
Arabidopsis Triphosphate Tunnel Metalloenzyme2 Is a Negative ...
13
Arabidopsis Triphosphate Tunnel Metalloenzyme2 Is a Negative Regulator of the Salicylic Acid- Mediated Feedback Ampli fication Loop for Defense Responses 1[W][OPEN] Huoi Ung, Wolfgang Moeder, and Keiko Yoshioka* Department of Cell and Systems Biology (H.U., W.M., K.Y.) and Center for the Analysis of Genome Evolution and Function (K.Y.), University of Toronto, Toronto, Ontario, Canada M5S 3B2 The triphosphate tunnel metalloenzyme (TTM) superfamily represents a group of enzymes that is characterized by their ability to hydrolyze a range of tripolyphosphate substrates. Arabidopsis (Arabidopsis thaliana) encodes three TTM genes, AtTTM1, AtTTM2, and AtTTM3. Although AtTTM3 has previously been reported to have tripolyphosphatase activity, recombinantly expressed AtTTM2 unexpectedly exhibited pyrophosphatase activity. AtTTM2 knockout mutant plants exhibit an enhanced hypersensitive response, elevated pathogen resistance against both virulent and avirulent pathogens, and elevated accumulation of salicylic acid (SA) upon infection. In addition, stronger systemic acquired resistance compared with wild-type plants was observed. These enhanced defense responses are dependent on SA, PHYTOALEXIN-DEFICIENT4, and NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1. Despite their enhanced pathogen resistance, ttm2 plants did not display constitutively active defense responses, suggesting that AtTTM2 is not a conventional negative regulator but a negative regulator of the amplification of defense responses. The transcriptional suppression of AtTTM2 by pathogen infection or treatment with SA or the systemic acquired resistance activator benzothiadiazole further supports this notion. Such transcriptional regulation is conserved among TTM2 orthologs in the crop plants soybean (Glycine max) and canola (Brassica napus), suggesting that TTM2 is involved in immunity in a wide variety of plant species. This indicates the possible usage of TTM2 knockout mutants for agricultural applications to generate pathogen-resistant crop plants. The triphosphate tunnel metalloenzyme (TTM) superfamily comprises a group of enzymes that are characterized by their ability to hydrolyze a range of tripolyphosphate (PPPi) substrates. All members of this superfamily use triphosphate substrates and re- quire a divalent cation cofactor for their activity, usu- ally Mg 2+ or Mn 2+ (Bettendorff and Wins, 2013). This superfamily contains two previously characterized groups of proteins: RNA triphosphatases and CYTH domain proteins (Iyer and Aravind, 2002; Gong et al., 2006). The CYTH domain was named after its two founding members, the adenylate cyclase CyaB from Aeromonas hydrophila and the mammalian thiamine triphosphatase (Iyer and Aravind, 2002). Despite low overall amino acid sequence similarity, all TTM family members possess a tunnel structure composed of eight antiparallel b-strands (b-barrel; Gallagher et al., 2006; Gong et al., 2006; Song et al., 2008; Moeder et al., 2013). The signature EXEXK motif (where X is any amino acid) located in the b-barrel has been shown to be im- portant for catalytic activity (Lima et al., 1999; Gallagher et al., 2006). The enzymatic and biological functions of most TTM family members are unknown. However, they seem to act on nucleotide and organophosphate substrates (Bettendorff and Wins, 2013) and acquired divergent biological functions in different taxonomic lineages (Iyer and Aravind, 2002). Known functions include adenylate cyclase for CyaB from A. hydrophila and Yersinia pestis adenylate cyclase IV from Y. pestis (Sismeiro et al., 1998; Gallagher et al., 2006), thiamine triphosphatase in mammals (Lakaye et al., 2004), and RNA triphospha- tase in fungi, protozoa, and some viruses (Shuman, 2002). In some instances, TTM proteins are fused to additional domains, such as a nucleotide kinase domain (Iyer and Aravind, 2002). Plants possess two types of TTM proteins: one type that comprises only the CYTH domain and another type with a CYTH domain fused to a phosphate- binding (P-loop) kinase domain (Iyer and Aravind, 2002). Arabidopsis (Arabidopsis thaliana), like most other plant species, codes for three TTM genes. We termed them AtTTM1, AtTTM2, and AtTTM3. AtTTM3 1 This work was supported by the Ontario Government (graduate student fellowships to H.U.) and by the Natural Science and Engi- neering Research Council of Canada, the Canadian Foundation for Innovation, and the Ontario Research Fund (Discovery Grant to K.Y.). * Address correspondence to [email protected]. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy de- scribed in the Instructions for Authors (www.plantphysiol.org) is: Keiko Yoshioka ([email protected]). [W] The online version of this article contains Web-only data. [OPEN] Articles can be viewed online without a subscription. www.plantphysiol.org/cgi/doi/10.1104/pp.114.248757 Plant Physiology Ò , October 2014, Vol. 166, pp. 1009–1021, www.plantphysiol.org Ó 2014 American Society of Plant Biologists. All Rights Reserved. 1009 www.plantphysiol.org on February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
Transcript of Arabidopsis Triphosphate Tunnel Metalloenzyme2 Is a Negative ...

Arabidopsis Triphosphate Tunnel Metalloenzyme2Is a Negative Regulator of the Salicylic Acid-Mediated Feedback Amplification Loop forDefense Responses1[W][OPEN]
Huoi Ung, Wolfgang Moeder, and Keiko Yoshioka*
Department of Cell and Systems Biology (H.U., W.M., K.Y.) and Center for the Analysis of Genome Evolutionand Function (K.Y.), University of Toronto, Toronto, Ontario, Canada M5S 3B2
The triphosphate tunnel metalloenzyme (TTM) superfamily represents a group of enzymes that is characterized by their abilityto hydrolyze a range of tripolyphosphate substrates. Arabidopsis (Arabidopsis thaliana) encodes three TTM genes, AtTTM1,AtTTM2, and AtTTM3. Although AtTTM3 has previously been reported to have tripolyphosphatase activity, recombinantlyexpressed AtTTM2 unexpectedly exhibited pyrophosphatase activity. AtTTM2 knockout mutant plants exhibit an enhancedhypersensitive response, elevated pathogen resistance against both virulent and avirulent pathogens, and elevated accumulationof salicylic acid (SA) upon infection. In addition, stronger systemic acquired resistance compared with wild-type plants wasobserved. These enhanced defense responses are dependent on SA, PHYTOALEXIN-DEFICIENT4, and NONEXPRESSOR OFPATHOGENESIS-RELATED GENES1. Despite their enhanced pathogen resistance, ttm2 plants did not display constitutivelyactive defense responses, suggesting that AtTTM2 is not a conventional negative regulator but a negative regulator of theamplification of defense responses. The transcriptional suppression of AtTTM2 by pathogen infection or treatment with SAor the systemic acquired resistance activator benzothiadiazole further supports this notion. Such transcriptional regulation isconserved among TTM2 orthologs in the crop plants soybean (Glycine max) and canola (Brassica napus), suggesting that TTM2 isinvolved in immunity in a wide variety of plant species. This indicates the possible usage of TTM2 knockout mutants foragricultural applications to generate pathogen-resistant crop plants.
The triphosphate tunnel metalloenzyme (TTM)superfamily comprises a group of enzymes that arecharacterized by their ability to hydrolyze a range oftripolyphosphate (PPPi) substrates. All members ofthis superfamily use triphosphate substrates and re-quire a divalent cation cofactor for their activity, usu-ally Mg2+ or Mn2+ (Bettendorff and Wins, 2013). Thissuperfamily contains two previously characterizedgroups of proteins: RNA triphosphatases and CYTHdomain proteins (Iyer and Aravind, 2002; Gong et al.,2006). The CYTH domain was named after its twofounding members, the adenylate cyclase CyaB fromAeromonas hydrophila and the mammalian thiaminetriphosphatase (Iyer and Aravind, 2002). Despite lowoverall amino acid sequence similarity, all TTM family
members possess a tunnel structure composed of eightantiparallel b-strands (b-barrel; Gallagher et al., 2006;Gong et al., 2006; Song et al., 2008; Moeder et al., 2013).The signature EXEXK motif (where X is any aminoacid) located in the b-barrel has been shown to be im-portant for catalytic activity (Lima et al., 1999; Gallagheret al., 2006).
The enzymatic and biological functions of most TTMfamily members are unknown. However, they seem toact on nucleotide and organophosphate substrates(Bettendorff and Wins, 2013) and acquired divergentbiological functions in different taxonomic lineages(Iyer and Aravind, 2002). Known functions includeadenylate cyclase for CyaB from A. hydrophila andYersinia pestis adenylate cyclase IV from Y. pestis (Sismeiroet al., 1998; Gallagher et al., 2006), thiamine triphosphatasein mammals (Lakaye et al., 2004), and RNA triphospha-tase in fungi, protozoa, and some viruses (Shuman, 2002).In some instances, TTM proteins are fused to additionaldomains, such as a nucleotide kinase domain (Iyer andAravind, 2002).
Plants possess two types of TTM proteins: one typethat comprises only the CYTH domain and anothertype with a CYTH domain fused to a phosphate-binding (P-loop) kinase domain (Iyer and Aravind,2002). Arabidopsis (Arabidopsis thaliana), like mostother plant species, codes for three TTM genes. Wetermed them AtTTM1, AtTTM2, and AtTTM3. AtTTM3
1 This work was supported by the Ontario Government (graduatestudent fellowships to H.U.) and by the Natural Science and Engi-neering Research Council of Canada, the Canadian Foundation forInnovation, and the Ontario Research Fund (Discovery Grant to K.Y.).
* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Keiko Yoshioka ([email protected]).
[W] The online version of this article contains Web-only data.[OPEN] Articles can be viewed online without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.114.248757
Plant Physiology�, October 2014, Vol. 166, pp. 1009–1021, www.plantphysiol.org � 2014 American Society of Plant Biologists. All Rights Reserved. 1009 www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from
Copyright © 2014 American Society of Plant Biologists. All rights reserved.

possesses only a CYTH domain, whereas AtTTM1 andAtTTM2 encode a nucleotide/uridine kinase domainfused to the CYTH domain (Moeder et al., 2013). Sofar, the exact biological function of TTM proteins inplants is not clear. We previously analyzed AtTTM3and found that it does not display adenylate cyclaseactivity, despite its annotation, but it does act on PPPiand with lower affinity, nucleotide triphosphates, re-leasing inorganic phosphate, similar to the TTM pro-teins from Clostridium thermocellum (CthTTM) andNitrosomonas europaea (NeuTTM; Keppetipola et al.,2007; Delvaux et al., 2011; Bettendorff and Wins, 2013;Moeder et al., 2013). Additionally, a transfer DNA(T-DNA) insertion knockout (KO) line of AtTTM3displayed a delay in root growth as well as reducedlength and number of lateral roots, suggesting a rolefor AtTTM3 in root development.
To gain insight into the biological function ofAtTTM1 and AtTTM2, we surveyed the Bio-Analyticresource (http://bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi;Winter et al., 2007) for any publicly available expressionanalysis data that might provide clues for the biologicalrole of these AtTTMs. The expression of AtTTM2 wassuppressed almost 2-fold after treatment with flg22,the well-studied pathogen-associated molecular pat-tern (PAMP) peptide, and infection with various vir-ulent and avirulent strains of Pseudomonas syringae(Supplemental Fig. S1). These data suggest the possibleinvolvement of AtTTM2 in pathogen defense responsesin plants.
The plant defense system has been studied exten-sively in the last two decades, and two levels of re-sistance responses have been reported. The first line ofdefense is basal immunity, which is triggered by therecognition of molecules that are conserved amongmany pathogens (the above-mentioned PAMPs), andthus, it is referred to as PAMP-triggered immunity.The second line of defense is a stronger response topathogen infection, which is mediated by resistance(R) genes that can recognize their cognate effectorsfrom the pathogen either directly or indirectly. Thisis known as effector-triggered immunity (Bent andMackey, 2007). The hypersensitive response (HR),which is characterized by apoptosis-like cell death atand around the site of pathogen entry, is one commondefense mechanism activated by R gene-mediatedpathogen recognition (Hammond-Kosack and Jones,1996; Heath, 2000). During HR development, an in-crease in salicylic acid (SA) and the accumulation ofpathogenesis-related (PR) proteins are observed (Vlotet al., 2008). Later, resistance against virulent patho-gens can also be seen in uninoculated systemic leaves.This phenomenon is called systemic acquired resis-tance (SAR) and confers a long-lasting, broad-rangeresistance to subsequent infection (Vlot et al., 2008;Shah and Zeier, 2013). Elevated SA levels and PR geneexpression can also be detected in uninoculated leavesthat exhibit SAR. Treatment with SA or synthetic SARactivators, such as benzothiadiazole (BTH), can alsotrigger SAR (Lawton et al., 1996; Vlot et al., 2008).
Recently, a number of metabolites that are involved inlong-distance signaling have been identified, such asmethyl salicylate, dehydroabietinal, azelaic acid, glycerol-3-P, and the Lys catabolite pipecolic acid (Shah and Zeier,2013).
Over the last two decades, significant effort has beenmade to identify components in the pathogen resis-tance signal transduction pathway. For instance, ISO-CHORISMATE SYNTHASE1 (ICS1) has been revealedto play a critical role in the biosynthesis of pathogen-induced SA. salicylic acid induction-deficient2 (sid2)/ics1mutants fail to produce elevated levels of SA afterpathogen infection and thus, are hypersensitive topathogens (Nawrath and Métraux, 1999; Wildermuthet al., 2001). NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1 (NPR1) is a key regulator of SA-mediated resistance, and npr1mutant plants fail to respondto exogenously supplied SA (Cao et al., 1994). The lipase-like proteins ENHANCED DISEASE SUSCEPTIBILITY1(EDS1) and PHYTOALEXIN-DEFICIENT4 (PAD4;Glazebrook et al., 1996; Parker et al., 1996) partici-pate in both basal and R protein-mediated defenseresponses (Falk et al., 1999; Jirage et al., 1999). EDS1
Figure 1. AtTTM2 is down-regulated after pathogen infection. A,qPCR analysis of AtTTM2 expression in H. arabidopsidis, isolateEmwa1-infected (Emwa1) or water-treated cotyledons (H2O) of 10-d-old Col-0 wild-type plants 7 d after infection. B, qPCR analysis ofAtTTM2 expression in uninfected true leaves of the same plants.Transcripts were normalized to AtEF1A. Each bar represents the meanof three independent experiments 6 SE. Each sample is a mix of 16seedlings. *, Statistical significance (Student’s t test) at P , 0.05. **,Statistical significance (Student’s t test) at P , 0.001.
1010 Plant Physiol. Vol. 166, 2014
Ung et al.
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
interacts with PAD4 and SENESCENCE ASSOCIATEDGENE101, and both EDS1 and PAD4 are required forHR formation and restriction of pathogen growth(Feys et al., 2001, 2005). A screen of mutants exhib-iting constitutive activation of resistance responsesalso identified components in defense. They show
heightened resistance, usually accompanied by ele-vated levels of SA and PR genes. These autoimmunemutants also frequently display spontaneous HR-likelesions, and thus, they are referred to as lesion mimicmutants (Moeder and Yoshioka, 2008; Hofius et al.,2009).
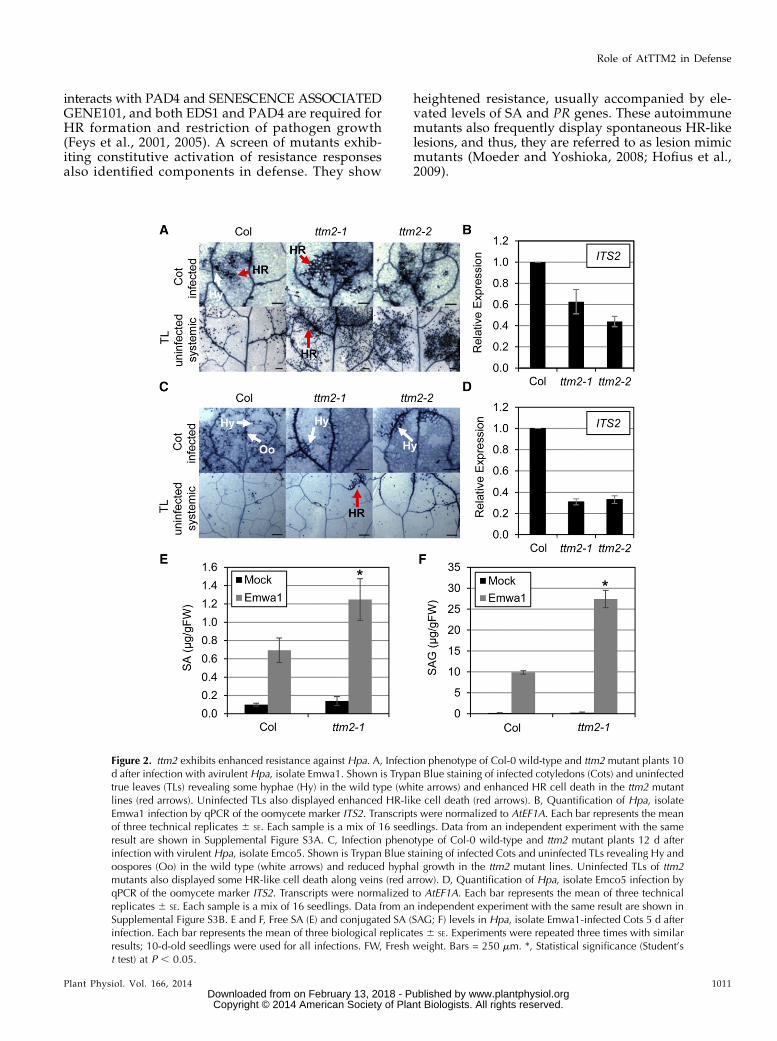
Figure 2. ttm2 exhibits enhanced resistance against Hpa. A, Infection phenotype of Col-0 wild-type and ttm2 mutant plants 10d after infection with avirulent Hpa, isolate Emwa1. Shown is Trypan Blue staining of infected cotyledons (Cots) and uninfectedtrue leaves (TLs) revealing some hyphae (Hy) in the wild type (white arrows) and enhanced HR cell death in the ttm2 mutantlines (red arrows). Uninfected TLs also displayed enhanced HR-like cell death (red arrows). B, Quantification of Hpa, isolateEmwa1 infection by qPCR of the oomycete marker ITS2. Transcripts were normalized to AtEF1A. Each bar represents the meanof three technical replicates 6 SE. Each sample is a mix of 16 seedlings. Data from an independent experiment with the sameresult are shown in Supplemental Figure S3A. C, Infection phenotype of Col-0 wild-type and ttm2 mutant plants 12 d afterinfection with virulent Hpa, isolate Emco5. Shown is Trypan Blue staining of infected Cots and uninfected TLs revealing Hy andoospores (Oo) in the wild type (white arrows) and reduced hyphal growth in the ttm2 mutant lines. Uninfected TLs of ttm2mutants also displayed some HR-like cell death along veins (red arrow). D, Quantification of Hpa, isolate Emco5 infection byqPCR of the oomycete marker ITS2. Transcripts were normalized to AtEF1A. Each bar represents the mean of three technicalreplicates 6 SE. Each sample is a mix of 16 seedlings. Data from an independent experiment with the same result are shown inSupplemental Figure S3B. E and F, Free SA (E) and conjugated SA (SAG; F) levels in Hpa, isolate Emwa1-infected Cots 5 d afterinfection. Each bar represents the mean of three biological replicates 6 SE. Experiments were repeated three times with similarresults; 10-d-old seedlings were used for all infections. FW, Fresh weight. Bars = 250 mm. *, Statistical significance (Student’st test) at P , 0.05.
Plant Physiol. Vol. 166, 2014 1011
Role of AtTTM2 in Defense
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Here, we show that AtTTM2 acts as a negative reg-ulator of plant immunity, likely at the positive ampli-fication loop of defense responses. KO mutants forAtTTM2 show enhanced pathogen resistance, whereasoverexpressors display enhanced susceptibility. The KOmutants do not show constitutive activation of defenseresponses like most autoimmune mutants but exhibitenhanced SAR on treatments with pathogens, suggest-ing that they are in a primed state. Furthermore, theexpression of TTM2 orthologs in canola (Brassica napus)and soybean (Glycine max) displays the same tran-scriptional down-regulation after BTH treatment, sug-gesting that the biological function of TTM2 in pathogendefense is conserved among agriculturally importantcrop plants.
RESULTS
AtTTM2 Is Down-Regulated after Pathogen Infection
Three genes, At1g73980, At1g26190, and At2g11890,are annotated as CYTH domain proteins in theArabidopsis genome and have been named AtTTM1,AtTTM2, and AtTTM3 (Moeder et al., 2013). Two al-lelic homozygous T-DNA insertion KO lines wereobtained for AtTTM2 (Salk_145897 [ttm2-1] andSalk_114669 [ttm2-2]). The T-DNA insertion positionswere found to be located in exon 3 and intron 5 inttm2-1 and ttm2-2, respectively (Supplemental Fig. S2A).Reverse transcription (RT) -PCR analysis showed thatboth lines are, indeed, KO mutants (Supplemental Fig.S2B). A morphological comparison showed no de-tectable difference in the size or shape of both ttm2 KOlines compared with wild-type Columbia-0 (Col-0;Supplemental Fig. S2C).
As mentioned, public microarray data revealed thedown-regulation of AtTTM2 during pathogen infec-tion (Supplemental Fig. S1). To confirm these results,quantitative real-time PCR (qPCR) was conducted onCol-0 wild-type plants that were infected with theoomycete pathogen Hyaloperonospora arabidopsidis (Hpa),isolate Emwa1. We observed a 2-fold reduction inAtTTM2 transcript levels in infected cotyledons com-pared with mock treatment (Fig. 1A), indicating theinvolvement of AtTTM2 in pathogen defense. Inter-estingly, AtTTM2 was also down-regulated in unin-fected systemic tissue of the same seedlings, indicatinga role for AtTTM2 in SAR as well (Fig. 1B).
ttm2 Exhibits Enhanced Resistance against Hpa
Because AtTTM2 is down-regulated after pathogeninfection, we asked whether ttm2 mutants show al-terations in defense-related phenotypes. Cotyledons of7-d-old to 10-d-old seedlings were infected with theHpa isolate, Emwa1, which is avirulent to the Col-0ecotype. It is notable that, although the Emwa1 isolateis considered to have an incompatible interaction withthe Col-0 ecotype, the resistance in this ecotype is not
perfect, and initial layers of mesophyll cells may showthe emergence of some hyphae (Fig. 2A, Cot infected).ttm2 lines, in addition to having fewer or no hyphae,also exhibited a greater manifestation of HR cell deathon infected tissue compared with the wild type, sug-gesting enhanced resistance (Fig. 2A, Cot infected).qPCR analysis also showed approximately 2-fold lessinternal transcribed spacer2 (ITS2) transcript levels, amarker to quantify oomycete infection (Quentin et al.,2009; Fig. 2B; Supplemental Fig. S3A), indicating lessgrowth of pathogens in ttm2 plants. We frequentlyobserved the formation of micro-HR-like cell death inuninfected systemic leaves of wild-type plants afteravirulent infection on cotyledons (Fig. 2A, TL un-infected systemic), similar to the findings of Alvarezet al. (1998). Interestingly, ttm2 plants displayed
Figure 3. ttm2 exhibits enhanced SAR. A, Primary infection of 10-d-oldcotyledons of Col-0 wild-type and ttm2 mutant plants was performedwith the avirulentHpa isolate Emwa1 (SAR+) or water (SAR2). After 7 d,a challenge infection was performed on systemic true leaves with Hpa,Noco2 (virulent). Hyphal structures were visualized 10 d later by TrypanBlue staining. B, Stained leaves were microscopically examined andassigned to different classes. Data shown are from two independentexperiments and taken from 50 plants each; Fisher exact probability testindicates a significant difference between SAR+ ttm2 lines and Col-0(P , 0.0001). The experiment was repeated three times with similarresults. Data from an independent experiment with a similar result areshown in Supplemental Figure S5. Bars = 250 mm.
1012 Plant Physiol. Vol. 166, 2014
Ung et al.
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

significantly enhanced HR cell death on the uninfectedsystemic true leaves (Fig. 2A, TL uninfected systemic).To determine whether this enhanced resistance was
specific to effector-triggered immunity or whetherit also affected PAMP-triggered immunity, infectionwith the virulent Hpa isolate, Emco5, was conducted.Trypan Blue analysis revealed little to no hyphae oninfected tissue of ttm2, whereas in wild-type plants,hyphal structures and oospore formation were clearlyvisible throughout the leaf (Fig. 2C, Cot infected).Consistent with this observation, ITS2 transcript levelsin infected cotyledons of ttm2 seedlings were morethan 2-fold lower compared with the wild type (Fig.2D; Supplemental Fig. S3B). Interestingly, we alsoobserved enhanced HR-like cell death along the veinsof uninfected systemic leaves of ttm2 seedlings (Fig.2C).Supplemental Figure S3C shows that ttm2 plants
also displayed enhanced resistance to the bacterialpathogen P. syringae DC3000 (AvrRps4). These dataindicate that ttm2 plants exhibited enhanced resistanceagainst both avirulent and virulent pathogens.SA has been shown to be a critical signaling mole-
cule in pathogen defense. In line with the resistancephenotype, a significant increase in free SA and itsconjugated form, salicylic acid glucoside (SAG), wasobserved in ttm2 plants upon pathogen infectioncompared with the wild type (Fig. 2, E and F). Takentogether, these data suggest that AtTTM2 is likely in-volved in SA-mediated defense signaling.
ttm2 Is Not a Lesion Mimic Mutant
To date, various autoimmune mutants have beenreported. They show enhanced resistance against var-ious pathogens and often exhibit activation of resis-tance responses, such as accumulation of SA andconstitutive PR gene expression without pathogen in-fection. One well-studied class of autoimmune mu-tants, called lesion mimic mutants, additionally exhibitsspontaneous cell death formation without pathogeninfection (Moeder and Yoshioka, 2008). To testwhether resistance responses are activated withoutpathogen infection in ttm2, Trypan Blue analysis onuninfected ttm2 seedlings was conducted and revealedno spontaneous cell death formation (SupplementalFig. S4A). Additionally, no elevated expression of thedefense marker gene PR1 (Laird et al., 2004) was ob-served in ttm2 seedlings without pathogen infection(Supplemental Fig. S4B). These data suggest that ttm2is not a lesion mimic or conventional autoimmunemutant but likely, a priming mutant that exhibits en-hanced resistance upon pathogen infection.
ttm2 Exhibits Enhanced SAR
The observation that AtTTM2 was also down-regulated in uninfected systemic leaves (Fig. 1B)combined with the enhanced HR cell death in ttm2seedlings (Fig. 2A) prompted us to investigate whetherttm2 is also affected in its SAR response. To assess
Figure 4. Involvement of PAD4,NPR1, and SA in ttm2-mediatedresistance. Infection phenotype ofthe Col-0 wild type, the Ws wildtype, pad4-1, sid2-1, npr1-1, andttm2 mutants, and correspondingdouble mutants 10 d after infectionwith avirulent Hpa, isolate Emwa1.Shown is Trypan Blue staining ofinfected cotyledons (Cots) and un-infected true leaves (TLs). Whitearrows indicate hyphal (Hy) growth,and red arrows indicate HR celldeath. Experiments were repeatedthree times with similar results;10-d-old seedlings were used for in-fection. Oo, Oospore. Bars = 250 mm.
Plant Physiol. Vol. 166, 2014 1013
Role of AtTTM2 in Defense
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
SAR, we first treated cotyledons of wild-type and ttm2plants with either water (SAR2) or the avirulent Hpaisolate, Emwa1 (SAR+). We then performed challengeinoculation using the aggressive virulent Hpa isolate,Noco2, on the upper systemic leaves (Fig. 3A;Supplemental Fig. S5A). We used very strong infectionconditions (i.e. 1 3 105 conidiospores) of the aggressiveisolate, Noco2, to see a clear difference between SAR-induced and noninduced groups. Thus, both wild-typeand ttm2 plants displayed comparable hyphae growthin water-treated plants (Fig. 3A, SAR2; SupplementalFig. S5A, lower). In contrast, Hpa-treated ttm2 plants(Fig. 3A, SAR+; Supplemental Fig. S5A, upper) revealeda stronger reduction in pathogen growth in systemicleaves compared with SAR+ wild-type plants. Stainedleaves were microscopically examined and assigned todifferent classes (Fig. 3B; Supplemental Fig. S5B). Fisherexact probability test indicated a significant differencebetween the ttm2 KO lines and the Col-0 wild type (P,0.0001). These data suggest that ttm2 mutants exhibitenhanced SAR.
The Enhanced Resistance Phenotype of ttm2 RequiresPAD4, ICS1, and NPR1
It has been shown that PAD4, SID2 (ICS1), andNPR1 play key roles in SA-dependent defense re-sponses (Glazebrook et al., 1996; Cao et al., 1997; Jirageet al., 1999; Nawrath and Métraux, 1999; Wildermuthet al., 2001). To investigate whether AtTTM2-mediatedresistance requires these signaling components, we per-formed epistatic analyses using double mutants of ttm2-2and pad4-1, sid2-1, or npr1-1. Col-0 and Wassilewskija(Ws) ecotypes are resistant and susceptible, respectively,to the Hpa isolate, Emwa1 (Fig. 4). As expected, the Col-0wild type exhibited resistance with some hyphae presenton the infected tissue along with punctate areas ofHR cell death in both infected and uninfected systemictissue, whereas the Ws wild type exhibited suscepti-bility with massive hyphal growth and oospore for-mation in infected tissue and no visible signs of HR inthe uninfected systemic leaves (Fig. 4, TL uninfected).pad4-1, sid2-1, and npr1-1 single mutants also exhibi-ted susceptibility with little or no visible HR (Fig. 4;Supplemental Fig. S6) but a great presence of hyphaeand in some cases, oospores (Fig. 4), as expected. Alldouble mutants with ttm2 exhibited similar suscepti-bility as pad4-1, sid2-1, and npr1-1 single mutants (Fig. 4;Supplemental Fig. S6). These data indicate that PAD4,ICS1, and NPR1 are all required for the enhancedresistance phenotype of ttm2.
AtTTM2 Expression Is Negatively Regulated by SA andPAMP Treatment
Because pathogen infection down-regulates thetranscription of AtTTM2 (Fig. 1), the effect of SA onAtTTM2 expression was tested. Col-0 wild-type plantswere sprayed with 100 mM SA and assessed 24 h later
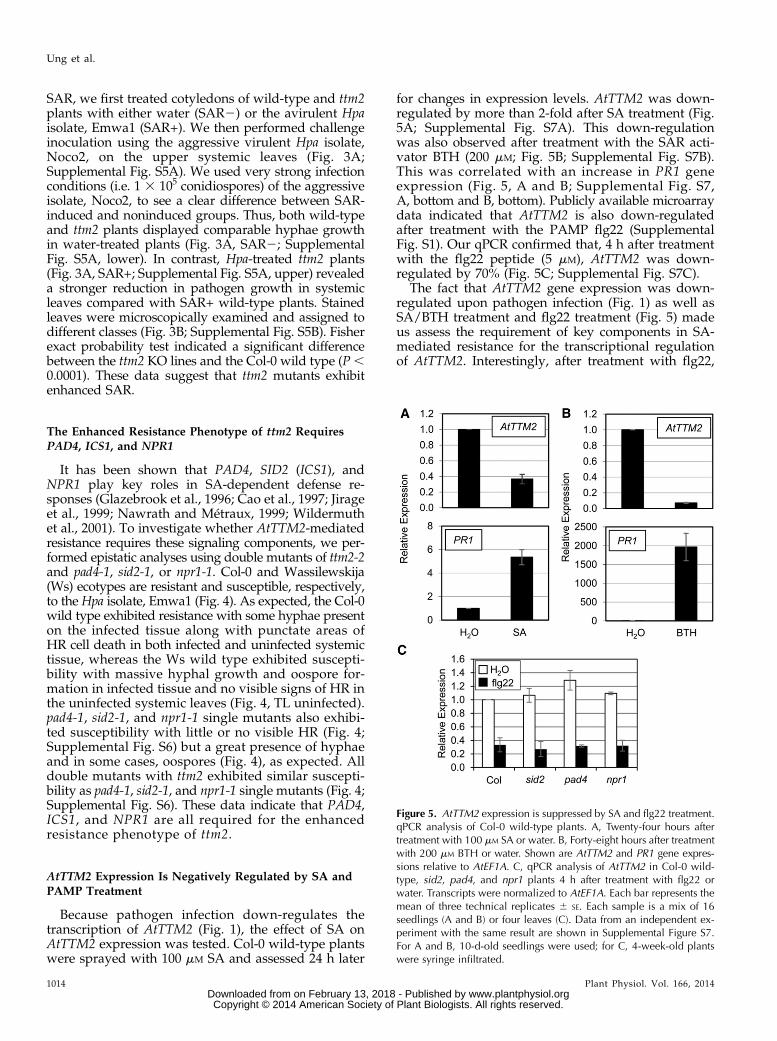
for changes in expression levels. AtTTM2 was down-regulated by more than 2-fold after SA treatment (Fig.5A; Supplemental Fig. S7A). This down-regulationwas also observed after treatment with the SAR acti-vator BTH (200 mM; Fig. 5B; Supplemental Fig. S7B).This was correlated with an increase in PR1 geneexpression (Fig. 5, A and B; Supplemental Fig. S7,A, bottom and B, bottom). Publicly available microarraydata indicated that AtTTM2 is also down-regulatedafter treatment with the PAMP flg22 (SupplementalFig. S1). Our qPCR confirmed that, 4 h after treatmentwith the flg22 peptide (5 mM), AtTTM2 was down-regulated by 70% (Fig. 5C; Supplemental Fig. S7C).
The fact that AtTTM2 gene expression was down-regulated upon pathogen infection (Fig. 1) as well asSA/BTH treatment and flg22 treatment (Fig. 5) madeus assess the requirement of key components in SA-mediated resistance for the transcriptional regulationof AtTTM2. Interestingly, after treatment with flg22,
Figure 5. AtTTM2 expression is suppressed by SA and flg22 treatment.qPCR analysis of Col-0 wild-type plants. A, Twenty-four hours aftertreatment with 100 mM SA or water. B, Forty-eight hours after treatmentwith 200 mM BTH or water. Shown are AtTTM2 and PR1 gene expres-sions relative to AtEF1A. C, qPCR analysis of AtTTM2 in Col-0 wild-type, sid2, pad4, and npr1 plants 4 h after treatment with flg22 orwater. Transcripts were normalized to AtEF1A. Each bar represents themean of three technical replicates 6 SE. Each sample is a mix of 16seedlings (A and B) or four leaves (C). Data from an independent ex-periment with the same result are shown in Supplemental Figure S7.For A and B, 10-d-old seedlings were used; for C, 4-week-old plantswere syringe infiltrated.
1014 Plant Physiol. Vol. 166, 2014
Ung et al.
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

sid2, pad4, and npr1 plants displayed the same level ofAtTTM2 down-regulation as wild-type plants (Fig. 5C;Supplemental Fig. S7C). A similar result was seen afterinfection with P. syringae ES4326 (Supplemental Fig.S8; Wang et al., 2008). Taken together, these data sug-gest that SA, PAD4, and NPR1 are not required for thetranscriptional down-regulation of AtTTM2 but are re-quired for the resistance phenotype of the ttm2mutants.
Overexpression of AtTTM2 Confers EnhancedSusceptibility to Pathogens
The observation that AtTTM2 is down-regulatedupon pathogen infection and SA/flg22 treatmentcombined with the fact that ttm2 plants displayenhanced disease resistance strongly suggest thatAtTTM2 is a negative regulator of disease resistance.Therefore, constitutive expression of AtTTM2 maylead to enhanced disease susceptibility. Thus, we cre-ated AtTTM2 overexpressor lines, where AtTTM2 ex-pression is driven by the strong Cauliflower mosaicvirus35S promoter. To detect differences in diseaseoutcome, we used relatively moderate infection con-ditions with the virulent Hpa isolate, Emco5. Weobserved elevated expression of AtTTM2 in three in-dependent transgenic lines, even after pathogen infec-tion (Fig. 6A; Supplemental Fig. S9). Although only 60%of Col-0 wild-type plants and 30% of ttm2 plantsexhibited heavy hyphal growth 10 d after infection,100% of the plants of the three overexpression linesshowed strong infection (Fig. 6, B and C; SupplementalFig. S9C). Fisher exact probability test indicated a sig-nificant difference between the overexpressor lines andthe Col-0 wild type (P, 0.001). This was also confirmedquantitatively by measuring the expression of theoomycete marker, ITS2 (Fig. 6D; Supplemental Fig. S9).These data strongly suggest that down-regulation ofAtTTM2 is, indeed, required for normal levels of diseaseresistance.
AtTTM2 Function Is Likely Conserved among DifferentPlant Species
Data from Phytozome (www.phytozome.net) indi-cated that TTM2 is highly conserved in a wide variety
Figure 6. Overexpression of AtTTM2 causes enhanced susceptibility.A, qPCR analysis of AtTTM2 in Hpa-infected cotyledons 10 d afterinfection. Transcripts were normalized to AtEF1A. Each bar representsthe mean of three technical replicates 6 SE. Each sample is a mix of 15seedlings. Data from an independent experiment are shown inSupplemental Figure S9. B, Trypan Blue staining of the Col-0 wild type,ttm2, and two independent 35S:AtTTM2 overexpressor lines (35S-2and 35S-5) 13 d after infection with Hpa, Emco5. Bars = 250 mm.
C, Quantitative assessment of infection. Stained leaves were micro-scopically examined and assigned to different classes. Data shownwere taken from 15 to 16 plants; Fisher exact probability test indicatesa significant difference between overexpressor lines and Col-0 (P ,0.001). The experiment was repeated three times with similar results.D, qPCR analysis of ITS2 in Hpa-infected cotyledons 10 d after in-fection. Transcripts were normalized to AtEF1A. Each bar representsthe mean of three technical replicates 6 SE. Each sample is a mix of 15seedlings. Data from an independent experiment are shown inSupplemental Figure S9. The analysis of a third independent line isshown in Supplemental Figure S9, B and C; 10-d-old seedlings wereused for all infections.
Plant Physiol. Vol. 166, 2014 1015
Role of AtTTM2 in Defense
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
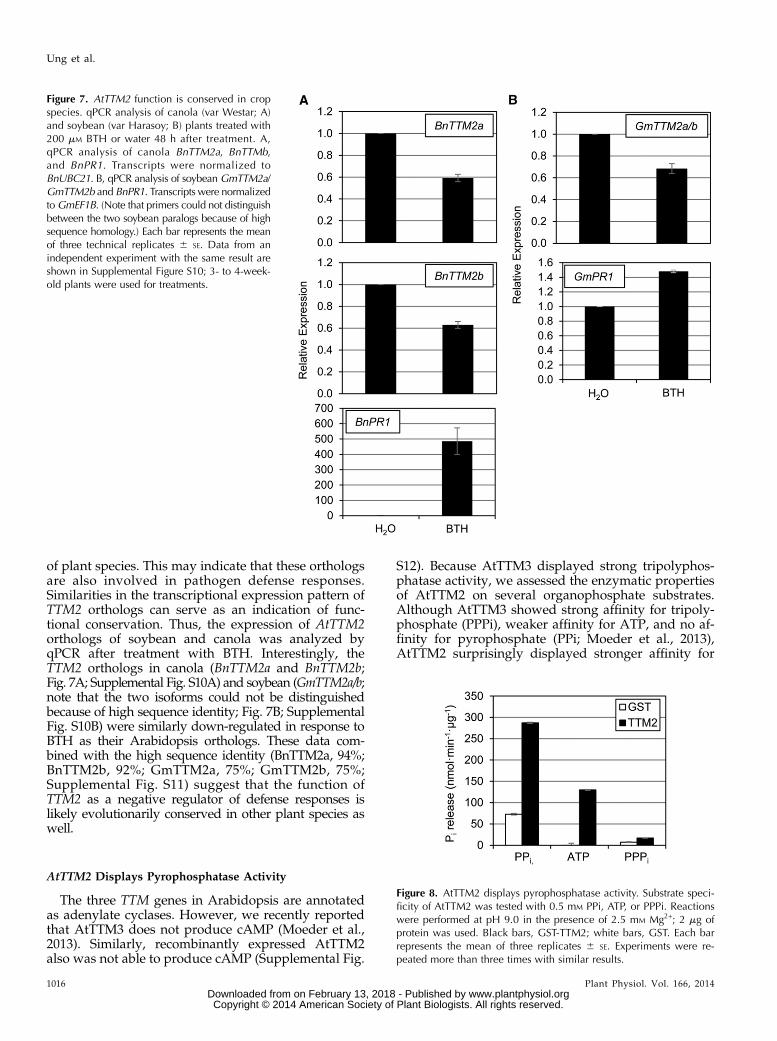
of plant species. This may indicate that these orthologsare also involved in pathogen defense responses.Similarities in the transcriptional expression pattern ofTTM2 orthologs can serve as an indication of func-tional conservation. Thus, the expression of AtTTM2orthologs of soybean and canola was analyzed byqPCR after treatment with BTH. Interestingly, theTTM2 orthologs in canola (BnTTM2a and BnTTM2b;Fig. 7A; Supplemental Fig. S10A) and soybean (GmTTM2a/b;note that the two isoforms could not be distinguishedbecause of high sequence identity; Fig. 7B; SupplementalFig. S10B) were similarly down-regulated in response toBTH as their Arabidopsis orthologs. These data com-bined with the high sequence identity (BnTTM2a, 94%;BnTTM2b, 92%; GmTTM2a, 75%; GmTTM2b, 75%;Supplemental Fig. S11) suggest that the function ofTTM2 as a negative regulator of defense responses islikely evolutionarily conserved in other plant species aswell.
AtTTM2 Displays Pyrophosphatase Activity
The three TTM genes in Arabidopsis are annotatedas adenylate cyclases. However, we recently reportedthat AtTTM3 does not produce cAMP (Moeder et al.,2013). Similarly, recombinantly expressed AtTTM2also was not able to produce cAMP (Supplemental Fig.
S12). Because AtTTM3 displayed strong tripolyphos-phatase activity, we assessed the enzymatic propertiesof AtTTM2 on several organophosphate substrates.Although AtTTM3 showed strong affinity for tripoly-phosphate (PPPi), weaker affinity for ATP, and no af-finity for pyrophosphate (PPi; Moeder et al., 2013),AtTTM2 surprisingly displayed stronger affinity for
Figure 8. AtTTM2 displays pyrophosphatase activity. Substrate speci-ficity of AtTTM2 was tested with 0.5 mM PPi, ATP, or PPPi. Reactionswere performed at pH 9.0 in the presence of 2.5 mM Mg2+; 2 mg ofprotein was used. Black bars, GST-TTM2; white bars, GST. Each barrepresents the mean of three replicates 6 SE. Experiments were re-peated more than three times with similar results.
Figure 7. AtTTM2 function is conserved in cropspecies. qPCR analysis of canola (var Westar; A)and soybean (var Harasoy; B) plants treated with200 mM BTH or water 48 h after treatment. A,qPCR analysis of canola BnTTM2a, BnTTMb,and BnPR1. Transcripts were normalized toBnUBC21. B, qPCR analysis of soybean GmTTM2a/GmTTM2b and BnPR1. Transcripts were normalizedtoGmEF1B. (Note that primers could not distinguishbetween the two soybean paralogs because of highsequence homology.) Each bar represents the meanof three technical replicates 6 SE. Data from anindependent experiment with the same result areshown in Supplemental Figure S10; 3- to 4-week-old plants were used for treatments.
1016 Plant Physiol. Vol. 166, 2014
Ung et al.
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

PPi, weaker activity for ATP, and almost no affinity forPPPi (Fig. 8). AtTTM2 was expressed as a glutathioneS-transferase (GST)-fusion protein. Protein extractedfrom Escherichia coli expressing the GST tag aloneconfirmed that the observed activities are not causedby contaminating bacterial proteins (Fig. 8). These datasuggest divergent biological functions of the AtTTMgenes, which is consistent with the different pheno-types observed in ttm2 and ttm3.
DISCUSSION
To understand the biological function of the TTM,AtTTM2, we have characterized the AtTTM2 KO mu-tants ttm2-1 and ttm2-2. Both lines displayed enhancedresistance against both virulent and avirulent patho-gens, because they exhibited lower growth of bothtypes of pathogens combined with an enhancement ofHR cell death. In addition, SAR was also enhanced inthese mutants. The enhanced resistance was depen-dent on the well-known defense signaling componentsSA, PAD4, and NPR1, which indicates that AtTTM2 isinvolved in the bona fide defense signaling pathwayand likely a negative regulator. Transcriptional sup-pression of AtTTM2 after pathogen infection, PAMPrecognition, or SA/BTH treatment further supportsthis notion. Interestingly, the enhanced pathogen re-sistance is only observed upon pathogen infection—nosignificant autoactivation of defense responses, such asspontaneous cell death formation and elevated levelsof basal SA or PR1 gene expression, were observed.This differentiates AtTTM2 mutants from the majorityof conventional autoimmune mutants (Moeder andYoshioka, 2008; Hofius et al., 2009).A similar phenomenon was reported in the Arabi-
dopsis mutants enhanced disease resistance1 (edr1) andedr2 (Frye and Innes, 1998; Tang et al., 2005a). EDR1 andEDR2 encode a CONSTITUTIVE TRIPLE RESPONSE1family Mitogen-activated protein kinase kinase ki-nase and an unknown protein with a pleckstrinhomology, a steroidogenic acute regulatory protein-related lipid-transfer, and a DOMAIN OF UN-KNOWN FUNCTION1336 (DUF), respectively (Fryeet al., 2001; Tang et al., 2005a, 2005b; Vorwerk et al.,2007). Both mutants were identified in the same screenfor decreased susceptibility against P. syringae DC3000without constitutive PR gene expression and alsoshow enhanced resistance against other pathogens,such as Erysiphe cichoracearum. Interestingly, bothmutants display stronger and faster defense responsesupon pathogen infection; however, no obvious auto-activation of defense was observed, just like for ttm2.These phenotypes were suppressed in mutants withdefects in the SA signal transduction pathway (e.g.sid2, pad4, npr1, and eds1) but not those with defects inthe ethylene/jasmonate pathway, suggesting that theyare hypersensitive to or have a lower threshold in ac-tivating the SA pathway (Frye et al., 2001; Tang et al.,2005a, 2005b; Vorwerk et al., 2007). The precise
molecular mechanisms of these mutants are not yetclear; however, the reported phenotypes are remark-ably similar to those of ttm2. The only outstandingdifference between ttm2 and edr2 is the enhanced SARphenotype in ttm2. As shown, ttm2 displayed strongenhancement of SAR, including HR cell death, in un-infected systemic leaves, but edr2-mediated enhance-ment of resistance does not occur in uninfectedsystemic leaves. This indicates that, although the mu-tant phenotypes are similar, the molecular mechanismbehind the phenomena is fundamentally different.
In terms of SAR, AGD2-LIKE DEFENSE RESPONSEPROTEIN1 (ALD1) was shown to be involved in bothlocal and systemic resistance (Song et al., 2004). ALD1is transcriptionally induced by pathogen infection aswell as BTH treatment in both inoculated and systemictissues. ald1 mutant plants have increased susceptibil-ity to avirulent pathogens and cannot activate SAR.The ALD1 aminotransferase is involved in the bio-synthesis of the SAR regulator pipecolic acid, whichaccumulates in local and systemic tissue of SAR-induced plants (Návarová et al., 2012). Pipecolic acid has
Figure 9. AtTTM2 is a negative regulator of the SA-mediated defenseamplification loop. Recognition of pathogens suppresses the tran-scription of AtTTM2 to amplify defense responses. At a later time point,production of SA leads to continuous transcriptional suppression ofAtTTM2, further amplifying the feedback loop. The KO mutants ofAtTTM2, thus, behave like in a primed state and show enhanced re-sistance on pathogen recognition. The mutant phenotype requires theknown defense signaling components ICS1, PAD4, and NPR1. ETI,Effector-triggered immunity; PTI, pathogen-associated molecular pattern-triggered immunity.
Plant Physiol. Vol. 166, 2014 1017
Role of AtTTM2 in Defense
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

been shown to mediate signal amplification that en-ables systemic SA accumulation, SAR establishment,and defense priming responses in SAR-induced plants.Considering that ttm2 also does not show constitutiveactivation of resistance and displays an SAR pheno-type, AtTTM2 may act by fine tuning the amplificationof defense responses in both inoculated and uninocu-lated leaves. Indeed, an SA-mediated feedback am-plification loop has been suggested for a long time(Shah, 2003). For instance, EDS1 and PAD4, which areimportant defense signaling components, are bothregulators and effectors of SA signaling, stronglysuggesting the existence of an SA-mediated feedbackamplification loop (Dong, 2004). Likewise, ACCELER-ATED CELL DEATH6, which is believed to work up-stream of SA biosynthesis, is transcriptionally inducedby BTH (Lu et al., 2003).
Thus, it can be hypothesized that recognition of path-ogen infection suppresses the expression of AtTTM2,which acts as a negative regulator of the amplificationloop, to facilitate a quick and strong resistance response.At a later time point, SA accumulation induced bypathogen infection further suppresses the expression ofAtTTM2 to boost the positive feedback amplification loopof defense responses. Transcriptional down-regulation ofAtTTM2 can already be seen 4 h after treatment withflg22 and 24 h after infection with P. syringae (Fig. 5C;Supplemental Fig. S8). Interestingly, AtTTM2 down-regulation was also observed in flg22-treated plants aswell as P. syringae-infected sid2, npr1, and pad4 mutantplants (Fig. 5C; Supplemental Figs. S7 and S8), indicatingthat the down-regulation is triggered upstream of PAD4.SA/BTH treatment causes AtTTM2 down-regulationthrough either an additional mechanism or feedbackthrough the SA amplification loop (Fig. 9). In this sce-nario, AtTTM2 plays a role to prevent accidental activa-tion of defense responses through the positive feedbackamplification loop in the absence of pathogens. Thus,ttm2 exhibits a primed mutant phenotype: it can induceresistance responses stronger than wild-type plants, butno constitutive activation of defense responses is ob-served. A model of this concept is presented in Figure 9.Although an SA-mediated feedback amplification loophas been discussed for quite some time (Shah, 2003), onlya few studies have identified components of this feedbackloop (Song et al., 2004; Raffaele et al., 2006; Roberts et al.,2013). Whether AtTTM2 negatively regulates defenseamplification by attenuating pipecolic acid biosynthesisremains to be determined. The molecular mechanism ofAtTTM2 will further our understanding of the SA-medi-ated feedback amplification loop.
All three Arabidopsis TTMs have been annotated asadenylate cyclases based on sequence similarity toCyaB from A. hydrophila (Iyer and Aravind, 2002).However, in this work and previous work, we haveshown that recombinantly expressed AtTTM3 andAtTTM2 do not show adenylate cyclase activity(Moeder et al., 2013; Supplemental Fig. S12). Rather,AtTTM3 exhibits strong tripolyphosphatase activitywith a strong affinity for PPPi. However, AtTTM2
showed the strongest affinity for PPi and only weakactivities for ATP and PPPi. Although the actual invivo substrates are currently unknown, the differencein the in vitro substrate preference between AtTTM3and AtTTM2 indicates distinct biological functions ofthese two TTM family members. Furthermore, in ad-dition to a CYTH domain, both AtTTM1 and AtTTM2,but not AtTTM3, possess a P-loop kinase domain intheir N termini. It is annotated as a uridine/cytidinekinase and has conserved Walker A, Walker B, and lidmodule motifs (Supplemental Fig. S11; Leipe et al.,2003). This indicates the possibility that AtTTM1 andAtTTM2 have dual enzymatic activities (both phos-phatase and kinase). Alternatively, the CYTH domainmay have lost its catalytic function in AtTTM1 andAtTTM2, and its function might be to bind and posi-tion their specific in vivo substrate for the kinase do-main (Iyer and Aravind, 2002). This idea is supportedby the fact that many of the conserved catalytic resi-dues of TTM proteins are altered in AtTTM1 andAtTTM2. The stereotypical EXEXK motif of CYTHproteins (including AtTTM3) is altered to TYILK.Furthermore, the majority of the conserved basic andacidic residues in the b-barrel is not conserved inAtTTM1 and AtTTM2 (Supplemental Fig. S11). Theseresidue changes are conserved among the TTM2orthologs in other plant species, indicating that theycontribute to the unusual catalytic activity of AtTTM2.Unlike all other described TTM proteins, which act ontriphosphate substrates, AtTTM2 prefers a diphos-phate (PPi). In any case, the study of in vivo substratesfor AtTTMs and the characterization of AtTTM1 willprovide additional insights into this group of proteinsin plants and the possible role of this phosphatase/kinase in pathogen defense responses. The analysisof AtTTM1 is currently in progress.
Genomic sequence analyses indicated that all threeTTM family members are conserved among most plantspecies, further indicating the distinct function of allthree TTMs in plants. Interestingly, transcriptionalsuppression of TTM2 by BTH was observed in soy-bean and canola, like in Arabidopsis, strongly indi-cating that the orthologs of TTM2 in these crop plantslikely also work as negative regulators of defense re-sponses. This raises the possibility that KO crop mu-tants for TTM2 will also show enhanced resistancesimilar to Arabidopsis ttm2 plants, providing a usefultool in agricultural biotechnology to generate pathogen-resistant crop plants.
MATERIALS AND METHODS
Plant Growth Conditions and Pathogen Assays
Arabidopsis (Arabidopsis thaliana; accession Col-0), canola (Brassica napusvar Westar), and soybean (Glycine max var Harasoy) plants were grown inSunshine Mix at 22°C, 60% relative humidity, and approximately 140 mE m22 s21
with a 9-h photoperiod; 7- to 10-d-old Arabidopsis plants were infected withHyaloperonospora arabidopsidis. Spore counts of 1 3 105 conidiospores mL21, 8 3 105
cells mL21, and 2 3 105 cells mL21 were used for Noco2, Emco5, and Emwa1isolates, respectively. Seedlings were then infected by drop inoculation and left
1018 Plant Physiol. Vol. 166, 2014
Ung et al.
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

at 16°C and .90% relative humidity for 7 to 10 d before disease assessment.Four-week-old plants were infiltrated with 13 105 colony forming units mL21 ofbacterial pathogen Pseudomonas syringae DC3000 tomato (AvrRps4), and bacterialgrowth was assessed at 0 and 3 d postinfiltration.
Cauliflower Mosaic Virus35S Transgenic Lines
The coding sequence of AtTTM2 was amplified from Arabidopsis Col-0complementary DNA (cDNA) using the primers 35S-TTM2-F and 35S-TTM2-R (Supplemental Table S1) and cloned into pBI121 (Clontech). The vectorwas transformed into Col-0 wild-type plants through Agrobacterium tume-faciens-mediated transformation using the floral dip method (Clough and Bent,1998).
RNA Extraction and RT-PCR
RNA extractionwas carried out using the TRIzol reagent (Life Technologies)according to the manufacturer’s instructions. RT-PCR was performed usingcDNA generated by SuperScript II Reverse Transcriptase (Life Technologies)according to the manufacturer’s instructions. Expression of PR1 was visual-ized by gel electrophoresis of samples after RT-PCR with PR1 primers (AtPR1-Fand AtPR1-R).
qPCR
qPCR was performed using Fast SYBR Green Master Mix (Life Technolo-gies). The expressions of Arabidopsis genes were normalized to the expressionof elongation factor 1a (AtEF1A), whereas the expressions of soybean andcanola genes were normalized to GmEF1B and ubiquitin conjugating enzyme21(BnUBC21), respectively. All primer sequences are listed in SupplementalTable S1.
Confirmation of T-DNA Insertion KO Lines
The SALK lines SALK_145897 (ttm2-1) and SALK_114669 (ttm2-2) wereobtained from the SALK Institute (Alonso et al., 2003). Homozygous plantswere isolated using gene-specific primers for ttm2-1 (897RP and 897LP) andttm2-2 (244RP and 244LP) combined with the T-DNA-specific primer LBb1-F.RT-PCR was then performed on cDNA from both ttm2 lines to confirm the KOstatus using the full-length TTM2 primers (190RT-F and 244RT-R). Expressionwas normalized to the expression of b-tubulin. Primer sequences are listed inSupplemental Table S1.
Epistatic Analysis
ttm2-2 was crossed with pad4-1 (Glazebrook et al., 1996; Jirage et al., 1999),ics1-1 (Wildermuth et al., 2001), and npr1-1 (Cao et al., 1997). Homozygousdouble mutants were isolated in the F2 generation.
SA, BTH, and flg22 Treatments
Seven- to 10-d-old Arabidopsis seedlings were treated with 100 mM SA or200 mM BTH. Treatments of canola and soybean plants were performed withthe same concentrations but on 3- to 4-week-old plants, which were sprayedwith the addition of 0.025% (v/v) Silwet. Treatment with 5 mM flg22 wasperformed on 3- to 4-week-old plants by syringe infiltration.
SAR Experiments
Seedlings were grown for 7 to 10 d and drop inoculated with either wateror 2 3 105 conidiospores mL21 of avirulent Hpa isolate, Emwa1. After trueleaves emerged 7 d later, a secondary infection on upper systemic leaveswith the virulent Hpa isolate, Noco2, was performed using 1 3 105 con-idiospores mL21 on all seedlings. Trypan Blue analysis was then performed 7to 10 d later.
Trypan Blue Staining
Trypan Blue staining was performed as previously described (Yoshiokaet al., 2001).
SA and SAG Measurements
Pooled tissue samples (n = 18) were collected 5 d after infection with theavirulent Hpa isolate, Emwa1, and frozen in liquid nitrogen. Endogenous SAand SAG were extracted and analyzed as previously described (Mosher et al.,2010).
Protein Expression in Escherichia coli
The coding region of AtTTM2 was cloned into pGEX-6P-1 from Arabi-dopsis Col-0 ecotype cDNA using the primers TTM2-TM-F and TTM2-TM-R,which excludes the annotated C-terminal transmembrane domain startingfrom D648. Plasmids were introduced into E. coli BL21 (DE3) and grownovernight in Luria-Bertani medium at 37°C. The overnight culture was used toseed a larger volume of autoinduction medium containing 13 NPS solution[25 mM (NH4)2SO4, 50 mM KH2PO4, and 50 mM Na2HPO4] and 13 5052 so-lution (0.05% [w/v] Glc, 0.2% [w/v] a-lactose, and 0.5% [v/v] glycerol), and itwas grown at 37°C for 3 to 4 h until optical density = 0.4. The temperature wasthen lowered to 18°C overnight before harvesting the cells by centrifugation at4°C (Studier, 2005).
Protein Extraction
E. coli cultures were centrifuged, and pellets were resuspended in 13phosphate-buffered saline (pH 7.5; 137 mM NaCl, 2.7 mM KCl, 10 mM
Na2HPO4, and 1.8 mM KH2PO4) containing 1 mM phenylmethylsulfonyl fluo-ride, 1 mM dithiothreitol, and 10 mg mL21 DNaseI. Cell suspensions wereincubated on ice for 30 min before cell lysis by French press at 1,000 poundsper square inch. Soluble fractions were obtained by centrifugation andsubjected to column purification using DE52 cellulose (Sigma) and GSHagarose (Sigma). Purified protein samples were eluted using 10 mM reducedglutathione.
Enzymatic Assays
Free phosphate released by AtTTM2 was measured with the MalachiteGreen assay (Bernal et al., 2005) as described by Moeder et al. (2013). The assayconditions were 0.5 mM PPi, ATP, or PPPi and 2.5 mM Mg2+, pH 9.0 at 37°C for30 min. cAMP formation was assayed in 25 mM Tris, pH 8, 1 mM ATP, and20 mM Mg2+ at 37°C for 30 min. HPLC analysis was an isocratic run with 20% (v/v)MeOH and 150 mM NaOAc, pH 5, on a Zorbax SB-C18 column (3.5 mm;Agilent).
Statistical Analysis
A two-tailed Student’s t test was performed for all comparisons betweentwo sample groups. Fisher’s exact test was performed for all comparisonsbetween two samples with multiple groups. A P value less than 0.05 was usedto denote significance.
Sequence data from this article can be found in the Arabidopsis GenomeInitiative or GenBank/EMBL databases under the following accession num-bers: AtTTM2 (At1g26190), Hpa-ITS2 (GU583836.1), PR1 (At2g14610), AtEF1A(At5g60390), b-tub (At5g23860), BnTTM2a (Bra011014), BnTTM2b (Bra012464),BnUBC21 (AC172883), BnPR1 (EF423806), GmTTM2a (Gm1g09660), GmTTM2b(Gm2g14110), GmEF1b (NM_001249608.1), and GmPR1 (XM_003545723.1).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Visualization of the expression pattern ofAtTTM2.
Supplemental Figure S2. T-DNA insertion line analysis.
Supplemental Figure S3. ttm2 exhibits enhanced pathogen resistance.
Supplemental Figure S4. ttm2 is not a lesion mimic mutant.
Supplemental Figure S5. ttm2 exhibits enhanced SAR.
Supplemental Figure S6. Epistatic analysis of ttm2.
Plant Physiol. Vol. 166, 2014 1019
Role of AtTTM2 in Defense
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Supplemental Figure S7. AtTTM2 expression is suppressed by SA andflg22 treatment.
Supplemental Figure S8. AtTTM2 down-regulation after Pseudomonassyringae infection does not require SA, NPR1, and PAD4.
Supplemental Figure S9. Overexpression of AtTTM2 causes enhanced sus-ceptibility.
Supplemental Figure S10. AtTTM2 function is conserved in crop species.
Supplemental Figure S11. Sequence alignment of TTM orthologs.
Supplemental Figure S12. AtTTM2 is not an adenylate cyclase.
Supplemental Table S1. Primer sequences.
Received August 15, 2014; accepted August 28, 2014; published September 2,2014.
LITERATURE CITED
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P,Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003)Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science301: 653–657
Alvarez ME, Pennell RI, Meijer PJ, Ishikawa A, Dixon RA, Lamb C (1998)Reactive oxygen intermediates mediate a systemic signal network in theestablishment of plant immunity. Cell 92: 773–784
Bent AF, Mackey D (2007) Elicitors, effectors, and R genes: the new par-adigm and a lifetime supply of questions. Annu Rev Phytopathol 45:399–436
Bernal C, Palacin C, Boronat A, Imperial S (2005) A colorimetric assay for thedetermination of 4-diphosphocytidyl-2-C-methyl-D-erythritol 4-phosphatesynthase activity. Anal Biochem 337: 55–61
Bettendorff L, Wins P (2013) Thiamine triphosphatase and the CYTH su-perfamily of proteins. FEBS J 280: 6443–6455
Cao H, Bowling SA, Gordon AS, Dong X (1994) Characterization of anArabidopsis mutant that is nonresponsive to inducers of systemic ac-quired resistance. Plant Cell 6: 1583–1592
Cao H, Glazebrook J, Clarke JD, Volko S, Dong X (1997) The ArabidopsisNPR1 gene that controls systemic acquired resistance encodes a novelprotein containing ankyrin repeats. Cell 88: 57–63
Clough SJ, Bent AF (1998) Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana.Plant J 16: 735–743
Delvaux D, Murty MRVS, Gabelica V, Lakaye B, Lunin VV, Skarina T,Onopriyenko O, Kohn G, Wins P, De Pauw E, et al (2011) A specificinorganic triphosphatase from Nitrosomonas europaea: structure andcatalytic mechanism. J Biol Chem 286: 34023–34035
Dong X (2004) The role of membrane-bound ankyrin-repeat protein ACD6in programmed cell death and plant defense. Sci STKE 2004: pe6
Falk A, Feys BJ, Frost LN, Jones JD, Daniels MJ, Parker JE (1999) EDS1, anessential component of R gene-mediated disease resistance in Arabi-dopsis has homology to eukaryotic lipases. Proc Natl Acad Sci USA 96:3292–3297
Feys BJ, Moisan LJ, Newman MA, Parker JE (2001) Direct interactionbetween the Arabidopsis disease resistance signaling proteins, EDS1and PAD4. EMBO J 20: 5400–5411
Feys BJ, Wiermer M, Bhat RA, Moisan LJ, Medina-Escobar N, Neu C,Cabral A, Parker JE (2005) Arabidopsis SENESCENCE-ASSOCIATEDGENE101 stabilizes and signals within an ENHANCED DISEASESUSCEPTIBILITY1 complex in plant innate immunity. Plant Cell 17:2601–2613
Frye CA, Innes RW (1998) An Arabidopsis mutant with enhanced resistanceto powdery mildew. Plant Cell 10: 947–956
Frye CA, Tang D, Innes RW (2001) Negative regulation of defense re-sponses in plants by a conserved MAPKK kinase. Proc Natl Acad SciUSA 98: 373–378
Gallagher DT, Smith NN, Kim SK, Heroux A, Robinson H, Reddy PT(2006) Structure of the class IV adenylyl cyclase reveals a novel fold.J Mol Biol 362: 114–122
Glazebrook J, Rogers EE, Ausubel FM (1996) Isolation of Arabidopsismutants with enhanced disease susceptibility by direct screening. Ge-netics 143: 973–982
Gong C, Smith P, Shuman S (2006) Structure-function analysis of Plas-modium RNA triphosphatase and description of a triphosphate tunnelmetalloenzyme superfamily that includes Cet1-like RNA triphospha-tases and CYTH proteins. RNA 12: 1468–1474
Hammond-Kosack KE, Jones JDG (1996) Resistance gene-dependent plantdefense responses. Plant Cell 8: 1773–1791
Heath MC (2000) Hypersensitive response-related death. Plant Mol Biol 44:321–334
Hofius D, Mundy J, Petersen M (2009) Self-consuming innate immunity inArabidopsis. Autophagy 5: 1206–1207
Iyer LM, Aravind L (2002) The catalytic domains of thiamine triphospha-tase and CyaB-like adenylyl cyclase define a novel superfamily of do-mains that bind organic phosphates. BMC Genomics 3: 33
Jirage D, Tootle TL, Reuber TL, Frost LN, Feys BJ, Parker JE, AusubelFM, Glazebrook J (1999) Arabidopsis thaliana PAD4 encodes a lipase-like gene that is important for salicylic acid signaling. Proc Natl Acad SciUSA 96: 13583–13588
Keppetipola N, Jain R, Shuman S (2007) Novel triphosphate phosphohydrolaseactivity of Clostridium thermocellum TTM, a member of the triphosphatetunnel metalloenzyme superfamily. J Biol Chem 282: 11941–11949
Laird J, Armengaud P, Giuntini P, Laval V, Milner JJ (2004) Inappropriateannotation of a key defence marker in Arabidopsis: will the real PR-1please stand up? Planta 219: 1089–1092
Lakaye B, Makarchikov AF, Wins P, Margineanu I, Roland S, Lins L,Aichour R, Lebeau L, El Moualij B, Zorzi W, et al (2004) Human re-combinant thiamine triphosphatase: purification, secondary structureand catalytic properties. Int J Biochem Cell Biol 36: 1348–1364
Lawton KA, Friedrich L, Hunt M, Weymann K, Delaney T, Kessmann H,Staub T, Ryals J (1996) Benzothiadiazole induces disease resistance inArabidopsis by activation of the systemic acquired resistance signaltransduction pathway. Plant J 10: 71–82
Leipe DD, Koonin EV, Aravind L (2003) Evolution and classification ofP-loop kinases and related proteins. J Mol Biol 333: 781–815
Lima CD, Wang LK, Shuman S (1999) Structure and mechanism of yeastRNA triphosphatase: an essential component of the mRNA cappingapparatus. Cell 99: 533–543
Lu H, Rate DN, Song JT, Greenberg JT (2003) ACD6, a novel ankyrinprotein, is a regulator and an effector of salicylic acid signaling in theArabidopsis defense response. Plant Cell 15: 2408–2420
Moeder W, Garcia-Petit C, Ung H, Fucile G, Samuel MA, Christendat D,Yoshioka K (2013) Crystal structure and biochemical analyses revealthat the Arabidopsis triphosphate tunnel metalloenzyme AtTTM3 is atripolyphosphatase involved in root development. Plant J 76: 615–626
Moeder W, Yoshioka K (2008) Lesion mimic mutants: a classical, yet stillfundamental approach to study programmed cell death. Plant SignalBehav 3: 764–767
Mosher S, Moeder W, Nishimura N, Jikumaru Y, Joo SH, Urquhart W,Klessig DF, Kim SK, Nambara E, Yoshioka K (2010) The lesion-mimicmutant cpr22 shows alterations in abscisic acid signaling and abscisicacid insensitivity in a salicylic acid-dependent manner. Plant Physiol152: 1901–1913
Návarová H, Bernsdorff F, Döring AC, Zeier J (2012) Pipecolic acid, anendogenous mediator of defense amplification and priming, is a criticalregulator of inducible plant immunity. Plant Cell 24: 5123–5141
Nawrath C, Métraux JP (1999) Salicylic acid induction-deficient mutants ofArabidopsis express PR-2 and PR-5 and accumulate high levels ofcamalexin after pathogen inoculation. Plant Cell 11: 1393–1404
Parker JE, Holub EB, Frost LN, Falk A, Gunn ND, Daniels MJ (1996)Characterization of eds1, a mutation in Arabidopsis suppressing resis-tance to Peronospora parasitica specified by several different RPP genes.Plant Cell 8: 2033–2046
Quentin M, Allasia V, Pegard A, Allais F, Ducrot PH, Favery B, Levis C,Martinet S, Masur C, Ponchet M, et al (2009) Imbalanced lignin bio-synthesis promotes the sexual reproduction of homothallic oomycetepathogens. PLoS Pathog 5: e1000264
Raffaele S, Rivas S, Roby D (2006) An essential role for salicylic acid inAtMYB30-mediated control of the hypersensitive cell death program inArabidopsis. FEBS Lett 580: 3498–3504
1020 Plant Physiol. Vol. 166, 2014
Ung et al.
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Roberts M, Tang S, Stallmann A, Dangl JL, Bonardi V (2013) Geneticrequirements for signaling from an autoactive plant NB-LRR intracel-lular innate immune receptor. PLoS Genet 9: e1003465
Shah J (2003) The salicylic acid loop in plant defense. Curr Opin Plant Biol6: 365–371
Shah J, Zeier J (2013) Long-distance communication and signal amplifi-cation in systemic acquired resistance. Front Plant Sci 4: 30
Shuman S (2002) What messenger RNA capping tells us about eukaryoticevolution. Nat Rev Mol Cell Biol 3: 619–625
Sismeiro O, Trotot P, Biville F, Vivares C, Danchin A (1998) Aeromonashydrophila adenylyl cyclase 2: a new class of adenylyl cyclases withthermophilic properties and sequence similarities to proteins from hy-perthermophilic archaebacteria. J Bacteriol 180: 3339–3344
Song J, Bettendorff L, Tonelli M, Markley JL (2008) Structural basis for thecatalytic mechanism of mammalian 25-kDa thiamine triphosphatase.J Biol Chem 283: 10939–10948
Song JT, Lu H, McDowell JM, Greenberg JT (2004) A key role for ALD1 inactivation of local and systemic defenses in Arabidopsis. Plant J 40: 200–212
Studier FW (2005) Protein production by auto-induction in high densityshaking cultures. Protein Expr Purif 41: 207–234
Tang D, Ade J, Frye CA, Innes RW (2005a) Regulation of plant defenseresponses in Arabidopsis by EDR2, a PH and START domain-containingprotein. Plant J 44: 245–257
Tang D, Christiansen KM, Innes RW (2005b) Regulation of plant diseaseresistance, stress responses, cell death, and ethylene signaling in Arabidopsisby the EDR1 protein kinase. Plant Physiol 138: 1018–1026
Vlot AC, Klessig DF, Park SW (2008) Systemic acquired resistance: theelusive signal(s). Curr Opin Plant Biol 11: 436–442
Vorwerk S, Schiff C, Santamaria M, Koh S, Nishimura M, Vogel J,Somerville C, Somerville S (2007) EDR2 negatively regulates salicylicacid-based defenses and cell death during powdery mildew infections ofArabidopsis thaliana. BMC Plant Biol 7: 35
Wang L, Mitra RM, Hasselmann KD, Sato M, Lenarz-Wyatt L, Cohen JD,Katagiri F, Glazebrook J (2008) The genetic network controlling theArabidopsis transcriptional response to Pseudomonas syringae pv. maculicola:roles of major regulators and the phytotoxin coronatine. Mol Plant MicrobeInteract 21: 1408–1420
Wildermuth MC, Dewdney J, Wu G, Ausubel FM (2001) Isochorismatesynthase is required to synthesize salicylic acid for plant defence. Nature414: 562–565
Winter D, Vinegar B, Nahal H, Ammar R, Wilson GV, Provart NJ (2007)An “Electronic Fluorescent Pictograph” browser for exploring and an-alyzing large-scale biological data sets. PLoS ONE 2: e718
Yoshioka K, Kachroo P, Tsui F, Sharma SB, Shah J, Klessig DF (2001)Environmentally sensitive, SA-dependent defense responses in the cpr22mutant of Arabidopsis. Plant J 26: 447–459
Plant Physiol. Vol. 166, 2014 1021
Role of AtTTM2 in Defense
www.plantphysiol.orgon February 13, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.


















