
Anti-multi drug resistant pathogen activity of ...
9
Korean Journal of Microbiology (2016) Vol. 52, No. 3, pp. 327-335 pISSN 0440-2413 DOI http://dx.doi.org/10.7845/kjm.2016.6033 eISSN 2383-9902 Copyright ⓒ 2016, The Microbiological Society of Korea Article Anti-multi drug resistant pathogen activity of siderochelin A, produced by a novel Amycolatopsis sp. KCTC 29142 Dong-Ryung Lee 1† , Jinhua Cheng 1,2† , Sung-Kwon Lee 1 , Hee-Jeon Hong 3 , Jaekyeong Song 4 , Seung Hwan Yang 5 * , and Joo-Won Suh 1,2 * 1 Center for Nutraceutical and Pharmaceutical Materials Myongji University, Yongin 17058, Republic of Korea 2 Division of Bioscience and Bioinformatics, College of Natural Science, Myongji University, Yongin 17058, Republic of Korea 3 Department of Biological and Medical Sciences, Faculty of Health and Life Sciences, Oxford Brookes University, Oxford OX3 0BP, United Kingdom 4 Division of Agricultural Microbiology, National Academy of Agricultural Science, Rural Development Administration, Jeonju 56050, Republic of Korea 5 Department of Biotechnology, Chonnam National University, Gwangju 59626, Republic of Korea Amycolatopsis sp. KCTC 29142 로부터 유래된 siderochelin A 의 다제 내성 균주에 대한 항균활성 이동령 1† ・ 성금화 1,2† ・ 이성권 1 ・ 홍희전 3 ・ 송재경 4 ・ 양승환 5 * ・ 서주원 1,2 * 1 명지대학교 농생명바이오식의약소재개발사업단, 2 명지대학교 생명과학정보학부 생명과학과, 3 옥스퍼드 브룩스대학 생의과학과, 4 국립농업과학원 농업미생물과, 5 전남대학교 생명산업공학과 (Received June 17, 2016; Revised August 3, 2016; Accepted August 9, 2016) ABSTRACT: A novel Amycolatopsis strain KCTC 29142 was isolated and characterized based on the polyphasic taxonomic analysis including morphological observation, phylogenetic analysis, physiological and chemotaxonomic characteristics. The ethyl acetate extract of strain KCTC 29142 culture broth showed strong antibacterial activity and the active compound was identified as siderochelin A, a ferrous-ion chelating compound. In this study, siderochelin A showed good activity against multi-drug resistant pathogens, including Acinetobacter baumanii, methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant Staphylococcus aureus (VRSA), and Escherichia coli (E. coli ). The minimum inhibitory activity against clinical isolates was also determined. Key words: Amycolatopsis, antibacterial activity, anti-multi drug resistant pathogens, Siderochelin A † These authors contributed equally to this work. *For correspondence. (S.H. Yang) E-mail: [email protected]; Tel.: +82-61-659-7306; Fax: +82-61-659-7309 (J.W. Suh) E-mail: [email protected]; Tel.: +82-31-330-6190; Fax: +82-31-321-7361 Multi drug-resistant pathogens (MDRPs) are a critical world- wide problem because of their resistance to a bevy of commer- cially available antibiotics and their high mortality rate. Available evidence showed that the proportion of Gram-positive and negative bacteria resistant to commonly used antibiotics is increasing (Lode, 2009; Lin and Lan, 2014). The mechanisms of this resistance are often complex, and include production of β -lactamases, upregulated efflux pumps, and target site mutations (Sakoulasand and Moellering, 2008). It is reported that patients with infections due to MDRP have higher costs than do patients with infections due to antimicrobial- susceptible organisms (Cosgrove, 2006). So it is necessary to develop new drugs for control of MDRP (Anderson, 2003). Iron is an essential bioelement for most bacteria, which involved in a variety of metabolic and informational cellular pathways (Miethke and Marahiel, 2007). Since the importance
Transcript of Anti-multi drug resistant pathogen activity of ...

Korean Journal of Microbiology (2016) Vol. 52, No. 3, pp. 327-335 pISSN 0440-2413DOI http://dx.doi.org/10.7845/kjm.2016.6033 eISSN 2383-9902Copyright ⓒ 2016, The Microbiological Society of Korea
Article
Anti-multi drug resistant pathogen activity of siderochelin A,
produced by a novel Amycolatopsis sp. KCTC 29142
Dong-Ryung Lee1†
, Jinhua Cheng1,2†
, Sung-Kwon Lee1, Hee-Jeon Hong
3, Jaekyeong Song
4, Seung Hwan Yang
5*, and
Joo-Won Suh1,2*
1Center for Nutraceutical and Pharmaceutical Materials Myongji University, Yongin 17058, Republic of Korea
2Division of Bioscience and Bioinformatics, College of Natural Science, Myongji University, Yongin 17058, Republic of Korea
3Department of Biological and Medical Sciences, Faculty of Health and Life Sciences, Oxford Brookes University, Oxford OX3
0BP, United Kingdom4Division of Agricultural Microbiology, National Academy of Agricultural Science, Rural Development Administration, Jeonju
56050, Republic of Korea5Department of Biotechnology, Chonnam National University, Gwangju 59626, Republic of Korea
Amycolatopsis sp. KCTC 29142로부터 유래된 siderochelin A의 다제 내성
균주에 대한 항균활성
이동령1† ・ 성금화
1,2† ・ 이성권
1 ・ 홍희전
3 ・ 송재경
4 ・ 양승환
5* ・ 서주원1,2*
1명지대학교 농생명바이오식의약소재개발사업단,
2명지대학교 생명과학정보학부 생명과학과,
3옥스퍼드 브룩스대학 생의과학과,
4국립농업과학원 농업미생물과,
5전남대학교 생명산업공학과
(Received June 17, 2016; Revised August 3, 2016; Accepted August 9, 2016)
ABSTRACT: A novel Amycolatopsis strain KCTC 29142 was isolated and characterized based on the polyphasic taxonomic analysis
including morphological observation, phylogenetic analysis, physiological and chemotaxonomic characteristics. The ethyl acetate extract of strain KCTC 29142 culture broth showed strong antibacterial activity and the active compound was identified as siderochelin A, a ferrous-ion chelating compound. In this study, siderochelin A showed good activity against multi-drug resistant pathogens, including
Acinetobacter baumanii, methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant Staphylococcus aureus (VRSA), and Escherichia coli (E. coli ). The minimum inhibitory activity against clinical isolates was also determined.
Key words: Amycolatopsis, antibacterial activity, anti-multi drug resistant pathogens, Siderochelin A
†These authors contributed equally to this work.
*For correspondence. (S.H. Yang) E-mail: [email protected];
Tel.: +82-61-659-7306; Fax: +82-61-659-7309
(J.W. Suh) E-mail: [email protected];
Tel.: +82-31-330-6190; Fax: +82-31-321-7361
Multi drug-resistant pathogens (MDRPs) are a critical world-
wide problem because of their resistance to a bevy of commer-
cially available antibiotics and their high mortality rate. Available
evidence showed that the proportion of Gram-positive and
negative bacteria resistant to commonly used antibiotics is
increasing (Lode, 2009; Lin and Lan, 2014). The mechanisms
of this resistance are often complex, and include production of
β-lactamases, upregulated efflux pumps, and target site mutations
(Sakoulasand and Moellering, 2008). It is reported that patients
with infections due to MDRP have higher costs than do patients
with infections due to antimicrobial- susceptible organisms
(Cosgrove, 2006). So it is necessary to develop new drugs for
control of MDRP (Anderson, 2003).
Iron is an essential bioelement for most bacteria, which
involved in a variety of metabolic and informational cellular
pathways (Miethke and Marahiel, 2007). Since the importance

328 ∙ Lee et al.
미생물학회지 제52권 제3호
of iron in the growth of bacteria, many studies has assessed the
potential viability of iron chelators as therapeutic agents against
various microbes (Chan et al., 2009; Kim and Shin, 2009).
Especially in the case of multi-drug resistant bacteria, which
are no longer effective for treatment by antibiotics, iron chelators
are assessed as alternative therapeutic agents (Thomson et al.,
2012).
Siderochelins, a ferrous-ion chelating agents, were first
discovered from fermentation broth of Nocardia sp. SC11340
(Liu et al., 1981). There are six analogues of siderochelin;
siderochelin A and B (from Nocardia sp. SC11340), siderochelin
C (from Actinomycetes sp.), and siderochelin D, E, F (from
Amycolatopsis sp. LZ149) (Liu et al., 1981; Mitscher et al.,
1984; Lu et al., 2015). These siderochelins showed a broad-
spectrum of antibacterial activity, however, the inhibitory
activity of siderochelins on MDRPs have not been studied yet.
Presently we report the phenotypic, physiological, chemo-
taxonomic, and phylogenetic characterization of a novel strain
KCTC 29142 isolated from a soil sample collected from Ami
Mountain, South Korea. The culture supernatant of this strain
displayed inhibitory activities on MDRPs. The active compound
was isolated and analyzed using 1H- and
13C-nulcear magnetic
resonance (NMR) and heteronuclear multiple bond correlation
(HMBC) spectral analysis, and finally identified as the ferrous-
ion chelating compound, siderochelin A. This is the first report
of anti-MDRP activities of siderochelin A, and suggests that
siderochelin A can be developed as a supplementary agent on
the treatment of infections caused by MDRPs.
Materials and Methods
Bacterial strain
Strain KCTC 29142 was isolated from the soil sample
collected from Ami Mountain in Chungcheongnam-do province,
Republic of Korea (latitude 36°50′50.4″N, 126°39′53.2″E)
from February 23 2007. One gram of soil was suspended in
0.85% NaCl, and 100 μl of the suspension was spread on
Humic acid-Vitamin (HV) agar medium, the plates were
incubated at 28°C for 2 weeks. The single colony was picked
using a sterile toothpick and streaked on Bennett’s agar medium
(glucose 10 g/L, peptone 2 g/L, yeast extract 1 g/L, beef extract
1 g/L, and agar 15 g/L) for cultivation. The strain was finally
stored in 20% (v/v) glycerol suspensions at -80°C.
Morphological, physiological, and biochemical cha-
racteristics
Culture characteristics of strain KCTC 29142 were observed
on International Streptomyces Project (ISP), Bennett’s and S
agar media (Shiring and Gottlieb, 1966; Baker, 1990). The
physiological and biochemical features of strain KCTC 29142
were determined according to previously described procedures
(Waksman and Henrici, 1943; Becker et al., 1965). Sugar
utilization was evaluated by using ISP9 medium supplemented
with 1% (w/v) of various sugars individually as sole carbon
source. Nitrogen source utilization was assessed by degradation
of casein, xanthine, gelatin, and tyrosine as described previously
(Gordon et al., 1974).
Phylogenetic analysis, DNA G+C content, and DNA-
DNA hybridization
Genomic DNA was extracted and purified using the ExgeneTM
cell SV kit (GeneAll). The 16S rRNA gene was amplified using
the universal primer set 27F/1492R (Lane, 1991) and sequenced
by Solgent. The resulting 16S rRNA gene sequence (1.5 kb)
was aligned with available 16S rRNA gene sequences using the
EzTaxon-e server (http://eztaxon-e.ezbiocloud.net/) (Kim et
al., 2012). The 16S rRNA gene sequences of closely related
type strains were obtained from the GenBank database and
multiple alignments were conducted with the CLUSTAL_X
program, version 1.8 (Thompson et al., 1997). The phylogenetic
tree was constructed with neighbor-joining (Saitou and Nei,
1987) and Tajima-Nei (Tajima and Nei, 1984) methods using
MEGA 4 software (Tamura et al., 2007). Robustness of
individual branches was evaluated by bootstrapping of 1,000
replications. The G+C contents of the total DNA and DNA-
DNA hybridization between the strain KCTC 29142 and the
reference type strains were performed as described before
(Lechevalier et al., 1986).
Isolation and identification of antibacterial compound
The supernatant from flask fermentation was obtained
following centrifugation and solvent partitioned in a step-wise

Anti-MDRP activity of siderochelin from a novel Amycolatopsis sp. ∙ 329
Korean Journal of Microbiology, Vol. 52, No. 3
Fig. 1. The 16S rRNA gene sequence-based phylogenetic tree generated using the neighbor-joining method shows the relationships of strain KCTC 29142
with other Amycolatopsis species. Numbers at nodes indicate percentage levels of bootstrap support based on a neighbor-joining analysis of 1,000
replications. The scale bar denotes 0.02 substitutions per nucleotide position.
Fig. 1. The 16S rRNA gene sequence-based phylogenetic tree generated using the neighbor-joining method shows the relationships of strain KCTC 29142
with other Amycolatopsis species. Numbers at nodes indicate percentage levels of bootstrap support based on a neighbor-joining analysis of 1,000
replications. The scale bar denotes 0.02 substitutions per nucleotide position.
fashion to purify the antibacterial compounds. The supernatant
was extracted using ethyl acetate (EtOAc; 1:1, v/v). The solvent
layer was evaporated to dryness to yield a yellowish extract.
The extract was dissolved in methanol, applied to a Sephadex
LH-20 gel filtration column, and eluted with methanol. Fractions
displaying activity against MRSA were pooled and subjected
to semi-preparative HPLC. The reverse-phase ODS column
(YMC-Pack ODS-A column, 250 × 10 mm, 5 μm, YMC) was
eluted with 18% acetonitrile containing 1% acetic acid at a flow
rate of 2 ml/min. The compound was monitored by ultraviolet
absorbance at 254 nm. Chemical structures of purified compound
was analyzed by nuclear magnetic resonance (NMR, 1D, 2D). 1H- and
13C-NMR spectra were obtained on an Advance DPX
500 MHz NMR spectrometer (Bruker), recorded in a deuterated
chloroform (CDCl3) solution.
Evaluation of the antibacterial activity
Antibacterial activity was measured by the disc diffusion
method. Mueller-Hinton agar medium was mixed with 0.1 ml
of bacterial suspension (105 CFU/ml). Paper discs which were
soaked with 30 μg of EtOAc extract or siderochelin A were
placed on the agar surface. The plates were incubated at 37°C
for 18 h, and the antibacterial activities were assessed by
measuring the diameter of inhibition zone (mm).
Determination of the minimum inhibitory concentration
(MIC)
MIC values of the siderochelin A against clinical isolates
were determined by agar dilution method. Siderochelin A was
added into the Mueller-Hinton (MH) agar at the concentration
ranging from 0.06–128.00 μg/ml by two-fold serial dilution.
Ten microliters of each test strain was inoculated onto the agar
surface by striking, and incubated at 28°C for 18 h. MIC was
expressed as the lowest concentration of inhibited growth of
visible colony on the agar surface.
Results and Discussion
Phylogenetic analysis, determination of DNA G+C
content, and DNA-DNA hybridization
The phylogenetic tree affiliated strain KCTC 29142 with the
genus Amylcolatopsis (Fig. 1). Phylogenetic relatedness based
on the 16S rRNA gene of KCTC 29142 indicated that the closest
relatives were Amycolatopsis keratiniphila subsp. nogabecina
DSM 44586T (98%), A. decaplanina DSM 44594
T (98.3%),
and A. orientalis IMSNU 20058T (97%). The DNA G+C content
of strain KCTC 29142 was 73.9 mol%. Members of genus
Amylcolatopsis have DNA G+C contents in the range 66–73

330 ∙ Lee et al.
미생물학회지 제52권 제3호
Fig. 2. Morphology of strain KCTC 29142 and the closely related strains.
Amycolatopsis orientalis IMSNU 20058, A. keratiniphila DSM44586, A.
decaplanina DSM 44594 and strains KCTC 29142 were grown on the
Bennett’s agar medium at 28°C for 7 days.
mol% (Tseng et al., 2006). The DNA-DNA hybridization
between strain KCTC 29142 and its nearest phylogenetic
neighbors, A. keratiniphila subsp. nogabecina DSM 44586T,
A. decaplanina DSM44594T, and A. orientalis IMSNU 20058
T,
were 28.91, 19.52, and 59.17%, respectively, which were
significantly lower than the threshold value (70%) for the
delineation of genomic species (Wayne et al., 1987). The
unique taxonomic position of strain KCTC 29142 determined
by a polyphasic taxonomic approach has prompted the present
suggestion of this strain as a new species of the genus
Amycolatopsis.
The genus Amycolatopsis was first described by Lechevalier
et al. (1986). It is a member of the family Pseudonocardiaceae,
in the phylum Actinobacteria and 65 species of the genus
Amycolatopsis have been recognized (Euzéby, 1997). Some
species of this genus were reported to produce epoxyquinomicin
C, vancomycin, rifomycin, rifamycin, and siderochelins (Sensi
et al., 1959; Matsumoto et al., 1997; Wink et al., 2003; Bala et
al., 2004; Lu et al., 2015).
Morphological, physiological, and biochemical cha-
racteristics
Strain KCTC29142 was Gram-positive, aerobic, catalase-
positive, and non-motile (data not shown). Aerial mycelia were
straight with a smooth surface. Morphology showed difference
when compared with the closely related strains on Bennett’s
medium: Aerial mycelia (front) and substrate mycelia (back)
of KCTC 29142 is white and pink, while those of A. orientalis
IMSNU 20058 are white and brownish yellow. Both A.
keratiniphila DSM 44586 and A. decaplanina DSM 44594
showed typical morphology of Amycolatopsis species, and the
colors are sand yellow (Fig. 2). Other culture characteristics of
strain KCTC 29142 on various media are summarized in
Supplementary data Table S1. Aerial mycelia were white on
most tested ISP media and vegetative mycelia were purple on
ISP3, purplish-pink on ISP4, violet on ISP7, pink on Bennett’s
medium, and beige or yellow on ISP2, ISP5, ISP6, and S
medium. Diffusible pink pigment was observed on Bennett’s
medium. The physiological and biochemical characteristics of
strain KCTC 29142 are compared with related type strains.
Strain KCTC 29142 can utilize glucose, arabinose, fructose,
mannitol, xylose and raffinose as sole carbon source, but not
cellulose, inositol, and sucrose. It can also degraded casein,
xanthine, gelatin, and tyrosine. Growth occurred at 10–37°C
(optimum, 28°C) and pH 5–10 (optimum, pH 7). The optimal
growth NaCl concentration was 0–2.5% (w/v, no growth on
5%), and was resistant to streptomycin (100 μg/ml), neomycin
(50 μg/ml), and vancomycin (50 μg/ml). These characteristics
were different with related type strains (Table 1), which
suggested it is an uncharacterized strain so far.
The phylum Actinobacteria, which includes the genus
Streptomyces, has proven particularly important as a source of
secondary metabolites include many antibiotics (Gao and Gupta,
2012). Previously, many studies have been reported on the
discovery of novel antibiotics from Actinobacteria which can
inhibit the MDRPs, but mostly are from Streptomyces species

Anti-MDRP activity of siderochelin from a novel Amycolatopsis sp. ∙ 331
Korean Journal of Microbiology, Vol. 52, No. 3
Table 1. Physiological and biochemical characteristics of strain KCTC
29142 and related Amycolatopsis species
Characteristics 1 2 3 4
Growth on sole carbon sources (1%, w/v)
D-Glucose + + + +
D-Arabinose + + + +
D-Fructose + + + +
D-Mannitol + + + +
D-Xylose + - + -
Raffinose + - - -
Cellulose - - + -
Inositol - + + -
Sucrose - - + -
Decomposition of
Casein + + + +
Xanthine + - - -
Gelatin +/- + - +/-
Tyrosine + + + +
Growth in
NaCl (2.5%) + + + +
NaCl (5.0%) - - - +
Growth at
4°C - - - -
10°C + + + +
28°C + + + +
37°C + + + +
45°C - - - -
Antibiotics resistant on
Tobramycin 50 μg/ml - - - +
Streptomycin 100 μg/ml + - - +
Neomycin 50 μg/ml + + - +
Vancomycin 50 μg/ml + + - +
Strains: 1, strain KCTC 29142; 2, Amycolatopsis orientalis IMSNU 20058T;
3, A. keratiniphila subsp. nogabecina DSM 44586T; 4, A. decaplanina
DSM 44594T. All data are from this study.
+, positive or present; -, negative or absent
Table 2. Chemical shifts 1H (500 MHz),
13C (125 MHz), and HMBC
correlations for anti-MDRP compound (in CDCl3)
Carbon No. δH δC HMBC
1 (CH) 8.16 dd (4.5, 1.5) 140.4 C2, C3, C5
2 (CH) 7.34 dd (8.5, 4.5) 126.1 C3, C4
3 (CH) 7.41 dd (8.5, 1.5) 128.0 C1, C4, C5
4 (C) 158.5
5 (C) 134.0
6 (C) 181.4
7 (C) 85.3
8 (CH2) 2.53 dd (13.5, 10.0) 40.9 C6, C7, C9, C10, C11
2.62 dd (13.5, 2.0)
9 (CH) 4.96 dd (10.0, 2.0) 70.6 C6, C10
10 (C) 173.2
11 (CH3) 1.69 s 28.6 C6, C7, C8
Fig. 3. Chemical structure of anti-MRSA compound isolated from strain
KCTC 29142.
(Procópio et al., 2012; Cheng et al., 2013). Recently, rare
actinomycetes were recognized as a potential storehouse for
novel antibiotics, and many studies were carried out to develop
novel antibiotics from rare actinomycetes (Tiwari and Gupta,
2012).
Chemotaxonomic analysis
Whole-cell lysate of strain KCTC 29142 was determined to
contain meso-diaminopimelic acid, arabinose, and galactose.
The predominant menaquinone was determined to be MK9(H4).
Phosphatidylethanolamine, phosphatidylglycerol, and phospha-
tidylinositol were the predominant phospholipids. The cellular
fatty acid compositions of strain KCTC 29142 and the three
reference strains are listed in Supplementary data Table S2. The
major cellular fatty acids of strain KCTC29142 were found to
be iso-C16:0 (19.5%), iso-C16:0 (16.4%), iso-C15:0 (8.5%), and C18:0
(7.8%). These results also suggested that the strain KCTC 29142
belong to the genus Amycolatopsis but have difference with
closely related strains.
Isolation and structure elucidation of anti-MRSA
compound produced by strain KCTC 29142
The supernatant from flask fermentation was extracted with
EtOAc and purified by several chromatographic purification

332 ∙ Lee et al.
미생물학회지 제52권 제3호
Table 3. Minimum inhibitory concentrations (MIC) of siderochelin A on clinical MRSA isolates
Strain Siderochelin A Norfloxacin Cephalothin Erythromycin Oxacillin Vancomycin
CCARM3040 64 4 128 >128 >128 2
CCARM3090 64 128 >128 >128 >64 4
CCARM3091 64 32 >128 >128 >64 4
CCARM3100 64 128 >128 >128 >128 1
CCARM3152 64 0.5 <0.25 16 8 0.5
CCARM3166 64 0.5 2 1 4 1
CCARM3197 64 1 0.5 0.5 0.5 0.5
CCARM3227 64 > 128 1 0.5 0.5 1
CCARM3231 64 8 16 0.5 16 1
CCARM3502 64 128 128 >128 >128 4
CCARM3568 64 > 128 128 >128 128 <1
CCARM3574 64 1 16 32 128 <1
CCARM3793 64 1 4 >128 4 2
CCARM, Culture Collection of Antimicrobial Resistant Microbes
procedures guided by anti-MRSA activity. The anti-MRSA
compound was finally obtained as a yellowish-colored crystal.
The molecular formula was determined as C11H13N3O3 according
to HR Q-TOF MS data (m/z 236.106 [M+H]+) and
1H- and
13C-NMR data (Supplementary data Fig. S1, Table 2). The
coincidence of 1H- and
13C-NMR spectroscopic properties of
the anti-MRSA compound with the data from previous reports
confirmed the compound as siderochelin A (Fig. 3). The
structure was verified by HMBC spectral data (Table 2).
Recently, 3 new siderochelins, siderochelin D, E, F were
isolated from Amycolatopsis sp. LZ149 (Lu et al., 2015). These
compounds, along with siderochelin A were reported to have
anti-mycobacterial activity. In our study, siderochelin A was
dominantly isolated from the culture broth.
Anti-MDRP activities of siderochelin A
Siderochelins, which chelate ferrous-ion, are produced from
Nocardia sp., Streptoalloteichus sp., and Amycolatopsis sp.
(Liu et al., 1981; Okuyama et al., 1982; Mitscher et al., 1984;
Lu et al., 2015). These reports also proposed that siderochelin
A has good antibacterial activity. However, the antibacterial
activity of siderochelin A on MDRPs had not been investigated
until the present study.
The MICs of siderochelin A on clinical isolates of MRSA are
summarized in Table 3. The clinical isolates were susceptible
only to vancomycin, and were resistant to either norfloxacin,
cephalothin, erythromycin, or oxacillin. However, siderochelin
A showed moderate inhibitory activity against all the clinical
isolates which is similar with previous reports (Liu et al., 1981;
Mitscher et al., 1984). The MIC for all these isolates was 64
μg/ml. This result implied that the inhibitory activity of
siderochelin A is not due to antibiotic mechanism, but may be
related with the high iron chelating activity which can limit the
growth of pathogens.
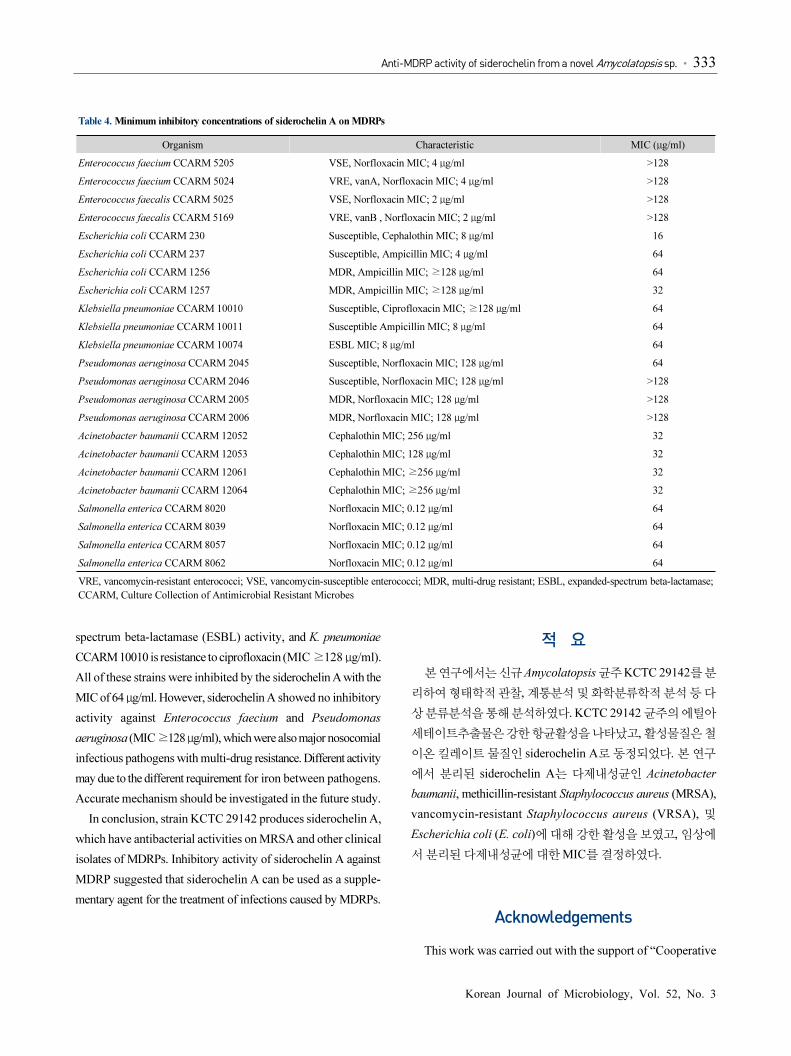
Inhibitory effects of siderochelin A on various human
infected MDRPs including Gram-positive and Gram-negative
bacteria were also investigated (Table 4). Siderochelin A
showed inhibitory activities against MDRPs, but differed in the
strains. Siderochelin A showed good activity to E. coli and
Acinetobacter baumannii. The MIC of siderochelin A to a
susceptible E. coli CCARM 230 and multi-drug resistant E. coli
CCARM 1257 are 16 and 32 μg/ml respectively. Siderochelin
A also showed good inhibitory activity against clinical isolates
of A. baumannii, which is an increasingly serious threat as a
nosocomial pathogen, and displays multi-drug resistance
(Kempf and Rolain, 2012). Even though the A. baumannii
isolates used in this study were resistant to cephalothin (MIC
128 to ≥256 μg/ml), they were susceptible to siderochelin A
(MIC 32 μg/ml). Siderochelin A showed moderate inhibitory
activity against Klebsiella pneumonia and Salmonella enterica
strains, the causative pathogen of pneumonia and salmonellosis
respectively. Klebsiella pneumoniae CCARM 10074 has extended-
Anti-MDRP activity of siderochelin from a novel Amycolatopsis sp. ∙ 333
Korean Journal of Microbiology, Vol. 52, No. 3
Table 4. Minimum inhibitory concentrations of siderochelin A on MDRPs
Organism Characteristic MIC (μg/ml)
Enterococcus faecium CCARM 5205 VSE, Norfloxacin MIC; 4 μg/ml >128
Enterococcus faecium CCARM 5024 VRE, vanA, Norfloxacin MIC; 4 μg/ml >128
Enterococcus faecalis CCARM 5025 VSE, Norfloxacin MIC; 2 μg/ml >128
Enterococcus faecalis CCARM 5169 VRE, vanB , Norfloxacin MIC; 2 μg/ml >128
Escherichia coli CCARM 230 Susceptible, Cephalothin MIC; 8 μg/ml 16
Escherichia coli CCARM 237 Susceptible, Ampicillin MIC; 4 μg/ml 64
Escherichia coli CCARM 1256 MDR, Ampicillin MIC; ≥128 μg/ml 64
Escherichia coli CCARM 1257 MDR, Ampicillin MIC; ≥128 μg/ml 32
Klebsiella pneumoniae CCARM 10010 Susceptible, Ciprofloxacin MIC; ≥128 μg/ml 64
Klebsiella pneumoniae CCARM 10011 Susceptible Ampicillin MIC; 8 μg/ml 64
Klebsiella pneumoniae CCARM 10074 ESBL MIC; 8 μg/ml 64
Pseudomonas aeruginosa CCARM 2045 Susceptible, Norfloxacin MIC; 128 μg/ml 64
Pseudomonas aeruginosa CCARM 2046 Susceptible, Norfloxacin MIC; 128 μg/ml >128
Pseudomonas aeruginosa CCARM 2005 MDR, Norfloxacin MIC; 128 μg/ml >128
Pseudomonas aeruginosa CCARM 2006 MDR, Norfloxacin MIC; 128 μg/ml >128
Acinetobacter baumanii CCARM 12052 Cephalothin MIC; 256 μg/ml 32
Acinetobacter baumanii CCARM 12053 Cephalothin MIC; 128 μg/ml 32
Acinetobacter baumanii CCARM 12061 Cephalothin MIC; ≥256 μg/ml 32
Acinetobacter baumanii CCARM 12064 Cephalothin MIC; ≥256 μg/ml 32
Salmonella enterica CCARM 8020 Norfloxacin MIC; 0.12 μg/ml 64
Salmonella enterica CCARM 8039 Norfloxacin MIC; 0.12 μg/ml 64
Salmonella enterica CCARM 8057 Norfloxacin MIC; 0.12 μg/ml 64
Salmonella enterica CCARM 8062 Norfloxacin MIC; 0.12 μg/ml 64
VRE, vancomycin-resistant enterococci; VSE, vancomycin-susceptible enterococci; MDR, multi-drug resistant; ESBL, expanded-spectrum beta-lactamase;
CCARM, Culture Collection of Antimicrobial Resistant Microbes
spectrum beta-lactamase (ESBL) activity, and K. pneumoniae
CCARM 10010 is resistance to ciprofloxacin (MIC ≥128 μg/ml).
All of these strains were inhibited by the siderochelin A with the
MIC of 64 μg/ml. However, siderochelin A showed no inhibitory
activity against Enterococcus faecium and Pseudomonas
aeruginosa (MIC ≥128 μg/ml), which were also major nosocomial
infectious pathogens with multi-drug resistance. Different activity
may due to the different requirement for iron between pathogens.
Accurate mechanism should be investigated in the future study.
In conclusion, strain KCTC 29142 produces siderochelin A,
which have antibacterial activities on MRSA and other clinical
isolates of MDRPs. Inhibitory activity of siderochelin A against
MDRP suggested that siderochelin A can be used as a supple-
mentary agent for the treatment of infections caused by MDRPs.
적 요
본 연구에서는 신규 Amycolatopsis 균주 KCTC 29142를 분
리하여 형태학적 관찰, 계통분석 및 화학분류학적 분석 등 다
상 분류분석을 통해 분석하였다. KCTC 29142 균주의 에틸아
세테이트추출물은 강한 항균활성을 나타났고, 활성물질은 철
이온 킬레이트 물질인 siderochelin A로 동정되었다. 본 연구
에서 분리된 siderochelin A는 다제내성균인 Acinetobacter
baumanii, methicillin-resistant Staphylococcus aureus (MRSA),
vancomycin-resistant Staphylococcus aureus (VRSA), 및
Escherichia coli (E. coli)에 대해 강한 활성을 보였고, 임상에
서 분리된 다제내성균에 대한 MIC를 결정하였다.
Acknowledgements
This work was carried out with the support of “Cooperative

334 ∙ Lee et al.
미생물학회지 제52권 제3호
Research Program for Agriculture Science & Technology
Development (Project No. PJ01128901)”, Rural Development
Administration, Republic of Korea. H.-J.H. was supported by
funding from the Royal Society, UK (51600.K5766/ROG) and
the Medical Research Council, UK (G0700141).
References
Anderson, D.I. 2003. Persistence of antibiotic resistant bacteria. Curr.
Opin. Microbiol. 6, 452–456.
Baker, D. 1990. Methods for the isolation, culture and characterization
of the Frankiaceae: soil actinomycetes and symbionts of acti-
norhizal plants. In Labeda, D.P. (eds), Isolation of Biotech-
nological Organisms from Nature, pp. 213–236. McGraw-Hill
Publishing Co., New York, USA.
Bala, S., Khanna, R., Dadhwal, M., Prabagaran, S.R., Shivaji, S.,
Cullum, J., and Lal, R. 2004. Reclassification of Amycolatopsis
mediterranei DSM 46095 as Amycolatopsis rifamycinica sp.
nov. Int. J. Syst. Evol. Microbiol. 54, 1145–1149.
Becker, B., Lechevalier, M.P., and Lechevalier, H.A. 1965. Chemical
composition of cell-wall preparations from strains of various
form-genera of aerobic actinomycetes. Appl. Microbiol. 13, 236
–243.
Chan, G.C., Chan, S., Ho, P.L., and Ha, S.Y. 2009. Effects of chelators
(deferoxamine, deferiprone and deferasirox) on the growth of
Klebsiella pneumoniae and Aeromonas hydrophila isolated
from transfusion-dependent thalassemia patients. Hemoglobin
33, 352–360.
Cheng, J., Jin, Y.Y., Yang, S.H., and Suh, J.W. 2013. Isolation and
characterization of anti-methicillin-resistant Staphylococcus
aureus/vancomycin-resistant Enterococcus compound from
Streptomyces bungoensis MJM2077. J. Korean Soc. Appl. Biol.
Chem. 56, 107–111.
Cosgrove, S.E. 2006. The relationship between antimicrobial resistance
and patient outcomes: mortality, length of hospital stay, and
health care costs. Clin. Infect. Dis. 42, 82–89.
Euzéby, J.P. 1997. List of bacterial names with standing in nomenclature:
a folder available on the internet. Int. J. Syst. Evol. Microbiol. 47,
590–592.
Gao, B. and Gupta, R.S. 2012. Phylogenetic framework and molecular
signatures for the main clades of the phylum Actinobacteria.
Microbiol. Mol. Biol. Rev. 76, 66–112.
Gordon, R.E., Barnett, D.A., Handerhan, J.E., and Pang, C.H.N. 1974.
Nocardia coeliaca, Nocardia autotrophica, and the Nocardin
strain. Int. J. Syst. Bacteriol. 24, 54–63.
Kempf, M. and Rolain, J.M. 2012. Emergence of resistance to
carbapenems in Acinetobacter baumannii in Europe, clinical
impact and therapeutic options. Int. J. Antimicrob. Agents 39,
105–114.
Kim, O.S., Cho, Y.J., Lee, K., Yoon, S.H., Kim, M., Na, H., Park, S.C.,
Jeon, Y.S., Lee, J.H., Yi, H., et al. 2012. Introducing EzTaxon-e,
a prokaryotic 16S rRNA gene sequence database with phylotypes
that represent uncultured species. Int. J. Syst. Evol. Microbiol.
62, 716–721.
Kim, C.M. and Shin, S.H. 2009. Effect of iron-chelator deferiprone on
the in vitro growth of staphylococci. J. Korean Med. Sci. 24, 289
–295.
Lane, D.J. 1991. 16S/23S rRNA sequencing. In Stackebrandt, E. and
Goodfellow, M. (eds.) Nucleic Acid Techniques in Bacterial
Systematics, pp. 115–176. Wiley, Chichester, UK.
Lechevalier, M.P., Prauser, H., Labeda, D.P., and Ruan, J.S. 1986.
Two new genera of nocardioform actinomycetes, Amycolata
gen. nov. and Amycolatopsis gen. nov. Int. J. Syst. Bacteriol. 36,
29–37.
Lin, M.F. and Lan, C.Y. 2014. Antimicrobial resistance in Acinetobacter
baumannii, from bench to bedside. World J. Clin. Cases 2, 787–
814.
Liu, W.C., Fisher, S.M., Wells, J.S. Jr., Ricca, C.S., Principe, P.A.,
Trejo, W.H., Bonner, D.P., Gougoutos, J.Z., Toeplitz, B.K., and
Sykes, R.B. 1981. Siderochelin, a new ferrous-ion chelating
agent produced by Norcardia. J. Antibiot. (Tokyo) 34, 791–799.
Lode, H.M. 2009. Clinical impact of antibiotic-resistant Gram-positive
pathogens. Clin. Microbiol. Infect. 15, 212–217.
Lu, C.H., Ye, F.W., and Shen, Y.M. 2015. Siderochelins with
anti-mycobacterial activity from Amycolatopsis sp. LZ149.
Chin. J. Nat. Med. 13, 69–72.
Matsumoto, N., Iinuma, H., Sawa, T., Takeuchi, T., Hirano, S.,
Yoshioka, T., and Ishizuka, M. 1997. Epoxyquinomicins A, B, C
and D, new antibiotics from Amycolatopsis. II. Effect on type II
collagen-induced arthritis in mice. J. Antibiot. (Tokyo) 50, 906–
911.
Miethke, M. and Marahiel, M.A. 2007. Siderophore-based iron
acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 71,
413–451.
Mitscher, L.A., Högberg, T., Drake, S.D., Burgstahler, A.W., Jackson,
M., Lee, B., Sheldon, R.I., Gracey, H.E., Kohl, W., and
Theriault, R.J. 1984. Isolation and structural determination of
siderochelin C, a fermentation product of an unusual Actinomycetes
sp. J. Antibiot. (Tokyo) 37, 1260–1263.
Okuyama, D., Nakamura, H., Naganawa, H., Takita, T., Umezawa, H.,
and Iitaka, Y. 1982. Isolation, racemization and absolute
configuration of siderochelin. J. Antibiot. (Tokyo) 35, 1240–
1242.
Procópio, R.E., Silva, I.R., Martins, M.K., Azevedo, J.L., and Araújo,
J.M. 2012. Antibiotics produced by Streptomyces. Braz. J.
Infect. Dis. 16, 466–471.
Saitou, N. and Nei, M. 1987. The neighbour-joining method, a new
method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4,
406–425.
Sakoulas, G. and Moellering, R.C. Jr. 2008. Increasing antibiotic
resistance among methicillin-resistant Staphylococcus aureus

Anti-MDRP activity of siderochelin from a novel Amycolatopsis sp. ∙ 335
Korean Journal of Microbiology, Vol. 52, No. 3
strains. Clin. Infect. Dis. 46, 360–367.
Sensi, P., Greco, A.M., and Ballotta, R. 1959. Rifomycin I. Isolation
and properties of rifomycin B and rifomycin complex. Antibiot.
Annu. 7, 262–270.
Shiring, E.B. and Gottlieb, D. 1966. Methods for characterization of
Streptomyces species. Int. J. Syst. Bacteriol. 16, 313–340.
Tajima, F. and Nei, M. 1984. Estimation of evolutionary distance
between nucleotide sequences. Mol. Biol. Evol. 1, 269–285.
Tamura, K., Dudley, J., Nei, M., and Kumar, S. 2007. MEGA4,
molecular evolutionary genetics analysis (MEGA) software
version 4.0. Mol. Biol. Evol. 24, 1596–1599.
Thompson, M.G., Corey, B.W., Si, Y., Craft, D.W., and Zurawski,
D.V. 2012. Antimicrobial activities of iron chelators against
common nosocomial pathogens. Antimicrob. Agents Chemother.
56, 5419–5421.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., and
Higgins, D.G. 1997. The CLUSTAL_X windows interface,
flexible strategies for multiple sequence alignment aided by
quality analysis tools. Nucleic Acids Res. 25, 4876–4882.
Tiwari, K. and, Gupta, R.K. 2012. Rare actinomycetes, a potential
storehouse for novel antibiotics. Crit. Rev. Biotechnol. 32, 108–
132.
Tseng, M., Yang, S.F., Li, W.J., and Jiang, C.L. 2006. Amycolatopsis
taiwanensis sp. nov., from soil. Int. J. Syst. Evol. Microbiol. 56,
1811–1815.
Waksman, S.A. and Henrici, A.T. 1943. The nomenclature and
classification of the Actinomycetes. J. Bacteriol. 46, 337–341.
Wayne, L.G., Brenner, D.J., Colwell, R.R., and Truper, H.G. 1987.
Report of the Ad hoc committee on reconciliation of approaches
to bacterial systematics. Int. J. Syst. Bacteriol. 37, 463–464.
Wink, J.M., Kroppenstedt, R.M., Ganguli, B.N., Nadkarni, S.R.,
Schumann, P., Seibert, G., and Stackebrandt, E. 2003. Three
new antibiotic producing species of the genus Amycolatopsis,
Amycolatopsis balhimycina sp. nov., A. tolypomycina sp. nov.,
A. vancoresmycina sp. nov., and description of Amycolatopsis
keratiniphila subsp. Keratiniphila subsp. nov. and A. keratiniphila
subsp. Nogabecina subsp. nov. Syst. Appl. Microbiol. 26, 38–46.


















