo
wer
hal
f – a
vera
ged
Bra
y-C
urtis
sim
ilarit
y in
dex
valu
es.
Rar
efie
d T
otal
T
ER
FA
I FL
O
PIC
SJ
G
SMG
N
at
TE
R
FAI
FLO
PI
C
SJG
SM
G
TE
R
44,6
29
,2
23,3
30
,2
31,9
24
,2
TE
R
14,7
11
,6
7,1
10,4
10
,1
9,3
FAI
0,43
50
25
,8
35,9
35
,2
28
FAI
0,51
22
8,
7 14
,1
13,5
13
,6
FLO
0,
35
0,37
37
,6
26,2
27
,7
22
FLO
0,
26
0,22
12
,2
8,2
7,9
8,2
PIC
0,
49
0,55
0,
51
52,9
35
,6
28,5
PI
C
0,53
0,
58
0,32
18
,6
11,2
11
,7
SJG
0,
42
0,43
0,
56
0,56
55
,1
28,7
SJ
G
0,61
0,
45
0,43
0,
53
18,5
11
,3
SMG
0,
33
0,23
0,
11
0,23
0,
19
53
SMG
0,
59
0,46
0,
16
0,43
0,
47
20,9
End
T
ER
FA
I FL
O
PIC
SJ
G
SMG
In
t T
ER
FA
I FL
O
PIC
SJ
G
SMG
T
ER
18
,2
12,5
11
,9
13
15,1
9,
3 T
ER
11
,2
4,9
4,1
6,6
6,4
5,3
FAI
0,37
19
12
,3
15,8
15
,5
9,6
FAI
0,28
9
4,7
6 6,
1 4,
7 FL
O
0,37
0,
43
19,7
13
,4
15,5
9,
5 FL
O
0,25
0,
46
5,5
4,3
4,1
4,1
PIC
0,
47
0,52
0,
56
21,8
18
11
PI
C
0,3
0,6
0,35
12
,4
6,1
5,5
SJG
0,
35
0,41
0,
58
0,56
26
,2
12,3
SJ
G
0,32
0,
53
0,46
0,
46
10,3
4,
9 SM
G
0,17
0,
09
0,09
0,
12
0,09
16
,3
SMG
0,
15
0,17
0,
23
0,24
0,
17
15,4

Biota associated with Juniperus brevifolia
35
Biota associated with Juniperus brevifolia
35
Fig. 11. Nonmetric Dimensional Scaling (NMDS) with Bray-Curtis similarities for the six islands using: Total - the full set of data; endemic native and introduced species data.
This separation of São Miguel from the remaining of the archipelago is even more clear when con-sidering only the endemic species and it is equally clear the dissimilarity between transects. This pattern is contrary to the apparent similarity ob-served between the remaining islands. The native communities seem to be fairly similar in each island and also across the archipelago, with the groups showing a great overlap, whereas for the introduced species, all the islands have fairly sim-ilar species compositions, although with some heterogeneity between transects in each island. This visual assessment of the NMDS ordina-tion plots can be statistically tested by using the allied statistical method of Analysis of Similari-ties (ANOSIM). When considering the whole community or only the endemic species, the com-positional dissimilarities seen in the ordination plots are confirmed by significantly high R2 val-ues (Total: R2 = 0.76, p = 0.001; endemic: R2 = 0.83, p = 0.001) obtained. The analysis of similar-
ities for the native species indicates that there are still significant dissimilarities between the species composition of each island, but the relatively lower R2 value hints at a more uniform distribu-tion of the species in the archipelago and/or at a higher intra-island dissimilarity (R2 = 0.39, p = 0.001). The ANOSIM for the introduced species indicates that there are no significant dissimilari-ties between islands/clusters and that the intra-island dissimilarity is high (R2 = 0.02, p = 0.328). Regarding the standardized data, we can observe that the general patterns shown by the full set of data are broadly maintained either with all the individuals or when divided by colonisation sta-tus, as can be seen by the example in Figure 12. In fact, some trends are even more noticeable, such as the differentiation of São Miguel from the remaining of the archipelago or the diametrical opposite position of Flores Island in relation to São Miguel (Fig. 12).

Nunes et al.
36
Biota associated with Juniperus brevifolia
37
Table 6 presents the results for the observed (un-rarefied) set of data together with mean R2 and the 95% confidence interval for the rarefied data. For the total number of species and for the en-demic species, the constructed confidence inter-vals (CIs) are relatively narrow and the observed R2 value is slightly higher than the upper quantile of the CI. The observed ANOSIM R2 for native species sits inside the CI for this category. For the introduced species the mean R2 is also quite low and the observed R2 actually falls under the lower quantile of the constructed CI.
Table 6. Summary table for ANOSIM results for the total of arthropod species, endemics, natives and intro-duced and for 999 ANOSIM rarefied replicates with the corresponding mean R2 and 95% confidence inter-val for each of the aforementioned categories.
Observed - R2 Signif. Total - 0,76 0,001 Endemic - 0,83 0,001 Native - 0,39 0,001 Introduced - 0,02 0,328 Rarefied replicates CI 95%
Mean R2 -
Total 0.52 - 0.74 0,63 - Endemic 0.58 - 0.79 0,69 - Native 0.09 - 0.41 0,24 - Introduced 0.04 - 0.31 0,17 -
Other taxa Juniperus brevifolia, existing in a wide range of elevation belts, accommodates a large number of taxa. Besides Arthropods, it is also an important substrate to other vascular species, to bryophytes (liverworts and mosses), to lichens and also to birds. The list of occurrence of all the document-ed taxa may be found in Appendix II, while the bibliographic references are mentioned in Appen-dix III. The endemic hemi-parasitic Arceuthobium azoricum is of particular interest among flowering species, and, among the five fern species prefer-entially epiphytes on J. brevifolia, two are con-sidered very rare, Ceradenia jungermannioides and Grammitis azorica. In the field, it is some-times possible to observe other vascular plants growing on Juniperus brevifolia, such as Lactuca watsoniana, Erica azorica, Myrsine africana or even seedlings of the Juniperus brevifolia itself.
There is a large number of bryophyte species growing on Juniperus brevifolia (105; 22.1%) (Appendix II.1). All of these are either endemic of the archipelago (one; 14.3%), the Macaronesia (five; 35.7%), the Iberian Macaronesia (four; 100%), Europe (seven; 58.3%) or native to the Azores (Appendix II). The proportion of species growing on Juni-perus brevifolia differs among the islands (c.f. Table 7), but apart from Graciosa (where the tree is no longer present) and Santa Maria (where ap-propriate references are lacking), all other islands have liverworts and mosses referred to this sub-strate. The islands of Pico (Homem 2005), São Jorge and Terceira (cf. Gabriel 2000; Gabriel & Bates 2005) present the highest richness values of species growing on this tree. The variation among these values is probably more related to the detail of studies published for each island, than to any real differences of the quality of the substrate. Recently, in inventories made with the Moveclim protocol (cf. Gabriel et al. 2014), many specimens were collected from Juniperus brevifolia and later works will probably even out these values. From the 105 bryophytes growing on Juniperus brevifolia, 30 species are considered as Conserva-tion Concern by the IUCN (Dierssen 2001) (cf. Appendix II). Vulnerable species include nine liverworts (Acrobolbus wilsonii, Aphanolejeunea azorica, A. sintenisii, Cephalozia crassifolia, Cheilolejeunea cedercreutzii, Leptoscyphus azoricus, Pallavicinia lyellii, Telaranea azorica, Tylimanthus laxus) and one moss (Daltonia sten-ophylla). Lichens are another taxonomic group that takes advantage of Juniperus brevifolia as a substrate (Appendix II.3). From the 781 lichens referred to the Azores (Aptroot et al. 2010), 106 were de-scribed as growing on this tree (Table 8). Apart from Dictyonema interruptum, one of the six Ba-sidiomycota referred to the archipelago, the 105 Ascomycota are distributed among the islands in an uneven way, most of the species from Terceira Island (Aptroot et al. 2009), but with the highest proportion in Flores Island, where lichens occur-ring on Juniperus brevifolia bark account for about one third of the species (37; 31.1%). Five lichen species occurring on Juniperus brevifolia are considered endemic of the archipelago (Nephroma hensseniae; N. venosum, Peltigera
Fi
g. 1
2.
Exam
ples
of N
onm
etric
Dim
ensio
nal S
calin
g (N
MDS
) with
Bra
y-Cu
rtis d
istan
ces f
or th
e six
islan
ds, u
sing
four
rand
omly
selec
ted tr
anse
cts p
er is
land,
for t
he to
tal se
t of s
pecie
s

Biota associated with Juniperus brevifolia
37
Biota associated with Juniperus brevifolia
37
Table 6 presents the results for the observed (un-rarefied) set of data together with mean R2 and the 95% confidence interval for the rarefied data. For the total number of species and for the en-demic species, the constructed confidence inter-vals (CIs) are relatively narrow and the observed R2 value is slightly higher than the upper quantile of the CI. The observed ANOSIM R2 for native species sits inside the CI for this category. For the introduced species the mean R2 is also quite low and the observed R2 actually falls under the lower quantile of the constructed CI.
Table 6. Summary table for ANOSIM results for the total of arthropod species, endemics, natives and intro-duced and for 999 ANOSIM rarefied replicates with the corresponding mean R2 and 95% confidence inter-val for each of the aforementioned categories.
Observed - R2 Signif. Total - 0,76 0,001 Endemic - 0,83 0,001 Native - 0,39 0,001 Introduced - 0,02 0,328 Rarefied replicates CI 95%
Mean R2 -
Total 0.52 - 0.74 0,63 - Endemic 0.58 - 0.79 0,69 - Native 0.09 - 0.41 0,24 - Introduced 0.04 - 0.31 0,17 -
Other taxa Juniperus brevifolia, existing in a wide range of elevation belts, accommodates a large number of taxa. Besides Arthropods, it is also an important substrate to other vascular species, to bryophytes (liverworts and mosses), to lichens and also to birds. The list of occurrence of all the document-ed taxa may be found in Appendix II, while the bibliographic references are mentioned in Appen-dix III. The endemic hemi-parasitic Arceuthobium azoricum is of particular interest among flowering species, and, among the five fern species prefer-entially epiphytes on J. brevifolia, two are con-sidered very rare, Ceradenia jungermannioides and Grammitis azorica. In the field, it is some-times possible to observe other vascular plants growing on Juniperus brevifolia, such as Lactuca watsoniana, Erica azorica, Myrsine africana or even seedlings of the Juniperus brevifolia itself.
There is a large number of bryophyte species growing on Juniperus brevifolia (105; 22.1%) (Appendix II.1). All of these are either endemic of the archipelago (one; 14.3%), the Macaronesia (five; 35.7%), the Iberian Macaronesia (four; 100%), Europe (seven; 58.3%) or native to the Azores (Appendix II). The proportion of species growing on Juni-perus brevifolia differs among the islands (c.f. Table 7), but apart from Graciosa (where the tree is no longer present) and Santa Maria (where ap-propriate references are lacking), all other islands have liverworts and mosses referred to this sub-strate. The islands of Pico (Homem 2005), São Jorge and Terceira (cf. Gabriel 2000; Gabriel & Bates 2005) present the highest richness values of species growing on this tree. The variation among these values is probably more related to the detail of studies published for each island, than to any real differences of the quality of the substrate. Recently, in inventories made with the Moveclim protocol (cf. Gabriel et al. 2014), many specimens were collected from Juniperus brevifolia and later works will probably even out these values. From the 105 bryophytes growing on Juniperus brevifolia, 30 species are considered as Conserva-tion Concern by the IUCN (Dierssen 2001) (cf. Appendix II). Vulnerable species include nine liverworts (Acrobolbus wilsonii, Aphanolejeunea azorica, A. sintenisii, Cephalozia crassifolia, Cheilolejeunea cedercreutzii, Leptoscyphus azoricus, Pallavicinia lyellii, Telaranea azorica, Tylimanthus laxus) and one moss (Daltonia sten-ophylla). Lichens are another taxonomic group that takes advantage of Juniperus brevifolia as a substrate (Appendix II.3). From the 781 lichens referred to the Azores (Aptroot et al. 2010), 106 were de-scribed as growing on this tree (Table 8). Apart from Dictyonema interruptum, one of the six Ba-sidiomycota referred to the archipelago, the 105 Ascomycota are distributed among the islands in an uneven way, most of the species from Terceira Island (Aptroot et al. 2009), but with the highest proportion in Flores Island, where lichens occur-ring on Juniperus brevifolia bark account for about one third of the species (37; 31.1%). Five lichen species occurring on Juniperus brevifolia are considered endemic of the archipelago (Nephroma hensseniae; N. venosum, Peltigera

Nunes et al.
38
Biota associated with Juniperus brevifolia
39
that hyperdiverse and biologically important groups like Hymenoptera and Diptera, that can be extremely abundant and diverse on tree canopies (Guilbert 1997; Kitching et al. 1997) were not considered in the BALA protocol (except for ants) can somewhat hamper direct comparisons. Focusing now on data from the Azores, the overall general patterns regarding species rich-ness and abundance agree with general patterns observed in previous studies for the communities of arthropods in Azorean native forests (see Ri-beiro et al. 2005; Gaspar et al. 2008), which is not surprising, since juniper samples are an important component of BALA project data. In all instances Araneae, Hemiptera, Lepidoptera and Coleoptera account for more than 75% of the species record-ed. Spiders were previously reported as the most abundant order overall (Gaspar et al. 2008) and also the one with the highest ratio of juveniles to adults (Gaspar et al. 2008). This is also true in the case of our study. This high abundance and diver-sity of spiders confirms the propensity of an ar-chitecturally complex tree as the J. brevifolia to accumulate species in general and spiders in par-ticular (Ribeiro et al. 2005; Ribeiro & Borges 2010). This high number of juvenile spiders, most of them of indigenous species, shows that source populations are building up and completing their life cycle on Juniper, taking advantage of the structural complexity of the host tree to protect their postures and juveniles (see e.g. Borges et al. 2008). Still regarding the spiders, Linyphiidae was the most diverse family of all the collected orders, something that could be related with the high dispersal capabilities of this family (Thomas et al. 2003) allowing for higher chances of arri-val/colonisation from the continental landmasses and for easier dispersal amongst the different is-lands (Borges & Wunderlich 2008; Cardoso et al. 2010; Carvalho & Cardoso 2014). The relatively high proportion of singletons found in this study is also in accordance with pre-vious findings (Gaspar et al. 2008). The amount of singletons for Terceira is considerably higher than in aforementioned study, but it is still lower than the values reported for herbivore singletons in canopies given in Ribeiro et al. (2005). Addi-tionally, the percentage of rare species as given
by binning the species into octaves (60%) was similar to the results obtained by Ribeiro & Bor-ges (2010) for juniper trees (60% of rare species), although these authors also present the rarity data pertaining to three feeding classes, with corre-sponding differences in the proportion of rare species. Our results also support these authors' findings regarding the fact that many rare species accumulate in the canopies of J. brevifolia. For native and introduced species, it may also be the case that these species have greater dispersal ca-pabilities and thus, could more easily end up as habitat-tourists (Borges et al. 2008). This is al-most certainly the case for most introduced spe-cies, which almost invariably present low abun-dances, indicating that the core populations are outside natural forests. It is also possible that this high number of (most likely) locally rare species could be a result of undersampling and/or a com-bination of phenological, methodological and spatial edge effects (Scharff et al. 2003; Cardoso et al. 2008). Concerning the functional groups of the col-lected specimens, both our study and Gaspar et al. (2008) agree on the dominance of predators and herbivores, in terms of abundance and species richness, with these accounting, on all instances, for more than 80% of the individuals or species (respectively) and, mainly in terms of species richness being the proportions of the different functional groups being quite constant through the archipelago. This is not dissimilar from the figures given by Winchester (1997) for temperate coniferous forests or by Krüger & McGavin (2001) for tropical savannah’s, and agrees with patterns of functional group proportion constancy in other parts of the world (Moran & Southwood 1982; Stork 1987, 1991). Herbivores were slight-ly more diverse and abundant than predators, but in our study, Terceira (for abundance) and São Miguel (for richness) deviate from this pattern. On the other hand, Borges et al. (2008) also found more predator than herbivore species on their samples from Terceira. One of the possible rea-sons might be the aforementioned propensity of juniper to accumulate spiders (Ribeiro & Borges 2010). For herbivores, especially chewers, lower diversity might also be related with the reduced number of food choices available on Juniperus (see Winchester 1997).
Nunes et al.
38
dissecta, Peltigera melanorrhiza and Thelotrema antoninii), while three species of the genus Usneaare considered Macaronesian endemic (Usnea geissleriana, U. macaronesica and U. subflam-mea). Besides plants and lichens, also birds use Juniperus brevifolia tree. It is possible to find explicit references in literature (Bannerman &
Bannerman 1966; Gabriel 2000; Cabral et al. 2005 and Moura 2014 - Appendix III) to the three subspecies of firecrest (Regulus regulus azoricus, R. r. inermis and R. r. sanctaemariae) and the Azores wood pigeon (Columba palumbus azori-ca) (Appendix II.4). All these subspecies are en-demic to the Azores.
Table 7. Number and proportion of bryophyte species occurring in Juniperus brevifolia and in all substrata, for each of the nine Azorean Islands and the Azores (total) for liverworts (Division Marchantiophyta) and mosses (Di-vision Bryophyta).
DivisionSubstrate Cor Flo Fai Pic Gra SJo Ter SMi SMa Total
1015.2%
109.3%
1513.8%
4940.5%
00.0%
1736.2%
5538.2%
75.5%
00.0%
6539.6%
66 107 109 121 47 131 144 127 74 164
Marchantiophyta
J. brevi olia
All substrata
BryophytaJ. brevi olia 3
2.7%7
4.2%6
3.4%23
14.4%0
0.0%11
13.3%29
13.6%2
0.9%0
0.0%40
12.9%All substrata 110 168 174 160 179 83 214 230 139 311
Table 8. Number and proportion of lichen species occurring in Juniperus brevifolia and in all substrata, for each of the nine Azorean Islands and the Azores (total) including Division Ascomycota and Division Basidiomycota.
Division Substrate Cor Flo Fai Pic Gra SJo Ter SMi SMa TotalAscomycotaJ. brevi olia 0
(0.0%)37
(31.1%)13
(6.6%)29
(12.8%)0
(0.0%)3
(2.0%)85
(18.9%)13
(3.4%)0
(0.0%)105
(13.5%)
All substrata 13 119 196 227 149 148 449 384 56 775
Basidiomycota
J. brevi olia 0(0.0%)
0(0.0%)
0(0.0%)
0(0.0%)
0(0.0%)
0(0.0%)
1(100%)
0(0.0%)
0(0.0%)
1(%)
All substrata 0 1 1 1 0 2 1 3 0 6
DISCUSSION
There is a large body of studies regarding arthro-pod canopy communities, encompassing a wide range of habitats and spectrum of host plants (Stork 1991; Winchester 1997; Kuria et al. 2010). In many of these studies there is a remarkable constancy in the proportions of certain functional
groups and orders (Moran & Southwood 1982; Krüger & McGavin 2001) and the most abundant and diverse orders in our study (Araneae, Hemip-tera, Lepidoptera) also tend to be amongst the most abundant and diverse in other parts of the world (Stork 1991; Floren & Linsenmair 1997). In our study, Coleoptera are an exception that will be discussed further below. Nevertheless, the fact

Biota associated with Juniperus brevifolia
39
Biota associated with Juniperus brevifolia
39
that hyperdiverse and biologically important groups like Hymenoptera and Diptera, that can be extremely abundant and diverse on tree canopies (Guilbert 1997; Kitching et al. 1997) were not considered in the BALA protocol (except for ants) can somewhat hamper direct comparisons. Focusing now on data from the Azores, the overall general patterns regarding species rich-ness and abundance agree with general patterns observed in previous studies for the communities of arthropods in Azorean native forests (see Ri-beiro et al. 2005; Gaspar et al. 2008), which is not surprising, since juniper samples are an important component of BALA project data. In all instances Araneae, Hemiptera, Lepidoptera and Coleoptera account for more than 75% of the species record-ed. Spiders were previously reported as the most abundant order overall (Gaspar et al. 2008) and also the one with the highest ratio of juveniles to adults (Gaspar et al. 2008). This is also true in the case of our study. This high abundance and diver-sity of spiders confirms the propensity of an ar-chitecturally complex tree as the J. brevifolia to accumulate species in general and spiders in par-ticular (Ribeiro et al. 2005; Ribeiro & Borges 2010). This high number of juvenile spiders, most of them of indigenous species, shows that source populations are building up and completing their life cycle on Juniper, taking advantage of the structural complexity of the host tree to protect their postures and juveniles (see e.g. Borges et al. 2008). Still regarding the spiders, Linyphiidae was the most diverse family of all the collected orders, something that could be related with the high dispersal capabilities of this family (Thomas et al. 2003) allowing for higher chances of arri-val/colonisation from the continental landmasses and for easier dispersal amongst the different is-lands (Borges & Wunderlich 2008; Cardoso et al. 2010; Carvalho & Cardoso 2014). The relatively high proportion of singletons found in this study is also in accordance with pre-vious findings (Gaspar et al. 2008). The amount of singletons for Terceira is considerably higher than in aforementioned study, but it is still lower than the values reported for herbivore singletons in canopies given in Ribeiro et al. (2005). Addi-tionally, the percentage of rare species as given
by binning the species into octaves (60%) was similar to the results obtained by Ribeiro & Bor-ges (2010) for juniper trees (60% of rare species), although these authors also present the rarity data pertaining to three feeding classes, with corre-sponding differences in the proportion of rare species. Our results also support these authors' findings regarding the fact that many rare species accumulate in the canopies of J. brevifolia. For native and introduced species, it may also be the case that these species have greater dispersal ca-pabilities and thus, could more easily end up as habitat-tourists (Borges et al. 2008). This is al-most certainly the case for most introduced spe-cies, which almost invariably present low abun-dances, indicating that the core populations are outside natural forests. It is also possible that this high number of (most likely) locally rare species could be a result of undersampling and/or a com-bination of phenological, methodological and spatial edge effects (Scharff et al. 2003; Cardoso et al. 2008). Concerning the functional groups of the col-lected specimens, both our study and Gaspar et al. (2008) agree on the dominance of predators and herbivores, in terms of abundance and species richness, with these accounting, on all instances, for more than 80% of the individuals or species (respectively) and, mainly in terms of species richness being the proportions of the different functional groups being quite constant through the archipelago. This is not dissimilar from the figures given by Winchester (1997) for temperate coniferous forests or by Krüger & McGavin (2001) for tropical savannah’s, and agrees with patterns of functional group proportion constancy in other parts of the world (Moran & Southwood 1982; Stork 1987, 1991). Herbivores were slight-ly more diverse and abundant than predators, but in our study, Terceira (for abundance) and São Miguel (for richness) deviate from this pattern. On the other hand, Borges et al. (2008) also found more predator than herbivore species on their samples from Terceira. One of the possible rea-sons might be the aforementioned propensity of juniper to accumulate spiders (Ribeiro & Borges 2010). For herbivores, especially chewers, lower diversity might also be related with the reduced number of food choices available on Juniperus (see Winchester 1997).

Nunes et al.
40
Biota associated with Juniperus brevifolia
41
pies, still maintain a high proportion of its origi-nal fauna. This pattern is not exclusive of J. brevifolia, but a characteristic of the native Azorean canopies in general, as further articles dealing with the remaining structurally important native trees should confirm. As such, it can be said that the canopies in the extant patches of native forest act as a small window to what was the original pristine Azorean arthropod fauna, at least above the 500 m altitude. BEETLES (COLEOPTERA) The near absence of beetles (Coleoptera) in the canopies of juniper is quite remarkable. Coleop-tera is the most diverse group of organisms, with approximately 360 000 described species world-wide (Bouchard et al. 2011). It is also a fact that of all the Macaronesia archipelagos, Azores has the lowest proportion of indigenous (native and endemic) beetle species (Triantis et al. 2010a). But this alone does not explain why only 153 individuals were collected, despite the fact that it was the third most diverse group. Coleoptera diversity in the Azores, even if low compared with that of the Canaries or with other Macaronesian archipelagos (Triantis et al. 2010a) is not that dissimilar from the World's average (Borges et al. 2005c). According to this same study, 31% of arthropod species in the Azores are beetles, this rising to 35% if only endemics are considered. Gaspar et al. (2008) also reports high beetle diversity for native forests of Azores (ap-prox. 30%) but low abundance, with beetles mak-ing up only 5% of the collected individuals. This pattern of relatively low diversity and low abun-dance of beetles for the native habitats of Azores is also alluded to by other authors (Gaspar et al. 2008; Amorim et al. 2012; Terzopoulou et al. 2015; Borges et al. in prep.) and is in sharp con-trast with canopy studies in other parts of the world, where this group can comprise more than a quarter of all collected individuals (Stork 1991; Wagner 1997). The percentage of Coleoptera species in our study was low (13%) compared with these studies. Moreover, an important per-centage of them are introduced and occurring at low number, thus they might just be habitat tour-ists. Also, it was previously demonstrated that a large portion of the beetle species present in the Azores are epigean ground-dwellers (Gaspar et al.
2008) or fungivores with cryptic lifestyles (Amo-rim et al. 2012) and thus, would not appear in the canopy, or appear only as habitat tourists. Never-theless, these facts alone do not explain the dis-proportionately small number of collected indi-viduals. We propose two possible explanations for this disharmony: i) the Azores lost most of its forest area in the last 600 years (Triantis et al. 2010b) leaving only some small high altitude patches. Survivors of this habitat destruction would have been pushed upwards to the higher altitudes and many species could have been una-ble to cope with the new and harsher climatic conditions. In this scenario, ground-dwelling spe-cies would have been less susceptible to these changes, as this stratum is a somewhat more sta-ble and sheltered habitat compared with the cano-pies, that are more exposed to the abiotic factors (Grimbacher & Stork 2007). Additionally, canopy beetle species have in general larger body-sizes than their ground-dwelling counterparts (Grim-bacher & Stork 2007), making then inherently more extinction-prone (Terzopoulou et al. 2015). If this hypothesis was true we should expect to find more species of beetles in canopies at lower altitudes, which is not the case, at least for sam-ples available from Pico Island (unpublished da-ta); ii) alternatively there is an eco-evolutionary explanation: trees of the genus Juniperus, as well as many other Cupressaceae are known to pro-duce several essential oils and other substances that are toxic or repellent to arthropods, beetles included (Carroll et al. 2010; Abad et al. 2013; Athanassiou et al. 2013). It can therefore be as-sumed that the probabilities of arrival to the ar-chipelago of species capable of feeding and re-producing on J. brevifolia or its continental rela-tive J. navicularis, at the right geologic moment, would be low. Also, it is possible that due to the fact that most of the archipelago's land area is younger than 1 Ma (Triantis et al. 2010a), most of the species that could potentially colonize juniper canopies would not have had enough time to spe-ciate and adapt to that particular habitat. In spite of this, of all the arthropod groups evaluated in Triantis et al. (2010b), Coleoptera were consid-ered to be at a higher extinction risk, and as such, it is possible that what we observe now in the canopies is a consequence of the aforementioned deforestation of the last 600 years or even a recur-
Nunes et al.
40
Also as previously reported (Borges et al. 2008; Gaspar et al. 2008), fungivores were the least well represented group, with only the aforementioned 12 individuals collected, divided between 3 spe-cies. This is not altogether surprising since the fungivorous species present in the archipelago belong to families composed mainly of specimens with small body-sizes and cryptic lifestyles. Compounding to this fact, the sampling was done on live tree specimens, whereas the adults and juveniles of these species live mainly on rotten wood, under tree bark or in the fruiting bodies of macrofungi (Watson & Dallwitz 2003; Amorim et al. 2012), thus reducing the chances of being col-lected using the present methodology. COLONISATION STATUS Looking at the colonisation status of the collected specimens, and keeping in mind that we focused in only one host tree species, our results are sub-stantially different from those of previous studies that take into account both, ground and canopy arthropods or the totality of the Azorean habitats. Borges et al. (2010b, with additional updated da-ta) gives 269 endemic arthropods for the Azores, out of a total of 2070 species and subspecies with a well defined colonisation status, which gives us a figure of approximately 13% of endemics. The same study reported that 47% of the arthropod species are introduced. Gaspar et al. (2008), sam-pling ground and canopy on native forest frag-ments and Borges et al. (2006) sampling the ground layer on Terceira forest fragments gives less grim figures, with approximately 25% of endemic species and 33% of introduced species. Florencio et al. (2013), working with epigean arthropods across several habitats reports 13.5% of endemics and 55% of introduced species. These results contrast with ours, as we found that the overall percentage of endemic species on ju-niper canopies was around 31% (and reaching 50% in Flores Island), whereas the percentage of introduced species was of 30%, but being lower than 20% in Flores, Faial and São Jorge (Fig. 4). Abundance of endemic and introduced species also differed from previous studies. Figures of almost 50% of endemics and 11% of introduced species are reported in Gaspar et al. (2008), while Florencio et al. (2013) states that 10 endemic and
10 introduced species accounted for 75% of all collected individuals. In our study, the percentage of introduced species on juniper canopies was extremely low, only less than 4% of individuals at archipelago level, but being as low as 1% on is-lands such as Flores and São Jorge (Fig. 7). This large difference in the relative proportions of abundance and diversity for introduced species in juniper canopies is particularly conspicuous when comparing figures 5 and 7. As it was seen in previous studies across dif-ferent habitats, with a higher or lower degree of anthropic disturbance, or even on the epigean soil component of the extant native forests (Borges et al. 2006, Gaspar et al. 2008; Cardoso et al. 2009; Meijer et al. 2011; Florencio et al. 2013), there is a high abundance and richness of introduced spe-cies. Only the canopies, in our case, of J. brevifo-lia seem to remain resistant to the establishment and dominance by introduced species. The data seems to indicate that introduced species are able to disperse to the juniper canopies either from the ground strata or from nearby disturbed habitats (Borges et al. 2008), but that, for now, are unable to gain a foothold and colonise this habitat. This might be due to the high structural complexity of the juniper canopies and its high predator (spider) load (Ribeiro & Borges 2010) or to some sort of hitherto unknown chemical deterrent (see next section). However, since this pattern is also ob-served for the canopies of other native host tree species (Ribeiro & Borges 2010), it can also be due to the fact that the canopies, with their higher diversity and abundance of native and endemic species, could represent an habitat with less emp-ty ecological niches or due to the fact that intro-duced generalist species are unable to properly access the nutritional resources present, to find shelter or the appropriate reproductive conditions. It can also be that this pattern is caused by the climatic harshness of the constant conditions of high humidity in the canopies of the Azorean high altitude forests remnants (see Grimbacher & Stork 2007). Whatever the true cause might be, from these and previous results it is possible to assume that despite the high proportion of introduced species in the Azores archipelago, the remnants of the native forests, and more specifically their cano-

Biota associated with Juniperus brevifolia
41
Biota associated with Juniperus brevifolia
41
pies, still maintain a high proportion of its origi-nal fauna. This pattern is not exclusive of J. brevifolia, but a characteristic of the native Azorean canopies in general, as further articles dealing with the remaining structurally important native trees should confirm. As such, it can be said that the canopies in the extant patches of native forest act as a small window to what was the original pristine Azorean arthropod fauna, at least above the 500 m altitude. BEETLES (COLEOPTERA) The near absence of beetles (Coleoptera) in the canopies of juniper is quite remarkable. Coleop-tera is the most diverse group of organisms, with approximately 360 000 described species world-wide (Bouchard et al. 2011). It is also a fact that of all the Macaronesia archipelagos, Azores has the lowest proportion of indigenous (native and endemic) beetle species (Triantis et al. 2010a). But this alone does not explain why only 153 individuals were collected, despite the fact that it was the third most diverse group. Coleoptera diversity in the Azores, even if low compared with that of the Canaries or with other Macaronesian archipelagos (Triantis et al. 2010a) is not that dissimilar from the World's average (Borges et al. 2005c). According to this same study, 31% of arthropod species in the Azores are beetles, this rising to 35% if only endemics are considered. Gaspar et al. (2008) also reports high beetle diversity for native forests of Azores (ap-prox. 30%) but low abundance, with beetles mak-ing up only 5% of the collected individuals. This pattern of relatively low diversity and low abun-dance of beetles for the native habitats of Azores is also alluded to by other authors (Gaspar et al. 2008; Amorim et al. 2012; Terzopoulou et al. 2015; Borges et al. in prep.) and is in sharp con-trast with canopy studies in other parts of the world, where this group can comprise more than a quarter of all collected individuals (Stork 1991; Wagner 1997). The percentage of Coleoptera species in our study was low (13%) compared with these studies. Moreover, an important per-centage of them are introduced and occurring at low number, thus they might just be habitat tour-ists. Also, it was previously demonstrated that a large portion of the beetle species present in the Azores are epigean ground-dwellers (Gaspar et al.
2008) or fungivores with cryptic lifestyles (Amo-rim et al. 2012) and thus, would not appear in the canopy, or appear only as habitat tourists. Never-theless, these facts alone do not explain the dis-proportionately small number of collected indi-viduals. We propose two possible explanations for this disharmony: i) the Azores lost most of its forest area in the last 600 years (Triantis et al. 2010b) leaving only some small high altitude patches. Survivors of this habitat destruction would have been pushed upwards to the higher altitudes and many species could have been una-ble to cope with the new and harsher climatic conditions. In this scenario, ground-dwelling spe-cies would have been less susceptible to these changes, as this stratum is a somewhat more sta-ble and sheltered habitat compared with the cano-pies, that are more exposed to the abiotic factors (Grimbacher & Stork 2007). Additionally, canopy beetle species have in general larger body-sizes than their ground-dwelling counterparts (Grim-bacher & Stork 2007), making then inherently more extinction-prone (Terzopoulou et al. 2015). If this hypothesis was true we should expect to find more species of beetles in canopies at lower altitudes, which is not the case, at least for sam-ples available from Pico Island (unpublished da-ta); ii) alternatively there is an eco-evolutionary explanation: trees of the genus Juniperus, as well as many other Cupressaceae are known to pro-duce several essential oils and other substances that are toxic or repellent to arthropods, beetles included (Carroll et al. 2010; Abad et al. 2013; Athanassiou et al. 2013). It can therefore be as-sumed that the probabilities of arrival to the ar-chipelago of species capable of feeding and re-producing on J. brevifolia or its continental rela-tive J. navicularis, at the right geologic moment, would be low. Also, it is possible that due to the fact that most of the archipelago's land area is younger than 1 Ma (Triantis et al. 2010a), most of the species that could potentially colonize juniper canopies would not have had enough time to spe-ciate and adapt to that particular habitat. In spite of this, of all the arthropod groups evaluated in Triantis et al. (2010b), Coleoptera were consid-ered to be at a higher extinction risk, and as such, it is possible that what we observe now in the canopies is a consequence of the aforementioned deforestation of the last 600 years or even a recur-

Nunes et al.
42
Biota associated with Juniperus brevifolia
43
scriptive analysis to the biota associated with the other structurally important trees from the Azorean indigenous forests. This holistic ap-proach also emphasises the necessity for studies targeting less studied and neglected groups.
ACKNOWLEDGMENTS
The authors of this manuscript would like to thank Enésima Mendonça for the shapefiles used in the construction of Figure 2 and to Sociedade de Gestão Ambiental e Conservação da Natureza – Azorina S.A. For the permission to use the sci-entific illustration of Figure 1. We are grateful to all researchers that collaborated in the field and laboratory during the BALA project funded by Direcção Regional dos Recursos Florestais (Pro-ject 17.01-080203): Álvaro Vitorino, Anabela Arraiol, Ana Rodrigues, Artur Serrano, Carlos Aguiar, Catarina Melo, Francisco Dinis, Genage André, Emanuel Barcelos, Fernando Pereira, Hu-go Mas, Isabel Rosário, João Amaral, Joaquín Hortal, Lara Dinis, Paula Gonçalves, Sandra Jar-roca, Sérvio Ribeiro and Luís Vieira. The Forest Services provided local support in each island. Acknowledgments are due to all the taxonomists who assisted in the identification of the mor-photypes: Andrew Polaszek, António Sousa, Ar-tur Serrano, Arturo Baz, Fernando Ilharco, Henrik Enghoff, Jordi Ribes, José Quartau, Jörg Wunder-lich, Mário Boieiro, Ole Karsholt, Richard Stras-sen, Volker Manhert and Virgílio Vieira. RN was funded by the Ph.D. Grant DRCT – M3.1.2/F/035/2011.
REFERENCES
Abad, M.K.R., B.A. Besheli & M.M. Sharif 2013. Toxicity and Repellency Effect of Arizona Cypress, Cupressus Arizonica Essential oil on three Coleopteran Stored Product Pests. International Journal of Agronomy and Plant Production 4(10): 2531-2536.
Adams, R.P. 2014. Junipers of the world: the genus Juniperus (4th edition). Trafford Publishing Co., Bloomington, Indiana, USA. 415 pp.
Aguiar, A.M.F. & O. Karsholt 2009. Lepidoptera: Systematic catalogue of the entomofauna of the Madeira Archipelago and Selvagens Islands 1.
Museu Municipal do Funchal. Boletim, Suplemento 9: 5-140.
Amorim, I.R., B.C. Emerson, P.A.V. Borges & R.K. Wayne 2012. Phylogeography and molecular phylogeny of Macaronesian island Tarphius (Coleoptera: Zopheridae): why are there so few species in the Azores? Journal of Biogeography 39: 1583-1595.
Aptroot, A., F. Schumm & R. Gabriel 2009. Lichens new to the Azores collected on Terceira. Herzogia, 22: 147-152.
Aptroot, A., A.F. Rodrigues, F. Schumm, S. Câmara & R. Gabriel 2010. List of lichens and lichenicolous fungi (Fungi). pp. 59-79, In: Borges, P.A.V., A. Costa, R. Cunha, R. Gabriel, V. Gonçalves, A. F. Martins, I. Melo, M. Parente, P. Raposeiro, P. Rodrigues, R.S. Santos, L. Silva, P. Vieira & V. Vieira (Eds.) A list of the terrestrial and marine biota from the Azores. Princípia, Cascais.
Aranda, S.C., R. Gabriel, P.A.V. Borges, A.M.C. Santos, E.B. Azevedo, J. Patiño, J. Hortal & J.M. Lobo 2014. Geographical, temporal and environmental determinants of bryophyte species richness in the Macaronesian islands. PLoSOne 9(7): e101786. Available from: http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0101786
Aranda, S.C., H. Hespanhol, N. Homem, P.A.V. Borges, J.M. Lobo & R. Gabriel 2015. The iterative process of plant species inventorying to obtain reliable biodiversity patterns. Botanical Journal of the Linnean Society 177(4): 491-503. DOI: 10.1111/boj.12259.
Athanassiou, C.G., N.G. Kavallieratos, E. Evergetis, A.M. Katsoula & S.A. Haroutounian 2013. Insecticidal efficacy of silica gel with Juniperus oxycedrus ssp. oxycedrus (Pinales: Cupressaceae) essential oil against Sitophilus oryzae (Coleoptera: Curculionidae) and Tribolium confusum (Coleoptera: Tenebrionidae). Journal of economic entomology 106(4): 1902-1910.
Azevedo, E.B., M.C. Rodrigues, & J.F. Fernandes 2004. Climatologia. Pp. 25-48 in: Forjaz, V.H. (Coord.) Atlas básico dos Açores. Observatório Vulcanológico e Geotérmico dos Açores, Ponta Delgada. 112 pp.
Borges, P.A.V. & V.K. Brown 1999. Effect of island geological age on the arthropod species richness of Azorean pastures. Biological Journal of the Linnean Society 66: 373-410.
Borges, P.A.V., A.M.R. Serrano & J.A. Quartau 2000. Ranking the Azorean Natural Forest Reserves for conservation using their endemic arthropods. Journal of Insect Conservation 4: 129-147.
Borges, P.A.V. & J. Wunderlich 2008. Spider
Nunes et al.
42
rent pattern with extinctions happening in the past due to some natural event. SIMILARITIES IN ARTHROPOD ISLAND COMMUNITIES The observed patterns of similarity between the islands were to be expected and are in accordance with previous works (e.g. Ribeiro et al. 2005). The islands from the Central group (Terceira, São Jorge, Pico and Faial) possess fairly similar communities, while Flores and, more conspicu-ously, São Miguel stand further apart. For native species the similarities between the islands are much stronger and the differences between São Miguel and Flores can likely be explained by geographical reasons, as both islands sit some 500 km apart on a WNW-ESE axis, and consequently will have different rates of species arrivals, either from continental landmasses or from other islands and archipelagos. The observed dissimilarity in species composition between São Miguel and the other islands is mainly due to the endemic spe-cies, and most likely derived by evolutionary pro-cesses. São Miguel is the oldest sampled island, meaning that there was more time for in situ spe-ciation, increasing the number of endemic and single island endemic species (Borges & Brown 1999; Whittaker et al. 2008; Borges & Hortal 2009). São Miguel is also the closest island to Santa Maria, the oldest island (and from which it would also be interesting to have comparison data), and a probable source of further endemic species (Borges & Hortal 2009; Amorim et al. 2012). The physical distance between the Eastern and Central groups would also help cement this dissimilarity by hindering the dispersal of the species. Regarding the introduced species, the lower R2 value obtained suggests a grouping of these spe-cies driven by factors acting on different scales other than the island/geological scale (namely historical introduction and habitat change histo-ry). Several studies indicate that introduced spe-cies increase the similarity between locations (Case 1996; Dormann et al. 2007; Florencio et al. 2013; to name a few), but in this particular case it seems that introduced species have not played yet a big role on the homogenisation of the canopy species assemblages through the archipelago, most likely due to the low number of introduced
specimens present in the juniper canopies, which is in itself a probable consequence of the (for now) low capacity of habitat penetration and col-onisation. Nevertheless, the introduced species assemblies are relatively similar throughout the juniper canopies in sampled Azorean islands. We also recognise the need to study previously neglected but ecologically important arthropod orders such as Hymenoptera and Diptera (this knowledge gap is nevertheless starting to be ad-dressed by currently ongoing studies in Terceira Island), as well as increasing the studies of other important taxonomic groups present in the native forests of the Azores, such as gastropods, fungi, epiphytes, birds and bats. OTHER BIOTAS The number of lichens and bryophytes listed as associated with the Azorean juniper is remarka-ble. Not only the number of endemic species is comparatively high, considering that neither of the groups has more than 10 Azorean endemic species (respectively 10 and seven; Borges et al., 2010a), but the number of conservation concern species for bryophytes is remarkable (30 out of 70; Gabriel et al. 2011). The complexity of the tree and the high diversity of micro-habitats cre-ated the conditions for epiphytic species to easily colonize all parts of the tree, from the bottom, trunk and branches. It is obvious that some is-lands are in need of further studies to complete list of the associated taxa, since there is a large difference among them in the number of species reported for Juniperus brevifolia.
FINAL REMARKS Contrary to other habitats in the archipelago, the canopies of indigenous trees in natural forests, and in this particular case, Juniper canopies still maintain a large proportion of native and endemic species, with a low penetration of exotics, and could therefore be considered as windows (albeit incomplete ones) to the original faunal communi-ties of the Azores. A comparison of the findings of this study with those regarding the other Azorean trees would allow a better look at the constancy in the community structure across host tree species and across the islands. It should also be stressed the necessity to do this kind of de-

Biota associated with Juniperus brevifolia
43
Biota associated with Juniperus brevifolia
43
scriptive analysis to the biota associated with the other structurally important trees from the Azorean indigenous forests. This holistic ap-proach also emphasises the necessity for studies targeting less studied and neglected groups.
ACKNOWLEDGMENTS
The authors of this manuscript would like to thank Enésima Mendonça for the shapefiles used in the construction of Figure 2 and to Sociedade de Gestão Ambiental e Conservação da Natureza – Azorina S.A. For the permission to use the sci-entific illustration of Figure 1. We are grateful to all researchers that collaborated in the field and laboratory during the BALA project funded by Direcção Regional dos Recursos Florestais (Pro-ject 17.01-080203): Álvaro Vitorino, Anabela Arraiol, Ana Rodrigues, Artur Serrano, Carlos Aguiar, Catarina Melo, Francisco Dinis, Genage André, Emanuel Barcelos, Fernando Pereira, Hu-go Mas, Isabel Rosário, João Amaral, Joaquín Hortal, Lara Dinis, Paula Gonçalves, Sandra Jar-roca, Sérvio Ribeiro and Luís Vieira. The Forest Services provided local support in each island. Acknowledgments are due to all the taxonomists who assisted in the identification of the mor-photypes: Andrew Polaszek, António Sousa, Ar-tur Serrano, Arturo Baz, Fernando Ilharco, Henrik Enghoff, Jordi Ribes, José Quartau, Jörg Wunder-lich, Mário Boieiro, Ole Karsholt, Richard Stras-sen, Volker Manhert and Virgílio Vieira. RN was funded by the Ph.D. Grant DRCT – M3.1.2/F/035/2011.
REFERENCES
Abad, M.K.R., B.A. Besheli & M.M. Sharif 2013. Toxicity and Repellency Effect of Arizona Cypress, Cupressus Arizonica Essential oil on three Coleopteran Stored Product Pests. International Journal of Agronomy and Plant Production 4(10): 2531-2536.
Adams, R.P. 2014. Junipers of the world: the genus Juniperus (4th edition). Trafford Publishing Co., Bloomington, Indiana, USA. 415 pp.
Aguiar, A.M.F. & O. Karsholt 2009. Lepidoptera: Systematic catalogue of the entomofauna of the Madeira Archipelago and Selvagens Islands 1.
Museu Municipal do Funchal. Boletim, Suplemento 9: 5-140.
Amorim, I.R., B.C. Emerson, P.A.V. Borges & R.K. Wayne 2012. Phylogeography and molecular phylogeny of Macaronesian island Tarphius (Coleoptera: Zopheridae): why are there so few species in the Azores? Journal of Biogeography 39: 1583-1595.
Aptroot, A., F. Schumm & R. Gabriel 2009. Lichens new to the Azores collected on Terceira. Herzogia, 22: 147-152.
Aptroot, A., A.F. Rodrigues, F. Schumm, S. Câmara & R. Gabriel 2010. List of lichens and lichenicolous fungi (Fungi). pp. 59-79, In: Borges, P.A.V., A. Costa, R. Cunha, R. Gabriel, V. Gonçalves, A. F. Martins, I. Melo, M. Parente, P. Raposeiro, P. Rodrigues, R.S. Santos, L. Silva, P. Vieira & V. Vieira (Eds.) A list of the terrestrial and marine biota from the Azores. Princípia, Cascais.
Aranda, S.C., R. Gabriel, P.A.V. Borges, A.M.C. Santos, E.B. Azevedo, J. Patiño, J. Hortal & J.M. Lobo 2014. Geographical, temporal and environmental determinants of bryophyte species richness in the Macaronesian islands. PLoSOne 9(7): e101786. Available from: http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0101786
Aranda, S.C., H. Hespanhol, N. Homem, P.A.V. Borges, J.M. Lobo & R. Gabriel 2015. The iterative process of plant species inventorying to obtain reliable biodiversity patterns. Botanical Journal of the Linnean Society 177(4): 491-503. DOI: 10.1111/boj.12259.
Athanassiou, C.G., N.G. Kavallieratos, E. Evergetis, A.M. Katsoula & S.A. Haroutounian 2013. Insecticidal efficacy of silica gel with Juniperus oxycedrus ssp. oxycedrus (Pinales: Cupressaceae) essential oil against Sitophilus oryzae (Coleoptera: Curculionidae) and Tribolium confusum (Coleoptera: Tenebrionidae). Journal of economic entomology 106(4): 1902-1910.
Azevedo, E.B., M.C. Rodrigues, & J.F. Fernandes 2004. Climatologia. Pp. 25-48 in: Forjaz, V.H. (Coord.) Atlas básico dos Açores. Observatório Vulcanológico e Geotérmico dos Açores, Ponta Delgada. 112 pp.
Borges, P.A.V. & V.K. Brown 1999. Effect of island geological age on the arthropod species richness of Azorean pastures. Biological Journal of the Linnean Society 66: 373-410.
Borges, P.A.V., A.M.R. Serrano & J.A. Quartau 2000. Ranking the Azorean Natural Forest Reserves for conservation using their endemic arthropods. Journal of Insect Conservation 4: 129-147.
Borges, P.A.V. & J. Wunderlich 2008. Spider

Nunes et al.
44
Biota associated with Juniperus brevifolia
45
Colwell, R.K., 2009. EstimateS: statistical estimation of species richness and shared species from samples, Version 8.2. User’s guide and application published online at http://purl.oclc.org/estimates.
Connor, S.E., J.F.N. van Leeuwen, T.M. Rittenour, W.O. van der Knaap, B. Ammann & S. Björck 2012. The ecological impact of oceanic island colonization – a palaeoecological perspective from the Azores. Journal of Biogeography 39(6): 1007-1023.
Connor, S.E., W.O. van der Knaap, J.F.N. van Leeuwen & P. Kuneš 2013. Holocene palaeoclimate and palaeovegetation on the islands of Flores and Pico. pp: 149-162 in: Fernández-Palacios, J.M., L. Nascimento, J.C. Hernández,, S. Clemente, A. González & J.P. Díaz-González (Eds.) Climate change perspectives from the Atlantic: Past, present and Future. Servicio de Publicaciones, Universidad de La Laguna. Tenerife, Canárias.
Dansereu, P. 1970. Macaronesian studies IV. Natural ecosystems of the Azores. Revue Canadienne de Géographie 24(1): 21-42.
Dierssen, K. 2001. Distribution, ecological amplitude and phytosociological characterization of European bryophytes. Bryophytorum Bibliotheca, 56: 1-289.
Dormann, C.F., O. Schweiger, I. Augenstein, D. Bailey, R. Billeter, G. de Blust, R. DeFilippi, et al. 2007. Effects of landscape structure and land-use intensity on similarity of plant and animal communities. Global Ecology and Biogeography 16(6): 774-787.
Dray, S. & A.B. Dufour 2007. The ade4 package: implementing the duality diagram for ecologists. Journal of Statistical Software 22(4): 1-20.
Elias, R.B. 2007. Ecologia das florestas de Juniperus dos Açores. PhD thesis, University of the Azores. Angra do Heroísmo, Azores, Portugal. 179pp.
Elias, R.B. & L. Silva 2008. Juniperus brevifolia (Seub.) Antoine. pp. 182–183. in: Martín, J.L., M.J. Arechavaleta, P.A.V. Borges & B. Faria (Eds.). Top 100 Las cien especies amenazadas prioritarias de gestión en la región europea biogeográfica de la Macaronesia. Consejeria de Medio Ambiente y Ordenación Territorial, Gobierno de Canarias. 500 pp.
Elias, R.B. & E. Dias 2009. Gap dynamics and regeneration strategies in Juniperus-Laurus forests of the Azores islands. Plant Ecology 200: 179–189.
Elias, R.B., E. Dias & F. Pereira 2011. Disturbance, regeneration and the spatial pattern of tree species in Azorean mountain forests. Community Ecology 12: 23–30.
Elias, R.B. & E. Dias 2014. The recognition of infraspecific taxa in Juniperus brevifolia
(Cupressaceae). Phytotaxa 188(5): 241-250. Fernández-Palacios, J.M., L. De Nascimento, R. Otto,
J.D. Delgado, E. García-del-Rey, J.R. Arévalo, et al. 2011. A reconstruction of Palaeo-Macaronesia, with particular reference to the long-term biogeography of the Atlantic island laurel forests. Journal of Biogeography 38: 226-246.
Ferreira, R.C., R. Piredda, F. Bagnoli, R. Bellarosa, M. Attimonelli, S. Fineschi, B. Schirone & M.C. Simeone 2011. Phylogeography and conservation perspectives of an endangered macaronesian endemic: Picconia azorica (Tutin) Knobl. (Oleaceae). European Journal of Forest Research 130: 181-195.
Floren, A., & K.E. Linsenmair 1997. Diversity and recolonization dynamics of selected arthropod groups on different tree species in a lowland rainforest in Sabah, Malaysia with special reference to Formicidae. pp 344-381. in Stork, N.E., J. Adis & R.K. Didham (Eds.). Canopy arthropods. Chapman & Hall. London. 567pp.
Florencio, M., P. Cardoso, J.M. Lobo, E.B. Azevedo & P.A.V. Borges. 2013. Arthropod assemblage homogenization in oceanic islands: the role of exotic and indigenous species under landscape disturbance. Diversity and Distributions 19: 1450-1460.
França, Z., J.V. Cruz, J.C. Nunes & V-H. Forjaz 2003. Geologia dos Açores: uma perspectiva actual. Açoreana 10: 11–140.
Frutuoso, G. 1978. Sixth book of the land memories (1859) [Livro sexto das saudades da terra] 1589, In Portuguese]. Clutural Institute of Ponta Delgada, Azores. Portugal. 432 pp.
Frutuoso, G. 1981. Fourth book of the land memories (1859) [Livro sexto das saudades da terra 1589, In Portuguese]. Clutural Institute of Ponta Delgada, Azores. Vol. II. 397 pp.
Frutuoso, G. 1981. Fourth book of the land memories (1859) [Livro sexto das saudades da terra 1589, In Portuguese]. Clutural Institute of Ponta Delgada, Azores. Vol. III.
Gabriel, R. 2000. Ecophysiology of Azorean forest bryophytes. Ph. D. thesis. Imperial College of Science, Technology and Medicine, London University. Silwood Park, Ascot, Berkshire. 310 pp.
Gabriel, R., & .W. Bates 2005. Bryophyte community composition and habitat specificity in the natural forests of Terceira, Azores. Plant Ecology 177(1): 125-144.
Gabriel, R., N. Homem, A. Couto, S.C. Aranda & P.A.V. Borges 2011. Azorean bryophytes: a preliminary review of rarity patterns. Açoreana 7: 149-206.
Nunes et al.
44
biodiversity patterns and their conservation in the Azorean archipelago, with description of new taxa. Systematics and Biodiversity 6: 249-282
Borges, P.A.V. & J. Hortal 2009. Time, area and isolation: factors driving the diversification of Azorean arthropods. Journal of Biogeography 36: 178-191.
Borges, P.A.V., C. Aguiar, J. Amaral, I.R. Amorim, G. André, A. Arraiol, A. Baz et al. 2005a. Ranking protected areas in the Azores using standardized sampling of soil epigean arthropods. Biodiversity and Conservation 14: 2029-2060.
Borges, P.A.V., R. Cunha, R. Gabriel, A.M. Martins, L. Silva & V. Vieira (Eds.) 2005b. A list of the terrestrial fauna (Mollusca and Arthropoda) and flora (Bryophyta, Pteridophyta and Spermatophyta) from the Azores. Direcção Regional do Ambiente & Universidade dos Açores, Horta, Angra do Heroísmo & Ponta Delgada. 318pp.
Borges, P.A., R.T.D. Cunha, R. Gabriel, A.M. Martins, L. Silva, V. Vieira et al. 2005c. Description of the terrestrial Azorean biodiversity. Pp: 21-68 in: Borges, P.A.V., R. Cunha, R. Gabriel, A.M. Martins, L. Silva & V. Vieira (Eds.). A list of the terrestrial fauna (Mollusca and Arthropoda) and flora (Bryophyta, Pteridophyta and Spermatophyta) from the Azores. Regional Environment Directorate of the Azores & University of the Azores, Horta, Angra do Heroísmo & Ponta Delgada. 318pp.
Borges, P.A.V., J.M. Lobo, E.B. Azevedo, C. Gaspar, C. Melo & L.V. Nunes 2006. Invasibility and species richness of island endemic arthropods: a general model of endemic vs. exotic species. Journal of Biogeography 33: 169-187.
Borges, P.A.V., K.I. Ugland, F.O. Dinis & C. Gaspar 2008. Insect and spider rarity in an oceanic island (Terceira, Azores): true rare and pseudo-rare species. pp. 47-70 in: Fattorini, S. (Ed.). Insect Ecology and Conservation. Research Signpost, Kerala, India. ISBN: 978-81-308-0297-8, 317 pp.
Borges, P.A.V., A. Costa, R. Cunha, R. Gabriel, V. Gonçalves, A.F. Martins, I. Melo, et al. (Eds.) 2010a. A list of the terrestrial and marine biota from the Azores. Princípia, Cascais, 432 pp.
Borges, P.A.V., A. Costa, R. Cunha, R. Gabriel, V. Gonçalves, A.F. Martins, I. Melo, et al. 2010b. Description of the Terrestrial and marine biodiversity of the Azores. Pp. 9-33 in: Borges, P.A.V., A. Costa, R. Cunha, R. Gabriel, V. Gonçalves, A.F. Martins, I. Melo, et al. (Eds). A list of the terrestrial and marine biota from the Azores. Princípia, Cascais, 432 pp.
Bouchard P., Y. Bousquet, A. Davies, M. Alonso-
Zarazaga, J. Lawrence, C. Lyal, A. Newton, et al. 2011. Family-Group Names In Coleoptera (Insecta). ZooKeys 88: 1-972.
Butchart, S. H., M. Walpole, B. Collen, A. van Strien, J.P. Scharlemann, R.E. Almond, J.E.M. Baillie, et al. 2010. Global biodiversity: indicators of recent declines. Science 328(5982): 1164-1168.
Cameron, R.A.D., K.A. Triantis, C.E. Parent, F. Guilhaumon, M.R. Alonso, M. Ibáñez, A.M.F. Martins, et al. 2013. Snails on oceanic islands: testing the general dynamic model of oceanic island biogeography using linear mixed effect models. Journal of Biogeography, 40: 117-130.
Cardoso, P., N. Scharff, C. Gaspar, SS. Henriques, R. Carvalho, P.H. Castro, J.B. Schmidt, et al. 2008. Rapid biodiversity assessment of spiders (Araneae) using semi-quantitative sampling: a case study in a Mediterranean forest. Insect Conservation and Diversity 1: 71-84.
Cardoso, P., J.M. Lobo, S.C. Aranda, F. Dinis, C. Gaspar. & P.A.V. Borges 2009. A spatial scale assessment of habitat effects on arthropod communities of an oceanic island. Acta Oecologica 35: 590-597.
Cardoso, P., M.A. Arnedo, K.A. Triantis & P.A.V. Borges 2010. Drivers of diversity in Macaronesian spiders and the role of species extinctions. Journal of Biogeography 37: 1034-1046.
Cardoso, P., T.L. Erwin, P.A.V. Borges & T.R. New 2011. The seven impediments in invertebrate conservation and how to overcome them. Biological Conservation 144: 2647-2655.
Carroll, J.F., N. Tabanca, M. Kramer, N.M. Elejalde, D.E. Wedge, U.R. Bernier, S. Zhang, et al. 2011. Essential oils of Cupressus funebris, Juniperus communis, and J. chinensis (Cupressaceae) as repellents against ticks (Acari: Ixodidae) and mosquitoes (Diptera: Culicidae) and as toxicants against mosquitoes. Journal of Vector Ecology 36(2): 258-268.
Carvalho, J.C. & P. Cardoso 2014. Drivers of beta diversity in Macaronesian spiders in relation to dispersal ability. Journal of Biogeography 41: 1859-1870.
Carvalho, J.C., P. Cardoso, F. Rigal, K.A. Triantis & P.A.V. Borges 2015. Modelling directional spatio-temporal processes in island biogeography. Ecology and Evolution. In press, DOI: 10.1002/ece3.1632.
Case, T J. 1996. Global patterns in the establishment and distribution of exotic birds. Biological Conservation 78(1): 69-96.
Clarke, K.R. 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology 18: 117-117.

Biota associated with Juniperus brevifolia
45
Biota associated with Juniperus brevifolia
45
Colwell, R.K., 2009. EstimateS: statistical estimation of species richness and shared species from samples, Version 8.2. User’s guide and application published online at http://purl.oclc.org/estimates.
Connor, S.E., J.F.N. van Leeuwen, T.M. Rittenour, W.O. van der Knaap, B. Ammann & S. Björck 2012. The ecological impact of oceanic island colonization – a palaeoecological perspective from the Azores. Journal of Biogeography 39(6): 1007-1023.
Connor, S.E., W.O. van der Knaap, J.F.N. van Leeuwen & P. Kuneš 2013. Holocene palaeoclimate and palaeovegetation on the islands of Flores and Pico. pp: 149-162 in: Fernández-Palacios, J.M., L. Nascimento, J.C. Hernández,, S. Clemente, A. González & J.P. Díaz-González (Eds.) Climate change perspectives from the Atlantic: Past, present and Future. Servicio de Publicaciones, Universidad de La Laguna. Tenerife, Canárias.
Dansereu, P. 1970. Macaronesian studies IV. Natural ecosystems of the Azores. Revue Canadienne de Géographie 24(1): 21-42.
Dierssen, K. 2001. Distribution, ecological amplitude and phytosociological characterization of European bryophytes. Bryophytorum Bibliotheca, 56: 1-289.
Dormann, C.F., O. Schweiger, I. Augenstein, D. Bailey, R. Billeter, G. de Blust, R. DeFilippi, et al. 2007. Effects of landscape structure and land-use intensity on similarity of plant and animal communities. Global Ecology and Biogeography 16(6): 774-787.
Dray, S. & A.B. Dufour 2007. The ade4 package: implementing the duality diagram for ecologists. Journal of Statistical Software 22(4): 1-20.
Elias, R.B. 2007. Ecologia das florestas de Juniperus dos Açores. PhD thesis, University of the Azores. Angra do Heroísmo, Azores, Portugal. 179pp.
Elias, R.B. & L. Silva 2008. Juniperus brevifolia (Seub.) Antoine. pp. 182–183. in: Martín, J.L., M.J. Arechavaleta, P.A.V. Borges & B. Faria (Eds.). Top 100 Las cien especies amenazadas prioritarias de gestión en la región europea biogeográfica de la Macaronesia. Consejeria de Medio Ambiente y Ordenación Territorial, Gobierno de Canarias. 500 pp.
Elias, R.B. & E. Dias 2009. Gap dynamics and regeneration strategies in Juniperus-Laurus forests of the Azores islands. Plant Ecology 200: 179–189.
Elias, R.B., E. Dias & F. Pereira 2011. Disturbance, regeneration and the spatial pattern of tree species in Azorean mountain forests. Community Ecology 12: 23–30.
Elias, R.B. & E. Dias 2014. The recognition of infraspecific taxa in Juniperus brevifolia
(Cupressaceae). Phytotaxa 188(5): 241-250. Fernández-Palacios, J.M., L. De Nascimento, R. Otto,
J.D. Delgado, E. García-del-Rey, J.R. Arévalo, et al. 2011. A reconstruction of Palaeo-Macaronesia, with particular reference to the long-term biogeography of the Atlantic island laurel forests. Journal of Biogeography 38: 226-246.
Ferreira, R.C., R. Piredda, F. Bagnoli, R. Bellarosa, M. Attimonelli, S. Fineschi, B. Schirone & M.C. Simeone 2011. Phylogeography and conservation perspectives of an endangered macaronesian endemic: Picconia azorica (Tutin) Knobl. (Oleaceae). European Journal of Forest Research 130: 181-195.
Floren, A., & K.E. Linsenmair 1997. Diversity and recolonization dynamics of selected arthropod groups on different tree species in a lowland rainforest in Sabah, Malaysia with special reference to Formicidae. pp 344-381. in Stork, N.E., J. Adis & R.K. Didham (Eds.). Canopy arthropods. Chapman & Hall. London. 567pp.
Florencio, M., P. Cardoso, J.M. Lobo, E.B. Azevedo & P.A.V. Borges. 2013. Arthropod assemblage homogenization in oceanic islands: the role of exotic and indigenous species under landscape disturbance. Diversity and Distributions 19: 1450-1460.
França, Z., J.V. Cruz, J.C. Nunes & V-H. Forjaz 2003. Geologia dos Açores: uma perspectiva actual. Açoreana 10: 11–140.
Frutuoso, G. 1978. Sixth book of the land memories (1859) [Livro sexto das saudades da terra] 1589, In Portuguese]. Clutural Institute of Ponta Delgada, Azores. Portugal. 432 pp.
Frutuoso, G. 1981. Fourth book of the land memories (1859) [Livro sexto das saudades da terra 1589, In Portuguese]. Clutural Institute of Ponta Delgada, Azores. Vol. II. 397 pp.
Frutuoso, G. 1981. Fourth book of the land memories (1859) [Livro sexto das saudades da terra 1589, In Portuguese]. Clutural Institute of Ponta Delgada, Azores. Vol. III.
Gabriel, R. 2000. Ecophysiology of Azorean forest bryophytes. Ph. D. thesis. Imperial College of Science, Technology and Medicine, London University. Silwood Park, Ascot, Berkshire. 310 pp.
Gabriel, R., & .W. Bates 2005. Bryophyte community composition and habitat specificity in the natural forests of Terceira, Azores. Plant Ecology 177(1): 125-144.
Gabriel, R., N. Homem, A. Couto, S.C. Aranda & P.A.V. Borges 2011. Azorean bryophytes: a preliminary review of rarity patterns. Açoreana 7: 149-206.

Nunes et al.
46
Biota associated with Juniperus brevifolia
47
island streams: the case of the Azores Archipelago. Fundamental and Applied Limnology / Archiv für Hydrobiologie 182(4): 337-350.
Ribeiro, S.P. & P.A.V. Borges 2010. Canopy habitat area effect on the arthropod species densities in the Azores: pondering the contribution of tourist species and other life histories. pp. 89-114 in Serrano, A.R.M., P.A.V. Borges, M. Boieiro & P. Oromí (Eds.). Terrestrial arthropods of Macaronesia – Biodiversity, Ecology and Evolution. Sociedade Portuguesa de Entomologia, Lisboa.
Ribeiro, S.P., P.A.V. Borges & C.S. Gaspar 2003. Ecology and evolution of the arborescent Erica azorica Hochst (Ericaceae). Arquipélago - Agricultural and Environmental Science 1: 41-50.
Ribeiro, S.P., P.A.V. Borges,, C. Gaspar, C. Melo, A.R.M. Serrano, J. Amaral, C. Aguiar, et al. 2005. Canopy insect herbivores in the Azorean Laurisilva forests: key host plant species in a highly generalist insect community. Ecography 28: 315-330.
Rodrigues P, R.J. Lopes, S.V. Drovetski, S. Reis, J.A. Ramos & R.T. Cunha 2013. Phylogeography and genetic diversity of the Robin (Erithacus rubecula) in the Azores islands. Journal of Ornithology 154(4): 889-900.
Rumeu, B., J. Caujapé-Castells, J-L. Blanco-Pastor, R. Jaén-Molina, M. Nogales, R.B. Elias & P. Vargas 2011. The colonization history of Juniperus brevifolia (Cupressaceae) in the Azores islands. PLoSOne 6: e27697. Available from: http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0027697
Santos, A.M.C., R.J. Whittaker, K.A. Triantis, P.A.V. Borges., O.R. Jones, D. Quicke & J. Hortal 2010. Are species-area relationships from entire archipelagos congruent with those of their constituent islands? Global Ecology and Biogeography 19: 527-540
Schäfer, H. 2005. Flora of the Azores – a field guide (2nd edition). Margraf Publishers, Weikersheim. 346 pp.
Schäfer, H., M. Moura, M.G.B. Maciel, L. Silva, F.J. Rumsey & M.A. Carine 2011. The Linnean shortfall in oceanic island biogeography: a case study in the Azores. Journal of Biogeography 38: 1345-1355.
Scharff, N., J.A. Coddington, C.R. Griswold, G. Hormiga & P.D.P. Bjørn 2003. When to quit? Estimating spider species richness in a northern European deciduous forest. Journal of Arachnology 31(2): 246-273.
Silva, L., G. Markin & J. Tavares 1995. Argyresthia atlanticella Rebel (Insecta, Lepidoptera) an excluded agent for Myrica faya Aiton (Myricaceae)
biocontrol. Arquipelago. Life and Marine Sciences 13A: 105-113.
Silva, L., R.B. Elias, M. Moura, H. Meimberg & E. Dias 2011. Genetic variability and differentiation among populations of the Azorean endemic gymnosperm Juniperus brevifolia: Baseline information for a conservation and restoration perspective. Biochemical Genetics 49: 715–734.
Silva, M.H.B.S.M. 2000. Some approaches to safeguard selected endangered endemic plants of the Azores. Ph.D. Thesis. Ghent University. Ghent, Belgium.
Silveira, L. 2013. Aprender com a História – Modos de Interacção com a Natureza na Ilha Terceira (Do Povoamento ao seculo XX). Cascais: Princípia Editora. 357 pp. ISBN: 978-989-716-042-4
Sjögren, E. 1978. Bryophyte vegetation in the Azores Islands. Memórias da Sociedade Broteriana Volume 26. Instituto Botânico da Universidade de Coimbra. 283pp.
Stork, N.E. 1987. Guild structure of arthropods from Bornean rain forest trees. Ecological Entomology 12(1): 69-80.
Stork, N.E. 1991. The composition of the arthropod fauna of Bornean lowland rain forest trees. Journal of Tropical Ecology 7(02) 161-180.
Summerville, K.S., M.J. Boulware, J.A. Veech & T.O. Crist 2003. Spatial variation in species diversity and composition of forest Lepidoptera in eastern deciduous forests of North America. Conservation Biology 17(4) 1045-1057.
Terzopoulou, S., F. Rigal, R.J. Whittaker, P.A.V. Borges & K.A. Triantis 2015. Drivers of extinction: the case of Azorean beetles. Biology Letters 11(6): 20150273.
Thomas, C.F.G., P. Brain & P.C. Jepson 2003. Aerial activity of linyphiid spiders: modelling dispersal distances from meteorology and behaviour. Journal of Applied Ecology 40(5): 912-927.
Triantis, K.A., P.A.V. Borges, J. Hortal & R.J. Whittaker 2010a. The Macaronesian province: patterns of species richness and endemism of arthropods. Pp. 45-64. in Serrano, A.R.M., P.A.V. Borges, M. Boieiro & P. Oromí (Eds.). Terrestrial arthropods of Macaronesia – Biodiversity, Ecology and Evolution. Portuguese Society of Entomology, Lisbon.
Triantis, K.A., P.A.V. Borges, R.J. Ladle, J. Hortal, P. Cardoso, C. Gaspar, F. Dinis, et al. 2010b. Extinction debt on oceanic islands. Ecography 33: 285-294.
Vieira, V., L. Silva, S. Dupont, R. Martins & D. Pombo, 1993. Lepidoptera and natural enemies of the Myrica faya of the São Jorge Island [Lepidópteros e Inimigos naturais de Myrica faya
Nunes et al.
46
Gaspar, C., P.A.V. Borges & K.J. Gaston 2008. Diversity and distribution of arthropods in native forests of the Azores archipelago. Arquipélago - Life and marine Sciences 25: 1-30.
Gaspar, C., K.J. Gaston, P.A.V. Borges & P. Cardoso 2011. Selection of priority areas for arthropod conservation in the Azores archipelago. Journal of Insect Conservation 15: 671–684.
Gaspar, C., P. Cardoso, P.A.V. Borges & K.J. Gaston 2014. Efficiency of sampling methods and effort to assess arthropod diversity in Azorean native forests. Arquipélago - Life and Marine Sciences 31: 21-36.
Gaston, K. J. 1994. Rarity. Springer Netherlands. 205 pp.
Gray, J.S., A. Bjoergesaeter & K.I. Ugland 2006. On plotting species abundance distributions. Journal of Animal Ecology 75: 752-756.
Grimbacher, P.S. & N.E. Stork 2007. Vertical stratification of feeding guilds and body size in beetle assemblages from an Australian tropical rainforest. Austral Ecology 32(1): 77-85.
Guilbert, E. 1997. Arthropod biodiversity in the canopy of New Caledonian forests. pp 265-277. in Stork, N.E., J. Adis & R.K. Didham (Eds.). Canopy arthropods. Chapman & Hall. London. 567pp.
Homem, N.F.M. 2005. Biodiversity of Briophytes in the natural forests of the Azores – Island Terceira and Pico [Biodiversidade de briófitos nas florestas naturais dos Açores – Ilhas Terceira e Pico; in Portuguese] MSc in Nature Management and Conservation. University of the Azores, Department of Agrarian Sciences, Angra do Heroísmo. 137pp.
Hooper, D.U., E.C. Adair, J.B. Cardinale, J.E. Byrnes, B.A. Hungate, K.L.Matulich, A. Gonzalez, et al. 2012. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486(7401): 105-108.
Kitching, R.L., H. Mitchell, G. Morse & C. Thebaud 1997. Determinants of species richness in assemblages of canopy arthropods in rainforests. pp131-150, in Stork, N.E., J. Adis & R.K. Didham (Eds.). Canopy arthropods. Chapman & Hall. London. 567pp.
Kruskal, J.B. 1964. Nonmetric multidimensional scaling: a numerical method. Psychometrika 29(2): 115-129.
Krüger, O. & C.G. McGavin 2001. Predator–prey ratio and guild constancy in a tropical insect community. Journal of Zoology 253(2): 265-273.
Kuria, S.K., M.H. Villet, T.M. Palmer & M.L. Stanton 2010. A comparison of two sampling methods for surveying mammalian herbivore impacts on beetle communities in the canopy of Acacia
drepanolobium in Kenya. African Entomology 18(1): 87-98.
MacArthur, R.H. & E.O. Wilson 1967. The theory of island biogeography (Vol. 1). Princeton University Press. 224 pp.
Marcelino, J.A.P., L. Silva, P. Garcia, E. Webber, & A.O. Soares 2013. Using species spectra to evaluate plant community conservation value along a gradient of anthropogenic disturbance. Environmental Monitoring and Assessement 185: 6221-6233.
Markin, J., L. Silva & F. Aguiar 1995. The insect fauna associated with the tree Myrica faya (Myricaceae) in macaronesian islands and on mainland Portugal. Boletim do Museu Municipal do Funchal 4: 411-420.
Matthews, T.J., M.K. Borregaard, K. Ugland, P.A.V. Borges, F. Rigal, P. Cardoso & R.J. Whittaker 2014. The gambin model provides a superior fit to species abundance distributions with a single free parameter: evidence, implementation and interpretation. Ecography 37: 1002–1011.
Meijer, S.S., R.J. Whittaker & P.A.V. Borges 2011. The effects of land-use change on arthropod richness and abundance on Santa Maria Island (Azores): unmanaged plantations favour endemic beetles. Journal of Insect Conservation 15: 505-522.
Moran, V.C. & T.R.E. Southwood 1982. The guild composition of arthropod communities in trees. The Journal of Animal Ecology 289-306.
Ogle, D. 2015. NCStats V. 0.4.4. - Functions to support learning in Northland College statistics courses. https://rforge.net/NCStats/index.html
Oksanen, J., F.G. Blanchet, R. Kindt, P. Legendre, P.R. Minchin, R.B. O'Hara, G.L. Simpson, et al. 2015. vegan: Community Ecology Package. R package version 2.2-1. http://CRAN.R-project.org/package=vegan
Platnick, N.I. 2014. The world spider catalog, version 15. American Museum of Natural History, online at http://research.amnh.org/entomology/spiders/catalog/index.html DOI: 10.5531/db.iz.0001.
R Core Team 2015. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/.
Rando J.C., J.A. Alcover, S.L. Olson & H. Pieper 2013. A new species of extinct scops owl (Aves: Strigiformes: Strigidae: Otus) from São Miguel island (Azores archipelago, north Atlantic ocean). Zootaxa 3647: 343–357.
Raposeiro P.M., S.H. Hughes & A.C. Costa 2013. Environmental drivers - spatial and temporal variation of macroinvertebrates communities in

Biota associated with Juniperus brevifolia
47
Biota associated with Juniperus brevifolia
47
island streams: the case of the Azores Archipelago. Fundamental and Applied Limnology / Archiv für Hydrobiologie 182(4): 337-350.
Ribeiro, S.P. & P.A.V. Borges 2010. Canopy habitat area effect on the arthropod species densities in the Azores: pondering the contribution of tourist species and other life histories. pp. 89-114 in Serrano, A.R.M., P.A.V. Borges, M. Boieiro & P. Oromí (Eds.). Terrestrial arthropods of Macaronesia – Biodiversity, Ecology and Evolution. Sociedade Portuguesa de Entomologia, Lisboa.
Ribeiro, S.P., P.A.V. Borges & C.S. Gaspar 2003. Ecology and evolution of the arborescent Erica azorica Hochst (Ericaceae). Arquipélago - Agricultural and Environmental Science 1: 41-50.
Ribeiro, S.P., P.A.V. Borges,, C. Gaspar, C. Melo, A.R.M. Serrano, J. Amaral, C. Aguiar, et al. 2005. Canopy insect herbivores in the Azorean Laurisilva forests: key host plant species in a highly generalist insect community. Ecography 28: 315-330.
Rodrigues P, R.J. Lopes, S.V. Drovetski, S. Reis, J.A. Ramos & R.T. Cunha 2013. Phylogeography and genetic diversity of the Robin (Erithacus rubecula) in the Azores islands. Journal of Ornithology 154(4): 889-900.
Rumeu, B., J. Caujapé-Castells, J-L. Blanco-Pastor, R. Jaén-Molina, M. Nogales, R.B. Elias & P. Vargas 2011. The colonization history of Juniperus brevifolia (Cupressaceae) in the Azores islands. PLoSOne 6: e27697. Available from: http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0027697
Santos, A.M.C., R.J. Whittaker, K.A. Triantis, P.A.V. Borges., O.R. Jones, D. Quicke & J. Hortal 2010. Are species-area relationships from entire archipelagos congruent with those of their constituent islands? Global Ecology and Biogeography 19: 527-540
Schäfer, H. 2005. Flora of the Azores – a field guide (2nd edition). Margraf Publishers, Weikersheim. 346 pp.
Schäfer, H., M. Moura, M.G.B. Maciel, L. Silva, F.J. Rumsey & M.A. Carine 2011. The Linnean shortfall in oceanic island biogeography: a case study in the Azores. Journal of Biogeography 38: 1345-1355.
Scharff, N., J.A. Coddington, C.R. Griswold, G. Hormiga & P.D.P. Bjørn 2003. When to quit? Estimating spider species richness in a northern European deciduous forest. Journal of Arachnology 31(2): 246-273.
Silva, L., G. Markin & J. Tavares 1995. Argyresthia atlanticella Rebel (Insecta, Lepidoptera) an excluded agent for Myrica faya Aiton (Myricaceae)
biocontrol. Arquipelago. Life and Marine Sciences 13A: 105-113.
Silva, L., R.B. Elias, M. Moura, H. Meimberg & E. Dias 2011. Genetic variability and differentiation among populations of the Azorean endemic gymnosperm Juniperus brevifolia: Baseline information for a conservation and restoration perspective. Biochemical Genetics 49: 715–734.
Silva, M.H.B.S.M. 2000. Some approaches to safeguard selected endangered endemic plants of the Azores. Ph.D. Thesis. Ghent University. Ghent, Belgium.
Silveira, L. 2013. Aprender com a História – Modos de Interacção com a Natureza na Ilha Terceira (Do Povoamento ao seculo XX). Cascais: Princípia Editora. 357 pp. ISBN: 978-989-716-042-4
Sjögren, E. 1978. Bryophyte vegetation in the Azores Islands. Memórias da Sociedade Broteriana Volume 26. Instituto Botânico da Universidade de Coimbra. 283pp.
Stork, N.E. 1987. Guild structure of arthropods from Bornean rain forest trees. Ecological Entomology 12(1): 69-80.
Stork, N.E. 1991. The composition of the arthropod fauna of Bornean lowland rain forest trees. Journal of Tropical Ecology 7(02) 161-180.
Summerville, K.S., M.J. Boulware, J.A. Veech & T.O. Crist 2003. Spatial variation in species diversity and composition of forest Lepidoptera in eastern deciduous forests of North America. Conservation Biology 17(4) 1045-1057.
Terzopoulou, S., F. Rigal, R.J. Whittaker, P.A.V. Borges & K.A. Triantis 2015. Drivers of extinction: the case of Azorean beetles. Biology Letters 11(6): 20150273.
Thomas, C.F.G., P. Brain & P.C. Jepson 2003. Aerial activity of linyphiid spiders: modelling dispersal distances from meteorology and behaviour. Journal of Applied Ecology 40(5): 912-927.
Triantis, K.A., P.A.V. Borges, J. Hortal & R.J. Whittaker 2010a. The Macaronesian province: patterns of species richness and endemism of arthropods. Pp. 45-64. in Serrano, A.R.M., P.A.V. Borges, M. Boieiro & P. Oromí (Eds.). Terrestrial arthropods of Macaronesia – Biodiversity, Ecology and Evolution. Portuguese Society of Entomology, Lisbon.
Triantis, K.A., P.A.V. Borges, R.J. Ladle, J. Hortal, P. Cardoso, C. Gaspar, F. Dinis, et al. 2010b. Extinction debt on oceanic islands. Ecography 33: 285-294.
Vieira, V., L. Silva, S. Dupont, R. Martins & D. Pombo, 1993. Lepidoptera and natural enemies of the Myrica faya of the São Jorge Island [Lepidópteros e Inimigos naturais de Myrica faya

Nunes et al.
48
APPE
NDIX
I Ta
ble 1
. List
of ar
thro
pod s
pecie
s asso
ciated
with
Junip
eus b
revif
olia i
n the
Azo
res. T
he cl
assif
icatio
n sys
tem fo
llows
the g
enera
l gui
delin
es pr
esen
ted in
Bor
ges e
t al. (
2010
a), w
ith th
e high
er tax
a list
ed in
a ph
yloge
netic
sequ
ence
, fro
m th
e les
s deri
ved t
o mor
e deri
ved g
roup
s. Th
e fam
ilies
, gen
era an
d spe
cies a
re lis
ted by
alph
abeti
cal o
rder.
Exc
eptio
ns to
this
are t
he A
rachn
idae
, who
fo
llow
the cl
assif
icatio
n of t
he “W
orld
Spide
r Cata
logue
” by P
latnic
k (20
14);
Coleo
ptera,
that
follo
w Bo
ucha
rd et
al. (
2011
) and
Lep
idopt
era, th
at fo
llow
Agui
ar &
Karsh
olt (
2006
). Th
e gen
era
and s
pecie
s are
neve
rthele
ss als
o list
ed by
alph
abeti
cal o
rder.
Spe
cies t
hat w
ere no
t ide
ntifie
d to a
t lea
st ge
nus l
evel
were
remov
ed fr
om th
is lis
t. Th
e col
onisa
tion s
tatus
of ea
ch sp
ecies
is
pres
ented
in th
e 5th co
lumn
of th
e list
as fo
llows
: E –
ende
mic;
N – n
ative
; I –
intro
duce
d. Th
e fun
ction
al gr
oup i
s give
n the
6th colum
n as f
ollo
ws: P
– pr
edato
r; H
– herb
ivore;
S –
sapr
ophy
te; F
– f
ungiv
ore;
Ex –
Exter
nal d
igesti
on an
d suc
king;
Ch –
Chew
ing a
nd cu
tting
; Pi -
Pier
cing a
nd su
cking
; Si -
Siph
oning
; No –
Not
feed
ing.
Flo,
Flor
es; F
ai, Fa
ial; P
ic, P
ico; S
Jo, S
ão Jo
rge;
Ter,
Terc
eira,
SMi, S
ão M
iguel.
Cl
ass
Orde
r Fa
mily
Sp
ecies
Co
lonisa
tion
St
atus
Fe
edin
g M
ode
Num
ber o
f In
divi
duals
Flo
Fai
Pic
Sjo
Ter
SMi
Arac
hnida
Ps
eudo
scor
pion
es
Neob
isiida
e Ne
obisi
um m
aroc
canu
m Be
ier
I P-
Ch
13
X X
Opili
ones
Ph
alang
iidae
Le
iobun
um bl
ackw
alli M
eade
N
P-Ch
14
X
X
X X
Arac
hnida
Ar
anea
e M
imeti
dae
Ero f
urca
ta (V
illers
) I
P-Ex
24
X X
X
Oeco
biida
e Oe
cobiu
s nav
us B
lackw
all
I P-
Ex
2
X X
Theri
diida
e Cr
yptac
haea
blatt
ea (U
rquh
art)
I P-
Ex
5
X
La
saeo
la oc
eanic
a Sim
on
E
P-Ex
99
X
X X
X X
Rh
omph
aea n
asica
(Sim
on)
I P-
Ex
2
X
Ru
gatho
des
acor
eens
is W
unde
rlich
E
P-
Ex
970
X
X X
X X
X
Steato
da gr
ossa
(C.L
. Koc
h)
I P-
Ex
11
X
X
Ther
idion
mus
ivivu
m Sc
hmidt
N
P-Ex
28
X
Liny
phiid
ae
Acor
igone
acor
eens
is (W
unde
rlich
) E
P-
Ex
167
X
X X
X X
X
Cana
ripha
ntes a
core
ensis
(Wun
derli
ch)
E
P-Ex
1
X
Erigo
ne at
ra B
lackw
all
I P-
Ex
31
X X
X X
X
Erigo
ne au
tumna
lis E
merto
n I
P-Ex
1
X
Mer
messu
s bry
antae
(Ivie
& B
arrow
s) I
P-Ex
1
X
Mer
messu
s tril
obatu
s (Em
erton
) I
P-Ex
1
X
Micr
oliny
phia
johns
oni (
Blac
kwall
) N
P-Ex
5
X
Mini
cia fl
ores
ensis
Wun
derli
ch
E
P-Ex
10
6
X
X X
X
Oe
dotho
rax f
uscu
s (Bl
ackw
all)
I P-
Ex
72
X
X X
X X
X
Palli
duph
antes
schm
itzi (
Kulcz
ynsk
i) N
P-Ex
4
X
Savig
niorrh
ipis a
core
ensis
Wun
derli
ch
E
P-Ex
57
58
X
X X
X X
X
Tenu
iphan
tes m
iguele
nsis
Wun
derli
ch
N P-
Ex
8
X
X
Te
nuiph
antes
tenu
is (B
lackw
all)
I P-
Ex
66
X
X X
X X
Walc
kena
eria
gran
dis (
Wun
derli
ch)
E
P-Ex
12
X
Nunes et al.
48
da Ilha de São Jorge]. Reports and Comunications of the Biology Department 21: 28:34. [Portuguese]
Wagner, T. 1997. The beetle fauna of different tree species in forests of Rwanda and East Zaire. Pp:169-183. in Stork, N.E., J. Adis & R.K. Didham (Eds.). Canopy arthropods. Chapman & Hall. London. 567pp.
Watson, L. & M.J. Dallwitz 2003. British insects: the families of Coleoptera. Version: 25th July 2012. http://delta-intkey.com
Whittaker, R.J., K.A. Triantis & R.J. Ladle 2008. A general dynamic theory of oceanic island biogeography. Journal of Biogeography 35(6): 977-994.
Whittaker, R., F. Rigal, P.A.V. Borges, P. Cardoso, S. Terzopoulou, F. Casanoves, L. Pla. et al. 2014. Functional biogeography of oceanic islands and the scaling of functional diversity in the Azores. PNAS - Proceedings of the National Academy of Sciences 111: 13709–13714.
Winchester, N.N. 1997. Canopy arthropods of coastal Sitka spruce trees on Vancouver Island, British Columbia, Canada. Pp: 151-168. in Stork, N.E., J.Adis & R.K. Didham (Eds). Canopy arthropods.Chapman & Hall. London. 567pp.
Received 9 Nov 2015. Accepted 30 Nov 2015,Published online 30 Dec 2015.

Biota associated with Juniperus brevifolia
49
APPE
NDIX
I Ta
ble 1
. List
of ar
thro
pod s
pecie
s asso
ciated
with
Junip
eus b
revif
olia i
n the
Azo
res. T
he cl
assif
icatio
n sys
tem fo
llows
the g
enera
l gui
delin
es pr
esen
ted in
Bor
ges e
t al. (
2010
a), w
ith th
e high
er tax
a list
ed in
a ph
yloge
netic
sequ
ence
, fro
m th
e les
s deri
ved t
o mor
e deri
ved g
roup
s. Th
e fam
ilies
, gen
era an
d spe
cies a
re lis
ted by
alph
abeti
cal o
rder.
Exc
eptio
ns to
this
are t
he A
rachn
idae
, who
fo
llow
the cl
assif
icatio
n of t
he “W
orld
Spide
r Cata
logue
” by P
latnic
k (20
14);
Coleo
ptera,
that
follo
w Bo
ucha
rd et
al. (
2011
) and
Lep
idopt
era, th
at fo
llow
Agui
ar &
Karsh
olt (
2006
). Th
e gen
era
and s
pecie
s are
neve
rthele
ss als
o list
ed by
alph
abeti
cal o
rder.
Spe
cies t
hat w
ere no
t ide
ntifie
d to a
t lea
st ge
nus l
evel
were
remov
ed fr
om th
is lis
t. Th
e col
onisa
tion s
tatus
of ea
ch sp
ecies
is
pres
ented
in th
e 5th co
lumn
of th
e list
as fo
llows
: E –
ende
mic;
N – n
ative
; I –
intro
duce
d. Th
e fun
ction
al gr
oup i
s give
n the
6th colum
n as f
ollo
ws: P
– pr
edato
r; H
– herb
ivore;
S –
sapr
ophy
te; F
– f
ungiv
ore;
Ex –
Exter
nal d
igesti
on an
d suc
king;
Ch –
Chew
ing a
nd cu
tting
; Pi -
Pier
cing a
nd su
cking
; Si -
Siph
oning
; No –
Not
feed
ing.
Flo,
Flor
es; F
ai, Fa
ial; P
ic, P
ico; S
Jo, S
ão Jo
rge;
Ter,
Terc
eira,
SMi, S
ão M
iguel.
Cl
ass
Orde
r Fa
mily
Sp
ecies
Co
lonisa
tion
St
atus
Fe
edin
g M
ode
Num
ber o
f In
divi
duals
Flo
Fai
Pic
Sjo
Ter
SMi
Arac
hnida
Ps
eudo
scor
pion
es
Neob
isiida
e Ne
obisi
um m
aroc
canu
m Be
ier
I P-
Ch
13
X X
Opili
ones
Ph
alang
iidae
Le
iobun
um bl
ackw
alli M
eade
N
P-Ch
14
X
X
X X
Arac
hnida
Ar
anea
e M
imeti
dae
Ero f
urca
ta (V
illers
) I
P-Ex
24
X X
X
Oeco
biida
e Oe
cobiu
s nav
us B
lackw
all
I P-
Ex
2
X X
Theri
diida
e Cr
yptac
haea
blatt
ea (U
rquh
art)
I P-
Ex
5
X
La
saeo
la oc
eanic
a Sim
on
E
P-Ex
99
X
X X
X X
Rh
omph
aea n
asica
(Sim
on)
I P-
Ex
2
X
Ru
gatho
des
acor
eens
is W
unde
rlich
E
P-
Ex
970
X
X X
X X
X
Steato
da gr
ossa
(C.L
. Koc
h)
I P-
Ex
11
X
X
Ther
idion
mus
ivivu
m Sc
hmidt
N
P-Ex
28
X
Liny
phiid
ae
Acor
igone
acor
eens
is (W
unde
rlich
) E
P-
Ex
167
X
X X
X X
X
Cana
ripha
ntes a
core
ensis
(Wun
derli
ch)
E
P-Ex
1
X
Erigo
ne at
ra B
lackw
all
I P-
Ex
31
X X
X X
X
Erigo
ne au
tumna
lis E
merto
n I
P-Ex
1
X
Mer
messu
s bry
antae
(Ivie
& B
arrow
s) I
P-Ex
1
X
Mer
messu
s tril
obatu
s (Em
erton
) I
P-Ex
1
X
Micr
oliny
phia
johns
oni (
Blac
kwall
) N
P-Ex
5
X
Mini
cia fl
ores
ensis
Wun
derli
ch
E
P-Ex
10
6
X
X X
X
Oe
dotho
rax f
uscu
s (Bl
ackw
all)
I P-
Ex
72
X
X X
X X
X
Palli
duph
antes
schm
itzi (
Kulcz
ynsk
i) N
P-Ex
4
X
Savig
niorrh
ipis a
core
ensis
Wun
derli
ch
E
P-Ex
57
58
X
X X
X X
X
Tenu
iphan
tes m
iguele
nsis
Wun
derli
ch
N P-
Ex
8
X
X
Te
nuiph
antes
tenu
is (B
lackw
all)
I P-
Ex
66
X
X X
X X
Walc
kena
eria
gran
dis (
Wun
derli
ch)
E
P-Ex
12
X

Nunes et al.
50
Clas
s Or
der
Fam
ily
Spec
ies
Colon
isatio
n
Stat
us
Feed
ing
Mod
e Nu
mbe
r of
Indi
vidu
als
Flo
Fai
Pic
Sjo
Ter
SMi
Trich
opso
cidae
Tr
ichop
socu
s clar
us (B
anks
) N
S-Ch
82
X
X X
X X
Trog
iidae
Ce
roba
sis cf
sp.1
E
S-Ch
55
7
X
X
X
Ce
roba
sis sp
.3 E
S-
Ch
542
X
Hemi
ptera
Ci
cade
llida
e Ap
hrod
es ha
milto
ni Qu
artau
& B
orge
s E
H-
Pi
1
X
Eu
pter
yx az
orica
Riba
ut E
H-
Pi
1
X
Cixii
dae
Cixiu
s azo
flore
si Re
mane
& A
sche
E
H-
Pi
752
X
Cixiu
s azo
pifajo
azofa
Rem
ane &
Asc
he
E
H-Pi
16
4
X
Ci
xius a
zopif
ajo az
ojo R
eman
e & A
sche
E
H-
Pi
239
X
Cixiu
s azo
pifajo
azop
ifajo
Rema
ne &
Asc
he
E
H-Pi
52
3
X
Ci
xius a
zoric
us az
oricu
s Lin
dberg
E
H-
Pi
3
X X
Ci
xius a
zoter
ceira
e Rem
ane &
Asc
he
E
H-Pi
81
5
X
Ci
xius i
nsula
ris L
indb
erg
E
H-Pi
12
32
X
De
lphac
idae
Meg
amelo
des q
uadr
imac
ulatus
(Sign
oret)
N
H-Pi
2
X
X
Mue
lleria
nella
sp.1
N H-
Pi
1
X
M
uelle
riane
lla sp
.3 N
H-Pi
1
X
Fl
atida
e Cy
phop
terum
adce
nden
s (He
rr.-S
chaff
.) N
H-Pi
80
4
X X
X X
X X
Anth
ocor
idae
Brac
hyste
les pa
rvico
rnis
(A. C
osta)
N
P-Pi
5
X
X
X
Ly
gaeid
ae
Kleid
ocer
ys er
icae (
Horv
áth)
N H-
Pi
33
X
X X
X X
X
M
icrop
hysid
ae
Loric
ula co
leoptr
ata (F
allén
) N
P-Pi
2
X
M
irida
e He
teroto
ma pl
anico
rnis
(Pall
as)
N P-
Pi
3
X
M
onalo
coris
filic
is (L
inna
eus)
N H-
Pi
6
X
X
X
Pi
nalit
us or
omii
J. Ri
bes
E
H-Pi
83
9
X X
X X
X X
Po
lymer
us co
gnatu
s (Fi
eber)
N
H-Pi
1
X
Na
bidae
Na
bis p
seud
oferu
s ibe
ricus
Rem
ane
N P-
Pi
1
X
Redu
viida
e Em
picor
is ru
brom
acula
tus (
Blac
kbur
n)
I P-
Pi
1
X
Psyll
idae
St
roph
ingia
harte
ni Ho
dkin
son
E
H-Pi
64
X
X X
X X
Trioz
idae
Tr
ioza
(Lau
ritrio
za) l
auris
ilvae
Hod
kinso
n N
H-Pi
21
0
X X
X X
X X
Aphi
didae
Am
phor
opho
ra ru
bi (K
alten
bach
) sen
su
latior
e N
H-Pi
1
X
Aphi
s cra
ccivo
ra K
och
N H-
Pi
1
X
Ap
his s
p.1
N H-
Pi
1
X
Rh
opalo
siphu
m ru
fiabd
omina
lis (S
asak
i) I
H-Pi
1
X
Toxo
ptera
aura
ntii (
Boye
r de F
onsc
olomb
e) I
H-Pi
4
X
Clas
s Or
der
Fam
ily
Spec
ies
Colon
isatio
n
Stat
us
Feed
ing
Mod
e Nu
mbe
r of
Indi
vidu
als
Flo
Fai
Pic
Sjo
Ter
SMi
Tetra
gnath
idae
Mete
llina
mer
ianae
(Sco
poli)
I
P-Ex
5
X
Sanc
us ac
oree
nsis
(Wun
derli
ch)
E
P-Ex
45
1
X X
X X
X X
Aran
eidae
Gi
bbar
anea
occid
ental
is W
unde
rlich
E
P-
Ex
706
X
X X
X X
X
Man
gora
acaly
pha (
Walc
kena
er)
I P-
Ex
2
X
X
Lyco
sidae
Pa
rdos
a aco
rens
is Si
mon
E
P-Ex
14
X
X
X X
Pisa
urida
e Pi
saur
a aco
reen
sis W
unde
rlich
E
P-
Ex
34
X
X
X X
X
Di
ctynid
ae
Embly
na ac
oree
nsis
Wun
derli
ch
E
P-Ex
30
X
X
La
thys d
entic
helis
(Sim
on)
N P-
Ex
4089
X X
X X
X X
Ni
gma p
uella
(Sim
on)
I P-
Ex
30
X
Cl
ubion
idae
Cheir
acan
thium
erra
ticum
(Walc
kena
er)
I P-
Ex
14
X
X
Cheir
acan
thium
flor
esen
se W
unde
rlich
E
P-
Ex
3
X
Ch
eirac
anthi
um jo
rgee
nse W
unde
rlich
E
P-
Ex
1
X
Cl
ubion
a dec
ora B
lackw
all
N P-
Ex
106
X X
X X
X
Club
iona t
erre
stris
Wes
tring
I
P-Ex
7
X
Th
omisi
dae
Xysti
cus c
or C
anes
trini
N P-
Ex
634
X
X X
X X
X
Sa
lticid
ae
Mac
aroe
ris ca
ta (B
lackw
all)
N P-
Ex
115
X
X X
X X
X
Neon
acor
eens
is W
unde
rlich
E
P-
Ex
3
X
X
Ps
eude
uoph
rys v
afra (
Blac
kwall
) I
P-Ex
1
X Di
plopo
da
Julid
a Ju
lidae
Om
matoi
ulus m
orele
tii (L
ucas
) I
H-Ch
17
7
X X
X X
X X
Chilo
poda
Li
thob
iomor
pha
Lith
obiid
ae
Litho
bius p
ilico
rnis
pilico
rnis
Newp
ort
N P-
Ch
1
X
Inse
cta
Micr
ocor
yphi
a M
achil
idae
Dilta
saxic
ola (W
omers
ley)
N S-
Ch
62
X
X X
X
Trigo
nioph
thalm
us bo
rges
i Men
des e
t al.
E
S-Ch
60
X
X
Bl
attod
ea
Polyp
hagid
ae
Zetha
vesti
ta (B
rullé
) N
S-Ch
15
09
X
X X
X X
X
Psoc
opter
a Ca
ecili
usida
e Va
lenzu
ela bu
rmeis
teri (
Brau
er)
N S-
Ch
40
X
X
X
Valen
zuela
flav
idus (
Step
hens
) N
S-Ch
19
X X
X X
X
Ectop
socid
ae
Ectop
socu
s brig
gsi M
cLac
hlan
I S-
Ch
165
X
X X
X X
X
Ectop
socu
s stra
uchi
End
erlein
N
S-Ch
2
X
X
El
ipso
cidae
El
ipsoc
us az
oricu
s Mein
ande
r E
S-
Ch
57
X
X X
X X
Elips
ocus
brinc
ki Ba
donn
el E
S-
Ch
778
X
X X
X X
Pe
ripso
cidae
Pe
ripso
cus p
haeo
pteru
s (St
ephe
ns)
N S-
Ch
4
X
Pe
ripso
cus s
ubfas
ciatu
s (Ra
mbur
) N
S-Ch
1
X
Ps
ocida
e At
lantop
socu
s adu
stus (
Hage
n)
N S-
Ch
3
X
X

Biota associated with Juniperus brevifolia
51
Clas
s Or
der
Fam
ily
Spec
ies
Colon
isatio
n
Stat
us
Feed
ing
Mod
e Nu
mbe
r of
Indi
vidu
als
Flo
Fai
Pic
Sjo
Ter
SMi
Trich
opso
cidae
Tr
ichop
socu
s clar
us (B
anks
) N
S-Ch
82
X
X X
X X
Trog
iidae
Ce
roba
sis cf
sp.1
E
S-Ch
55
7
X
X
X
Ce
roba
sis sp
.3 E
S-
Ch
542
X
Hemi
ptera
Ci
cade
llida
e Ap
hrod
es ha
milto
ni Qu
artau
& B
orge
s E
H-
Pi
1
X
Eu
pter
yx az
orica
Riba
ut E
H-
Pi
1
X
Cixii
dae
Cixiu
s azo
flore
si Re
mane
& A
sche
E
H-
Pi
752
X
Cixiu
s azo
pifajo
azofa
Rem
ane &
Asc
he
E
H-Pi
16
4
X
Ci
xius a
zopif
ajo az
ojo R
eman
e & A
sche
E
H-
Pi
239
X
Cixiu
s azo
pifajo
azop
ifajo
Rema
ne &
Asc
he
E
H-Pi
52
3
X
Ci
xius a
zoric
us az
oricu
s Lin
dberg
E
H-
Pi
3
X X
Ci
xius a
zoter
ceira
e Rem
ane &
Asc
he
E
H-Pi
81
5
X
Ci
xius i
nsula
ris L
indb
erg
E
H-Pi
12
32
X
De
lphac
idae
Meg
amelo
des q
uadr
imac
ulatus
(Sign
oret)
N
H-Pi
2
X
X
Mue
lleria
nella
sp.1
N H-
Pi
1
X
M
uelle
riane
lla sp
.3 N
H-Pi
1
X
Fl
atida
e Cy
phop
terum
adce
nden
s (He
rr.-S
chaff
.) N
H-Pi
80
4
X X
X X
X X
Anth
ocor
idae
Brac
hyste
les pa
rvico
rnis
(A. C
osta)
N
P-Pi
5
X
X
X
Ly
gaeid
ae
Kleid
ocer
ys er
icae (
Horv
áth)
N H-
Pi
33
X
X X
X X
X
M
icrop
hysid
ae
Loric
ula co
leoptr
ata (F
allén
) N
P-Pi
2
X
M
irida
e He
teroto
ma pl
anico
rnis
(Pall
as)
N P-
Pi
3
X
M
onalo
coris
filic
is (L
inna
eus)
N H-
Pi
6
X
X
X
Pi
nalit
us or
omii
J. Ri
bes
E
H-Pi
83
9
X X
X X
X X
Po
lymer
us co
gnatu
s (Fi
eber)
N
H-Pi
1
X
Na
bidae
Na
bis p
seud
oferu
s ibe
ricus
Rem
ane
N P-
Pi
1
X
Redu
viida
e Em
picor
is ru
brom
acula
tus (
Blac
kbur
n)
I P-
Pi
1
X
Psyll
idae
St
roph
ingia
harte
ni Ho
dkin
son
E
H-Pi
64
X
X X
X X
Trioz
idae
Tr
ioza
(Lau
ritrio
za) l
auris
ilvae
Hod
kinso
n N
H-Pi
21
0
X X
X X
X X
Aphi
didae
Am
phor
opho
ra ru
bi (K
alten
bach
) sen
su
latior
e N
H-Pi
1
X
Aphi
s cra
ccivo
ra K
och
N H-
Pi
1
X
Ap
his s
p.1
N H-
Pi
1
X
Rh
opalo
siphu
m ru
fiabd
omina
lis (S
asak
i) I
H-Pi
1
X
Toxo
ptera
aura
ntii (
Boye
r de F
onsc
olomb
e) I
H-Pi
4
X

Nunes et al.
52
Clas
s Or
der
Fam
ily
Spec
ies
Colon
isatio
n
Stat
us
Feed
ing
Mod
e Nu
mbe
r of
Indi
vidu
als
Flo
Fai
Pic
Sjo
Ter
SMi
Drep
anos
iphida
e Th
eriap
his t
rifoli
i (M
onell
) N
H-Pi
1
X
La
chnid
ae
Cina
ra ju
niper
i (De
Gee
r) N
H-Pi
30
85
X
X X
X X
X
Thys
anop
tera
Phlae
othrip
idae
Apter
ygoth
rips n
.sp. ?
E
H-
Pi
1
X
Eu
rythr
ips tr
istis
Hood
I
H-Pi
1
X
Hoplo
thrips
corti
cis (D
e Gee
r) N
F-Pi
3
X
X
Hoplo
thrips
ulmi
(Fab
ricius
) I
F-Pi
4
X
Th
ripida
e Ap
tinoth
rips r
ufus H
alida
y I
H-Pi
2
X
X
Cera
tothr
ips er
icae (
Halid
ay)
N H-
Pi
1
X
He
liothr
ips h
aemo
rrhoid
alis (
Bouc
hé)
I H-
Pi
5
X
X
He
rcino
thrips
bicin
ctus (
Bagn
all)
I H-
Pi
1
X
Th
rips f
lavus
Sch
rank
N H-
Pi
4
X
X
Ne
urop
tera
Heme
robi
idae
Heme
robiu
s azo
ricus
Tjed
er E
P-
Pi
149
X
X X
X X
X
Coleo
ptera
Ca
rabida
e Oc
ys ha
rpalo
ides (
Audin
et-Se
rvill
e) N
P-Ch
3
X
X
X
Hy
drop
hilid
ae
Cerc
yon h
aemo
rrhoid
alis (
Fabr
icius
) I
S-Ch
1
X
St
aphy
linida
e Al
eoch
ara b
ipustu
lata (
Linn
aeus
) I
P-Ch
2
X X
Aloc
onota
sulci
frons
(Step
hens
) N
P-Ch
1
X
Amisc
ha an
alis (
Grav
enho
rst)
I P-
Ch
4
X
At
heta
dryo
char
es Is
raelso
n E
P-
Ch
21
X
X
X
Athe
ta fun
gi (G
raven
horst
) I
F-Ch
5
X
Gabr
ius n
igritu
lus (
Grav
enho
rst)
I P-
Ch
1
X
Ph
loeos
tiba a
zoric
a (Fa
uvel)
E
P-
Ch
1
X
Pr
oteinu
s atom
arius
Eric
hson
N
P-Ch
1
X
Suniu
s pro
pinqu
us (B
risou
t) N
P-Ch
1
X
Cr
yptop
hagid
ae
Cryp
topha
gus s
p.1
I S-
Ch
2
X
Cr
yptop
hagu
s sp.6
I
S-Ch
1
X
Silva
nidae
Cr
yptam
orph
a des
jardin
sii (G
uérin
-M
énev
ille)
I
P-Ch
7
X
Ph
alacri
dae
Stilbu
s tes
taceu
s (Pa
nzer)
N
S-Ch
1
X
Ni
tiduli
dae
Meli
gethe
s aen
eus (
Fabr
icius
) I
H-Ch
2
X
X
Meli
gethe
s sp.2
I
H-Ch
2
X
X
La
tridi
idae
Carto
dere
nodif
er (W
estw
ood)
I
S-Ch
1
X
Sc
raptii
dae
Anas
pis pr
oteu
s (W
ollas
ton)
N H-
Ch
47
X
X X
X
Ch
ryso
melid
ae
Chae
tocne
ma ho
rtens
is (F
ourcr
oy)
I H-
Ch
1
X

Biota associated with Juniperus brevifolia
53
Clas
s Or
der
Fam
ily
Spec
ies
Colon
isatio
n
Stat
us
Feed
ing
Mod
e Nu
mbe
r of
Indi
vidu
als
Flo
Fai
Pic
Sjo
Ter
SMi
Curcu
lionid
ae
Calac
alles
subc
arina
tus (I
sraels
on)
E
H-Ch
18
X X
X X
X X
Ph
loeos
inus g
iller
forsi
Brigh
t E
H-
Ch
6
X X
X X
Ps
eudo
phloe
opha
gus t
enax
(Woll
aston
) N
H-Ch
23
X X
X X
X X
Tr
ichop
tera
Limn
ephi
lidae
Li
mnep
hilus
atlan
ticus
Nyb
om ?
E
P-Ch
51
X X
X X
X X
Le
pido
ptera
Ti
neid
ae
Oino
phila
v-fla
va (H
awor
th)
I H-
No
3
X
X X
Ypon
omeu
tidae
Ar
gyre
sthia
atlan
ticell
a Reb
el E
H-
Ch/S
i 84
83
X
X X
X X
X
Bl
astob
asida
e Bl
astob
asis
sp.1
N H-
Si
1
X
Bl
astob
asis
sp.3
N H-
Si
4
X
Ne
omar
iania
sp.1
I H-
Si
2
X
Tortr
icida
e Rh
opob
ota na
evan
a (Hü
bner)
I
H-Ch
/Si
92
X
X X
X X
X
Cr
ambid
ae
Eudo
nia lu
teusa
lis (H
amps
on) ?
E
H-
Si
13
X X
Scop
aria
coec
imac
ulalis
Warr
en ?
E
H-Si
21
X X
X X
X
Sc
opar
ia se
miam
plalis
Warr
en ?
E
H-Si
2
X
Ge
ometr
idae
As
cotis
fortu
nata
azor
ica P
inker
E
H-Ch
26
X
X
X X
Cy
cloph
ora a
zore
nsis
(Pro
ut)
E
H-Ch
/Si
239
X
X X
X X
Nycte
rose
a obs
tipata
(Fab
ricius
) N
H-Ch
31
X
X X
X
Xa
nthor
hoe i
naeq
uata
(Warr
en)
E
H-Ch
11
7
X X
X X
X
Noctu
idae
Mes
apam
ea st
orai
(Reb
el)
E
H-Si
2
X X
Hym
enop
tera
Form
icida
e La
sius g
rand
is Fo
rel
N P/
H-Ch
38
8
X X
X
X X

Nunes et al.
54
Cla
ss/O
rder
Fa
mily
T
axa
Col
on.
IUC
N
Cor
Fl
o Fa
i Pi
c SJ
o T
er
SMi
Cal
ypog
eia
nees
iana
(C. M
assa
l. et
Car
estia
) Mül
l. Fr
ib.
H
Mni
olom
a fu
scum
(Leh
m.)
R. M
. Sch
ust.
R
L
H
L L/
H
Cep
halo
ziac
eae
Cep
halo
zia
bicu
spid
ata
(L.)
Dum
ort.
H
Cep
halo
zia
cras
sifo
lia (L
inde
nb. e
t Got
tsch
e) F
ulfo
rd
V
L/H
L
H
C
epha
lozi
a lu
nulif
olia
(Dum
ort.)
Dum
ort.
L
H
Now
ellia
cur
vifo
lia (D
icks
.) M
itt.
L/
H
L L/
H
L
O
dont
osch
ism
a de
nuda
tum
(Mar
t.) D
umor
t.
H
L
O
dont
osch
ism
a pr
ostr
atum
(Sw
.) Tr
evis
.
L
H
L L/
H
Cep
halo
ziel
lace
ae
Cep
halo
ziel
la d
ivar
icat
a (S
m.)
Schi
ffn.
H
Geo
caly
cace
ae
Geo
caly
x gr
aveo
lens
(Sch
rad.
) Nee
s
H
L/
H
Sa
ccog
yna
vitic
ulos
a (L
.) D
umor
t. EU
R
L
L/H
L
H
Her
berta
ceae
H
erbe
rtus
send
tner
i (N
ees)
Lin
db.
R
L
L/H
L/H
Le
pido
ziac
eae
Bazz
ania
azo
rica
H. B
uch
et H
. Per
ss.
END
R
L/H
L
L/H
Lepi
dozi
a cu
pres
sina
(Sw
.) Li
nden
b. su
bsp.
pin
nata
(H
ook.
) Póc
s
L/H
L
L/H
Lepi
dozi
a re
ptan
s (L
.) D
umor
t.
L
H
L H
Lepi
dozi
a st
uhlm
anni
i Ste
ph.
H
Tela
rane
a az
oric
a (H
. Buc
h et
H. P
erss
.) Pó
cs e
x Sc
hum
acke
r et V
áña
MA
C
V
L L/
H
L L/
H
L
Tela
rane
a eu
ropa
ea E
ngel
et M
err.
R
L/H
L/H
Lo
phoc
olea
ceae
C
hilo
scyp
hus f
ragr
ans
(Mor
is e
t De
Not
.) J.
J. E
ngel
et R
. M
. Sch
ust.
L/
H
H
Lept
oscy
phus
azo
ricu
s (H
. Buc
h et
H. P
erss
.) G
rolle
EU
R
V
H
L/H
L
Lept
oscy
phus
cun
eifo
lius
(Hoo
k.) M
itt.
H
Pl
agio
chila
ceae
Pl
agio
chila
bifa
ria
(Sw
.) Li
nden
b.
L L/
H
L L/
H
Pl
agio
chila
exi
gua
(Tay
lor)
Tay
lor
L
H
L
H
L
Pl
agio
chila
long
ispi
na L
inde
nb. e
t Got
tsch
e
H
Pl
agio
chila
pun
ctat
a (T
aylo
r) T
aylo
r
H
Pseu
dole
pico
leac
eae
Blep
haro
stom
a tr
icho
phyl
lum
(L.)
Dum
ort.
L L/
H
L/
H
Scap
ania
ceae
Ba
rbilo
phoz
ia a
ttenu
ata
(Mar
t.) L
oesk
e
L
H
D
iplo
phyl
lum
alb
ican
s (L
.) D
umor
t.
H
L H
Loph
ozia
ven
tric
osa
(Dic
ks.)
Dum
ort.
aggr
.
L
H
Sc
apan
ia c
urta
(Mar
t.) D
umor
t.
L
Scap
ania
gra
cilis
Lin
db.
L
L L/
H
L L/
H
Sc
apan
ia n
emor
ea (L
.) G
rolle
H
Sc
apan
ia u
ndul
ata
(L.)
Dum
ort.
L
H
Met
zger
iale
s A
neur
acea
e Ri
ccar
dia
cham
edry
folia
(With
.) G
rolle
H
H
Ricc
ardi
a pa
lmat
a (H
edw
.) C
arru
th.
H
M
etzg
eria
ceae
M
etzg
eria
furc
ata
(L.)
Dum
ort.
L
L L
H
Met
zger
ia le
pton
eura
Spr
uce
L/
H
L/
H
APP
END
IX II
List
of o
ther
spec
ies a
ssoc
iate
d w
ith J
unip
eus b
revi
folia
(Seu
b.) A
ntoi
ne.
Caut
iona
ry n
ote:
The
follo
win
g lis
ts do
not
gen
eral
ly re
flect
the
com
plet
e di
strib
utio
n of
the
spec
ies i
n th
e di
ffere
nt A
zore
an is
land
s sin
ce th
ey w
ere
asse
mbl
ed b
earin
g in
min
d th
e co
nnec
tion
with
Jun
iper
us b
revi
folia
; the
refo
re, t
hey
may
be
pres
ent i
n ot
her i
sland
s, in
diff
eren
t sub
strat
es (c
heck
Bor
ges
et a
l. 20
10a
for c
ompl
ete
chec
k-lis
ts). T
he p
rese
nt li
sts al
so ex
pose
the
diffe
rent
col
lect
ion
effo
rt m
ade
amon
g di
ffere
nt ta
xono
mic
al g
roup
s and
am
ong
islan
ds; F
lo, F
lore
s; Fa
i, Fa
ial;
Pic,
Pico
; SJo
, São
Jo
rge;
Ter
, Ter
ceira
, SM
i, Sã
o M
igue
l. II
.1 V
ASC
ULA
R P
LAN
TS
(All
Pter
idop
hyta
spec
ies a
re e
piph
ytes
. Arc
euth
obiu
m a
zoric
um is
the
hem
ipar
asite
of J
. bre
vifo
lia)
Tabl
e 1. L
ist o
f vas
cula
r pla
nts a
ssoc
iate
d w
ith Ju
nipe
rus b
revi
folia
. Col
onisa
tion
statu
s for
each
spec
ies (
Colo
n.) d
istin
guish
es am
ong:
EN
D, A
zore
an en
dem
ic an
d N
AT, n
ativ
e spe
cies
.
Div
ision
Cl
ass
Ord
er
Fam
ily
Spec
ies
Col
onisa
tion
Stat
us
Cor
Fl
o Fa
i Pi
c SJ
o Te
r SM
i Pt
erid
ophy
ta
Poly
podi
opsid
a H
ymen
ophy
llale
s H
ymen
ophy
llace
ae
Hym
enop
hyllu
m tu
nbrig
ense
(L.)
Sm.
NAT
x
x x
x x
x x
H
ymen
ophy
llum
wils
onii
Hoo
k. (L
.) Sm
. N
AT
x x
x x
x x
x
Poly
podi
ales
D
ryop
terid
acea
e El
apho
glos
sum
sem
icyl
indr
icum
(Bow
dich
) Ben
l N
AT
x
x x
x x
x
Po
lypo
diac
eae
Cera
deni
a ju
nger
man
nioi
des (
Klo
tzsc
h) L
.E.
Bish
op
NAT
x
x
x
G
ram
miti
s azo
rica
(Sw.
) H. S
chae
f. EN
D
x
x
x
M
agno
lioph
yta
Mag
nolio
psid
a Sa
ntal
ales
Sa
ntal
acea
e Ar
ceut
hobi
um a
zoric
um W
iens
& H
awks
wor
th
END
x
x x
x x
II.2
.1 B
RYO
PHYT
ES -
DIV
ISIO
N M
AR
CH
AN
TIO
PHY
TA
Tabl
e 2.
1. L
ist o
f liv
erw
orts
asso
ciat
ed w
ith Ju
nipe
rus b
revi
folia
. Rec
ords
com
ing
from
lite
ratu
re (L
; che
ck A
ppen
dix
III) a
nd/o
r Her
bariu
m (H
). C
olon
isatio
n sta
tus f
or
each
spe
cies
(Co
lon.
) di
sting
uish
es a
mon
g: E
ND
, Azo
rean
end
emic
; M
ac,
Mac
aron
esia
n en
dem
ic;
IB-M
AC,
Ibe
rian-
Mac
aron
esia
n en
dem
ic a
nd E
UR,
Eur
opea
n en
dem
ic; a
ll ot
her s
peci
es a
re c
onsid
ered
nat
ive;
IUCN
, men
tions
con
serv
atio
n co
ncer
n sp
ecie
s (D
iers
sen,
200
1) a
nd d
istin
guis
hes
amon
g: V
, vul
nera
ble;
R, r
are;
RT,
re
gion
ally
thre
aten
ed; a
nd T
, tax
onom
ic p
robl
ems.
Cla
ss/O
rder
Fa
mily
Ta
xa
Col
on.
IUC
N
Cor
Fl
o Fa
i Pi
c SJ
o Te
r SM
i Ju
nger
man
niop
sida
Jung
erm
anni
ales
A
crob
olba
ceae
Ac
robo
lbus
wils
onii
Nee
s
V
H
H
L
Tylim
anth
us la
xus (
Lehm
. & L
inde
nb.)
Spru
ce
V
H
L/
H
Ade
lant
hace
ae
Adel
anth
us d
ecip
iens
(Hoo
k.) M
itt.
L/
H
L/
H
L
Caly
poge
iace
ae
Caly
poge
ia a
zoric
a Bi
schl
. M
AC
R
L
Caly
poge
ia fi
ssa
(L.)
Radd
i
H
L/
H
Ca
lypo
geia
mue
lleria
na (S
chiff
n.) M
üll.
Frib
.
L
L/H
L
L/H

Biota associated with Juniperus brevifolia
55
Cla
ss/O
rder
Fa
mily
T
axa
Col
on.
IUC
N
Cor
Fl
o Fa
i Pi
c SJ
o T
er
SMi
Cal
ypog
eia
nees
iana
(C. M
assa
l. et
Car
estia
) Mül
l. Fr
ib.
H
Mni
olom
a fu
scum
(Leh
m.)
R. M
. Sch
ust.
R
L
H
L L/
H
Cep
halo
ziac
eae
Cep
halo
zia
bicu
spid
ata
(L.)
Dum
ort.
H
Cep
halo
zia
cras
sifo
lia (L
inde
nb. e
t Got
tsch
e) F
ulfo
rd
V
L/H
L
H
C
epha
lozi
a lu
nulif
olia
(Dum
ort.)
Dum
ort.
L
H
Now
ellia
cur
vifo
lia (D
icks
.) M
itt.
L/
H
L L/
H
L
O
dont
osch
ism
a de
nuda
tum
(Mar
t.) D
umor
t.
H
L
O
dont
osch
ism
a pr
ostr
atum
(Sw
.) Tr
evis
.
L
H
L L/
H
Cep
halo
ziel
lace
ae
Cep
halo
ziel
la d
ivar
icat
a (S
m.)
Schi
ffn.
H
Geo
caly
cace
ae
Geo
caly
x gr
aveo
lens
(Sch
rad.
) Nee
s
H
L/
H
Sa
ccog
yna
vitic
ulos
a (L
.) D
umor
t. EU
R
L
L/H
L
H
Her
berta
ceae
H
erbe
rtus
send
tner
i (N
ees)
Lin
db.
R
L
L/H
L/H
Le
pido
ziac
eae
Bazz
ania
azo
rica
H. B
uch
et H
. Per
ss.
END
R
L/H
L
L/H
Lepi
dozi
a cu
pres
sina
(Sw
.) Li
nden
b. su
bsp.
pin
nata
(H
ook.
) Póc
s
L/H
L
L/H
Lepi
dozi
a re
ptan
s (L
.) D
umor
t.
L
H
L H
Lepi
dozi
a st
uhlm
anni
i Ste
ph.
H
Tela
rane
a az
oric
a (H
. Buc
h et
H. P
erss
.) Pó
cs e
x Sc
hum
acke
r et V
áña
MA
C
V
L L/
H
L L/
H
L
Tela
rane
a eu
ropa
ea E
ngel
et M
err.
R
L/H
L/H
Lo
phoc
olea
ceae
C
hilo
scyp
hus f
ragr
ans
(Mor
is e
t De
Not
.) J.
J. E
ngel
et R
. M
. Sch
ust.
L/
H
H
Lept
oscy
phus
azo
ricu
s (H
. Buc
h et
H. P
erss
.) G
rolle
EU
R
V
H
L/H
L
Lept
oscy
phus
cun
eifo
lius
(Hoo
k.) M
itt.
H
Pl
agio
chila
ceae
Pl
agio
chila
bifa
ria
(Sw
.) Li
nden
b.
L L/
H
L L/
H
Pl
agio
chila
exi
gua
(Tay
lor)
Tay
lor
L
H
L
H
L
Pl
agio
chila
long
ispi
na L
inde
nb. e
t Got
tsch
e
H
Pl
agio
chila
pun
ctat
a (T
aylo
r) T
aylo
r
H
Pseu
dole
pico
leac
eae
Blep
haro
stom
a tr
icho
phyl
lum
(L.)
Dum
ort.
L L/
H
L/
H
Scap
ania
ceae
Ba
rbilo
phoz
ia a
ttenu
ata
(Mar
t.) L
oesk
e
L
H
D
iplo
phyl
lum
alb
ican
s (L
.) D
umor
t.
H
L H
Loph
ozia
ven
tric
osa
(Dic
ks.)
Dum
ort.
aggr
.
L
H
Sc
apan
ia c
urta
(Mar
t.) D
umor
t.
L
Scap
ania
gra
cilis
Lin
db.
L
L L/
H
L L/
H
Sc
apan
ia n
emor
ea (L
.) G
rolle
H
Sc
apan
ia u
ndul
ata
(L.)
Dum
ort.
L
H
Met
zger
iale
s A
neur
acea
e Ri
ccar
dia
cham
edry
folia
(With
.) G
rolle
H
H
Ricc
ardi
a pa
lmat
a (H
edw
.) C
arru
th.
H
M
etzg
eria
ceae
M
etzg
eria
furc
ata
(L.)
Dum
ort.
L
L L
H
Met
zger
ia le
pton
eura
Spr
uce
L/
H
L/
H

Nunes et al.
56
II.2
.2 B
RY
OPH
YTE
S - D
IVIS
ION
BR
YO
PHY
TA
Tabl
e 2.
2. L
ist o
f mos
ses a
ssoc
iate
d w
ith J
unip
erus
bre
vifo
lia in
the
vario
us A
zore
an is
land
s (Fl
o, F
lore
s; Fa
i, Fa
ial;
Pic,
Pic
o; S
Jo, S
ão Jo
rge;
Ter
, Ter
ceira
, SM
i, Sã
o M
igue
l). R
ecor
ds c
omin
g fro
m li
tera
ture
(L; c
heck
App
endi
x III
) and
/or H
erba
rium
(H).
Col
onisa
tion
statu
s for
eac
h sp
ecie
s (Co
lon.
) dist
ingu
ishes
am
ong:
EN
D,
Azo
rean
end
emic
; Mac
, Mac
aron
esia
n en
dem
ic; I
B-M
AC,
Iber
ian-
Mac
aron
esia
n en
dem
ic a
nd E
UR,
Eur
opea
n en
dem
ic; a
ll ot
her s
peci
es a
re c
onsid
ered
nat
ive;
IUCN
, m
entio
ns c
onse
rvat
ion
conc
ern
spec
ies (
Die
rsse
n, 2
001)
and
dist
ingu
ishes
am
ong:
V, v
ulne
rabl
e; R
, rar
e; R
T, re
gion
ally
thre
aten
ed; a
nd T
, tax
onom
ic p
robl
ems.
Cla
ss/O
rder
Fa
mily
Ta
xa
Col
on
IUC
N
Cor
Flo
Fai
Pic
SJo
Ter
SMi
Bryo
psid
a D
icra
nale
s D
icra
nace
ae
Dic
ranu
m c
anar
iens
e H
ampe
ex
Mül
l. H
al.
L
Dic
ranu
m fl
agel
lare
Hed
w.
H
Dic
ranu
m sc
opar
ium
Hed
w.
L L
L
D
icra
num
scot
tianu
m T
urn.
L L
L/H
L
L/H
Le
ucob
ryac
eae
Cam
pylo
pus c
ygne
us (H
edw
.) Br
id.
K
H
Ca
mpy
lopu
s fle
xuos
us (H
edw
.) Br
id.
L
L H
Cam
pylo
pus p
yrifo
rmis
(Sch
ultz
) Brid
.
H
Ca
mpy
lopu
s sha
wii W
ilson
R
L
Leuc
obry
um a
lbid
um (P
. Bea
uv.)
Lind
b.
L
Le
ucob
ryum
juni
pero
ideu
m (B
rid.)
Mül
l. H
al.
H
H
D
iphy
scia
les
Dip
hysc
iace
ae
Dip
hysc
ium
folio
sum
(Hed
w.)
D. M
ohr
H
ooke
riale
s D
alto
niac
eae
Dal
toni
a ste
noph
ylla
Mitt
.
V
L
Leuc
omia
ceae
Te
trasti
chiu
m fo
ntan
um (M
itt.)
Card
ot
IB-M
AC
R
L
L
Te
trasti
chiu
m v
irens
(Car
dot)
S. P
. Chu
rchi
ll IB
-MA
C
R
H
Pilo
trich
acea
e Cy
clod
icty
on la
etev
irens
(Hoo
k. e
t Tay
lor)
Mitt
.
R
L
L/H
Hyp
nale
s A
mbl
yste
giac
eae
Hyg
roam
blys
tegi
um h
umile
(P. B
eauv
.) V
ande
rp.,
Gof
finet
& H
eden
äs
H
Br
achy
thec
iace
ae
Brac
hyth
eciu
m ru
tabu
lum
(Hed
w.)
Schi
mp.
H
Ki
ndbe
rgia
pra
elon
ga (H
edw
.) O
chyr
a
L
H
yloc
omia
ceae
H
yloc
omiu
m sp
lend
ens (
Hed
w.)
Schi
mp.
L
Hyo
com
ium
arm
oric
um (B
rid.)
Wijk
et M
arg.
L
Pleu
roziu
m sc
hreb
eri (
Brid
.) M
itt.
H
H
ypna
ceae
An
doa
berth
elot
iana
(Mon
t.) O
chyr
a M
AC
R
L/
H
L/
H
H
ypnu
m a
ndoi
Sm
ith
H
Hyp
num
cup
ress
iform
e H
edw
.
L
L
L
H
H
ypnu
m ju
tland
icum
Hol
men
et E
. War
ncke
H
H
ypnu
m u
ncin
ulat
um Ju
r. EU
R RT
L
L L
L/H
L
L/H
Pseu
dota
xiph
yllu
m e
lega
ns (B
rid.)
Z. Iw
ats.
L
L L
Pseu
dota
xiph
yllu
m la
etev
irens
(Kop
pe e
t Dül
l) H
eden
äs
EUR
R
H
L/
H
Le
mbo
phyl
lace
ae
Isot
heci
um p
rolix
um (M
itt.)
Stec
h, S
im-S
im, T
angn
ey
et D
.Qua
ndt
MA
C
R
L
L/H
L
L/H
Cla
ss/O
rder
Fa
mily
T
axa
Col
on.
IUC
N
Cor
Fl
o Fa
i Pi
c SJ
o T
er
SMi
Palla
vici
nial
es
Palla
vici
niac
eae
Palla
vici
nia
lyel
lii (H
ook.
) Car
ruth
.
V
L/
H
Pore
llale
s Fr
ulla
niac
eae
Frul
lani
a az
oric
a Si
m-S
im e
t al.
IB-
MA
C
H
Frul
lani
a m
icro
phyl
la (G
otts
che)
Pea
rson
EU
R
L
H
Fr
ulla
nia
tam
aris
ci (L
.) D
umor
t.
L
L/H
L/
H
L/H
L
L/H
Frul
lani
a te
neri
ffae
(F. W
eber
) Nee
s
L
L L
L/H
H
Le
jeun
eace
ae
Apha
nole
jeun
ea a
zori
ca (V
. Allo
rge
et A
st) P
ócs
et
Ber
neck
er
V
L
L/H
Apha
nole
jeun
ea m
icro
scop
ica
(Tay
lor)
A. E
vans
L/H
H
Ap
hano
leje
unea
sin
teni
sii S
teph
.
V
L
H
H
C
heilo
leje
unea
ced
ercr
eutz
ii (H
. Buc
h et
H. P
erss
.) G
rolle
M
AC
V
H
L/
H
C
olol
ejeu
nea
min
utis
sim
a (S
m.)
Schi
ffn.
L
Col
ura
caly
ptri
folia
(Hoo
k.) D
umor
t.
RT
H
H
D
repa
nole
jeun
ea h
amat
ifolia
(Hoo
k.) S
chiff
n.
L
L L/
H
H
L/H
Har
pale
jeun
ea m
olle
ri (S
teph
.) G
rolle
L
L
L/H
H
Leje
unea
flav
a (S
w.)
Nee
s su
bsp.
moo
rei (
Lind
b.) R
. M.
Schu
st.
L
L/
H
Leje
unea
lam
acer
ina
(Ste
ph.)
Schi
ffn.
L
H
H
L/H
Leje
unea
pat
ens
Lind
b.
H
L/H
Mar
ches
inia
mac
kaii
(Hoo
k.) G
ray
H
Po
rella
ceae
Po
rella
can
arie
nsis
(F. W
eber
) Bry
hn
EUR
T
L
Rad
ulac
eae
Radu
la a
quile
gia
(Hoo
k. f.
et T
aylo
r) G
otts
che
et a
l.
L L
L/H
L/H
Radu
la c
arri
ngto
nii J
. B. J
ack
R
H
H
Radu
la h
oltii
Spr
uce
EUR
R
L
Mar
chan
tiops
ida
Mar
chan
tiale
s D
umor
tiera
ceae
D
umor
tiera
hir
suta
(Sw
.) N
ees
subs
p. h
irsu
ta
R
L
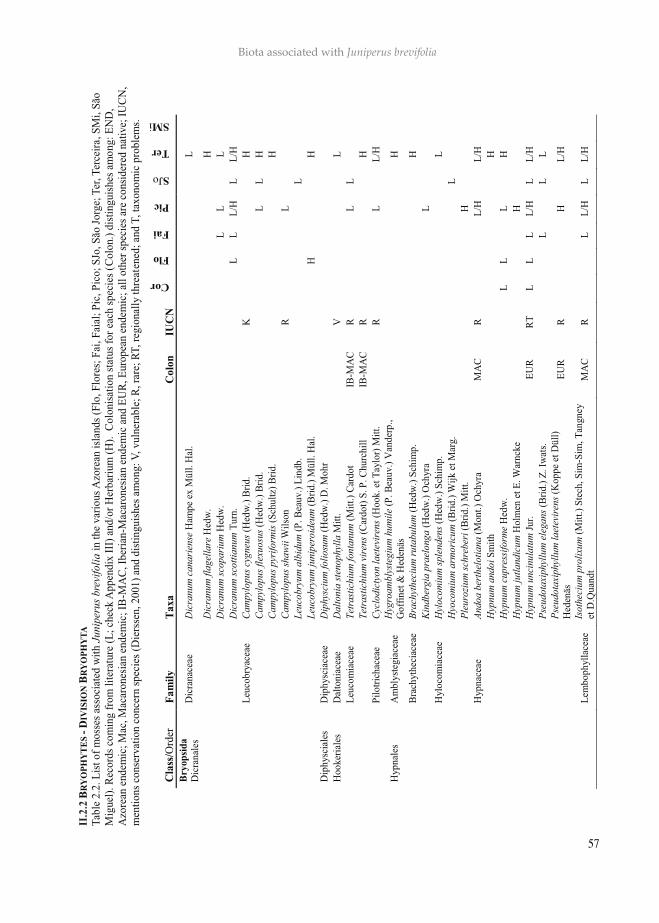
Biota associated with Juniperus brevifolia
57
II.2
.2 B
RY
OPH
YTE
S - D
IVIS
ION
BR
YO
PHY
TA
Tabl
e 2.
2. L
ist o
f mos
ses a
ssoc
iate
d w
ith J
unip
erus
bre
vifo
lia in
the
vario
us A
zore
an is
land
s (Fl
o, F
lore
s; Fa
i, Fa
ial;
Pic,
Pic
o; S
Jo, S
ão Jo
rge;
Ter
, Ter
ceira
, SM
i, Sã
o M
igue
l). R
ecor
ds c
omin
g fro
m li
tera
ture
(L; c
heck
App
endi
x III
) and
/or H
erba
rium
(H).
Col
onisa
tion
statu
s for
eac
h sp
ecie
s (Co
lon.
) dist
ingu
ishes
am
ong:
EN
D,
Azo
rean
end
emic
; Mac
, Mac
aron
esia
n en
dem
ic; I
B-M
AC,
Iber
ian-
Mac
aron
esia
n en
dem
ic a
nd E
UR,
Eur
opea
n en
dem
ic; a
ll ot
her s
peci
es a
re c
onsid
ered
nat
ive;
IUCN
, m
entio
ns c
onse
rvat
ion
conc
ern
spec
ies (
Die
rsse
n, 2
001)
and
dist
ingu
ishes
am
ong:
V, v
ulne
rabl
e; R
, rar
e; R
T, re
gion
ally
thre
aten
ed; a
nd T
, tax
onom
ic p
robl
ems.
Cla
ss/O
rder
Fa
mily
Ta
xa
Col
on
IUC
N
Cor
Flo
Fai
Pic
SJo
Ter
SMi
Bryo
psid
a D
icra
nale
s D
icra
nace
ae
Dic
ranu
m c
anar
iens
e H
ampe
ex
Mül
l. H
al.
L
Dic
ranu
m fl
agel
lare
Hed
w.
H
Dic
ranu
m sc
opar
ium
Hed
w.
L L
L
D
icra
num
scot
tianu
m T
urn.
L L
L/H
L
L/H
Le
ucob
ryac
eae
Cam
pylo
pus c
ygne
us (H
edw
.) Br
id.
K
H
Ca
mpy
lopu
s fle
xuos
us (H
edw
.) Br
id.
L
L H
Cam
pylo
pus p
yrifo
rmis
(Sch
ultz
) Brid
.
H
Ca
mpy
lopu
s sha
wii W
ilson
R
L
Leuc
obry
um a
lbid
um (P
. Bea
uv.)
Lind
b.
L
Le
ucob
ryum
juni
pero
ideu
m (B
rid.)
Mül
l. H
al.
H
H
D
iphy
scia
les
Dip
hysc
iace
ae
Dip
hysc
ium
folio
sum
(Hed
w.)
D. M
ohr
H
ooke
riale
s D
alto
niac
eae
Dal
toni
a ste
noph
ylla
Mitt
.
V
L
Leuc
omia
ceae
Te
trasti
chiu
m fo
ntan
um (M
itt.)
Card
ot
IB-M
AC
R
L
L
Te
trasti
chiu
m v
irens
(Car
dot)
S. P
. Chu
rchi
ll IB
-MA
C
R
H
Pilo
trich
acea
e Cy
clod
icty
on la
etev
irens
(Hoo
k. e
t Tay
lor)
Mitt
.
R
L
L/H
Hyp
nale
s A
mbl
yste
giac
eae
Hyg
roam
blys
tegi
um h
umile
(P. B
eauv
.) V
ande
rp.,
Gof
finet
& H
eden
äs
H
Br
achy
thec
iace
ae
Brac
hyth
eciu
m ru
tabu
lum
(Hed
w.)
Schi
mp.
H
Ki
ndbe
rgia
pra
elon
ga (H
edw
.) O
chyr
a
L
H
yloc
omia
ceae
H
yloc
omiu
m sp
lend
ens (
Hed
w.)
Schi
mp.
L
Hyo
com
ium
arm
oric
um (B
rid.)
Wijk
et M
arg.
L
Pleu
roziu
m sc
hreb
eri (
Brid
.) M
itt.
H
H
ypna
ceae
An
doa
berth
elot
iana
(Mon
t.) O
chyr
a M
AC
R
L/
H
L/
H
H
ypnu
m a
ndoi
Sm
ith
H
Hyp
num
cup
ress
iform
e H
edw
.
L
L
L
H
H
ypnu
m ju
tland
icum
Hol
men
et E
. War
ncke
H
H
ypnu
m u
ncin
ulat
um Ju
r. EU
R RT
L
L L
L/H
L
L/H
Pseu
dota
xiph
yllu
m e
lega
ns (B
rid.)
Z. Iw
ats.
L
L L
Pseu
dota
xiph
yllu
m la
etev
irens
(Kop
pe e
t Dül
l) H
eden
äs
EUR
R
H
L/
H
Le
mbo
phyl
lace
ae
Isot
heci
um p
rolix
um (M
itt.)
Stec
h, S
im-S
im, T
angn
ey
et D
.Qua
ndt
MA
C
R
L
L/H
L
L/H

Nunes et al.
58
Cla
ss/O
rder
Fa
mily
Ta
xa
Col
on
IUC
N
Cor
Flo
Fai
Pic
SJo
Ter
SMi
Myu
riace
ae
Myu
rium
hoc
hste
tteri
(Sch
imp.
) Kin
db.
L
H
L
H
Nec
kera
ceae
N
ecke
ra in
term
edia
Brid
. IB
-MA
C
RT
L
Th
amno
bryu
m a
lope
curu
m (H
edw
.) N
ieuw
l.
L
Pl
agio
thec
iace
ae
Plag
ioth
eciu
m n
emor
ale
(Mitt
.) A
. Jae
ger
L
H
Se
mat
ophy
llace
ae
Sem
atop
hyllu
m su
bstr
umul
osum
(Ham
pe) B
ritto
n
L
H
Thui
diac
eae
Thui
dium
tam
aris
cinu
m (H
edw
.) Sc
him
p.
L/
H
L H
Orth
otric
hale
s O
rthot
richa
ceae
Zy
godo
n co
noid
eus (
Dic
ks.)
Hoo
k. e
t Tay
lor
L Po
lytr
icho
psid
a Po
lytri
chal
es
Poly
trich
acea
e Po
lytr
icha
stru
m fo
rmos
um (H
edw
.) G
.L.S
m.
H
H
Poly
tric
hum
com
mun
e H
edw
.
H
Sp
hagn
opsi
da
Spha
gnal
es
Spha
gnac
eae
Spha
gnum
pal
ustr
e L.
H
H
Spha
gnum
subn
itens
Rus
sow
et W
arns
t.
H
II
.3.1
LIC
HE
NS
- DIV
ISIO
N A
SCO
MY
CO
TA
Tabl
e 3.
1. L
ist
of l
iche
ns (
Asc
omyc
ota)
ass
ocia
ted
with
Jun
iper
us b
revi
folia
in
the
vario
us A
zore
an i
slan
ds (
Flo,
Flo
res;
Fai
, Fai
al;
Pic,
Pic
o; S
Jo, S
ão J
orge
; Ter
, Te
rcei
ra, S
Mi,
São
Mig
uel).
Rec
ords
com
ing
from
lite
ratu
re (L
; che
ck A
ppen
dix
III)
and
/or H
erba
rium
(H).
Col
onis
atio
n st
atus
for e
ach
spec
ies
(Col
on.)
dist
ingu
ishe
s am
ong
Azo
rean
end
emic
s (EN
D) a
nd M
acar
ones
ian
ende
mic
s (M
AC
); al
l oth
er sp
ecie
s are
con
side
red
nativ
e.
C
lass
/ O
rder
Fa
mily
Ta
xa
Col
on
Flo
Fai
Pic
SJo
Ter
SM
i A
rtho
niom
ycet
es
Arth
onia
les
Chr
ysot
hric
acea
e C
hrys
othr
ix c
ande
lari
s (L.
) J. R
. Lau
ndon
, 198
1
L
Chr
ysot
hrix
chr
ysop
htha
lma
(P. J
ames
) P. J
ames
& J.
R. L
aund
on, 1
981
L
E
urot
iom
ycet
es
Ince
rtae
sedi
s St
rigul
acea
e St
rigul
a ni
tidul
a M
ont.,
184
2
L
Pyre
nula
les
Pyre
nula
ceae
Py
renu
la a
cutis
pora
Kal
b &
Haf
elln
er, 1
992
H
Ver
ruca
riale
s V
erru
caria
ceae
N
orm
andi
na p
ulch
ella
(Bor
rer)
Nyl
., 18
61
L
Le
cano
rom
ycet
es
Agy
riale
s A
gyria
ceae
Pl
acyn
thie
lla d
asae
a (S
tirt.)
Tøn
sber
g, 1
992
L
Trap
elia
cor
ticol
a Co
ppin
s & P
. Jam
es, 1
984
L
L
Tr
apel
iops
is fl
exuo
sa (F
r.) C
oppi
ns &
P. J
ames
, 198
4
L
L
Trap
elio
psis
pse
udog
ranu
losa
Cop
pins
& P
. Jam
es, 1
984
H
/L
G
yale
ctal
es
Gya
lect
acea
e C
oeno
goni
um p
inet
i (Sc
hrad
.) Lü
ckin
g &
Lum
bsch
, 200
4
L
L
Leca
nora
les
Can
dela
riace
ae
Can
dela
ria
conc
olor
(Dic
ks.)
Stei
n, 1
879
L
Cla
doni
acea
e C
lado
nia
borb
onic
a N
yl.,
1868
L
Cla
ss/O
rder
Fa
mily
Ta
xa
Col
on
IUC
N
Cor
Flo
Fai
Pic
SJo
Ter
SMi
M
yuria
ceae
M
yuri
um h
ochs
tette
ri (S
chim
p.) K
indb
.
L
H
L H
N
ecke
race
ae
Nec
kera
inte
rmed
ia B
rid.
IB-M
AC
R
T
L
Tham
nobr
yum
alo
pecu
rum
(Hed
w.)
Nie
uwl.
L
Plag
ioth
ecia
ceae
Pl
agio
thec
ium
nem
oral
e (M
itt.)
A. J
aege
r
L
H
Sem
atop
hylla
ceae
Se
mat
ophy
llum
subs
trum
ulos
um (H
ampe
) Brit
ton
L
H
Th
uidi
acea
e Th
uidi
um ta
mar
isci
num
(Hed
w.)
Schi
mp.
L/H
L
H
O
rthot
richa
les
Orth
otric
hace
ae
Zygo
don
cono
ideu
s (D
icks
.) H
ook.
et T
aylo
r
L
Poly
tric
hops
ida
Poly
trich
ales
Po
lytri
chac
eae
Poly
tric
hast
rum
form
osum
(Hed
w.)
G.L
.Sm
.
H
H
Poly
tric
hum
com
mun
e H
edw
.
H
Sp
hagn
opsi
da
Spha
gnal
es
Spha
gnac
eae
Spha
gnum
pal
ustr
e L.
H
H
Spha
gnum
subn
itens
Rus
sow
et W
arns
t.
H
II
.3.1
LIC
HE
NS
- DIV
ISIO
N A
SCO
MY
CO
TA
Tabl
e 3.
1. L
ist
of l
iche
ns (
Asc
omyc
ota)
ass
ocia
ted
with
Jun
iper
us b
revi
folia
in
the
vario
us A
zore
an i
slan
ds (
Flo,
Flo
res;
Fai
, Fai
al;
Pic,
Pic
o; S
Jo, S
ão J
orge
; Ter
, Te
rcei
ra, S
Mi,
São
Mig
uel).
Rec
ords
com
ing
from
lite
ratu
re (L
; che
ck A
ppen
dix
III)
and
/or H
erba
rium
(H).
Col
onis
atio
n st
atus
for e
ach
spec
ies
(Col
on.)
dist
ingu
ishe
s am
ong
Azo
rean
end
emic
s (EN
D) a
nd M
acar
ones
ian
ende
mic
s (M
AC
); al
l oth
er sp
ecie
s are
con
side
red
nativ
e.
C
lass
/ O
rder
Fa
mily
Ta
xa
Col
on
Flo
Fai
Pic
SJo
Ter
SM
i A
rtho
niom
ycet
es
Arth
onia
les
Chr
ysot
hric
acea
e C
hrys
othr
ix c
ande
lari
s (L.
) J. R
. Lau
ndon
, 198
1
L
Chr
ysot
hrix
chr
ysop
htha
lma
(P. J
ames
) P. J
ames
& J.
R. L
aund
on, 1
981
L
E
urot
iom
ycet
es
Ince
rtae
sedi
s St
rigul
acea
e St
rigul
a ni
tidul
a M
ont.,
184
2
L
Pyre
nula
les
Pyre
nula
ceae
Py
renu
la a
cutis
pora
Kal
b &
Haf
elln
er, 1
992
H
Ver
ruca
riale
s V
erru
caria
ceae
N
orm
andi
na p
ulch
ella
(Bor
rer)
Nyl
., 18
61
L
Le
cano
rom
ycet
es
Agy
riale
s A
gyria
ceae
Pl
acyn
thie
lla d
asae
a (S
tirt.)
Tøn
sber
g, 1
992
L
Trap
elia
cor
ticol
a Co
ppin
s & P
. Jam
es, 1
984
L
L
Tr
apel
iops
is fl
exuo
sa (F
r.) C
oppi
ns &
P. J
ames
, 198
4
L
L
Trap
elio
psis
pse
udog
ranu
losa
Cop
pins
& P
. Jam
es, 1
984
H
/L
G
yale
ctal
es
Gya
lect
acea
e C
oeno
goni
um p
inet
i (Sc
hrad
.) Lü
ckin
g &
Lum
bsch
, 200
4
L
L
Leca
nora
les
Can
dela
riace
ae
Can
dela
ria
conc
olor
(Dic
ks.)
Stei
n, 1
879
L
Cla
doni
acea
e C
lado
nia
borb
onic
a N
yl.,
1868
L

Biota associated with Juniperus brevifolia
59
Cla
ss/O
rder
Fa
mily
Ta
xa
Col
on
IUC
N
Cor
Flo
Fai
Pic
SJo
Ter
SMi
M
yuria
ceae
M
yuri
um h
ochs
tette
ri (S
chim
p.) K
indb
.
L
H
L H
N
ecke
race
ae
Nec
kera
inte
rmed
ia B
rid.
IB-M
AC
R
T
L
Tham
nobr
yum
alo
pecu
rum
(Hed
w.)
Nie
uwl.
L
Plag
ioth
ecia
ceae
Pl
agio
thec
ium
nem
oral
e (M
itt.)
A. J
aege
r
L
H
Sem
atop
hylla
ceae
Se
mat
ophy
llum
subs
trum
ulos
um (H
ampe
) Brit
ton
L
H
Th
uidi
acea
e Th
uidi
um ta
mar
isci
num
(Hed
w.)
Schi
mp.
L/H
L
H
O
rthot
richa
les
Orth
otric
hace
ae
Zygo
don
cono
ideu
s (D
icks
.) H
ook.
et T
aylo
r
L
Poly
tric
hops
ida
Poly
trich
ales
Po
lytri
chac
eae
Poly
tric
hast
rum
form
osum
(Hed
w.)
G.L
.Sm
.
H
H
Poly
tric
hum
com
mun
e H
edw
.
H
Sp
hagn
opsi
da
Spha
gnal
es
Spha
gnac
eae
Spha
gnum
pal
ustr
e L.
H
H
Spha
gnum
subn
itens
Rus
sow
et W
arns
t.
H
II
.3.1
LIC
HE
NS
- DIV
ISIO
N A
SCO
MY
CO
TA
Tabl
e 3.
1. L
ist
of l
iche
ns (
Asc
omyc
ota)
ass
ocia
ted
with
Jun
iper
us b
revi
folia
in
the
vario
us A
zore
an i
slan
ds (
Flo,
Flo
res;
Fai
, Fai
al;
Pic,
Pic
o; S
Jo, S
ão J
orge
; Ter
, Te
rcei
ra, S
Mi,
São
Mig
uel).
Rec
ords
com
ing
from
lite
ratu
re (L
; che
ck A
ppen
dix
III)
and
/or H
erba
rium
(H).
Col
onis
atio
n st
atus
for e
ach
spec
ies
(Col
on.)
dist
ingu
ishe
s am
ong
Azo
rean
end
emic
s (EN
D) a
nd M
acar
ones
ian
ende
mic
s (M
AC
); al
l oth
er sp
ecie
s are
con
side
red
nativ
e.
C
lass
/ O
rder
Fa
mily
Ta
xa
Col
on
Flo
Fai
Pic
SJo
Ter
SM
i A
rtho
niom
ycet
es
Arth
onia
les
Chr
ysot
hric
acea
e C
hrys
othr
ix c
ande
lari
s (L.
) J. R
. Lau
ndon
, 198
1
L
Chr
ysot
hrix
chr
ysop
htha
lma
(P. J
ames
) P. J
ames
& J.
R. L
aund
on, 1
981
L
E
urot
iom
ycet
es
Ince
rtae
sedi
s St
rigul
acea
e St
rigul
a ni
tidul
a M
ont.,
184
2
L
Pyre
nula
les
Pyre
nula
ceae
Py
renu
la a
cutis
pora
Kal
b &
Haf
elln
er, 1
992
H
Ver
ruca
riale
s V
erru
caria
ceae
N
orm
andi
na p
ulch
ella
(Bor
rer)
Nyl
., 18
61
L
Le
cano
rom
ycet
es
Agy
riale
s A
gyria
ceae
Pl
acyn
thie
lla d
asae
a (S
tirt.)
Tøn
sber
g, 1
992
L
Trap
elia
cor
ticol
a Co
ppin
s & P
. Jam
es, 1
984
L
L
Tr
apel
iops
is fl
exuo
sa (F
r.) C
oppi
ns &
P. J
ames
, 198
4
L
L
Trap
elio
psis
pse
udog
ranu
losa
Cop
pins
& P
. Jam
es, 1
984
H
/L
G
yale
ctal
es
Gya
lect
acea
e C
oeno
goni
um p
inet
i (Sc
hrad
.) Lü
ckin
g &
Lum
bsch
, 200
4
L
L
Leca
nora
les
Can
dela
riace
ae
Can
dela
ria
conc
olor
(Dic
ks.)
Stei
n, 1
879
L
Cla
doni
acea
e C
lado
nia
borb
onic
a N
yl.,
1868
L

Nunes et al.
60
Cla
ss /
Ord
er
Fam
ily
Taxa
C
olon
Fl
o Fa
i Pi
c SJ
o T
er
SMi
Loba
ria
vire
ns (W
ith.)
J. R.
Lau
ndon
, 198
4
L
Pseu
docy
phel
laria
aur
ata
(Ach
.) V
ain.
, 189
0
L
Pseu
docy
phel
laria
cro
cata
(L.)
Vai
n., 1
898
L
Ps
eudo
cyph
ella
ria in
tric
ata
(Del
ise)
Vai
n., 1
898
L
L
Ps
eudo
cyph
ella
ria la
cera
ta D
egel
., 19
41
L
St
icta
can
arie
nsis
(Ach
.) B
ory
ex D
elis
e, 1
822
L
St
icta
fulig
inos
a (D
icks
.) A
ch.,
1803
L
L
Stic
ta li
mba
ta (S
m.)
Ach
., 18
03
L
Nep
hrom
atac
eae
Nep
hrom
a he
nsse
niae
P. J
ames
& F
. J. W
hite
, 198
7 EN
D
L
L
Nep
hrom
a ve
nosu
m D
egel
., 19
41
END
L L
L
L
Pann
aria
ceae
Fu
scop
anna
ria a
tlant
ica
P. M
. Jør
g. &
P. J
ames
, 200
5
H
Pelti
gera
ceae
Pe
ltige
ra d
isse
cta
Purv
is, P
. Jam
es &
Viti
k., 1
993
END
L
L
L
Pe
ltige
ra m
elan
orrh
iza
Purv
is, P
. Jam
es &
Viti
k., 1
993
END
L
L L
Pe
rtusa
riale
s Pe
rtusa
riace
ae
Och
role
chia
inve
rsa
(Nyl
.) J.
R. L
aund
on, 1
963
L
Pe
rtus
aria
am
ara
(Ach
.) N
yl.,
1873
L L
L
Ince
rtae
sedi
s In
certa
e se
dis
Ince
rtae
sedi
s Li
chen
odip
lis le
cano
rae
(Vou
aux)
Dyk
o &
D. H
awks
w.,
1979
L
II.3
.2 L
iche
ns -
Div
isio
n B
asid
iom
ycot
a Ta
ble
3.2.
Lis
t of l
iche
ns (B
asid
iom
ycot
a) a
ssoc
iate
d w
ith J
unip
erus
bre
vifo
lia in
the
vario
us A
zore
an is
land
s (Fl
o, F
lore
s; F
ai, F
aial
; Pic
, Pic
o; S
Jo, S
ão Jo
rge;
Ter
, Te
rcei
ra, S
Mi,
São
Mig
uel).
Rec
ords
com
ing
from
lite
ratu
re (L
; che
ck A
ppen
dix
III)
. T
he sp
ecie
s is c
onsi
dere
d na
tive
to th
e Azo
res.
Cla
ss
Ord
er
Fam
ily
Taxa
C
olon
Fl
o Fa
i Pi
c SJ
o Te
r SM
i B
asid
iom
ycet
es
Ster
eale
s M
erul
iace
ae
Dic
tyon
ema
inte
rrup
tum
(Car
mic
h. e
x H
ook.
) Par
mas
to, 1
979
L
L
II
.4 B
irds
– P
hyllu
m C
hord
ata
Tabl
e 4.
Lis
t of b
irds a
ssoc
iate
d w
ith J
unip
erus
bre
vifo
lia in
the
vario
us A
zore
an is
land
s (Fl
o, F
lore
s; F
ai, F
aial
; Pic
, Pic
o; S
Jo, S
ão Jo
rge;
Ter
, Ter
ceira
, SM
i, Sã
o M
igue
l). R
ecor
ds c
omin
g fr
om li
tera
ture
(L; c
heck
App
endi
x II
I). A
ll su
bspe
cies
are
Azo
rean
end
emic
(EN
D).
C
lass
O
rder
Fa
mily
Ta
xa
Col
on
Cor
Fl
o Fa
i Pi
c SJ
o Te
r SM
i SM
a Av
es
Colu
mbi
form
es
Colu
mbi
dae
Col
umba
pal
umbu
s azo
rica
Har
tert,
190
5 EN
D
L L
L L
L L
L L
Pa
sser
iform
es
Sylv
iidae
Re
gulu
s reg
ulus
azo
ricu
s See
bohm
, 188
3 EN
D
L
Re
gulu
s reg
ulus
iner
mis
Mur
phy
& C
hapi
n, 1
929
END
L
L L
L L
L
Regu
lus r
egul
us sa
ncta
emar
iae V
aurie
, 195
4 EN
D
L
Cla
ss /
Ord
er
Fam
ily
Taxa
C
olon
Fl
o Fa
i Pi
c SJ
o T
er
SMi
Parm
otre
ma
perl
atum
(Hud
s.) M
. Cho
isy,
195
2
L
Parm
otre
ma
retic
ulat
um (T
aylo
r) M
. Cho
isy,
195
2
L
Parm
otre
ma
robu
stum
(Deg
el.)
Hal
e, 1
974
L
H
/L
Pa
rmot
rem
a su
bisid
iosu
m (M
üll.
Arg
.) H
ale,
197
4
L
L
Plat
ism
atia
gla
uca
(L.)
W. L
. Cul
b. &
C. F
. Cul
b., 1
968
L
L
U
snea
cor
nuta
Kör
b., 1
859
L
L L
L
L
U
snea
flam
mea
Stir
t., 1
881
L
U
snea
gei
ssle
rian
a P.
Cle
rc, 2
006
MA
C
L
Usn
ea h
irta
(L.)
Web
er e
x F.
H. W
igg.
, 178
0
L
Usn
ea k
rogi
ana
P. C
lerc
, 200
6
L
H/L
Usn
ea m
acar
ones
ica
P. C
lerc
, 200
6 M
AC
L
U
snea
rubi
cund
a St
irt.,
1881
L
Usn
ea su
bfla
mm
ea P
. Cle
rc, 2
006
MA
C
L
L
Usn
ea su
bsca
bros
a N
yl. e
x M
otyk
a, 1
937
L
Phys
ciac
eae
Buel
lia e
rube
scen
s Arn
old,
187
3
L
Het
erod
erm
ia ja
poni
ca (M
. Sat
ô) S
win
scow
& K
rog,
197
6
L
L
Het
erod
erm
ia le
ucom
ela
(Fée
) Sw
insc
ow &
Kro
g, 1
976
L
L
H
eter
oder
mia
lute
scen
s (K
urok
.) Fo
llman
n, 1
974
L
Py
xine
sore
diat
a (A
ch.)
Mon
t., 1
842
L
Pilo
carp
acea
e By
ssol
oma
leuc
oble
phar
um (N
yl.)
Vai
n., 1
926
L
By
ssol
oma
subd
isco
rdan
s (N
yl.)
P. Ja
mes
, 197
1
L L
Mic
area
lign
aria
(Ach
.) H
edl.,
189
2
L
H/L
L
Mic
area
pra
sina
Fr.,
182
5
L L
L
R
amal
inac
eae
Baci
dina
api
ahic
a (M
üll.
Arg
.) V
ezda
, 199
1
L
Ram
alin
a pe
ruvi
ana
Ach
., 18
10
L
Ster
eoca
ulac
eae
Lepr
aria
inca
na (L
.) A
ch.,
1803
L
Lepr
aria
lobi
fican
s Nyl
., 18
73
H
/L
Le
prar
ia u
mbr
icol
a Tø
nsbe
rg, 1
992
L
O
stro
pale
s G
omph
illac
eae
Gom
phill
us c
alyc
ioid
es (D
elis
e ex
Dub
y) N
yl.,
1855
L
L
Gya
lideo
psis
mus
cico
la P
. Jam
es &
Vez
da, 1
972
L
L
Gra
phid
acea
e Fi
ssur
ina
triti
cea
(Nyl
.) St
aige
r, 20
02
L
L L
H
/L
L
Thel
otre
mat
acea
e Ra
mon
ia a
zori
ca P
. Jam
es &
Pur
vis,
1993
L
L
L
Thel
otre
ma
anto
nini
i Pur
vis &
P. J
ames
, 199
5 EN
D
L
L
H/L
Thel
otre
ma
isid
ioid
es (B
orre
r) R
. San
t., 1
980
L
L L
Th
elot
rem
a le
padi
num
(Ach
.) A
ch.,
1803
L
H/L
L
Pelti
gera
les
Coc
coca
rpia
ceae
C
occo
carp
ia p
alm
icol
a (S
pren
g.) A
rv. &
D. J
. Gal
low
ay, 1
979
L
L
Col
lem
atac
eae
Lept
ogiu
m c
yane
scen
s (A
ch.)
Kör
b., 1
877
L
L
Loba
riace
ae
Loba
ria
pulm
onar
ia (L
.) H
offm
., 17
96
L

Biota associated with Juniperus brevifolia
61
Cla
ss /
Ord
er
Fam
ily
Taxa
C
olon
Fl
o Fa
i Pi
c SJ
o T
er
SMi
Loba
ria
vire
ns (W
ith.)
J. R.
Lau
ndon
, 198
4
L
Pseu
docy
phel
laria
aur
ata
(Ach
.) V
ain.
, 189
0
L
Pseu
docy
phel
laria
cro
cata
(L.)
Vai
n., 1
898
L
Ps
eudo
cyph
ella
ria in
tric
ata
(Del
ise)
Vai
n., 1
898
L
L
Ps
eudo
cyph
ella
ria la
cera
ta D
egel
., 19
41
L
St
icta
can
arie
nsis
(Ach
.) B
ory
ex D
elis
e, 1
822
L
St
icta
fulig
inos
a (D
icks
.) A
ch.,
1803
L
L
Stic
ta li
mba
ta (S
m.)
Ach
., 18
03
L
Nep
hrom
atac
eae
Nep
hrom
a he
nsse
niae
P. J
ames
& F
. J. W
hite
, 198
7 EN
D
L
L
Nep
hrom
a ve
nosu
m D
egel
., 19
41
END
L L
L
L
Pann
aria
ceae
Fu
scop
anna
ria a
tlant
ica
P. M
. Jør
g. &
P. J
ames
, 200
5
H
Pelti
gera
ceae
Pe
ltige
ra d
isse
cta
Purv
is, P
. Jam
es &
Viti
k., 1
993
END
L
L
L
Pe
ltige
ra m
elan
orrh
iza
Purv
is, P
. Jam
es &
Viti
k., 1
993
END
L
L L
Pe
rtusa
riale
s Pe
rtusa
riace
ae
Och
role
chia
inve
rsa
(Nyl
.) J.
R. L
aund
on, 1
963
L
Pe
rtus
aria
am
ara
(Ach
.) N
yl.,
1873
L L
L
Ince
rtae
sedi
s In
certa
e se
dis
Ince
rtae
sedi
s Li
chen
odip
lis le
cano
rae
(Vou
aux)
Dyk
o &
D. H
awks
w.,
1979
L
II.3
.2 L
iche
ns -
Div
isio
n B
asid
iom
ycot
a Ta
ble
3.2.
Lis
t of l
iche
ns (B
asid
iom
ycot
a) a
ssoc
iate
d w
ith J
unip
erus
bre
vifo
lia in
the
vario
us A
zore
an is
land
s (Fl
o, F
lore
s; F
ai, F
aial
; Pic
, Pic
o; S
Jo, S
ão Jo
rge;
Ter
, Te
rcei
ra, S
Mi,
São
Mig
uel).
Rec
ords
com
ing
from
lite
ratu
re (L
; che
ck A
ppen
dix
III)
. T
he sp
ecie
s is c
onsi
dere
d na
tive
to th
e Azo
res.
Cla
ss
Ord
er
Fam
ily
Taxa
C
olon
Fl
o Fa
i Pi
c SJ
o Te
r SM
i B
asid
iom
ycet
es
Ster
eale
s M
erul
iace
ae
Dic
tyon
ema
inte
rrup
tum
(Car
mic
h. e
x H
ook.
) Par
mas
to, 1
979
L
L
II
.4 B
irds
– P
hyllu
m C
hord
ata
Tabl
e 4.
Lis
t of b
irds a
ssoc
iate
d w
ith J
unip
erus
bre
vifo
lia in
the
vario
us A
zore
an is
land
s (Fl
o, F
lore
s; F
ai, F
aial
; Pic
, Pic
o; S
Jo, S
ão Jo
rge;
Ter
, Ter
ceira
, SM
i, Sã
o M
igue
l). R
ecor
ds c
omin
g fr
om li
tera
ture
(L; c
heck
App
endi
x II
I). A
ll su
bspe
cies
are
Azo
rean
end
emic
(EN
D).
C
lass
O
rder
Fa
mily
Ta
xa
Col
on
Cor
Fl
o Fa
i Pi
c SJ
o Te
r SM
i SM
a Av
es
Colu
mbi
form
es
Colu
mbi
dae
Col
umba
pal
umbu
s azo
rica
Har
tert,
190
5 EN
D
L L
L L
L L
L L
Pa
sser
iform
es
Sylv
iidae
Re
gulu
s reg
ulus
azo
ricu
s See
bohm
, 188
3 EN
D
L
Re
gulu
s reg
ulus
iner
mis
Mur
phy
& C
hapi
n, 1
929
END
L
L L
L L
L
Regu
lus r
egul
us sa
ncta
emar
iae V
aurie
, 195
4 EN
D
L

Nunes et al.
62
APPENDIX III List of references mentioning different taxa associated with Juniperus brevifolia BRYOPHYTES
1. Allorge, P. & Allorge, V. 1950. Hépatiques recoltées par P. et V. Allorge aux îles Açores en 1937. Révue Bryologique et Lichénologique, 19: 90-118.
2. Allorge, P. & Allorge, V. 1952. Mousses recoltées par P. et V. Allorge aux îles Açores en 1937. Révue Bryologique et Lichénologique, 21: 50-95.
3. Allorge, P. & Persson, H. 1938a. Contribution à la flore hepaticologique des îles Açores. Annales Bryologici, 11: 6-14.
4. Allorge, V. & Allorge, P. 1938. Sur la répartition et l'écologie des hépatiques epiphylles aux Açores. Boletim da Sociedade Broteriana, 2ª série, 13: 211-236.
5. Allorge, V. & Allorge, P. 1948. Végétation bryologique de l'île de Flores (Açores). Révue Bryologique et Lichénologique, 17: 126-164.
6. Bates, J. 2000. Introduction to the Azores and its Bryophytes. Bulletin of the British Bryological Society, 76: 21-23. 7. Bates, J.W. & Gabriel, R. 1997. Sphagnum cuspidatum and S. imbricatum ssp. affine new to Macaronesia, and
other new island records for Terceira, Azores. Journal of Bryology, 19 (3): 645-648. 8. Buch, H. & Persson, H. 1941. Bryophyten von den Azoren und Madeira. Societas Scientiarum Fennica.
Commentationes Biologicae, 8 (7): 1-15. 9. Frahm, J.-P. 2004. A Guide to Bryological Hotspots in Europe. Archive for Bryology, 3: 4-14. 10. Gabriel, R. 1994b. Briófitos da Ilha Terceira (Açores). Ecologia, distribuição e vulnerabilidade de espécies
seleccionadas. M. Sc. thesis. Departamento de Ciências Agrárias. Universidade dos Açores. Angra do Heroísmo. 11. Richards, P.W. 1937. A collection of bryophytes from the Azores. Annales Bryologici, 9: 131-138. 12. Schumacker, R. & J. Vána 1999. Two new liverworts for Europe in Macaronesia: Odontoschisma prostratum (Sw.)
Trevis. on the Azores and Jungermannia callithrix Lindenb. & Gottsche on the Azores and Madeira. Tropical Bryology, 17:115-127.
13. Silveira, T. 1937. Flora briológica. Espécies novas para os Açores. Açoreana, 1: 240-247. 14. Sjögren, E. 1978. Bryophyte vegetation in the Azores Islands. Memórias da Sociedade Broteriana, 26: 1-273. 15. Sjögren, E. 1993. Bryophyte flora and vegetation on the island of Corvo (Azores). Arquipélago, Life and Marine
Sciences, 11A: 17-48. 16. Sjögren, E. 1997. Report on investigations of the bryoflora and bryovegetation in 1997 on the Azorean island of
Terceira. LIFE project. Departamento de Ciências Agrárias. Angra do Heroísmo. 17. Sjögren, E. 2001. Distribution of Azorean bryophytes up to 1999, their island distribution and information on their
presence elsewhere, including Madeira and the Canary Islands. Boletim do Museu Municipal do Funchal, Sup. Nº 7: 1-89.
18. Sjögren, E. 2003. Azorean Bryophyte Communities - A Revision of Diferencial Species. Arquipélago, Life and Marine Sciences 20A: 1-29.
19. Sjögren, E. 2005c. Revisão da colecção de briófitos recolhidos pelo casal Allorge nos Açores em 1937 e por outros autores do final do século XIX e início do século XX e depositados no Museu Carlos Machado (Ponta Delgada, São Miguel). Manuscript.
20. Sjögren, E. 2006. Bryophytes (musci) unexpectedly rare or absent in the Azores. Arquipélago, Life and Marine Sciences, 23A: 1-17.
21. SMNH - Swedish Museum of Natural History (2006). Moss Register. (http://andor.nrm.se/fmi/xsl/kbo/publFinditems.xsl?-token.nav=items&-view&-db=kbo_mossregister&-token.languagecode=en-GB)
LICHENS
1. Abbayes, H. des 1946. Les Cladonia (lichens) des îles Açores. Portugalia Acta Biologia, Série B, 1: 243-254. 2. Abbayes, H. des 1948. Lichens des îles Açores récoltés en 1937 par V. et P. Allorge. Revue Bryologique et
Lichénologique, 17: 105-112. 3. Aptroot, A. 1989. Contribution to the Azores lichen flora. Lichenologist, 21 (1): 59-65.

Biota associated with Juniperus brevifolia
63
4. Arvidsson, L. 1990. Additions to the lichen flora of the Azores. Bibliotheca Lichenologica, 38: 13-27. 5. Berger, F. & F. Priemetzhofer 2008. Neufunde und interessante Nachweise von Flechten und flechtenbewohnenden
Pilzen von den Azoren. Herzogia 21 (manuscrito) (pp1-53). 6. Clerc, P. 2006. Synopsis of Usnea (lichenized Ascomycetes) from the Azores with additional information on the
species in Macaronesia. Lichenologist, 38 (3): 191-212. 7. James, P.W. & F.J. White 1987. Studies on the genus Nephroma 1. The European and Macaronesian species.
Lichenologist, 19 (3): 215-268. 8. Jorgensen, P.M. & P.W. James 2001. A new Erioderma taxon from the Azores. Lichenologist, 33 (6): 483-485. 9. Purvis, O.W. & P.W. James 1993. Studies on the lichens of the Azores. Part 1 - Caldeira do Faial. Arquipélago. Life
and Marine Sciences, 11A: 1-15. 10. Purivs, O.W., P.W. James & C.W. Smith 1995. Studies on the lichens of the Azores. Part 3. Macrolichens of relict
cloud forests. Boletim do Museu Municipal do Funchal, 4: 599-619. 11. PURVIS, O. W., JAMES, P. W., SMITH, C. W. & DIAS, E. (1998). Studies in the lichens of the Azores. Part 5. A
comparison between the lichen floras associated with Juniperus brevifolia in relict woodland in selected sites on Terceira and Flores. Boletim do Museu Municipal do Funchal, Sup. 5: 333-348.
12. Purvis, O.W., P.M. Jorgensen & P.W. JAMES 1995. The lichen genus Thelotrema Ach. in Europe. Bibliotheca Lichenologica, 58: 335-360.
13. Santos, N. & A.F. Rodrigues 2005. Resultados da identificação das amostras do Cimo do Pico da Vara. Documento de autor, 2 pp. Angra do Heroísmo.
14. Santos, N. 2005. Elaboração de uma ferramenta promocional da ZPEPVRG (Zona de Protecção Especial do Pico da Vara e Ribeira do Guilherme) numa perspectiva de educação ambiental. Relatório de Estágio de Licenciatura em Engenharia do Ambiente. Departamento de Ciências Agrárias, Angra do Heroísmo.
15. SMNH - Swedish Museum of Natural History (2007). Lichen database. 12 pp. http://andor.nrm.se/fmi/xsl/kbo/publResultitems.xsl
BIRDS 1. Bannerman, D.A. & W.M. BANNERMAN 1966. Birds of the Atlantic Islands. Volume 3. Oliver & Boyd. London.
262 pp. 2. Cabral, M.J. (coord.); J. Almeida, P.R. Almeida, T. Delliger, N. Ferrand de Almeita, M.E. Oliveira, J.M. Palmeirim,
A.I. Queirós, L. Rogado, M. Santos-Reis (eds.) 2005. Livro Vermelho dos Vertebrados de Portugal. Instituto da Conservação da Natureza. ICNB. Lisboa. 659 pp.
3. Gabriel, R. 2000. Ecophysiology of Azorean forest bryophytes. Ph. D. thesis. Imperial College of Science, Technology and Medicine, London University. Silwood Park, Ascot, Berkshire. 310 pp.
4. Moura, N.B. 2014. Caracterização dos habitats propícios à conservação da Estrelinha-de-Santa-Maria (Regulus regulus sanctae-mariae). Mestrado em Gestão e Conservação da Natureza. Universidade dos Açores, Departamento de Ciências Agrárias. Angra do Heroísmo. 65 pp.

Nunes et al.
64
Table 2. Results of post-hoc Chi-square tests (with Bonferroni adjustment) for the proportion of abundance per order at island level, for all species, endemics, natives and introduced spe-cies.
Total Endemic
comparisonAdjusted p-value comparison
Adjusted p-value
TER vs FAI 0.00062 TER vs FAI 0.00000TER vs FLO 0.00017 TER vs FLO 0.00102TER vs PIC NS TER vs PIC NSTER vs SJG 0.00002 TER vs SJG 0.00000TER vs SMG 0.00123 TER vs SMG 0.00000FAI vs FLO NS FAI vs FLO NSFAI vs PIC NS FAI vs PIC NSFAI vs SJG 0.00030 FAI vs SJG NSFAI vs SMG 0.00002 FAI vs SMG 0.00000FLO vs PIC 0.00002 FLO vs PIC NSFLO vs SJG NS FLO vs SJG NSFLO vs SMG 0.00000 FLO vs SMG 0.00000PIC vs SJG NS PIC vs SJG NSPIC vs SMG 0.00000 PIC vs SMG 0.00000SJG vs SMG 0.00000 SJG vs SMG 0.00000
Native Introduced
comparisonAdjusted p-value comparison
Adjusted p-value
TER vs FAI 0.00127 TER vs FAI 0.00000TER vs FLO NS TER vs FLO 0.00000TER vs PIC NS TER vs PIC 0.00000TER vs SJG NS TER vs SJG 0.00000TER vs SMG NS TER vs SMG 0.00030FAI vs FLO 0.00284 FAI vs FLO -FAI vs PIC NS FAI vs PIC NSFAI vs SJG 0.00000 FAI vs SJG NSFAI vs SMG 0.00000 FAI vs SMG 0.00000FLO vs PIC NS FLO vs PIC 0.00487FLO vs SJG 0.00003 FLO vs SJG -FLO vs SMG 0.00104 FLO vs SMG 0.00000PIC vs SJG 0.00102 PIC vs SJG 0.00031PIC vs SMG 0.00558 PIC vs SMG 0.00001SJG vs SMG NS SJG vs SMG 0.00000
APPENDIX IV Results of chi-square tests for all pairs of populations, with resulting p-values ad-justed with Bonferroni method (for inflation due to multiple comparisons); FAI -Faial Island, FLO – Flores Island, PIC – Pico Island, SJG – São Jorge Island, SMG –São Miguel Island, TER- Terceira Island.
Table 1. Results of post-hoc Chi-square tests (with Bonferroni adjustment) for the proportion of abundance per colonisation status for endemic, native and introduced species.
Colonisation statuscomparison Adjusted p-valueFLO vs FAI 0.00000FLO vs PIC 0.03033FLO vs SJG NSFLO vs TER 0.00000FLO vs SMG 0.00107FAI vs PIC NSFAI vs SJG 0.00278FAI vs TER NSFAI vs SMG NSPIC vs SJG NSPIC vs TER NSPIC vs SMG NSSJG vs TER 0.00272SJG vs SMG NSTER vs SMG NS

Biota associated with Juniperus brevifolia
65
Table 2. Results of post-hoc Chi-square tests (with Bonferroni adjustment) for the proportion of abundance per order at island level, for all species, endemics, natives and introduced spe-cies.
Total Endemic
comparisonAdjusted p-value comparison
Adjusted p-value
TER vs FAI 0.00062 TER vs FAI 0.00000TER vs FLO 0.00017 TER vs FLO 0.00102TER vs PIC NS TER vs PIC NSTER vs SJG 0.00002 TER vs SJG 0.00000TER vs SMG 0.00123 TER vs SMG 0.00000FAI vs FLO NS FAI vs FLO NSFAI vs PIC NS FAI vs PIC NSFAI vs SJG 0.00030 FAI vs SJG NSFAI vs SMG 0.00002 FAI vs SMG 0.00000FLO vs PIC 0.00002 FLO vs PIC NSFLO vs SJG NS FLO vs SJG NSFLO vs SMG 0.00000 FLO vs SMG 0.00000PIC vs SJG NS PIC vs SJG NSPIC vs SMG 0.00000 PIC vs SMG 0.00000SJG vs SMG 0.00000 SJG vs SMG 0.00000
Native Introduced
comparisonAdjusted p-value comparison
Adjusted p-value
TER vs FAI 0.00127 TER vs FAI 0.00000TER vs FLO NS TER vs FLO 0.00000TER vs PIC NS TER vs PIC 0.00000TER vs SJG NS TER vs SJG 0.00000TER vs SMG NS TER vs SMG 0.00030FAI vs FLO 0.00284 FAI vs FLO -FAI vs PIC NS FAI vs PIC NSFAI vs SJG 0.00000 FAI vs SJG NSFAI vs SMG 0.00000 FAI vs SMG 0.00000FLO vs PIC NS FLO vs PIC 0.00487FLO vs SJG 0.00003 FLO vs SJG -FLO vs SMG 0.00104 FLO vs SMG 0.00000PIC vs SJG 0.00102 PIC vs SJG 0.00031PIC vs SMG 0.00558 PIC vs SMG 0.00001SJG vs SMG NS SJG vs SMG 0.00000

Nunes et al.
66
67
Gomes-Pereira, J.N., I. Tojeira, L.P. Ribeiro, R.S. Santos, F. Dias, F. Tempera 2015. First in situ observations of soft bottom megafauna from the Cascais Canyon head. Arquipelago. Life and Marine Sciences 32: 67-74.
We report the first in situ observations of soft bottom megafauna from the Cascais Canyon head. Observations were collected opportunistically during three technical dives with the ROV Luso between 460-805 m at two locations distanced 1,230 m. The habitats were clas-sified as upper bathyal fine mud. The soft bottom fauna was dominated by burrows of Nephrops norvegicus reaching up to 2.9 burrows/m2, a common habitat along the Portu-guese continental margin. To our knowledge, densities are the highest ever reported for depths below 300 m. The ichthyofauna at the upper Cascais Canyon is a mixture of lower shelf and upper bathyal species, including Phycis blennoides, Scyliorhynus canicula, Coe-lorhynchus labiatus/occa and Chimaera monstrosa. Bait release attracted Myxine glutinosa. Surveys in other geological settings of the Cascays Canyon are required to understand more comprehensively the diversity of its sessile and vagile biodiversity.
Key words: Atlantic, Canyon, soft bottom megafauna, ichthyofauna, Nephrops norvegicus
Gomes-Pereira1, J.N., I. Tojeira, L.P. Ribeiro, F. Dias, Task Group for the Extension of the Continental Shelf (EMEPC), Rua Costa Pinto 165, Paço de Arcos 2770-047, Portugal. R.S. Santos & F. Tempera 1MARE – Marine and Environmental Research Centre, IMAR, Department of Oceanography and Fisheries of the University of the Azores, 9901-862 Horta, Azores, Portugal.
INTRODUCTION
Canyons play key roles in geological and biogeo-chemical processes (Gardner 1989; Arzola et al. 2008). In the last decade there was an increasing acknowledgment of the importance of submarine Canyons for biodiversity (Morell 2007; Tyler et al. 2009). In southwestern Europe, the Portuguese continental shelf and slope are cut by several can-yons, with the Nazaré Canyon amongst the largest and most studied in the world (Tyler et al. 2009). The Cascais canyon is located approximately 120 km to the south of the Nazaré Canyon, cut-ting into the Portuguese shelf edge at depths of ~175 m. It is the steepest of the Portuguese Can-yons and one of the shortest attaining the abyssal plain, with water depths exceeding 2500 m, in a mere 20 km (Lastras et al. 2009). In contrast with
the Nazaré canyon, which has no rivers of signifi-cance in its vicinity, the Cascais canyon is located off the Tagus mouth – the longest river of the Iberian Peninsula and the one draining the second largest Iberian basin (e.g. Milliman et al., 1995). This makes the Cascais Canyon more prone to anthropogenic impacts originating both from pol-lutants collected throughout the Tagus drainage basin and from human activities based on the neighbouring densely populated and industrial-ized Lisbon metropolitan area. Despite this proximity to a major northeastern Atlantic European capital (i.e. Lisbon), the mega-faunal communities from the Cascais canyon, like other canyons on the Portuguese continental mar-gin, have been poorly studied, with few studies reporting in-situ observations of its biological communities.
Table 3. Results of post-hoc Chi-square tests (with Bonferroni adjustment) for the proportion of abundance per different functional groups at island level.
Functional groupcomparison Adjusted p-valueTER vs FAI 0.00053TER vs FLO 0.03419TER vs PIC NSTER vs SJG NS
TER vs SMG NSFAI vs FLO NSFAI vs PIC NSFAI vs SJG NS
FAI vs SMG 0.00608FLO vs PIC NSFLO vs SJG NS
FLO vs SMG NSPIC vs SJG NS
PIC vs SMG NSSJG vs SMG 0.02355
Table 4. Results of post-hoc Chi-square tests (with Bonferroni adjustment) for the proportion of abundance per different feeding modes at island level.
Feeding modecomparison Adjusted p-valueTER vs FAI 0.01233TER vs FLO 0.00064TER vs PIC NSTER vs SJG 0.00002
TER vs SMG 0.03018FAI vs FLO NSFAI vs PIC NSFAI vs SJG 0.00045
FAI vs SMG 0.00006FLO vs PIC NSFLO vs SJG NS
FLO vs SMG 0.00000PIC vs SJG NS
PIC vs SMG 0.00001SJG vs SMG 0.00000

67
67
Gomes-Pereira, J.N., I. Tojeira, L.P. Ribeiro, R.S. Santos, F. Dias, F. Tempera 2015. First in situ observations of soft bottom megafauna from the Cascais Canyon head. Arquipelago. Life and Marine Sciences 32: 67-74.
We report the first in situ observations of soft bottom megafauna from the Cascais Canyon head. Observations were collected opportunistically during three technical dives with the ROV Luso between 460-805 m at two locations distanced 1,230 m. The habitats were clas-sified as upper bathyal fine mud. The soft bottom fauna was dominated by burrows of Nephrops norvegicus reaching up to 2.9 burrows/m2, a common habitat along the Portu-guese continental margin. To our knowledge, densities are the highest ever reported for depths below 300 m. The ichthyofauna at the upper Cascais Canyon is a mixture of lower shelf and upper bathyal species, including Phycis blennoides, Scyliorhynus canicula, Coe-lorhynchus labiatus/occa and Chimaera monstrosa. Bait release attracted Myxine glutinosa. Surveys in other geological settings of the Cascays Canyon are required to understand more comprehensively the diversity of its sessile and vagile biodiversity.
Key words: Atlantic, Canyon, soft bottom megafauna, ichthyofauna, Nephrops norvegicus
Gomes-Pereira1, J.N., I. Tojeira, L.P. Ribeiro, F. Dias, Task Group for the Extension of the Continental Shelf (EMEPC), Rua Costa Pinto 165, Paço de Arcos 2770-047, Portugal. R.S. Santos & F. Tempera 1MARE – Marine and Environmental Research Centre, IMAR, Department of Oceanography and Fisheries of the University of the Azores, 9901-862 Horta, Azores, Portugal.
INTRODUCTION
Canyons play key roles in geological and biogeo-chemical processes (Gardner 1989; Arzola et al. 2008). In the last decade there was an increasing acknowledgment of the importance of submarine Canyons for biodiversity (Morell 2007; Tyler et al. 2009). In southwestern Europe, the Portuguese continental shelf and slope are cut by several can-yons, with the Nazaré Canyon amongst the largest and most studied in the world (Tyler et al. 2009). The Cascais canyon is located approximately 120 km to the south of the Nazaré Canyon, cut-ting into the Portuguese shelf edge at depths of ~175 m. It is the steepest of the Portuguese Can-yons and one of the shortest attaining the abyssal plain, with water depths exceeding 2500 m, in a mere 20 km (Lastras et al. 2009). In contrast with
the Nazaré canyon, which has no rivers of signifi-cance in its vicinity, the Cascais canyon is located off the Tagus mouth – the longest river of the Iberian Peninsula and the one draining the second largest Iberian basin (e.g. Milliman et al., 1995). This makes the Cascais Canyon more prone to anthropogenic impacts originating both from pol-lutants collected throughout the Tagus drainage basin and from human activities based on the neighbouring densely populated and industrial-ized Lisbon metropolitan area. Despite this proximity to a major northeastern Atlantic European capital (i.e. Lisbon), the mega-faunal communities from the Cascais canyon, like other canyons on the Portuguese continental mar-gin, have been poorly studied, with few studies reporting in-situ observations of its biological communities.

Gomes-Pereira et al.
68
In situ observations from Cascais Canyon
69
Figure 1. a-b) Cascais Canyon area on the European continental margin, Portuguese Coast; c-d) surveyed sites A and B; (image d) with vertical exaggeration of 2x; source: Estrutura de Missão para a Extensão da Plataforma Continental, EMEPC 2015). Table 1. Location of Dives in the Cascais Canyon head
Start Start position Depth (m) Site DiveCode Date Time Bottom time Lat; Long Min-Max A L15D01 25-05-2015 18:54:38 01:57:19 38,470042; 9,468358 795-802
A L15D03 27-05-2015 09:27:20 01:39:18 38,470184; 9,468489 797-805
B L15D02 26-05-2015 11:00:38 00:56:03 before bait 02:20:48 after bait 38,469550; 9,454483 461-491
68
We report benthic megafaunal observations rec-orded opportunistically with an ROV during technical operations in the upper reach of the Cascais Canyon, including a description of habi-tats and a list of invertebrate megafauna and fish fauna.
METHODS STUDY AREA The head of the Cascais Canyon is located at 38.507°N; 9.480°W approximately 23 km to the south of the town of Cascais. Its head has a 4.2 km width, a V-shaped morphology, and is incised down to 1350 m (Lastras et al. 2009). ROV OBSERVATIONS Three dives with the ROV Luso were performed in May 2015 in the Cascais Canyon head with technical objectives related to practicing the de-ployment and retrieval of equipment on the sea-floor. ROV Luso is a working class Bathysaurus XL (Calado et al. 2008) from the Task Group for the Extension of Continental Shelf (EMEPC), and was operated from the NRP Almirante Gago Coutinho during the EMEPC/PEPC/LUSO/2015. Digital HD video was recorded and used to de-scribe faunal occurrences. Faunal occurrences were described to the lowest feasible taxa (Hayward & Ryland 1995). The camera is an Ar-gus HD-SDI camera, comprising a Argus HD Camera Housing with a Sony FCB-H11 camera (10x optical zoom; focal distance 5.1-51mm). Two parallel scaling lasers distanced ca. 63.5 cm apart projected on the seafloor, were used for measurements and area estimates. The software ImageJ was used for image analysis. The ROV dives were performed in two locations, distanced 1.230 m. Two dives were performed at site A, between 795-805 m (dives L15D01 and L15D03), and one dive in site B, ranging 460-470 m (L15D02) (Table 1, Fig. 1). A total bottom time of 7.5h was recorded. The imagery collected during the ROV opera-tions on the seafloor was annotated in order to extract scientific megafauna observations. Lists of sessile and mobile invertebrates as well as fishes were compiled for each dive. Habitat description notes were limited due to high turbidity and bot-tom currents. A video transect with the ROV
front camera in oblique view was executed at Site B to obtain densities of invertebrate megafauna. The ROV travelled at ca. 05m/s distanced 0.5 to 1.5 m from the seafloor. Area estimations were calculated using the parallel laser lines projected on the seafloor following (Dias et al. 2015). The video transect was sub-sampled by extracting still images at regular 10-second intervals between 11:07:00 and 11:11:20 (hh:mm:ss). Images of poor quality and with overlapping seafloor sec-tions were excluded. A total of 25 images were analysed, representing on average 0.49 m2 (STD=0.11) of seafloor per image. Densities of Nephrops norvegicus were esti-mated by counting burrows on the seafloor (see Morello et al. 2007 for a review; Aguzzi & Sardà 2008). The burrows were ascribed to Nephrops norvegicus, which were observed at the burrow entrances in several occasions (Fig 2b). Other aspects considered were the shape and appearance of burrow openings, the size and angle of tunnels, the geometric relationship between openings, as well as the presence of features such as tracks next to the openings (Chapman & Rice 1971; Atkinson 1974; Chapman 1980; Tuck et al. 1994; Marrs et al. 1996). Generally, each burrow was assumed to be occupied and contain a single ani-mal (Sardà & Aguzzi 2012). However, where the above-mentioned criteria suggested that clusters of openings were related and represented a single burrow system, individual openings were not enumerated and a single burrow was counted. Burrows of uncertain occupancy, in a state of collapse or with partially-blocked openings indi-cating abandonment were also ignored as it is known that unoccupied burrows rapidly degrade and collapse (Marrs e al., 1996). These criteria render conservative estimates. In an attempt to record further fish species, fish attraction was stimulated by releasing bait and shutting down all the ROV's light and mechanical systems (dive L15D03; site A). The bait was composed of rotten Trachurus trachurus and was held in latex gloves. The bait was released using the ROV arm to rip the glove apart, and the lights were switched off for 30 minutes. The lights were then switched on, a second portion of the bait was released, and all ichthyofauna observed was reg-istered.

In situ observations from Cascais Canyon
69
In situ observations from Cascais Canyon
69
Figure 1. a-b) Cascais Canyon area on the European continental margin, Portuguese Coast; c-d) surveyed sites A and B; (image d) with vertical exaggeration of 2x; source: Estrutura de Missão para a Extensão da Plataforma Continental, EMEPC 2015). Table 1. Location of Dives in the Cascais Canyon head
Start Start position Depth (m) Site DiveCode Date Time Bottom time Lat; Long Min-Max A L15D01 25-05-2015 18:54:38 01:57:19 38,470042; 9,468358 795-802
A L15D03 27-05-2015 09:27:20 01:39:18 38,470184; 9,468489 797-805
B L15D02 26-05-2015 11:00:38 00:56:03 before bait 02:20:48 after bait 38,469550; 9,454483 461-491

Gomes-Pereira et al.
70
In situ observations from Cascais Canyon
71
Figure 2. a) Site A, b) site B with Nephrops norvegicus at the entrance of a burrow; c) Cerianthus sp. (site A); d) two Bonnelia viridis (Site A/B), e) Chimaera monstrosa (Site B), f) Scyliorhynus canicula (Site B), g) Coe-lorhynchus labiatus, h) Myxine glutinosa. demonstrated that deep-sea biodiversity and eco-system functioning may also be impaired by commercial trawling fisheries frequently targeting the shelf areas upstream from submarine canyons and the canyon rims themselves (Palanques 2006;
Martín et al. 2008; Martín 2014). By enhancing the input of re-suspended particles into canyons and eventually triggering sediment gravity flows, such activities extend trawling impacts deeper from the fishing grounds, into bathyal sectors of
70
RESULTS
The seafloor at both site A and B was dominated by clay. At the deeper site (site A) the sediment layer reached a thickness of at least 0.5 m (Fig. 2a). High suspension was generated by particulate matter constantly arriving to the seafloor. Bonellia viridis was the most frequently ob-served epibenthic sessile species with densities up to 3.63 ind.m-2 (average 0.224 burrows.m-2; SD=0.812). The seafloor was heavily bioturbated, particularly at the shallowest site (site B). Occu-pied and old burrows of Nephrops norvegicus made up a significant part of the major bioturba-tion signs observed. These reached 2.902 bur-rows.m-2 (average 0.826 burrows.m-2 with SD=1.168). Henslow's swimming crabs Polybius henslowii were observed at the seafloor on both sites (Fig. 2c), as well as during the ROV descent in dive
L15D03. Euphasiacea were attracted to the ROV lights in the three dives, limiting the illumination of the seafloor. Ichthyofauna records were the most varied, yielding at least eight taxa. All fish were consid-ered to have been attracted to the ROV, as these occurred with the ROV inactive on the seafloor or appeared swimming in the ROV field of view (Lorance & Trenkel 2006). Exception was a one Schyliorhinus canicula individual that was ob-served resting on the seafloor (Fig. 2f). After bait release, and 30 min. of light absence, the ichthyofaunal observed in the vicinity of the ROV was S. canicula and Myxine glutinosa. S. canicula was observed biting the latex glove with bait remains. M. glutinosa entered the ROV front drawer possibly in search or attracted by bait re-mains.
Table 2. Soft bottom megafauna (number of individuals) observed in situ in the Cascais Canyon head (TL – Total Length in cm; * faunal occurrences only after bait release).
Site A Site B TL Order Lower taxa L15D0 L15D03 L15D02 (cm) Annelidae Cerianthus sp. - - 1 n.a. Crustacea Nephrops norvegicus - - 2 n.a. Decapoda undetermined (shrimp) - - 1 n.a. Echiura Bonellia viridis - - 9 n.a. Actinopterigyii Anguiliforme undet. 1 3 - n.a. Coelorhynchus occa/labiatus 1 1 1 40 Chimaera monstrosa - 1 1 79.5 Macrouridae - - 1 n.a. Myxine glutinosa - - 1* n.a. Phycis blennoides - - 1 33 Scyliorhinus canicula - - 1+2* 44.5 Synaphobranchus kaupi 1 - - n.a. Synaphobranchus sp. 1 n.a.
DISCUSSION
The soft bottom invertebrate fauna in the Cascais Canyon head is dominated by Nephrops norvegi-cus, inhabiting burrows excavated in the muddy sediment. This is a common habitat in the Portu-guese shelf and slope areas between 90 and 800 m depth (Marta-Almeida et al. 2008). The species supports one of the most valuable fisheries from the Northeast Atlantic to the Mediterranean
(ICES 2012) including part of the Portuguese fleet operating on the shelf. It is worth noting that on none of the sites were these burrows associat-ed with seapens as in the OSPAR priority list habitat “Seapens and burrowing megafauna communities” (Commission 2008). It is unclear if the absence of large epibenthic corals results from a lack of natural conditions for the development of these organisms. A series of works on Mediterranean canyons

In situ observations from Cascais Canyon
71
In situ observations from Cascais Canyon
71
Figure 2. a) Site A, b) site B with Nephrops norvegicus at the entrance of a burrow; c) Cerianthus sp. (site A); d) two Bonnelia viridis (Site A/B), e) Chimaera monstrosa (Site B), f) Scyliorhynus canicula (Site B), g) Coe-lorhynchus labiatus, h) Myxine glutinosa. demonstrated that deep-sea biodiversity and eco-system functioning may also be impaired by commercial trawling fisheries frequently targeting the shelf areas upstream from submarine canyons and the canyon rims themselves (Palanques 2006;
Martín et al. 2008; Martín 2014). By enhancing the input of re-suspended particles into canyons and eventually triggering sediment gravity flows, such activities extend trawling impacts deeper from the fishing grounds, into bathyal sectors of

Gomes-Pereira et al.
72
In situ observations from Cascais Canyon
73
other anonymous reviewers. J.N.G.P. was sup-ported by a doctoral grant from the Regional Di-rectorate for Education, Science and Culture, of the Regional Government of the Azores (M3.1.2/F/062/2011).
REFERENCES
Aguzzi, J., A. Bozzano & F. Sardà 2004. First observations on Nephrops norvegicus (L.) burrow densities on the continental shelf off the Catalan coast (western Mediterranean). Crustaceana 77: 299-310.
Aguzzi, J. & F. Sardà 2008. A history of recent advancements on Nephrops norvegicus behavioral and physiological rhythms. Reviews in Fish Biology and Fisheries 18: 235-248.
Arzola, R.G., R.B. Wynn, G. Lastras, D.G. Masson & P.P.E. Weaver 2008. Sedimentary features and processes in the Nazaré and Setúbal submarine canyons, west Iberian margin. Marine Geology 250: 64-88.
Atkinson, R.J.A. 1974. Spatial distribution of Nephrops burrows. Estuarine and Coastal Marine Science 2: 171-176.
Calado, A., N. Lourenço, P. Madureira, R. Costa, M. Abreu & E. Team 2008. LUSO: Portugal's new 6000 m rated ROV for deep sea research, AGU Fall Meeting Abstracts, p. 1271.
Campbell, N., L. Allan, A. Weetman & H. Dobby 2009. Investigating the link between Nephrops norvegicus burrow density and sediment composition in Scottish waters. ICES Journal of Marine Science: Journal du Conseil 66: 2052-2059.
Chapman, C.J. 1980. Ecology of juvenile and adult Nephrops. The biology and management of lobsters 2: 143-178.
Chapman, C.J. & A.L. Rice 1971. Some direct observations on the ecology and behaviour of the Norway lobster Nephrops norvegicus. Marine Biology 10: 321-329.
Commission, O. 2008. OSPAR List of Threatened and/or Declining Species and Habitats. Reference.
Curdia, J., S. Carvalho, A. Ravara, J.D. Gage, A.M. Rodrigues & V. Quintino 2004. Deep macrobenthic communities from Nazaré submarine canyon (NW Portugal). Scientia Marina 68: 171-180.
Davies, J.S., K.L. Howell, H.A. Stewart, J. Guinan & N. Golding 2014. Defining biological assemblages (biotopes) of conservation interest in the submarine canyons of the South West Approaches (offshore United Kingdom) for use in marine habitat mapping. Deep Sea Research Part II: Topical
Studies in Oceanography 104: 208-229. Dias, F., J. Gomes-Pereira, I. Tojeira, M. Souto, A.
Afonso, A. Calado, P. Madureira & A.S. Campos 2015. Area estimation of deep-sea surfaces from oblique still images. PloS one.
Gage, J.D., P.A. Lamont & P.A. Tyler 1995. Deep-Sea Macrobenthic Communities at Contrasting Sites off Portugal, Preliminary Results: I Introduction and Diversity Comparisons. Internationale Revue der Gesamten Hydrobiologie und Hydrographie 80: 235-250.
Gardner, W.D. 1989. Baltimore Canyon as a modern conduit of sediment to the deep sea. Deep Sea Research Part A. Oceanographic Research Papers 36: 323-358.
Hayward, P.J. & J.S. Ryland 1995. Handbook of the marine fauna of North-West Europe. Oxford University Press.
Huvenne, V.A. & J.S. Davies 2014. Towards a new and integrated approach to submarine canyon research. Deep-Sea Research Part II: 1-5.
ICES, A.C.o.F.M. 2012. ICES SGNEPS REPORT 2012 Report of the Study Group on Nephrops Surveys (SGNEPS), in: ICES (Ed.), Ancona, Italy, p. 36p.
Sea Macrobenthic Communities at Contrasting Sites off Portugal, Preliminary Results: II Spatial Dispersion. Internationale Revue der gesamten Hydrobiologie und Hydrographie 80: 251-265.
Lastras, G., R.G. Arzola, D.G. Masson, R.B. Wynn, V.A.I. Huvenne, V. Hühnerbach & M. Canals 2009. Geomorphology and sedimentary features in the Central Portuguese submarine canyons, Western Iberian margin. Geomorphology 103: 310-329.
Lorance, P. & V.M. Trenkel 2006. Variability in natural behaviour, and observed reactions to an ROV, by mid-slope fish species. Journal of Experimental Marine Biology and Ecology 332: 106-119.
Marrs, S.J., R.J.A. Atkinson, C.J. Smith & J.M. Hills 1996. Calibration of the towed underwater TV technique for use in stock assessment of Nephrops norvegicus. EC DGXIV Final Report, Study Project 94: 069.
Marta-Almeida, M., J. Dubert, A. Peliz, A. dos Santos & H. Queiroga 2008. A modelling study of Norway lobster (Nephrops norvegicus) larval dispersal in southern Portugal: predictions of larval wastage and self-recruitment in the Algarve stock. Canadian Journal of Fisheries and Aquatic Sciences 65: 2253-2268.
Martín, J., P. Puig, A. Palanques, P. Masqué & J. García-Orellana 2008. Effect of commercial
72
deep-sea canyons. From our observations, no trawl marks or litter items were observed at both sites. The presence of suitable sediment is considered a key factor for Nephrops habitat selection and distribution. However the relationship between Nephrops burrow density and sediments appears to be non-linear and stock-specific (Campbell et al. 2009). Existing data generally show density declining with increasing depth (see review in Aguzzi et al. 2004). In fact, burrow densities at 460-470 meters depth in the Cascais canyon (0.826 burrows.m-2) are among the highest re-ported at such depths. Reported densities below 300 m are generally below 0.2 burrows.m-2
(Smith et al. 2003). The application of the area estimation method of Dias et al. (2015) represents an alternative suitable approach for surveying grounds of Nephrops (Morello et al. 2007). However, a com-bination of methods may still be most adequate (Sardà & Aguzzi 2012). The ichthyofauna at the upper Cascais Canyon comprises a mixture of lower shelf and upper bathyal species. The distribution of S. canicula extends from shelf habitats down to 400 m (Whitehead et al. 1989), and was only observed at the shallower site. Phycis blennoides is also most-ly a shelf species, ranging from 150-300 meters and was observed at 467 m. Coelorhynchus labi-atus/occa, which normally range from 580-2,200 m (Whitehead et al. 1989), were observed at both shallow and deep locations (Fig. 2g). The species was observed in all dives on three consecutive days. In two occasions, C. labiatus approached the ROV just as it reached the seafloor, being strongly attracted to the platform. Other species were observed in the two dives, Chimaera mon-strosa (on both shallow and deep sites; normal depth range 300-500 m), and anguilliform fishes, only present at the deeper site. Rather sparse information is available on can-yon benthic communities (Paterson et al. 2011; Huvenne & Davies 2014). This is true even for macrofaunal communities, the best studied group in the submarine canyons from northeast Atlantic continental margins with data restricted to a few Canyons: Capbreton Canyon (Rallo et al. 1994), Cap-Ferret (Sorbe 1999), Nazaré canyon (Curdia
et al. 2004; Paterson et al. 2011) and the Setúbal canyon (Gage et al. 1995; Lamont et al. 1995; Paterson et al. 2011). In the Cascais Canyon, Pat-erson et al. (2011) addressed macrofaunal diversi-ty and assemblage structure using polychaetes with conclusions hampered by the small-scale heterogeneity derived from box corers. The meg-afaunal of canyon upper region has been ad-dressed in other areas, such as in the South West UK (Davies et al. 2014), with bottoms of mud and sand composed mostly of burrowing fauna, including ophiuroids and cerianthids, with resem-blances to the continental shelf communities. This report adds to the growing efforts to un-derstand canyon biodiversity and map marine habitats and associated species. The habitat ob-served conforms simultaneously to EUNIS level 3 A6.5 Deep-sea mud as well as to EUNIS level 4 A6.81 Canyons, channels, slope failures and slumps on the continental slope (http://eunis.eea.europa.eu). Such double nature is a good illustration of the ambiguity affecting many deep-sea habitats in the current version of EUNIS classification, where the attributed class depends on following the substrate-based path or the geomorphologic one (habitat complexes in section X of the classification). The soft bottom fauna here reported should extend throughout the Canyon head, as similar geomorphology is expected throughout the can-yon head. Some reliefs known to occur in the area where not explored, notably a ~80 m high es-carpment crossing the canyon axis at 920 m water depth (Lastras et al., 2009). Different faunal settings are expected to occur as well as certain taxonomic groups not detected in this survey (Tecchio et al. 2013). Further studies should focus on different habitat features and deeper areas of the Cascais Canyon to extend current knowledge on species, habitats and canyon biodi-versity.
ACKNOWLEDGMENTS
The authors wish to acknowledge the ROV pilots and the support of all the Task Group for the Ex-tension of Continental Shelf (EMEPC) team dur-ing at sea operations. The manuscript was greatly improved with the comments of J. Aguzzi and

In situ observations from Cascais Canyon
73
In situ observations from Cascais Canyon
73
other anonymous reviewers. J.N.G.P. was sup-ported by a doctoral grant from the Regional Di-rectorate for Education, Science and Culture, of the Regional Government of the Azores (M3.1.2/F/062/2011).
REFERENCES
Aguzzi, J., A. Bozzano & F. Sardà 2004. First observations on Nephrops norvegicus (L.) burrow densities on the continental shelf off the Catalan coast (western Mediterranean). Crustaceana 77: 299-310.
Aguzzi, J. & F. Sardà 2008. A history of recent advancements on Nephrops norvegicus behavioral and physiological rhythms. Reviews in Fish Biology and Fisheries 18: 235-248.
Arzola, R.G., R.B. Wynn, G. Lastras, D.G. Masson & P.P.E. Weaver 2008. Sedimentary features and processes in the Nazaré and Setúbal submarine canyons, west Iberian margin. Marine Geology 250: 64-88.
Atkinson, R.J.A. 1974. Spatial distribution of Nephrops burrows. Estuarine and Coastal Marine Science 2: 171-176.
Calado, A., N. Lourenço, P. Madureira, R. Costa, M. Abreu & E. Team 2008. LUSO: Portugal's new 6000 m rated ROV for deep sea research, AGU Fall Meeting Abstracts, p. 1271.
Campbell, N., L. Allan, A. Weetman & H. Dobby 2009. Investigating the link between Nephrops norvegicus burrow density and sediment composition in Scottish waters. ICES Journal of Marine Science: Journal du Conseil 66: 2052-2059.
Chapman, C.J. 1980. Ecology of juvenile and adult Nephrops. The biology and management of lobsters 2: 143-178.
Chapman, C.J. & A.L. Rice 1971. Some direct observations on the ecology and behaviour of the Norway lobster Nephrops norvegicus. Marine Biology 10: 321-329.
Commission, O. 2008. OSPAR List of Threatened and/or Declining Species and Habitats. Reference.
Curdia, J., S. Carvalho, A. Ravara, J.D. Gage, A.M. Rodrigues & V. Quintino 2004. Deep macrobenthic communities from Nazaré submarine canyon (NW Portugal). Scientia Marina 68: 171-180.
Davies, J.S., K.L. Howell, H.A. Stewart, J. Guinan & N. Golding 2014. Defining biological assemblages (biotopes) of conservation interest in the submarine canyons of the South West Approaches (offshore United Kingdom) for use in marine habitat mapping. Deep Sea Research Part II: Topical
Studies in Oceanography 104: 208-229. Dias, F., J. Gomes-Pereira, I. Tojeira, M. Souto, A.
Afonso, A. Calado, P. Madureira & A.S. Campos 2015. Area estimation of deep-sea surfaces from oblique still images. PloS one.
Gage, J.D., P.A. Lamont & P.A. Tyler 1995. Deep-Sea Macrobenthic Communities at Contrasting Sites off Portugal, Preliminary Results: I Introduction and Diversity Comparisons. Internationale Revue der Gesamten Hydrobiologie und Hydrographie 80: 235-250.
Gardner, W.D. 1989. Baltimore Canyon as a modern conduit of sediment to the deep sea. Deep Sea Research Part A. Oceanographic Research Papers 36: 323-358.
Hayward, P.J. & J.S. Ryland 1995. Handbook of the marine fauna of North-West Europe. Oxford University Press.
Huvenne, V.A. & J.S. Davies 2014. Towards a new and integrated approach to submarine canyon research. Deep-Sea Research Part II: 1-5.
ICES, A.C.o.F.M. 2012. ICES SGNEPS REPORT 2012 Report of the Study Group on Nephrops Surveys (SGNEPS), in: ICES (Ed.), Ancona, Italy, p. 36p.
Sea Macrobenthic Communities at Contrasting Sites off Portugal, Preliminary Results: II Spatial Dispersion. Internationale Revue der gesamten Hydrobiologie und Hydrographie 80: 251-265.
Lastras, G., R.G. Arzola, D.G. Masson, R.B. Wynn, V.A.I. Huvenne, V. Hühnerbach & M. Canals 2009. Geomorphology and sedimentary features in the Central Portuguese submarine canyons, Western Iberian margin. Geomorphology 103: 310-329.
Lorance, P. & V.M. Trenkel 2006. Variability in natural behaviour, and observed reactions to an ROV, by mid-slope fish species. Journal of Experimental Marine Biology and Ecology 332: 106-119.
Marrs, S.J., R.J.A. Atkinson, C.J. Smith & J.M. Hills 1996. Calibration of the towed underwater TV technique for use in stock assessment of Nephrops norvegicus. EC DGXIV Final Report, Study Project 94: 069.
Marta-Almeida, M., J. Dubert, A. Peliz, A. dos Santos & H. Queiroga 2008. A modelling study of Norway lobster (Nephrops norvegicus) larval dispersal in southern Portugal: predictions of larval wastage and self-recruitment in the Algarve stock. Canadian Journal of Fisheries and Aquatic Sciences 65: 2253-2268.
Martín, J., P. Puig, A. Palanques, P. Masqué & J. García-Orellana 2008. Effect of commercial

Gomes-Pereira et al.
74
75
Lino, S., J.R. Xavier, R.S. Santos & A. Colaço 2015. Sponge perforating lace coral with anticancer activity. Arquipelago. Life and Marine Sciences 32: 75-77.
Silvia Lino (e-mail: [email protected]), Ricardo S. Santos & Ana Colaço, MARE – Marine and Environmental Sciences Centre, and Department of Oceanography and Fisheries, Uni-versity of the Azores, PT-9901-862 Horta, Portugal; Joana R. Xavier, Centre for Geobiolo-gy and Department of Biology, University of Bergen, Thor Møhlensgate 53A/B, NO-5020, Bergen, Norway.
This short note reports results from a pilot study to investigate new anticancer agents from deep-sea corals in which colonizing sponges were en-countered. The pure white stylasterid coral fans of Errina dabneyi (Pourtalès, 1871) are a conspicu-ous feature on the upper bathyal slopes in Azorean waters and can be found in depths from 215 to more than 500 m (Wisshak et al. 2009; Braga-Henriques et al. 2013). From the 26 spe-cies of Errina known worldwide (most from deeper waters) (Cairns 1983), E. dabneyi seems to be endemic to the Azores Archipelago and the adjacent Mid-Atlantic Ridge region (Zibrowius & Cairns 1992; Braga-Henriques et al. 2013). The anticancer activity of the lipid extract from the lace coral Errina dabneyi (Figure 1A) was being investigated as part of a broader ongoing bio prospecting research on deep-sea inverte-brates from the Azores area. Deep-sea fisheries in the Azores use longline gears at depths where cold-water corals are common (Braga-Henriques et al. 2011; Pham et al. 2014). The specimens studied were collected accidently (bycatch) at Princess Alice Seamount (near the Azorean cen-tral group of islands) at 200 m in April 2012 by a local commercial fishery fleet. Observers onboard froze samples immediately after collection and kept them at -20ºC until being extracted. A sub-sample was taken and preserved for the biological reference collection at University of the Azores (DOP/UAc COLETA), with ID number - DOP
9044. When processing Errina dabneyi, we noted that part of the coral (from the base to the middle of the primary branches) had been bored by what appeared to be a sponge in the interior of the skeleton (Figure 1B). Since it was impossible to fully separate the sponge from the coral, the part of the coral that was colonized was treated as an independent sample (CS; Figure 1c). For identifi-cation purposes, a small fragment of sponge tis-sue was excised, digested in sodium hypochlorite, and washed in a water-ethanol series. The ob-tained spicules were then mounted in Canada balsam, observed and measured under a micro-scope. The sponge was identified as Thoosa ar-mata Topsent, 1888 (Demospongiae, Astrophori-da), a species previously known to occur associ-ated with deep-sea corals in the Azores (Topsent 1904). Thoosa is one of several demosponge gen-era known for their ability to excavate calcareous substrates (see review in Cairns 1983), a genus of demosponge in the family Thoosidae. This genus is known for boring holes in corals (Schoenberg 2008) as can be seen in Figure 1B. Total lipids were extracted using a modified Bligh and Dyer method as described in Lino et al. (2013). Cyto-toxicity essays were performed on HCT-116 hu-man colon adenocarcinoma (tumorous) cell line. Briefly, cells were inoculated into plates at 2,5 104 cells/ml (150 μl/well), and incubated overnight 37ºC and 5% CO2 before treatment. Series of dilutions in DMSO of each of the
74
trawling on the deep sedimentation in a Mediterranean submarine canyon. Marine Geology 252: 150–155.
Martín, J., Puig, P., Palanques, A., Ribó, M. 2014. Trawling-induced daily sediment resuspension in the flank of a Mediterranean submarine canyon. Deep Sea Research Part B 104: 174-183.
Morell, V. 2007. Into the deep: first glimpse of Bering Sea canyons heats up fisheries battle. Science 318: 181-182.
Morello, E.B., C. Froglia & R.J.A. Atkinson 2007. Underwater television as a fishery-independent method for stock assessment of Norway lobster (Nephrops norvegicus) in the central Adriatic Sea (Italy). ICES Journal of Marine Science: Journal du Conseil 64: 1116-1123.
Palanques, A. 2006. Evidence of sediment gravity flows induced by trawling in the Palamós (Fonera) submarine canyon (northwestern Mediterranean). Deep Sea Research Part A. Oceanographic Research Papers 53.
Paterson, G.L.J., A.G. Glover, M.R. Cunha, L. Neal, H.C. de Stigter, K. Kiriakoulakis, D.S.M. Billett, G.A. Wolff, A. Tiago & A. Ravara 2011. Disturbance, productivity and diversity in deep-sea canyons: a worm's eye view. Deep Sea Research Part B: Topical Studies in Oceanography 58: 2448-2460.
Rallo, A., L. Garcia-Arberas & I. Isasi 1994. Fauna macrobéntica de los fondos del cañón de Capbretón: análisis faunístico de poliquetos, crustáceos y cnidarios y caracterización de puntos de muestreo según estos descriptores. Cahiers de biologie marine 35: 69-90.
Sardà, F. & J. Aguzzi 2012. A review of burrow counting as an alternative to other typical methods
of assessment of Norway lobster populations. Reviews in Fish Biology and Fisheries 22: 409-422.
Smith, C.J., S.J. Marrs, R.J.A. Atkinson, K.N. Papadopoulou & J.M. Hills 2003. Underwater television for fisheries-independent stock assessment of Nephrops norvegius from the Aegean (eastern Mediterranean) Sea. Marine Ecology Progress Series 256: 161-170.
Sorbe, J.C. 1999. Deep-sea macrofaunal assemblages within the benthic boundary layer of the Cap-Ferret Canyon (Bay of Biscay, NE Atlantic). Deep Sea Research Part II: Topical Studies in Oceanography 46: 2309-2329.
Tecchio, S., E. Ramírez-Llodra, J. Aguzzi, A. Sanchez-Vidal, M.M. Flexas, F. Sardà & J.B. Company 2013. Seasonal fluctuations of deep megabenthos: Finding evidence of standing stock accumulation in a flux-rich continental slope. Progress in Oceanography 118: 188-198.
Tuck, I.D., R.J.A. Atkinson & C.J. Chapman 1994. The structure and seasonal variability in the spatial distribution of Nephrops norvegicus burrows. Ophelia 40: 13-25.
Tyler, P.A., T. Amaro, R. ArzoLa, M.R. Cunha, H. dE StigTer, A. Gooday, V. Huveene, J. Ingels, K. Kiriakoulakis & G. Lastras Membrive 2009. Europe's grand canyon: Nazaré submarine canyon. Oceanography, vol. 22(1): 52-57.
Whitehead, P.J., M.L. Bauchot, J.C. Hureau, J. Nielsen & T.E. (Eds.) 1989. Fishes of the Northeast Atlantic and the Mediterranean. UNESCO, Paris.
Received 31 Oct 2015. Accepted 10 Dec 2015. Published online 30 Dec 2015.

Sponge with anticancer activity
75
75
Lino, S., J.R. Xavier, R.S. Santos & A. Colaço 2015. Sponge perforating lace coral with anticancer activity. Arquipelago. Life and Marine Sciences 32: 75-77.
Silvia Lino (e-mail: [email protected]), Ricardo S. Santos & Ana Colaço, MARE – Marine and Environmental Sciences Centre, and Department of Oceanography and Fisheries, Uni-versity of the Azores, PT-9901-862 Horta, Portugal; Joana R. Xavier, Centre for Geobiolo-gy and Department of Biology, University of Bergen, Thor Møhlensgate 53A/B, NO-5020, Bergen, Norway.
This short note reports results from a pilot study to investigate new anticancer agents from deep-sea corals in which colonizing sponges were en-countered. The pure white stylasterid coral fans of Errina dabneyi (Pourtalès, 1871) are a conspicu-ous feature on the upper bathyal slopes in Azorean waters and can be found in depths from 215 to more than 500 m (Wisshak et al. 2009; Braga-Henriques et al. 2013). From the 26 spe-cies of Errina known worldwide (most from deeper waters) (Cairns 1983), E. dabneyi seems to be endemic to the Azores Archipelago and the adjacent Mid-Atlantic Ridge region (Zibrowius & Cairns 1992; Braga-Henriques et al. 2013). The anticancer activity of the lipid extract from the lace coral Errina dabneyi (Figure 1A) was being investigated as part of a broader ongoing bio prospecting research on deep-sea inverte-brates from the Azores area. Deep-sea fisheries in the Azores use longline gears at depths where cold-water corals are common (Braga-Henriques et al. 2011; Pham et al. 2014). The specimens studied were collected accidently (bycatch) at Princess Alice Seamount (near the Azorean cen-tral group of islands) at 200 m in April 2012 by a local commercial fishery fleet. Observers onboard froze samples immediately after collection and kept them at -20ºC until being extracted. A sub-sample was taken and preserved for the biological reference collection at University of the Azores (DOP/UAc COLETA), with ID number - DOP
9044. When processing Errina dabneyi, we noted that part of the coral (from the base to the middle of the primary branches) had been bored by what appeared to be a sponge in the interior of the skeleton (Figure 1B). Since it was impossible to fully separate the sponge from the coral, the part of the coral that was colonized was treated as an independent sample (CS; Figure 1c). For identifi-cation purposes, a small fragment of sponge tis-sue was excised, digested in sodium hypochlorite, and washed in a water-ethanol series. The ob-tained spicules were then mounted in Canada balsam, observed and measured under a micro-scope. The sponge was identified as Thoosa ar-mata Topsent, 1888 (Demospongiae, Astrophori-da), a species previously known to occur associ-ated with deep-sea corals in the Azores (Topsent 1904). Thoosa is one of several demosponge gen-era known for their ability to excavate calcareous substrates (see review in Cairns 1983), a genus of demosponge in the family Thoosidae. This genus is known for boring holes in corals (Schoenberg 2008) as can be seen in Figure 1B. Total lipids were extracted using a modified Bligh and Dyer method as described in Lino et al. (2013). Cyto-toxicity essays were performed on HCT-116 hu-man colon adenocarcinoma (tumorous) cell line. Briefly, cells were inoculated into plates at 2,5 104 cells/ml (150 μl/well), and incubated overnight 37ºC and 5% CO2 before treatment. Series of dilutions in DMSO of each of the

Lino et al.
76
77
(IC50 values) was automatically calculated by SoftMax Pro software. Results for anticancer ac-tivity showed that the extract from Errina dab-neyi did not show any activity but the extract from the coral sample harbouring the sponge was active presenting IC50 values of 25.29 and 32.72 μg/mL for the two essays. To our knowledge this is the first time that anticancer activity is reported in a coral perforating sponge. Considering the interesting preliminary results on the anticancer activity for the sponge and tak-ing in consideration the high bio eroding ratio observed in Errina dabneyi specimens deposited in the collection at University of the Azores, more consistent studies should be made to address this matter.
ACKNOWLEDGMENTS
The authors would like to thank the observer onboard Daniel Pereira for bringing the samples in good preservation conditions. We are grateful to Professor William Fenical at Center for Marine Biotechnology and Biomedicine, Scripps Institu-tion of Oceanography, University of California San Diego, who kindly accepted Silvia Lino as collaborator at his lab to investigate the natural marine products of our samples and to Lauren Paul for performing the cytotoxicity essays. We thank Valentina Matos and Andreia Braga-Henriques for assisting with sampling and coral taxonomic identification. Silvia Lino was sup-ported by PhD grant from Fundação para a Ciên-cia e Tecnologia, (FCT - Ministry of Science and Technology of Portugal), with reference SFRH/BD/72154/2010.
REFERENCES
Braga-Henriques, A., M. Carreiro-Silva, F. Porteiro, V. Matos, Í. Sampaio, O. Ocaña, & S.P. Ávila 2011. The association between a deep-sea gastropod Pedicularia sicula (Caenogastropoda: Pedicularii-dae) and its coral host Errina dabneyi (Hydrozoa; Stylasteridae) in the Azores. ICES Journal of Marine Sciences 68: 399–407.
Braga-Henriques, A., F. M Porteiro, P.A. Ribeiro, V. de Matos, I. Sampaio, O. Ocaña, & R.S. Santos 2013. Diversity, distribution and spatial structure of the cold-water coral fauna of the Azores (NE Atlantic). Biogeosciences 10: 4009–4036.
Cairns, S.D. 1983. A generic revision of the Stylasterina (Coelenterata: Hydrozoa). Part 1. Description of the genera. Bulletin of Marine Science 33: 427–508
Lino, S., B. Solé, V.de Matos, R Pereira, J. Xavier, R. Santos & A. Colaço 2013. Lipid extractions from marine deep-sea invertebrates: modifications to the Bligh & Dyer method to increase lipid yields. Arquipelago. Life and Marine Sciences 31: 9-14.
Pham C.K., H. Diogo, G. Menezes, F. Porteiro, A. Braga-Henriques, F. Vandeperre & T. Morato. 2014. Regulated bottom longline helps achieving sustainability of deep-sea fisheries. Scientific Reports. 4:4837.
Schoenberg, C.H.L. 2008. A history of sponge erosion: from past myths and hypotheses to recent approaches. Pp. 165-202 in: Wishak, M. & L. Tapanila (Eds). Current developments in bioerosion. Erlangen Earth Conference Series. Berlin, Heidelberg: Springer-Verlag.
Topsent, E. 1904. Spongiaires des Açores. Résultats des campagnes scientifiques accomplies par le Prince Albert I. Monaco 25: 1-280, pls 1-18.
Wisshak, M., M. López Correa, H. Zibrowius, J. Jakobsen & A. Freiwald 2009. Skeletal reorganisation affects geochemical signals, exemplified in the stylasterid hydrocoral Errina dabneyi (Azores Archipelago). Marine Ecology Progress Series 397: 197-208.
Zibrowius, H. & S.D. Cairns 1992. Revision of the northeast Atlantic and Mediterranean Stylasteridae (Cnidaria: Hydrozoa). Mémoires du muséum national d'histoire naturelle A 153:1–136.
Received 12 Dez 2014. Accepted 06 Feb 2015, Published online 13 February 2015.
.
76
Fig. 1. a) Errina dabneyi specimen deposited in the reference collection at the University of the Azores DOP/UAç COLETA); b) Errina dabneyi specimen where it is possible to see the sponge inside (pointed by the black arrow); c) Specimen of coral E. dabneyi plus sponge (CS) investigated for their anticancer potential in this study - the white arrows indicate the holes done by the Thoosa sponge in the coral skeleton in order to connect with the exteri-or and which indicates the presence of the sponge within the coral.
extracts (Errina dabneyi (C) and Errina plus sponge (CS)), were added to the cells followed by further incubation at 37ºC for 72 h. Each extract was essayed in duplicate. Cell viability was de-termined by a colorimetric [3-(4,5-dimethyl-thiazol-2-yl)-5-(3-carboxymethoxyphen-yl)-2-(4-sulfophenyl)-2H-tetrazolium] (MTS) assay. After the exposure time, 20 μL of MTS/ PMS (phena-zine methosulfate - electron coupling agent) solu-tion was added to each well. Plates were incubat-
ed for 3 h before absorbance was measured at 490 nm using an EMax® Endpoint ELISA Plate Reader. Cells in DMSO were used as negative control, and cells treated with Etoposide (VP-16) were used as positive control. Relative cellular survival was determined by using the measured optical density (OD) and was calculated as fol-lows: (% Survival for sample) = [(OD of sam-ple)/(Average of negative control)] x100. Con-centration that killed 50% of the cancer cells

Sponge with anticancer activity
77
77
(IC50 values) was automatically calculated by SoftMax Pro software. Results for anticancer ac-tivity showed that the extract from Errina dab-neyi did not show any activity but the extract from the coral sample harbouring the sponge was active presenting IC50 values of 25.29 and 32.72 μg/mL for the two essays. To our knowledge this is the first time that anticancer activity is reported in a coral perforating sponge. Considering the interesting preliminary results on the anticancer activity for the sponge and tak-ing in consideration the high bio eroding ratio observed in Errina dabneyi specimens deposited in the collection at University of the Azores, more consistent studies should be made to address this matter.
ACKNOWLEDGMENTS
The authors would like to thank the observer onboard Daniel Pereira for bringing the samples in good preservation conditions. We are grateful to Professor William Fenical at Center for Marine Biotechnology and Biomedicine, Scripps Institu-tion of Oceanography, University of California San Diego, who kindly accepted Silvia Lino as collaborator at his lab to investigate the natural marine products of our samples and to Lauren Paul for performing the cytotoxicity essays. We thank Valentina Matos and Andreia Braga-Henriques for assisting with sampling and coral taxonomic identification. Silvia Lino was sup-ported by PhD grant from Fundação para a Ciên-cia e Tecnologia, (FCT - Ministry of Science and Technology of Portugal), with reference SFRH/BD/72154/2010.
REFERENCES
Braga-Henriques, A., M. Carreiro-Silva, F. Porteiro, V. Matos, Í. Sampaio, O. Ocaña, & S.P. Ávila 2011. The association between a deep-sea gastropod Pedicularia sicula (Caenogastropoda: Pedicularii-dae) and its coral host Errina dabneyi (Hydrozoa; Stylasteridae) in the Azores. ICES Journal of Marine Sciences 68: 399–407.
Braga-Henriques, A., F. M Porteiro, P.A. Ribeiro, V. de Matos, I. Sampaio, O. Ocaña, & R.S. Santos 2013. Diversity, distribution and spatial structure of the cold-water coral fauna of the Azores (NE Atlantic). Biogeosciences 10: 4009–4036.
Cairns, S.D. 1983. A generic revision of the Stylasterina (Coelenterata: Hydrozoa). Part 1. Description of the genera. Bulletin of Marine Science 33: 427–508
Lino, S., B. Solé, V.de Matos, R Pereira, J. Xavier, R. Santos & A. Colaço 2013. Lipid extractions from marine deep-sea invertebrates: modifications to the Bligh & Dyer method to increase lipid yields. Arquipelago. Life and Marine Sciences 31: 9-14.
Pham C.K., H. Diogo, G. Menezes, F. Porteiro, A. Braga-Henriques, F. Vandeperre & T. Morato. 2014. Regulated bottom longline helps achieving sustainability of deep-sea fisheries. Scientific Reports. 4:4837.
Schoenberg, C.H.L. 2008. A history of sponge erosion: from past myths and hypotheses to recent approaches. Pp. 165-202 in: Wishak, M. & L. Tapanila (Eds). Current developments in bioerosion. Erlangen Earth Conference Series. Berlin, Heidelberg: Springer-Verlag.
Topsent, E. 1904. Spongiaires des Açores. Résultats des campagnes scientifiques accomplies par le Prince Albert I. Monaco 25: 1-280, pls 1-18.
Wisshak, M., M. López Correa, H. Zibrowius, J. Jakobsen & A. Freiwald 2009. Skeletal reorganisation affects geochemical signals, exemplified in the stylasterid hydrocoral Errina dabneyi (Azores Archipelago). Marine Ecology Progress Series 397: 197-208.
Zibrowius, H. & S.D. Cairns 1992. Revision of the northeast Atlantic and Mediterranean Stylasteridae (Cnidaria: Hydrozoa). Mémoires du muséum national d'histoire naturelle A 153:1–136.
Received 12 Dez 2014. Accepted 06 Feb 2015, Published online 13 February 2015.
.


Eagle ray aggregations
79
79
Afonso, P. & N. Vasco-Rodrigues 2015. Summer aggregations of the common eagle ray, Myliobatis aquila. Arquipelago. Life and Marine Sciences 32: 79-82.
Pedro Afonso (email: [email protected]): IMAR-UAz - Institute of Marine Research at the University of the Azores. Dept. of Oceanography and Fisheries, PT-9901-862 Horta; MARE – Marine and Environmental Sciences Centre, University of the Azores, Dept. of Oceanography and Fisheries, 9901-862 Horta, Açores, Portugal; Nuno Vasco-Rodrigues, MARE – Marine and Environmental Sciences Centre, ESTM, Polytechnic Institute of Lei-ria, 2520-641 Peniche, Portugal; Flying Sharks, Rua Jorge Castilho 1613, 7C, 1900-272 Lisboa, Portugal.
The aggregation of multiple individuals is a long known behaviour in a variety of elasmobranchs, including reef sharks (Speed et al. 2011), ham-merhead sharks (Klimley & Nelson 1984), whale sharks (Hoffmayer et al. 2007), devilrays (Ward-Page et al. 2013; Sobral & Afonso 2014), sting-rays (Semeniuk & Rotley 2008), eagle rays (Sil-liman & Gruber 1999) and cownose rays (Smith et al. 1987). Various functions have been hypoth-esised to explain these aggregations, including feeding (e.g. Wilson et al. 2001; Rohner et al. 2013), courtship or mating (Whitney et al. 2004; Dudgeon et al. 2008), and cleaning stations (Dewar et al. 2008). However, apart from the obvious cases whenever individuals directly en-gage in reproductive activities, the social func-tions that these aggregations might serve are not understood or even described. During the summer of 2014, while conducting regular fish visual surveys, an aggregation of up to at least 30 common eagle rays was observed at Radares point, a site located on eastern point of the Monte da Guia marine protected area (MPA), in the Azorean island of Faial, mid-north Atlantic (Fig. 1 and 2). Individuals were identified based on morphological characteristics (Fisher et al. 1981) and previous citations for the region (Santos et al. 1997; Barreiros & Gadig 2011).
Fig. 1. Location of the aggregation site (dark cloud) on the perimeter of the Monte da Guia Marine Reserve, Faial Island, Azores (dashed line).

Afonso & Vasco-Rodrigues
80
80
Fig. 2. Common eagle rays aggregating at Radares point, Faial Island.
The aggregation was observed in four consecutive dives spanning eight days (Table 1). Upon the first sighting, another six dedicated dives were made until November. Dives were conducted at various times of the day, from dawn to dusk. In-dividuals were observed almost daily during the week around the full moon of August (08, 10, 11 and 14-08-2014). The aggregation held 20 to 30 observable individuals at one given dive, which measured an estimated 30 to 90 cm disc width (DW) (Fig. 2). The rays were always encountered within a localized area (ca. 1 ha) of the reef (Fig. 1), in spite of the prospective transects done in the areas surrounding the aggregation site. The site lies at the transition from the rocky to the sandy bottom at 40 m depth just off the point, but the animals where swimming in mid-water (10-30 m depth). All individuals appeared to be females as no claspers were seen sticking out of the fins
Table 1. Details of observations on the aggregation site; * indicates full moon.
Date Time Nº indiv. Size range (DW, cm)
08/08/2014 16:00 25 30-90 10/08/2014* 18:30 30 30-90 11/08/2014 07:30 20 30-90 14/08/2014 10:00 20 30-90 03/09/2014 14:00 0 -
08/09/2014* 08:00 0 - 25/11/2014 09:00 0 -
trailing edge, which would be expected in sub-adult and adult males (Capapé et al. 2007). There was no apparent close interaction between indi-viduals, feeding, or cleaning activity. Right after this period, the aggregation disappeared.

Eagle ray aggregations
81
81
The common eagle ray is a relatively rare elas-mobranch in coastal areas of the Azores, where it was only observed in 1.6% of over 500 transects for fish counts across the archipelago and over 15 years, and there was never an occasion when more than one individual had been observed (P. Afonso, personal observation). McEachran & Séret (1990) state in their review of the family that the species "is often found in groups" with no further data on locations, numbers or behaviour. Barreiros & Gadig (2011) also refer that "this species can be observed in reproductive aggrega-tions in the summers, in one specific cave, in Ilhéu das Cabras, Terceira Island" but provide no information to ascertain the function of the ag-gregation. Similar aggregations were also ob-served in the summer at an offshore reef in Gra-ciosa Island, apparently also constituted only by females of various sizes (Rolando Oliveira, pers. comm.). The fact that no particular behaviour was observed makes it impossible to even speculate about the nature of the aggregation, although it seems that there are no obvious feeding or para-site cleaning functions. As such, it remains possi-ble that these aggregations serve social functions. It is particularly intriguing that only females were confirmedly observed, although it remains possi-ble that immature males were in the group but went unnoticed. Thus, the aggregation might serve reproductive functions, namely related to egg deposition. Capapé et al. (2007) refer that this species breeds in August and September in the northwestern Mediterranean. Indeed, recruits of this species (ca. 10-15 cm DW) were observed from July to the end of summer in the neighbour nursery area of Porto Pim, just around the perime-ter of Monte da Guia (Figure 1; P. Afonso un-published data), so it appears that the reproduc-tive schedule in the Azores matches that of the Mediterranean, as in most coastal fishes. This would explain why the aggregation is exclusively composed of mature females. The nature of the aggregation and its potential significance for the conservation of this species should be investigated given its status as ‘data deficient’ and vulnerable population characteris-tics.
ACKNOWLEDGEMENTS
The authors would like to thank C. Moura and M. Schmiing for dive support and M. Simões for the help given in photo treatment. This paper is a contribution to the MoniZec project (DRCT/M212/I/018/2011).
REFERENCES
Barreiros, J. P. & O.B.F. Gadig 2011. Sharks and Rays from the Azores: An illustrated catalogue. Instituto Açoriano de Cultura, Angra do Heroísmo, Terceira, Azores. 187pp.
Capapé, C., O. Guélorget, Y. Vergne & J.P. Quignard 2007. Reproductive biology of the common Eagle Ray Myliobatis aquila (Chondrichthyes: Myliobatidae) from the Coast of Languedoc (Southern France, Northern Mediterranean). Vie milieu: Life Environment 57 (3): 1-6.
Dewar, H., P. Mous, M. Domeier, A. Muljadi, J. Pet & J. Whitty 2008. Movements and site fidelity of the giant manta ray, Manta birostris, in the Komodo Marine Park, Indonesia. Marine Biology 155: 121–133.
Dudgeon, C.L., M.J. Noad & J.M. Lanyon 2008. Abundance and demography of a seasonal aggregation of zebra sharks Stegostoma fasciatum. Marine Ecology Progress Series 368: 269–281.
Fisher, W., G. Bianchi & W.B. Scott (Eds) 1981. FAO species identification sheets for fisheries purposes. Eastern Central Atlantic; Fishing areas 34, 47 (in part). Canada Funds-in-Trust. Ottawa, Department of Fisheries and Oceans Canada, by arrangement with the Food and Agriculture Organization of the United Nations, vol. 5.
Hoffmayer, E.R., J.S. Franks, W.B. Driggers, K.J. Oswald, K.J. & J.M. Quattro 2007. Observations of a feeding aggregation of whale sharks, Rhincodon typus, in the north central Gulf of Mexico. Gulf and Caribbean Research 19(2): 69–73.
Klimley, A.P. & D.R. Nelson 1984. Diel movement patterns of the scalloped hammerhead shark (Sphyrna lewini) in relation to El Bajo Espiritu Santo: a refuging central-position social system. Behavioural Ecology and Sociobiology 15: 45-54.
McEachran, J.D. & B. Séret 1990. Myliobatididae. Pp.67-70 in: Quero, J.C., J.C. Hureau, C. Karrer, A. Post. & L. Saldanha (Eds) Check-list of the fishes of the eastern tropical Atlantic (CLOFETA). JNICT, Lisbon; SEI, Paris; and UNESCO, Paris. Vol. 1. 519 pp.
80
Fig. 2. Common eagle rays aggregating at Radares point, Faial Island.
The aggregation was observed in four consecutive dives spanning eight days (Table 1). Upon the first sighting, another six dedicated dives were made until November. Dives were conducted at various times of the day, from dawn to dusk. In-dividuals were observed almost daily during the week around the full moon of August (08, 10, 11 and 14-08-2014). The aggregation held 20 to 30 observable individuals at one given dive, which measured an estimated 30 to 90 cm disc width (DW) (Fig. 2). The rays were always encountered within a localized area (ca. 1 ha) of the reef (Fig. 1), in spite of the prospective transects done in the areas surrounding the aggregation site. The site lies at the transition from the rocky to the sandy bottom at 40 m depth just off the point, but the animals where swimming in mid-water (10-30 m depth). All individuals appeared to be females as no claspers were seen sticking out of the fins
Table 1. Details of observations on the aggregation site; * indicates full moon.
Date Time Nº indiv. Size range (DW, cm)
08/08/2014 16:00 25 30-90 10/08/2014* 18:30 30 30-90 11/08/2014 07:30 20 30-90 14/08/2014 10:00 20 30-90 03/09/2014 14:00 0 -
08/09/2014* 08:00 0 - 25/11/2014 09:00 0 -
trailing edge, which would be expected in sub-adult and adult males (Capapé et al. 2007). There was no apparent close interaction between indi-viduals, feeding, or cleaning activity. Right after this period, the aggregation disappeared.

Afonso & Vasco-Rodrigues
82
82
Rohner, C.A., S.J Pierce, A.D. Marshall, S.J. Weeks, M.B. Bennett. & A.J. Richardson 2013. Trends in sightings and environmental influences on a coastal aggregation of manta rays and whale sharks. Marine Ecology Progress Series 482: 153-168.
Santos, R.S., F.M. Porteiro & J.P. Barreiros 1997. Marine Fishes of the Azores: annotated checklist and bibliography. Arquipelago. Life and Marine Sciences. Supplement 1. 244 pp.
Semeniuk, C. & K. Rothley 2008. Costs of group-living for a normally solitary forager: effects of provisioning tourism on southern stingrays Dasyatis americana. Marine Ecology Progress Series 357: 271-282.
Silliman, W.R. & S.H. Gruber 1999. Behavioral biology of the spotted eagle ray, Aetobatus narinari (Euphrasen, 1790), in Bimini, Bahamas; an interim report. Bahamas Journal of Science 7:13–20.
Sobral, A.F. & P. Afonso 2014. Occurrence of mobulids in the Azores, central North Atlantic. Journal of the Marine Biological Association of the United Kingdom 94 (8): 1671-1675.
Speed, C.W., M.G. Meekan, I.C. Field, C.R. McMahon, J.D. Stevens, F. McGregor, C. Huveneer, Y. Berger & C.J.A. Bradshaw 2011. Spatial and temporal movement patterns of a multi-
species coastal reef shark aggregation. Marine Ecology Progress Series 429: 261-275.
Ward-Paige, C.A., B. Davis & B.Worm, 2013. Global Population Trends and Human use Patterns of Manta and Mobula Rays. Plos One 8(9): e74835.
Whitney, N.M., J.H.L. Pratt & J.C. Carrier 2004. Group courtship, mating behaviour and siphon sac function in the whitetip reef shark, Triaenodon obesus. Animal Behaviour 68: 1435–1442.
Wilson, S.G., J.G. Taylor & A.F. Pearce 2001. The Seasonal Aggregation of Whale Sharks at Ningaloo Reef, Western Australia: Currents, Migrations and the El Niño/Southern Oscillation. Environmental Biology of Fishes 61: 1-11.
Received 24 Apr 2015. Accepted 26 Jun 2015, Published online 13 Jul 2015.

Gymnothorax polygonius in Ascension Island
83
83
Richardson, A.J. & A. Brito 2015. The polygon moray, Gymnothorax polygonius (Poey, 1875): A new record from Ascension Island, South Atlantic. Arquipelago. Life and Marine Sciences 32: 83-85.
Andrew J. Richardson (email: [email protected]) Conservation Depart-ment, Ascension Island Government, Ascension Island, South Atlantic ASCN 1ZZ; Alberto Brito, Grupo de investigación BIOECOMAC (U. D. Ciencias Marinas), Universidad de La Laguna, Tenerife, Islas Canarias.
Ascension Island (07°57 S, 14°22 W) is an isolat-ed volcanic edifice in the South Atlantic. The closest land, St. Helena is approximately 700 miles south with the closest land mass of any size being West Africa, 1000 miles to the east. Approximately 1500 miles west of Ascension Island is the coast of Brazil. As such, the fish community of the island is a mixture of eastern and western Atlantic species, although richness is relatively low compared with other similar loca-tions such as Cape Verde (Wirtz et al. 2014). De-spite the apparent remoteness of Ascension Island there have been several key publications on the ichthyofauna for the island and surrounding ma-rine zone. The fish community of Ascension was first summarized by Cadenat & Marchal (1963), with a comprehensive checklist of shallow-water
fish species published by Lubbock (1980) record-ing 71 species. A report on the benthic and near-benthic fish community, from seamounts around Ascension, was compiled in Trunov (2006). The most recent review of the ichthyofauna of Ascen-sion can be found in Wirtz et al. (2014), a study listing 173 fish species, including 12 members of the family Muraenidae. The extensive rocky ma-rine habitat of Ascension Island is often consid-ered a key factor behind the considerable abun-dance of moray eels around the island. On 21 Feburary 2015 an unidentified moray eel was caught by a recreational angler who had been fishing in deep water from a vessel off the west-ern coast of Ascension Island (Fig. 1). The spec-imen did not match any previous identification records of Muraenidae from Ascension Island.
Fig. 1. Gymnothorax polygonius specimen, Ascension Island (21-02-2015)
83
Richardson, A.J. & A. Brito 2015. The polygon moray, Gymnothorax polygonius (Poey, 1875): A new record from Ascension Island, South Atlantic. Arquipelago. Life and Marine Sciences 32: 83-85.
Andrew J. Richardson (email: [email protected]) Conservation Depart-ment, Ascension Island Government, Ascension Island, South Atlantic ASCN 1ZZ; Alberto Brito, Grupo de investigación BIOECOMAC (U. D. Ciencias Marinas), Universidad de La Laguna, Tenerife, Islas Canarias.
Ascension Island (07°57 S, 14°22 W) is an isolat-ed volcanic edifice in the South Atlantic. The closest land, St. Helena is approximately 700 miles south with the closest land mass of any size being West Africa, 1000 miles to the east. Approximately 1500 miles west of Ascension Island is the coast of Brazil. As such, the fish community of the island is a mixture of eastern and western Atlantic species, although richness is relatively low compared with other similar loca-tions such as Cape Verde (Wirtz et al. 2014). De-spite the apparent remoteness of Ascension Island there have been several key publications on the ichthyofauna for the island and surrounding ma-rine zone. The fish community of Ascension was first summarized by Cadenat & Marchal (1963), with a comprehensive checklist of shallow-water
fish species published by Lubbock (1980) record-ing 71 species. A report on the benthic and near-benthic fish community, from seamounts around Ascension, was compiled in Trunov (2006). The most recent review of the ichthyofauna of Ascen-sion can be found in Wirtz et al. (2014), a study listing 173 fish species, including 12 members of the family Muraenidae. The extensive rocky ma-rine habitat of Ascension Island is often consid-ered a key factor behind the considerable abun-dance of moray eels around the island. On 21 Feburary 2015 an unidentified moray eel was caught by a recreational angler who had been fishing in deep water from a vessel off the west-ern coast of Ascension Island (Fig. 1). The spec-imen did not match any previous identification records of Muraenidae from Ascension Island.
Fig. 1. Gymnothorax polygonius specimen, Ascension Island (21-02-2015)
82
Rohner, C.A., S.J Pierce, A.D. Marshall, S.J. Weeks, M.B. Bennett. & A.J. Richardson 2013. Trends in sightings and environmental influences on a coastal aggregation of manta rays and whale sharks. Marine Ecology Progress Series 482: 153-168.
Santos, R.S., F.M. Porteiro & J.P. Barreiros 1997. Marine Fishes of the Azores: annotated checklist and bibliography. Arquipelago. Life and Marine Sciences. Supplement 1. 244 pp.
Semeniuk, C. & K. Rothley 2008. Costs of group-living for a normally solitary forager: effects of provisioning tourism on southern stingrays Dasyatis americana. Marine Ecology Progress Series 357: 271-282.
Silliman, W.R. & S.H. Gruber 1999. Behavioral biology of the spotted eagle ray, Aetobatus narinari (Euphrasen, 1790), in Bimini, Bahamas; an interim report. Bahamas Journal of Science 7:13–20.
Sobral, A.F. & P. Afonso 2014. Occurrence of mobulids in the Azores, central North Atlantic. Journal of the Marine Biological Association of the United Kingdom 94 (8): 1671-1675.
Speed, C.W., M.G. Meekan, I.C. Field, C.R. McMahon, J.D. Stevens, F. McGregor, C. Huveneer, Y. Berger & C.J.A. Bradshaw 2011. Spatial and temporal movement patterns of a multi-
species coastal reef shark aggregation. Marine Ecology Progress Series 429: 261-275.
Ward-Paige, C.A., B. Davis & B.Worm, 2013. Global Population Trends and Human use Patterns of Manta and Mobula Rays. Plos One 8(9): e74835.
Whitney, N.M., J.H.L. Pratt & J.C. Carrier 2004. Group courtship, mating behaviour and siphon sac function in the whitetip reef shark, Triaenodon obesus. Animal Behaviour 68: 1435–1442.
Wilson, S.G., J.G. Taylor & A.F. Pearce 2001. The Seasonal Aggregation of Whale Sharks at Ningaloo Reef, Western Australia: Currents, Migrations and the El Niño/Southern Oscillation. Environmental Biology of Fishes 61: 1-11.
Received 24 Apr 2015. Accepted 26 Jun 2015, Published online 13 Jul 2015.
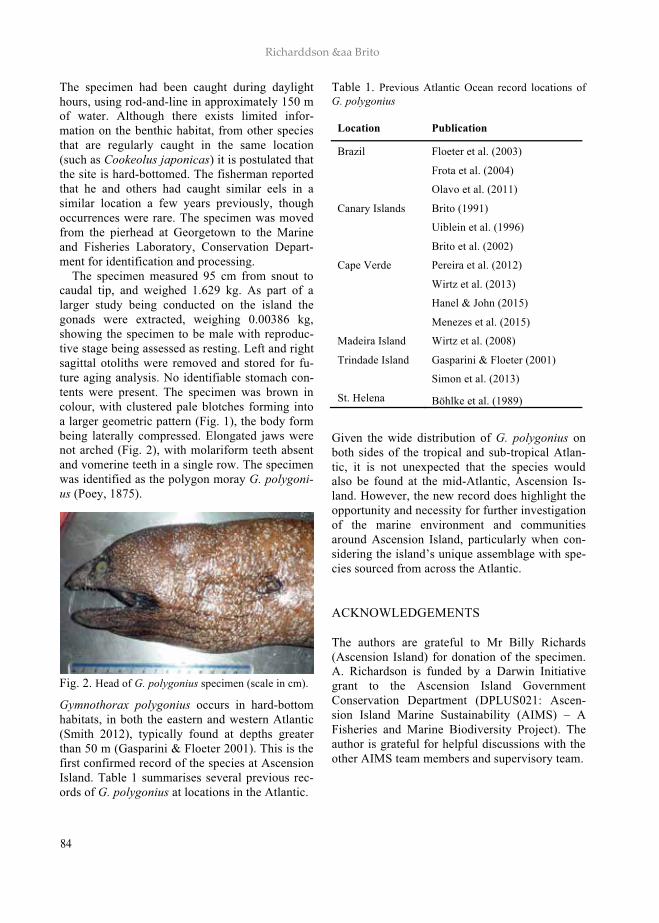
Richarddson &aa Brito
84
85
REFERENCES
Böhlke, E. B., J. E. McCosker & J. E. Böhlke 1989. Family Muraenidae. Pp. 104-206 in: Böhlke, E.B. (Ed.) Fishes of the Western North Atlantic. Part 9, vol. 1. Orders Anguilliformes and Saccopharyngiformes. Memoir Sears Foundation for Marine Research. New Haven. 655 pp.
Brito, A. 1991. Catálogo de los peces de las Islas Canarias. Francisco Lemus Editor, La Laguna. 230 pp.
Brito, A., P. J. Pascual, J. M. Falcón, A. Sancho & G. González 2002. Peces de las Islas Canarias. Catálogo comentado e ilustrado. Francisco Lemus Editor, La Laguna. 419 pp.
Cadenat, J. & E. Marchal 1963. Poissons. Résultats des campagnes océanographiques de la Reine-Pokou aux îles Sainte-Héléne et Ascension. Bulletin de l’Institut Francais d’Afrique Noire (A) Sciences Naturelles 25: 1235–1315
Floeter, S.R. & J.L. Gasparini 2003. The southwestern Atlantic reef fish fauna: composition and zoogeographic patterns. Journal of Fish Biology 56: 1099-1114
Frota, L.O., P.A.S. Costa & A.C. Braga 2004. Length-weight relationships of marine fishes from the central Brazilian coast. NAGA, WorldFish Center Quarterly 27: 20-26.
Gasparini, J.L. & S.R. Floeter 2001 The shore fishes of Trindade Island, western South Atlantic. Journal of Natural History 35: 1639-1656.
Hanel, R. & John, H.C. 2015. A revised checklist of Cape Verde Islands sea fishes. Journal of Applied Ichthyology 31: 135-169.
Lubbock, R. 1980. The shore fishes of Ascension Island. Journal of Fish Biology 17: 283-303.
Menezes, G.M., O. Tariche, M.R. Pinho, M.F. Sigler & H.M. Silva 2015. Structure and zonation of demersal and deep-water fish assemblages off the Cape Verde archipelago (northeast-Atlantic) as sampled by baited longlines. Deep Sea Research Part I. 102: 118-134.
Olavo, G. P.A. Costa, A.S. Martins & B.P. Ferreira 2011. Shelf edge reefs as priority areas for conservation of reef fish diversity in the tropical Atlantic. Aquatic Conservation: Marine and Freshwater Ecosystems 21: 199-209.
Pereira, J.N., A. Simas, A. Rosa, A. Aranha, S. Lino, E. Constantino, V. Monteiro, O. Tariche & G. Menezes 2012. Weight–length relationships for 27 demersal fish species caught off the Cape Verde archipelago (eastern North Atlantic). Journal of Applied Ichthyology 28: 156-159.
Simon, T., R.M. Macieira & J.C. Joyeux 2013. The shore fishes of the Trindade–Martin Vaz insular complex: an update. Journal of Fish Biology 82: 2113-2127.
Smith, D.G. 2012. A checklist of the moray eels of the world (Teleostei: Anguilliformes: Muraenidae). Zootaxa 3474: 1-64.
Uiblein, F., F. Bordes & R. Castillo 1996. Diversity, abundance and depth distribution of demersal deepwater fishes off Lanzarote and Fuerteventura, Canary Islands. Journal of Fish Biology 49. Supplement sA: 75-90.
Wirtz, P., R. Fricke & M.J. Biscoito 2008. The coastal fishes of Madeira Island–new records and an annotated check-list. Zootaxa 1715: 1-26.
Wirtz, P., A. Brito, J.M. Falcón, R. Freitas, R. Fricke, V. Monteiro, F. Reiner & O. Tariche 2013. The coastal fishes of the Cape Verde Islands–new records and an annotated check-list. Spixiana 36: 113-142.
Wirtz, P., J. Bingeman, J. Bingeman, R. Fricke, T.J. Hook & J. Young 2014. The fishes of Ascension Island, central Atlantic Ocean – new records and an annotated checklist. Journal of the Marine Biological Association of the United Kingdom, 16 pp.
Received 9 Jul 2015. Accepted 21 Jul 2015, Published online 14 Sep 2015.
84
The specimen had been caught during daylight hours, using rod-and-line in approximately 150 m of water. Although there exists limited infor-mation on the benthic habitat, from other species that are regularly caught in the same location (such as Cookeolus japonicas) it is postulated that the site is hard-bottomed. The fisherman reported that he and others had caught similar eels in a similar location a few years previously, though occurrences were rare. The specimen was moved from the pierhead at Georgetown to the Marine and Fisheries Laboratory, Conservation Depart-ment for identification and processing. The specimen measured 95 cm from snout to caudal tip, and weighed 1.629 kg. As part of a larger study being conducted on the island the gonads were extracted, weighing 0.00386 kg, showing the specimen to be male with reproduc-tive stage being assessed as resting. Left and right sagittal otoliths were removed and stored for fu-ture aging analysis. No identifiable stomach con-tents were present. The specimen was brown in colour, with clustered pale blotches forming into a larger geometric pattern (Fig. 1), the body form being laterally compressed. Elongated jaws were not arched (Fig. 2), with molariform teeth absent and vomerine teeth in a single row. The specimen was identified as the polygon moray G. polygoni-us (Poey, 1875).
Fig. 2. Head of G. polygonius specimen (scale in cm).
Gymnothorax polygonius occurs in hard-bottom habitats, in both the eastern and western Atlantic (Smith 2012), typically found at depths greater than 50 m (Gasparini & Floeter 2001). This is the first confirmed record of the species at Ascension Island. Table 1 summarises several previous rec-ords of G. polygonius at locations in the Atlantic.
Table 1. Previous Atlantic Ocean record locations of G. polygonius
Location Publication
Brazil Floeter et al. (2003)
Frota et al. (2004)
Olavo et al. (2011)
Canary Islands Brito (1991)
Uiblein et al. (1996)
Brito et al. (2002)
Cape Verde Pereira et al. (2012)
Wirtz et al. (2013)
Hanel & John (2015)
Menezes et al. (2015)
Madeira Island Wirtz et al. (2008)
Trindade Island Gasparini & Floeter (2001)
Simon et al. (2013)
St. Helena Böhlke et al. (1989)
Given the wide distribution of G. polygonius on both sides of the tropical and sub-tropical Atlan-tic, it is not unexpected that the species would also be found at the mid-Atlantic, Ascension Is-land. However, the new record does highlight the opportunity and necessity for further investigation of the marine environment and communities around Ascension Island, particularly when con-sidering the island’s unique assemblage with spe-cies sourced from across the Atlantic.
ACKNOWLEDGEMENTS
The authors are grateful to Mr Billy Richards (Ascension Island) for donation of the specimen. A. Richardson is funded by a Darwin Initiative grant to the Ascension Island Government Conservation Department (DPLUS021: Ascen-sion Island Marine Sustainability (AIMS) – A Fisheries and Marine Biodiversity Project). The author is grateful for helpful discussions with the other AIMS team members and supervisory team.
84
The specimen had been caught during daylight hours, using rod-and-line in approximately 150 m of water. Although there exists limited infor-mation on the benthic habitat, from other species that are regularly caught in the same location (such as Cookeolus japonicas) it is postulated that the site is hard-bottomed. The fisherman reported that he and others had caught similar eels in a similar location a few years previously, though occurrences were rare. The specimen was moved from the pierhead at Georgetown to the Marine and Fisheries Laboratory, Conservation Depart-ment for identification and processing. The specimen measured 95 cm from snout to caudal tip, and weighed 1.629 kg. As part of a larger study being conducted on the island the gonads were extracted, weighing 0.00386 kg, showing the specimen to be male with reproduc-tive stage being assessed as resting. Left and right sagittal otoliths were removed and stored for fu-ture aging analysis. No identifiable stomach con-tents were present. The specimen was brown in colour, with clustered pale blotches forming into a larger geometric pattern (Fig. 1), the body form being laterally compressed. Elongated jaws were not arched (Fig. 2), with molariform teeth absent and vomerine teeth in a single row. The specimen was identified as the polygon moray G. polygoni-us (Poey, 1875).
Fig. 2. Head of G. polygonius specimen (scale in cm).
Gymnothorax polygonius occurs in hard-bottom habitats, in both the eastern and western Atlantic (Smith 2012), typically found at depths greater than 50 m (Gasparini & Floeter 2001). This is the first confirmed record of the species at Ascension Island. Table 1 summarises several previous rec-ords of G. polygonius at locations in the Atlantic.
Table 1. Previous Atlantic Ocean record locations of G. polygonius
Location Publication
Brazil Floeter et al. (2003)
Frota et al. (2004)
Olavo et al. (2011)
Canary Islands Brito (1991)
Uiblein et al. (1996)
Brito et al. (2002)
Cape Verde Pereira et al. (2012)
Wirtz et al. (2013)
Hanel & John (2015)
Menezes et al. (2015)
Madeira Island Wirtz et al. (2008)
Trindade Island Gasparini & Floeter (2001)
Simon et al. (2013)
St. Helena Böhlke et al. (1989)
Given the wide distribution of G. polygonius on both sides of the tropical and sub-tropical Atlan-tic, it is not unexpected that the species would also be found at the mid-Atlantic, Ascension Is-land. However, the new record does highlight the opportunity and necessity for further investigation of the marine environment and communities around Ascension Island, particularly when con-sidering the island’s unique assemblage with spe-cies sourced from across the Atlantic.
ACKNOWLEDGEMENTS
The authors are grateful to Mr Billy Richards (Ascension Island) for donation of the specimen. A. Richardson is funded by a Darwin Initiative grant to the Ascension Island Government Conservation Department (DPLUS021: Ascen-sion Island Marine Sustainability (AIMS) – A Fisheries and Marine Biodiversity Project). The author is grateful for helpful discussions with the other AIMS team members and supervisory team.
84
The specimen had been caught during daylight hours, using rod-and-line in approximately 150 m of water. Although there exists limited infor-mation on the benthic habitat, from other species that are regularly caught in the same location (such as Cookeolus japonicas) it is postulated that the site is hard-bottomed. The fisherman reported that he and others had caught similar eels in a similar location a few years previously, though occurrences were rare. The specimen was moved from the pierhead at Georgetown to the Marine and Fisheries Laboratory, Conservation Depart-ment for identification and processing. The specimen measured 95 cm from snout to caudal tip, and weighed 1.629 kg. As part of a larger study being conducted on the island the gonads were extracted, weighing 0.00386 kg, showing the specimen to be male with reproduc-tive stage being assessed as resting. Left and right sagittal otoliths were removed and stored for fu-ture aging analysis. No identifiable stomach con-tents were present. The specimen was brown in colour, with clustered pale blotches forming into a larger geometric pattern (Fig. 1), the body form being laterally compressed. Elongated jaws were not arched (Fig. 2), with molariform teeth absent and vomerine teeth in a single row. The specimen was identified as the polygon moray G. polygoni-us (Poey, 1875).
Fig. 2. Head of G. polygonius specimen (scale in cm).
Gymnothorax polygonius occurs in hard-bottom habitats, in both the eastern and western Atlantic (Smith 2012), typically found at depths greater than 50 m (Gasparini & Floeter 2001). This is the first confirmed record of the species at Ascension Island. Table 1 summarises several previous rec-ords of G. polygonius at locations in the Atlantic.
Table 1. Previous Atlantic Ocean record locations of G. polygonius
Location Publication
Brazil Floeter et al. (2003)
Frota et al. (2004)
Olavo et al. (2011)
Canary Islands Brito (1991)
Uiblein et al. (1996)
Brito et al. (2002)
Cape Verde Pereira et al. (2012)
Wirtz et al. (2013)
Hanel & John (2015)
Menezes et al. (2015)
Madeira Island Wirtz et al. (2008)
Trindade Island Gasparini & Floeter (2001)
Simon et al. (2013)
St. Helena Böhlke et al. (1989)
Given the wide distribution of G. polygonius on both sides of the tropical and sub-tropical Atlan-tic, it is not unexpected that the species would also be found at the mid-Atlantic, Ascension Is-land. However, the new record does highlight the opportunity and necessity for further investigation of the marine environment and communities around Ascension Island, particularly when con-sidering the island’s unique assemblage with spe-cies sourced from across the Atlantic.
ACKNOWLEDGEMENTS
The authors are grateful to Mr Billy Richards (Ascension Island) for donation of the specimen. A. Richardson is funded by a Darwin Initiative grant to the Ascension Island Government Conservation Department (DPLUS021: Ascen-sion Island Marine Sustainability (AIMS) – A Fisheries and Marine Biodiversity Project). The author is grateful for helpful discussions with the other AIMS team members and supervisory team.

Gymnothorax polygonius in Ascension Island
85
85
REFERENCES
Böhlke, E. B., J. E. McCosker & J. E. Böhlke 1989. Family Muraenidae. Pp. 104-206 in: Böhlke, E.B. (Ed.) Fishes of the Western North Atlantic. Part 9, vol. 1. Orders Anguilliformes and Saccopharyngiformes. Memoir Sears Foundation for Marine Research. New Haven. 655 pp.
Brito, A. 1991. Catálogo de los peces de las Islas Canarias. Francisco Lemus Editor, La Laguna. 230 pp.
Brito, A., P. J. Pascual, J. M. Falcón, A. Sancho & G. González 2002. Peces de las Islas Canarias. Catálogo comentado e ilustrado. Francisco Lemus Editor, La Laguna. 419 pp.
Cadenat, J. & E. Marchal 1963. Poissons. Résultats des campagnes océanographiques de la Reine-Pokou aux îles Sainte-Héléne et Ascension. Bulletin de l’Institut Francais d’Afrique Noire (A) Sciences Naturelles 25: 1235–1315
Floeter, S.R. & J.L. Gasparini 2003. The southwestern Atlantic reef fish fauna: composition and zoogeographic patterns. Journal of Fish Biology 56: 1099-1114
Frota, L.O., P.A.S. Costa & A.C. Braga 2004. Length-weight relationships of marine fishes from the central Brazilian coast. NAGA, WorldFish Center Quarterly 27: 20-26.
Gasparini, J.L. & S.R. Floeter 2001 The shore fishes of Trindade Island, western South Atlantic. Journal of Natural History 35: 1639-1656.
Hanel, R. & John, H.C. 2015. A revised checklist of Cape Verde Islands sea fishes. Journal of Applied Ichthyology 31: 135-169.
Lubbock, R. 1980. The shore fishes of Ascension Island. Journal of Fish Biology 17: 283-303.
Menezes, G.M., O. Tariche, M.R. Pinho, M.F. Sigler & H.M. Silva 2015. Structure and zonation of demersal and deep-water fish assemblages off the Cape Verde archipelago (northeast-Atlantic) as sampled by baited longlines. Deep Sea Research Part I. 102: 118-134.
Olavo, G. P.A. Costa, A.S. Martins & B.P. Ferreira 2011. Shelf edge reefs as priority areas for conservation of reef fish diversity in the tropical Atlantic. Aquatic Conservation: Marine and Freshwater Ecosystems 21: 199-209.
Pereira, J.N., A. Simas, A. Rosa, A. Aranha, S. Lino, E. Constantino, V. Monteiro, O. Tariche & G. Menezes 2012. Weight–length relationships for 27 demersal fish species caught off the Cape Verde archipelago (eastern North Atlantic). Journal of Applied Ichthyology 28: 156-159.
Simon, T., R.M. Macieira & J.C. Joyeux 2013. The shore fishes of the Trindade–Martin Vaz insular complex: an update. Journal of Fish Biology 82: 2113-2127.
Smith, D.G. 2012. A checklist of the moray eels of the world (Teleostei: Anguilliformes: Muraenidae). Zootaxa 3474: 1-64.
Uiblein, F., F. Bordes & R. Castillo 1996. Diversity, abundance and depth distribution of demersal deepwater fishes off Lanzarote and Fuerteventura, Canary Islands. Journal of Fish Biology 49. Supplement sA: 75-90.
Wirtz, P., R. Fricke & M.J. Biscoito 2008. The coastal fishes of Madeira Island–new records and an annotated check-list. Zootaxa 1715: 1-26.
Wirtz, P., A. Brito, J.M. Falcón, R. Freitas, R. Fricke, V. Monteiro, F. Reiner & O. Tariche 2013. The coastal fishes of the Cape Verde Islands–new records and an annotated check-list. Spixiana 36: 113-142.
Wirtz, P., J. Bingeman, J. Bingeman, R. Fricke, T.J. Hook & J. Young 2014. The fishes of Ascension Island, central Atlantic Ocean – new records and an annotated checklist. Journal of the Marine Biological Association of the United Kingdom, 16 pp.
Received 9 Jul 2015. Accepted 21 Jul 2015, Published online 14 Sep 2015.

87
Weissmann, J. & H. Schaefer 2015. Honeybees (Apis mellifera) collect latex of Azores bellflowers (Azorina vidalii, Campanulaceae). Arquipelago. Life and Marine Sciences 32: 87-89.
Julie A. Weissmann (email: [email protected]) and Hanno Schaefer, Technische Universität München, Dept. Ecology & Ecosystem Management, Plant Biodiversity Re-search, Emil-Ramann Str. 2, D-85354 Freising, Germany.
The Azores bellflower (Azorina vidalii (H.C.Wats.) Feer, synonym: Campanula vidalii H.C.Wats.) is a small shrub in the bellflower family, Campanulaceae, which is endemic to the Azores archipelago. Its habitats are coastal rocks and cliffs, often with elevated nitrogen levels (sewage water, rubbish) and close to settlements. Azorina is found on all nine islands of the Azores archipelago but is most common on the western islands of Flores and Corvo (Schaefer 2003). The pollination biology of Azorina is still a bit of a mystery: while flower morphology would fit best to bird pollination, birds have never been ob-served visiting its flowers and today’s avifauna of the archipelago does not include any obvious candidate species. The first author therefore per-formed an extensive field study in the summer of 2015, mainly on Corvo but also on Flores and Pico islands, in order to identify the pollinators of this enigmatic endemic. During this fieldwork, we observed on August 6 at the beach of Fajã Grande, Flores Island, sev-eral honeybees on an old Azorina plant. The bees did not visit the open flowers of the plant but instead went directly for the lower parts of the stems. They landed on the stem and then searched for small wounds where latex had recently been produced by the plant. They actively collected the sticky substance with their proboscis, middle and hind legs (Fig. 1a) and then flew off, often having trouble to get away from the gluey surface
(Fig. 1b). During 30 minutes of observation time (5:58-6:28 PM), we saw five honeybee workers collecting latex. No other insects showed interest in the latex, even though the flowers received visits by bumblebees and several dipterans. Bellflower latex coagulates very rapidly (Bauer et al. 2014) and its main function is probably sealing of wounds and defence against herbivores (Konno 2011). Sytwala et al. (2015) analysed latex of Azorina and 22 other Campanulaceae and found in all but one species protease activity, more specifically serine proteases. This indicates that the latex has antimicrobial properties. Back in the hive, the bees very likely turned the latex into propolis, a mixture of plant substances, bees wax and pollen used mainly by honeybees, sting-less bees and megachilid bees (Willmer 2011) to coat the inside of the hive and cells and anything that cannot be transported outside the hive, e.g. carcasses of small mammals. Propolis is the main way for a bee colony to achieve social immunity (Simone-Finstrom & Spivak 2010). The bees collect a variety of plant exudates including cuticular waxes, mucilages, gums, resins, and latices. Propolis varies considerably in compo- sition depending on the locally available plants but has usually significant antibacterial, antifun-gal, antiviral, cytotoxic, antioxidant and anti-inflammatory activity (Bankova et al. 2014; Kujumgiev et al. 1999).

Apis mellifera collects latex from Azorina vidalii
87
87
Weissmann, J. & H. Schaefer 2015. Honeybees (Apis mellifera) collect latex of Azores bellflowers (Azorina vidalii, Campanulaceae). Arquipelago. Life and Marine Sciences 32: 87-89.
Julie A. Weissmann (email: [email protected]) and Hanno Schaefer, Technische Universität München, Dept. Ecology & Ecosystem Management, Plant Biodiversity Re-search, Emil-Ramann Str. 2, D-85354 Freising, Germany.
The Azores bellflower (Azorina vidalii (H.C.Wats.) Feer, synonym: Campanula vidalii H.C.Wats.) is a small shrub in the bellflower family, Campanulaceae, which is endemic to the Azores archipelago. Its habitats are coastal rocks and cliffs, often with elevated nitrogen levels (sewage water, rubbish) and close to settlements. Azorina is found on all nine islands of the Azores archipelago but is most common on the western islands of Flores and Corvo (Schaefer 2003). The pollination biology of Azorina is still a bit of a mystery: while flower morphology would fit best to bird pollination, birds have never been ob-served visiting its flowers and today’s avifauna of the archipelago does not include any obvious candidate species. The first author therefore per-formed an extensive field study in the summer of 2015, mainly on Corvo but also on Flores and Pico islands, in order to identify the pollinators of this enigmatic endemic. During this fieldwork, we observed on August 6 at the beach of Fajã Grande, Flores Island, sev-eral honeybees on an old Azorina plant. The bees did not visit the open flowers of the plant but instead went directly for the lower parts of the stems. They landed on the stem and then searched for small wounds where latex had recently been produced by the plant. They actively collected the sticky substance with their proboscis, middle and hind legs (Fig. 1a) and then flew off, often having trouble to get away from the gluey surface
(Fig. 1b). During 30 minutes of observation time (5:58-6:28 PM), we saw five honeybee workers collecting latex. No other insects showed interest in the latex, even though the flowers received visits by bumblebees and several dipterans. Bellflower latex coagulates very rapidly (Bauer et al. 2014) and its main function is probably sealing of wounds and defence against herbivores (Konno 2011). Sytwala et al. (2015) analysed latex of Azorina and 22 other Campanulaceae and found in all but one species protease activity, more specifically serine proteases. This indicates that the latex has antimicrobial properties. Back in the hive, the bees very likely turned the latex into propolis, a mixture of plant substances, bees wax and pollen used mainly by honeybees, sting-less bees and megachilid bees (Willmer 2011) to coat the inside of the hive and cells and anything that cannot be transported outside the hive, e.g. carcasses of small mammals. Propolis is the main way for a bee colony to achieve social immunity (Simone-Finstrom & Spivak 2010). The bees collect a variety of plant exudates including cuticular waxes, mucilages, gums, resins, and latices. Propolis varies considerably in compo- sition depending on the locally available plants but has usually significant antibacterial, antifun-gal, antiviral, cytotoxic, antioxidant and anti-inflammatory activity (Bankova et al. 2014; Kujumgiev et al. 1999).

Weissmann & Schaefer
88
89
Because of this, propolis has also been used for centuries in traditional medicine as remedy to treat e.g., wounds, burns, and sore throats (Bankova 2005). Phytochemical analyses of propolis and the determination of its pollen con-tent can help to classify it in different types (Bankova et al. 2014; Kumazawa et al. 2003). Since different plants can be used by the bees depending on the local flora and the season, the composition and activity of propolis and propolis products are in general quite variable (Bankova et al. 2014) and detailed knowledge about the source plants is important. Falcão et al. (2013) compared propolis samples from different parts of Portugal including Azores and Madeira and found them to differ significantly from region to region with Azores samples especially rich in flavones. We hypothesize that this difference is at least partly due to Azorean endemic plants like Azorina and perhaps the Azores juniper (Juniperus brevifolia) as latex and resin sources and encourage for more bee observation studies because this direct evi-dence is the most reliable approach to understand different composition and properties of propolis.
ACKNOWLEDGEMENTS
We thank the Direcção Regional do Ambiente of the Azores for research permit no. SAI/DRA/2015/2116, proc. 116.14.06/62, Tânia Pipa, Barbara Ambros, Carlos Silva and Nuno Oliveira (SPEA) for help with logistics and all sorts of challenges on Corvo, and Fernando Ferreira (Parque Natural da Ilha do Corvo) for general support.
REFERENCES
Bankova, V. 2005. Recent trends and important developments in propolis research. eCAM 2: 29-32.
Bankova, V., M. Popova & B. Trusheva 2014. Propolis volatile compounds: chemical diversity and biological activity: a review. Chemistry Central Journal 8: 28.
Bauer, G., S. N. Gorb, M.- C. Klein, A. Nellesen, M. von Tapavicza & T. Speck 2014. Comparative study on plant latex particles and latex coagulation in Ficus benjamina, Campanula glomerata and three Euphorbia species. PLoS ONE 9(11): e113336. doi:10.1371/journal.pone.0113336
Falcão, S.I., A. Tomás, N. Vale, P. Gomes, C. Freire & M. Vilas-Boas 2013. Phenolic quantification and botanical origin of Portuguese propolis. Industrial Crops and Products 49: 805-812.
Konno, K. 2011. Plant latex and other exudates as plant defense systems: Roles of various defense chemicals and proteins contained therein. Phytochemistry 72: 1510-1530.
Kujumgiev, A., I. Tsvetkova, Y. Serkedjieva, V. Bankova, R. Christov. & S. Popov 1999. Antibacterial, antifungal and antiviral activity of propolis from different geographic origin. Journal of Ethnopharmacy 64: 235-240.
Kumazawa, S., M. Yoneda, I. Shibata, J. Kanaeda,
T.
Hamasaka & T. Nakayama 2003. Direct evidence for the plant origin of Brazilian propolis by the observation of Honeybee behavior and phytochemical analysis. Chemical and Pharmaceutical Bulletin 51: 740-742.
Schaefer, H. 2003. Chorology and diversity of the Azorean flora. Dissertationes Botanicae 374: part I: 1-130, part II: 1-580.
Simone-Finstrom, M. & M. Spivak 2010. Propolis and bee health: the natural history and significance of resin use by honey bees. Apidologie 41: 295-311.
Sytwala, S., A. Domsalla & M.F. Melzig 2015. Investigation of plant latices of Asteraceae and Campanulaceae regarding proteolytic activity. Plant Physiology and Biochemistry 97: 117-123.
Willmer, P. 2011. Pollination and Floral Ecology. Princeton University Press, 778 pp.
Received 13 Nov 2015. Accepted 19 Nov 2015, Published online 26 Nov 2015.
88
Fig. 1. a) Honeybee worker collecting latex on old stem of Azorina vidalii, Flores Island, Azores; b) Honeybee worker trying to fly off an Azorina vidalii stem with proposcis still attached to the plant stem by white latex string (photographs by J.A. Weissmann).

Apis mellifera collects latex from Azorina vidalii
89
89
Because of this, propolis has also been used for centuries in traditional medicine as remedy to treat e.g., wounds, burns, and sore throats (Bankova 2005). Phytochemical analyses of propolis and the determination of its pollen con-tent can help to classify it in different types (Bankova et al. 2014; Kumazawa et al. 2003). Since different plants can be used by the bees depending on the local flora and the season, the composition and activity of propolis and propolis products are in general quite variable (Bankova et al. 2014) and detailed knowledge about the source plants is important. Falcão et al. (2013) compared propolis samples from different parts of Portugal including Azores and Madeira and found them to differ significantly from region to region with Azores samples especially rich in flavones. We hypothesize that this difference is at least partly due to Azorean endemic plants like Azorina and perhaps the Azores juniper (Juniperus brevifolia) as latex and resin sources and encourage for more bee observation studies because this direct evi-dence is the most reliable approach to understand different composition and properties of propolis.
ACKNOWLEDGEMENTS
We thank the Direcção Regional do Ambiente of the Azores for research permit no. SAI/DRA/2015/2116, proc. 116.14.06/62, Tânia Pipa, Barbara Ambros, Carlos Silva and Nuno Oliveira (SPEA) for help with logistics and all sorts of challenges on Corvo, and Fernando Ferreira (Parque Natural da Ilha do Corvo) for general support.
REFERENCES
Bankova, V. 2005. Recent trends and important developments in propolis research. eCAM 2: 29-32.
Bankova, V., M. Popova & B. Trusheva 2014. Propolis volatile compounds: chemical diversity and biological activity: a review. Chemistry Central Journal 8: 28.
Bauer, G., S. N. Gorb, M.- C. Klein, A. Nellesen, M. von Tapavicza & T. Speck 2014. Comparative study on plant latex particles and latex coagulation in Ficus benjamina, Campanula glomerata and three Euphorbia species. PLoS ONE 9(11): e113336. doi:10.1371/journal.pone.0113336
Falcão, S.I., A. Tomás, N. Vale, P. Gomes, C. Freire & M. Vilas-Boas 2013. Phenolic quantification and botanical origin of Portuguese propolis. Industrial Crops and Products 49: 805-812.
Konno, K. 2011. Plant latex and other exudates as plant defense systems: Roles of various defense chemicals and proteins contained therein. Phytochemistry 72: 1510-1530.
Kujumgiev, A., I. Tsvetkova, Y. Serkedjieva, V. Bankova, R. Christov. & S. Popov 1999. Antibacterial, antifungal and antiviral activity of propolis from different geographic origin. Journal of Ethnopharmacy 64: 235-240.
Kumazawa, S., M. Yoneda, I. Shibata, J. Kanaeda,
T.
Hamasaka & T. Nakayama 2003. Direct evidence for the plant origin of Brazilian propolis by the observation of Honeybee behavior and phytochemical analysis. Chemical and Pharmaceutical Bulletin 51: 740-742.
Schaefer, H. 2003. Chorology and diversity of the Azorean flora. Dissertationes Botanicae 374: part I: 1-130, part II: 1-580.
Simone-Finstrom, M. & M. Spivak 2010. Propolis and bee health: the natural history and significance of resin use by honey bees. Apidologie 41: 295-311.
Sytwala, S., A. Domsalla & M.F. Melzig 2015. Investigation of plant latices of Asteraceae and Campanulaceae regarding proteolytic activity. Plant Physiology and Biochemistry 97: 117-123.
Willmer, P. 2011. Pollination and Floral Ecology. Princeton University Press, 778 pp.
Received 13 Nov 2015. Accepted 19 Nov 2015, Published online 26 Nov 2015.

Composição — Departamento de Oceanografia e Pescas, Universidade dos Açores, HortaExecução gráfica — COINGRA, Lda. Parque Industrial da R. Grande — S. Miguel
350 ex. — 2015 — Depósito legal n.º 33098/89

Composição — Departamento de Oceanografia e Pescas, Universidade dos Açores, HortaExecução gráfica — COINGRA, Lda. Parque Industrial da R. Grande — S. Miguel
350 ex. — 2015 — Depósito legal n.º 33098/89


ARQUIPELAGO Life and Marine Sciences SCOPE ARQUIPELAGO - Life and Marine Sciences, publishes annually original scientific articles, short communications and reviews on the terrestrial and marine environment of Atlantic oceanic islands and seamounts. PUBLISHER University of the Azores Rua da Mãe de Deus, 13A PT – 9501-801 Ponta Delgada, Azores, Portugal. EDITOR Helen Rost Martins Phone: + 351 292 207 400 / 428 - Fax: +351 292 207 811 E-mail: [email protected] INTERNET RESOURCES http://www.arquipelago.info Journal information, instructions to authors and free access to all papers. FINANCIAL SUPPORT Secretaria Regional do Mar, Ciência e Tecnologia, do Governo dos Açores e OMA – Observatório do Mar dos Açores, Horta.
EDITORIAL SECRETARIAT Helen R. Martins, Ruth Higgins, José Gomes-Pereira, Emmanuel Arand. EDITORIAL COMMITTEE Paulo A.V. Borges, Angra do Heroísmo; José Azevedo, Ponta Delgada; João M. Gonçalves, Horta.
ADVISORY BOARD Miguel A. Alcaraz, Barcelona, Spain; Alan B. Bolten, Florida, USA; António B. de Sousa, Lisboa, Portugal; Richard D.M. Nash, Bergen, Norway; Erik Sjögren, Uppsala, Sweden; Charles H.J.M. Fransen, Leiden, Netherlands; George R. Sedberry, Georgia, USA; Hanno Schäfer, Munich, Germany; Tony Pitcher, Vancouver, Canada; João Canning Clode, Funchal, Portugal.
Indexed in: Aquatic Sciences and Fisheries Abstracts (ASFA), BIOSIS Previews,
Zoological Record, Directory of Open Access Journals (DOAJ), Web of Science.
Cover design: Emmanuel Arand. Photo: Myliobatis aquila, Faial Island - Nuno Vasco-Rodrigues.
EXCHANGEThe journal is distributed on exchange basis through the editor.
PURCHASE Separate numbers of the Journal can be purchased from the publisher.
OFFPRINTS Authors receive PDF files of their papers. Orders of paper copies can be sent with the proofs if desired.
INSTRUCTIONS TO AUTHORS General. MSS should be submitted to the editor preferably by e-mail ([email protected]). The text should be ready for printing and carefully checked for all errors, including linguistic ones, before submission. English is the recommended language. Choose Font Times New Roman, size 12, double spaced paragraph, and insert page numbers. MSS that are not in accordance with these and the following instructions will be returned for revision. All MSS will be peer-reviewed by two referees prior to their acceptance by the Editor.
Style. 1) Title and 2) Author's name, followed by 3) Abstract (not exceeding 200 words), up to five key words, and (4) the author's name and professional postal address; 5) The main text; 6) Acknowledgements; 7) References; 8) Appendices; 9) Tables with legends; and the 10) List of figures with legends. For research articles the text (5) must be organized as follows: Introduction; Material and methods; Results; Discussion. The Abstract (3) should convey the scope of the paper, emphasizing the results and conclusions, not merely describing its contents. Limit the authors’ professional address (4) to official filiations only.
References. (7) Should correspond with the examples given below:
a) Book: Etgen, W.M. & P.M. Reaves 1978. Dairy Cattle Feeding and Management (6th edition). John Wiley & Sons Inc. New York. 638 pp.
b) Chapter from a book: O’Dor, R., H. O. Pörtner & R. E. Shadwick 1990. Squid as elite athletes: locomotory, respiratory, and circulatory integration. Pp. 481-503 in: Gilbert, D.L., W.J. Adelman & J.M. Arnold (Eds). Squid As Experimental Animals. Plenum Press, New York-London. 516 pp.
c) Article from a journal: Bentley, M.G., P.J.W. Olive, P.R. Garwood & N.H. Wright 1984. The spawning and spawning mechanism of Nephtys caeca (Fabricius, 1780) and Nephtys homebergi Savigny, 1818 (Annelida: Polychaeta). Sarsia 69: 63-68.
d) Electronic article, from online-only Journal: Woo, K.L. 2006. Testing Visual Sensitivity to the Speed and Direction of Motion in Lizards. Journal of Visualized Experiments [Internet]. Available from: http://www.jove.com/index/details.stp?id=127 (cited 18 February 2007).
Use ampersand (&) for all joint authorships in the reference list and for double authorships in the text. Use “et al.” in the text for references with three or more authors and in the reference list for more than seven authors. Avoid multiple citations in text, preferably including the earliest and most recent work. The citation list should be ordered alphabetically by author’s name and year of publication, followed by increasing number of co-authors. Journal names should be written in full.
Figures and Tables. All figures and large tables should be presented with Font Times New Roman, and separately. Photographs should be clear, have high contrast, and in digital format (tiff or jpg) with 300 dpi, with at least 15x15 cm. No original figures should be submitted before the MS has been accepted.
E-mail Attachment. All text file formats are accepted. Attachments must be identified with the author’s last name, the file name and extension, as follows: Bianchi_text.doc, Bianchi_fig1.tif, Bianchi_table1.xls.
For a complete list of format specifications and instructions to authors consult our website at: http://www.arquipelago.info

ISSN 0873-4704
ARQUIPELAGO - Life and Marine Sciences
No. 32 – 2015
CONTENTS: PAGE GABRIEL, DANIELA, W.E. SCHMIDT, D.M. KRAYESKY, D.J. HARRIS & S. FREDERICQ The crustose red algal genus Peyssonnelia (Peyssonneliales, Rhodophyta) in the Azores: from five to one species 1 PAIVA, L.S., E.M.C. LIMA, A.I. NETO & J.A.B. BAPTISTA Screening for angiotensin I-converting enzyme (ACE) inhibitory activity of enzymatic hydrolysates obtained from Azorean macroalgae 11 NUNES, R., R. GABRIEL, R.B. ELIAS, F. RIGAL, A.O. SOARES, P. CARDOSO & P.A.V. BORGES Arthropods and other Biota associated with the Azorean Trees and Shrubs: Juniperus brevifolia 19 GOMES-PEREIRA, J.N., I. TOJEIRA, L.P. RIBEIRO, R.S. SANTOS, F. DIAS & F. TEMPERA First in situ observations of soft bottom megafauna from the Cascais Canyon head 67
SHORT COMMUNICATIONS: LINO, S., J.R. XAVIER, R.S. SANTOS & A. COLAÇO Sponge perforating lace coral with anticancer activity 75
AFONSO, P. & N. VASCO-RODRIGUES Summer aggregations of the common eagle ray, Myliobatis aquila 79 RICHARDSON, A.J. & A. BRITO The polygon moray, Gymnothorax polygonius (Poey, 1875): A new record from Ascension Island, South Atlantic 83 WEISSMANN, J. & H. SCHAEFER Honeybees (Apis mellifera) collect latex of Azores bellflowers (Azorina vidalii, Campanulaceae) 87
ARQ
UIPELA
GO
- LIFE AN
D M
ARIN
E SCIEN
CES - N
O. 32 - 2015


















