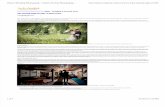
Animal Model of Cheetah
23
Animal model of Cheetah No. Characteristics 1. Distinctly different from other cats in anatomy and behaviour: fastest animal on land, it is the felid version of the greyhound. 2 Long-legged, slender bodies are built for speed not power or strength. 3. Much smaller and lighter than lions. 4 Cheetahs on average are 8 cm taller than leopards; male leopards normally outweigh male cheetahs. 5. But female leopards usually weigh less than female cheetahs. 6. Jaws not as large and strong as leopard or lion. 7. Blunt claws are semi-retractable and straighter than other cats. 8. Black ‘tear marks’ run from copper-coloured eyes to mouth. 9. Single, round, solid black or blackish-brown spots cover Brief introduction of the studied animal models.
-
Upload
fareez-hassan -
Category
Documents
-
view
21 -
download
5
description
animal adaptation
Transcript of Animal Model of Cheetah

Animal model of Cheetah
No. Characteristics
1. Distinctly different from other cats in anatomy and behaviour: fastest animal on
land, it is the felid version of the greyhound.
2 Long-legged, slender bodies are built for speed not power or strength.
3. Much smaller and lighter than lions.
4 Cheetahs on average are 8 cm taller than leopards; male leopards normally
outweigh male cheetahs.
5. But female leopards usually weigh less than female cheetahs.
6. Jaws not as large and strong as leopard or lion.
7. Blunt claws are semi-retractable and straighter than other cats.
8. Black ‘tear marks’ run from copper-coloured eyes to mouth.
9. Single, round, solid black or blackish-brown spots cover most of body.
10. Not good tree-climbers though they use sloping trees for vantage points.
Brief introduction of the studied animal models.

Body size and proportions:
(Male Avg: 45-55 kg)
(Female Avg: 35-40 kg)Pacinian CO
Total length Tail Shoulder height Mass
180- 220 cm 65-90 cm 70-90 cm (/Avg: 78-80 cm) 35-65 kg
No. Characteristics
11. Diurnal, hunting in early morning and late afternoon.
12. Eyes specialised for day vision; can apparently see detail up to 5-1/2 km away.
13. Specialised to prey on the fleetest antelope.
14. Hunt prey smaller themselves though male coalitions will sometimes hunt larger
animals.
15. Stalk within 50m or less of prey before racing at high speed for a few hundred
meters and using momentum of speed to knock prey down.
16. Almost never scavenge but often have their kills stolen by other predators,
especially lions and hyenas.
17. Generally solitary animals though males often live together in coalitions.
18. Gestation period is between 90 and 95 days.
19. Litters range from 1 to 9 cubs (average 3 to 5 cubs).
20. Females look after their young alone - cubs do not reach independence until 18
months of age.
21. After separation from mother, cubs may associate in sibling group for up to 6
months.
22. Average life expectancy in captivity is 10 to 12 years (up to 16); in the wild, few
survive beyond 7 or 8 years.
23. Cannot roar, but purrs when content.
24. Prefers open habitat with broken cover in order to get close to its prey. Avoids
forest or woodland with thick understorey.
25. Sparsely distributed in sub-Sahara Africa with more living in unprotected areas
than in parks and reserves.
26. With approximately 9,000 to 12,000 left in the wild, is one of the most
endangered big cats in the world.
27. The least aggressive of the big cats, cheetahs avoid confrontation and are not a
threat to humans.

The cheetah is built for speed and her survival is dependent on this gift. That is why she
lets other large predators steal her food rather than fight for it, as it avoids injury. It is more
efficient to chase and kill again. Apparently over a 2.5 mile chase in 1938 two mongrel dogs
brought a cheetah to bay. The cheetah rarely runs more than a quarter mile. It’s all about
cheetah speed.
Thus, high-speed locomotion is essential for the survival and success of many species
for both prey capture and escape from predatory attacks (Alexander, 2003). The
cheetah (Acinonyx jubatus) is widely acknowledged as the fastest living land mammal,
capable of speeds up to 29 ms−1 (Sharp, 1997). The greyhound (Canis familiaris) has been
selectively bred for high-speed locomotion yet can only attain speeds of 17 ms−1 during a
race (Usherwood & Wilson, 2005), despite both animals having a similar mass and gross
morphology.
Through the examination of the musculoskeletal anatomy of an animal it is possible to
gain insight into its locomotor capabilities. Measurements of muscle mass enable
calculations of muscle volume, to which power is directly proportional (Zajac, 1989). The
internal architecture of a muscle, example are fibre lengths and pennation angles, can also
be measured to calculate the maximal isometric force (Fmax) that a muscle can achieve.
However, to truly understand how muscle anatomy corresponds to an animal’s locomotor
abilities, muscle moment arms must be considered. The moment arm of a muscle is defined
as the perpendicular distance from the joint centre of rotation to the line of action of the
muscle, which can vary with changes in joint angle (Landsmeer, 1961; An et al. 1981; Spoor
& van Leeuwen, 1992). Through measuring the muscle’s moment arm we can convert the
linear forces of a muscle to rotational joint moments, which act to resist external forces and
move the limb.
One of the major functions of locomotor muscle is to support the body weight of the
animal by resisting the ground reaction force joint moments during stance. To support body
weight, an impulse must be applied to the ground (during stance) that is equal in magnitude
to the product of the animal’s body weight and stride time (Alexander & Jayes, 1978).
With increasing speed, an animal’s duty factor (proportion of the stride that the feet are in
contact with the ground) decreases, and therefore to maintain the impulse required to
support body weight, the peak ground reaction force must increase (Witte et al. 2006). It is

therefore crucial for high-speed predators such as the cheetah to be able to generate large
muscle joint torques to resist this force. Another requirement of high-speed locomotion is the
ability to swing the limb rapidly and reposition it for the next stride.
This is dependent on several factors: the limb’s inertia (Lee et al. 2004), the internal
architecture of the muscles (a long-fibred muscle has more sarcomeres in series enabling it
to contract at a high velocity), and the muscle moment arms (a short moment arm enables
greater changes in the joint rotation for a given change in muscle length).
The hindlimb anatomy of other high-speed animals has been studied in detail (Payne
et al. 2005; Smith et al. 2006, 2007; Williams et al. 2008a; Williams et al. 2008b), and
several common anatomical features have been noted. In all cases, a proximal to distal
reduction in muscle mass was observed, with many of the distal muscles being in series with
long tendons. This will reduce the inertia of the limb and therefore the amount of muscular
work required to swing the limb. A trend for a large volume of powerful hip extensor
musculature, which will aid acceleration, was also apparent (Usherwood & Wilson,
2005 Williams 2008b; Williams 2009b).
Acquisition of nutrients in relation to their metabolisms and energy for movement.
Unique nutritional requirements of felids must be recognized in feeding captive
cheetahs, including the need for high protein and fat diets, inclusion of dietary vitamin A (as
retinol), arachidonic acid, taurine, and niacin. General reviews of felid nutrition (Scott, 1968;
MacDonald et al., 1984) provide indirect evidence that the domestic cat remains the best
model for establishing dietary composition parameters (National Research Council, 1984) for
the cheetah, but a detailed summary of feeding habits with chemical analysis of natural prey
items would supply useful comparative data.
Composition should closely adhere to nutrient specifications as outlined above.
Recent studies with zoo felids have reported excesses of vitamin A and phytoestrogens, and
deficiencies of taurine in various commercial preparations. Responsive manufacturers are
correcting these potential health problems by reducing retinol to levels of approximately
15,000 lU/kg (dry basis), minimizing the addition of estrogenic plant materials, and
increasing the addition of taurine, particularly in heat processed meat products. No vitamin

supplements should be necessary with properly formulated and stored commercial diets.The
commercial preparations are formulated from the comparative dietary requirements and
received some field-testing for varying periods. The advantage of the commercial diets is
that they are readily available, require little or no labor in preparation, and are assumed to be
formulated with a sound nutritional basis. Economics determine the components of these
diets as the ingredients vary with the change in cost of producing the diet. Thus, the
guaranteed analysis remains the same, but the diet may vary in raw ingredients. The
guaranteed analysis label does not guarantee that the Ingredients of the diet are actually
utilized or available for utilization by the cheetah.
Movement in the environment (gravity, buoyancy, aerodynamics and hydrodynamics).
Speed
* Every facet of the cheetah's anatomy has been honed to serve one purpose, the chase,
and it represents the most efficient biomechanical design for speed in a quadruped.
* The cheetah's body extends and recoils in rhythmic sequence when chasing its prey, its
supple spine alternatively arching and stretching.
* Small, streamlined head; long, straight, light limbs; powerful hind legs; large, deep chest
and narrow waist; flexible shoulders and spine; long muscular tail; and semi-retractable
claws all combine to make it the fastest mammalian sprinter on earth.
* Cheetahs can attain speeds up to 110 to 120 km/h (approximately 70 to 75 mph) though
average speed during a chase is more like 85 km/h. A human sprinter runs 100 metres at
roughly 37 km/h.
* Cheetahs can accelerate from 0 to 80 km/h (50 mph) in just 3 seconds.
* Gazelles can reach speeds up to 90 km/h but have more stamina than cheetahs.

Stride
* The key to the cheetah's ability to reach such high speeds is the distance covered in each
stride (stride length) and the tempo at which they take a stride (stride rate):
A stride can be defined as the distance an animal covers once every foot has hit the
ground once.
As the cheetah speeds up, the distance covered per stride increases but the time
taken for a complete stride remains the same at a little under a third of a second
(about three and a half strides per second).
At top speed, a stride can measure just under 10 metres (32 feet), allowing the
cheetah to cover over 34 metres (112 feet) in 1 second.
The higher speed at which the limbs are moved provides enough energy to raise
the body into the air and propel it forward out of contact with the ground.
For more than half of every stride, the cheetah is airborne and as speed increases,
the floating phases increase in length, thus increasing the stride length.
* An increased stride length results from:
Elongated legs: the femur and tibia, and the humerus and radius differ very little in
length (as is true in greyhound dogs). Other felid species that depend on strength
rather than speed have proportionally shorter lower limbs than upper.
The scapula is pushed in to give slightly more length to the front legs.
Longest and most flexible spine of any large cat; half the muscle mass on a
cheetah is packed onto its spine, which allows it flex and extend.
Long, narrow, rectangular scapula as opposed to wide and fan-shaped scapula in other
felids.
Hip bones that pivot in their sockets.

Two phases per cycle when body is floating above the ground: extending body fully
as the fore limbs are protracted (extended) and flexing it as the hind limbs are
brought forward (flexed).
Hind legs that push sequentially rather than together.
* An increased stride rate results from:
Limb bones that are thinner and lighter than other cats. Bones of lower limbs and
paws are slender and light.
Muscles that do most of work are bunched high on each leg, close to body, and
light ligaments transfer muscular action down limb without need for heavy muscles
on lower legs.
To ensure stability and strength at high speeds, fibrous ligaments tightly bind fine
bones of the lower leg (fibula and tibia) in a rigid unit.
It’s a compromise that restricts the amount of rotation of the lower limb, crucial for
resisting stresses during the chase and allowing the cheetah to make sudden turns
at high speed.
As a result, the paws aren’t very good at anything other than running.
* A cheetah's back is hyper-extended going into the extended phase of a stride, allowing
the hind legs to push against the ground longer and the fore limbs to reach out further.
This phase is longer than the flexed phase.
* In the flexed phase of a stride, the back bows so deeply that the hind legs actually come
forward ahead of where the fore limbs last landed; thus, extra ground is covered between
strides.
* A cheetah can change lead the instant before its forefoot strikes the ground, so it can turn
immediately after zigzagging prey. Its long muscular tail acts as a counterbalancing
rudder or stabiliser when making fast, sharp turns so it doesn't roll over or spin out.

Shoulders
* The cheetah’s shoulders or scapula are free to swing back and forth or rotate in broad,
fluid arcs, increasing the length of the fore limbs and thus each stride.
* Shoulder structure:
A small clavicle or collar bone is embedded between 2 muscles but has no bony
attachment to the body.
Without attachment via the clavicle, shoulders and forelegs have no bony connection to
the body.
Body is supported between 2 forelegs in a muscular sling that extends on each side
from inside the scapula to the middle of the chest.
Support of body in muscular sling allows forelegs to swing freely in bigger arc and
also acts as a shock absorber to cushion landing after leap
Cheetah’s scapula is more elongated than any other cat species and the muscle
attachment on the inner surface of the scapula is unusually deep and narrow to
facilitate running (in the leopard, it is wide and shallow to facilitate climbing).
Cheetah's shoulder anatomy is a compromise: scapula is free to rotate, lengthening
each stride, but a small clavicle provides support when cheetah drags prey down
with forepaws.

Feet and Claws
* A cheetah's feet show several modifications:
A cheetah's pads are less rounded than other cats and resemble a dog's;
they are small and tough with edges that aid in gripping.
Digital pads and metacarpal pad are extremely hard and pointed at the
front, possibly as an adaptation for sudden braking and sharp turning.
Palmar pads bear a pair of longitudinal ridges instead of the usual slight
impressions, the functional equivalent of tyre treads, serving as anti-skid
devices.
Pad accompanying large, curved dewclaw is hard and calloused, rather
than soft in other cat species.
* A cheetah's partially retractable claws differ from other cats:
Cheetahs retain the physical apparatus to retract their claws, with the same
arrangement of muscles and ligaments that extends and withdraws claws
in all cat species.
However, changes in lower limbs to ensure stability at high speed result in
diminished action so retraction is only partial.
Cheetahs also lack the sheath of skin that protects and hides the claws of most
cat species.
As a result, their long, straight claws are not significantly retracted in the
rest position and are thus clearly visible on top of each digit.

Much blunting is caused by fully extending claws during chase for increased
traction. Claws act like cleats on track shoes, providing more grip on the
ground and aiding the cheetah's swift, sideways jinking movements as well as
helping acceleration.
There are two other species of cats that cannot fully retract their claws - the Asian
Fishing Cat and the Flat-headed cat.
Head and Teeth
* The cheetah's high domed skull is designed to give its jaws a vise-like grip
capable of maintaining a tight throttle-hold for 20 minutes or more. A steep angle
to the face raises the top of the skull and eyes, helping clamping action of jaws.
* A reduction in the size of the roots of the upper canines allows a larger nasal
aperture for increased air intake during exertion and for panting through the nose
while strangling its prey.
* Canines are short, rounded and sharp, with little space between them and the rest of
the teeth.
* Canines don't need to be long as their function is to hold prey by the throat during
strangulation. Except for small prey, cheetahs don't use their canines for severing
the spinal cord at the nape of the neck like other cats in the typical felid neck bite.
* More posterior teeth are large with expanded cusps to aid in crushing grasp on throat
of prey.
* Flat face and reduced muzzle length allow the large eyes to be positioned for
maximum binocular vision.

Endurance
* A cheetah's propulsion comes almost entirely from muscular force, mostly from its
strongly muscled hind legs, restricting its stamina and causing it to tire quickly.
* Cheetahs can only maintain top speed for between 300 to 500 m. Most chases
average 300 m in length and last about 20 seconds (rarely more than 1 minute).
* If a cheetah fails to catch its prey after 300 m, its breathing rate goes up to 150
per minute and it has to cool down for 30 minutes or more.
* A cheetah's body has been measured at 40 C during a 400 m sprint. Cheetahs
haven't evolved the evaporative heat release mechanisms of gazelles though
panting is a means of cooling down.
* Heavy, rapid breathing after a chase is indicative of enormous energy expenditure
and oxygen debt. Breathing rate during a chase may increase ten-fold from what it
was at rest.
* Breathing rates for cheetahs have been recorded at:
16 breaths per minute for a resting male.
156 breaths per minute for a male after a chase.
140 breaths per minute for a female after a chase.
195 breaths per minute after field immobilization.
* A cheetah's normal heart rate is 100 beats per minute. The average heart rate for
a 25-year-old human is 60 to 70 beats per minute (120 max, 185 extreme).

* Nasal passages, lungs, heart, liver, arteries, and adrenals are all enlarged:
Large, broad nostrils and sinus cavities allow more oxygen exchange and
also allow cheetah to pant through their noses while strangling prey.
Large lungs move oxygen more quickly throughout the body.
Big heart increases respiratory rate, enabling blood to move from the lungs to
the muscles rapidly.
Highly muscular arteries allow for maximum delivery of oxygen.
Sensory reception against environmental stimuli.
Focusing on Mechanoreception of Cheetah
Specialized cell or organ that can transform mechanical stimuli, such as pressure
changes into electrical signals that can be interpreted by the rest of the nervous system.
All organisms, and probably all cells have the ability to sense and respond to
mechanical stimuli.
Mechanoreception is very important for cell volume control of cheetah, and the
sense of touch, hearing and balance palys a critical role in regulating blood pressure in
Cheetah. There are two types of main mechanoreceptor: ENaC (Epithelial Sodium
Channels)and TRP (transient receptor potential) channels.
Both are attached to cytoskeleton and to extracellular anchoring proteins, pulling
on the ion channel and causing conformation change that alters the movement of ion

across the membrane.Changing the membrane potential of the cell allowing the cell to
transduce mechanical signals into electrical signals.
Touch and pressure receptor for Cheetah.
Mechanoreceptors that detect touch and pressure can be grouped into three classes.
Baroreceptors detect pressure changes in the walls of blood vessels, part of the heart,
and in the digestive, reproductive, and urinary tracts of vertebrates. While
proprioceptors monitor the position of the body.
Cheetah also has vertebrate tactile receptors which are widely dispersed. It is isolated
sensory cell embedded in the skin. Some of these receptors are simply free nerve
endings that are interspersed among the epidermal cells of the skin, whereas others a
re associated with accessory structures. Merkels disks are free nerve endings that are
associated with an enlarge epidermal cell called the Merkel cell. This receptor have very
small receptor field, and are used for fine tactile discrimination. Both the free nerve
endings and Merkel’s disks are slowly adapting tonic receptors that are most sensitive
to indenation of the skin, and are thus important for sensing light touch and pressure on
the surface of the skin. We use the Merkel’s disks in our skin when we perform tasks
such as reading that require very fine discrimination.
Figure 6.13
The nerve endings of the root hair plexus, which are wrapped around the base of hair
follicles, monitor movements across the body surface. When a hair is displace, the
movement of the hair follicle causes the sensory nerve endings to stretch, stimulating
mechanoreceptor protein on dendritic membrane. These receptors are rapidly adapting
phasic receptors, so they are most sensitive to change in movement. For example you
can often sense when an insect crawls across your skin, but you may not detect an
insect that is not moving.

Pacinian corpuscles are located deep within the skin and in the muscles, joints and
internal organs. At almost a millimeter in length, they are actually visible to the naked
eye in sections of the skin, and a typical human hand contain as 400 of these receptor.
Pacinian corpuscles contain sensory dendrite surrounded by up to 70 layers of tissue
with a viscous gel between them. These layers, called lamellae change shape,changing
the shape of the sensory dendrite and initiating a change in the membrane potential.
The viscous gel quickly returns to its original position, even in the presence of
continuous pressure, returning the membrane potential to its resting level. As a result,
the sensation of pressure disappears even though the pressure is still present at the
surface of the skin.
When the pressure is removed, the connective tissue layers return to their normal
shape, pulling on the nerve ending, which causes another change in membrane
potential, and another stimulus. Thus, Pacinian corpuscles are rapidly adapting sensory
receptors that are sensitive to both the beginning and the end of stimulus. This property
makes Pacinian Corpuscles especially sensitive to vibrations. So, when you feel your
cell phone vibrating, it is your Pacinian corpuscles that detect that incoming call.
Pacinian corpuscles have relatively large receptive fields, and thus do not allowed for
fine-scale discrimination of touch sensations.
Ruffini corpuscles are located in the connective tissue of the skin and with the
connective tissue of the skin and with the connective tissue of the limbs and joints. They
are sensitive to stretching of the skin and movement of the joints as we move
around.Ruffini corpuscles work together with other proprioceptors to help an animal
determine the location of its body in space. When it hit the snooze button on your alarm
clock without even opening your eyes, it is your ruffini corpuscles that help Cheetah do
so.
Vetebrate proprioceptors monitor body position
In addition to touch and pressure receptors such as Ruffini corpuscles , there are three
major groups of veterbrates proprioceptors associated with the joints and limbs:

1. Muscle spindles on the surface of skeletal muscles monitor the length of the
muscle. Each muscle spindle consists of modified musclefibers called intrafusal
fibers enclose in a connective tissue capsule.
2. Golgi tendon organ are located at the junction between a skeletal muscle and a
tendon. These receptors are stimulated by changes in tension in the tendon.
3. Joint capsule receptors are located in the capsule that enclose the joints. Several
types of receptors are in this category, including receptors similar to free nerve
endings, Parcinian corpuscles, and golgi tendon organ. These receptors detect
pressure , tension, and movement in the joint.
4. Proprioceptors typically do not aapt to stimuli, and thus constantly send
information to the central nervous system regarding body position. Another class
of more rapidly adapting receptors is responsible for detecting movement, and
provides the sense of kinaesthetic.
Regulation of endocrine systems against predation and stress response.
Thermal physiology in maintenance of constant body temperature.
Cheetah are the fastest terrestrial mammals. During a hunt, cheetah metabolism
and heat production increases by more than fifty-fold. Treadmill studies (Taylor and

Rowntree 1973) conclude that neither evaporative nor non-evaporative heat loss
increase during a run, so all metabolic heat is stored.
If cheetah indeed do use this heat-storage strategy, the duration of a sprint (and
hence hunting success) would be determined by the amount of heat the cheetah can
store without thermal compromise. Example, body temperature was measured and
activity for every 5 min, using biologging, in six free-living cheetah in Namibia. Test
conducted whether free-living cheetah employed heat storage during hunts, and
whether hunts were thermally limited.
There are no evidence found that hunts of free-living cheetah were thermally
limited. At maximal activity levels, body temperature averaged less than 39ºC, much
less than the 40.5ºC thermal limit to exercise reported for captive cheetah (Taylor and
Rowntree 1973) and within the daily variation of body temperature of free-living
cheetah. One of our male cheetah came close to maximum hunt distance (500m) and
speed (100km.h-1; upper left panel), yet his rate of body temperature rise(0.02ºC.min-1)
was a fraction of the estimated 1.6ºC.min-1 required to store the heat produced by
cheetah running at 110km.h-1.
.
Based on 15 hunts per cheetah, we conclude that the increase in body
temperature following successful hunts was twice that of unsuccessful hunts, despite no
difference in activity levels. Therefore physical exertion of the hunt contributed little to
the increase in body temperature. It may be that the energy required subduing prey
further increases the heat production or that the act of suffocating the prey may prevent
the panting required to dissipate the heat produced.
However, because the increase in body temperature was evident in cheetah not
directly involved in the hunt, it is proposed that the increased body temperature
following a hunt may reflect stress or excitement, and is elevated following hunting
success.