
Anderson 1967; Calow 1984!.
36
CHAPTER V ENERGETICS OF FEEDING AND REPRODUCTION FOR MALES AND FEMALES OF THREE SPECIES OF HAWAIIAN SUTTERFLYFISHES INTRODUCTION An animal's fitness depends on acquiring energy by feeding, and partitioning this energy between reproduction and other metabolic demands Calow 1985!. Many foraging models use energy intake from feeding as the major currency in the determination of fitness Pyke et al. 1976!, These analyses assume a relationship between energy intake and reproductive fitness. Each animal, however, must partition usable energy among the demands of current reproduction, maintenance activities, growth, and storage for future survival and reproduction. In addition, there is an unavoidable loss of energy in the form of heat and unutilized waste products. Zf energy is a limited resource, then energy budgets of animals will be sensitive to natural selection Anderson 1967; Calow 1984!. Animals can meet their energy budgets by adjusting their energy intake by changes in foraging behavior, or by adjusting how this energy is partitioned. An individual animal's activity levels, foraging behavior, and partitioning of energy may all differ between the reproductive and non-reproductive seasons Elliot 1979; Soofiani 275
Transcript of Anderson 1967; Calow 1984!.

CHAPTER V
ENERGETICS OF FEEDING AND REPRODUCTION FOR MALES AND FEMALES
OF THREE SPECIES OF HAWAIIAN SUTTERFLYFISHES
INTRODUCTION
An animal's fitness depends on acquiring energy by feeding, and
partitioning this energy between reproduction and other metabolic
demands Calow 1985!. Many foraging models use energy intake from
feeding as the major currency in the determination of fitness Pyke et
al. 1976!, These analyses assume a relationship between energy intake
and reproductive fitness. Each animal, however, must partition usable
energy among the demands of current reproduction, maintenance
activities, growth, and storage for future survival and reproduction.
In addition, there is an unavoidable loss of energy in the form of
heat and unutilized waste products. Zf energy is a limited resource,
then energy budgets of animals will be sensitive to natural selection
Anderson 1967; Calow 1984!.
Animals can meet their energy budgets by adjusting their energy
intake by changes in foraging behavior, or by adjusting how this
energy is partitioned. An individual animal's activity levels,
foraging behavior, and partitioning of energy may all differ between
the reproductive and non-reproductive seasons Elliot 1979; Soofiani
275

and Hawkins 1985; Wootton 1985! . The energetic trade-o ff between
current reproduction and growth and survival for future reproduction
is a basic premise of life history theory Wi1.1iams 1966; Stearns
1976!.
Energy budgets may also differ between the sexes. Females are
expected to contribute more energy into each reproductive effort than
a male Bateman 1908; Trivers 1972!, a situation resulting from
anisogamy, where the female's energy investment in ova is greater tl
the male's investment in sperm. Although males may contribute
significant amounts of energy to offspring in terms of parental care,
it is the initial inequality of anisogamy which is considered to be
the basis of many social systems Trivers 1972; Wilson 1975; Emlen and
Oring 1977!. In situations without parental care, such as occurs in
broadcast spawning reef fishes, the inequality of male vs. female
investment in offspring is expected to be a predominant factor
affecting mating systems.
Schoener �969, 1971, 1983! broadly categorized animal foraging
strategies as lying along a continuum from foragi~g "time minimizers"
to "energy maximizers". Time minimizers limit their feeding time to
the minimum necessary to meet their daily metabolic requirements.
This maximizes the time available for other activities. Ebersole
�980! has termed this an energy maintenance strategy. In contrast,
energy maximizers forage to maximize their daily rade of energy
intake. Energy in excess of that needed for metabolic maintenance may
be channeled into increased reproduction or stored for future use.
276

Schoener �971! proposed that in many species, males tend to be
foraging time minimizers, while females tend to be energy maximizers.
Finally, different species wi.th different energy budgets are
expected to have differing patterns of energy intake and partitioning,
These differences may be reflected in different social systems.
Studies of energy budgets in fishes have concentrated on
freshwater species, predominantly salmonids, under hatchery or
laboratory conditions Elliot 1976, 1979; Brett and Groves 1979;
Brafield 1985!. Field studies of several temperate freshwater fishes
reveal seasonal differences in energy intake and partitioning
reviewed by Soofiani and Hawkins 1985!. Evidence for sexual
differences in energy budgets is less common Newsome et al. 1975;
Diana 1983; Wootton 1985!. Very few studies of reef fishes have
measured energy intake and partitioning from an ecological perspective
Muir and Niimi 1972; Brett and Groves 1979!. For several reef fish
species, where sexual differences in foraging behavior are known,
females feed more than males: e.g. acanthurids Robertson et al.
1979!, pomacentrids Kbersole 1980!, labrids Ross 1983, Green et al.
1984! and scarids Warner and Downs 1977!. Hoffman �983! found that
in protogynous gi~da~s spp. females reduced their foraging time when
they changed sex to become males.
It is generally assumed that additional energy intake by female
fish is channeled into increased egg production. This assumption has
rarely been tested Wootton 1985!. Hirshfield �980! showed that
increased food rations led to higher female fecundity in the Japanese
277

fecundity per spawn was limited by female size, however, food ration
was the most important factor determining the number of spawnings per
breeding season Wootton 1977, 1985!.
The present study compares energy intake and energy partitioning
hy mains and fsmalss of tmo coral-fasdfng huttarflyfishss, ~chs codon
"* I
6b
coral and supplements its diet with algae and polychaetes Hobson
1974; Hourigan Chapter III! . These species have similar social
systems, consisting of long term, apparently monogamous pairs in which
both sexes defend a shared feeding territory Tricas 1985, 1986;
Hourigan 1986a, Chapter III!. The findings from these two species are
compared to data collected during the breeding season of a third
butterflyfish, ~ergo i~ ~~, which exhibits a haremic social
system Hourigan 1986b, Chapter IV!, It is an omnivore which feeds
primarily on polychaetes and algae Hobson 1974, Hourigan Chapter IV!,
All three species are broadcast spawners, which release gametes into
the water column and show no parental care. A series of eight
experiments were conducted to determine seasonal and sexual
differences in energy intake and absorption efficiencies, and the
partitioning of this energy between somatic, storage, and reproductive
tissues. These results are compared to field observations of feeding
and activity in these species.
278

METHODS AND RESULTS
Z. Study sites and field collections:
Methods:
C
coral reefs at Puako, Hawaii Lat 19 58'N, Lon 155 51'W; for a
complete description of the habitats and observation methods see Hayes
et al. 1982; and Hourigan Chapter lI and ill!. The third species,
~Cltaetod freebll, eas rare at Puako. Therefore, observatfons and
collections were conducted in similar habitats at Kahe Pt. Lat 21
21'N, Lon 158 8'W! off the leeward coast of the island of Oahu
made at Kahe Pt. and at coral poor areas at Portlock, Oahu Lat 2lo
16'N, Lon 157 4'W!,
collected by spearing at Puako during three times of the year: the
non-breeding season in fa11 November 1980!, the height of the
breeding season in spring March 1981!, and the end of the breeding
season in summer July, 1981!. individual C ~f em~ were collected
at Kahe Pt, Oahu during the spring March 1981!. Additional
November 1983. Each fish was either dissected immediately, or
labeled, double bagged and frozen for later analysis.
279

In the laboratory individual fishes were thawed, and standard
length SL! and wet weight were measured. Fishes were then dissected,
and the liver, gonads, and fat deposits surrounding the intestine
removed and weighed. Three small sub-samples from each ovary were
removed, weighed, fixed in l0% formalin and preserved in 70% alcohol.
Numbers of vitellogenic eggs vere counted and staged according to the
methods of Hourigan and Kelley �985!. The remaining tissues as well
as food material from the pouchlike stomach, and portions of feces
from the rectal region were saved to be utilized in the experiments
described below.
Results.'
female ~ ~emb~ was collected. Pairs were composed of one male and
one female, both reproductively mature. Length-weight relationships
for males and females of the same species were similar, indicating
C
size. In both paired species, males vere usually larger than their
pair-mates paired t-tests, p�.05 for both species!. The mean size
"c
a The haphazard collection of C
~fremb precluded similar comparisons. Observations of harems
280

Figure 5.1a Length-weight relationship for males and females of three. b! ~
uadrimaculatus, and c! C f~em~b

20
0 sL mm!
282

Figure 5. lb Length-weight relationship for males and f'emales of three
283
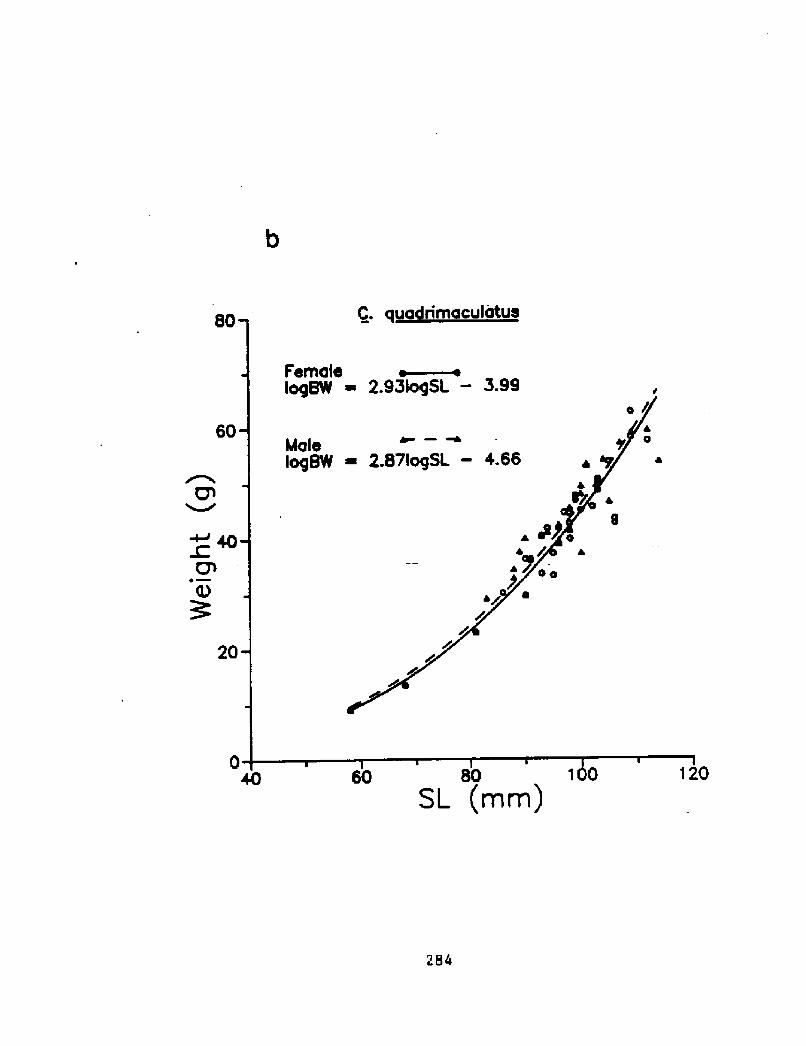
~ ~ ~ ~ ~ I ~80
60
20 sL mm!284

Figure 5, lc Length-weight relationship for males and females of threespecies of Hawaiian butterflyfishes: c! ~ ~i~~.
285

20 sL mm!286

indicated that males were visibly larger than their associated
females.
Gonosomatic indices CSI 100 X ratio of wet weight of gonad to
total wet weight of body! of both males and females of the paired
species were highest during the spring Table 5.1!. Comparisons of
GSIs of fish with very different body sixes are difficult due to
associated body-gonad allometries deVlaming et al. 1982!, There
were, however, no significant differences within each species among
the body sizes of fish in the three seasonal samples Independent one-
way ANOVAs, p�,5!, Fecundity, defined as the the total number of
vitellogenic oocytes yolked eggs! contained in the ovary of a female,
was also highest in spring, as was the median ova diameter Table
5.1!. Almost all vitellogenic oocytes were of a similar size and
stage of development. Hydrated eggs were only found on two occasions,
c- ~
prior to the new moon. Spawning was also observed on that occasion.
Examination of the two ovaries containing hydrated eggs showed that
77% to 82% of large vitellogenic eggs had undergone hydration
synchronously. This allo~ed estimation of the batch fecundity,
defined as numbers of eggs spawned at one time, estimated as 79.54 of
the total number of vitellogenic eggs Table S.l!, assuming that the
two remaining species spawned a similar proportion of all vitellogenic
oocytas. ~Chaeeoda ~ambit had the largest estimated batch fecundity
Preliminary histological observations of the testes of all three
species revealed no evidence of protogynous sex change. Testes
287

eIjl
CD w
OO
6 hlru W
I
ev e
CVCV
no4
CjjC 0 0 0IIIIjj
rCD O
OO
~ CDCV IP
~ ~wO
O I-mg
O
Ill
CVO
I
O
Cl WIVI M
~ ~n O
CJj
III
aS IC
II
4 II
c 0 0ClCj
V
O
< coP! Mvl IM 4!
tel
H OEV CV
OOIS
0
I ICS
OIO 4>
cv O
O O
IIO CD
OSj
0 U C
e
C i
0CJCl
CjCJj
ISCJ
Cj
C OSC
kJ ~C CSEPn ~0 IS
IS WC0
~ W
Cj~ kJ
Cj
Cj
~ ~~ O
I4Cj Ig3
V!
OI
O
CD M~ ~~ ~
OOc
0 0kJILIIS
I I I
OCVCV
Ch M
O Iw lh
r
CD
CV 0
OO~ ~
A H
~ 0CIS ~ ~aA
K CfJ
lg toaA
K CJj VICI
OuC CjCj e
Cj
Ijj A
kJ~ e
~ eC 00Cj C
U IjjILI K W
O OVl
CV~ O
OI
o
VkJ 'CI
Ijj 'UkJ C 4g N CJj
CI C
CO
CjClC
IU
IS
e
0kJ
IIIC
kJ0 CI
ILI
CjC0 CjCS ~ w
Ijj wIj! aa Vw oo~
IS W3
~ U~~ IjjC 4 'Cj
E kJ4W IS4 III eE kJ~0 mCJ ~ C
C o V Ijlgp III C
ECI
0 0
Ijj0WCC
Sj CI
IIIPlo
kJ CC kJ gjCJ IS~CJ
Ijj
SjCj ~ 0 MC
KIjj III~ " > Cj CI
Cj M
ew IS«gl IS 0 USj kJ IScol WC
Ijj4 kJ ISx
eg 4kJe IsISCII
Ijj AlCfl IS ~ j IS< ca O «I <
~ ~IS 4 CJ
288

contained no atretic oocytes, nor- was a membrane lined ovarian lumen
remnant present.
In addition to the adult fishes, six non-territorial 9
collected in March 1981. These fish were significantly smaller than
territorial individuals of the same species independent t- tests for
males and females, p�,01!, Both males and females were represented,
but neither were sexually mature i.e. ovaries did not contain
vitellogenic oocytes, and testes did not contain live sperm!.
IZ. Field and Laboratory Kxperiments.
The following experiments were designed to estimate rates of
energy intake in the field, in order to determine how this energy was
partitioned, and to estimate spawning rates of females.
Experiment 81.: Energy intake per feeding bite.
Methods:
in the field by divers using hand nets. These fishes were transported
to the Hawaii Institute of Marine Biology in aerated buckets and
placed in large �00 1! tanks with flow-through filtered seawater and
plastic tubes for shelter. Live coral colonies coral heads! were
placed in each tank for food, and the fish were left undisturbed for
289

at lease 24 hrs. All coral colonies used in these experiments were
collected from Kahe Pt. and different locations in Kaneohe Bay,
Individual, healthy colonies were removed and transported back to the
laboratory where they were kept in an outdoor water table wi.th free
flowing sea water. Fresh corals were collected on the day before each
experiment,
Prior to the experiment, an individual was transferred to a 40 1
aquarium along with a freshly collected coral colony of a particular
species, Only fish which fed on the offered coral colony were used.
After 30 min the coral colony was removed, and the fish starved for 24
hrs to allow gut clearance and acclimatization to the test aquarium.
On the following day, a new coral colony of the same species
previously offered, was placed in the test aquarium, separated from
the fish by a clear partition. After one hour, the coral polyps had
expanded. and the partition was removed. Each experimental fish was
observed for 30 min following its first bite on the coral. All
feeding bites were counted by an observer sitting quietly 2 m from the
aquarium, or recorded on videotape for subsequent counting. The
behavior of fishes did not appear to differ during early trials of
these two recording methods, so all subsequent observations were
conducted with an observer present. Eight replicate trials with C
were conducted, with each of the three most abundant
Hawaiian coral species ~Peri ~abate, giHHi~e ~coa ~es and
290

After' 30 min, the experimental fish was removed from the
aquarium, sacrificed immediately, weighed and measured SL! . The fish
was dissected, its sex determined, and stomach contents were removed.
The length of the gut was measured, and the distance cleared by food
from the px'evious day was measured. The stomach contents vere
inspected, weighed, then dried to constant weight in a 60 C oven.
Each sample of dried stomach contents vas ground and homogenized
using a mortar and pestle. The homogenate was further subdivided into
sub-samples which were formed into pellets, weighed, and either ashed
at 500 C and re-weighed to determine ash free dry weights AFDW!, or
combusted in a Phillipson oxygen microbomb calorimeter, as describedR
by Paine �971!, to determine caloric content. In general, four sub-
samples of each dried tissue were analyzed for AFDW, and four to five
sub-samples � to 20 mg each! were analyzed for caloric content. In
this manner, dry weights, ash-free dry weights and caloric values were
obtained for the samples. These were then calculated on a per-bite
basis.
Results.
o
introduced to the aquarium. The amounts of each coral ingested are
summarized in Table 5.2. More than 95% of the material ingested in 30
min remained in the pouchlike stomach. Organic content and caloric
content cal/mg AFDW; 1 cal - 4.81 joules! wex'e slightly higher for
stomach contents of fish feeding on ~ ~scend ina than for those of
291

CI
C C CIhlE
hl
hl
J U E~ P4
CV
I
CVO OCO h
C hlhl
I
OQ
C0CVm U C
4J
8
Cl4JhI
0
hl4 4I
W 'l5O
CV
Q
8 CIhI
lh
Dh
E h!hI
4 0 ClU
U
0
CI C0
C0
~0' a.g 0 Ehlr 0
U hlCI
'0
W 00
ul hl
a r ~IU
~~ 0
hIgp C ClCC4
hl~BCCI
hl4 Ihhl 0JJrACj8 4
hlU ~
CIC
00
W hl ~hl
~ CO
8 CI ~ Nhl
C CI~g~
hl"eCU U
~ Cl C 0 Uhl
CI
eI C
I E
O gW Ch
Ch O
W Cfl
Cl O
M R
4! Oh
~ A&R
oI COW
~ hI MAI W R
I I
R N' ~cL
CO R
OR
CV
CI
Pl QO g
I OR
eoOI nOI Q gI OCh
hlJJ
Cl
ooce E
hl C3
II
CVI r
hI
hl4! W hl
CIC 40
C5 hI0 U
U 4W W C40
g
4~ hl 0
kJ 0 Uhl
T4
CIIJ hl
UC
Ch0
hlhl
SChl C4~CCI
hl
C
hI
08 ~0 CIW U
0 hhhl
CQ4
g hl
IXIhl rgl~ 1~I
8
0U
CIU4hl
40
CI
04
0
IfJhl
Uhl
C 0-hl
hl
CI4 0
C
hlW W
00C
CIhl Cl
aIh
06 mC CI
kJ
292

fish feeding on the two pi~~ spp. Net intake in calories per bite
was higher for fish feeding on g ~me ~na than those feeding on
2 ' I'
feed.
Examination of the stomach contents of fishes feeding on P
medea ~ showed complete oral disks with tentacular rings. Only a
few such rings were visible in the stomachs of fish feeding on ~~eg
spp.. No calcareous material was ingested. Examination of the corals
themselves revealed that most polyps on g<~~s spp. were still intact
with perhaps only portions of one polyp removed per bite, After a
single bite, polyps in a 10-15 mm radius of the bite retracted
completely into the calyces within 15 seconds. Additional polyps were
partially retracted in a radius of 20-30 mm around the bite. If left
undisturbed, polyps would come out after 3 to 6 min.
Experiment 82. Energy intake per feeding bite: Field observations.
Methods:
To determine energy intake per feeding bite in the field, two
subsequently followed for 30 min after they had begun feeding. The
number of feeding bites by each fish on different substrata were
counted. The fishes were then speared, placed in plastic bags and
293

returned to the laboratory on ice. Fishes were weighed and measured,
and their stomach contents removed, dried and processed as in
Experiment ¹1. These energetic values could then be compared ta
values from Experiment ¹1 and field observations of feeding.
Results:
s
colo :' c
ncc
the hard substratum. There was some feeding on g ~i~L. The third
species, ~ ~<~3~ fed on the hard substratum, in crevices, in the
Ecl
After 30 min of feeding, 95% of the newly ingested food remained
in the stomach. Inspection of stomach contents revealed that C
contents included large numbers of zooxanthellae and both intact and
discharged nematocysts, Stomach contents of C. u drimaculatus
contained coral material as well as polychaetes, small crustaceans and
algae. The stoaach contents of t ~frembli' contained only
polychaetes, crustaceans, algae and sea urchin pedicillaria. Females
of all three species had more material in their stomachs, and a
greater calculated energy intake than did males Table 5.3!.
294

tU
ICI
Ce
E 0
I
OO
4 ICI0
CV
O ICV
CV
00C
'0 Q QLN
CD
Cl
C
a5 IIE WO
R CIR
O
O
I
CQ
CItp cv
IItd M
KO
IW 0
O 44Oc0
lIl
JJ
'W 04 0
IIW
O
ClV
ICl
C VCl
A Cl
295
C ILI
SlO 0
0
0
Pl ~ Mhh W
lII
IO
CJ h4
ul
~ / ym
CClClIQ Clu '0CN
QV V
0 AIm
lCI S
Ol
Q 44J
V 0 c5V

Experiment 4 3. Rate of food clearance through the gut,
Methods:
Energy absorption from food intake depends in part on the time
that food remains in the gut. A rough measure of the rate of food
clearance through the gut was determined by sacrificing fish at 6, 12,
24 and 48 hrs after they were fed, and measuring the amount of the
stomach and intestine which was cleared of food material. Two male
The guts were removed, the length of the intestine was measured, and
the portion which was cleared of food was recorded.
Results:
After 6 hrs, the stomach was empty, as well as the first 32% SD
4.6%! of the intestine. After 12 hours, 82% SD 8.5%! of the
intestine had been cleared of food, After 24 hrs 88% SD 2. 3%! of the
intestine was cleared. Same fecal material was still retained in this
last 12% even after 48 hrs. There were no consistent differences
between males and females. Sample sizes in this experiment were small
from experiment 1 and all three species from
experiment 2 had similar percentages of cleared areas corresponding to
the last time they had fed.
296

Experiment «4. Estimation of caloric intake and absorpt.ion.
Methods:
In this experiment, the caloric intake and relative absorption
assimilation! efficiency of male and female fishes were determined.
Food in the stomachs of teleosts is sub! ected to the action of HC1 and
peptidases, but little or no absorption occurs there Kapoor et al.
1975!, Thus as feeding is relatively continuous Hourigan Chapter
III! and food mo~es at a constant rate through the gut Experiment
«3!, the energy content of the diets of fishes can be estimated by
analyzing the energy content of the stomach contents Talbot 1985!.
In butterflyfishes, the stomach is followed by a very long intestine
where most absorption of nutrients occurs. At the distal end of this
intestine is a short rectal region. The energy content. of material in
this region probably approximates that voided in the feces.
t
were collected by spearing in November 1980, March 1981 and in July
1981, at Puako, for a total sample of 48 fish of each species. Pair-
mates were collected at. the same time, but different pairs were
collected at different times of the day. Six individual male and six
female C Stern'blti vere collected in the afternoon on several
different days in March 1981 at Kahe Pt.. Fishes were frozen
immediately, and subsequently partially thawed before analysis. This
allowed removal of the stomach contents and feces with little
contamination by material from the remaining portion of the intestine
Talbot, 1985!. Samples of stomach contents and feces from each fish
297

vere dried and analyzed as in Experiment ¹1. In this manner the
organic content and caloric content of the food and feces vas
estimated.
Results:
in their stomachs than did the males with whom they were paired. The
the weight of stomach contents of their mates, speared at the same
time N - 24; Paired t-test p<0.001!. Stomach contents of male C
4
their mates N 24, Paired t-test p�.001!. This vas true even
though males vere on average larger than thei,r mates. Because male
and female 9 fZyg~ were not collected at the same- time, similar
comparisons of stomach content weights could not be made.
Stomach contents were of similar composition to those in
Experiment ¹2. For the paired species there were no significant
differences between pair members in the percent organic content of the
stomach contents organic density: 100 X AFDW/dry weight! ot' in the
energetic content per gram -caloric density expressed as cal/mg AFDW;
paired t test p ! 0.5; Table 5.4!. This is in accordance vith
observations that both males and females fed on similar foods. There
were no differences between male and female g ~~ in the organic
or caloric densities of stomach material t-test p > 0.25! .
298

0IC I
II I
40C»0 ~
~ 1 I
I.' iI
I IZs I
0W IIIS IfaR I
IIIV II
a I
~ CV ~04» »V
0 IC IC.s I
I I0»l I
~ l
lPJ CflIPII 4
I0 4 II
0» I~ Ig ~ I
4J
4I ~0'0 $
alU »
g CI
0
Ie ~ I
0» I4 Ig» I4 I 0 I
»I I»I~ 0
lh 0PgCU0 I
4
th e e 4C tv +
$»C&glhgIV
tV» Q P Q Vl
eve je jCO Ch 0!
IC» D + CI »C~ ~ ~ ~ t ~
g W e IC e ICev je jSC CO CC
Clr»ICCIelVWgg+$
» tV Al 'A»C IC~ ~ ~ ~
9 i:H"t" I'
4 hs> is%CC»O»%PJ V
» 4 ill %
ICICLE»C»C Vl
CI CC
lV »C % %~ ~
ES4" I'
g ha&0
~ Q +~ CIo
CICC EIJlv'
gaCl Ch
» CII g CIC»Ill » A
=i I
~K 2
II
5I
II I
+ 4
I hg
IIIJ
4 II ~
~gf gs
I
I ~ 4I 0 ~C»
aa4 0
~ 4~ I
I
I Z3I CJ

All three species had very long intestines. The ratio of body
length SL! to intestine length measured fram the pyloric sphincter
and 1:6.67 SD 1.31! for C frem~,
Within each species, there were no differences in these ratios between
the sexes Independent t-tests, p ! 0.8!, nor did there appear to be
differences in the shapes of male and female guts, The fecal
material at t! end of the gut had very little recognizable material
except. for la; e numbers of apparently undigested sooxanthellae in the
guts of the two coral-feeding species. Again, for each species there
vere no significant differences between males and females in the
percent organic content of the feces.
A coefficient of absorption efficiency assimilation efficiency!
of organic material, Aa was calculated using the equation adapted
from Fange and Groves l979!:
0 organic content of fecesA - 100 X 1o 0 organic content of stomach contents
A similar equation gives the calaric absorption efficiency, A
Calories per rag dry wt. of fecesA 100 X 1
c Calories per mg dry wt. of stomach contents
There was no difference between males and females within the same
species in either measure of efficiency Table 5.4!. For the two
paired species there were no significant differences in energy density
of the food intake or in absorption efficiencies between seasons.
300

There was, however, a great difference in the absorption ef ficiencies
of ~ ~i~ compared to the the remaining two species One way
ANOVA, p<0,01!, ~hae i~do ~m~b appeared to absorb two to three
times as much organic material from its diet than did the other
species, resulting in a higher net energy intake.
Experiment ¹5, Estimation of caloric partitioning among different
tissues by males and females of the three species.
Methods:
In this experiment, the energetic content of the body tissues of
the fishes of all three species used in Experiment ¹4 N - 48 C
f "MJU
After the guts were removed, the remaining tissues were divided into
gonads, liver, gut mesenteries with attached fat deposits, and the
remaining carcass. A tissue sample from the dorsal musculature was
also removed. The remaining carcass was dried, weighed, then ashed
for a determination of the ash-free dry weight. Each of the other
tissues was dried, weighed, homogenized and formed into pellets as
described above.
Ash-free dry weights and caloric contents were determined from 5
sub-samples of each tissue as described in Experiment ¹1, Because of
the small size of testes, fewer sub-samples �-5! were used. Since
the fat deposits yielded liquids when dried, the ash-free portions
301

were assumed to be lipid, and were given a value of 9,45 cal/mg AFDW
for comparative purposes. This value is the energy physiologically
available in lipids Brett and Groves 1979!.
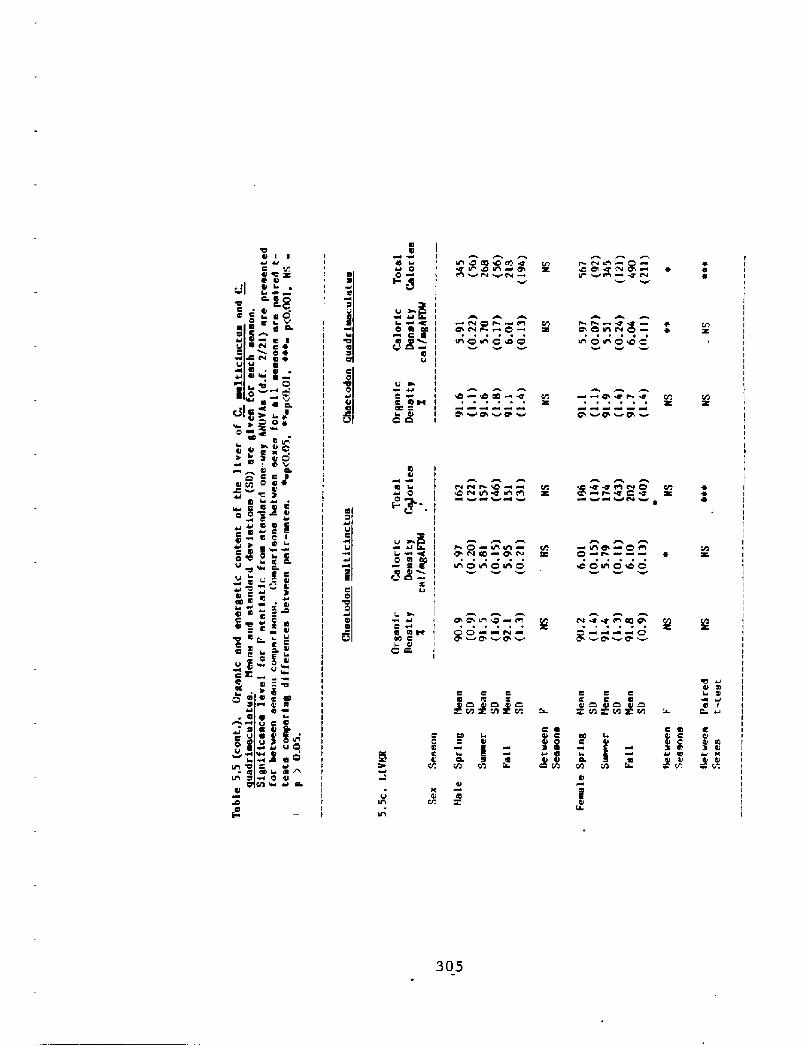
Results:
Tissues were divided into three ma]or categories: reproductive
tissues the ovaries and testes!, storage tissues the fat stores of
the intestinal membranes and the liver!, and other somatic tissues
represented by the sample of muscle tissue and the rest of the body!.
The distribution of oz'genic content and energy among these different
tissues is shown in Table 5.5 and 5,6, Organic density is represented
as a percentage ratio of ash-free dry weight AFDW! to total dry
weight of the tissue, and represents the percent of organic material
in the tissue. Caloric density is presented as cal/mg AFDV and
zepresents the energy content of organic material in the tissue. In
addition, the percentage of the total organic content of the animal
which is comprised by that tissue Fig. 5.2! provides a rough
approximation of the percent of net organic intake devoted to that
tissue. In the case of reproductive tissues, this provides a first
approximation of reproductive effort RE! for comparative purposes.
Reproductive tissues showed the greatest difference between the
sexes in tissue energy content. The ovaries of females contained 8 to
15 times as many calories as did the testes of males, even though male
302

44~ is
0 pl W gu uCa
V pm
~ S V~ Qs
O ~$Pvw»
80
Ca I &4 OP
Vl g $gh CVCI O u
U K I
-1= I O no
~ ~ e4 Vw ~ IU0
~ -.=e~4 ~ 4,
0 C ~w e»4Uii 04I Q 4 a
4 4,48UR "O
& cv el
c-IcI all IZ8
v 54 4
0 Clvt g
4 4
FORO~Oo88COIQ
WO~
4O4
Vu
IhOO040
UII
4 IIU IC rs
4 /It
a.
IVI Vl Pl
c -I
, 4 44I0. u
a a ~
4QkQJS 00 4 C 4
CI4
k~4ee0 4u4
4I 4~ v
<I e0CI 4~ 0 o4 w'
4Ig4
w 'e 4 I
4 0c 4Vmu C u4 e '4Qu~VeueV Q 4
4 hLeC0 V4 ewVZ~
44444
C ~ 04 0
4
g IC J
I
g �
8,eg
V0g CIiJ Ig w I44 % I4 C4 w44 I I
M O IJCO
c el 4
naia I'v
lv cv IVI ca
R
RR I
Pal g-C
O!W 44NA
Vl A CV tl A QI~ ~ ~ ~ ~ ~We w'QI V
mrvgma
C 0 C4 4 4g cn r Pn g Pn ca.
I
I I I
I I I I
I I I I I I I I I I I I I I II I I I I II I I I II
0I
V cl
I I
I V
I II 4I I II I I 4I 4I 4I '4I I V
I I 4I I I
0
II.'
4w
4 4+c
c
44
4
u44 4
~ CI' 0
4 4
C
C 0V
4CI u
44
V44 0
C ~uw0 0
w L4C
0V
I
3g3

Iu
3'8
4u0F I I
4vlC I
I0 IEJ 'I
Itj % I I0» I
Nit I& C IO8
4 C u CC4!au~e ~ 0
c eve
Swg .O
4 0la0
CJ
OOOV r4 M 0! 4' OO04u 0&
I I I I II I I
0 W Im 0, II «!W I0 4 A I
I I0 I I
I I I IC r I4 4 Q I
I
e V
u C 0 04!G
c 4cl C0 g
CC
0. 0 C0
we+ I 00pg 04 e.
w + g 04 u u, 0
C
caucus
C
~aeceI,3,'
~ a 4'~~ueg~'uefm.
0 Swuu u~-u-4 COIll '0 0 u~ 4 44 NAcniwoaVJ w u 4.
a 4
O O «cv n «
«Zae ON~ ~ ~I7 ~Ch»
c 4 cp A $ A gi A
OOOO
rv
C t4 hl
Om«CMPI«COP
C CC 0 CPQkRkR
III0 I
g IC IV! II0 I IIIC I4! I
I0 I0 I
!I!IIIIIIIIIIIIII0 !
0 !I
C41Cl. !!C !0 II!C !4 I IC I00
Iu I
1 0 IIC I IIIC I
SC0 Iu 4 ICRP. II!III
304
4II
0III
II4 IR Ic 4 III
V ! II!II
4»» II
e I I4PC Cl' 4 0
V L 0 «&O0! CV >&OR
«O«O IC O
CI « IoIC C teO»
0 «Ig 4
4
Cl MU 4C0 0
4CWVu e4 cCR a
4g~
e4 4Cu 404Cl
O
O O4 I4I I
II Oe0
0 I
I I I I I4 p Ijl~ I
& «92v'04
u 0
C 4 4 0 Iu II t I
ett I
I I
I I
$20,4 4P
u C 4 4C 4
O
C CC Cgl Ct p
K.
uO
c alC44
ta4 I 4C6
C 0 V4re CQ
g 'at x
colR,P
eel CO0 v4 4 4 0.C 4
C «4 «4 ~eel C ~
0 ~o»«C cvV V 44 ~
441 4«O
4 4,o/c
a
Q 0X w
ClC C0 44c 4
42C4 4U C4 0
R
U4JJ
!a
0 $
Ol0
444V Cc 44
«0C/I w
!III 4I eII eII
!II C!I 4IIII C
0I00
1t
4 V C V g C0
0 4CO
IIIIIIIIIIIIIIIII
II
W W Ct IC K' We«0I a%MN Mhl
Cl «< Ch~ ~ t
gv p4 % 'at ««v'~~Mat sAW
1 O«e~
lA O IA O IA
p ~ah«OI7l~M ChVl«P«N~
0 Olh~ $sA OvlOC!'V U'
IA 0I «O tie
M w w M co
I» »
»
CII
IV! ChX R
»»
III
Ich I» I I
II
à IZ Z III!IIIII
JJe e II 0JJ I0 I IC I
IC 4 C IIt! C 4 4 Ix 4 4 4e m K IiS P' tt ~ I IIII
IIII
305

~ I4
400
I
1 I<wK lIg v0 4 ICIC 5 'IDP» C IIP I
I I IIP K I IC m Ia ACePA C IoS
Cl > ~L4., ~84 d4 0
I=4% ca
CP g44IP 44 I=
4PPP4PPPP4
O a r
O gC
4 III'II
C C I1C I0 I III
1C I4 rp" I41 IIQ C II
4 Iaa 044 0
I 4I g
4f
Cl 44 W0
44 ~C
CL gC0
~ p1 M
vOv
CP Pcv Ch wOvO
. P O v O IC hl
0 WSWO
4 4I 4AlIC-
CCl $ CP
C
~R>40 4
5.
4 A4
C0 g C
«I44= I0 4 4
44
4
4I
4 4IP,'
IP,
ww a 404' c 4ewe4 or
u 0kJ 4Il 4 I
4
4 ww g ego 4 4
PP CAwaOCa I»0 ap~~w a
4Iaa 4p4 4 C
CL
Cl44 L =0 400 ~4 4 C
o 4 4JJ 4
4 C 44IPIJ
wwg
C L 0 /, w
I I I I I II I I I
I I I I 4I 0I I 0I UI I ClI C
I I I I I II I I I I I'I
4I I UI IPI CI I ''0I I PP
I I C4
1 I I I I II I I
I I I I I ! I I
I I I I I I
5-O"~SRvOPrtOvo
PL W A C4 V V~ ~ ~R-~-z"
I I I I I I
I I I I I I I
FHRRg~ ~ ~«Ov OvO
APPI WW O» Pl lflK a rt ~ CV hl eChwW~~ ~
I I I I I I I I
g e g IL Q Ch PPPvOvOlhO'V 'v
C C C
kÃ3EQPSEl
4 4 Cl 4
306

Ff gure 5, 2, Partitioning of organic material in different tissues of'C-~'" C-~ u tus, during the
fall, spring, and summer. Values represent the mean percent + 95%confidence limits! of. the total organic content of the body which iscomprised by the organic content of a particular tissue. The organiccontent of the tissues was determined from their ash-free dry weights.The tissues are: a! the gonads, b! the viceral fat. For each season,the sample for each species consisted of eight fish of each sex.
307

308
M %see awaaerSeason
Qyyhy QuasarSeason
8P4ay SeamerSeason
f4 %!thy herseSeason

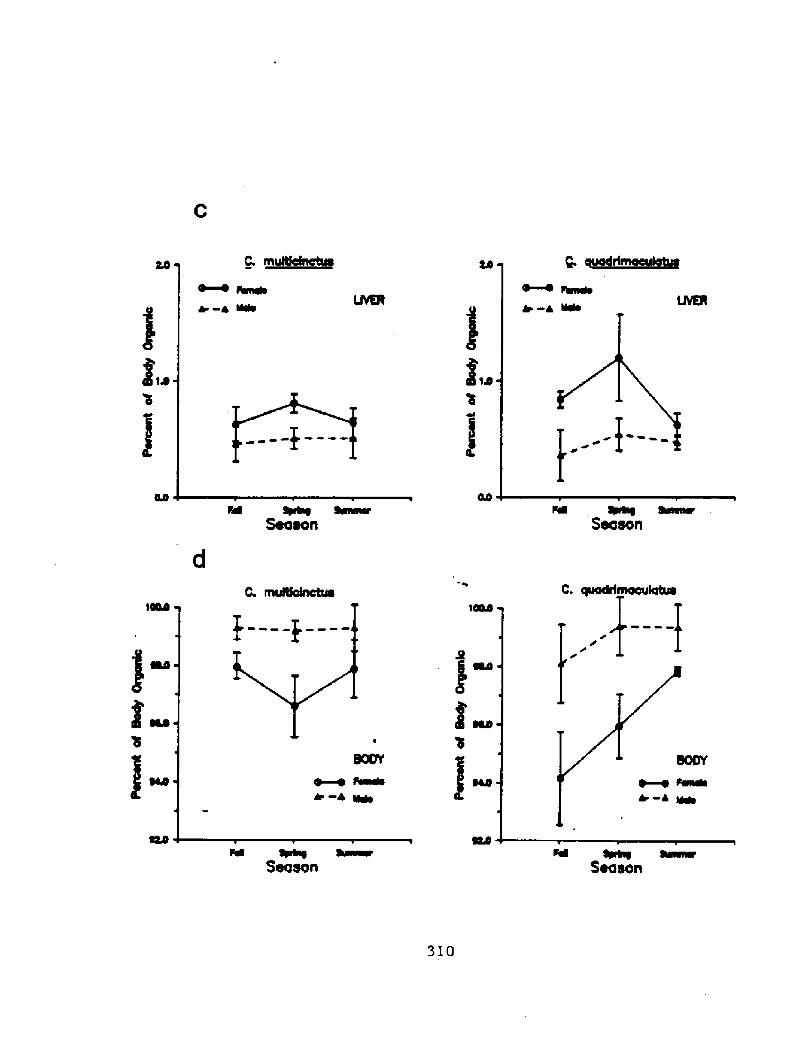
Figure 5.2 cont.! Partitioning of organic material in different
during the fall, spring, and summer. Values represent the meanpercent + 95% confidence limits! of the total organic content of thebody which is comprised by the organic content of a particular tissue.The organic content of the tissues was determined from their ash- freedry weights. The tissues are: c! the liver,: and d! the remaining bodytissues. For each season, the sample for each species consisted ofeight fish of each sex.
309
%ghee SeeeerSeason
310


















