
AP-2 transcription factor family member expression, activity, and regulation in human epidermal...
13
ORIGINAL ARTICLE Claudia Popa . Alison L. Dahler . Magdalena M. Serewko-Auret . Chung F. Wong . Louise Smith . Liam M. Barnes . Geoff M. Strutton . Nicholas A. Saunders AP-2 transcription factor family member expression, activity, and regulation in human epidermal keratinocytes in vitro Received February 26, 2004; accepted in revised form April 6, 2004 Abstract The AP-2 transcription factor family is pre- sumed to play an important role in the regulation of the keratinocyte squamous differentiation program; how- ever, limited functional data are available to support this. In the present study, the activity and regulation of AP-2 were examined in differentiating human epidermal keratinocytes. We report that (1) AP-2 transcriptional activity decreases in differentiated keratinocytes but re- mains unchanged in differentiation-insensitive squa- mous cell carcinoma cell lines, (2) diminished AP-2 transcriptional activity is associated with a loss of spe- cific DNA-bound AP-2 complexes, and (3) there is an increase in the ability of cytoplasmic extracts, derived from differentiated keratinocytes, to phosphorylate AP-2a and AP-2b when cells differentiate. In contrast, extracts from differentiation-insensitive squamous cell carcinoma cells are unable to phosphorylate AP-2 pro- teins. Finally, the phosphorylation of recombinant AP-2a by cytosolic extracts from differentiated kera- tinocytes is associated with decreased AP-2 DNA-bind- ing activity. Combined, these data indicate that AP-2 trans-activation and DNA-binding activity decrease as keratinocytes differentiate, and that this decreased ac- tivity is associated with an enhanced ability to phos- phorylate AP-2a and b. Key words AP-2 human epidermal keratinocytes transcription factor Introduction The epithelial cells of the epidermis (keratinocytes) maintain the integrity of the epidermal barrier by fol- lowing a tightly regulated, multistep program of squamous differentiation (Fuchs, 1990, 1994, 1998; By- rne et al., 1994; Fuchs and Byrne, 1994; Byrne, 1997; Nemes and Steinert, 1999). During squamous differen- tiation, proliferative, undifferentiated keratinocytes, located within the basal layer, are converted to non- proliferative, differentiated keratinocytes and ultimately to anuclear corneocytes (Fuchs, 1990, 1998; Byrne et al., 1994; Fuchs and Byrne, 1994; Byrne, 1997; Nemes and Steinert, 1999). As cells migrate through the differenti- ating suprabasal layers, their morphological and bio- chemical characteristics alter in a temporally and spatially specific fashion. Initially, squamous differen- tiation requires repression of proliferation-associated genes such as E2F1, cdc2, and p53 or keratins K5 and K14, followed by induction of squamous-specific genes such as transglutaminase type I (TGaseI), involucrin, loricrin, and filaggrin, and the keratins K1 and K10 (Fuchs and Green, 1980; Rothnagel and Rogers, 1984; Claudia Popa Alison L. Dahler Magdalena M. Serewko- Auret Chung F. Wong Louise Smith Liam M. Barnes Nicholas A. Saunders ( . *) Epithelial Pathobiology Group Cancer Biology Program Centre for Immunology and Cancer Research University of Queensland Building 1, R Wing, Level 4 Princess Alexandra Hospital Brisbane, Queensland, Australia, 4102 Tel: 617 3240 5936 Fax: 617 3240 5946 E-mail: [email protected] Geoff M. Strutton Department of Pathology Princess Alexandra Hospital Brisbane, Queensland, Australia, 4102 Nicholas A. Saunders Department of Physiology and Pharmacology University of Queensland St. Lucia, Queensland, Australia, 4072 U.S. Copyright Clearance Center Code Statement: 0301–4681/2004/7205–185 $ 15.00/0 Differentiation (2004) 72:185–197 r International Society of Differentiation 2004
-
Upload
claudia-popa -
Category
Documents
-
view
212 -
download
0
Transcript of AP-2 transcription factor family member expression, activity, and regulation in human epidermal...

ORIGINAL ARTICLE
Claudia Popa . Alison L. Dahler . Magdalena M.
Serewko-Auret . Chung F. Wong . Louise Smith .
Liam M. Barnes . Geoff M. Strutton .
Nicholas A. Saunders
AP-2 transcription factor family member expression, activity,and regulation in human epidermal keratinocytes in vitro
Received February 26, 2004; accepted in revised form April 6, 2004
Abstract The AP-2 transcription factor family is pre-sumed to play an important role in the regulation of thekeratinocyte squamous differentiation program; how-ever, limited functional data are available to supportthis. In the present study, the activity and regulation ofAP-2 were examined in differentiating human epidermalkeratinocytes. We report that (1) AP-2 transcriptionalactivity decreases in differentiated keratinocytes but re-mains unchanged in differentiation-insensitive squa-mous cell carcinoma cell lines, (2) diminished AP-2transcriptional activity is associated with a loss of spe-cific DNA-bound AP-2 complexes, and (3) there is anincrease in the ability of cytoplasmic extracts, derivedfrom differentiated keratinocytes, to phosphorylateAP-2a and AP-2b when cells differentiate. In contrast,
extracts from differentiation-insensitive squamous cellcarcinoma cells are unable to phosphorylate AP-2 pro-teins. Finally, the phosphorylation of recombinantAP-2a by cytosolic extracts from differentiated kera-tinocytes is associated with decreased AP-2 DNA-bind-ing activity. Combined, these data indicate that AP-2trans-activation and DNA-binding activity decrease askeratinocytes differentiate, and that this decreased ac-tivity is associated with an enhanced ability to phos-phorylate AP-2a and b.
Key words AP-2 � human epidermal keratinocytes �transcription factor
Introduction
The epithelial cells of the epidermis (keratinocytes)maintain the integrity of the epidermal barrier by fol-lowing a tightly regulated, multistep program ofsquamous differentiation (Fuchs, 1990, 1994, 1998; By-rne et al., 1994; Fuchs and Byrne, 1994; Byrne, 1997;Nemes and Steinert, 1999). During squamous differen-tiation, proliferative, undifferentiated keratinocytes,located within the basal layer, are converted to non-proliferative, differentiated keratinocytes and ultimatelyto anuclear corneocytes (Fuchs, 1990, 1998; Byrne et al.,1994; Fuchs and Byrne, 1994; Byrne, 1997; Nemes andSteinert, 1999). As cells migrate through the differenti-ating suprabasal layers, their morphological and bio-chemical characteristics alter in a temporally andspatially specific fashion. Initially, squamous differen-tiation requires repression of proliferation-associatedgenes such as E2F1, cdc2, and p53 or keratins K5 andK14, followed by induction of squamous-specific genessuch as transglutaminase type I (TGaseI), involucrin,loricrin, and filaggrin, and the keratins K1 and K10(Fuchs and Green, 1980; Rothnagel and Rogers, 1984;
Claudia Popa � Alison L. Dahler � Magdalena M. Serewko-Auret � Chung F. Wong � Louise Smith � Liam M. Barnes �Nicholas A. Saunders ( .*)Epithelial Pathobiology GroupCancer Biology ProgramCentre for Immunology and Cancer ResearchUniversity of QueenslandBuilding 1, R Wing, Level 4Princess Alexandra HospitalBrisbane, Queensland, Australia, 4102Tel: 617 3240 5936Fax: 617 3240 5946E-mail: [email protected]
Geoff M. StruttonDepartment of PathologyPrincess Alexandra HospitalBrisbane, Queensland, Australia, 4102
Nicholas A. SaundersDepartment of Physiology and PharmacologyUniversity of QueenslandSt. Lucia, Queensland, Australia, 4072
U.S. Copyright Clearance Center Code Statement: 0301–4681/2004/7205–185 $ 15.00/0
Differentiation (2004) 72:185–197 r International Society of Differentiation 2004

Eichner et al., 1986; Hohl et al., 1991; Younus andGilchrest, 1992; Saunders et al., 1993a, 1993b; Eckertet al., 1997a; Candi et al., 1998). To a largeextent, the ordered regulation of gene expression, asso-ciated with the different stages of squamous differ-entiation, is mediated by alterations in the activityof specific transcriptional activator/repressor proteins(Magnaldo et al., 1993; Eckert and Welter, 1996; Byrne,1997; Eckert et al., 1997b).
A number of transcription factors that are expressedin the epidermis have been implicated in the regulationof squamous differentiation. However, these transcrip-tion factors are often expressed in a variety of other celltypes, raising the question of how their squamous-specific functions may be mediated. Current thinkingsuggests that differentiation specificity is conferredthrough the activities of specific transcription factor fam-ily members acting in combination with one another (Le-ask et al., 1990, 1991; Snape et al., 1990, 1991; Andersenet al., 1993, 1997; Magnaldo et al., 1993; Faus et al.,1994; Eckert and Welter, 1996; Fischer et al., 1996;Morasso et al., 1996; Andreoli et al., 1997; Eckert et al.,1997b; Feng et al., 1997; Kachinskas et al., 1997;Ma et al., 1997; Maytin and Habener, 1998; Rossi et al.,1998; Agarwal et al., 1999; DiSepio et al., 1999; Maytinet al., 1999; Segre et al., 1999; Balasubramanian et al.,2000; Jang et al., 2000; Kaufman and Fuchs, 2000;Li et al., 2000, 2001; Radoja et al., 2000; Sinha et al.,2000; Mazina et al., 2001; Sinha and Fuchs, 2001;Kaufman et al., 2002). One such factor is the AP-2transcription factor family. AP-2 was one of the firsttranscription factors to be cloned and is associated withthe development of neural crest-derived tissues as wellas the epidermis (Mitchell et al., 1987, 1991; Williamset al., 1988; Mitchell and Tjian, 1989; Moser et al., 1995,1997a, 1997b; Oulad-Abdelghani et al., 1996; Schorleet al., 1996; Zhang et al., 1996; Hilger-Eversheim et al.,2000). It consists of five related factors named AP-2a(previously known as AP-2), AP-2b, AP-2g, AP-2d,and AP-2e (Mitchell et al., 1987; Williams et al., 1988;Moser et al., 1993, 1995; Chazaud et al., 1996; Oulad-Abdelghani et al., 1996; Zhao et al., 2001) that bind asdimers (homo- or hetero-) to an AP-2-specific responseelement GCCN3GGC (Imagawa et al., 1987; Mitchellet al., 1987; Williams and Tjian, 1991a, 1991b; Wank-hade et al., 2000). AP-2 family members can also bindto sequences other than the consensus AP-2 site, sug-gesting that AP-2 may mediate transcriptional controlover a larger number of genes than was previouslythought (Gee et al., 1998; Mohibullah et al., 1999;Hilger-Eversheim et al., 2000; Zhao et al., 2001).
There are several lines of evidence to suggest an im-portant role for the AP-2 transcription factor family inepidermal differentiation. For example, AP-2 expres-sion has been reported in cultured keratinocytes andkeratinocyte cell lines (BALB/MK2 cells, SCC9,SCC13, and HaCaTs) as well as in the developing and
adult epidermis (Leask et al., 1990, 1991; Mitchell et al.,1991; Chazaud et al., 1996; Schorle et al., 1996; Wanneret al., 1996; Maytin et al., 1999; Mazina et al., 2001).Furthermore, AP-2 binding sites have been identifiedin the promoters of many keratinocyte-specific genesincluding the keratins K5, K14, K1, K3, K6b, andK17 (Leask et al., 1990, 1991; Byrne and Fuchs, 1993;Magnaldo et al., 1993; Byrne et al., 1994; Milisavljevicet al., 1996; Chen et al., 1997; Wanner et al., 1997; Sinhaet al., 2000; Sinha and Fuchs, 2001; Kaufman et al.,2002), TGaseI (Saunders et al., 1993b; Mariniello et al.,1995), involucrin (Kachinskas et al., 1997), cystatin A(Takahashi et al., 1998, 2000), and superoxide dismutase(St Clair et al., 2002; Xu et al., 2002), and in some in-stances these sites have been shown to be functional(Leask et al., 1991; Snape et al., 1991; Magnaldo et al.,1993; Margalit et al., 1993; Byrne et al., 1994;Milisavljevic et al., 1996; Kachinskas et al., 1997; Sinhaet al., 2000; Takahashi et al., 2000; Kaufman et al., 2002).However, expression of these putative AP-2-responsivegenes is not restricted to a specific layer of the epidermis,suggesting that if AP-2 is involved in their regulation,then the regulation of AP-2 activity must be complex.
It is currently difficult to attribute a specific functionto AP-2 in squamous differentiating tissues. For in-stance, although the expression patterns of the AP-2a,b, and g genes have been described in the developingmouse embryo, and the presence of four isoforms isknown to occur in the adult mouse (Mitchell et al.,1991; Moser et al., 1995, 1997a; Oulad-Abdelghaniet al., 1996; Nottoli et al., 1998), only AP-2a expressionhas been localized to a particular cellular compartment(basal layer) in the newborn (Mitchell et al., 1991) andadult (Maytin et al., 1999) mouse epidermis. In con-trast, the reports (presented by two groups) on AP-2expression in human epidermis appear to be contradic-tory because in vitro AP-2a expression was observed inboth sub-confluent, proliferating keratinocytes as wellas in confluent keratinocytes (Mazina et al., 2001), whilein vivo expression of AP-2a was reported to be in basalkeratinocytes only (Takahashi et al., 1998; Oyama et al.,1999, 2002). To date, only a limited number of studieshave addressed issues relating to the DNA-bindingactivity and trans-activation ability of AP-2 in humankeratinocytes (Sinha et al., 2000; Mazina et al., 2001;Kaufman et al., 2002; Oyama et al., 2002). The presentstudy provides an analysis of AP-2 expression, DNA-binding activity, trans-activation ability, and the phos-phorylation of AP-2 in differentiating keratinocytes.
Methods
Cell culture
Human epidermal keratinocytes (HEKs) were isolated from neo-natal foreskins and cultured as previously described (Jones et al.,
186

1997). To maintain keratinocytes in the proliferative state, theywere trypsinized and serially passaged before reaching confluence.To induce squamous differentiation, HEKs were treated with 50 ng/ml 12-O-tetradecanoyl phorbol-13-acetate (TPA; Fluka, Sydney,Australia), 100U/ml interferon-g (IFNg; Life Technologies, Syd-ney, Australia), or maintained as confluent cultures for 48 hr (Sa-unders and Jetten, 1994; Saunders et al., 1998, 1999a, 1999b; Popaet al., 1999; Dicker et al., 2000a, 2000b; Dahler et al., 2001). Thetongue-derived squamous cell carcinoma cell line, SCC25, waspurchased from American Type Culture Collection (ATCC; Man-assas, VA) and cultured as previously described (Jones et al., 1997).The COLO-16 cells and KJD cells are squamous cell carcinoma celllines derived from a metastatic SCC of the skin and SV40 trans-formed, respectively, and were cultured as previously described(Serewko et al., 2002).
RNA isolation and RT-PCR analysis
Keratinocyte sscDNA was produced as described (Jones et al.,1997). The expression of AP-2a, AP-2b, AP-2g, TGaseI, cdc2, andactin were determined by polymerase chain reaction (PCR) ampli-fication under conditions of linearity with respect to cycle numberand template concentration (Jones et al., 1997; Popa et al., 1999;Dicker et al., 2000a). The following primer sets were used for theamplification of each gene. AP-2a primer sequences were: forwardprimer, 50-GCC AGG AGT CTG GGC TCC TGC-30 and reverseprimer, 50-CCT GCA GGC AGA TTT AAT CCT-30, which gen-erated a PCR product of 561 bp. AP-2b primer sequences were:forward primer, 50-GGC TCG GTG TCC CAA GGA-30 and re-verse primer, 50-GAA GTC ACC CAT TTG GGA-30, which gen-erated a PCR product of 500 bp. AP-2g primer sequences were:forward primer, 50-GGA GCA GCA ATG GGA ATC-30 and re-verse primer, 50-AGG AGA GTC ACA TGA GCG-30, which gen-erated a PCR product of 843 bp. TGaseI, cdc2, and actin primersequences have been described (Dahler et al., 1998; Popa et al.,1999). PCR amplifications were carried out under the followingconditions: denaturation at 941C for 40 sec, annealing at 551C (AP-2a, AP-2g, actin, and cdc2), 581C (AP-2b), or 651C (TGaseI) for40 sec, and extension at 721C for 45 sec. The PCR products werefractionated by agarose gel electrophoresis and transferred onto anylon membrane (Schleicher and Schuell Inc., Keene, NH) as pre-viously described (Sambrook et al., 1989). The membranes werethen probed with the appropriate 32P-labeled probes and washed athigh stringency (Popa et al., 1999; Saunders et al., 1999b). Theimage was analyzed on an SF phosphorimager (Molecular Dy-namics, Sydney, Australia) and inequalities in reverse transcriptionand PCR amplification were normalized to actin expression of thesame sample.
Nuclear extract preparations and EMSA studies
Nuclear extracts were prepared as described (Jones et al., 1997;Dahler et al., 1998; Serewko et al., 2002). Electromobility shift as-says were performed by incubating radioactively labeled, double-stranded AP-2 oligonucleotide consensus sequence (50-GAT CGAACT GAC CGC CCG CGG CCC GT-30; the consensus sequencefor the actual binding site for AP-2 is indicated in bold; Promega,Sydney, Australia) with 2mg nuclear extract in 1� gel shift buffer(4% glycerol [v/v], 1mM MgCl2, 0.5mM EDTA, 0.5mM DTT,50mMNaCl, 10mM Tris-Cl [pH 7.5], and 50mg/ml poly [dI.dC]-[dI.dC]; final reaction volume 10ml). This AP-2 oligonucleotide,derived from the human metallothionein IIA promoter region, wasradioactively labeled with T4 polynucleotide kinase (New EnglandBiolabs, Beverley, MA) as per the manufacturer’s instructions and30,000–60,000 cpm/ml was used in binding reactions. In specific andnonspecific competition experiments, an excess of 100� unlabeledAP-2 oligonucleotide or cdxA oligonucleotide (50-AGA TCT GGTACC ATT TAA GCC CTC GAG ATC TA-30; Margalit et al.,
1993) was added, respectively, and incubated for 15min. In someinstances, binding assays were performed using 0.1mg of the humanrecombinant AP-2 protein (Promega). In other experiments, thebinding reactions included 50mM ATP as controls for the phos-phorylation of AP-2 proteins. The reaction was then loaded onto a4% polyacrylamide gel and electrophoresed at 200 volts for 1.5 hr.The gel was dried under vacuum and exposed to x-ray film.
Western blotting
Nuclear protein (10 mg) was fractionated on a 12% SDS-polyacryl-amide gel containing a 3% SDS-polyacrylamide stacking gel, thentransferred onto a PVDF membrane (Millipore Corporation, Bed-ford, MA). The membrane was blocked with 3% skim milk/PBST(15mM sodium phosphate, 150mMNaCl, and 0.5% Tween 20 [pH7.3]) for 3 hr and probed with either 1:2,000, 1:500, or 1:500 rabbitpolyclonal antibody raised against human AP-2a, b, or g (SantaCruz Biotechnologies, Santa Cruz, CA), respectively, at 41C over-night. Following three 5-min washes in PBST, the membranes wereincubated with 1:3,000 horseradish peroxidase (HRP)-conjugatedanti-rabbit IgG raised in sheep (Amrad Operations Pty Ltd, Mel-bourne, Australia) for 1 hr at room temperature. The membraneswere washed for 5min in PBST three times and the signal wasdetected by chemiluminescence (Dahler et al., 2001). Western blotswere then scanned and the digital image was quantified by Image-QuaNT (Molecular Dynamics).
Immunohistochemistry
To detect the protein expression and localization of the various AP-2isoforms, immunohistochemical staining was carried out on paraf-fin-embedded sections containing normal human neonatal andadult skin. The sections were prepared as described (Serewko et al.,2002). Following a 2-hr blocking period in 3% milk/PBST, sectionswere incubated for 1 hr with a rabbit polyclonal antibody againsthuman AP-2a (1:1,000), AP-2b (1:30), or AP-2g (1:300). The sec-tions were then incubated in a 1:1,500 dilution of biotinylated goatanti-rabbit IgG (DAKO Corporation, Carpinteria, CA) for 30minfollowed by a 1:300 concentration of HRP-conjugated streptavidinfor 30min (DAKO Corporation). Slides were washed three timesfor 5min each with PBST following each antibody incubation.Visualization was performed by reaction with diaminobenzidine(DAKO Corporation) and reactions were terminated in water.Control sections were incubated with blocking peptides corre-sponding to each AP-2 isoform (Santa Cruz Biotechnologies)or normal rabbit IgG (DAKO Corporation). These two controlmethods gave identical results.
Transfection studies
Proliferative or differentiated keratinocytes were transiently trans-fected and assayed for reporter gene activity 48 hr post-transfection.To determine AP-2 activity, keratinocytes were transfected with theA2BCAT4 reporter construct containing three copies of the AP-2response element from the human metallothionein IIA gene (Will-iams et al., 1988) driving the expression of the CAT gene from anE1B minimal promoter. To normalize for transfection efficiency,keratinocytes were co-transfected with the b-actin luciferase report-er construct. As a control, the reporter activity of the BCAT4 con-struct, which does not contain the AP-2 response elements, wassubtracted from all values obtained with the A2BCAT4 reporter.To further validate the trans-activation ability of AP-2, the aboveexperiments were repeated using a second AP-2 binding site-con-taining reporter construct, p21-Luc (Zeng et al., 1997). A p21-Lucconstruct lacking the AP-2 binding sites was also used as controland b-actin-CAT was used to normalize for transfection efficiency.
187

Transfections were carried out in 10-cm2 wells with either 1 mgA2BCAT4 or p21-Luc, 0.3 mg b-actin luciferase or b-actin-CAT,respectively, and 4mg lipofectamine reagent (Life Technologies)using previously described methods (Dicker et al., 2000a; Dahleret al., 2001). The media was replaced 5 hr later with either completeSFM (keratinocyte serum-free medium; Invitrogen, Sydney, Aus-tralia) or complete SFM containing the differentiation-inducingagent TPA. In other experiments, the squamous cell carcinoma cellline SCC25, COLO-16, and KJDs were transiently transfected andassayed for AP-2 reporter gene activity following transfection witheffectine using methods described by the manufacturers (Life Tech-nologies). CAT reporter activity and luciferase activity were esti-mated as described (Dicker et al., 2000a, 2000b).
Generation of recombinant AP-2 proteins and phosphorylationreactions
Recombinant, biotin-tagged AP-2 proteins were generated by clon-ing full-length AP-2a, b, and g cDNA (PCR amplified from thehuman AP-2a pCMX-PL1 vector and the mouse AP-2b pCMX-PL1 and AP-2g pCMX-PL vectors) into the PinPoint Xa-1 T-vector(Promega). Expression of biotinylated AP-2 proteins was verified asper the manufacturer’s protocol. To confirm that the bacterialsupernatants contained the recombinant, biotinylated AP-2a, b,and g isoforms, equal amounts of the respective supernatants wereelectrophoresed on a 10% SDS-PAGE gel and immunodetectionwas performed using the isoform-specific antibodies describedabove (Fig. 1). Figure 1 also serves to confirm the isoformspecificity of the AP-2 antibodies.
In phosphorylation experiments, 20ml of bacterial supernatantwas incubated with 10ml TetraLink Tetrameric Avidin Resin(Promega), rotated gently at 41C for 4 hr, and washed four timeswith NETN buffer (20mM Tris-Cl [pH 8.0], 1mM EDTA, 0.5%NP-40, and 100mM NaCl). The protein-bound TetraLink was re-suspended in 10ml NETN buffer and incubated with nuclear extractor cytoplasmic extract obtained either from 50% or 2-day confluentkeratinocytes. The reaction was made up to 40 ml using kinasebuffer (50mM Tris-HCl [pH 7.4], 10mM MgCl2, 1mM DTT,50 mMATP, and 5ml/10ml [g-32P]ATP [3,000 Ci/mmol]) and incu-bated at room temperature for 20min. The reaction was terminatedby the addition of 10ml 10� STOP buffer (77mM Na2HPO4,23mM NaH2PO4, 100mM Na pyrophosphate, 100mM EDTA,and 1mg/ml BSA). The sample was washed three times with NETNbuffer, resuspended in 4� protein sample buffer, and boiled for3min to release the proteins. The samples were then elect-rophoresed on a 10% SDS-PAGE gel, transferred onto PVDF
membrane, and exposed to x-ray film. In other experiments, thephosphorylation reaction was incubated with or without thephosphatase inhibitors NaF (5mM) and Na orthovanadate (0.05mM).
Results
AP-2 expression in human epidermal keratinocytes
The expression of AP-2 was investigated in an in vitrosystem of primary HEK cultures in which the differen-tiation status could be manipulated (Saunders et al.,1993a, 1998a, 1999b; Saunders and Jetten, 1994; Joneset al., 1997; Dahler et al., 1998, 2001; Popa et al., 1999;Dicker et al., 2000a). In the present study, mRNA levelswere determined by RT-PCR. The robustness of theRT-PCR method was confirmed by a decrease in themRNA level of the proliferation-specific gene cdc2, anda concomitant increase in the mRNA level of the dif-ferentiation-specific gene TGaseI, in response to differ-entiation stimuli (Fig. 2; Dahler et al., 1998; Popa et al.,1999; Dicker et al., 2000b). The mRNA expression lev-els of three AP-2 isoforms (a, b, and g) were examined invitro and shown to be present in both proliferative cul-tures of HEKs and in HEKs differentiated by variousstimuli (Fig. 2). The induction of differentiation was notassociated with consistent alterations in the mRNAlevels of any of the three AP-2 isoforms examined (Fig.2). In accordance with the mRNA data, Western blotanalysis of nuclear extracts from proliferating and dif-ferentiated HEKs also revealed no substantial changesin the protein levels for AP-2a, b, and g irrespective ofthe differentiation status (Fig. 3).
The expression of AP-2 protein isoforms was alsoinvestigated in vivo by immunohistochemical staining ofhuman skin. Consistent with the in vitro studies, AP-2a,b, and g protein expression was observed throughoutthe epidermis in both the proliferative, basal layer aswell as the suprabasal differentiating layers (Figs. 4A–4C). This is in accordance with an in vitro study inwhich AP-2a protein expression was shown to occur inboth the basal and superficial confluent cells of humankeratinocytes (Mazina et al., 2001) but differs from thebasal cell-specific staining reported by Oyama et al.(1999, 2002). In vivo, expression of all three AP-2 iso-forms (a, b, and g) was primarily localized to the nu-cleus of the keratinocytes although staining of thecytoplasm was also visible (Fig. 4).
AP-2 activity during squamous differentiation
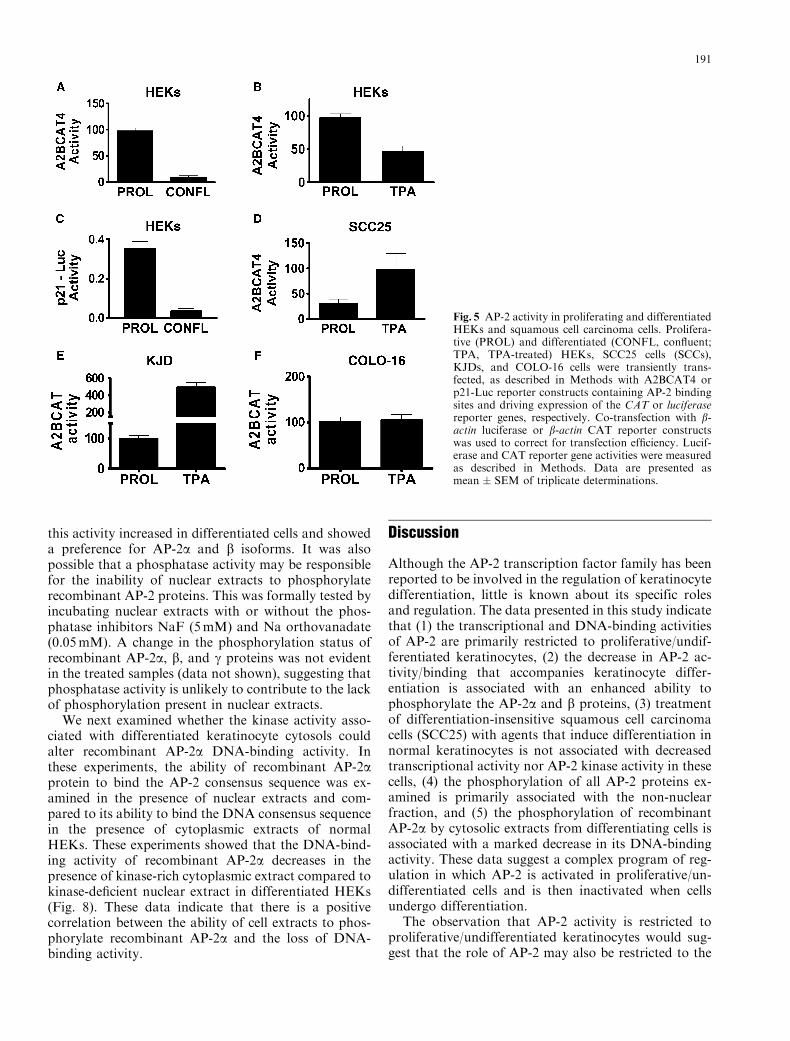
Alterations in the activity of AP-2 were investigated byintroducing a reporter construct, A2BCAT4, containingthree AP-2 consensus binding sites, linked to the CATreporter gene, into cultured HEKs. In contrast to themRNA and protein data, these results clearly demon-strate a marked decrease in AP-2 reporter activity in
Fig. 1 Expression of recombinant AP-2a, b, and gproteins. Equalamounts of recombinant biotin-tagged (bt-tagged) AP-2a, b, and gproteins were electrophoresed on a 10% SDS-PAGE gel, trans-ferred onto PVDF membrane, and probed with the respective hu-man polyclonal AP-2 antibodies as described in Methods. Proteinloading was similar between samples as shown by the streptavidin-horseradish peroxidase antibody (SA-HRP).
188

cells induced to differentiate following growth to con-fluence or treatment with TPA (Figs. 5A, 5B). A similardecrease in AP-2 trans-activation was observed in dif-ferentiated keratinocytes when a second AP-2-respon-sive reporter, consisting of the p21 promoter linked tothe luciferase gene, was used (Fig. 5C). In both in-stances, reporter constructs with mutated AP-2 consen-sus sites were assayed for activity (data not shown) andthese values were subtracted from the values obtainedusing the AP-2-responsive constructs. These data indi-cate that AP-2 activity is primarily restricted to prolif-erative, undifferentiated keratinocytes. An associationbetween keratinocyte proliferation and AP-2 activitywas further examined in three differentiation-resistantsquamous cell carcinoma cell lines: SCC25, COLO-16,and SV40-transformed KJD cells (Jones et al., 1997;Dicker et al., 2000a, 2000b; Serewko et al., 2002).Treatment of these cell lines with the differentiation-inducing agent TPA was used because squamous cellcarcinoma cells are resistant to TPA-induced growthinhibition and differentiation (Jones et al., 1997; Dickeret al., 2000a, 2000b; Serewko et al., 2002). A reductionin the AP-2 reporter activity did not occur in TPA-treated SCC25, KJD, or COLO-16 cells when comparedto untreated cells (Figs. 5D–5F). In fact, A2BCAT ac-tivity was increased approximately 4-fold in the SCC25and KJD cell lines in response to TPA but remained
Fig. 2 AP-2a, b, and gmRNA expression in differentiating HEKs.Total RNA was extracted from HEKs induced to differentiate bytreatment with TPA (50 ng/ml), IFNg (100U/ml), or growth toconfluence (CONFL) and 2mg was used for RT-PCR analysis. ThemRNA levels of AP-2a, b, and g were estimated by phosphorimageanalysis of Southern blots of PCR products. Expression levels forthe proliferation-specific marker, cdc2, and differentiation-specificmarker, TGaseI, are also shown. Data are presented as a percentageof the expression level of proliferating HEKs (except TGaseI ) andas mean � SEM of no less than three experiments. All samples arenormalized to actin expression. Points reflect various times between0 and 48 hr post-treatment. For confluent cells, points are presentedas 50%, 80%, and 100% confluence.
Fig. 3 Western blot analysis of AP-2a, b, and g in HEKs. Nuclearprotein was extracted from proliferating (P) HEKs or HEKs dif-ferentiated by treatment with TPA (T5 50 ng/ml, 48 hr), IFNg(I5 100U/ml, 48 hr), or growth to confluence for 48 hr (C). Proteinwas electrophoresed and then transferred to PVDF membranewhich was probed using either rabbit anti-human AP-2a, b, or gpolyclonal antibodies and visualized by chemiluminescence. Proteinloading equalities are shown by the ratio between AP-2 isoform andamido black staining (ratios reported under bands).
189

unaltered in the COLO-16 cells. These data provideclear evidence that the loss of appropriate AP-2 regu-lation is associated with the squamous cell carcinomaphenotype.
The decrease in AP-2 activity in the presence of per-sistent AP-2a, b, and g expression raised the question ofwhat was the molecular basis for the decrease in AP-2activity in differentiated keratinocytes. One mechanismby which such a decrease may occur is by decreasedbinding of the AP-2 proteins to their AP-2 consensus-binding site. We thus examined the binding of AP-2proteins to the AP-2 consensus site in proliferating anddifferentiating keratinocytes by EMSA analysis. AP-2DNA-binding activity in HEK nuclear extracts could beresolved into three specific complexes (a–c), of whichonly complexes ‘‘a’’ and ‘‘c’’ were consistently decreasedin differentiated cells (Fig. 6). These complexes werecompeted for by unlabeled AP-2 probe but not by thenonspecific probe (Fig. 6). In contrast, other complexes(x and y; Fig. 6) were not so easily competed for by theunlabeled AP-2 probe and most likely represent non-specific bands. These results demonstrate that there is aclose correlation between decreases in AP-2 trans-acti-vation ability and decreases in DNA-binding activity(complexes a and c) of AP-2 in cells induced to differ-entiate. Furthermore, multiple specific AP-2-bindingcomplexes are visible, suggesting that AP-2 associateseither with other cofactors or as homo- or heterodimersin combination with other AP-2 isoforms. The associ-ation of AP-2a and g with other DNA-binding cofac-tors has recently been reported to occur in keratinocytes(Kaufman et al., 2002; Braganca et al., 2003).
Post-translational modification of AP-2 proteins
Having demonstrated that both AP-2 reporter activityand DNA-binding activity decrease in differentiatedkeratinocytes, we sought to identify whether thesechanges were associated with alterations in the phos-phorylation of AP-2 proteins during keratinocyte dif-ferentiation. However, traditional analyses of in vivophosphorylation status by immunoprecipitation of AP-2 were unsuccessful using available AP-2 antibodies.We therefore carried out experiments using biotin-tagged AP-2a, b, and g proteins, which were incubatedwith HEK nuclear or cytoplasmic extracts in the pres-ence of 32P-labeled ATP. Our data demonstrate that thephosphorylation status of recombinant AP-2a, b, and gproteins increases in the presence of cytoplasmic ex-tracts from keratinocytes but is not evident in the pres-ence of nuclear extracts from HEK cells (Fig. 7).Moreover, differentiation was associated with a markedincrease in recombinant AP-2a phosphorylation activityand to a lesser extent AP-2b (Fig. 7). In contrast, bothnuclear and cytoplasmic extracts from the squamouscell carcinoma cell line SCC25 show little, if any, phos-phorylation of recombinant AP-2 proteins (data notshown). These data indicate that cytosolic extracts ofnormal keratinocytes contain a kinase that can phos-phorylate recombinant AP-2 isoforms. Significantly,
Fig. 4 Immunohistochemical staining of AP-2a b, and g in normalskin. Human skin sections were stained for either AP-2a (A), b (B),or g (C) using either rabbit anti-human AP-2a, b, or g polyclonalantibodies and visualized by DAB staining as described in Meth-ods. Control slides (shown as insets) showed no immunoreactivityand were prepared by incubating sections in the presence of rabbitIgG antibody or blocking peptide. Negative controls using AP-2a,b, or g peptides (not shown) were similar to the IgG controls.Magnification, 25� .
190
this activity increased in differentiated cells and showeda preference for AP-2a and b isoforms. It was alsopossible that a phosphatase activity may be responsiblefor the inability of nuclear extracts to phosphorylaterecombinant AP-2 proteins. This was formally tested byincubating nuclear extracts with or without the phos-phatase inhibitors NaF (5mM) and Na orthovanadate(0.05mM). A change in the phosphorylation status ofrecombinant AP-2a, b, and g proteins was not evidentin the treated samples (data not shown), suggesting thatphosphatase activity is unlikely to contribute to the lackof phosphorylation present in nuclear extracts.
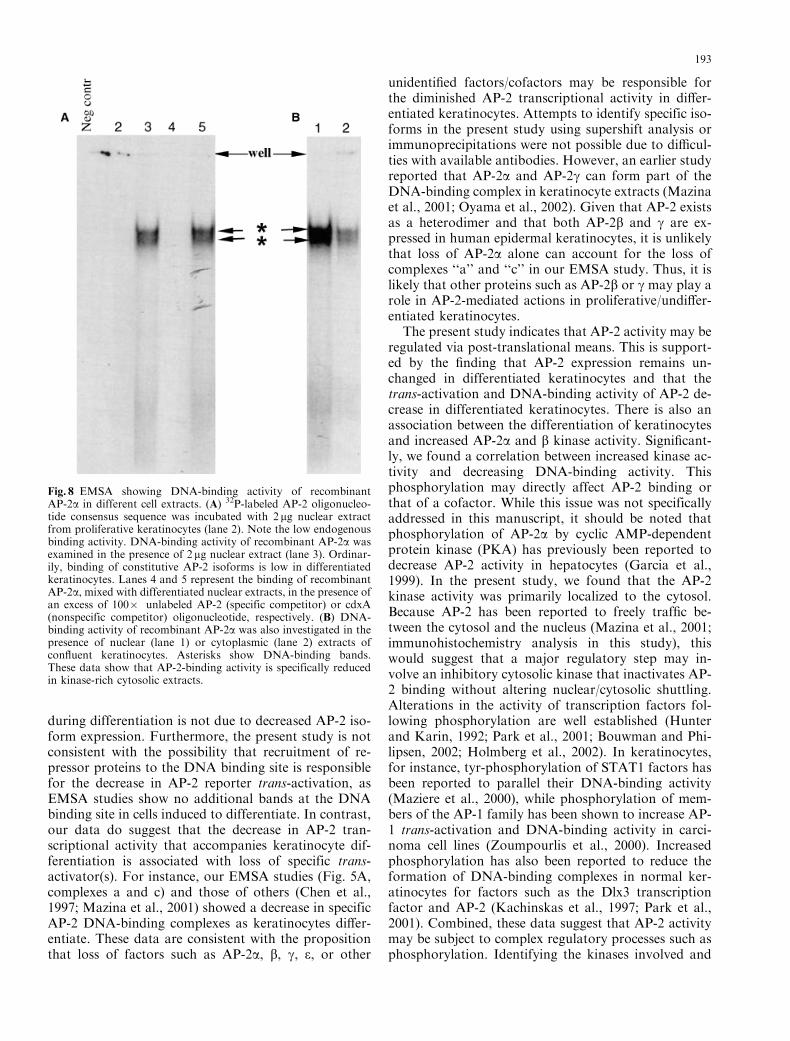
We next examined whether the kinase activity asso-ciated with differentiated keratinocyte cytosols couldalter recombinant AP-2a DNA-binding activity. Inthese experiments, the ability of recombinant AP-2aprotein to bind the AP-2 consensus sequence was ex-amined in the presence of nuclear extracts and com-pared to its ability to bind the DNA consensus sequencein the presence of cytoplasmic extracts of normalHEKs. These experiments showed that the DNA-bind-ing activity of recombinant AP-2a decreases in thepresence of kinase-rich cytoplasmic extract compared tokinase-deficient nuclear extract in differentiated HEKs(Fig. 8). These data indicate that there is a positivecorrelation between the ability of cell extracts to phos-phorylate recombinant AP-2a and the loss of DNA-binding activity.
Discussion
Although the AP-2 transcription factor family has beenreported to be involved in the regulation of keratinocytedifferentiation, little is known about its specific rolesand regulation. The data presented in this study indicatethat (1) the transcriptional and DNA-binding activitiesof AP-2 are primarily restricted to proliferative/undif-ferentiated keratinocytes, (2) the decrease in AP-2 ac-tivity/binding that accompanies keratinocyte differ-entiation is associated with an enhanced ability tophosphorylate the AP-2a and b proteins, (3) treatmentof differentiation-insensitive squamous cell carcinomacells (SCC25) with agents that induce differentiation innormal keratinocytes is not associated with decreasedtranscriptional activity nor AP-2 kinase activity in thesecells, (4) the phosphorylation of all AP-2 proteins ex-amined is primarily associated with the non-nuclearfraction, and (5) the phosphorylation of recombinantAP-2a by cytosolic extracts from differentiating cells isassociated with a marked decrease in its DNA-bindingactivity. These data suggest a complex program of reg-ulation in which AP-2 is activated in proliferative/un-differentiated cells and is then inactivated when cellsundergo differentiation.
The observation that AP-2 activity is restricted toproliferative/undifferentiated keratinocytes would sug-gest that the role of AP-2 may also be restricted to the
Fig. 5 AP-2 activity in proliferating and differentiatedHEKs and squamous cell carcinoma cells. Prolifera-tive (PROL) and differentiated (CONFL, confluent;TPA, TPA-treated) HEKs, SCC25 cells (SCCs),KJDs, and COLO-16 cells were transiently trans-fected, as described in Methods with A2BCAT4 orp21-Luc reporter constructs containing AP-2 bindingsites and driving expression of the CAT or luciferasereporter genes, respectively. Co-transfection with b-actin luciferase or b-actin CAT reporter constructswas used to correct for transfection efficiency. Lucif-erase and CAT reporter gene activities were measuredas described in Methods. Data are presented asmean � SEM of triplicate determinations.
191

proliferative/undifferentiated keratinocyte. This is sup-ported by the inability of the growth inhibitor-resistantsquamous cell carcinoma cell lines SCC25, COLO-16,and KJD cells to decrease AP-2 reporter activity in re-sponse to known differentiation stimuli such as TPA.Earlier studies have also reported an association be-tween AP-2 factors, cellular proliferation, and themaintenance of the undifferentiated state in other celltypes as well as in keratinocytes (Mitchell et al., 1991;Byrne et al., 1994; Chen et al., 1997; Buehner et al.,1998; Bisgrove and Godbout, 1999; Maytin et al., 1999;Sinha et al., 2000; Takahashi et al., 2000; Kaufmanet al., 2002; Oyama et al., 2002; Pfisterer et al., 2002;Jager et al., 2003). For example, loss of AP-2a has been
associated with the decreased proliferation of embry-onic fibroblasts (Pfisterer et al., 2002), while mice de-ficient for AP-2g display an embryonic lethal phenotypedue to decreased proliferation of placental cells (Wer-ling and Schorle, 2002). AP-2 has also been shown toactivate the proliferation-associated receptor tyrosinekinase c-erb-B2 (Bosher et al., 1996), and AP-2g isoverexpressed in many mammary tumors and mamma-ry tumor cell lines (Turner et al., 1998). More recently,it has been reported that AP-2 factors are involved inthe inhibition of differentiation-specific genes (Chenet al., 1997; Pfisterer et al., 2002; Jager et al., 2003).Hence, the present study and earlier studies stronglysupport the contention that the AP-2 transcription fac-tor family can play both pro-proliferative roles as wellas be involved in actively maintaining the undifferenti-ated cell phenotype. Despite these data, specific targetgenes for the AP-2 isoforms in proliferative/undifferen-tiated keratinocytes remain unknown.
Given that AP-2 activity is decreased during kera-tinocyte differentiation and that AP-2 activity was notdecreased in similarly treated differentiation-resistantSCC cells, it was of interest to examine the molecularbasis for the regulation of AP-2 activity. Our mRNAand protein expression studies in vitro and in vivo sug-gest that this decrease in AP-2 transcriptional activity
Fig. 6 EMSA showing DNA-binding activity of AP-2. 32P-labeledAP-2 oligonucleotide consensus sequence was incubated with 2 mgnuclear extract. In specific (Sp. Comp.) and nonspecific (Nonsp.Comp.) competition experiments, an excess of 100� unlabeled AP-2or cdxA oligonucleotide was added, respectively. The reaction wasseparated by 4% polyacrylamide gel electrophoresis, dried, andexposed to film. Labels P and C show DNA binding in proliferativeHEKs or HEKs grown to confluence (differentiated), respectively.Labels a–c show the multiple specific DNA-binding complexes;x and y denote nonspecific complexes.
Fig. 7 Phosphorylation of recombinant AP-2a, b, and g proteins byhuman epidermal keratinocyte extracts. Equal amounts of recom-binant AP-2a, b, and g proteins were incubated for 20min at RTwith nuclear (NE) or cytoplasmic extract (CE) obtained from pro-liferating (PROL) or differentiated (CONFL) HEKs. Followingwashes in NETN buffer, the samples were incubated in phosphor-ylation reaction buffer as described in the Methods. Proteins werethen subjected to SDS-PAGE electrophoresis, blotted, and exposedto x-ray film. To check for loading equivalence the blot was probedwith HRP-conjugated streptavidin.
192
during differentiation is not due to decreased AP-2 iso-form expression. Furthermore, the present study is notconsistent with the possibility that recruitment of re-pressor proteins to the DNA binding site is responsiblefor the decrease in AP-2 reporter trans-activation, asEMSA studies show no additional bands at the DNAbinding site in cells induced to differentiate. In contrast,our data do suggest that the decrease in AP-2 tran-scriptional activity that accompanies keratinocyte dif-ferentiation is associated with loss of specific trans-activator(s). For instance, our EMSA studies (Fig. 5A,complexes a and c) and those of others (Chen et al.,1997; Mazina et al., 2001) showed a decrease in specificAP-2 DNA-binding complexes as keratinocytes differ-entiate. These data are consistent with the propositionthat loss of factors such as AP-2a, b, g, e, or other
unidentified factors/cofactors may be responsible forthe diminished AP-2 transcriptional activity in differ-entiated keratinocytes. Attempts to identify specific iso-forms in the present study using supershift analysis orimmunoprecipitations were not possible due to difficul-ties with available antibodies. However, an earlier studyreported that AP-2a and AP-2g can form part of theDNA-binding complex in keratinocyte extracts (Mazinaet al., 2001; Oyama et al., 2002). Given that AP-2 existsas a heterodimer and that both AP-2b and g are ex-pressed in human epidermal keratinocytes, it is unlikelythat loss of AP-2a alone can account for the loss ofcomplexes ‘‘a’’ and ‘‘c’’ in our EMSA study. Thus, it islikely that other proteins such as AP-2b or g may play arole in AP-2-mediated actions in proliferative/undiffer-entiated keratinocytes.
The present study indicates that AP-2 activity may beregulated via post-translational means. This is support-ed by the finding that AP-2 expression remains un-changed in differentiated keratinocytes and that thetrans-activation and DNA-binding activity of AP-2 de-crease in differentiated keratinocytes. There is also anassociation between the differentiation of keratinocytesand increased AP-2a and b kinase activity. Significant-ly, we found a correlation between increased kinase ac-tivity and decreasing DNA-binding activity. Thisphosphorylation may directly affect AP-2 binding orthat of a cofactor. While this issue was not specificallyaddressed in this manuscript, it should be noted thatphosphorylation of AP-2a by cyclic AMP-dependentprotein kinase (PKA) has previously been reported todecrease AP-2 activity in hepatocytes (Garcia et al.,1999). In the present study, we found that the AP-2kinase activity was primarily localized to the cytosol.Because AP-2 has been reported to freely traffic be-tween the cytosol and the nucleus (Mazina et al., 2001;immunohistochemistry analysis in this study), thiswould suggest that a major regulatory step may in-volve an inhibitory cytosolic kinase that inactivates AP-2 binding without altering nuclear/cytosolic shuttling.Alterations in the activity of transcription factors fol-lowing phosphorylation are well established (Hunterand Karin, 1992; Park et al., 2001; Bouwman and Phi-lipsen, 2002; Holmberg et al., 2002). In keratinocytes,for instance, tyr-phosphorylation of STAT1 factors hasbeen reported to parallel their DNA-binding activity(Maziere et al., 2000), while phosphorylation of mem-bers of the AP-1 family has been shown to increase AP-1 trans-activation and DNA-binding activity in carci-noma cell lines (Zoumpourlis et al., 2000). Increasedphosphorylation has also been reported to reduce theformation of DNA-binding complexes in normal ker-atinocytes for factors such as the Dlx3 transcriptionfactor and AP-2 (Kachinskas et al., 1997; Park et al.,2001). Combined, these data suggest that AP-2 activitymay be subject to complex regulatory processes such asphosphorylation. Identifying the kinases involved and
Fig. 8 EMSA showing DNA-binding activity of recombinantAP-2a in different cell extracts. (A) 32P-labeled AP-2 oligonucleo-tide consensus sequence was incubated with 2 mg nuclear extractfrom proliferative keratinocytes (lane 2). Note the low endogenousbinding activity. DNA-binding activity of recombinant AP-2a wasexamined in the presence of 2mg nuclear extract (lane 3). Ordinar-ily, binding of constitutive AP-2 isoforms is low in differentiatedkeratinocytes. Lanes 4 and 5 represent the binding of recombinantAP-2a, mixed with differentiated nuclear extracts, in the presence ofan excess of 100� unlabeled AP-2 (specific competitor) or cdxA(nonspecific competitor) oligonucleotide, respectively. (B) DNA-binding activity of recombinant AP-2a was also investigated in thepresence of nuclear (lane 1) or cytoplasmic (lane 2) extracts ofconfluent keratinocytes. Asterisks show DNA-binding bands.These data show that AP-2-binding activity is specifically reducedin kinase-rich cytosolic extracts.
193

the molecular basis for their activation will provide im-portant insight into the control of squamous differen-tiation. Given that extracts from the squamous cellcarcinoma cell line SCC25 show little, if any, phos-phorylation of recombinant AP-2 proteins and no al-terations in DNA-binding activity in response tonormal differentiation stimuli, this would also suggestthat identification of AP-2 regulation may give signif-icant insight into squamous neoplasia.
Acknowledgments The authors would like to thank ProfessorTrevor Williams (University of Colorado, Health Science Center,Denver, CO) for the generous gift of the A2BCAT and BCAT4reporter constructs. The authors are grateful to Associate ProfessorW. el-Deiry (Departments of Medicine, Genetics, and Pharmacol-ogy, University of Pennsylvania School of Medicine, Philadelphia,PA) for providing the p21-luciferase constructs and Dr. R. Buettner(Institute of Pathology, Medical School RWTH, Aachen, Germa-ny) for providing the hAP-2a pCMX-PL1, mAP-2b pCMX-PL1,and mAP-2g-pSG5 vectors.
Contract grant sponsor: Queensland Cancer Fund; contractgrant number: 98/QCFN001G. Contract grant sponsor: The Asso-ciation for International Cancer Research and The Australian Na-tional Health and Medical Research Committee; contract grantnumber: 301075. Contract grant sponsor: The Garnett Passe andRodney Williams Memorial Foundation. Contract grant sponsor:Australian Postgraduate Award. Contract grant sponsor: UQ Post-doctoral Research Fellow Award.
References
Agarwal, C., Efimova, T., Welter, J.F., Crish, J.F. and Eckert, R.L.(1999) CCAAT/enhancer-binding proteins. A role in regulationof human involucrin promoter response to phorbol ester. J BiolChem 274:6190–6194.
Andersen, B., Schonemann, M.D., Flynn, S.E., Pearse, R.V. II,Singh, H. and osenfeld, M.G. (1993) Skn-1a and Skn-1i: twofunctionally distinct Oct-2-related factors expressed in epidermis.Science 260:78–82.
Andersen, B., Weinberg, W.C., Rennekampff, O., McEvilly, R.J.,Bermingham, J.R. Jr., Hooshmand, F., Vasilyev, V., Hansbrough,J.F., Pittelkow, M.R., Yuspa, S.H. and Rosenfeld, M.G. (1997)Functions of the POU domain genes Skn-1a/i and Tst-1/Oct-6/SCIP in epidermal differentiation. Genes Dev 11:1873–1884.
Andreoli, J.M., Jang, S.I., Chung, E., Coticchia, C.M., Steinert,P.M. and Markova, N.G. (1997) The expression of a novel, ep-ithelium-specific ets transcription factor is restricted to the mostdifferentiated layers in the epidermis. Nucleic Acids Res 25:4287–4295.
Balasubramanian, S., Agarwal, C., Efimova, T., Dubyak, G.R.,Banks, E., Welter, J. and Eckert, R.L. (2000) Thapsigargin sup-presses phorbol ester-dependent human involucrin promoter ac-tivity by suppressing CCAAT-enhancer-binding protein alpha(C/EBPa) DNA binding. Biochem J 350(Pt 3): 791–796.
Bisgrove, D.A. and Godbout, R. (1999) Differential expression ofAP-2a and AP-2b in the developing chick retina: repression ofR-FABP promoter activity by AP-2. Dev Dyn 214:195–206.
Bosher, J.M., Totty, N.F., Hsuan, J.J., Williams, T. and Hurst,H.C. (1996) A family of AP-2 proteins regulates c-erbB-2 ex-pression in mammary carcinoma. Oncogene 13:1701–1707.
Bouwman, P. and Philipsen, S. (2002) Regulation of the activityof Sp1-related transcription factors. Mol Cell Endocrinol 195:27–38.
Braganca, J., Eloranta, J.J., Bamforth, S.D., Ibbitt, J.C., Hurst,H.C. and Bhattacharya, S. (2003) Physical and functional inter-
actions between AP-2 transcription factors, p300/CBP, and CIT-ED2. J Biol Chem 12:12.
Buehner, A.W., Stolz, W. and Buettner, R. (1998) AP-2, the rel-evant AP-2 isoform for skin differentiation? J Invest Dermatol110:539.
Byrne, C. (1997) Regulation of gene expression in developing ep-idermal epithelia. Bioessays 19:691–698.
Byrne, C. and Fuchs, E. (1993) Probing keratinocyte and differen-tiation specificity of the human K5 promoter in vitro and intransgenic mice. Mol Cell Biol 13:3176–3190.
Byrne, C., Tainsky, M. and Fuchs, E. (1994) Programming gene ex-pression in developing epidermis. Development 120:2369–2383.
Candi, E., Tarcsa, E., Digiovanna, J.J., Compton, J.G., Elias,P.M., Marekov, L.N. and Steinert, P.M. (1998) A highly con-served lysine residue on the head domain of type II keratins isessential for the attachment of keratin intermediate filaments tothe cornified cell envelope through isopeptide crosslinking bytransglutaminases. Proc Natl Acad Sci USA 95:2067–2072.
Chazaud, C., Oulad-Abdelghani, M., Bouillet, P., Decimo, D.,Chambon, P. and Dolle, P. (1996) AP-2.2, a novel gene related toAP-2, is expressed in the forebrain, limbs and face during mouseembryogenesis. Mech Dev 54:83–94.
Chen, T.T., Wu, R.L., Castro-Munozledo, F. and Sun, T.T. (1997)Regulation of K3 keratin gene transcription by Sp1 and AP-2 indifferentiating rabbit corneal epithelial cells. Mol Cell Biol17:3056–3064.
Dahler, A.L., Jones, S.J., Dicker, A.J. and Saunders, N.A. (1998)Keratinocyte growth arrest is associated with activation of atranscriptional repressor element in the human cdk1 promoter.J Cell Physiol 177:474–482.
Dahler, A.L., Cavanagh, L.L. and Saunders, N.A. (2001) Suppres-sion of keratinocyte growth and differentiation by transforminggrowth factor b1 involves multiple signaling pathways. J InvestDermatol 116:266–274.
Dicker, A.J., Popa, C., Dahler, A.L., Serewko, M.M., Hilditch-Maguire, P.A., Frazer, I.H. and Saunders, N.A. (2000a) E2F-1induces proliferation-specific genes and suppresses squamousdifferentiation-specific genes in human epidermal keratinocytes.Oncogene 19:2887–2894.
Dicker, A.J., Serewko, M.M., Dahler, A.L., Khanna, K.K., Kaur,P., Li, A., Strutton, G.M. and Saunders, N.A. (2000b) Func-tional characterization of cultured cells derived from an intraep-idermal carcinoma of the skin (IEC-1). Exp Cell Res 258:352–360.
DiSepio, D., Bickenbach, J.R., Longley, M.A., Bundman, D.S.,Rothnagel, J.A. and Roop, D.R. (1999) Characterization ofloricrin regulation in vitro and in transgenic mice. Differentiation64:225–235.
Eckert, R.L. and Welter, J.F. (1996) Transcription factor regula-tion of epidermal keratinocyte gene expression. Mol Biol Rep23:59–70.
Eckert, R.L., Crish, J.F. and Robinson, N.A. (1997a) The epider-mal keratinocyte as a model for the study of gene regulation andcell differentiation. Physiol Rev 77:397–424.
Eckert, R.L., Crish, J.F., Banks, E.B. and Welter, J.F. (1997b) Theepidermis: genes on—genes off. J Invest Dermatol 109:501–509.
Eichner, R., Sun, T.T. and Aebi, U. (1986) The role of keratinsubfamilies and keratin pairs in the formation of human epider-mal intermediate filaments. J Cell Biol 102:1767–1777.
Faus, I., Hsu, H.J. and Fuchs, E. (1994) Oct-6: a regulator of ker-atinocyte gene expression in stratified squamous epithelia. MolCell Biol 14:3263–3275.
Feng, X., Peng, Z.H., Di, W., Li, X.Y., Rochette-Egly, C.,Chambon, P., Voorhees, J.J. and Xiao, J.H. (1997) Suprabasalexpression of a dominant-negative RXR a mutant in transgenicmouse epidermis impairs regulation of gene transcription andbasal keratinocyte proliferation by RAR-selective retinoids.Genes Dev 11:59–71.
Fischer, D.F., Gibbs, S., van De Putte, P. and Backendorf, C.(1996) Interdependent transcription control elements regulate the
194

expression of the SPRR2A gene during keratinocyte terminaldifferentiation. Mol Cell Biol 16:5365–5374.
Fuchs, E. (1990) Epidermal differentiation: the bare essentials.J Cell Biol 111(6 Pt 2): 2807–2814.
Fuchs, E. (1994) Epidermal differentiation and keratin gene ex-pression. Princess Takamatsu Symp 24:290–302.
Fuchs, E. (1998) Beauty is skin deep: the fascinating biology of theepidermis and its appendages. Harvey Lect 94:47–77.
Fuchs, E. and Byrne, C. (1994) The epidermis: rising to the surface.Curr Opin Genet Dev 4:725–736.
Fuchs, E. and Green, H. (1980) Changes in keratin gene expressionduring terminal differentiation of the keratinocyte. Cell 19:1033–1042.
Garcia, M.A., Campillos, M., Marina, A., Valdivieso, F. andVazquez, J. (1999) Transcription factor AP-2 activity is modu-lated by protein kinase A-mediated phosphorylation. FEBS Lett444:27–31.
Gee, M.S., Sarkisian, C.J. and el-Deiry, W.S. (1998) Identificationof a novel AP-2 consensus DNA binding site. Biochem BiophysRes Commun 243:307–316.
Hilger-Eversheim, K., Moser, M., Schorle, H. and Buettner, R.(2000) Regulatory roles of AP-2 transcription factors in verte-brate development, apoptosis and cell-cycle control. Gene 260:1–12.
Hohl, D., Mehrel, T., Lichti, U., Turner, M.L., Roop, D.R. andSteinert, P.M. (1991) Characterization of human loricrin. Struc-ture and function of a new class of epidermal cell envelope pro-teins. J Biol Chem 266:6626–6636.
Holmberg, C.I., Tran, S.E., Eriksson, J.E. and Sistonen, L. (2002)Multisite phosphorylation provides sophisticated regulation oftranscription factors. Trends Biochem Sci 27:619–627.
Hunter, T. and Karin, M. (1992) The regulation of transcription byphosphorylation. Cell 70:375–387.
Imagawa, M., Chiu, R. and Karin, M. (1987) Transcriptionfactor AP-2 mediates induction by two different signal-trans-duction pathways: protein kinase C and cAMP. Cell 51:251–260.
Jager, R., Werling, U., Rimpf, S., Jacob, A. and Schorle, H. (2003)Transcription factor AP-2g stimulates proliferation and apopto-sis and impairs differentiation in a transgenic model. Mol CancerRes 1:921–929.
Jang, S.I., Karaman-Jurukovska, N., Morasso, M.I., Steinert, P.M.and Markova, N.G. (2000) Complex interactions betweenepidermal POU domain and activator protein 1 transcriptionfactors regulate the expression of the profilaggrin gene innormal human epidermal keratinocytes. J Biol Chem 275:15295–15304.
Jones, S.J., Dicker, A.J., Dahler, A.L. and Saunders, N.A. (1997)E2F as a regulator of keratinocyte proliferation: implications forskin tumor development. J Invest Dermatol 109:187–193.
Kachinskas, D.J., Qin, Q., Phillips, M.A. and Rice, R.H. (1997)Arsenate suppression of human keratinocyte programming.Mutat Res 386:253–261.
Kaufman, C.K. and Fuchs, E. (2000) It’s got you covered. NF-kBin the epidermis. J Cell Biol 149:999–1004.
Kaufman, C.K., Sinha, S., Bolotin, D., Fan, J. and Fuchs, E. (2002)Dissection of a complex enhancer element: maintenance of ker-atinocyte specificity but loss of differentiation specificity. MolCell Biol 22:4293–4308.
Leask, A., Rosenberg, M., Vassar, R. and Fuchs, E. (1990) Reg-ulation of a human epidermal keratin gene: sequences and nu-clear factors involved in keratinocyte-specific transcription.Genes Dev 4:1985–1998.
Leask, A., Byrne, C. and Fuchs, E. (1991) Transcription factor AP2and its role in epidermal-specific gene expression. Proc Natl AcadSci USA 88:7948–7952.
Li, M., Indra, A.K., Warot, X., Brocard, J., Messaddeq, N., Kato,S., Metzger, D. and Chambon, P. (2000) Skin abnormalitiesgenerated by temporally controlled RXRa mutations in mouseepidermis. Nature 407:633–636.
Li, M., Chiba, H., Warot, X., Messaddeq, N., Gerard, C.,Chambon, P. and Metzger, D. (2001) RXR-a ablation in skinkeratinocytes results in alopecia and epidermal alterations.Development 128:675–688.
Ma, S., Rao, L., Freedberg, I.M. and Blumenberg, M. (1997)Transcriptional control of K5, K6, K14, and K17 keratingenes by AP-1 and NF-kB family members. Gene Expr 6:361–370.
Magnaldo, T., Vidal, R.G., Ohtsuki, M., Freedberg, I.M. and Blu-menberg, M. (1993) On the role of AP2 in epithelial-specific geneexpression. Gene Expr 3:307–315.
Margalit, Y., Yarus, S., Shapira, E., Gruenbaum, Y. and Fainsod,A. (1993) Isolation and characterization of target sequences ofthe chicken CdxA homeobox gene. Nucleic Acids Res 21:4915–4922.
Mariniello, L., Qin, Q., Jessen, B.A. and Rice, R.H. (1995) Ker-atinocyte transglutaminase promoter analysis. Identification of afunctional response element. J Biol Chem 270:31358–31363.
Maytin, E.V. and Habener, J.F. (1998) Transcription factors C/EBP a, C/EBP b, and CHOP (Gadd153) expressed during thedifferentiation program of keratinocytes in vitro and in vivo.J Invest Dermatol 110:238–246.
Maytin, E.V., Lin, J.C., Krishnamurthy, R., Batchvarova, N., Ron,D., Mitchell, P.J. and Habener, J.F. (1999) Keratin 10 gene ex-pression during differentiation of mouse epidermis requires tran-scription factors C/EBP and AP-2. Dev Biol 216:164–181.
Maziere, C., Dantin, F., Dubois, F., Santus, R. and Maziere, J.(2000) Biphasic effect of UVA radiation on STAT1 activity andtyrosine phosphorylation in cultured human keratinocytes. FreeRadic Biol Med 28:1430–1437.
Mazina, O.M., Phillips, M.A., Williams, T., Vines, C.A., Cherr,G.N. and Rice, R.H. (2001) Redistribution of transcription fac-tor AP-2a in differentiating cultured human epidermal cells.J Invest Dermatol 117:864–870.
Milisavljevic, V., Freedberg, I.M. and Blumenberg, M. (1996)Characterization of nuclear protein binding sites in the promoterof keratin K17 gene. DNA Cell Biol 15:65–74.
Mitchell, P.J. and Tjian, R. (1989) Transcriptional regulation inmammalian cells by sequence-specific DNA binding proteins.Science 245:371–378.
Mitchell, P.J., Wang, C. and Tjian, R. (1987) Positive and negativeregulation of transcription in vitro: enhancer-binding protein AP-2 is inhibited by SV40 T antigen. Cell 50:847–861.
Mitchell, P.J., Timmons, P.M., Hebert, J.M., Rigby, P.W. andTjian, R. (1991) Transcription factor AP-2 is expressed in neuralcrest cell lineages during mouse embryogenesis. Genes Dev5:105–119.
Mohibullah, N., Donner, A., Ippolito, J.A. and Williams, T. (1999)SELEX and missing phosphate contact analyses reveal flexibilitywithin the AP-2[a] protein: DNA binding complex. Nucleic AcidsRes 27:2760–2769.
Morasso, M.I., Markova, N.G. and Sargent, T.D. (1996) Regula-tion of epidermal differentiation by a Distal-less homeodomaingene. J Cell Biol 135(6 Pt 2): 1879–1887.
Moser, M., Pscherer, A., Bauer, R., Imhof, A., Seegers, S., Kersc-her, M. and Buettner, R. (1993) The complete murine cDNAsequence of the transcription factor AP-2. Nucleic Acids Res21:4844.
Moser, M., Imhof, A., Pscherer, A., Bauer, R., Amselgruber, W.,Sinowatz, F., Hofstadter, F., Schule, R. and Buettner, R. (1995)Cloning and characterization of a second AP-2 transcriptionfactor: AP-2 b. Development 121:2779–2788.
Moser, M., Ruschoff, J. and Buettner, R. (1997a) Comparativeanalysis of AP-2 a and AP-2 b gene expression during murineembryogenesis. Dev Dyn 208:115–124.
Moser, M., Pscherer, A., Roth, C., Becker, J., Mucher, G., Zerres,K., Dixkens, C., Weis, J., Guay-Woodford, L., Buettner, R. andFassler, R. (1997b) Enhanced apoptotic cell death of renal ep-ithelial cells in mice lacking transcription factor AP-2b. GenesDev 11:1938–1948.
195

Nemes, Z. and Steinert, P.M. (1999) Bricks and mortar of the ep-idermal barrier. Exp Mol Med 31:5–19.
Nottoli, T., Hagopian-Donaldson, S., Zhang, J., Perkins, A. andWilliams, T. (1998) AP-2-null cells disrupt morphogenesis of theeye, face, and limbs in chimeric mice. Proc Natl Acad Sci USA95:13714–13719.
Oulad-Abdelghani, M., Bouillet, P., Chazaud, C., Dolle, P. andChambon, P. (1996) AP-2.2: a novel AP-2-related transcriptionfactor induced by retinoic acid during differentiation of P19 em-bryonal carcinoma cells. Exp Cell Res 225:338–347.
Oyama, N., Iwatsuki, K., Homma, Y. and Kaneko, F. (1999) In-duction of transcription factor AP-2 by inflammatory cytokinesin human keratinocytes. J Invest Dermatol 113:600–606.
Oyama, N., Takahashi, H., Tojo, M., Iwatsuki, K., Iizuka, H.,Nakamura, K., Homma, Y. and Kaneko, F. (2002) Differentproperties of three isoforms (a, b, and g) of transcription factorAP-2 in the expression of human keratinocyte genes. Arch De-rmatol Res 294:273–280.
Park, G.T., Denning, M.F. and Morasso, M.I. (2001) Phosphor-ylation of murine homeodomain protein Dlx3 by protein kinaseC. FEBS Lett 496:60–65.
Pfisterer, P., Ehlermann, J., Hegen, M. and Schorle, H. (2002) Asubtractive gene expression screen suggests a role of transcriptionfactor AP-2 a in control of proliferation and differentiation.J Biol Chem 277:6637–6644.
Popa, C., Dicker, A.J., Dahler, A.L. and Saunders, NA. (1999)Cytochrome P450, CYP26AI, is expressed at low levels in humanepidermal keratinocytes and is not retinoic acid-inducible. Br JDermatol 141:460–468.
Radoja, N., Komine, M., Jho, S.H., Blumenberg, M. and Tomic-Canic, M. (2000) Novel mechanism of steroid action in skinthrough glucocorticoid receptor monomers. Mol Cell Biol20:4328–4339.
Rossi, A., Jang, S.I., Ceci, R., Steinert, P.M. and Markova, N.G.(1998) Effect of AP1 transcription factors on the regulation oftranscription in normal human epidermal keratinocytes. J InvestDermatol 110:34–40.
Rothnagel, J.A. and Rogers, G.E. (1984) Transglutaminase-medi-ated cross-linking in mammalian epidermis. Mol Cell Biochem58:113–119.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecularcloning: a laboratory manual. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, New York, pp. 9.39–39.55.
Saunders, N.A. and Jetten, A.M. (1994) Control of growth regu-latory and differentiation-specific genes in human epidermal ker-atinocytes by interferon g. Antagonism by retinoic acid andtransforming growth factor b 1. J Biol Chem 269:2016–2022.
Saunders, N.A., Smith, R.J. and Jetten, A.M. (1993a) Regulationof proliferation-specific and differentiation-specific genes duringsenescence of human epidermal keratinocyte and mammaryepithelial cells. Biochem Biophys Res Commun 197:46–54.
Saunders, N.A., Bernacki, S.H., Vollberg, T.M. and Jetten, A.M.(1993b) Regulation of transglutaminase type I expression insquamous differentiating rabbit tracheal epithelial cells and hu-man epidermal keratinocytes: effects of retinoic acid and phorbolesters. Mol Endocrinol 7:387–398.
Saunders, N.A., Dicker, A.J., Jones, S.J. and Dahler, A.L. (1998)E2F1 messenger RNA is destabilized in response to a growthinhibitor in normal human keratinocytes but not in a squamouscarcinoma cell line. Cancer Res 58:1646–1649.
Saunders, N.A., Popa, C., Serewko, M.M., Jones, S.J., Dicker, A.J.and Dahler, A.L. (1999a) Histone deacetylase inhibitors: novel an-ticancer agents. Expert Opin Investig Drugs 8:1611–1621.
Saunders, N., Dicker, A., Popa, C., Jones, S. and Dahler, A.(1999b) Histone deacetylase inhibitors as potential anti-skin can-cer agents. Cancer Res 59:399–404.
Schorle, H., Meier, P., Buchert, M., Jaenisch, R. and Mitchell, P.J.(1996) Transcription factor AP-2 essential for cranial closure andcraniofacial development. Nature 381:235–238.
Segre, J.A., Bauer, C. and Fuchs, E. (1999) Klf4 is a transcriptionfactor required for establishing the barrier function of the skin.Nat Genet 22:356–360.
Serewko, M.M., Popa, C., Dahler, A.L., Smith, L., Strutton, G.M.,Coman, W., Dicker, A.J. and Saunders, N.A. (2002) Alterationsin gene expression and activity during squamous cell carcinomadevelopment. Cancer Res 62:3759–3765.
Sinha, S. and Fuchs, E. (2001) Identification and dissection of anenhancer controlling epithelial gene expression in skin. Proc NatlAcad Sci USA 98:2455–2460.
Sinha, S., Degenstein, L., Copenhaver, C. and Fuchs, E. (2000)Defining the regulatory factors required for epidermal gene ex-pression. Mol Cell Biol 20:2543–2555.
Snape, A.M., Jonas, E.A. and Sargent, T.D. (1990) KTF-1, a tran-scriptional activator of Xenopus embryonic keratin expression.Development 109:157–165.
Snape, A.M., Winning, R.S. and Sargent, T.D. (1991) Transcrip-tion factor AP-2 is tissue-specific in Xenopus and is closely relatedor identical to keratin transcription factor 1 (KTF-1). Develop-ment 113:283–293.
St Clair, D.K., Porntadavity, S., Xu, Y. and Kiningham, K. (2002)Transcription regulation of human manganese superoxide di-smutase gene. Methods Enzymol 349:306–312.
Takahashi, H., Asano, K., Kinouchi, M., Ishida-Yamamoto, A.,Wuepper, K.D. and Iizuka, H. (1998) Structure and transcrip-tional regulation of the human cystatin A gene. The 12-O-tet-radecanoylphorbol-13-acetate (TPA) responsive element-2 site(� 272 to � 278) on cystatin A gene is critical for TPA-depend-ent regulation. J Biol Chem 273:17375–17380.
Takahashi, H., Oyama, N., Itoh, Y., Ishida-Yamamoto, A.,Kaneko, F. and Iizuka, H. (2000) Transcriptional factor AP-2gincreases human cystatin A gene transcription of keratinocytes.Biochem Biophys Res Commun 278:719–723.
Turner, B.C., Zhang, J., Gumbs, A.A., Maher, M.G., Kaplan, L.,Carter, D., Glazer, P.M., Hurst, H.C., Haffty, B.G. andWilliams, T. (1998) Expression of AP-2 transcription factorsin human breast cancer correlates with the regulation ofmultiple growth factor signalling pathways. Cancer Res 58:5466–5472.
Wankhade, S., Yu, Y., Weinberg, J., Tainsky, M.A. and Kannan,P. (2000) Characterization of the activation domains ofAP-2 family transcription factors. J Biol Chem 275:29701–29708.
Wanner, R., Zhang, J., Henz, B.M. and Rosenbach, T. (1996) AP-2gene expression and modulation by retinoic acid during keratin-ocyte differentiation. Biochem Biophys Res Commun 223:666–669.
Wanner, R., Zhang, J., Dorbic, T., Mischke, D., Henz, B.M.,Wittig, B. and Rosenbach, T. (1997) The promoter of theHaCaT keratinocyte differentiation-related gene keratin 4 con-tains a functional AP-2 binding site. Arch Dermatol Res289:705–708.
Werling, U. and Schorle, H. (2002) Transcription factor gene AP-2g essential for early murine development. Mol Cell Biol 22:3149–3156.
Williams, T. and Tjian, R. (1991a) Characterization of a dime-rization motif in AP-2 and its function in heterologous DNA-binding proteins. Science 251:1067–1071.
Williams, T. and Tjian, R. (1991b) Analysis of the DNA-bindingand activation properties of the human transcription factorAP-2. Genes Dev 5:670–682.
Williams, T., Admon, A., Luscher, B. and Tjian, R. (1988) Cloningand expression of AP-2, a cell-type-specific transcription factorthat activates inducible enhancer elements. Genes Dev 2(12A):1557–1569.
Xu, Y., Porntadavity, S. and St Clair, D.K. (2002) Transcriptionalregulation of the human manganese superoxide dismutase gene:the role of specificity protein 1 (Sp1) and activating protein-2(AP-2). Biochem J 362(Pt 2): 401–412.
196

Younus, J. and Gilchrest, B.A. (1992) Modulation of mRNA levelsduring human keratinocyte differentiation. J Cell Physiol152:232–239.
Zeng, Y.X., Somasundaram, K. and el-Deiry, W.S. (1997) AP2inhibits cancer cell growth and activates p21WAF1/CIP1 ex-pression. Nat Genet 15:78–82.
Zhang, J., Hagopian-Donaldson, S., Serbedzija, G., Elsemore, J.,Plehn-Dujowich, D., McMahon, A.P., Flavell, R.A. and Will-iams, T. (1996) Neural tube, skeletal and body wall defects inmice lacking transcription factor AP-2. Nature 381:238–241.
Zhao, F., Satoda, M., Licht, J.D., Hayashizaki, Y. and Gelb, B.D.(2001) Cloning and characterization of a novel mouse AP-2transcription factor, AP-2d, with unique DNA bind-ing and transactivation properties. J Biol Chem 276:40755–40760.
Zoumpourlis, V., Papassava, P., Linardopoulos, S., Gillespie, D.,Balmain, A. and Pintzas, A. (2000) High levels of phosphoryl-ated c-Jun, Fra-1, Fra-2 and ATF-2 proteins correlate withmalignant phenotypes in the multistage mouse skincarcinogenesis model. Oncogene 19:4011–4021.
197


















