
Anti-CD47 antibody–mediated phagocytosis of cancer.pdf
6
Anti-CD47 antibody–mediated phagocytosis of cancer by macrophages primes an effective antitumor T-cell response Diane Tseng a,b,1 , Jens-Peter Volkmer a,b , Stephen B. Willingham a,b , Humberto Contreras-Trujillo a,b , John W. Fathman a,b , Nathaniel B. Fernhoff a,b , Jun Seita a,b , Matthew A. Inlay a,b , Kipp Weiskopf a,b , Masanori Miyanishi a,b , and Irving L. Weissman a,b,1 a Institute for Stem Cell Biology and Regenerative Medicine and b the Ludwig Cancer Center, Stanford University Medical Center, Stanford, CA 94305 Contributed by Irving L. Weissman, April 4, 2013 (sent for review February 12, 2013) Mobilization of the T-cell response against cancer has the potential to achieve long-lasting cures. However, it is not known how to harness antigen-presenting cells optimally to achieve an effective antitumor T-cell response. In this study, we show that anti-CD47 antibody–mediated phagocytosis of cancer by macrophages can ini- tiate an antitumor T-cell immune response. Using the ovalbumin model antigen system, anti-CD47 antibody–mediated phagocytosis of cancer cells by macrophages resulted in increased priming of OT-I T cells [cluster of differentiation 8-positive (CD8 + )] but decreased priming of OT-II T cells (CD4 + ). The CD4 + T-cell response was charac- terized by a reduction in forkhead box P3-positive (Foxp3 + ) regulatory T cells. Macrophages following anti-CD47–mediated phagocytosis primed CD8 + T cells to exhibit cytotoxic function in vivo. This re- sponse protected animals from tumor challenge. We conclude that anti-CD47 antibody treatment not only enables macrophage phagocytosis of cancer but also can initiate an antitumor cytotoxic T-cell immune response. A ntigen presentation is the process by which innate immune cells such as macrophages and dendritic cells (antigen-pre- senting cells, APC) acquire antigens and present them to T cells to initiate the adaptive immune response. How APCs shape the im- mune response by both degrading antigens and preserving antigens for presentation to T cells has been a longstanding area of interest (1). Recently, the mechanism of antigen recognition by APCs has been shown to affect the preference of MHC I versus MHC II antigen-presentation pathways. For instance, mannose receptor- mediated endocytosis on dendritic cells has been associated with MHC I antigen presentation, whereas scavenger receptor-mediated endocytosis has been associated with MHC II presentation (2). Moreover, the functional outcomes of antigen presentation have been shown to be context dependent. For instance, targeting anti- gens to DEC-205 using monoclonal antibodies induced tolerance under noninflammatory conditions but mediated immunogenicity under activating conditions by cluster of differentiation 40 ligand (CD40L) (3). Harnessing APCs to enhance the antitumor T-cell response offers an exciting strategy for cancer immunotherapy. The ability of the T-cell immune response to be mobilized successfully against cancer has been demonstrated through preclinical and clinical studies of anti-CTLA4 antibody for T-cell activation (4). Phagocytosis by macrophages relies on the cell’s recognition of prophagocytic (“eat me”) and antiphagocytic (“don’t eat me”) sig- nals on target cells. Anti-CD47 blocking monoclonal antibodies (mAbs) induce macrophage phagocytosis of cancer cells by inhibiting an important antiphagocytic signal, allowing propha- gocytic signals to dominate (5, 6). CD47 is highly expressed on cancer cells as compared with normal cells (5, 6) and interacts with the ligand signal regulatory protein α (SIRP-α) on macro- phages (7). This interaction results in phosphorylation of immu- noreceptor tyrosine-based inhibition (ITIM) motifs on SIRP-α’s cytoplasmic tail and the recruitment of Src homology phosphatase- 1 (SHP-1) and SHP-2 phosphatases, which is thought to block phagocytosis by preventing myosin-IIA accumulation at the phago- cytic synapse (8–12). We have demonstrated the therapeutic efficacy of anti-CD47 blocking mAbs against xenograft human cancers growing in immunodeficient mice, including cancers such as leukemia (5, 13), lymphoma (14), and multiple myeloma (15), solid tumors, including breast, colon, prostate, and bladder cancers, and sarcomas (6, 16). Whether the adaptive immune response also can be recruited against the cancer after anti-CD47 mAb treatment has not been tested, because the immunodeficient mice used to establish the xenograft models lack T, B, and NK cells. In this study, we tested the hypothesis that anti-CD47 antibody–mediated phagocytosis of cancer cells can facilitate an antitumor T-cell immune response. Results Macrophages Phagocytose Cancer Cells in the Presence of Anti-CD47 Blocking Antibody. To follow the immune response to a model tu- mor antigen, the human colon cancer cell line DLD1 was trans- fected with a lentiviral vector for expressing cytoplasmic ovalbumin (cOVA) and GFP (DLD1-cOVA-GFP) (Fig. S1). DLD1-cOVA- GFP cancer cells express CD47 and can be recognized by both CD47 mAbs, clones B6H12 and 2D3 (Fig. S1). Anti-CD47 B6H12 (blocking) mAb blocks the interaction between CD47 and SIRP-α, whereas anti-CD47 2D3 (non-blocking) antibody binds CD47 but does not block its interaction with SIRP-α. Macrophages phagocytose DLD1-cOVA-GFP cancer cells in the presence of anti-CD47 B6H12, but not anti-CD47 2D3 mAbs, demonstrat- ing that phagocytosis is dependent on the blockade of CD47/ SIRPα interactions and not entirely due to antibody opsoniza- tion effects (Fig. 1 and Fig. S2). Anti-CD47 mediated phago- cytosis of DLD1-cOVA-GFP cancer cells by macrophages leads to cross-presentation of ovalbumin peptide onto MHC-I, as assessed by staining for the SIINFEKL-H2kb complex on the cell surface (Fig. S3). Costimulatory molecule CD86 is up- regulated, but not coinhibitory molecule B7-H1 (Fig. S4). Anti- CD47 B6H12–mediated phagocytosis of cancer cells leads to macrophage release of proinflammatory cytokines. For example, IL-12p40, TNF-α, regulated upon activation normal T cell ex- pressed and secreted (RANTES), and monocyte chemotactic protein-3 (MCP-3) cytokine levels increase after anti-CD47 B6H12-mediated phagocytosis (Fig. S5). Next, the ability of the APCs, macrophages and dendritic cells, were tested for phagocytic activity in response to anti-CD47 mAbs. Com- pared to dendritic cells, macrophages effectively phagocytose DLD1-cOVA-GFP cancer cells in the presence of anti-CD47 Author contributions: D.T., J.-P.V., S.B.W., and I.L.W. designed research; D.T., J.-P.V., S.B.W., H.C.-T., J.W.F., and N.B.F. performed research; D.T., J.-P.V., and K.W. contributed new reagents/analytic tools; D.T., J.-P.V., S.B.W., J.W.F., N.B.F., J.S., M.A.I., M.M., and I.L.W. analyzed data; and D.T., J.-P.V., and I.L.W. wrote the paper. Conflict of interest statement: I.L.W. owns Amgen Inc. stock and is a Director of Stem Cells, Inc. Freely available online through the PNAS open access option. See Commentary on page 10886. 1 To whom correspondence may be addressed. E-mail: [email protected] or [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1305569110/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1305569110 PNAS | July 2, 2013 | vol. 110 | no. 27 | 11103–11108 MEDICAL SCIENCES SEE COMMENTARY
-
Upload
rui-freitas -
Category
Documents
-
view
25 -
download
5
Transcript of Anti-CD47 antibody–mediated phagocytosis of cancer.pdf

Anti-CD47 antibody–mediated phagocytosis of cancerby macrophages primes an effective antitumorT-cell responseDiane Tsenga,b,1, Jens-Peter Volkmera,b, Stephen B. Willinghama,b, Humberto Contreras-Trujilloa,b,John W. Fathmana,b, Nathaniel B. Fernhoffa,b, Jun Seitaa,b, Matthew A. Inlaya,b, Kipp Weiskopfa,b,Masanori Miyanishia,b, and Irving L. Weissmana,b,1
aInstitute for Stem Cell Biology and Regenerative Medicine and bthe Ludwig Cancer Center, Stanford University Medical Center, Stanford, CA 94305
Contributed by Irving L. Weissman, April 4, 2013 (sent for review February 12, 2013)
Mobilization of the T-cell response against cancer has the potentialto achieve long-lasting cures. However, it is not known how toharness antigen-presenting cells optimally to achieve an effectiveantitumor T-cell response. In this study, we show that anti-CD47antibody–mediated phagocytosis of cancer bymacrophages can ini-tiate an antitumor T-cell immune response. Using the ovalbuminmodel antigen system, anti-CD47 antibody–mediated phagocytosisof cancer cells bymacrophages resulted in increased priming of OT-IT cells [cluster of differentiation 8-positive (CD8+)] but decreasedpriming of OT-II T cells (CD4+). The CD4+ T-cell response was charac-terizedbya reduction in forkheadboxP3-positive (Foxp3+) regulatoryT cells. Macrophages following anti-CD47–mediated phagocytosisprimed CD8+ T cells to exhibit cytotoxic function in vivo. This re-sponse protected animals from tumor challenge. We conclude thatanti-CD47 antibody treatment not only enables macrophagephagocytosis of cancer but also can initiate an antitumor cytotoxicT-cell immune response.
Antigen presentation is the process by which innate immunecells such as macrophages and dendritic cells (antigen-pre-
senting cells, APC) acquire antigens and present them to T cells toinitiate the adaptive immune response. How APCs shape the im-mune response by both degrading antigens and preserving antigensfor presentation to T cells has been a longstanding area of interest(1). Recently, the mechanism of antigen recognition by APCs hasbeen shown to affect the preference of MHC I versus MHC IIantigen-presentation pathways. For instance, mannose receptor-mediated endocytosis on dendritic cells has been associated withMHC I antigen presentation, whereas scavenger receptor-mediatedendocytosis has been associated with MHC II presentation (2).Moreover, the functional outcomes of antigen presentation havebeen shown to be context dependent. For instance, targeting anti-gens to DEC-205 using monoclonal antibodies induced toleranceunder noninflammatory conditions but mediated immunogenicityunder activating conditions by cluster of differentiation 40 ligand(CD40L) (3). Harnessing APCs to enhance the antitumor T-cellresponse offers an exciting strategy for cancer immunotherapy. Theability of the T-cell immune response to be mobilized successfullyagainst cancer has been demonstrated through preclinical andclinical studies of anti-CTLA4 antibody for T-cell activation (4).Phagocytosis by macrophages relies on the cell’s recognition of
prophagocytic (“eat me”) and antiphagocytic (“don’t eat me”) sig-nals on target cells. Anti-CD47 blocking monoclonal antibodies(mAbs) induce macrophage phagocytosis of cancer cells byinhibiting an important antiphagocytic signal, allowing propha-gocytic signals to dominate (5, 6). CD47 is highly expressed oncancer cells as compared with normal cells (5, 6) and interactswith the ligand signal regulatory protein α (SIRP-α) on macro-phages (7). This interaction results in phosphorylation of immu-noreceptor tyrosine-based inhibition (ITIM) motifs on SIRP-α’scytoplasmic tail and the recruitment of Src homology phosphatase-1 (SHP-1) and SHP-2 phosphatases, which is thought to blockphagocytosis by preventing myosin-IIA accumulation at the phago-cytic synapse (8–12). We have demonstrated the therapeutic
efficacy of anti-CD47 blocking mAbs against xenograft humancancers growing in immunodeficient mice, including cancers such asleukemia (5, 13), lymphoma (14), and multiple myeloma (15), solidtumors, including breast, colon, prostate, and bladder cancers, andsarcomas (6, 16). Whether the adaptive immune response also canbe recruited against the cancer after anti-CD47 mAb treatment hasnot been tested, because the immunodeficientmice used to establishthe xenograft models lack T, B, andNK cells. In this study, we testedthe hypothesis that anti-CD47 antibody–mediated phagocytosisof cancer cells can facilitate an antitumor T-cell immune response.
ResultsMacrophages Phagocytose Cancer Cells in the Presence of Anti-CD47Blocking Antibody. To follow the immune response to a model tu-mor antigen, the human colon cancer cell line DLD1 was trans-fected with a lentiviral vector for expressing cytoplasmic ovalbumin(cOVA) and GFP (DLD1-cOVA-GFP) (Fig. S1). DLD1-cOVA-GFP cancer cells express CD47 and can be recognized by bothCD47 mAbs, clones B6H12 and 2D3 (Fig. S1). Anti-CD47 B6H12(blocking) mAb blocks the interaction between CD47 and SIRP-α,whereas anti-CD47 2D3 (non-blocking) antibody binds CD47but does not block its interaction with SIRP-α. Macrophagesphagocytose DLD1-cOVA-GFP cancer cells in the presence ofanti-CD47 B6H12, but not anti-CD47 2D3 mAbs, demonstrat-ing that phagocytosis is dependent on the blockade of CD47/SIRPα interactions and not entirely due to antibody opsoniza-tion effects (Fig. 1 and Fig. S2). Anti-CD47 mediated phago-cytosis of DLD1-cOVA-GFP cancer cells by macrophages leadsto cross-presentation of ovalbumin peptide onto MHC-I, asassessed by staining for the SIINFEKL-H2kb complex on thecell surface (Fig. S3). Costimulatory molecule CD86 is up-regulated, but not coinhibitory molecule B7-H1 (Fig. S4). Anti-CD47 B6H12–mediated phagocytosis of cancer cells leads tomacrophage release of proinflammatory cytokines. For example,IL-12p40, TNF-α, regulated upon activation normal T cell ex-pressed and secreted (RANTES), and monocyte chemotacticprotein-3 (MCP-3) cytokine levels increase after anti-CD47B6H12-mediated phagocytosis (Fig. S5). Next, the ability ofthe APCs, macrophages and dendritic cells, were tested forphagocytic activity in response to anti-CD47 mAbs. Com-pared to dendritic cells, macrophages effectively phagocytoseDLD1-cOVA-GFP cancer cells in the presence of anti-CD47
Author contributions: D.T., J.-P.V., S.B.W., and I.L.W. designed research; D.T., J.-P.V., S.B.W.,H.C.-T., J.W.F., and N.B.F. performed research; D.T., J.-P.V., and K.W. contributed newreagents/analytic tools; D.T., J.-P.V., S.B.W., J.W.F., N.B.F., J.S., M.A.I., M.M., and I.L.W.analyzed data; and D.T., J.-P.V., and I.L.W. wrote the paper.
Conflict of interest statement: I.L.W. owns Amgen Inc. stock and is a Director of StemCells, Inc.
Freely available online through the PNAS open access option.
See Commentary on page 10886.1To whom correspondence may be addressed. E-mail: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1305569110/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1305569110 PNAS | July 2, 2013 | vol. 110 | no. 27 | 11103–11108
MED
ICALSC
IENCE
SSE
ECO
MMEN
TARY
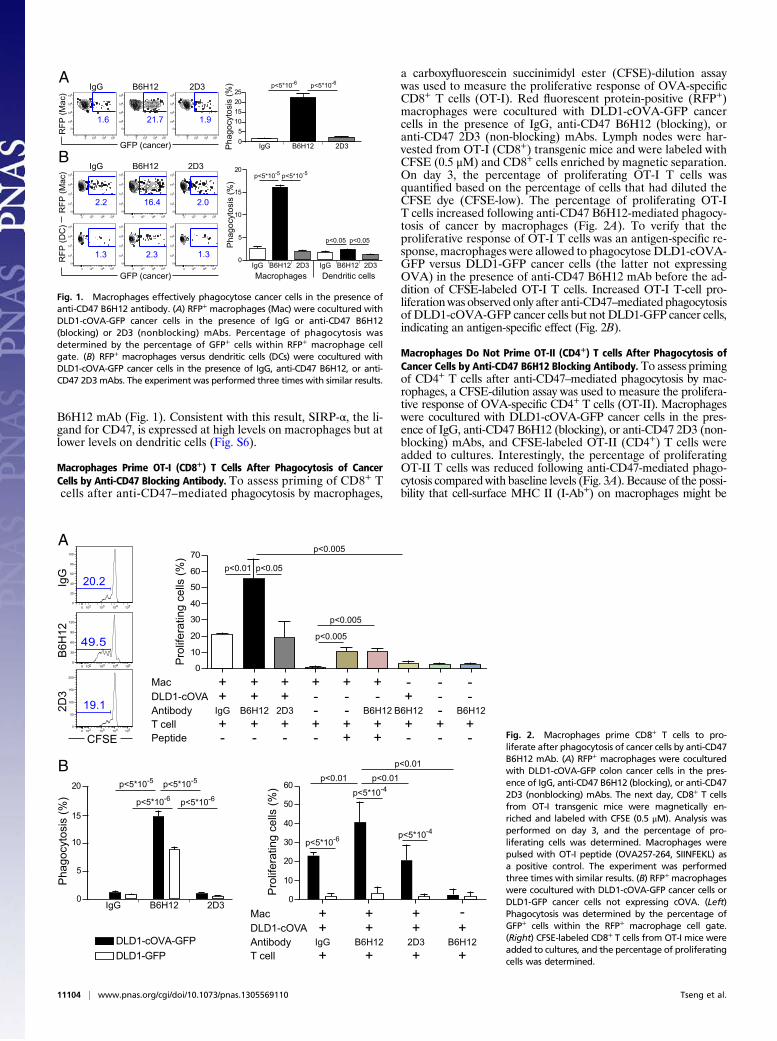
B6H12 mAb (Fig. 1). Consistent with this result, SIRP-α, the li-gand for CD47, is expressed at high levels on macrophages but atlower levels on dendritic cells (Fig. S6).
Macrophages Prime OT-I (CD8+) T Cells After Phagocytosis of CancerCells by Anti-CD47 Blocking Antibody. To assess priming of CD8+ Tcells after anti-CD47–mediated phagocytosis by macrophages,
a carboxyfluorescein succinimidyl ester (CFSE)-dilution assaywas used to measure the proliferative response of OVA-specificCD8+ T cells (OT-I). Red fluorescent protein-positive (RFP+)macrophages were cocultured with DLD1-cOVA-GFP cancercells in the presence of IgG, anti-CD47 B6H12 (blocking), oranti-CD47 2D3 (non-blocking) mAbs. Lymph nodes were har-vested from OT-I (CD8+) transgenic mice and were labeled withCFSE (0.5 μM) and CD8+ cells enriched by magnetic separation.On day 3, the percentage of proliferating OT-I T cells wasquantified based on the percentage of cells that had diluted theCFSE dye (CFSE-low). The percentage of proliferating OT-IT cells increased following anti-CD47 B6H12-mediated phagocy-tosis of cancer by macrophages (Fig. 2A). To verify that theproliferative response of OT-I T cells was an antigen-specific re-sponse, macrophages were allowed to phagocytose DLD1-cOVA-GFP versus DLD1-GFP cancer cells (the latter not expressingOVA) in the presence of anti-CD47 B6H12 mAb before the ad-dition of CFSE-labeled OT-I T cells. Increased OT-I T-cell pro-liferation was observed only after anti-CD47–mediated phagocytosisof DLD1-cOVA-GFP cancer cells but not DLD1-GFP cancer cells,indicating an antigen-specific effect (Fig. 2B).
Macrophages Do Not Prime OT-II (CD4+) T cells After Phagocytosis ofCancer Cells by Anti-CD47 B6H12 Blocking Antibody. To assess primingof CD4+ T cells after anti-CD47–mediated phagocytosis by mac-rophages, a CFSE-dilution assay was used to measure the prolifera-tive response of OVA-specific CD4+ T cells (OT-II). Macrophageswere cocultured with DLD1-cOVA-GFP cancer cells in the pres-ence of IgG, anti-CD47 B6H12 (blocking), or anti-CD47 2D3 (non-blocking) mAbs, and CFSE-labeled OT-II (CD4+) T cells wereadded to cultures. Interestingly, the percentage of proliferatingOT-II T cells was reduced following anti-CD47-mediated phago-cytosis compared with baseline levels (Fig. 3A). Because of the possi-bility that cell-surface MHC II (I-Ab+) on macrophages might be
IgG
RFP
(DC
)R
FP (M
ac)
RFP
(Mac
)
GFP (cancer)
A
B0 103 104 105
0
102
103
104
105
1.6
0 103 104 105
0
102
103
104
105
21.7
0 103 104 105
0
102
103
104
105
1.9
0 103 104 105
0
102
103
104
105
2.20 103 104 105
0
102
103
104
105
16.40 103 104 105
0
102
103
104
105
2.0
0 103 104 105
0
102
103
104
105
1.30 103 104 105
0
102
103
104
105
2.30 103 104 105
0
102
103
104
105
1.3
B6H12 2D3
IgG B6H12 2D3
Macrophages Dendritic cells
GFP (cancer)
0
5
10
15
20
Pha
gocy
tosi
s (%
)
IgG B6H12 2D3 IgG B6H12 2D3
p<0.05 p<0.05
p<5*10-5 p<5*10-5
Pha
gocy
tosi
s (%
)
IgG B6H12 2D305
10152025
p<5*10-6 p<5*10-6
Fig. 1. Macrophages effectively phagocytose cancer cells in the presence ofanti-CD47 B6H12 antibody. (A) RFP+ macrophages (Mac) were cocultured withDLD1-cOVA-GFP cancer cells in the presence of IgG or anti-CD47 B6H12(blocking) or 2D3 (nonblocking) mAbs. Percentage of phagocytosis wasdetermined by the percentage of GFP+ cells within RFP+ macrophage cellgate. (B) RFP+ macrophages versus dendritic cells (DCs) were cocultured withDLD1-cOVA-GFP cancer cells in the presence of IgG, anti-CD47 B6H12, or anti-CD47 2D3 mAbs. The experiment was performed three times with similar results.
0 102 103 104 1050
20
40
60
80
100
20.2
0 102 103 104 1050
30
60
90
120
49.5
0 102 103 104 1050
50
100
150
200
19.1
B6H
122D
3Ig
G
CFSE
0
10
20
30
40
50
60
70
B6H12
Pro
lifer
atin
g ce
lls (%
)
B6H12 B6H12DLD1-cOVAAntibodyT cellPeptide
Mac
2D3 B6H12IgG -
+ + + + + + - - -+ + + - - - + - -
+ + + + + + + + +- -
- - - - + + - - -
p<0.005
p<0.005
p<0.01 p<0.05
p<0.005
DLD1-cOVA-GFPDLD1-GFP
Pha
gocy
tosi
s (%
)
0
5
10
15
20
IgG B6H12 2D3
p<5*10-5 p<5*10-5
p<5*10-6 p<5*10-6
A
B
0
10
20
30
40
50
60
p<5*10-6
p<5*10-4
p<5*10-4
Pro
lifer
atin
g ce
lls (%
)
B6H12DLD1-cOVAAntibodyT cell
Mac
2D3IgG B6H12
+ + + -+ + + +
+ + + +
p<0.01 p<0.01p<0.01
Fig. 2. Macrophages prime CD8+ T cells to pro-liferate after phagocytosis of cancer cells by anti-CD47B6H12 mAb. (A) RFP+ macrophages were coculturedwith DLD1-cOVA-GFP colon cancer cells in the pres-ence of IgG, anti-CD47 B6H12 (blocking), or anti-CD472D3 (nonblocking) mAbs. The next day, CD8+ T cellsfrom OT-I transgenic mice were magnetically en-riched and labeled with CFSE (0.5 μM). Analysis wasperformed on day 3, and the percentage of pro-liferating cells was determined. Macrophages werepulsed with OT-I peptide (OVA257-264, SIINFEKL) asa positive control. The experiment was performedthree times with similar results. (B) RFP+ macrophageswere cocultured with DLD1-cOVA-GFP cancer cells orDLD1-GFP cancer cells not expressing cOVA. (Left)Phagocytosis was determined by the percentage ofGFP+ cells within the RFP+ macrophage cell gate.(Right) CFSE-labeled CD8+ T cells from OT-I mice wereadded to cultures, and the percentage of proliferatingcells was determined.
11104 | www.pnas.org/cgi/doi/10.1073/pnas.1305569110 Tseng et al.
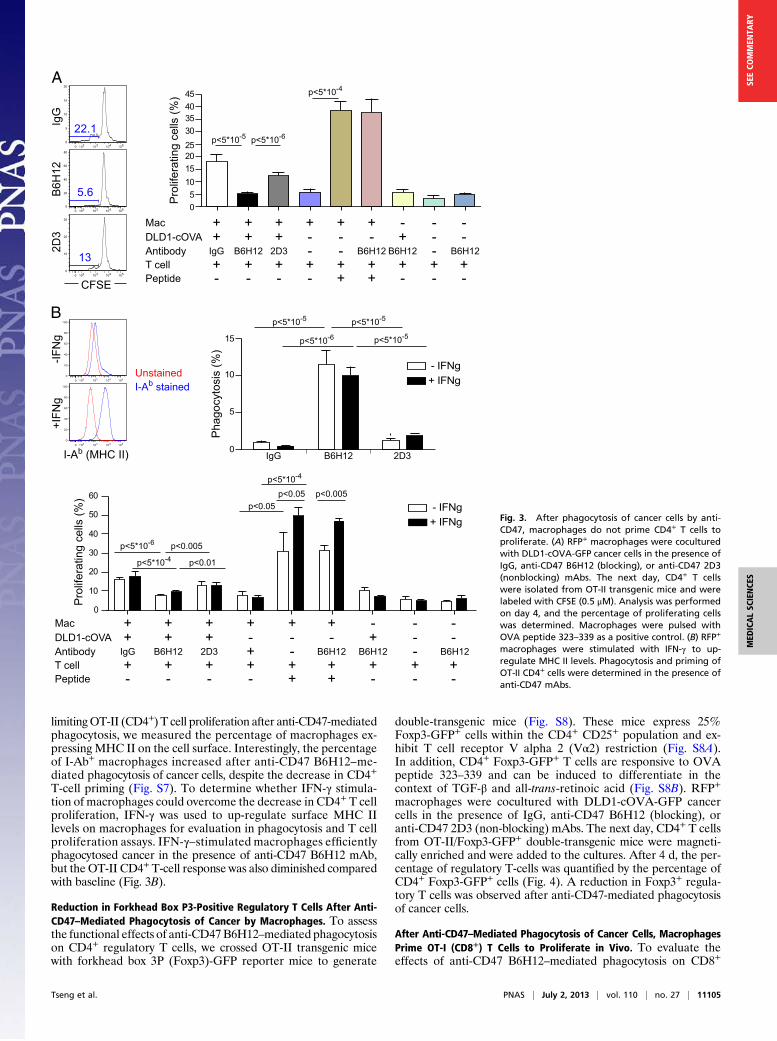
limitingOT-II (CD4+) T cell proliferation after anti-CD47-mediatedphagocytosis, we measured the percentage of macrophages ex-pressing MHC II on the cell surface. Interestingly, the percentageof I-Ab+ macrophages increased after anti-CD47 B6H12–me-diated phagocytosis of cancer cells, despite the decrease in CD4+
T-cell priming (Fig. S7). To determine whether IFN-γ stimula-tion of macrophages could overcome the decrease in CD4+ T cellproliferation, IFN-γ was used to up-regulate surface MHC IIlevels on macrophages for evaluation in phagocytosis and T cellproliferation assays. IFN-γ–stimulated macrophages efficientlyphagocytosed cancer in the presence of anti-CD47 B6H12 mAb,but theOT-II CD4+T-cell response was also diminished comparedwith baseline (Fig. 3B).
Reduction in Forkhead Box P3-Positive Regulatory T Cells After Anti-CD47–Mediated Phagocytosis of Cancer by Macrophages. To assessthe functional effects of anti-CD47B6H12–mediated phagocytosison CD4+ regulatory T cells, we crossed OT-II transgenic micewith forkhead box 3P (Foxp3)-GFP reporter mice to generate
double-transgenic mice (Fig. S8). These mice express 25%Foxp3-GFP+ cells within the CD4+ CD25+ population and ex-hibit T cell receptor V alpha 2 (Vα2) restriction (Fig. S8A).In addition, CD4+ Foxp3-GFP+ T cells are responsive to OVApeptide 323–339 and can be induced to differentiate in thecontext of TGF-β and all-trans-retinoic acid (Fig. S8B). RFP+
macrophages were cocultured with DLD1-cOVA-GFP cancercells in the presence of IgG, anti-CD47 B6H12 (blocking), oranti-CD47 2D3 (non-blocking) mAbs. The next day, CD4+ T cellsfrom OT-II/Foxp3-GFP+ double-transgenic mice were magneti-cally enriched and were added to the cultures. After 4 d, the per-centage of regulatory T-cells was quantified by the percentage ofCD4+ Foxp3-GFP+ cells (Fig. 4). A reduction in Foxp3+ regula-tory T cells was observed after anti-CD47-mediated phagocytosisof cancer cells.
After Anti-CD47–Mediated Phagocytosis of Cancer Cells, MacrophagesPrime OT-I (CD8+) T Cells to Proliferate in Vivo. To evaluate theeffects of anti-CD47 B6H12–mediated phagocytosis on CD8+
A
B
B6H
122D
3Ig
G
CFSE
0
5
10
15
20
22.1
0 102 103 104 1050
20
40
60
80
5.6
0 102 103 104 1050
10
20
30
13 B6H12
Pro
lifer
atin
g ce
lls (%
)
B6H12 B6H12DLD1-cOVAAntibodyT cellPeptide
Mac
2D3 B6H12IgG -
+ + + + + + - - -+ + + - - - + - -
+ + + + + + + + +- -
- - - - + + - - -
05
10152025303540
p<5*10-5 p<5*10-6
p<5*10-445
I-Ab (MHC II) IgG B6H12 2D30
5
10
15
- IFNg+ IFNg
p<5*10-5 p<5*10-5
p<5*10-5p<5*10-6
Pha
gocy
tosi
s (%
)
p<5*10-6
p<5*10-4
p<5*10-4
p<0.01
p<0.005
- IFNg+ IFNg
Pro
lifer
atin
g ce
lls (%
)
0
10
20
30
40
50
60
B6H12 B6H12 B6H12DLD1-cOVAAntibodyT cellPeptide
Mac
2D3 B6H12IgG +
+ + + + + + - - -+ + + - - - + - -
+ + + + + + + + +- -
- - - - + + - - -
p<0.005p<0.05p<0.05
Unstained I-Ab stained
Fig. 3. After phagocytosis of cancer cells by anti-CD47, macrophages do not prime CD4+ T cells toproliferate. (A) RFP+ macrophages were coculturedwith DLD1-cOVA-GFP cancer cells in the presence ofIgG, anti-CD47 B6H12 (blocking), or anti-CD47 2D3(nonblocking) mAbs. The next day, CD4+ T cellswere isolated from OT-II transgenic mice and werelabeled with CFSE (0.5 μM). Analysis was performedon day 4, and the percentage of proliferating cellswas determined. Macrophages were pulsed withOVA peptide 323–339 as a positive control. (B) RFP+
macrophages were stimulated with IFN-γ to up-regulate MHC II levels. Phagocytosis and priming ofOT-II CD4+ cells were determined in the presence ofanti-CD47 mAbs.
Tseng et al. PNAS | July 2, 2013 | vol. 110 | no. 27 | 11105
MED
ICALSC
IENCE
SSE
ECO
MMEN
TARY

T-cell priming in vivo, OT-I (CD8+) T cells (CD45.2) were CFSE-labeled and adoptively transferred to CD45.1 recipient mice (Fig.5A). The next day, RFP+ macrophages were cocultured withDLD1-cOVA-GFP cancer cells in the presence of IgG or anti-CD47 B6H12 mAb. Macrophages were isolated by magnetic en-richment, and phagocytosis was verified by FACS analysis beforesubcutaneous (subQ) transfer into the footpad. After 4 d, thepopliteal lymph node was analyzed for the percentage of pro-liferating cells (CFSE-low) within the CD45.2+ gate. There wasan increase in proliferating OT-I T cells in mice receiving mac-rophages that phagocytose cancer cells via anti-CD47 B6H12mAb (Fig. 5B).
Macrophages Prime an Antitumor CD8+ T-Cell Response in Vivo AfterAnti-CD47–Mediated Phagocytosis of Cancer Cells. We next evalu-ated the functional effects of OT-I (CD8+) T-cell priming afteranti-CD47–mediated phagocytosis of cancer cells bymacrophages.To assess the efficacy of CD8+ T-cell killing of OVA peptide-displaying targets, CD8+T cells were isolated fromOT-I transgenicmice and were i.v. transferred to recipient mice (Fig. 6A). RFP+macrophages were cocultured with DLD1-cOVA-GFP cancer cellsin vitro in the presence of IgG or anti-CD47 B6H12mAb. After 2-hincubation, macrophages were isolated and injected into the foot-pad. After 4 d, mice were challenged with target cells (CD45.1splenocytes) to assess cytotoxic activity. CFSE-high splenocyteswere pulsed with 1 μM OVA class I-restricted peptide (OVA257-264, SIINFEKL) to make them targets for OT-I cytotoxic T cellsand then were mixed in a 1:1 ratio with non–peptide-pulsedCFSE-low cells before i.v. transfer. Analysis of draining lymphnodes 16 h later showed increased cell killing of peptide-pulsedCFSE-high lymphocytes in mice receiving macrophages that hadphagocytosed cancer cells by anti-CD47 B6H12 mAb (Fig. 6A).Next, the ability of CD8+ effector T cells to prime an antitumor
immune response was evaluated. CD8+T cells fromOT-Imice weretransferred into recipient animals (Fig. 6B). Macrophages werecocultured with DLD1-cOVA-GFP cancer cells in vitro in thepresence of IgG or anti-CD47 B6H12 mAb; then the macro-phages were transferred into the footpad on days 1 and 10. Ani-mals were challenged with EG.7 (OVA-expressing EL4) cancercells on day 14, and tumor growth wasmonitored over time. CD8+T cells primed by macrophages following anti-CD47-mediatedphagocytosis protected mice from tumor challenge (Fig. 6B).
DiscussionRecent studies from the I.L.W. laboratory have shown that ma-lignant cancers universally up-regulate the “don’t eat me” signalCD47, presumably in their progression to allow escape fromendogenous “eat me” signals that were induced as part ofprogrammed cell death and programmed cell removal (5, 6, 13,14, 16–18). These results indicate a role for human CD47-block-ing antibodies in cancer therapy via induced phagocytosis. Ourprevious experiments involved xenotransplantation of primaryhuman cancers into immunodeficient mice. We now have exam-ined the possible role of anti-CD47–enabled phagocytosis in an-tigen presentation of tumor peptides to T cells of the adaptiveimmune system. Here we have demonstrated that CD47 serves as
an “invisibility cloak” for both innate and adaptive immunity.Treatment with anti-CD47 blocking mAbs led to adaptive T-cellimmune responses, thereby providing an additional mechanism ofaction for anti-CD47 antibodies. OVA-specific OT-I (CD8+) andOT-II (CD4+) T-cell clones were used to follow the outcomes ofantigen presentation by macrophages after anti-CD47–mediatedphagocytosis of cancer cells engineered to express cytoplasmicOVA. Using in vitro and in vivo assays, we show that antigens arepresented to CD8+ T cells effectively, but the proliferative re-sponse of OT-II CD4+ T cells to loaded macrophages was di-minished compared with baseline levels. The baseline level ofOT-I CD8+ proliferation and OT-II CD4+ proliferation was 20%(Figs. 2 and 3), likely because of OVA released from cancer cellsthat become endocytosed or pinocytosed by macrophages andthen processed for presentation to both MHC I and MHC IIpathways. Together, our results suggest that anti-CD47–mediatedphagocytosis of cancer cells results in the presentation of negativesignals to CD4+ T cells and positive signals to CD8+ T cells. Inaddition, the CD4+ T-cell response was characterized by reducedregulatory T cells. This reduction might be attributed either todecreased proliferation of regulatory T cells in response to pep-tide or to less efficient regulatory T-cell differentiation. The invivo priming of an antitumor T-cell response by macrophagesafter anti-CD47–mediated phagocytosis of cancer protects micefrom tumor challenge. Anti-CD47 mAbs may represent a thera-peutic strategy for overcoming the regulatory T-cell contributionto immune evasion by cancer and initiating an effective antitumorcytotoxic T-cell response.In this system macrophages are the primary APCs that phago-
cytose cancer in response to anti-CD47 antibody and present an-tigen to CD8+ T cells. Perhaps this observation is due to higherlevels of SIRP-α expressed on macrophages than on dendriticcells or because macrophages phagocytose whole cells more ef-fectively. Because subsets of dendritic cells have been reported tovary in their levels of SIRP-α expression (19), it is possible thatother dendritic cell subsets may more effectively phagocytose can-cer in response to anti-CD47 antibody in vivo. While dendritic cellsare well known for their function in antigen presentation, in thisstudy we show that macrophages also have the capacity to stimulatea CD8+ T-cell immune response in the context of blocking theCD47-SIRPalpha axis. Prior to this work, the ability to presentexogenous antigens to CD8+ T cells has been largely attributed totheCD8+ subtype ofDCs.Understanding the roles of tissue-specificmacrophages and dendritic cells in response to anti-CD47 mAbs
Foxp
3-G
FP (%
)
IgG B6H12 2D3
p<0.05 p<0.05IgGFo
xp3-
GFP
B6H12 2D3
CD40 102 103 104 105
0
102
103
104
105 6.1
0 102 103 104 105
0
102
103
104
105 0.8
0 102 103 104 105
0
102
103
104
105 3.7
012345
Fig. 4. A reduction in Foxp3+ regulatory T cells occurs after anti-CD47B6H12–mediated phagocytosis of cancer cells by macrophages. RFP+ mac-rophages were cocultured with DLD1-cOVA-GFP cancer cells in the presenceof IgG or anti-CD47 B6H12 (blocking) or 2D3 (nonblocking) mAbs. The nextday, CD4+ T cells from OT-II/Foxp3-GFP+ transgenic mice were magneticallyenriched and were added to cultures. On day 4, the percentage of CD4+
Foxp3-GFP+ cells was quantified.
0 102 103 104 1050
10
20
30
17.3
0 102 103 104 1050
20
40
60
80
100
80.2
0 102 103 104 105
0
102
103
104
105
0.5
0 102 103 104 105
0
102
103
104
105
1.7
B6H
12Ig
G
SSC-A
CD
45.2
CFSE IgG B6H120
25
50
75
100
Pro
lifer
atin
g ce
lls (%
)
p<0.05
OT I(CD45.2)
Purify CD8 T cellsCFSE label
Co-culture with cancer(IgG vs anti-CD47)
Mac-1 enrichmentfor macrophages
C57BL/6(CD45.1)
FootpadInjection
A
B
BMDM
AnalyzePopiteal LN
Day 0
Day 1
Day 5
Fig. 5. After anti-CD47–mediated phagocytosis of cancer cells, macrophagesprime CD8+ T cells in vivo. (A) Experimental setup. BMDM, bone marrow-derived macrophages. (B) Adoptively transferred CFSE+ OT-I T cells were ana-lyzed in the draining lymph node by gating on CD45.2+ cells. The percentage ofproliferating cells was determined by gating on the CFSE-low population.n = 5 mice per group.
11106 | www.pnas.org/cgi/doi/10.1073/pnas.1305569110 Tseng et al.

warrants further investigation both in mouse and human systems(20–22).Antibody-mediated uptake of antigens via Fcγ receptor-medi-
ated endocytosis by dendritic cells can prime CD4+ and CD8+
T-cell responses in some circumstances (23, 24). In contrast, anti-CD47–mediated phagocytosis by macrophages predominantlyprimes CD8+ T cells. This predominant CD8+ T cell response isnot explained entirely by opsonization of cancer cells, becauseboth the anti-CD47 clones B6H12 and 2D3 bind DLD1-cOVA-GFP cancer cells, but only B6H12 is capable of inducing phago-cytosis through its ability to block the interaction between CD47on cancer cells and SIRP-α on macrophages. The preferentialactivation of CD8+ T cells may rely on routing phagocytosedcancer antigens toward theMHC I pathway or might be explainedby SIRP-α’s role as a negative regulator of macrophage function.Engagement of SIRP-α by CD47 leads to phosphorylation ofSIRP-α’s cytoplasmic ITIM motifs by SHP-1 and SHP-2. Thispathway is well described to inhibit macrophage phagocytosis, butmore recently, SIRP-α also has been shown to attenuate macro-phage activation by LPS by sequestering SHP-2, which is needed foractivation of the MAPK and NF-κB pathways (25). In addition,inhibition of SHP-1 in dendritic cells has been reported to enhanceCD8+ and CD4+ T-cell responses and reduce regulatory T cells,a response that protected mice against tumor challenge (26). SIRP-α also may negatively regulate cross-presentation by a mechanismnot yet understood, perhaps involving recruitment of cross-pre-sentation machinery to phagosomes. Alternatively, prophagocyticsignals such as calreticulin have been described to elicit antitumorimmunity (27–29). It may be that the unmasking of such propha-gocytic signals may route antigens preferentially toward the cross-presentation pathway.Other groups examining the effect of SIRP-α on dendritic cells
interacting with CD47 on T cells have reported T-cell activationor inhibition, depending on the context (30–33). In contrast, ourexperiments were designed to study the impact of antigen pre-sentation after anti-CD47 mAb–mediated phagocytosis of can-cer cells by macrophages. In our system, anti-CD47 B6H12 mAbdid not have a direct effect on T-cell activation or inhibition(Figs. 2 and 3).The involvement of both the innate and adaptive immune sys-
tems in the mechanism of action of anti-CD47 antibody has sev-eral clinical implications. First, our findings suggest a novel role ofanti-CD47 blocking mAbs as a vaccination strategy to enhanceCD8+ effector T cells recognizing antigens on phagocytosed
target cells. Second, in designing clinical trial protocols for testinganti-CD47 therapy in patients, immune monitoring of T cells maybe important for understanding clinical response to treatment andclinical outcomes. Third, anti-CD47 mAbs might be used clinicallyin combination with adoptive T-cell therapy or T-cell–activatingantibodies to enhance the adaptive immune response against tu-mor antigens andminimize toxicity.We conclude that anti-CD47–mediated phagocytosis of cancer not only functions in directlyclearing cancer cells but also can initiate an antitumor T-cell re-sponse to eliminate cancers. Patients receiving anti-CD47 therapymay benefit from both the innate and adaptive immune responsesagainst cancer.
Materials and MethodsMice. Mice, including C57BL/Ka (CD45.2), C57BL/Ka (CD45.1), and C57BL/Ka
Rosa26-mRFP1 mice, were bred and maintained at the Stanford UniversityResearch Animal Facility in accordance with the Administrative Panel onLaboratory Animal Care. All the animals were housed in sterile micro-insulators and were given water and rodent chow ad libitum. OT-I TCRtransgenic mice, OT-II TCR transgenic mice, and Foxp3-GFP mice were pur-chased from the Jackson Laboratory. OT-I mice have transgenic TCRs specificfor OVA257-264 in the context of H2-kb. OT-II mice have TCRs specific forOVA323-339 in the context of IAb. Both OT-I and OT-II mice have transgenicVα2 Vβ5 TCRs.
Molecular Biology. cOVA was cloned from pCI-neo-cOVA (plasmid 25097;AddGene) and was shuttled into the lentiviral pCDH-EF1-MCS-T2A-copGFPvector (System Biosciences) using EcoRI and BamHI restriction sites. Lentiviralproduction and concentration were accomplished using standard protocols.
Generation of Macrophages and Dendritic Cells. Whole bone marrow cellswere isolated from C57BL/Ka (CD45.2) or C57BL/Ka Rosa26-mRFP1 mice.Macrophages were generated by incubating whole bone marrow in mac-rophage colony-stimulating factor (10 ng/mL) for 7 d and harvesting theadherent fraction. Dendritic cells were generated in granulocyte-macro-phage colony stimulating factor (GM-CSF) (1,000 U/mL); on days 2 and 4 cellswere washed and medium was replaced with fresh cytokine-containingmedium. Nonadherent cells were replated on day 6 and harvested on day 7.
In Vitro Phagocytosis Assay. For the in vitro phagocytosis assay, 2 × 104
macrophages or dendritic cells per well were plated in a 96-well ultra-low-adherent plate (Corning), along with 2 × 104 cancer cells (DLD1-cOVA-GFP)in serum-free RPMI medium.
The indicated antibodies (10 μg/mL) were added and incubated for 4 h at37°. Macrophages were washed twice and analyzed using a BD LSR FortessaAnalyzer. The percentage of phagocytosis was calculated as the percentage
CFSE
IgG
0 102 103 104 1050
20
40
60 44.255.6B6H12
0
20
40
60 12.687.3
Cel
l kill
ing
(%)
IgG B6H12
p<0.005
01020304050A
B
Time (days)0 1 10 14
MacsubQ
MacsubQ
EG.7 tumorchallenge
OT I T cells iv
6 7
***
Tum
or v
olum
e (m
m3 )
8 9 10 11 12 13 14 15
*
***
*
Days post tumor challenge
350300250200150100500
IgGanti-CD47
Fig. 6. After anti-CD47–mediated phagocytosis ofcancer cells, macrophages prime an antitumor CD8+
T-cell response in vivo. (A) After anti-CD47–mediatedphagocytosis of cancer cells, macrophages primeeffector cytotoxic T cells. CD8+ T cells were isolatedfrom OT-I transgenic mice and were transferred i.v.to recipient mice. Macrophages (Mac) were cocul-tured with DLD1-cOVA-GFP cancer cells in vitro inthe presence of IgG or anti-CD47 B6H12 (blocking)mAb. Macrophages were isolated by magneticseparation and were transferred subcutaneously(subQ) on the next day. After 4 d, target cells(CD45.1 splenocytes) were labeled as CFSE-high(10 μM) or -low (1 μM). CFSE-high cells were pulsedwith 1 μM OVA class I-restricted peptide (SIINFEKL)to make them targets for OT-I cytotoxic T-cell func-tion. CFSE-high (peptide-pulsed) and -low (unpulsed)cells were mixed in a 1:1 ratio and transferred i.v.Draining lymph nodes were analyzed 16 h laterto determine the percentage of CFSE-high versusCFSE-low cells. The percentage of cell killing wasdetermined as described in Materials and Methods.n = 10 mice. (B) After anti-CD47–mediated phagocytosis of cancer cells, macrophages prime an antitumor CD8+ T-cell response. OT-I CD8+ T cells weretransferred i.v. to recipient mice. Macrophages were cocultured with DLD1-cOVA-GFP cancer cells in vitro in the presence of IgG or anti-CD47 B6H12 mAb, andthen macrophages were transferred on days 1 and 10. Animals were challenged with EG.7 (EL4 mouse lymphoma cells expressing ovalbumin) cancer cells onday 14, and tumor growth was monitored over time. n = 5 mice per group. *P < 0.05; **P < 0.01.
Tseng et al. PNAS | July 2, 2013 | vol. 110 | no. 27 | 11107
MED
ICALSC
IENCE
SSE
ECO
MMEN
TARY

of GFP+ cells within RFP+ macrophages or F4/80+ macrophages. For in vivotransfer assays, 5 × 105 macrophages and cancer cells were cocultured in thepresence of control IgG1 or anti-CD47 B6H12 mAb (10 μg/mL) and were in-cubated for 2 h. Macrophages then were separated from the cultures duringanti–Mac-1 magnetic beads (Miltenyi Biotec).
T Cell Priming Assay. For in vitro T cell priming assays, 104 macrophages werecocultured overnight with equal numbers of DLD1-cOVA-GFP cancer cells inserum-free RPMI medium. The next day, equal volume of RPMI+ 20% (vol/vol) FCS was added to the cultures. Peripheral lymph nodes were harvestedfrom OT-I or OT-II TCR transgenic mice and labeled with 0.5 mM CFSE(Molecular Probes). T cells were isolated using biotinylated anti-CD8 or anti-CD4 antibodies, followed by enrichment with anti-biotin magnetic beads(Miltenyi Biotec). Then 5 × 104 T cells were added to the cultures and ana-lyzed on day 3 (for OT-I T cells) or day 4 (for OT-II T cells). For in vivo T cellpriming assays, 2 × 106 CFSE-labeled OT-I T cells (CD45.2) were adoptivelytransferred i.v. into recipient mice (CD45.1). Macrophages were isolatedfrom coculture with cancer cells as previously described and were injectedinto the footpads of mice. Popliteal lymph nodes were analyzed on day 4 forCFSE dilution within CD45.2+ cells.
Antibodies and Flow Cytometry Analysis. Mouse anti-human anti-CD47 mAbB6H12 (IgG1) was obtained from Bio-XCell. Mouse anti-human anti-CD47mAb 2D3 (IgG1) and mouse IgG1 mAb were obtained from eBioscience. Forverification of binding of anti-CD47 B6H12 and 2D3 to DLD1-cOVA-GFPcancer cells, the cells were labeled with a saturating concentration of anti-CD47 antibody, followed by phycoerythrin-conjugated donkey-anti-mouseIgG (H&L) (eBioscience). Data were acquired using a BD LSR Fortessa Ana-lyzer and analyzed using FlowJo software.
In Vivo Cell-Killing Assay. In brief, splenocytes from C57BL/Ka (CD45.1) micewere labeled with 10 μM CFSE (CFSE-high) or 1 μM CFSE (CFSE-low). CFSE-high splenocytes then were pulsed in a six-well plate with 1 μM SIINFEKLpeptide for 1 h. Cells then were mixed in a 1:1 ratio with non–peptide-pulsedCFSE-low cells before i.v. transfer. To account for variation in the CFSE-high/-lowratio in the absence of peptide-specific lysis, control mice received CFSE-high and -low splenocytes not pulsed with SIINFEKL peptide and mixed in
a 1:1 ratio. Draining lymph nodes were analyzed 16 h later. The percentageof cytotoxicity was calculated as (1 − percentage of CFSE-high/percentage ofCFSE-low) normalized to the ratio in control mice.
Tumor Challenge. CD8-enriched OT-I T cells (1 × 106) were adoptively trans-ferred i.v. into recipient C57BL/Ka mice. Macrophages from syngeneic C57BL/Ka mice were cocultured with DLD1-cOVA-GFP cancer cells as previouslydescribed and then were isolated by magnetic enrichment and injected intothe footpads of mice. The tumor cell line E.G7 (EL.4 cells expressing thechicken OVA cDNA) was used for tumor challenge of mice (Amercan TypeCulture Collection). E.G7 cells (1 × 105) were injected s.c. into the righthindlimb of the mice in a 1:1 ratio with regular Matrigel. Tumor size wasmeasured every day using fine calipers, and tumor volume was calculatedbased on length × width × height × π/6.
Cytokine Assay.Macrophages were cocultured overnight with equal numbersof DLD1-cOVA-GFP cancer cells in serum-free RPMI medium. The next day,supernatants were harvested and submitted to the Stanford Human ImmuneMonitoring Core for cytokine analysis by mouse 26-plex Luminex assay(Affymetrix).
ACKNOWLEDGMENTS. We thank Theresa Storm and Libuse Jerabek for ex-cellent laboratory management, Tejaswitha Naik for antibody conjugation,Aaron McCarty for mouse breeding and management, Patty Lovelace andJennifer Ho for technical assistance with flow cytometry, the Stanford Hu-man Immune Monitoring Core and Yael Rosenberg-Hasson for assistancewith the Luminex assay, and Suparna Dutt for intellectual discussions. Fund-ing support for this work was provided by the Student Training and Re-search in Tumor Immunology (STaRT) Program of the Cancer ResearchInstitute (D.T.), the Virginia and D. K. Ludwig Fund for Cancer Research(I.L.W., D.T., M.M.), the Joseph and Laurie Lacob Gynecologic/Ovarian Can-cer Fund (J.V.P.), and by Grants R01 CA86017 (I.L.W.), P01 CA139490 (I.L.W.,S.W., H.C.-T., K.W.), P30 CA124435 (I.L.W.), and F30 CA168059 (K.W.) fromthe National Institutes of Health. D.T. is the recipient of a Howard HughesMedical Institute medical student fellowship. We also thank the StanfordProgram in Cancer Biology and StanfordMedical Scientist Training Programfor their support (D.T. and K.W.).
1. Unanue ER (1972) The regulatory role of macrophages in antigenic stimulation. AdvImmunol 15:95–165.
2. Burgdorf S, Kautz A, Böhnert V, Knolle PA, Kurts C (2007) Distinct pathways of an-tigen uptake and intracellular routing in CD4 and CD8 T cell activation. Science316(5824):612–616.
3. Bonifaz LC, et al. (2004) In vivo targeting of antigens to maturing dendritic cells viathe DEC-205 receptor improves T cell vaccination. J Exp Med 199(6):815–824.
4. Callahan MK, Wolchok JD, Allison JP (2010) Anti-CTLA-4 antibody therapy: Immunemonitoring during clinical development of a novel immunotherapy. Semin Oncol37(5):473–484.
5. Majeti R, et al. (2009) CD47 is an adverse prognostic factor and therapeutic antibodytarget on human acute myeloid leukemia stem cells. Cell 138(2):286–299.
6. Willingham SB, et al. (2012) The CD47-signal regulatory protein alpha (SIRPa) in-teraction is a therapeutic target for human solid tumors. Proc Natl Acad Sci USA109(17):6662–6667.
7. Brown EJ, Frazier WA (2001) Integrin-associated protein (CD47) and its ligands. TrendsCell Biol 11(3):130–135.
8. Oldenborg PA, et al. (2000) Role of CD47 as a marker of self on red blood cells. Science288(5473):2051–2054.
9. Kharitonenkov A, et al. (1997) A family of proteins that inhibit signalling throughtyrosine kinase receptors. Nature 386(6621):181–186.
10. Fujioka Y, et al. (1996) A novel membrane glycoprotein, SHPS-1, that binds the SH2-domain-containing protein tyrosine phosphatase SHP-2 in response to mitogens andcell adhesion. Mol Cell Biol 16(12):6887–6899.
11. Timms JF, et al. (1998) Identification of major binding proteins and substrates for theSH2-containing protein tyrosine phosphatase SHP-1 in macrophages. Mol Cell Biol18(7):3838–3850.
12. Tsai RK, Discher DE (2008) Inhibition of “self” engulfment through deactivation ofmyosin-II at the phagocytic synapse between human cells. J Cell Biol 180(5):989–1003.
13. Chao MP, et al. (2011) Therapeutic antibody targeting of CD47 eliminates humanacute lymphoblastic leukemia. Cancer Res 71(4):1374–1384.
14. Chao MP, et al. (2011) Extranodal dissemination of non-Hodgkin lymphoma requiresCD47 and is inhibited by anti-CD47 antibody therapy. Blood 118(18):4890–4901.
15. Kim D, et al. (2012) Anti-CD47 antibodies promote phagocytosis and inhibit thegrowth of human myeloma cells. Leukemia 26(12):2538–2545.
16. Edris B, et al. (2012) Antibody therapy targeting the CD47 protein is effective ina model of aggressive metastatic leiomyosarcoma. Proc Natl Acad Sci USA 109(17):6656–6661.
17. Chao MP, et al. (2010) Anti-CD47 antibody synergizes with rituximab to promotephagocytosis and eradicate non-Hodgkin lymphoma. Cell 142(5):699–713.
18. Chao MP, et al. (2010) Calreticulin is the dominant pro-phagocytic signal on multiplehuman cancers and is counterbalanced by CD47. Sci Transl Med 2(63):63ra94.
19. Saito Y, et al. (2010) Regulation by SIRPα of dendritic cell homeostasis in lymphoidtissues. Blood 116(18):3517–3525.
20. Asano K, et al. (2011) CD169-positive macrophages dominate antitumor immunity bycrosspresenting dead cell-associated antigens. Immunity 34(1):85–95.
21. Harshyne LA, Watkins SC, Gambotto A, Barratt-Boyes SM (2001) Dendritic cells acquireantigens from live cells for cross-presentation to CTL. J Immunol 166(6):3717–3723.
22. Allan RS, et al. (2006) Migratory dendritic cells transfer antigen to a lymph node-resident dendritic cell population for efficient CTL priming. Immunity 25(1):153–162.
23. Harbers SO, et al. (2007) Antibody-enhanced cross-presentation of self antigen breaksT cell tolerance. J Clin Invest 117(5):1361–1369.
24. Rafiq K, Bergtold A, Clynes R (2002) Immune complex-mediated antigen presentationinduces tumor immunity. J Clin Invest 110(1):71–79.
25. Kong XN, et al. (2007) LPS-induced down-regulation of signal regulatory protein alphacontributes to innate immune activation in macrophages. J Exp Med 204(11):2719–2731.
26. Ramachandran IR, et al. (2011) The phosphatase SRC homology region 2 domain-containing phosphatase-1 is an intrinsic central regulator of dendritic cell function. JImmunol 186:3934–3945.
27. Basu S, Srivastava PK (1999) Calreticulin, a peptide-binding chaperone of the endoplasmicreticulum, elicits tumor- and peptide-specific immunity. J Exp Med 189(5):797–802.
28. Nair S, et al. (1999) Calreticulin displays in vivo peptide-binding activity and can elicitCTL responses against bound peptides. J Immunol 162(11):6426–6432.
29. Obeid M, et al. (2007) Calreticulin exposure dictates the immunogenicity of cancer celldeath. Nat Med 13(1):54–61.
30. Ticchioni M, et al. (2001) Integrin-associated protein (CD47/IAP) contributes to T cellarrest on inflammatory vascular endothelium under flow. FASEB J 15(2):341–350.
31. Reinhold MI, Lindberg FP, Kersh GJ, Allen PM, Brown EJ (1997) Costimulation of T cellactivation by integrin-associated protein (CD47) is an adhesion-dependent, CD28-independent signaling pathway. J Exp Med 185(1):1–11.
32. Waclavicek M, et al. (1997) T cell stimulation via CD47: Agonistic and antagonisticeffects of CD47 monoclonal antibody 1/1A4. J Immunol 159(11):5345–5354.
33. Latour S, et al. (2001) Bidirectional negative regulation of human T and dendritic cellsby CD47 and its cognate receptor signal-regulator protein-alpha: Down-regulation ofIL-12 responsiveness and inhibition of dendritic cell activation. J Immunol 167(5):2547–2554.
11108 | www.pnas.org/cgi/doi/10.1073/pnas.1305569110 Tseng et al.


















