
Architecture of Trees in the Rain Forest of morobe district, new guinea
8
Architecture of Trees in the Rain Forest of Morohe District, New Guinea Francis Halle institut de Botanique, Universite de Montpellier, France ABSTRACT The architectural spectrum of the rain-forest appears to be qualitatively the same in New Guinea as previously re- ported in tropical Africa or America. However, the study of Asiatic and Melanesian trees leads to a broader understand- ing of the architectural range in pantropical families of flowering plants, and suggests an approach to the understanding of many problems in taxonomy, phylogeny, biogeography, and forest ecology. The architecture of trees in the tropics, a new subject for botanical research, has already been ap- plied to taxonomy, as, for example, in the Euphor- biaceae (Halle 1971). In the present paper, I wish to introduce another aspect of tree architecture, which is biogeographical rather than taxonomical. The introduction of the topic of tree architecture by Halle and Oldeman (1970) was mainly founded upon African and American records, and was com- paratively poor in original data about Asiatic trees. Taking into account the particular richness of the Asiatic flora, and its well-known importance for tropical botany, filling such a gap was a preliminary essential for the satisfactory formulation of the prob- lem of architectural evolution in trees. A visit to Australian New Guinea in 1972 provided an op- portunity to make a first study of the architecture of Asiatic and Melanesian trees, the main results of which are summarized below.1 All the architectural models previously dis- covered in Africa and America were found easily in the lowland rain forest around Lae, and in the sub- montane rain forest around Bulolo, Morobe District. A significant, if negative, result is that— contrary to a priori views—no new architectural models were found during my visit to Asia. Thus, as far as I know, the architectural spectrum of the rain forest appears to be qualitatively the same in all parts of the tropics. Of course, the possibility of quantita- tive variation within the spectrum cannot be denied, some models being favored in some countries either for historical reasons or by environmental condi- tions, but this supposition is speculative, as a study which would reveal this still remains to be done. 11 wish to express my gratitude to John S. Womersley, head of the Division of Botany at Lae, to the members of his staff, particularly Mark J. E. Coode, for their help in my field-work, and to Anna Cook and Bokinane, for their assistance. Even if the total number of known models has not been augmented, this study of Asian trees leads to a better understanding of the architectural range in several pantropical families of flowering plants. From the 76 New Guinea species from 45 families whose form was studied, I extract 11 which were observed in particularly good circumstances. Schuwmcmsia heningsii K. Schum. OCHNACEAE Cf. NGF 40163 from Red Hill, near Lae, Morobe District. Leeuwenberg’ s Model. This very common small tree, usually less than 10 meters high, is one of the finest plants around Lae at flowering time. Figure 1 a and b represents a well-grown flowering specimen. All the serial axes are orthotropic and equivalent to one another, the only variation being a quantitative one such that, when the branching increases, the size of the suc- cessive axes, and the length of their leaves, progres- sively decreases, as advanced by Corner (1949) in his Durian Theory’ . Apical inflorescences appear simultaneously on all the upper axes, the lower ones remaining sterile from then on. Owing to these architectural features, Schuurmansia heningsii be- longs to Leeuwenberg’ s model, as defined by Halle and Oldeman (1970). Apart from the characteris- tics of the model itself, Schummansia heningsii of- fers some individual morphogenetic features, such as the stilt-roots supporting the seedling axis, and the periodic growth which is indicated by the series of bud-scale scars at each level of temporarily inter- rupted growth. This is the first record of Leeuwen- berg’ s model in Ochnaceae. However, in the same family, the African Lophira data Banks possesses an architecture related to the present one, although its aerial axes, after the first growth, are no longer equivalent to one another, since some of them grow upright to form a s’ .mpodial trunk (Kwan Koriba’ s model). BIOTROPICA 6(1): 43-50 1974 43
-
Upload
arbor-culture -
Category
Documents
-
view
75 -
download
1
Transcript of Architecture of Trees in the Rain Forest of morobe district, new guinea

Architecture of Trees in the Rain Forest of Morohe District, New GuineaFrancis Halle
institut de Botanique, Universite de Montpellier, France
ABSTRACTThe architectural spectrum of the rain-forest appears to be qualitatively the same in New Guinea as previously reported in tropical Africa or America. However, the study of Asiatic and Melanesian trees leads to a broader understanding of the architectural range in pantropical families of flowering plants, and suggests an approach to the understanding of many problems in taxonomy, phylogeny, biogeography, and forest ecology.
The a r ch i t e c tu r e o f t re e s in the tropics, a new subject for botanical research, has already been applied to taxonomy, as, for example, in the Euphor- biaceae (Halle 1971). In the present paper, I wish to introduce another aspect of tree architecture, which is biogeographical rather than taxonomical. The introduction of the topic of tree architecture by Halle and Oldeman (1970) was mainly founded upon African and American records, and was comparatively poor in original data about Asiatic trees. Taking into account the particular richness of the Asiatic flora, and its well-known importance for tropical botany, filling such a gap was a preliminary essential for the satisfactory formulation of the problem of architectural evolution in trees. A visit to Australian New Guinea in 1972 provided an opportunity to make a first study of the architecture of Asiatic and Melanesian trees, the main results of which are summarized below.1
All the architectural models previously discovered in Africa and America were found easily in the lowland rain forest around Lae, and in the submontane rain forest around Bulolo, Morobe District. A significant, if negative, result is that— contrary to a priori views— no new architectural models were found during my visit to Asia. Thus, as far as I know, the architectural spectrum of the rain forest appears to be qualitatively the same in all parts of the tropics. Of course, the possibility of quantitative variation within the spectrum cannot be denied, some models being favored in some countries either for historical reasons or by environmental conditions, but this supposition is speculative, as a study which would reveal this still remains to be done.
11 wish to express my gratitude to John S. Womersley, head of the Division of Botany at Lae, to the members of his staff, particularly Mark J. E. Coode, for their help in my field-work, and to Anna Cook and Bokinane, for their assistance.
Even if the total number of known models has not been augmented, this study of Asian trees leads to a better understanding of the architectural range in several pantropical families of flowering plants. From the 76 New Guinea species from 45 families whose form was studied, I extract 11 which were observed in particularly good circumstances.Schuwmcmsia heningsii K. Schum. OCHNACEAE Cf. NGF 40163 from Red Hill, near Lae, Morobe District. Leeuwenberg’s Model.This very common small tree, usually less than 10 meters high, is one of the finest plants around Lae at flowering time. Figure 1 a and b represents a well-grown flowering specimen. All the serial axes are orthotropic and equivalent to one another, the only variation being a quantitative one such that, when the branching increases, the size of the successive axes, and the length of their leaves, progressively decreases, as advanced by Corner (1949) in his Durian Theory’. Apical inflorescences appear simultaneously on all the upper axes, the lower ones remaining sterile from then on. Owing to these architectural features, Schuurmansia heningsii belongs to Leeuwenberg’s model, as defined by Halle and Oldeman (1970). Apart from the characteristics of the model itself, Schummansia heningsii offers some individual morphogenetic features, such as the stilt-roots supporting the seedling axis, and the periodic growth which is indicated by the series of bud-scale scars at each level of temporarily interrupted growth. This is the first record of Leeuwenberg’s model in Ochnaceae. However, in the same family, the African Lophira data Banks possesses an architecture related to the present one, although its aerial axes, after the first growth, are no longer equivalent to one another, since some of them grow upright to form a s’.mpodial trunk (Kwan Koriba’s model).
BIOTROPICA 6(1): 43-50 1974 43

44 Halle

Barringtonia edulis Seem. LECYTHIDACEAE Botanical Gardens. Lae, S 2752.Leeuwenberg’s Model.An interesting problem is raised by the existence of the same architectural model in a quite unrelated family. Figure 2 shows Barringtonia edulis, from New Britain, New Ireland, and New Guinea (Ma- dang), growing in the Lae Botanical Gardens. This small tree, usually less than 15 meters high, is easy to recognize by its large pendulous apical inflorescences. Besides the basic features of Leeuwenberg’s model, the tree displays an obvious periodic growth; each aerial axis is built by a succession of flushes, the first ones remaining vegetative, the last one becoming inflorescential. According to Payens (1967), periodic growth is a constant feature in the genus Barringtonia which contains about 40 species. Apart from the lack of stilt-roots, the architecture is nearly the same as in Schuurmansia\ however, as there is no link between appearance and architecture, these two trees are physiognomically quite different. This is the first record of Leeuwenberg’s model in Lecy- thidaceae; although, the African Cornbretodendron africanum (Welw.) Exell. belongs to the Kwan- Koriba model.
Barringtonia calyptrocalyx K. Schum. LECYTHIDACEAECf. Botanical Gardens Lae, T. 872. F. Halle 1944, Garagos River, Morobe District. Corner’s Model.It was surprising to discover the same architecture in two unrelated families; it is no less strange that two qualitatively different models can be found within the same genus. Barringtonia calyptrocalyx is a small tree of the submontane rain forest along the road from Lae to Bulolo. As shown in figure 3, this species is monocaulous with lateral inflorescences, and therefore belongs to Corner’s model. Payens (1967) has already described "a cvcadoid habit bound up with cauliflory” in three species of the genus Barringtonia, all of them from New Guinea: B. papuana, B. papeh, and B. calyptrocalyx. The periodic growth of the genus Barringtonia is also evident in the monocaulous species; from the morphogenetic point of view, the only essential difference between B. edulis and B. calyptrocalyx seems to be the position of the inflorescences.
Pithecellobiu77i hansemanii (F. Muell.) Mohl. LE GUMIN OS AE-MIMOSOIDE AE NGF 8677, J. S. Womersley and L. J. Brass Norman by Island, Papua.NGF 10468, K. J. White, Oomsis, Morobe District New Guinea.Botanical Gardens Lae, E 2005. Corner’s Model. Corner’s model is always present in the tropica rain forest, and is even common in some countries such as Gabon (Halle and Halle 1965). In Ne^ Guinea, monocaulous trees seem to be particular! frequent. Two more examples may be describe in addition to that of Barringtonia calyptrocalyx The possible occurrence of monocaulous tree within the Leguminosae has been considered by Cor ner (1954). Figure 4 a and b represents a smal monocaulous tree of the subfamily Mimosoideae Pithecellobium hansemanii is cauliflorous, but, con trary to Barringtonia, there is no evidence of peri odic growth, all the leaf scars being equal. Notab! as a rare feature in Corner’s model are the long in ternodes, correlated with a slender trunk, bending a the top. The specimens of figure 4 are 7 meter tall, with a basal diameter not exceeding 5 cm ant large bipinnate leaves more than 120 cm long. It i probable that Pithecellobium hansemanii is not thi only arborescent Leguminosae belonging to thi architectural model. A tree of Halmaheira Islanc (Indonesia), Archidendron beguinii de Wit, ha been described by de W it (1942) as unbranched carrying a rich foliage and inflorescences on the ole wood. Archidendron F. Muell. is a primitive genu among the Mimosoideae, being one of the few hav ing several (5-10) carpels.
Semecarpus magnifica K. Schum. ANACARDIA CEAEF. Halle 1959, from Lae, Morobe District. Corner’ Model.This small tree, up to 10 meters high, is very com mon in the rain forest undergrowth around Lae, a low altitude. Figure 5 shows that the species obvi ously belongs to Corner’s model, with a striking peri odic growth. A similar figure has been supplied b; van Royen (1964) in his study of the Papuan anc New Guinean Anacardiaceae. Semecarpus magnifi ca is not the first record of this architectural mode
FIGURES 1—4. Figure la Schuurmansia heningsii K. Schum., Ochnaceae; a small tree, 7 m high, at flowering tim< in July. Figure lb The top of the first aerial axis, at flowering time. From the secondary forest, Lae. Figure 1 Barringtonia edulis Seem., Lecythidaceae; a young tree, 2 m high, with the large pendulous apical inflorescences Botanical Gardens, Lae. Figure 3 Barringtonia calyptrocalyx K. Schum., Lecythidaceae; a small monocaulous tree 5 m high. From the submontane rain forest along the Lae-Bulolo road. Figure 4a Pithecellobium hansemanii (F Muell.) Mohl., Leguminosae-Mimosoideae; a monocaulous tree, 7 m high, with bending top. Figure 4b P. hanse manii in which the withered top has been replaced by a single shoot. Botanical Gardens, Lae.
Architecture of Trees in New Guinea 4E

Barringtonia edulis Seem. LECYTHIDACEAE Botanical Gardens, Lae, S 2752.Leeuwenberg’s Model.An interesting problem is raised by the existence of the same architectural model in a quite unrelated family. Figure 2 shows Barringtonia edulis, from New Britain, New Ireland, and New Guinea (Ma- dang), growing in the Lae Botanical Gardens. This small tree, usually less than 15 meters high, is easy to recognize by its large pendulous apical inflorescences. Besides the basic features of Leeuwenberg’s model, the tree displays an obvious periodic growth; each aerial axis is built by a succession of flushes, the first ones remaining vegetative, the last one becoming inflorescential. According to Payens (1967), periodic growth is a constant feature in the genus Barringtonia which contains about 40 species. Apart from the lack of stilt-roots, the architecture is nearly the same as in Schuurmansia\ however, as there is no link between appearance and architecture, these two trees are physiognomically quite different. This is the first record of Leeuwenberg’s model in Lecy- thidaceae; although, the African Combretodendron africanum (Welw.) Exell. belongs to the Kwan- Koriba model.
Barringtonia calyptrocalyx K. Schum. LECYTHIDACEAECf. Botanical Gardens Lae, T. 872. F. Halle 1944, Garagos River, Morobe District. Corner’s Model.It was surprising to discover the same architecture in two unrelated families; it is no less strange that two qualitatively different models can be found within the same genus. Barringtonia calyptrocalyx is a small tree of the submontane rain forest along the road from Lae to Bulolo. As shown in figure 3, this species is monocaulous with lateral inflorescences, and therefore belongs to Corner’s model. Payens (1967) has already described "a cvcadoid habit bound up with cauliflory” in three species of the genus Barringtonia. all of them from New Guinea: B. papuana. B. papeh, and B. calyptrocalyx. The periodic growth of the genus Barringtonia is also evident in the monocaulous species; from the morphogenetic point of view, the only essential difference between B. edulis and B. calyptrocalyx seems to be the position of the inflorescences.
Pithecellobium hansemanii (F. Muell.) Mohl. LE- GUMIN OS AE-MIMOSOIDE AE NGF 8677, J. S. Womersley and L. J. Brass Norman- by Island, Papua.NGF 10468, K. J. White, Oomsis, Morobe District, New Guinea.Botanical Gardens Lae, E 2005. Corner’s Model. Corner’s model is always present in the tropical rain forest, and is even common in some countries, such as Gabon (Halle and Halle 1965). In New Guinea, monocaulous trees seem to be particularly frequent. Two more examples may be described in addition to that of Barringtonia calyptrocalyx. The possible occurrence of monocaulous trees within the Leguminosae has been considered by Corner (1954). Figure 4 a and b represents a small monocaulous tree of the subfamily Mimosoideae; Pithecellobium hansemanii is cauliflorous, but, contrary to Barringtonia, there is no evidence of periodic growth, all the leaf scars being equal. Notable as a rare feature in Corner’s model are the long internodes, correlated with a slender trunk, bending at the top. The specimens of figure 4 are 7 meters tall, with a basal diameter not exceeding 5 cm and large bipinnate leaves more than 120 cm long. It is probable that Pithecellobium hansemanii is not the only arborescent Leguminosae belonging to this architectural model. A tree of Halmaheira Island (Indonesia), Archidendron beguinii de Wit, has been described by de W it (1942) as unbranched, carrying a rich foliage and inflorescences on the old wood. Archidendron F. Muell. is a primitive genus among the Mimosoideae, being one of the few having several (5-10) carpels.
Semecarpus magnifica K. Schum. ANACARDIA- CEAEF. Halle 1959, from Lae, Morobe District. Corner’s Model.This small tree, up to 10 meters high, is very common in the rain forest undergrowth around Lae, at low altitude. Figure 5 shows that the species obviously belongs to Corner’s model, with a striking periodic growth. A similar figure has been supplied by van Royen (1964) in his study of the Papuan and New Guinean Anacardiaceae. Semecarpus magnifica is not the first record of this architectural model
FIGURES 1—4. Figure la Schuurmansia heningsii K. Schum., Ochnaceae; a small tree, 7 m high, at flowering time in July. Figure lb The top of the first aerial axis, at flowrering time. From the secondary' forest, Lae. Figure 2 Barringtonia edulis Seem., Lecythidaceae; a young tree, 2 m high, with the large pendulous apical inflorescences. Botanical Gardens, Lae. Figure 3 Barringtonia calyptrocalyx K. Schum., Lecythidaceae; a small monocaulous tree, 5 m high. From the submontane rain forest along the Lae-Buloio road. Figure 4a Pithecellobium ha?isemanii (F. Muell.) Mohl., Leguminosae-Mimosoideae; a monocaulous tree, 7 m high, with bending top. Figure 4b P. hansemanii in w'hich the withered top has been replaced by a single shoot. Botanical Gardens, Lae.
Architecture of Trees in New Guinea 45

in Anacardiaceae. The rain forest of Equatorial Africa includes the monocaulous Tricboscypha fer- ruginea Engl., which displays the same periodic growth and the same cauliflory (Halle and Halle 1965).
Campnosperma brevipetiolata Volk. ANACARDIACEAENGF 3167, J. S. Womersley, from Lae, Morobe D istrict.F. Halle 1949, from Oomsis Hill, Markham River, Morobe District. Aubreville’s Model.Although belonging to the same family as Semecar- pus, Campnosperma brevipetiolata is an example of a quite different architectural model. This tall tree, up to 50 meters high, is common in the swamp- forest around Lae and grows as shown in figure 6. The branches are grouped in tiers, because of the periodic activity of the apical meristem of the mono- podial trunk; these branches are plagiotropic, hence the widely spreading crown described by van Royen (1964a). The branch is itself sympodial; each segment of the sympodium ends by bending upright, the next segment arising laterally from the curve. The inflorescences are axillary, growing from the upright slow-growing terminal part of each segment. Campnosperma paniculata typically belongs to Aubreville’s model; this is the first record in Anacardiaceae. Otherwise this model is dominant among the big trees of Sapotaceae (Palaquimn, Nispero) and Combretaceae (Terminalia).
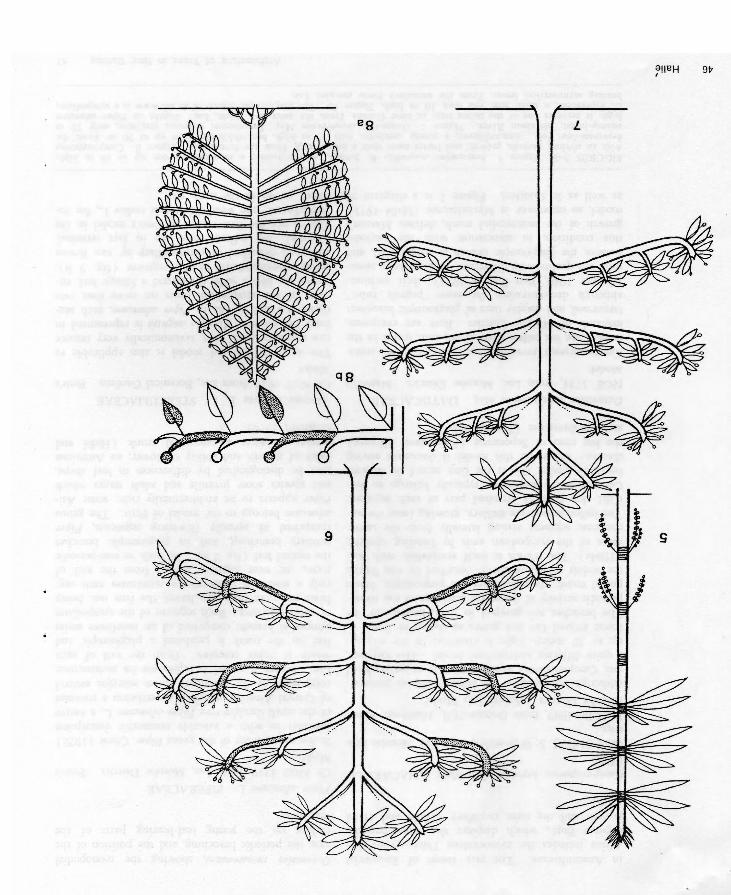
Octomeles sumatranum Miq. DATISCACEAE NGF 5724, from Lae, Morobe District. Massart’s Model.Campnosperma brevipetiolata and Octomeles sumatranum are the tallest trees of the rain forest in the lowlands of Morobe District. Both are evergreen, buttressed, and display tiers of plagiotropic branches; although demonstrating the same "pagoda habit” (Corner 1940, Van Steenis 1953), their architectures are qualitatively different. In Octomeles sumatranum, the plagiotropic branch is monopodial, and this condition, in association with the periodic growth of the monopodial trunk, defines Massart’s model, so important in Myristicaceae (Halle 1971) as well as in Conifers. Figure 7 is a diagram of
Octomeles sumatranum, showing the monopodial axis, the periodic branching, and the position of the catkins on the young leaf-bearing parts of the branches.
Piper aduncum L. PIPERACEAECf. NGF 4719, from Lae, Morobe District. Petit’sModel.In his recent study of the genus Piper. Chew (1972) provides us with a valuable taxonomic description of the small shrubby tree Piper aduncum L., a native of Central America, which is nevertheless a constant component of the secondary forest margins around Lae. Figure 8 a and b represents its architecture, which is rather complex. From the axil of each leaf on the trunk is produced a plagiotropic and sympodial branch, comprised of an indefinite series of short segments. Each segment of the sympodium bears no more than two leaves, the first one being only a scale; an inflorescence terminates each segment, the next segment arising from the axil of the second leaf (fig. 8 b). By both its non-periodic primary branching, and its plagiotropic branches comprised of apically flowering segments, Piper aduncum belongs to the model of Petit. The genus Piper appears to be architecturally rich: some Asiatic species show juvenile and adult stages which may be distinguished by differences in leaf shape, habit of growth and ability to flower; an American species possesses a sympodial trunk (Halle and Oldeman 1970).
Abroma augusta L. f. STERCULIACEAECf. NGF 9824, from Lae, Botanical Gardens. Petit’sModel.The same architectural model is also applicable to this shrub or small tree, taxonomically very remote from Piperaceae. Abroma augusta is represented in figure 9 a and b. As in Piper aduncum, each segment of the sympodium bears no more than two leaves, the first a scale, the second a foliage leaf, ensuring photosynthesis in the segment (fig. 9 b). Although described as extra-axillary by van Royen (1964b), the inflorescences are in fact terminal. This is not the first record of Petit’s model in the Sterculiaceae; the weed Waltheria indica L., for instance, grows in the same way.
FIGURES 5—8. Figure 5 Semecarpus magnified K. Schum., Anacardiaceae; a monocaulous tree up to 10 m high, with an obvious periodic growth, and leaves more than a meter long. From the forest, Lae. Figure 6 Campnosperma brevipetiolata Volk., Anacardiaceae; a young specimen, only 15 m high, but which can grow up to 50 m. From the swamp-forest, Markham River. Figure 7 Octomeles sumatranum Miq., Datiscaceae; a young specimen, only 10 m high. It becomes one of the tallest trees in New Guinea. From the secondary forest, Lae. Figure 8a Piper aduncum L., Piperaceae; a small tree, less than 10 m high. Figure 8b The plagiotropic branch of P. aduncum is a sympodium, bearing asymmetrical leaves. From the secondary forest margins, Lae.
Architecture of Trees in New Guinea 47

48 Halle

Medusanthera laxiflora (Miers.) Howard ICA- CINACEAECf. NGF 1571, from Lae, Morobe District. Roux's Model.This small tree is common in the lowland rain forest of the Morobe District; its abundance in the forest margins around the Botanical Gardens in Lae is noticeable. The architecture of Medusanthera laxiflora, as shown by figure 10, is simple. The seedling meristem produces a trunk with spirally arranged leaves; from the axil of each leaf is produced a plagiotropic and monopodial branch, which has a distichous phyllotaxis and bears axillary flowers. The above-mentioned architectural features are typically those of Roux’s model. Most of the families of Dicotyledons possess some species belonging to this model, which is one of the most frequent in the tropical rain forest, probably because of its efficiency in catching the poor light of the forest undergrowth; nevertheless, despite the architectural richness of the Icacinaceae, Medusanthera laxiflora provides the first record of this model for the family.
Citronella suaveolens (BL) Howard ICACINACEAECf. NGF 7405, from Bulolo River valley, Morobe District. Botanical Gardens Lae, S. 1863. Nozeran’s Model.The Icacinaceae are architecturally so rich that, with the study of another genus, more often than not, another model appears; this is the case, for instance, with Citronella. Citronella suaveolens is a small tree, widely distributed from Java to New Guinea (Sleumer 1969); its complex and unusual architecture is shown in Figure 11 a, b, and c. The seedling meristem produces a segment of trunk, with spirally arranged leaves, and then disappears abruptly, aborting by parenchymatization (fig. 11 b). The axillary meristems of the three most distal leaves form a tier of plagiotropic flowering branches with distichous
phyllotaxis. Upright growth can begin again, as the distal leaves of the trunk-segment possess not only one but two axillary meristems; in effect, the second axillary meristem of one of the distal leaves builds a proleptic trunk-segment, in turn ending with a new apical tier of sylleptic (Tomlinson and Gill 1973) and plagiotropic axes (fig. 11 c). This process repeats itself indefinitely. Due to its sympodial trunk, of which the segments are crowned by a plagiotropic tier, Citronella suaveolens belongs to Nozeran’s model, already described in the Euphor- biaceae (Halle 1971).
The small family Icacinaceae, with only 250 species, seems to be, from the architecmral point of view, one of the richest families of flowering plants, since seven models have so far been recognized within it.
CONCLUSIONSThrough these descriptions, I wish to suggest that tree architecture can claim to be of interest to botanists. Architectural knowledge of the tree is useful for the taxonomist, since the model is a specific stable structure^ valuable-Jor. diagnosis, and some- times more successful . than floral characters7~Ton sBeaHHdentiflcation in the tropical forest. For the evoIunomstTl^ as each familypossesses itsown architecmral spectrum, probably correlated with its evolutionary level. It is also useful for the bio- geographer as it remains to be elucidated why the same family— or even the same genus— can generate different architecmral models in different countries although with the same tropical humid climate. Finally, the value of tree architecture for the ecologist is demonstrated by Oldeman (1972). Using the Guianese forest as an example, he shows that the richness in ecological niches is determined by the rain forest architecture, which in its turn is itself dependent upon the architecture of its trees.
FIGURES 9—1L Figure 9a Abroma augusta L., Sterculiaceae; a shrubby small tree, 3 m high. Figure 9b The plagiotropic branch of A. augusta is a sympodium; the inflorescences are terminal. From the secondary forest margins, Markham River. Figure 10a Medusanthera laxiflora (Miers.) Howard, Icacinaceae; a tree, 10 m high; the axillary inflorescences arising mainly from the defoliated parts of the branches. Figure 10b Each leaf of the trunk of M. laxiflora possesses two axillary meristems; the first one, pushed aside from the axil, produces a sylleptic branch; the second one, in the middle of the axil, remains dormant. From the forest, Lae. Figure 11a Citronella suaveolens (BI.) Howard, Icacinaceae; three stages in the growth of this small tree, 10 m high. Figure 11b The abortion of the orthotropic meristem of C. suaveolens and the growth of the plagiotropic tier; the sylleptic branch is produced laterally from the axil of its subtending leaf: cf. Medusanthera. Figure 11c The orthotropic and proleptic shoot of C. suaveolens is built by the second meristem, from the middle of the axil. Botanical Gardens, Lae.
Architecture of Trees in New Guinea 49


















