
Fetal Alcohol Syndrome and Fetal Alcohol Spectrum Disorder ...

Sylvain Mayeur,1 Jean-Sébastien Wattez,1 Marie-Amélie Lukaszewski,1
Simon Lecoutre,1 Laura Butruille,1 Anne Drougard,2 Delphine Eberlé,1
Bruno Bastide,3 Christine Laborie,1 Laurent Storme,1 Claude Knauf,2 Didier Vieau,1
Christophe Breton,1 and Jean Lesage1
Apelin Controls Fetal and NeonatalGlucose Homeostasis and Is Alteredby Maternal UndernutritionDiabetes 2016;65:554–560 | DOI: 10.2337/db15-0228
The adequate control of glucose homeostasis duringboth gestation and early postnatal life is crucial for thedevelopment of the fetoplacental unit and adaptivephysiological responses at birth. Growing evidencesindicate that apelin and its receptor, APJ, which areexpressed across a wide range of tissues, exert impor-tant roles in glucose homeostasis in adults. However,little is known about the function of the apelinergicsystem during gestation. In this study, we evaluated theactivity of this system in rats, the role of apelin in fetaland neonatal glucose homeostasis, and its modula-tion by maternal food restriction. We found that 1) theapelinergic system was expressed at the fetoplacentalinterface and in numerous fetal tissues, 2) ex vivo, theplacenta released high amounts of apelin in late gesta-tion, 3) intravenous apelin injection in mothers increasedthe transplacental transport of glucose, and 4) intraper-itoneal apelin administration in neonates increasedglucose uptake in lung and muscle. Maternal foodrestriction drastically reduced apelinemia in both mothersand growth-restricted fetuses and altered the expres-sion of the apelinergic system at the fetoplacental in-terface. Together, our data demonstrate that apelincontrols fetal and neonatal glucose homeostasis andis altered by fetal growth restriction induced by maternalundernutrition.
Apelin is a regulatory peptide, identified as an endogenousligand of the apelin receptor named APJ (1). The apelingene encodes a 77–amino-acid preproprotein that generates
several molecular isoforms such as apelin-36, apelin-17,and apelin-13 by posttranslational processing (2,3).Moreover, the N-terminal glutamate of apelin-13 can beposttranslationally modified, thus creating the pyroglutamateapelin-13 ([pyr-1]-apelin-13), which is more protectedfrom exopeptidase degradation (4). Apelin and APJ havea widespread distribution in the body and are involved invarious physiological functions. Apelin and APJ mRNAsare expressed in heart, lung, placenta, mammary gland,several regions of the central nervous system, adipocytes,and the gastrointestinal tract (5,6). Depending on thecell type studied, APJ activation results in the activation ofseveral intracellular effectors such as extracellular signal–regulated kinases, protein kinase B (PKB or Akt), andp70S6 kinase and in the inhibition of cAMP production(7,8).
Recently, apelin has been extensively described as abeneficial factor regarding glucose metabolism and asendowed with antidiabetes properties (9,10). Studiesdemonstrated that both short- and long-term apelintreatments improve insulin sensitivity in obese and in-sulin-resistant mice mainly by increasing glucose uptakein skeletal muscle (9–11). Apelin also modulates insulinsecretion and increases pancreatic islet cell mass andb-cell insulin content in mice (12). During prenatal devel-opment, previous studies have revealed that APJ defi-ciency in mice causes early embryonic defects and leadsto embryonic lethality due to growth retardation and car-diac malformations (13). However, the effect of apelin onglucose homeostasis in utero remains unknown. To gain
1Unité Environnement Périnatal et Santé, Equipe d’accueil 4489, Équipe MalnutritionMaternelle et Programmation des Maladies Métaboliques, Université de Lille,Villeneuve d’Ascq, France2INSERM U1048, Équipe no. 3, Institut de Médecine Moléculaire de Rangueil,Université Paul Sabatier, Toulouse, France3Activité Physique, Muscle et Santé, Equipe d’accueil 4488, Université de Lille,Villeneuve d’Ascq, France
Corresponding author: Jean Lesage, [email protected].
Received 18 February 2015 and accepted 24 November 2015.
This article contains Supplementary Data online at http://diabetes.diabetesjournals.org/lookup/suppl/doi:10.2337/db15-0228/-/DC1.
© 2016 by the American Diabetes Association. Readers may use this article aslong as the work is properly cited, the use is educational and not for profit, andthe work is not altered.
554 Diabetes Volume 65, March 2016
METABOLISM

further insight into apelin function during gestation and tostudy the effects of apelin on fetal and neonatal glucosehomeostasis, we formulated the aims of the current study:to investigate in rats 1) the kinetics of apelin plasma levelsin mother/fetus pairs, 2) the ex vivo placental apelin re-lease, 3) the gene expression levels of apelin and APJ in thefetoplacental unit, 4) the effect of maternal apelin admin-istration on transplacental glucose transfer, and 5) the ef-fect of apelin administration to neonates on glucose uptakein several tissues. We subsequently studied the apelinergic(apelin and APJ) system in intrauterine growth–restricted(IUGR) fetuses from rat mothers that received only 30% ofthe food intake of control mothers (FR30 model).
RESEARCH DESIGN AND METHODS
AnimalsExperiments were conducted in accordance with theinstitutional guidelines for the use of laboratory animalsand were approved by the animal ethics committee (fromUniversité de Lille). Adult Wistar rats (Janvier) werehoused at 22°C with a 12-h light/dark cycle with free accessto a chow diet (16% protein, 3% fat, and 60% carbohy-drates). Females were mated with a male. Embryonic day0 (E0) was defined the following day if spermatozoa werefound in vaginal smears. Pregnant females were dividedinto two groups: 1) a control group (n = 50) in whichdams were fed ad libitum and 2) a food-restricted group(n = 27) in which females received 30% of the foodintake of control mothers from E1 to E21. Five virgin adultfemales were used for apelinemia determination.
Plasma and Tissue CollectionsTail blood samples were collected at 9:00 A.M. in virgin andE7 females to measure apelinemia. At E13, E17, and E21,pregnant rats were killed by decapitation. Placentas,fetuses, and mesometrial triangles (Meso-tr) were collectedby caesarean section. Maternal and fetal plasma aliquotswere stored at 220°C. Placentas and Meso-tr were frozenin liquid N2 and stored at280°C. Some E21 fetuses (n = 5)were used to study the tissue distribution of apelin andAPJ mRNAs using reverse transcriptase quantitative real-time PCR (RT-qPCR) analysis. In fetuses, 17 selected tis-sues were dissected, frozen, and stored at 280°C.
Endocrine and Circulating ParametersCommercially available ELISA/enzyme immunosorbent as-say kits were used to measure plasma insulin (catalog no.EIA-2943; DRG International) and apelin (catalog no.EK-057-23, which assayed all isoforms of apelin from apelin-12to apelin-36; Phoenix Pharmaceuticals) levels. Blood glucosewas measured using a glucometer (Roche Diagnostics).
RT-qPCR AnalysisMethods for RT-qPCR analysis have been previouslydescribed (14). RT-qPCR was performed with LightCycler480 SYBR Green I Master and a LightCycler 480 instru-ment (Roche). Primers for apelin, APJ, GLUT1, and GLUT3genes reported in Supplementary Table 1 were designed
using the Primer Premier software (Premier Biosoft In-ternational). Several housekeeping genes (Gapdh, Ppib,and Hprt1) were used for the normalization and weredescribed previously (14).
Placental Apelin SecretionAfter cesarean section, E17 and E21 placentas (n = 24)were collected, rinsed in saline, and incubated for 24 hin dish plates containing 2 mL of DMEM (Gibco). Dishplates were placed at 37°C with 95% O2 and 5% CO2 and95% humidity. Samples of the medium were collectedafter 2, 6, and 24 h of incubation for determination ofapelin concentrations.
Placental Transport of 2-deoxy-D-[3H]glucoseE17 pregnant females (n = 10) were anesthetized usingisoflurane and 10 min later were implanted with a cath-eter in the jugular vein. An intravenous injection of 50 mCi2-deoxy-D-[3H]glucose ([3H]-2DG) (NEN LifeScience) witheither saline or 600 pmol/kg [pyr-1]-apelin-13 (Bachem[U.K.] Limited) was performed. After 30 min, placentasand fetuses were rapidly collected, weighed, and frozen.Total [3H]-2DG content in placental and fetal homogenateswas measured.
Effect of Apelin-13 on Glycemia of NeonatesE21 pregnant females (n = 9) were killed by decapitation.and fetuses were collected by cesarean section. Neonateswere immediately weighed and injected with either0.1 mL i.p. saline or different doses of [pyr-1]-apelin-13(from 1 to 40 nmol/kg i.p.). After 30 min, blood glucosewas measured. Plasma samples from neonates injectedwith saline and with 10, 15, 20, and 40 nmol/kg apelinwere also used for insulinemia determination.
In Vivo [3H]-2DG Incorporation in Neonatal TissuesAn injection of 50 mCi i.p. [3H]-2DG in saline or associatedwith [pyr-1]-apelin-13 (15 nmol/kg i.p.) was performed inrat neonates at birth (n = 14). After 30 min, rats weredecapitated and seven selected tissues were collected for[3H]-2DG uptake determination.
Statistical AnalysisResults are reported as mean 6 SEM. Statistical analyseswere performed using one-way ANOVA and the Dunnetttest. A P value of ,0.05 was considered significant.
RESULTS
Plasma and Expression Levels of Apelin and APJin the Rat Fetoplacental UnitMaternal plasma apelin concentrations were decreasedat E7, increased from E7 to E17, and reduced at term(Fig. 1A). Fetal plasma apelin concentrations were two-fold higher than maternal levels at E17, whereas theywere similar at E21 (Fig. 1B). Apelin mRNA levels werehigher in the Meso-tr compared with placenta at E13and thereafter reduced until term of gestation in bothtissues (Fig. 1C). APJ mRNA levels were similar in thesetwo tissues at E13 (Fig. 1D). In the placenta, APJ expres-sion was increased at E17 versus the E13 level and
diabetes.diabetesjournals.org Mayeur and Associates 555
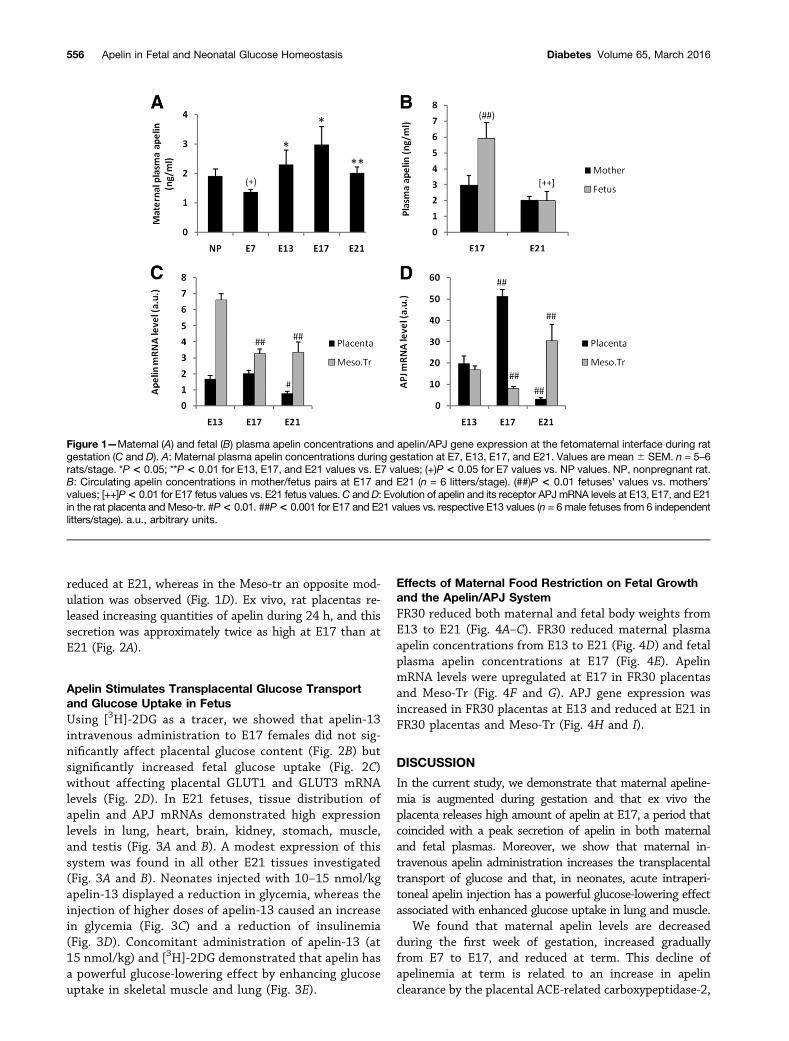
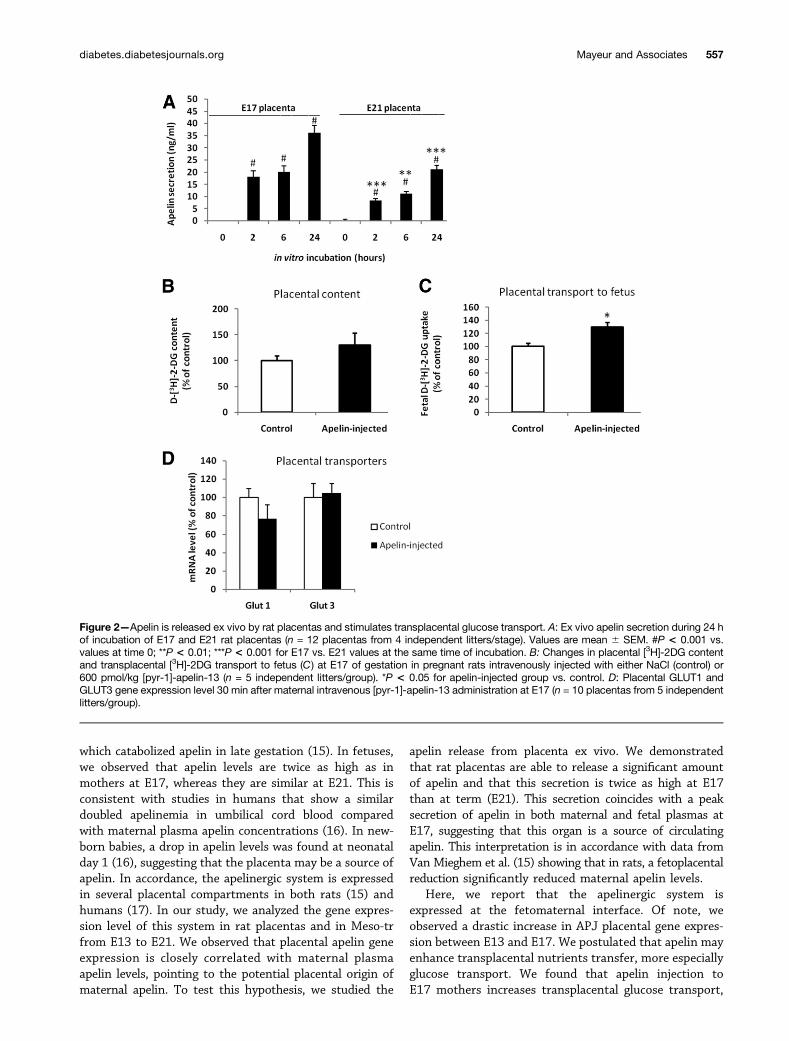
reduced at E21, whereas in the Meso-tr an opposite mod-ulation was observed (Fig. 1D). Ex vivo, rat placentas re-leased increasing quantities of apelin during 24 h, and thissecretion was approximately twice as high at E17 than atE21 (Fig. 2A).
Apelin Stimulates Transplacental Glucose Transportand Glucose Uptake in FetusUsing [3H]-2DG as a tracer, we showed that apelin-13intravenous administration to E17 females did not sig-nificantly affect placental glucose content (Fig. 2B) butsignificantly increased fetal glucose uptake (Fig. 2C)without affecting placental GLUT1 and GLUT3 mRNAlevels (Fig. 2D). In E21 fetuses, tissue distribution ofapelin and APJ mRNAs demonstrated high expressionlevels in lung, heart, brain, kidney, stomach, muscle,and testis (Fig. 3A and B). A modest expression of thissystem was found in all other E21 tissues investigated(Fig. 3A and B). Neonates injected with 10–15 nmol/kgapelin-13 displayed a reduction in glycemia, whereas theinjection of higher doses of apelin-13 caused an increasein glycemia (Fig. 3C) and a reduction of insulinemia(Fig. 3D). Concomitant administration of apelin-13 (at15 nmol/kg) and [3H]-2DG demonstrated that apelin hasa powerful glucose-lowering effect by enhancing glucoseuptake in skeletal muscle and lung (Fig. 3E).
Effects of Maternal Food Restriction on Fetal Growthand the Apelin/APJ SystemFR30 reduced both maternal and fetal body weights fromE13 to E21 (Fig. 4A–C). FR30 reduced maternal plasmaapelin concentrations from E13 to E21 (Fig. 4D) and fetalplasma apelin concentrations at E17 (Fig. 4E). ApelinmRNA levels were upregulated at E17 in FR30 placentasand Meso-Tr (Fig. 4F and G). APJ gene expression wasincreased in FR30 placentas at E13 and reduced at E21 inFR30 placentas and Meso-Tr (Fig. 4H and I).
DISCUSSION
In the current study, we demonstrate that maternal apeline-mia is augmented during gestation and that ex vivo theplacenta releases high amount of apelin at E17, a period thatcoincided with a peak secretion of apelin in both maternaland fetal plasmas. Moreover, we show that maternal in-travenous apelin administration increases the transplacentaltransport of glucose and that, in neonates, acute intraperi-toneal apelin injection has a powerful glucose-lowering effectassociated with enhanced glucose uptake in lung and muscle.
We found that maternal apelin levels are decreasedduring the first week of gestation, increased graduallyfrom E7 to E17, and reduced at term. This decline ofapelinemia at term is related to an increase in apelinclearance by the placental ACE-related carboxypeptidase-2,
Figure 1—Maternal (A) and fetal (B) plasma apelin concentrations and apelin/APJ gene expression at the fetomaternal interface during ratgestation (C and D). A: Maternal plasma apelin concentrations during gestation at E7, E13, E17, and E21. Values are mean 6 SEM. n = 5–6rats/stage. *P < 0.05; **P < 0.01 for E13, E17, and E21 values vs. E7 values; (+)P < 0.05 for E7 values vs. NP values. NP, nonpregnant rat.B: Circulating apelin concentrations in mother/fetus pairs at E17 and E21 (n = 6 litters/stage). (##)P < 0.01 fetuses’ values vs. mothers’values; [++]P< 0.01 for E17 fetus values vs. E21 fetus values.C andD: Evolution of apelin and its receptor APJmRNA levels at E13, E17, and E21in the rat placenta and Meso-tr. #P< 0.01. ##P< 0.001 for E17 and E21 values vs. respective E13 values (n = 6 male fetuses from 6 independentlitters/stage). a.u., arbitrary units.
556 Apelin in Fetal and Neonatal Glucose Homeostasis Diabetes Volume 65, March 2016
which catabolized apelin in late gestation (15). In fetuses,we observed that apelin levels are twice as high as inmothers at E17, whereas they are similar at E21. This isconsistent with studies in humans that show a similardoubled apelinemia in umbilical cord blood comparedwith maternal plasma apelin concentrations (16). In new-born babies, a drop in apelin levels was found at neonatalday 1 (16), suggesting that the placenta may be a source ofapelin. In accordance, the apelinergic system is expressedin several placental compartments in both rats (15) andhumans (17). In our study, we analyzed the gene expres-sion level of this system in rat placentas and in Meso-trfrom E13 to E21. We observed that placental apelin geneexpression is closely correlated with maternal plasmaapelin levels, pointing to the potential placental origin ofmaternal apelin. To test this hypothesis, we studied the
apelin release from placenta ex vivo. We demonstratedthat rat placentas are able to release a significant amountof apelin and that this secretion is twice as high at E17than at term (E21). This secretion coincides with a peaksecretion of apelin in both maternal and fetal plasmas atE17, suggesting that this organ is a source of circulatingapelin. This interpretation is in accordance with data fromVan Mieghem et al. (15) showing that in rats, a fetoplacentalreduction significantly reduced maternal apelin levels.
Here, we report that the apelinergic system isexpressed at the fetomaternal interface. Of note, weobserved a drastic increase in APJ placental gene expres-sion between E13 and E17. We postulated that apelin mayenhance transplacental nutrients transfer, more especiallyglucose transport. We found that apelin injection toE17 mothers increases transplacental glucose transport,
Figure 2—Apelin is released ex vivo by rat placentas and stimulates transplacental glucose transport. A: Ex vivo apelin secretion during 24 hof incubation of E17 and E21 rat placentas (n = 12 placentas from 4 independent litters/stage). Values are mean 6 SEM. #P < 0.001 vs.values at time 0; **P < 0.01; ***P < 0.001 for E17 vs. E21 values at the same time of incubation. B: Changes in placental [3H]-2DG contentand transplacental [3H]-2DG transport to fetus (C) at E17 of gestation in pregnant rats intravenously injected with either NaCl (control) or600 pmol/kg [pyr-1]-apelin-13 (n = 5 independent litters/group). *P < 0.05 for apelin-injected group vs. control. D: Placental GLUT1 andGLUT3 gene expression level 30 min after maternal intravenous [pyr-1]-apelin-13 administration at E17 (n = 10 placentas from 5 independentlitters/group).
diabetes.diabetesjournals.org Mayeur and Associates 557

Figure 3—Gene expression levels of apelin and APJ in E21 rat tissues (A and B) and effect of apelin on glucose homeostasis in ratneonates. n = 6 male fetuses. C and D: Effects of intraperitoneal administration of different doses of [pyr-1]-apelin-13 to neonates onglycemia and plasma insulin concentrations (n = 5–7 rats/treatment group). Values are mean 6 SEM. *P < 0.05; ***P < 0.001 for apelin-injected group vs. NaCl-injected group. E: Effect of a dose of 15 nmol/kg i.p. [pyr-1]-apelin-13 on [3H]-2DG content in rat neonatal tissues.*P < 0.05 for apelin-injected group vs. control (n = 7 neonates/group). a.u., arbitrary units; BAT, brown adipose tissue; Umb., umbilical.
558 Apelin in Fetal and Neonatal Glucose Homeostasis Diabetes Volume 65, March 2016

suggesting that maternal apelin levels control fetal glu-cose supply. This was observed without a change in placen-tal glucose transporters expressions. In rat placentas,apelin was detected in perivascular smooth muscle of thelabyrinth, suggesting that apelin may exert vasoactiveaction in the placenta (15), as it has been extensivelyreported for apelin in the periphery (18–20). We proposethat apelin may induce a vasodilatation of placental ves-sels, which would enhance transplacental glucose transfer,
or indirectly may modulate maternal blood pressure, result-ing in this enhanced placental transport.
We observed high apelin levels in fetuses, whichsuggests that apelin may be important for fetal develop-ment. In E21 fetuses, the apelin and APJ tissue distribu-tion was closely comparable with that in adults (5,6,21),with higher expressions found in lung, heart, muscle,brain, kidney, stomach, and testis. Injection of apelininto neonates was able to reduce glycemia at a low dose
Figure 4—Effect of maternal 70% food restriction (FR30) during gestation on maternal and fetal body weight (A–C) and apelinemia (D andE ) and on the expression of the apelinergic system at the fetomaternal interface (F–I). A–C: n = 10 litters/group. Values are mean 6 SEM.*P < 0.05; **P < 0.01; ***P < 0.001 for FR30 vs. control (C) group. C–F: Quantification of mRNA levels of the apelin/APJ system in placentaand Meso-tr. *P < 0.05; **P < 0.01 for FR30 vs. control group (n = 6 male fetuses from 6 independent litters/developmental stage).
diabetes.diabetesjournals.org Mayeur and Associates 559

and, inversely, to increase glycemia at a high dose byinhibiting insulin release. These results are in accordancewith the hypoglycemic effect of physiologic low doses ofapelin in mice (11) and with the inhibitory role of highpharmacologic doses of apelin on insulin release (22). Wedemonstrate that the hypoglycemic effect of apelin is as-sociated with an increase in lung and muscle glucose up-takes. Similarly, in mice, apelin was shown to exert apowerful glucose-lowering effect associated with enhancedglucose utilization in skeletal muscle (11).
Finally, we studied the apelinergic system in IUGRfetuses from FR30 mothers. FR30 drastically reducesmaternal apelin levels from E13 to E21 as well as fetalapelin levels at E17. In accordance, plasma apelinconcentrations were found to be reduced by food re-striction in adult rodents and humans (9,21). In ourFR30 model, we found that at the fetomaternal interface,apelin gene expression is upregulated at E17. This phe-nomenon may be a compensatory response of IUGRfetuses to increase their glucose supply to improve theirgrowth. Altogether, our data demonstrate that apelin is anew hormone implicated in fetal and neonatal glucosehomeostasis.
Acknowledgments. The authors thank Anne Dickes-Coopman, ValérieMontel, and Yseult Lesage for technical help.Funding. This work was supported by grants from the French Ministry ofEducation and the French National Research Agency (project no. ANR-06-PNRA-022)and by a grant from the Appert Institute.Duality of Interest. No potential conflicts of interest relevant to this articlewere reported.Author Contributions. S.M., J.-S.W., M.-A.L., S.L., L.B., A.D., B.B., C.L.,C.K., and J.L. designed the experiments, researched data, and wrote themanuscript. D.E., L.S., D.V., and C.B. interpreted the results and drafted themanuscript. J.L. is the guarantor of this work and, as such, had full access to allthe data in the study and takes responsibility for the integrity of the data and theaccuracy of the data analysis.
References1. Tatemoto K, Hosoya M, Habata Y, et al. Isolation and characterization of anovel endogenous peptide ligand for the human APJ receptor. Biochem BiophysRes Commun 1998;251:471–4762. Lee DK, Cheng R, Nguyen T, et al. Characterization of apelin, the ligand forthe APJ receptor. J Neurochem 2000;74:34–413. Carpéné C, Dray C, Attané C, et al. Expanding role for the apelin/APJ systemin physiopathology. J Physiol Biochem 2007;63:359–373
4. Habata Y, Fujii R, Hosoya M, et al. Apelin, the natural ligand of the orphanreceptor APJ, is abundantly secreted in the colostrum. Biochim Biophys Acta1999;1452:25–355. Hosoya M, Kawamata Y, Fukusumi S, et al. Molecular and functionalcharacteristics of APJ. Tissue distribution of mRNA and interaction with theendogenous ligand apelin. J Biol Chem 2000;275:21061–210676. Kawamata Y, Habata Y, Fukusumi S, et al. Molecular properties of apelin:tissue distribution and receptor binding. Biochim Biophys Acta 2001;1538:162–1717. Masri B, Knibiehler B, Audigier Y. Apelin signalling: a promising pathwayfrom cloning to pharmacology. Cell Signal 2005;17:415–4268. D’Aniello C, Lonardo E, Iaconis S, et al. G protein-coupled receptor APJ andits ligand apelin act downstream of Cripto to specify embryonic stem cells towardthe cardiac lineage through extracellular signal-regulated kinase/p70S6 kinasesignaling pathway. Circ Res 2009;105:231–2389. Castan-Laurell I, Dray C, Knauf C, Kunduzova O, Valet P. Apelin, a promisingtarget for type 2 diabetes treatment? Trends Endocrinol Metab 2012;23:234–24110. Chaves-Almagro C, Castan-Laurell I, Dray C, Knauf C, Valet P, Masri B. Apelinreceptors: from signaling to antidiabetic strategy. Eur J Pharmacol 2015;763:149–15911. Dray C, Knauf C, Daviaud D, et al. Apelin stimulates glucose utilization innormal and obese insulin-resistant mice. Cell Metab 2008;8:437–44512. Chen H, Zheng C, Zhang X, et al. Apelin alleviates diabetes-associated endo-plasmic reticulum stress in the pancreas of Akita mice. Peptides 2011;32:1634–163913. Charo DN, Ho M, Fajardo G, et al. Endogenous regulation of cardiovascularfunction by apelin-APJ. Am J Physiol Heart Circ Physiol 2009;297:H1904–H191314. Mayeur S, Lancel S, Theys N, et al. Maternal calorie restriction modulatesplacental mitochondrial biogenesis and bioenergetic efficiency: putative in-volvement in fetoplacental growth defects in rats. Am J Physiol Endocrinol Metab2013;304:E14–E2215. Van Mieghem T, van Bree R, Van Herck E, Pijnenborg R, Deprest J, VerhaegheJ. Maternal apelin physiology during rat pregnancy: the role of the placenta. Pla-centa 2010;31:725–73016. Malamitsi-Puchner A, Gourgiotis D, Boutsikou M, Baka S, Hassiakos D,Briana DD. Circulating apelin concentrations in mother/infant pairs at term. ActaPaediatr 2007;96:1751–175417. Cobellis L, De Falco M, Mastrogiacomo A, et al. Modulation of apelin andAPJ receptor in normal and preeclampsia-complicated placentas. Histol Histo-pathol 2007;22:1–818. Ashley EA, Powers J, Chen M, et al. The endogenous peptide apelin potentlyimproves cardiac contractility and reduces cardiac loading in vivo. CardiovascRes 2005;65:73–8219. Japp AG, Cruden NL, Amer DAB, et al. Vascular effects of apelin in vivo inman. J Am Coll Cardiol 2008;52:908–91320. Tatemoto K, Takayama K, Zou MX, et al. The novel peptide apelin lowers bloodpressure via a nitric oxide-dependent mechanism. Regul Pept 2001;99:87–9221. Butruille L, Drougard A, Knauf C, et al. The apelinergic system: sexual di-morphism and tissue-specific modulations by obesity and insulin resistance infemale mice. Peptides 2013;46:94–10122. Ringström C, Nitert MD, Bennet H, et al. Apelin is a novel islet peptide.Regul Pept 2010;162:44–51
560 Apelin in Fetal and Neonatal Glucose Homeostasis Diabetes Volume 65, March 2016


















